76 F-- THE CONTROL OF ALDOSTERONE SECRETION J. P.F-OGHLAN, B.Sc. Research Biochemist, Ionic Research Laboratory D. A.7-DENTON, M.B., B.S. Sentior MjCedical Reseairch Fellowc, National Health and Alledical Reseairch Counclil, Ionic Resetarch LaboratorY J. R..rODING, MI.B., B.S. S'enior Research Fellozc, ItFool Indulstry Flund of the Commonlwea!th, Ionic Research Laboratory R. D. WRIGHT, M.S., D.Sc., F.R.A.C.P. Professor of Physiology (Fromn the Department of Physiology, University of MClelbourne, l/ictori(I, Altstralial) SOME GENERAL CONSIDERATIONS Clinical There is evidence that increased secretion of aldosterone is a contributory cause of oedema occurring in kidney, liver and heart disease. The term ' contributory ' is used in the sense of imply- ing that a number of factors act concurrently in the developed clinical state, and it would be unusual for oedema formation to be simply and com- mensurately attributable to any one of them alone. Thus administration to normal men of doses of aldosterone considerably larger than the inferred daily secretion in man with congestive cardiac failure did not produce oedema.1 Oedema is not a characteristic sign in the aldosterone secreting tumours which were first described by Conn.2, 3 Similarly with adrenalectomized dogs, Gross and Lichtlen' found that very large doses of aldosterone did not produce oedema. On the other hand, Davis, Pechet, Ball and Goodkinds found that aldo- sterone secretion was increased in dogs with experimental right heart lesions which caused congestive failure, and in dogs with thoracic inferior vena cava constriction producing ascites. If such dogs were adrenalectomized, administra- tion of adequate electrolyte-active hormone was essential for oedema formation to occur.6 The developed clinical syndromes are similar to many other physiological responses and pathological states in so far as the ' cause ' is identifiable as the concurrent action of a number of factors which are severally necessary and jointly sufficient. In the particular instance of cardiac oedema, substantial studies have been made to evaluate other factors postulated in the ' forward ' and ' backward failure theories, and we do not propose to ente this discussion here. It is this multiplicity of concomitant and conse quent phenomena in clinical conditions whic Inakes elucidation so difficult. As an example w may consider the enormous excretion of aldo sterone by nephrotic patients. Using promising new modification of the original isoto dilution method7, 8 of estimating aldosteron secretion, Ulick, Laragh and Lieberman9 foun up to 6,600 T/day. The etiology of nephrosis i unknown and therefore the direct effect of th etiological conditions on the adrenal cannot b tested. It is possible that the nephrotic kidne produces a substance which activates the adren but this hypothesis cannot be tested until suc material is identifiable by assay. Through un known mechanisms the excess secretion mav resul from disturbance of electrolyte levels or dis tribution or from abnormal distribution of bod fluid. Abnormal metabolic degradation of aldo sterone by the liver or other tissues may cau excess secretion through some feed-back mecha nism to the adrenal. The elucidation of an clinical condition is impossible without under standing every phenomenon in the disea mechanism to the stage where each can b separately identified both qualitatively an quantitatively. Experimental Physiology The principal physiological questions involvee in the control of aldosterone secretion are, (i) th by copyright. on February 1, 2022 by guest. Protected http://pmj.bmj.com/ Postgrad Med J: first published as 10.1136/pgmj.36.412.76 on 1 February 1960. Downloaded from
Transcript
76
F--
THE CONTROL OFALDOSTERONE SECRETION
J. P.F-OGHLAN, B.Sc.Research Biochemist, Ionic Research Laboratory
D. A.7-DENTON, M.B., B.S.Sentior MjCedical Reseairch Fellowc, National Health and Alledical Reseairch Counclil, Ionic Resetarch LaboratorY
J. R..rODING, MI.B., B.S.S'enior Research Fellozc, ItFool Indulstry Flund of the Commonlwea!th, Ionic Research Laboratory
R. D. WRIGHT, M.S., D.Sc., F.R.A.C.P.Professor of Physiology
(Fromn the Department of Physiology, University of MClelbourne, l/ictori(I, Altstralial)
SOME GENERAL CONSIDERATIONSClinical
There is evidence that increased secretion ofaldosterone is a contributory cause of oedemaoccurring in kidney, liver and heart disease. Theterm ' contributory ' is used in the sense of imply-ing that a number of factors act concurrently in thedeveloped clinical state, and it would be unusualfor oedema formation to be simply and com-mensurately attributable to any one of them alone.Thus administration to normal men of doses ofaldosterone considerably larger than the inferreddaily secretion in man with congestive cardiacfailure did not produce oedema.1 Oedema is not acharacteristic sign in the aldosterone secretingtumours which were first described by Conn.2, 3Similarly with adrenalectomized dogs, Gross andLichtlen' found that very large doses of aldosteronedid not produce oedema. On the other hand, Davis,Pechet, Ball and Goodkinds found that aldo-sterone secretion was increased in dogs withexperimental right heart lesions which causedcongestive failure, and in dogs with thoracicinferior vena cava constriction producing ascites.If such dogs were adrenalectomized, administra-tion of adequate electrolyte-active hormone wasessential for oedema formation to occur.6 Thedeveloped clinical syndromes are similar to manyother physiological responses and pathologicalstates in so far as the ' cause ' is identifiable as theconcurrent action of a number of factors whichare severally necessary and jointly sufficient. In theparticular instance of cardiac oedema, substantialstudies have been made to evaluate other factors
postulated in the ' forward ' and ' backwardfailure theories, and we do not propose to entethis discussion here.
It is this multiplicity of concomitant and consequent phenomena in clinical conditions whicInakes elucidation so difficult. As an example wmay consider the enormous excretion of aldosterone by nephrotic patients. Usingpromising new modification of the original isotodilution method7, 8 of estimating aldosteronsecretion, Ulick, Laragh and Lieberman9 founup to 6,600 T/day. The etiology of nephrosis iunknown and therefore the direct effect of thetiological conditions on the adrenal cannot btested. It is possible that the nephrotic kidneproduces a substance which activates the adrenbut this hypothesis cannot be tested until sucmaterial is identifiable by assay. Through unknown mechanisms the excess secretion mav resulfrom disturbance of electrolyte levels or distribution or from abnormal distribution of bodfluid. Abnormal metabolic degradation of aldosterone by the liver or other tissues may cauexcess secretion through some feed-back mechanism to the adrenal. The elucidation of anclinical condition is impossible without understanding every phenomenon in the diseamechanism to the stage where each can bseparately identified both qualitatively anquantitatively.Experimental PhysiologyThe principal physiological questions involvee
in the control of aldosterone secretion are, (i) th
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
IIbnar' 196o C'()(;1IIAN, I)EN'I'()N, (;01)IN(; and \VRI(;Ht'I' ('oltrol of .lldosteroc Scrlctlol 7
tomical site of the receptor (or receptors)lating aldosterone secretion, (ii) the exact
ture of the stimulus to it, (iii) the site of origin,de of transmission and nature of the stimulus toadrenal, and (iv) the manner of the adrenal
ponse. There apears to be some conflictopinion as to the present status of know-ledge ination to these questions. 'IThe opening sentencethe Year Book of Endocrinology (1958-59)
itorial dealing w-ith the adrenal cortex states,ne of the most fascinating developments of thet year is the discovery of the vway the body
ntrols the release of aldosterone.' Thoughain of the studies cited in support of thistement are evidently of great importance, it is,our view-, an over-optimistic assessment of theesent positioni. In fact on many crucial issuesere are flatly contradictory findings in recentrature. This is as likely to be a source ofcultv and confusion for someone seeking a
neral review of the field as it is for those directlyncer.ed in experimental investigation of theestionis cited above. For this reason it may belpful at the outset to state four main factors inperimental approach, the consideration ofhich may identify the causes of some con-cting findings.actor I--'/1( Operation of Measurei,entThe primary aim of animal experiment is to haventrol of all the relevant coinditions and be able tod the effect of controlled variation of one or morethese conditions. It is essential that the processmclasurement shoild niot iltrodluce ain iuncontrolledundesired 'variable. In all cases, it may beumed that the base-line levels w-ill be found ille confident conscious animial. For Nwork on thenscious animnal, training to indifference to theserver and what he does is essential. NWrhen thenctional tissue for investigation is anatomicallyp-seated and difficult of access, surgical pro-dures of considerable complexity over a longriod may be required in order to contrive theessarv direct access in the conlscious undis-bed state.Pavlov was the first person to enunciate clearlye principle and take advantage of aseptic surgeryimplement this approach. He recognized
at the crude damage to the physiology of theganism -which occurs in the acute surgicalerimenit unider anaesthesia may evoke a chainreactions Nwhich frequently overshadows or
dicallv modifies the normal function underutiny. As Verney has pointed out, initialeriments with postpituitary extract made onesthetized animals led to the belief that theysiological role of this substance w-as to promoteuresis. In the course of his own work, Verneyl'
found that a normal *w-ater diuresis did not occur inanaesthetized or frightened dogs, and we haveshown that normal responses to rapid changes of\Na balance do not occur in anaesthetized orfrightened sheep.1l An outstanding example ofsuccessful implementation of this principle ofapproach Nwas the development by Lockett,O'Connor and Verney-'2 of the renal artery looppreparation permitting direct access to theartcrial supply and venous drainage of the kidneyin a conscious dog.
HowN-ever, in advocating this principle, parti-cularlv where it is recognized that higher nervousactivity plays an important role in the physio-logical regulation concerned, it obviously cannotbe oxerlooked that facts of great importance havebeen discovered by the method of acute experi-ment. Apart from examples such as Harvey'sobservations by vivisection, and the discovery ofsecretin, the early observation by Verney andStarling13 of the profuse urine flosw in the heart-lung-kidney preparation was the point of departurefor Vernev's analysis of the control of secretion ofantidiuretic hormone. 'rhe simple three organpreparation provided the basic fact of absence ofan inhibitory influenice, but it is extremely unlikelythat the discovery that the physiological stimulusto antidiuretic hormone control was contemporarysmall variation in the osmotic pressure of carotidartery blood could have been made except by thesurvival type of experiment. In other publica-tions14 1-we have set out in detail the desideratafor valid determination of the cause and manner ofsecretion of a hormone by an endocrine gland.They are:
I. All observations should be made on consciousulndisturbed animals in normal relationship withtheir environment. Thus the results derived arethose holding for a normal animal.
2. The physiological stresses causing a responsedependent upon the gland should be preciselydefined and standardized, and be capable ofgraded variation.
3. 'The arterial blood supply of the endocrinegland under investigation should be exclusivelyaccessible so that the effect of supposed activeagents can be tested by direct local injection.
4. 'The nerve supply of the gland, when rele-vant, should be accessible for stimulation orblocking.
5. 'rhe venous effluent exclusively from thegland should be accessible.
6. 'l'here should be one or more biologicalsystems for quantitative assay of the activity of thevenous effluent.
7. The chemical assay of the venous effluentmust be developed until it detects quantitativelyeaich substance known to be active on the bio-
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
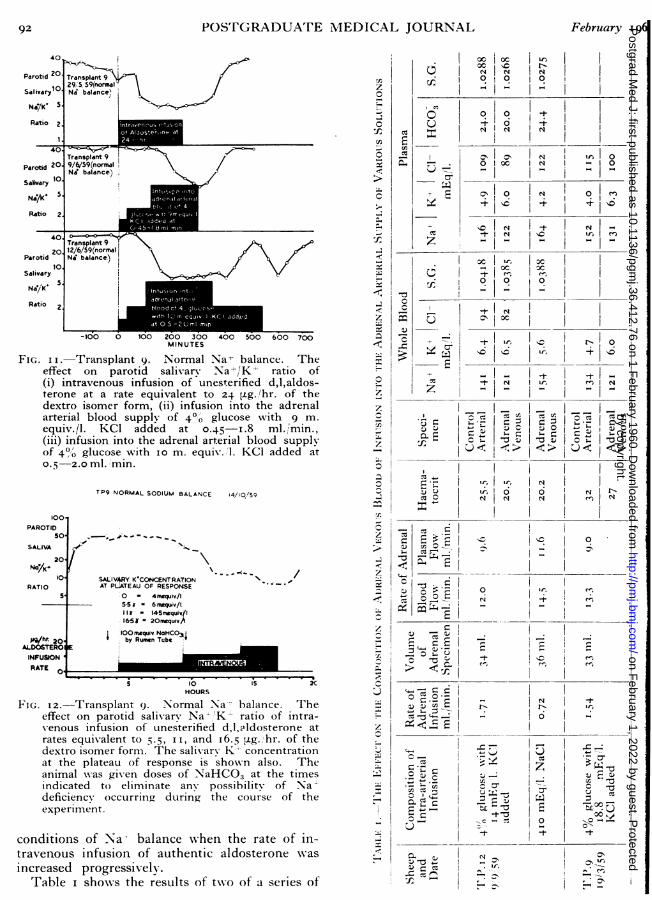
1(;. I.--Transplant 12. A photograph show-ing the skin loop containing thecarotid artery, jugular vein, and the left adrenal gland. A pnleunlatic cuffinflated to 300 Ilm. of I1g. has been applied to the lo,;, cranial to thetransplant. A needle attached to polythene tubing has been inserted inttothe carotid artery caudal to the transplant, and an infusio,n is being nsmadeinto the arterial blood supply of the adrenal gland. T'he transplant opera-tion was carried out two y-ears earlier.
logical inldicator, anrd the o\verall effect of thesesubstances must correspond quantitatively to theaction of the entire venous effluent.Some experimental procedlures ainLnl,, to mieet
the desiderata. It would be ideal for investigationof aldosterone regulation if it w-ere possible to havea continuous record of the adrenal blood flow andthe arteriovenous difference in aldosterone con-centration in a conscious animal ullder appropriatephysiological stress. T'Ihe amount of bloodremoved for analysis should not, of itself, stimulatesecretion unless desired. In circumstances whereit is impossible to contrive this end or as a matter ofconvenience, an alternative would be to have acontinuous index of the concentration of aldo-sterone in peripheral blood. This concentrationis dependent upon the rate of secretion, the rate ofdestruction and excretion, and the volume of fluidin Nwhich the hormone is distributed. Formally,the possibility of storage has to be considered also,and this includes the question of protein binding.In the instance of concurrent chemical and bio-logical assay of peripheral blood, a differencecould be attributable to the fact that protein-bound steroid Nwas biologically inactivNe. W\\hereasvwith cortisol this is an important consideration,evidence to date is against storage or significantprotein binding of aldosterone. If onie assumesthat the rate of degradation andt excretioni of aldo-sterone is proportional to the peripheral concetn-
tratiotn, the short half-life of aldosterone woulresult in the peripheral concentration following insmoothed-out fashion variations in rate of secre-tion. In that chemical assay of aldosterone iperipheral blood requires large volumes andistill iln the developmental stage, the better alternative is a biological indicator.
'I'he parotid gland of the ruminant is such aindicator,' respondingi with approximatcly 6o t8o minutes delay to starting or stopping aipsilateral intra-arterial infusion of aldosterone.Corrections for parotid secretion rate variationiand increased sensitivity of the gland in Nadepletion'l are necessary to make the salivaNa' K'a satisfactory index of peripheral blooaldosterone. Notwithstanding the limitations, thsalivary N'a K is a valuable continuous indeof the secretion of aldosterone in the consciouanimal. ''he great advantage of the parotifistula preparation in the ruminant is that desideratI, 2 and 6 are met. With respect to 2, the fistuladepletes the animal of Na' rapidly, and the degreof deficiency can be controlled by administerinNaHCO', by rumeni tube or allowing the animal aappropriate a)nount of NaHCOa or NaCI tdrink.'8. I!
To meet the desiderata 3, 4 and 5 outlined and,in particular, to determine whether the adrenal istimulated directly by the local changes of ionicoInpositioii of arterial blood, an adrenial transplant
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
truary 196o CO(;HLAN, DENTOI'N, (;ODING( and WRIGT(;I': ('oCltrol of .-Alosterolc Secretio, 7()ration has been developed in this laboratory.left adrenal gland of the MIerino sheep isplanted to a combined carotid artery-jugularskin loop in the neck.20 Thus the composi-of adrenal arterial blood may be locallyd and the adrenal steroid output can betly measured in the adrenal venous blood of
conscious undisturbed animal (Fig. i).The implementation of the desiderata has re-ed a number of facts about adrenal secretion*ch would have been difficult to determine in avincing manner in anll other way, e.g., changesionic composition of adrenal arterial bloodctly affect aldosterone secretion. However'c changes do not account for the known varia-n of adrenal secretion seen with change of Nance, and it is clear that some additional factorfactors act upon the adrenal via the bloodlam. hen the transplants w-ere used for-circulation experiments, it 'was shown thatblood of a Na--depleted adrenalectomizedal stimulated the adrenal transplant of ap in normal Na' balance to produce electro--active adrenal steroid.'4The new method of aldosterone assay in bloodeloped by Kliman and Petersolfi'2, 22, 23 permitsrate assay of small serial samples of adrenaldso that parotid salivary Na /K-' ratio may beelated with aldosterone production over longiods of time without loss of blood and itstained aldosterone becoming an undesiredable in the experimental situation. Thecurrent use of both indices, i.e. episodicmical assay of adrenal venous blood, and con-uous biological assay of peripheral blood, toe extent circumvents the shortcomings result-from the use of either method alone. \NWith theival type of experiment there is the overallantage that the trained animal can be used asown control over a period of years, and thets of chromatographic assay can be compareder identical conditions with the effect ofous rates of intravenous injection of aldo-ne itself and other steroids. Thus theysis outlined in desideratum 7 can be maderously.rocedures for meeting desideratum 5 have beenribed by Hume and Nelson24 and by WeaverEik-Nes.'25vestigation by, acute experiment. Mlost of theculties of interpretation arising from defects of[gn in these regards are exemplified in therts of Farrell and co-workers.26-32 Usingesthetized or decerebrated animals, the totalus outflow from one adrenal is collected for aber of hours to provide one sample for a singlemical analysis. Sometimes specimens areled. No baseline for the individual animal is
Blood Adrenalloss A a B C A DdE - Aldosterone
t Secretion
t Rx
Nadeficiency I
Anaesthesiaand trauma
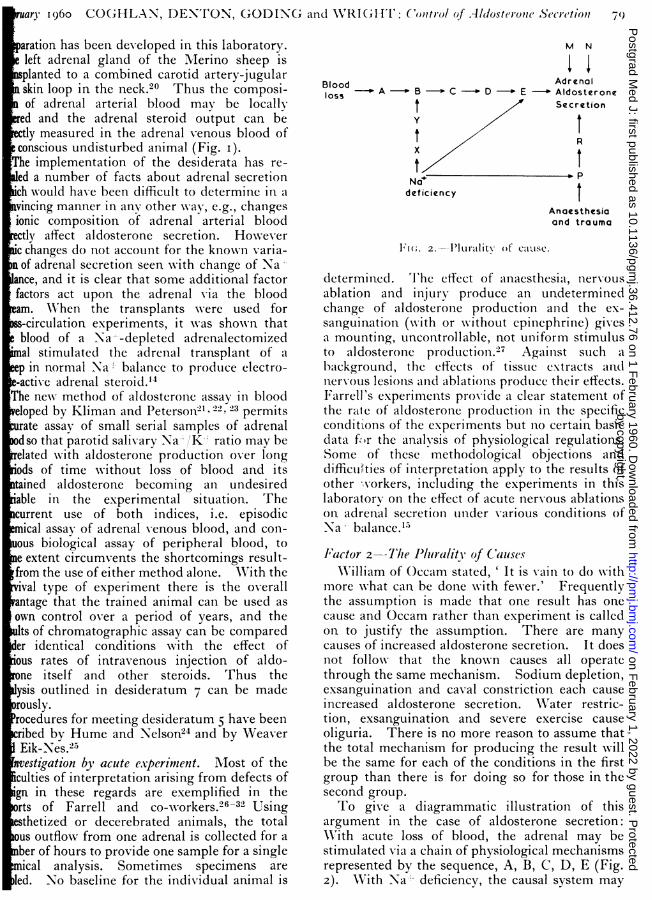
l1"(;. 2.--I'lutl1litV Of catsc.
determined. 'I'he effect of anaesthesia, nervousablation and injury produce an undeterminedchange of aldosterone production and the ex-sanguinlation (w-ith or w-ithout epinephrine) givesa mounting, uncontrollable, not uniforin stimulusto aldosterone production.-' Agaiiist such abackground, the effects of tissue extracts alndnlerv-ous lesions and ablatioins produce their effects.Farrell's experimenits provide a clear statement ofthe rale of aldosterone productionl in the specificconditions of the experiments but no certain basicdata f~,r the analysis of physiological regulations.Some of these methodological objections anddifficuilties of interpretation apply to the results ofother ,-orkers, including the experiments in thislaboratory on the effect of acute nervous ablationson adrenal secretion under various conditions ofNa balance.l-
I'actor 2--7The P]Irality of CauLsesNWilliam of ()ccam stated,' It is vain to do with
more w-hat can be done w-ith fewer.' Frequentlythe assumption is made that one result has onecause and Occam rather than experiment is calledon to justify the assumption. There are manycauses of increased aldosterone secretion. It doesnot follow that the known causes all operatethrough the same mechanism. Sodium depletion,exsanguination and caval constriction each causeincreased aldosterone secretion. Wrater restric-tion, exsanguination and severe exercise causeoliguria. There is no more reason to assume thatthe total mechanism for producing the result willbe the same for each of the conditions in the firstgroup than there is for doing so for those in thesecond group.To give a diagrammatic illustration of this
argument in the case of aldosterone secretion:With acute loss of blood, the adrenal may bestimulated X-ia a chain of physiological mechanismsrepresented by the sequence, A, B, C, D, E (Fig.2). WVith Na-- deficiency, the causal system may
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
involve action on the adrenal by more than onepathway and, as a corollary, interruption of onepathway may not preclude action via another. Inthe two instances, the operation of the causal chainmight require the existence of certain standingconditions (represented as M and N). Assumingthis analysis to be representative of the state ofaffairs in the conscious undisturbed animal, it isformally possible that the combination of, e.g.,Na' deficiency or exsanguination with anaesthesiaand surgical injury could institute a causal chainwhich did not operate under other circumstancesand which would continue to act despite disruptionof the normal physiological chain. This is shownas P, R, on the diagram.John Mill made the following statement: '
It is not true ... that one effect must be connectedwith only one cause, or assemblage of conditions;that each phenomenon can be produced only inone way. There are often several independentmodes in wXhich the same phenomenon could haveoriginated. One fact may be the consequent inseveral invariable sequences; it may follo, w-ithequal uniformity, any one of several antecedents,or collection of antecedents. Many causes mayproduce mechanical motion: many causes mayproduce some kinds of sensations: many causesmay produce death. A given effect may really beproduced by a certain cause, and yet be perfectlycapable of being produced without it.'3
Another possible source of misapprehension isthe transference of notions derived from theresults of one operational set of conditions to fillin gaps in a superficially similar but actuallydifferent set of conditions. It must be uncertainthat the effect of mid-collicular decerebration onthe aldosterone secretion in the Na depletedanimal will be the same as its effect on aldosteronesecretion resulting from exsanguination eventhough both investigations use the anaesthetic-traumatized animal. Section or ablation in thenervous system may produce acute effects byparalysis, irritation or release; long term resultsare modified by compensation and plasticity.Comparisons must therefore be on the basis ofprecisely defined anatomical, chronological andphysiological similarity and variation.The possibility of variation in physiological
organization for one functional result in highermammalian types must be considered also, butit is unlikely that such a basic mechanism as theregulation of ionic content wvould show any radicaldifferences.
Factor 3-Chemical MethodsA third reason for conflicting findings is varia-
tion of reliability of chemical methods for detectionof aldosterone in biological fluids. This is
CH20 H OH CH OHc=O 0 CH
H0 CHO
O 0
A B
H2,OHc=O
CH20H 0
00H H
FI(;. 3.--Chemical formulae.
particularly so in the case of meCasuremenitaldosterone in urine.34 3I ,29
I'actor 4-The Criteria for Hormonal A ctizvit'tExtracts of Biological MaterialA putative hormone may be isolated from bloo
excreta, or the supposed site of origin. Tcriteria for the full categorization of its role ashormone are (i) demonstration of its presencethe site of origin, (ii) the disappearance of tphysiological effect attributed to it upon removof the site of origin, (iii) demonstration of activiby infusion of the material exclusively into tharterial blood supply of the target organ. Thieliminates the possibility of indirect action Xviaintermediate chain of causation. (ixv) Demonstration of transmission by the blood stream atconcentration commensurate wvith the elicitiphysiological stress.
Tissue extracts active on an organ are not necesarily in a hormonic relation to that organ, e.bronchial extract may activate gastric secretiobecause of the histamine content.
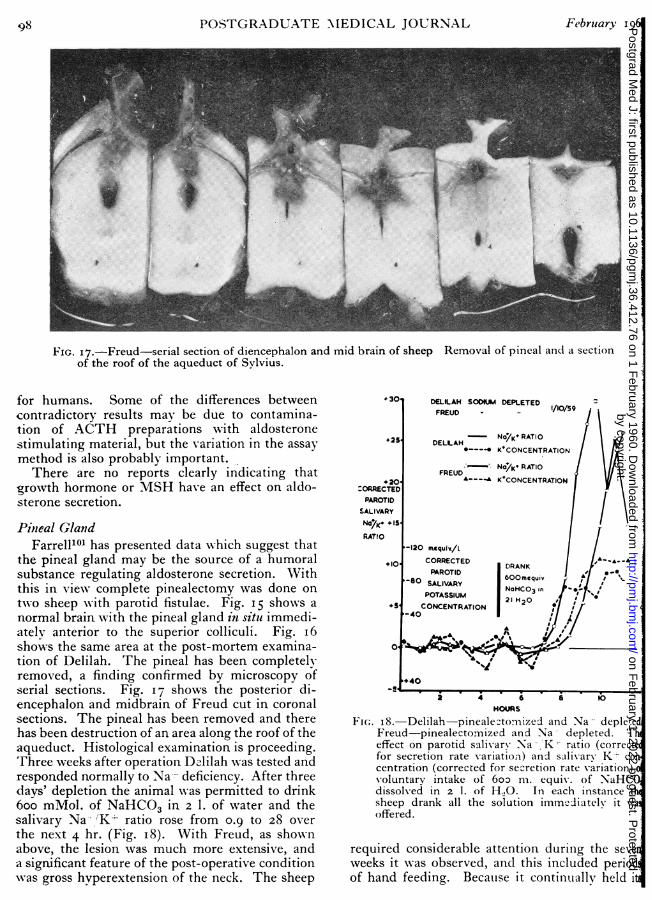
Deane, Shaw and Greep36 shoNwed that variatioof Na-L and K-i- balance caused cytological changof the zona glomerulosa indicative of altereactivity. 'IThere is evidence from the studies ofnumber of workers that there is a hornonstimulus to aldosterone secretion. Farrell2!' hsuggested the name ' glomerulotropic hormoneor ' glomerulotropin' for the substance asexercises its trophic action on the glomerulosHowever, there is no experimental evidence to dathat this material has any trophic activity as defineabove, i.e., removal of the site of origin an
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
ebruary 1i960 COGHLAN, DENTON, GODING and WRIGHT: Control of Aldosterone Secretioni 8r
monstration of zona glomerulosa atrophy in theival preparation. At this preliminary stage, it
Fght be better to name the postulated substanceglomerulosa stimulating hormone (GSH).'These general observations introduce the de-iled sections which follow. Several excellentview articles which have appeared in the lasto year29, 40. 37, 38 39 cover most aspects ofonological development in the field so Xwe will
t recapitulate in detail here.
MICAL STRUCTURE AND PROPERTIESF ALDOSTERONEAldosterone is unique among the adrenalroids in possessing an aldehyde grouping onsition I8 (Fig. 3, A). In aqueous solution itts largely as the i8 - o - Ii hemiacetal
ig. 3, B).41 42The provisional formula of a metabolite of aldo-rone43 which occurs in human urine in greaterounts than free aldosterone is shown (Fig. 3c).
teroid compounds structurally related to thediac glycosides and with the property of block-g the action of aldosterone on the renal tubuleve been developed. The structure of two suchmpounds is shown (Fig. 3, D). In the steroidC5233, R is CH3, in steroid SC8Io9 (Spiro-tone), R is H.44-47Other compounds have the property of blocking
production of adrenal steroids, includingsterone.48 49 Examples are amphenoneand 2-methyl-I ,2-bis-(3-pyridyl)- I-propanoneU4885), the latter having selective action on-beta-hydroxylation.50ction of AldosteroneThe biological actions of aldosterone cause a
in Na+ and a fall in K+ in the extracellularid (E.C.F.). There is retention of Na+ andcretion of K,51, 52, 53 by the kidney, and a fallthe Na+/K+ ratio of saliva and faeces.1 Inte experiments54 it has been reported to cause a
in Na(E.C.F.)/Na(I.C.F.) and a rise in(I.C.F.)/K(E.C.F.) ratio in muscle and brain,though in chronic primary hyperaldosteronisme reverse occurs. This paradoxical situation is
idered later.
fect of Lack of AldosteroneIf aldosterone secretion is stopped or the peri-eral action is blocked there are the followingects-an increase in urinary Na+ excretion and arease in K+ excretion, a rise in Na+/K+ ratiosweat55 and saliva56, 57, a rise of intracellular Na.d a fall of intracellular K+, and a fall of E.C.F.a+/K+ ratio. Oedema and ascites associatedith raised aldosterone production may be reducedeatly.58, 47 Clinical conditions of selective aldo-
sterone deficiency are being recognized, parti-cularly following removal of adenomata in cases ofConn's syndrome.The effect of deficiency of electrolyte-active
secretion appears to differ between animal types,e.g. many adrenalectomized humans can be main-tained on cortisone alone, whereas, with anadrenalectomized sheep on its normal diet, thewithdraw-al of electrolyte-active hormone causesdeath within four days.57 In a sheep with severeadrenal insufficiency, aldosterone administrationdoes not seem to be as effective as a life-savingprocedure as DOCA. Swingle59 found thataldosterone was ineffective in the resuscitation ofthe adrenally-insufficient dog.
hydroxy-pregnane-20-one and 3-t- I -6- x-di-hydroxy-allopregnane-2o-one have been isolated.60The former was recovered from the urine ofpatients with salt-losing adrenogenital syndrome.In this condition, a very low output of aldosteroneoccurs despite Na'X deficit.61, 62, 63:Clinical Conditions which have been Reported toCause Increased Aldosterone ExecretionThese can be broadly sub-divided into eight
(iii) Conditions associated with oedema orascites (congestive cardiac failure, cir-rhosis of the liver, nephrotic syndrome,hypoproteinaemia, idiopathic oedema).
(iv) Conditions associated with disturbances ofother endocrine systems (hyperthyroid-ism, hypothyroidism, pregnancy, insulinshock).
(v) Adrenal cortical hyperplasia and tumours(Conn's syndrome).
(vi) Conditions involving the C.N.S. (midbrainvascular accidents, cyclical insanity,anxiety states).
(vii) Change of position and activity.(viii) Others including carcinoma of lung with
or without adrenal metastases.
Relative Electrolyte Activity of Aldosterone Com-pared with other SteroidsMany naturally occurring steroids, e.g. cortisol,
cortisone, corticosterone, testosterone, proges-terone, if given in large doses cause Na+ retention.Desaulles63a showed that aldosterone was 2,000times as effective as corticosterone in causing
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
I:(c;. 4.-Basil, adrenalectornized and Na deficient.The effect on parotid salivary Na'K' ratioof intravenous infusion of unesterified d,l,aldo-sterone in isotonic saline at rates equivalent to7.5, 15, and 3Otg. hr. of dextro isonier form. Thearrows mark the time of cessation of infusion ineach instance. The mean salivary K- concentrationat the plateau of response is recorded also.
GEORGE 24 7 sa2.e't,,? J': J,-JJ.' ,
P04otid
ohvaryNa/Kratio
20
mg hr
:ymsi-yccinat?,rocortison?
rsng. D O C Aintrovcnou$1y +intra muesc
2 4 6 8 10 (2HCaRS
FIGc;. 5.--George, adrenalectomized and Na- deficient.The effect on parotid salivars Na-'K-- ratio of(i) intravenous injection of 20 mg. of hydrocortisonehemi-succinate folloNwed by continuous infusionat the rate of io and then 20 mg. hr.; (ii) intra-muscular injection of 5 mg. of DOCA.
urinary Na retention. We have found that i.%v.aldosterone is over i,ooo times more potent thancortisol in causing a fall in salivary Na-- K" ratio(Figs. 4 and 5). When given intramuscularlvaldosterone is 40 times as effective as DOCA(Fig. 6).The Normal Secretion Rate, C'oncenttration inPeripheral Blood and .MletabolismThe secretion rate of aldosterone in normal
human subjects has been estimated by Ayreset al.64 to be 170 to I90 y/day, Ulick et al."' foundI5 oto 300 y/day, Peterson23 330 to 400oo day.This agrees approximately with an amount of
GEORGE
20
salrary
Na;'K 2r .ratio I .- d.. Aldosterone,y
MSg im.x----xDOCA. V
oS . 20mg. im
2 4 6 8 10HOUR S
FlI(;. 6.--(;eorge, N-a depleted and adrenalectonmizl'he effect on parotid salivary Na,' K' ratio(i) intramuscular injection of 1mg. of d,l,aldosterom(onoacetate; (ii) intramuscular injection of 20-of desoNxvcorticosterone acetate.
250 , reported by MIach et al."51 as the daily rplacement dose for maintenance of patients wiAddison's disease. The concentration of aldsterone in peripheral blood in normal subjects w0.o4 to o.08 I00oo mi.,23 while Ayres et al.8 report0o.o03 1I00 ml.
If tritium-labelled aldosteroiie is injected innlormal subjects, approximately 70 per cent of tadministered dose appears in the urine in the fi24 hours. The free steroids make up 4 to 8cent., metabolites conjugated as glucuronid30 to 40 per cent., and those released after pHincubation, Io to 20 per cent.'3 The tetrahymetabolite (Fig. 3, C) Nwhich occurs as a conjugw-ith glucuronic acid accounts for I0o to I 5 per ceof the total radioactivity appearing in the urin
Methods of Estimating AldosteroneC'hromatographyThe initial isolation of aldosteronie was effect
bv the application of column and paper chromatgraphic techniques to the fractionation ofadrenal extracts into their Xvarious adrenocorticoicomponents.41 As no specific chemical methodavailable which can reliably determine aldosteroin a mixture of corticosteroids, paper chromagraphic separations have'been used for the isoltion of aldosterone from biological fluids in a fosufficiently pure to permit accurate quantitationone or other of the non-specific methods.The basic chromatographic systems gener
used are those of Bush65 and Zaffaroni.66 Mimodifications have been introduced by maworkers. There is 1no univ-ersal accord as tomost suitable combination of chromatograpsystems to achieve the high degree of purirequired, but the failure of one or two systeto give reliable resolution is well docInented.3aa 67, 68. 69 Probably the best systemschromatography and derivative formationassist purification can be selected only in the lig
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
uary I960 COGHLAN, DENTON, GODING and WRIGHT: Conttrol of Aldosterone Secretionl 83
nsiderable experience with the particulares of experimental animal, and the type ofcal material being extracted.
ports on the many chromatographic combina-employed have not always included compre-ive details of recovery experiments.2",69 Theding method available to date appears to be
ouble isotope derivative assay and the doubledilution derivative assay of Kliman and
sn.21, 22, 23. 70
ey summarize the method as follows: ' Ale labelling technique which has provedin this laboratory for the assay of many
ids is applicable to the selective assay ofterone. Dried extracts of plasma or urinecetylated with tritium-labelled acetic an-ide in the presence of pyridine and benzene toert aldosterone quantitatively to the tritium-ed diacetate. A measured amount of authen-dosterone diacetate-C'4 is added to eachle, and the double-labelled steroid purifiedaper chromatography. After tw-o chromato-hies, the samples are treated with chromic acidacial acetic acid to form a monoacetate oxida-product, and are subjected to a final chro-aphy to separate the aldosterone from other-labelled materials. The tritium and
n-14 content of the purified steroid ised by simultaneous counting in a liquidlation spectrometer. The aldosteronent in the original extract can be calculatedthe determination of the amount of carbon-14tor lost during the purification, the yield ofradioactivity, and the specific activity of the-labelled acetic anhydride. An efficient
can be done with 5 to io ml. of urine or 2 toadrenal vein plasma. This method will
rely determine o.oi jg of aldosterone, andthe accuracy characteristic of isotopenques.'ntly it has been reported that, by using
gnated glass paper, a rapid and satisfactorytion of small quantities of steroids includingterone can be achieved.7' Possibly, in con-hon with one or other of the isotopic methods,time could be saved provided the same highof purification can be achieved. Further
ess along these lines will be of great interest.
luaye methods normally in use depend on aldo-ne causing Na+ retention and increased K-tion by the kidney. The usual experimentals have been adrenalectomized mice, rats
dogs.72-76 The material to be assayed isd into animals recently adrenalectomized
which are loaded with Na'. The alterationa+ and K- excretion is estimated either by
flame photometry77 or by radio-active isotopemethods.78 This change is compared with theeffect of a standard dose of DOCA or authenticaldosterone.The advantage of the use of the rat or mouse is
that minute amounts of aldosterone mav be as-sayed. These animals, however, excrete smallvolumes of urine, and the resultant change inelectrolNyte excretion due to aldosterone is not large.The method does not permit use of the animal asits own control, and depends on statistical analysisof difference between the control and the experi-mental group. Thorn et al.34 state that because itis necessary to remove interfering steroids69 beforeproceeding to bioassay there may be large lossesof aldosterone which Nwould obviously affect thereproducibility of the results. Difficulties ofinterpretation of results using a rat bioassay areevident in the paper of Orti et al.76 in which themethod was used to demonstrate the presence ofan aldosterone stimulating substance in urine.The effect of aldosterone on the parotid salivary
Na-";K- ratio of the sheep will be discussed morefully in a subsequent section. For bioassaypurposes, an infusion can be made into theipsilateral carotid artery loop. It has been shownthat in the Na-- depleted adrenally insufficientsheep as little as o.5 -,.`hr. of aldosterone causes anunequivocal fall in salivarv Na+'K- ratio after adelay of 90 to 120 minutes. A vascular isolationprocedure contriving that the ipsilateral carotidartery supplies the parotid gland only has leendevised in the sheep. It is possible that this pre-paration will give reliable results with a smallerrate of aldosterone infusion. A possible objectionto the use of ipsilateral intracarotid infusion forthe purpose of comparing the biological activitiesof different pure steroids resides in the likelihoodof very different degrees of protein binding. Thetime of transit between the tip of an intracarotidneedle and the parotid gland is probably I to 2sec.,10 and if this time were inadequate for thecompletion of binding, the procedure could give aphysiologically erroneous estimate of the electro-lvte activity of a steroid such as cortisol which isnormally 80 to 95 per cent. bound.79-8' 23 Thesame objection does not hold for the use of theprocedure for bioassay of electrolyte activity inwhole blood.
Methods of Assessing Secretion ofAldosterone in Biological Experiments-Methods Depending on Direct Access to the AdrenalGland
In vitro studies of steroid secretion have beenmade by many workers.82-86 Giroud et al.87 haveconfirmed earlier evidence of Dean et al.36 thatelectrolyte active corticosteroid is secreted princi-
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
pally by the zona glomerulosa. On the otherhand, Lane and de Bodo88, 89 found in the dogthat there is not the same atrophy of the fasciculataand preservation of zona glomerulosa after hypo-physectomy as in the rat. The results obtainedfrom in vitro preparations must be accepted withcaution when they purport to yield informationrelating to normal physiological control of adrenalsecretion. However, this method might provideuseful information when ' screening' extracts ofpossible aldosterone-stimulating activity, providedthat such extracts are subsequently tested in thewhole animal under the conditions set out above(Factor 4).Mulrow et al.86 have summarized some explana-
tions possible when crude extracts cause increasedsecretion of aldosterone with in vitro preparations.The action may be due to (a) a true physiologicalstimulus to the adrenal gland, (b) provision of anessential ingredient in the substrate, (c) the actionof ACTH (which in turn could be either regulatory,or a standing condition necessary for some meta-bolic step under in vizvo conditions, or (d) a non-specific effect.
In vivo methods are much to be preferred.Adrenal vein blood has been obtained under condi-tions of acute surgery, or recent surgical inter-vention. 90-97 22, 98-101 The direct effect of theseprocedures is to increase aldosterone output wellabove basal rate and if, in addition, amounts ofblood are withdrawn which are large in proportionto the animal's circulating blood volume,'02 thesecretion of aldosterone will approach the maxi-mum adrenal output. Hence, the main factdemonstrable in the latter instance is that anexperimental procedure has reduced aldosteronesecretion through interference with pathwayswhich subserve the stimulus (trauma plus grossblood loss). However, the result may be difficult tointerpret, as the blood loss may cause also pro-gressive reduction of adrenal blood flow which, ifsevere, might limit aldosterone production. InFarrell's experiments on selective destruction ofareas of the C.N.S. of cats,ll9 there is a suggestiverelation between adrenal blood flow and aldo-sterone output. In the experiments on decerebra-tion and bioassay of extracts the adrenal blood flowhas not been reported.28Hume and Nelson24 developed a method of
chronic cannulation of the adrenolumbar veinin the dog which permits serial collection of bloodat a sufficient interval after surgery to allow theassumption that the immediate effects have dis-appeared. Howvever, they report values of 17-hydroxy cor' icosteroid output which are sporadi-cally as high as with ACTH infusion, and thisraises the question whether the animal is invariablyundisturbed by the procedure of pulling the intra-
abdominal ' choker' which occludes the vebetween the adrenal and the vena cava. Weaand Eik-Nes,25 using Teflon prosthesis, repsurgical preparations which permit access toadrenal vein in conscious animals. Howevthere is in this preparation no exclusive accessthe arterial supply of the adrenal gland, andsamples obtained do not permit estimatesadrenal secretion rates, as the rate of blood flcannot be measured.Another interesting approach to the probl
has been made by Masonil03 who has been ableinsert a No. 7 or No. 8 catheter into the riadrenal vein in 9 out of 20 conscious humsubjects.
In I939, Levy and Blalock104 made subcutanetransplants of the intact adrenal gland intoneck of the dog. These animals survived for loperiods and several had normal litters. From tone could conclude that the transplanted glsecreted normally, even under ' stress.' Thtransplants, however, allowed limited access oto the venous outflow from the gland, and no accto the arterial inflow.To meet the desiderata outlined above and,
particular to determine whether the adrenalstimulated directly by the local changes of iocomposition of arterial blood, an adrenal transplpreparation has been developed in this laboratThe left adrenal gland of the Merino sheeptransplanted to a combined carotid artery-jugvein skin loop in the neck.20 The left adreusually receives a small artery from the renal artand the adrenal vein drains into the left renal veThe renal vessels are anastomosed end-to-swith the carotid and jugular vein. The kidis discarded. Subsequently the right adregland is removed. An animal with an adretransplant has the same time relations of paroresponse to changes of Na+ balance as one wiboth adrenals in their normal position. Whepneumatic cuff applied to the vascular loop crato the transplant is inflated to 300 mm. Hg.,carotid artery supplies essentially only the adreand simultaneous application caudal to the adreof a cuff inflated to 30 mm. Hg. contrives thacannula in the jugular vein drains only adrevein blood. Thus the composition of adrearterial blood may be locally altered and the adresteroid output can be directly measured inadrenal venous blood of the conscious undisturanimal.
Methods Depending on the Concentration ofAldosterone in Peripheral Blood
Chromatography. Until the advent of raactive isotope methods,64 measurement of alsterone in peripheral blood has not been a pract
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
Wruary 1960 COGHLAN, DENTON, GODING( and WRIGHT: Control of Aldosteronle Secretiotn 85dure. With the Kliman and Petersonod2l aldosterone may be estimated in 50 to
ml. of peripheral plasma.23 Peterson reportsal levels of 0o.o04 to o.o8 y(/Ioo ml.; o.o8 to
I5 y/'oo ml. in patients with cirrhosis on lowa+ diet. The biological half-time of aldosterone
0o.5 to o.8 hr. for patients with Addison'sase, and normal men on low Na+ intake, and
3 hr. for patients with liver cirrhosis. Thermal 'turnover rate' was 330 to 400 y/day,easing to 56o y/day on low Na+diet, and was
ooo y/day for cirrhotic patients on low Na- diet.
Estimation by biological effects. Large changesplasma Na--/K- ratio may indicate deficiencyexcess of electrolyte-active adrenal secretion.nn discovered that DOCA reduced the Na/'K-'
tio in human sweat, and assumed that the loweat Na+/,K-' ratio observed during acclimatiza-n to heat was due to increased production ofectrolyte-active hormone by the adrenal gland.rawley and Thorn56 showed that DOCA reducedeNa-'/K-- ratio in human mixed saliva. Simpsond Taitl05 and August, Nelson and Thorn1owed that aldosterone caused a lowering ofman salivary Na 'K-- ratio. Observations oneat and saliva have given information onripheral aldosterone levels under varying physio-gical conditions and in disease states and, indeed,re largely responsible for Conn diagnozingnfidently his early cases of primary hyperaldo-eronism.However, I'horn et al.34 stated they found anence of correlation betNween aldosterone excre-n and salivary Na' +K' ratio in man. Theults show a large fluctuation in day to daydosterone excretion, as against a smooth curvelating salivary Na ,'K- ratio to degree of Na*pletion. It is open to question which methodassessing change of aldosterone secretion is theore reliable. The acute effects of ACTH,dosterone, sodium and potassium on parotidretion were studied in the dog by Langleyal.106 Intravenous injection of ACTH (8o U.)d aldosterone (75 y,) caused the pilocarpineulated salivary Na-'/K- ratio to rise. ACTH
used the secretion rate to rise, and aldosteroneused it to fall slightly. The ACTH effect mayve been associated with the change in salivarycretion rate. The effect of aldosterone found
not large, and apparently the observationsere made 60 minutes after injection. Ipsilateraltracarotid infusion of steroids in this laboratorynever had an effect on salivary Na-;'K - ratio
der 60 min., and the full response may belayed for as long as 240 minutes.107 A similare delay in the effect of aldosterone on urinarymposition upon infusion into the renal arterv
was reported by Barger et al.'08 Accordingly theresults of Langley et al. have no significance.The problem in using the salivary Na-J/K+ in
man is that it is a mixed secretion while concentra-tions of Na. and K- are not large and thereforethe variation in composition in response to aldo-sterone is small. Further, the Na+ concentrationvaries with secretion rate. These objections donot hold in the case of measurement of the parotidsalivary Na+/K- ratio in the ruminant.
Salivary secretion in a ruminant like the sheepis continuous, and 4 to io 1. of a simple solution ofinorganic ions are produced by the parotid glandseach day. The salivary Na+/K' ratio is normallyabout I80'6 30, and as a result of aldosteroneadministration or Na' depletion it may fall aslow as 20/I60 - O.I2. If a permanent unilateralparotid fistula is made, it is easy to collect serialsamples of adequate volume for assay by flamephotometry, and the scale of change induced isvery large relative to any analytical error involvedin photometry. The evidence justifying use ofthe parotid salivary Na-, K ratio as an index ofaldosterone secretion follows:
I. There is no parotid response to Na- de-ficiency if a sheep is adrenalectomized and isadrenally insufficient, and very little response if itis given only sufficient DOCA to maintain it whenin normal Na balance. A normal parotidresponse does occur if the sheep is given a four-eight fold increase of DOCA dosage.l7
2. Bilateral adrenalectomy of an anaesthetizedNa -- depleted sheep causes the salivary Na-/'K4-ratio to rise to normal after a latent period of90 to I20 min."
3. Physiological adrenalectomy. I1n a Nadeficient sheep with an adrenal transplant thedrainage of the total adrenal Xvenous effluentcauses the salivary Na-,iK-- ratio to rise to normalafter a latent period of go to 120 min. The re-injection of the adrenal v-enous effluent into theipsilateral carotid artery loop or intravenouslycauses the salivary Na '-- to decrease to the levelholding before the physiological adrenal-ectomy.20. 14The evidence that the effective component of
the adrenal secretion is aldosterone is:(i) McDonald and Reich'o"09 have shown that in
normal Na- balance the transplanted adrenalgland secretes 40o to 600 -, hr. of cortisol and smallamounts of corticosterone and cortisone. With thischromatographic method aldosterone was notdetected in 75 ml. of adrenal vein plasma. InNa deficiency aldosterone was always detected,being secreted in rates varying from 6 to 24 -'hr.In moderate Na- deficiency there was no changeof cortisol or corticosterone secretion, but in grossdeficiency the rate of secretion of both was
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
86 POSTGRADUAT'rE .MEDICAL JOURNAL Februlary i961moderately increased. Cortisol ranged from330 y/hr. to 1.2 mg./hr. and corticosterone from5 -f/hr. to 73 y/hr. The maximum values forcortisol and corticosterone were about one-thirdof those found during ACTH stimulation. Thusthe significant consistent difference betweennormal Na' balance and Na- deficiency was thedetection of aldosterone, and the results indicatethat the salivary response was due to this steroidor some undetected steroid. In their studies onsteroid output in the adrenal -ein blood of dogsRosnagle and Farrell1l° found that Na? deficiencydoubled aldosterone secretion, but did not affectcortisol. In man, Crabbe, Reddy, Ross and Thorn"'have shown that Na-- deprivation causes a con-sistent rise in urinary aldosterone secretion andhas no effect on urinary cortisol secretion. In sofar as urinary excretion of these compounds maybe taken as reflecting adrenal secretion, the resultsindicate that the physiological adaptations to Na+deficiency in man attributable to electrolyte-activeadrenal secretion are caused by increased aldo-sterone secretion and are unrelated to cortisolsecretion.
(ii) Intravenous infusion of authentic aldo-sterone in Na---deficient adrenally-insufficientsheep at rates corresponding to the range detectedby chromatographic procedures causes a com-mensurate decrease in salivary Nab-:'K-- ratio tothe extent observed in Na-- deficiency (Fig. 4).Intravenous infusion of cortisol at rates io timesgreater than secretion rate detected by chromato-graphic analyses of adrenal venous blood duringNa+ deficiency has no effect on salivary Na4 /'K-ratio (Fig. 5).This was examined also by observingthe effect of intravenous infusion of these steroidsduring rapid correction of Na deficiency bysystemic administration of 260m\IM. of NaC1.It was found that infusion of aldosterone at20 , 'hr. blocked completely the usual large rapidrise of salivary Na-'-'K- ratio which occurs 120min. after commencing correction of Na' de-ficiency. Aldosterone at I 0 y,/hr. delayed andreduced the effect, whereas cortisol infusion atI2 mg./ihr. caused little, if any, reduction ofresponse. A factor contributing to these large dif-ferences in electrolyte activity, which indicate thatit is unlikely that cortisol has any causal role in thesalivary response to Na?- deficiency, is that cortisolis 80 to 95 per cent. bound to protein in the peri-pheral plasma, whereas aldosterone is not sig-nificantly bound."2
In a sheep in normal Nat balance large fluctua-tions in secretion rate have very little influence onsalivary Na+,/K+ ratio, irrespective of whether thechanges follow psychic stimuli or rumination orwhether they are contrived by reduction of parotidblood flow.16 In a Na~--depleted sheep there is a
highly significant relation between parotid salivNa-/K- ratio and secretion rate. This effect hto be taken into account when using the salivaNa-'-K' ratio as an index of aldosterone secretirate. This can be done satisfactorily in the mtype of experimental context we have used fexamining the control of adrenal secretionaldosterone, viz. the adrenal response to rapicorrection of Na- deficiency in a depletanimal. 11. 17, 14. 15 Prior to the experimentprocedure there is a control period of 200 to 3minutes during which the relation between secrtion rate and salivary Na-/K+ ratio is determineWhen the salivary Na-/K= ratio or salivary Kor Na concentration is graphed against an aritmetic ordinate scale there is a highly significlinear correlation with secretion rate. During texperiment (e.g. rapid correction of Na+ deficienor local alteration of the ionic compositionadrenal arterial blood) the position of each poirelative to this line is determined. A sustaindeviation from the control period line during tperiod of experiment is interpreted as reflectingchange of aldosterone secretion. In the case of tsalivarv Na -'K:- ratio, a rise above the line reflecan inhibition of secretion; a fall indicates stimultion. The results are graphed as parotid salivaNa+/'K+ ratio ' corrected for secretion rate varition' and for reasons set out elsewhere'4 the Kconcentration ' corrected for secretion rate varition' is included also on the figure (Fig. I8Whereas changes of Na- balance do not alsignificantly the sensitivity of the biological idicator over the time scale of this type of expement, the chronic effect of such changes on tsensitivity of the parotid to aldosterone have totaken into account in other types of experimenFor example, a standard dose of aldosterone caua much larger decrease of salivary Na±/K` ratin a Na --depleted adrenalectomized sheep thandoes in a sheep in normal Na- balance (Figs.and I2). Reference to these figures indicates ththe parotid salivary Na '/K-L ratio gives a reliaspeedy reproducible index of the peripheral aldsterone level. The evidence from the resultsstopping, starting and altering the rate of intrvenous infusion of authentic aldosterone indicatthat, provided due allowance is made for time 1of biological action, the salivary Na+/K+ rareflects commensurately changes of adrenal aldsterone secretion rate (Fig. I2). This would appdirectly in experiments where the only chaninduced is local alteration in the ionic environmeof the adrenal gland, but may require soquantitative qualification in short-term expements where the volume over which aldosteronedistributed is rapidly changed.
Muscle biopsy. The Nal/K - ratio or the N
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
Wuary g196o C(OGH)(;I.AN, DEN'I'()N, (;()DING and \RIGHT: Control of .Aldosterone S'ecretioi S7
K- concentrations in mtuscle have beeii usedindex of aldosterone levels in peripheralThe results of Woodbury and Koch34
been mentioned previously. Gornall"l foundased aldosterone excretion was associated within Na (E.C.F.) Na(I.C.F.) and a fall of K
.F.)/K(I.C.F.) in w-omen during the thirdester of pregnancyr. However, there is someculty in reconciling these findings with thosenn's syndrome, w-here measurement of in-lular electrolytes show-ed a rise in Na- and ain K-+ 3. 113. 11t 'There are at least tw-oible causes of this difference:During the course of a prolonged surgicaldure, large electrol-te shifts occur K isfrom cells and Na - moves in.117 Hence it isble that biopsy specimens be taken at theopportunity during an operation.) If the rate of aldosterone secretion has beenfor prolonged periods, urinary K loss causes
deficit."'
ds Depending oin Excretion oJ Aldosteronene. Nearly all the recent work publishedhecontrol of aldosterone secretion in man, hasnded on the estimation of free aldosterone in. specimens of urine. This has usually beened out by chromatography, but some workers,bly Llaurado, have used bioassay procedures
well. The validity of the assumption thatges in urinary aldosterone output reflectges in the rate of aldosterone secretion hasdiscussed previousl ' 14 and will not be
ated in detail here. The main points are (i)a small proportion of secreted aldosterone,4 to 8 per cent, is detectable in the urine,
small changes in renal tubular reabsorption ofterone would lead to large changes in aldo-ne excretion, (iii) different methods of hydro-of conjugates yield different results. In orderparate aldosterone from interfering substances,tlosses are incurred which must affect theility of the method. This applies parti-ly when it is intended to use bioassay pro-res to estimate aldosterone which hasseparated by chromatographic means.34
nfurth and Venning69 consider that fourmatographic systems are necessary for separa-of aldosterone from human urine. Duncan,le and Bartter120 have advanced reasons foring that change of urinary aldosteronetion reflects change of plasma concentration.may be legitimnate under many conditions butt be questionable when there are acute altera-of renal haemodynamics or when tubular
tion changes are induced by pitressin orcurial diuretics.
n an important paper by Ulick, Laragh and
Lieberman:' a inew method is presented whichovercomes some objections to urinary steroidestimations. The method is based on the dis-covery of a urinary metabolite (Fig. 3, C) of aldo-sterone43 Nwhich occurs in larger amounts than freealdosterone. 'rThe method depends on the deter-mination of the specific activity of this tetrahydrometabolite-a measure which indicates the dilu-tion of an intravenous dose of I y of tritiumlabelled aldosterone. The validity of the estimateof daily secretion rate depends on the followingassumptions, (i) the labelled aldosterone mixesrapidlv with the unlabelled aldosterone in thebody, (ii) the subsequent fate of both forms ofaldosterone is the same, (iii) the pool size of aldo-sterone remains constant during the study, i.e. rateof disappearance is equal to the rate of secretion,(iv) the proportion of aldosterone converted to themetabolite remains unchanged during the study,(v) the metabolite is not produced by the degrada-tion of any other steroid. Variations in the rate ofhepatic inactivation or of renal clearance do noteffect the determination, unless they result inalterations of adrenal secretion rate. This promisesto be a very useful me.thod for clinical studies, andfor some experimental work. It does not appearsuitable, in its present form, for observation ofchanges of adrenal secretion rate over a timecoursewhich is much less than 24 hr.
Faeces. Orti, Ralli et al.76 have reportedexperimental results depending on estimation ofaldosterone in the faeces of rats. This was doneon the basis that the main route of excretion in thisspecies wvas via the faeces.
secretion in man and in all mammals investigatedto date. In man the ability to respond to Na+'depletion by increased urinary aldosterone excre-tion has been used as a test of normal aldosteroneregulation.l21
Replacement of SNa in the deficient animal orman decreases aldosterone secretion. With Na+deficient sheep, the rapid inhibition of adrenalaldosterone secretion which follows a systemicdose of Na-- has provided an excellent physio-logical test system for the analysis of control ofadrenal secretion. The important feature is thatthe ' inbuilt' biological indicator, the salivaryNa*/!K- ratio, exhibits a large quantitative changeover a short time scale, a situation analogous tothe inhibition of an established water diuresis inthe experimental animal which was used byVerneyl° to estimate the anti-diuretic hormonereleased by intracarotid infusion.
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
In sheep, Na- deficiency due to loss of parotidsaliva usually causes a fall of plasma Na+ concentra-tion. For example, 20 specimens of blood drawnfrom P.F.I under control conditions with adequateNa' intake had a plasma Na+ concentration of148 + i.6 (S.D.) mEq/l. In eight externalelectrolyte balance studies in which free access towater was permitted and in which Na+ deficit of500 to 90o mEq. occurred, the Na+ concentrationof 4I specimens of blood was I43 + 3.I (S.D.)mEq./l. The Na-+ concentration of ten specimensdrawn during the period 24 to 72 hr. following thereplacement of Na-' supplement was I50 + 3.I(S.D.) mEq/l. The plasma Na:- concentrationof I5 specimens taken from P.F.I3, P.F.i7 andP.F.33, whilst on adequate Na+ supplement was'47 + 3-7 (S.D.) mEq/l., of 21 specimens takenduring Na- depletion 140 + 5.8 (S.D.) mEq/l.and of ten specimens taken during the first 72hr. of replacement of Nat- supplement I52 ± 3.I(S.D.) mEq 1. In gross Na- deficiency (700to I,ooo mEq.) the plasma Na+ may decrease to120 mEq/l. or less. There is usually a rise ofplasma K-'- concentration in Na+ deficiency. Themean plasma K' concentration of 25 specimens ofarterial blood taken from P.F.I3, P.F.I6 andP.F.33 whilst in normal Na+ balance was 4.4 + 0.4(S.D.) mEq/l. Eleven specimens taken fromthese sheep during Na-- deficiency had a K*concentration of 5.6 -1 o.8 (S.D.) mEq/1.Many other changes occur, e.g. decrease in pH,
plasma HC03-, intracellular Na2- concentration,plasma and extracellular volume, blood pressureand presumably right arterial pressure, cardiac out-put and total body Xwater. There is a rise inhaematocrit and blood viscosity.When Na deficiency is wholly or party corrected
rapidly by the standard procedure of systemicinfusion of 60 to 70 ml. of 4 M NaCl during aperiod of 40 to 50 min., there is a 15 to 30 mEq/l.rise of plasma Na concentration, a fall of 0.5 to 2.0mEqil. of K' concentration, as well as a sub-stantial reversal towards normal of the deviationof haemodynamic conditions caused by Na'deficiency.
In considering the increased aldosterone secre-tion resulting from Na depletion, the initialquestion is ,whether this is due to decrease ofplasma Na4'- concentration. Though there is usuallya fall as indicated above, in the course of a singlebalance experiment the plasma Na - concentrationand salivary Na+-'K ratio often do not show acorrespondence suggestive of a commensuratecause and effect relation. The fall of salivary ratiois progressive, whereas the plasma Na* mayfluctuate considerably, a finding reflecting con-current operation of the osmo-regulating mechan-ism. The significant fall of plasma Na-;- in severe
20. waterdepleted
l0
Parotid 5g g'~ d
Salivary2,
Ratio0'5
250.ParotidSalivary
200 .,'
Concentration
waterdepletedr---X
152 /
148 X
Na+144.1440
Period 595 595IWithout, ....-NaHCO3 y da'yNa+ me'v/-600.
Balance¢-200.
+200. S'-5 0 5 10 15 20
days
FIIG. 7.-P.F. I. Comiparison of the effect on (iparotid salivary Na~' K- ratio, (ii) parotisalivary Na -rt K' concentration, (iii) plasmNa- concentration, (iv) external \Na'- balanof a standard episode of Na- depletio(o o) with one in which wuater intake vseverely restricted during 'he first 7 davysNa4 depletion (X - X). Na- depletiocommenced on day- o in both instances, anwas produced by withholding the daily Nasupplement of 595 m. equiv. of NaHCO0On the morning of the 7th day the animal wpermitted free access to wvater and drank 3.litres during the first 5 minutes.
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
puary I96o COGHLAN, DENTON, GODING and WRIGHT: Control of .ldosterone Secretion 89
JACK ,87se/(No dp/tW)
.10
140Plasma S
-8 Na'
O4 20pattributableto secreti on rate) of intra
infusion of 2 1. of I02 n.equiv./l. NaCI -204m.equiv. of Nad. The effectonplasmaNarcon-
a ltiontravenou
centration, and the dosage of pitressin emplovedto prevent a water diuresis is shown also.
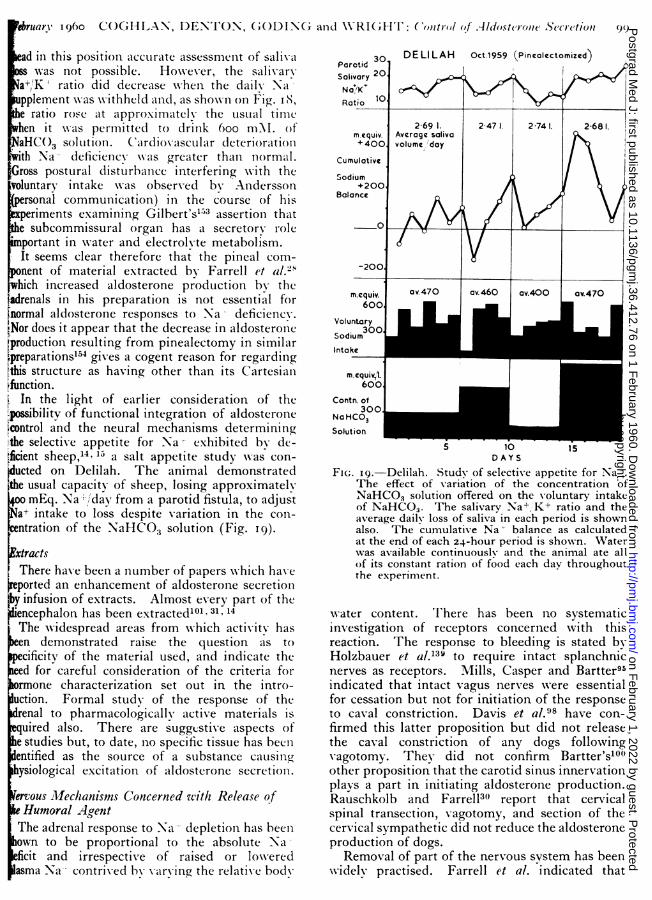
fciency indicates' accommodationi ' of the osmo-.eptors, a process which is possibly dependenttheir partaking in the changes of intracellularmposition occurring in this state..The following experiments are pertinent to thistes2on:Jack,sheepwas cocurretldepleted ofbothparotidswater, so thK rating Na- deficiency therewasattriseof plasma Nato s concentration instead of theual fall. Nevertheless, the salivarv Na,-/Kio fell, indicating increased aldosterone secre-n.Fig. 7shoons this fallof salivaryNaCa20Km.equiv,theriseffectofplasmaNa'concentration during+ deficiency, and the factthat a largessin empoluntaryciake of indicater(3 1.) permitted on the osmorningthe seventhday, caused a large fa llof plasmapo+concentration,but little effectontheissaliary+/K-rtion .
In other expesothat duriments Na deficiency theas cor-rised by the infusia concentrationic solution(2 1.
2 mMl. NaCIevertheless,)and simultaeously theto-regulal tingsystem as prevented fromtsing a rise of plasma Na- concentration byministration of pitressin. Though in this ex-+ deficiency, and thefsalivaryNaK ratiooccurredI8e of water (1in.insteadoftheusual 120min., it isar that adrenal secretion of aldosterone wasbited, althoughno ris ge f of plasmaNaco0-
ntration occurred (Fig. 8).In examining the possible role of increasedsmaK+concentratittle effect onin causithegaldosteroneretion sheep NAere given an artificial diet withnrylow K+ content. Na deficiency(600 to 700q.)causedby the salivary NaKhypotonic ratioto fall,ugh therewas a fallofplasma K -concentration byough there was a fall of plasma K* concentration
instead of the usual rise.122 In these experimentsthere Xwas also a rise of the plasma Na+/K+ ratioinstead of the usual fall. The extent of decrease ofsalivary Na-/K- ratio was not always as great aswith uncomplicated Na- deficiency, and in-vestigation is proceeding to determine whetherthis results from a circumstance peculiar to thisexperiment, i.e. deficiency of K- supply to thebiological indicator, or from a smaller adrenalresponse than usual.
In the dog high K-- intake, if it leads to in-creased plasma K- concentration, causes a rise inaldosterone excretion.123 In man, if the dietary Nais reduced whilst on low K- intake, the rise inaldosterone excretion may not be as great as thatoccurring with low Na- intake alone.l24 120 Singerand Stack-Dunne94 have shown that K- deficiencyreduces the amount of aldosterone detected inadrenal vein blood relative to control specimenscollected under anaesthesia.
Observation of the blood pH and the plasmaHCO, concentration during the course of changesin Na-- balance gives little reason to suppose thatthere is causal connection with the adrenalresponse,122 and it is a common finding thatinfusion of 4 NI NaCI, which inhibits adrenalsecretion in Na- deficiency, does not produceany change in pH or plasma HCOa over the timeduring which a full parotid response occurs.These results as they stand, particularly as far
as plasma Na'- and K- concentration changes areconcerned, might appear to suggest that thesefactors had little or no influence on adrenal secre-tion of aldosterone. They could be taken asevidence in favour of a hypothesis that change ofvolume of the extracellular or intravascular com-partments or some consequence of it causes aldo-sterone secretion in Na-- deficiency and in ainumber of other clinical conditions. Before con-sidering the logical basis of this argument we wishto cite some further experiments.
In sheep, when adrenal secretion is inhibited bythe standard procedure for correcting Na-deficiency, some 6o to 70 ml. of 4 M NaCl aregiven. It is most unlikely that the effect producedis attributable to change of total body water and,in any case, there is no effect if the same volume of2.7 MVI glucose is given.1' Hence, if the effect wereattributable to volume changes, it would depend onredistribution of fluid between compartments, thehypertonic Na- solution causing an expansion ofextracellular and plasma volumes. If this is thecausal mechanism, it is surprising that an equalvolume of 2.7 MI glucose has no effect, since,because of slo,w entry into cells, its osmotic effectw-ould be quite large over the I20 min. followingcommencement of infusion. Gross water deficiencyvcausing considerable reduction of plasma volume
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
POSTGRADUATE \IEDICAL JOURNAL. February 1 91PF 33 9-10,'7"59
+16
*14Corrcted
Porot,d.10
Saivaory + 8
No'K' 6 Drank Drcnk2'421 2 /
.+4 NaHCO)+2
H, 0 525 mequ,01-2
wm qunfq 155Plasma s N Plasma
30N' HCO;145 3
fonctntn. HCO,x .... ..........- .. ' concentn
208050,'Whol3
blood, 1 45 1-030. 34
ox -Hmat
2 4 6 e lo 12 14 16 18
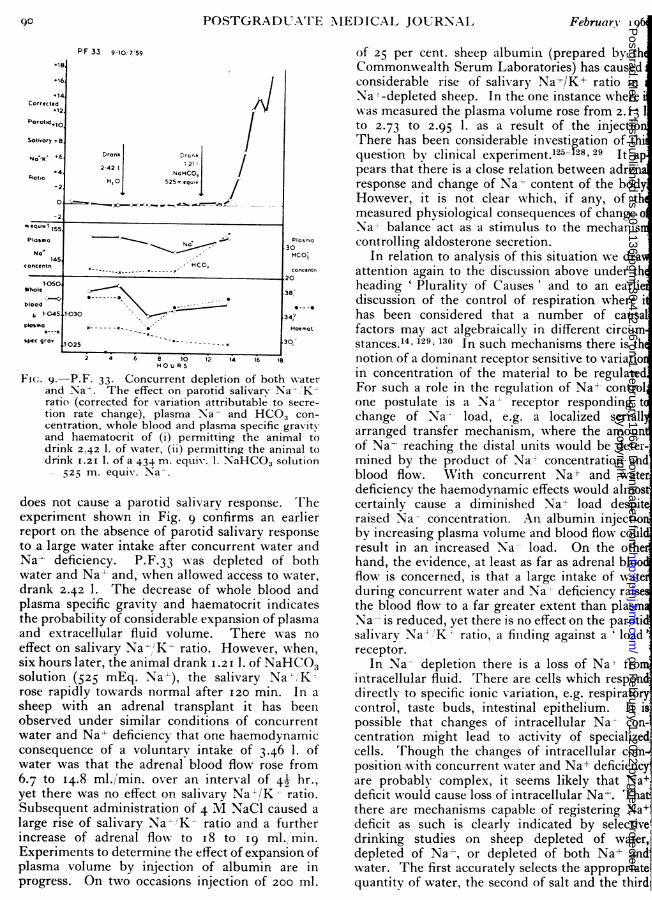
F(c;. 9.-P.F. 33. Concurrent depletion of both w-aterand Na-. The effect on parotid salivary Na':;K-ratio (corrected for variation attributable to secre-tion rate change), plasma Na- and HCO3 con-centration, whole blood and plasma specific gravityand haematocrit of (i) permitting the animal todrink 2.42 1. of water, (ii) permitting the animal todrink I.2I 1. of a 434 m. equivl.l. NaHCO3 solution
525 m. equiv. Na'.
does not cause a parotid salivary response. Theexperiment shown in Fig. 9 confirms an earlierreport on the absence of parotid salivary responseto a large water intake after concurrent water andNa- deficiency. P.F.33 was depleted of bothwater and Na-' and, when allowed access to water,drank 2.42 1. The decrease of whole blood andplasma specific gravity and haematocrit indicatesthe probability of considerable expansion of plasmaand extracellular fluid volume. There was noeffect on salivary Na-' K- ratio. However, when,six hours later, the animal drank 1.2iI. of NaHCO3solution (525 mEq. Na-l), the salivary Na-';Krose rapidly towards normal after I20 min. In asheep with an adrenal transplant it has beenobserved under similar conditions of concurrentwater and Na4- deficiency that one haemodynamicconsequence of a voluntary intake of 3.46 1. ofwater was that the adrenal blood flow rose from6.7 to I4.8 ml./min. over an interval of 49 hr.,yet there was no effect on salivary Na-l/K ratio.Subsequent administration of 4 M NaCl caused alarge rise of salivary Na- KX ratio and a furtherincrease of adrenal flow to I8 to 19 ml.,'min.Experiments to determine the effect of expansion ofplasma volume by injection of albumin are inprogress. On two occasions injection of 200 ml.
of 25 per cent. sheep albumin (prepared by thCommonwealth Serum Laboratories) has causedconsiderable rise of salivary Na<+/K+ ratio inNa-'-depleted sheep. In the one instance wherewas measured the plasma volume rose from 2. 3 1to 2.73 to 2.95 1. as a result of the injectioThere has been considerable investigation of thiquestion by clinical experiment.1l25-28, 29 It apears that there is a close relation between adrenresponse and change of Na- content of the bodyHowever, it is not clear which, if any, of thmeasured physiological consequences of change oNa-' balance act as a stimulus to the mechaniscontrolling aldosterone secretion.
In relation to analysis of this situation we draattention again to the discussion above under thheading ' Plurality of Causes' and to an earliediscussion of the control of respiration where ihas been considered that a number of causalfactors may act algebraically in different circumstances. 4, 129.130 In such mechanisms there is thnotion of a dominant receptor sensitive to variatioin concentration of the material to be regulatedFor such a role in the regulation of Na+- controlone postulate is a Na+ receptor responding tchange of Na- load, e.g. a localized seriallarranged transfer mechanism, where the amounof Na- reaching the distal units would be deter-Imined by the product of Na+ concentration and,blood flow. With concurrent Na+ and waterdeficiency the haemodynamic effects would almostcertainly cause a diminished Na+ load despit'raised Na- concentration. An albumin injectioniby increasing plasma volume and blood flow couldresult in an increased Na load. On the othedhand, the evidence, at least as far as adrenal bloocflow is concerned, is that a large intake of wateduring concurrent water and Na+- deficiency raisethe blood flow to a far greater extent than plasmNa-- is reduced, yet there is no effect on the parotisalivary NaL K ratio, a finding against a 'loadreceptor.
In Na- depletion there is a loss of Na' fronmintracellular fluid. There are cells which responddirectlyN to specific ionic variation, e.g. respiratorcontrol, taste buds, intestinal epithelium. It is,possible that changes of intracellular Na' con-centration might lead to activity of specialized,cells. Though the changes of intracellular com-position with concurrent water and Na+ deficiencyare probably complex, it seems likely that Naideficit would cause loss of intracellular Na+. That-there are mechanisms capable of registering Na+deficit as such is clearly indicated by selectivedrinking studies on sheep depleted of water,depleted of Na+, or depleted of both Nal and1water. The first accurately selects the appropriatelquantity of water, the second of salt and the thirdt
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
uar3, I960 (COGHI,AN, I)ENTON, GODING and WRIGHT'I': C'ontrol of .4ldosteronle Secretionl 91
th water and salt.'4 There may be dominanttors for each deficit which may act separatel-gether, but the accuracy of ingestion clearl-
cates that centres with ' memory' and ' com-g' capacity w-ould operate from such
ysers. The aldosterone response is commen-e and continuous in relation to the absoluteum deficit and in these respects is somew-hatrent from the ADH response, which is inrtion to the saltw -ater ratio and whichmodates.
further possibility not necessarily excluded byexistence of others is that the responsive tissuetlocalized and that a metabolic consequence ofletion of a Na reservoir, such as bone, causesrelease a chemical agent capable of specifically
:ting the adrenal glomerulosa.nsideration of the difficulties of analysis lede view that a decisive discrimination Nwould
it from devising a method permitting access toarterial supply of the adrenal gland in theious animal. The primary object would be
determine whether or not local changes in thecomposition of the arterial blood influencedterone secretion. Also critical stipulations
cerned in the establishment of a physiologicalfor a putative hormone might be met. Theonstration that a sheep with its sole adrenald transplanted to the neck has the sameinses to alteration of Na balance as a normalal has shown that the adrenal nerve supply isessential for this physiological response.uring the past three years an extensive series ofriments has been made in this laboratory oneffect of local alterations of the ionic environ-t of the adrenal gland.'14 15 A full report will belished shortly. A summary of some aspectsws.our main types of experiments have been made:Local reduction of Na concentration and
ease of K-- concentration of adrenal arterialAn example of the main result of this
is is shown on Fig. io. Transplant 9 was innal Na balance: the salivary secretion rated between I and 4 ml./min. To ensure that
effect could be attributable to Na deficiencyrvening in the course of the experiment theal was given a large intake of Na- via rumenduring the experiment.
) Decreasing the Na - concentration of adrenalial blood by 5 per cent. glucose infusion orion of 4 per cent. glucose with io mM.'1.1 added did not affect the salivary Na ,'K-
i) Increasing the K- concentration had little,y, effect on the salivary Na+/K+ ratio.
ii) The concurrent decrease of Na+ concentra-and increa:;e of K-- concentration of adrenal
T P 9 Normal Sodium Balance100
Parotid CO0°x
Salivary CO x C ? _ A
20N a/K
10Ratio
300m.equiv.Nc lOOmequiv. Na 100m.equiv. N2 into rumen into rumen into rumen
O. -mI/minRate ot 1 - AA
Infusion into ·2
Adrenal Arterial
Blood Supply 0
2
1 00
-200 0 +200 +400 +600MINUTES
Fi(;. o.--'l'ransplant 9. Normal Na- balance. 'Theeffect on parotid salivary Na' K ratio ofinfusion into the adrenal arterial supply at 0.2-I.8 ml. minm. of the following solutions:--(i) 5,, glucose, (ii) 40o glucose with to nm.equiv. i. NaCi added, (iii) I41 m. equiv. l. NaCI-with I5 m. equiv. 1. ICI added, (iv) 4`0 glucosewith I3.2 m. equiv.l. KCI added. In eachexperiment the animal Nwas given the dosesof NaHCO:, at the time shown to eliminateany possibility of Na depletion occurringduring the course of the experiment.
arterial blood had an unequivocal effect on salivaryNa' K- ratio wNithin I20 minutes of commencingthe intra-arterial infusion. The ratio fell to therange of 4 to 5. A second feature of the particularexperiment shoivlnon this figure is that the ratiorose again upon halving the rate of infusion. Fig.I i records similar experiments. In this instance(middle section) the ratio commenced to fall 120min. after beginning the intra-adrenal infusion,and rose 6o to 70 min. after stopping it. Theeffect may be compared with that caused undersimilar experimental conditions by the intravenousinfusion of 24 hr. of authentic aldosterone. Inthe lower section of Fig. i I an experiment isshown in which the parotid response has occurredmore rapidly. The effect produced by halvingthe rate of adrenal arterial infusion is very in-teresting, and is suggestive of an ' undamped'system.
Fig. 12 is of considerable importance in that itshows the dose-response relation under these
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
20 12/6/59(normalParotid Ni balance.)SalivaryNia/K' 5
Ratio
-100 0 100 200 300 400 500 600 700MINUTES
FIG. I I.-Transplant 9. Normal Na- balance. Theeffect on parotid salivary Na /K'- ratio of(i) intravenous infusion of unesterified d,l,aldos-terone at a rate equivalent to 24 ,tg.,/hr. of thedextro isomer form, (ii) infusion into the adrenalarterial blood supply of 4'0 glucose with 9 m.equiv./l. KC1 added at 0.45-1.8ml./'min.,(iii) infusion into the adrenal arterial blood supplyof 4%0 glucose with io m. equiv./l. KC1 added at0.5-2.0 ml. ,min.
TP9 NORMAL SODIUM BALANCE 14/i0/S9
100PAROTID
50 -;- -,', ' 0-
SALIVA20.
Nd/- -- - -
10 SALIV_RY K'CONCENTRATION /RATIO AT PLATEAU OF RESPONSE
S 0 - 4mgquiv./l5 5 -= 6 nmquiv/lI r " 145n quiv/I1651 = 2On qui.A
muiOOnuquiv. NaHC03sg/hr 20 by Rumen TubeCALDOSTEROINFUSIONRATE 0
5 10 52CXHOURS
F'I. I2.-Transplant 9. Normal Na-- balance. Theeffect on parotid salivary Na- 'K' ratio of intra-venous infusion of unesterified d,l,aldosterone atrates equivalent to 5.5, i, and I6.5 ptg. hr. of thedextro isomer form. The salivary- K concentrationat the plateau of response is shoNwn also. Theanimal -was given doses of NaHCO3 at the timesindicated to eliminate any possibility of Nadeficiency occurring during the course of theexperiment.
conditions of Na- balance when the rate of in-travenous infusion of authentic aldosterone Nwasincreased progressively.
Table i shows the results of tNwo of a series of
·o ocI
0o0 __ _
, O I00 _ __ -
IN1
0.I
o
I0 oo X
3'
0 0 0 '
2 IC s
-1-IIr
i
;-
I - . --
,C - "1-
z 0;U z -
, oV - I-° I.
!|,-- § ,I
0 -- .I
c .
O'E (j - rY I:
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
rbruary 196o CO(;HLAN, DENTON, GODINC; and WRIGHT: Conltrol of Aldosterone Secretion 93
eriments in which the effect of such infusionthe composition of adrenal arterial blood hasn measured by collection of the venous effluentm an indNwelling polyethylene cannula. Theange of plasma K-- concentration produced wasthe order observed often during Na:- deficiency.oreover, as shown on Fig. i i, infusion at half thete with only Io mrM\. 1. KC1 added to the fluideffective. The decrease of plasma Na - con-ntration produced Nwas large (2I to 24 mEq 1.)
ut it has been shown that the same salivaryponse follows if an infusion of 2 per cent.
lucose with 75 mMI. 1. NaCl + 15 mMl. 1. KC1given (cf. Fig. I). This would reduce Na+ncentration by approximately io to I2 mEq/l.The specificity of the stimulation of adrenalcretion constitutes control of a number ofnditions; e.g. the possibility that the effect isue to glucose, temperature changes, osmotic orcotic effects is largely eliminated in that thesee common to most of the procedures.2. Decrease of the Na - concentration and increase
f KT concentration during rapid correction ofNa -eincy. These experiments are complementary
those cited in Section I. The aim was torevent locally the rise of Na- concentration andecrease of K4- concentration of adrenal arteriallood which occurs when Na- deficiency is rapidlyrrected by systemic administration of 4 M NaCI.indicated earlier (see Fig. 4 and I2) the parotid
and in Na+ deficiency is more sensitive to aldo-erone, and in this context better quantitativefinition of the role of various ionic changes isssible. Confirming the results in Section I, it
as been sho,wn that:(i) Decreasing the Na- concentration of adrenalerial blood (5 per cent. glucose infusion) doest affect the parotid response to correction of Na--ficiency.(ii) Increasing the K- concentration of adrenalerial blood (infusion of 136 mM. NaCl with
5 mM. /1. KC1 added) caused reduction of therotid response.(iii) Concurrent decrease of the Na+ concentra-on and increase of K- concentration of adrenalerial blood (infusion of 4 per cent. glucose with
o to I5 mM./l. KC1 added) caused a large re-uction of parotid response, the effect being inter-ediate between that produced by intravenoususion of authentic aldosterone at io and 20
/hr.3. Local increase of the Nat concentration ofrenal arterial blood during NVa+ deficiency. Thea+ concentration was increased progressivelyver the usual time interval of correction of Na+ficiency by systemic administration of 4 MaCl (i.e. 40 to 60 min.) and then held at thislateau for I80 to 240 min. Increase of the plasma
Na-- concentration also causes a decrease of plasmaK- concentration (Table i). The results of over30 experiments in this series have shown that localincrease of Na- concentration of adrenal blood byIo to I5 mEq'l. has little if any effect on parotidsalivary Na+/'K ratio. Increase bv 20 to 25mEq/1. has a significant effect, but it is in no waycomparable to that occurring when the same changeof Na- concentration is caused by systemic dosageof Na-.
4. Local increase of ¥Va-t concentration of adrenalarterial blood as NVa-- deficiency commences. Tento I; hours after the last intake of Na-', theparotid salivary Na' /K- ratio of a fistulated sheepwhich is secreting saliva at Ioo to 200 ml. hr.begins to decrease. Experiments have been madein which the Na+ concentration of adrenal arterialblood has been increased at this time. The effecthas been to prevent the usual fall of salivaryNa+,lK ratio. When the infusion is stopped, theratio falls rapidly within 90 to I20 minutes. Noeffect occurs if the same infusion is given systemi-cally. The investigation is still in progress. Aparticularly interesting feature emerging is that ifthe adrenal infusion is begun just after thesalivary Na--lK--- ratio begins to fall the ratio risesto normal and remains normal whilst the infusioncontinues. If, however, the infusion is stoppedand the parotid salivary Na -/K- ratio falls, recom-mencement of the adrenal infusion some hourslater does not cause the ratio to rise again. Thisresult, consistent with the findings in Section 3,will, if confirmed in the necessary series of experi-ments, raise the interesting possibility that in-creased adrenal arterial Na+ concentration can pre-vent the action upon the adrenal of anotherhumoral factor at the onset of Na+ deficiency, butonce this factor has acted upon the adrenal for anadequate period, the effect cannot be reversed byincreased adrenal arterial Na+ concentration.
Summarizing, the results of the large series ofexperiments in Section 3, and those in Section 4,indicate formally by the method of difference thatsome factor or factors other than simple change ofNa-- and K+ concentration act upon the adrenalgland to cause the response to Na+ deficiency.As the transplanted gland is denervated, this factoris a humoral stimulus, and the evidence at presentis against its being ACTH. The demonstration ofadrenal response to concurrent decrease of plasmaNa+ and the increase of plasma K+ concentrationraises interesting questions. The extent of changenecessary for this effect, i.e. approximately ImEq/l. rise of K and io mEq/l. fall of Na+,is well within the range of change observed insevere Na+; deficiency. Investigations so far aresuggestive that there is a threshold of ionic changefor this effect. If this be so, it adds further weight
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
9+ POST(GRADUATE MEDICAL JOURNAL February I91to the importance of the observation of manyworkers that there may be an adrenal response toNa-- deficiency, though there is little or no changeof plasma ionic composition. On the other hand,the response to local ionic change is quantita-tively significant in terms of aldosterone equivalentand experiments are proceeding using the Klimanand Peterson assay to fulfil desideratum 7. It-ould seem to be of considerable clinical import-ance that the extent of concurrent deviation fromnormal of these two ions required to produce thiseffect is within the range of plasma changeobserved in patients when severe fluid loss resultsfrom diarrhoea, intestinal fistulae, intestinal suc-tion, burns, and in patients with shock, renalfailure, diabetic acidosis and other medical andsurgical emergencies. In these situations theplasma ionic changes could be an importantmechanism evoking maximal aldosterone secretion.
At this stage of the investigation of aldosteronecontrol it is of great interest also to consider theproblem in its general biological context, as thismight be provocative of experiments which throwlight on the mechanism of control in higheranimals. As Chester Jonesm32 has pointed out, ithas not yet been established at what point in thephylogenetic system adrenal secretion of aldo-sterone emerges. His recalculation of the data ofHartmann cited in his book, 'The AdrenalCortex,'34 is suggestive that adrenalectomv inanimals as primitive as elasmobranchs may causea rise in plasma K+ concentration. The character-istic effect of adrenalectomy (decrease of plasmaNa-/K-:- ratio) is seen in the toad. One mightpostulate that, at the earliest stage of emergenceof aldosterone control, the evoking stimulus wasdecrease of plasma Na and increase of plasma K-concentration acting directly on cortical tissuewhen these changes proceeded to an extent whichthreatened the characteristic differential of ioniccomposition between intracellular and extracellularfluid. With subsequent evolution a considerablymore comprehensive system of control emergedwhich operates long before circulatory deteriora-tion or other physiological effect causes severefunctional embarrassment to such vital character-istics. This added mechanism, emerging inassociation with the great development of thenervous system, was functionally integrated withparallel development in animals of the capacity tomanifest behaviour appropriate to the correction ofspecific chemical deficiencies.132 14. 15 The func-tional end-result in the higher mammalian speciesis a specific hormonal control of aldosteronesecretion14, 22, 29 and a specific and quantitativelyaccurate appetite for Na- salts. As a legacy ofearly phylogenetic development there remainsalso, as a second line mechanism, the capacity of
the adrenal cortex to respond directly to concurrealteration of plasma Na- and K4- concentratiowhen the changes proceed to the extent seensevere fluid loss, electrolyte depletion and ciculatory deterioration.
Changes of Intravascular or Extracellular VoluWe have considered some experimental aspe
of this problem in the previous section. A coprehensive presentation by Bartter and discussiof recent data was included in the I959 edition' Recent Progress in Hormone Research '100 andwrill not recapitulate here. Farrell's article' Physiological Reviews '29 summarizes the resultsmany of the experiments involving alterationbody fluid volume by administration of water apitressin.
Constriction of the Inferior I ena Cava as a Stimulto 4Aldosterone SecretionConstriction of the thoracic inferior vena ca
results in an increase of aldosterone secretion.5,22The following experiments throw some lig
on the physiological basis of this effect:(a) In an earlier study by Ball and Davis
the diameter of the abdominal vena cava was costricted by 25 to 50 per cent. immediately abovthe renal vessels in order to determine whethincreased adrenal and renal venous pressure wouhave the same effect. There was no changealdosterone secretion and it was inferred tvenous congestion of the liver was the causeincreased aldosterone secretion after caval costriction. Subsequently it was shown that if tinferior vena cava were constricted below the livto an extent to cause femoral venous pressurerise above 20 cm. of water increased aldosterosecretion occurred.97 The effect of thoracic cavconstriction was largely abolished if the animXwere encased in an abdominal plaster cast.136
(b) The infusion of dextran in amounts sufficieto maintain and increase circulating blood volfailed to prevent the rise of aldosterone secretiodue to caval constriction.97 On the other hanBartter'00 reports that infusion of very laramounts of blood will reverse the effect of caconstriction.
Apart from loss of fluid from the vascular copartment, a possible explanation of the effectthoracic caval constriction on aldosterone secretiois that the hepatic venous congestion might increase the rate of degradation of aldosterone anby reducing the peripheral feed-back on a regulatory centre, cause a stimulation of adrenal secretioAlternatively, the congested liver may releasesubstance which acts directly or indirectly upothe adrenal gland.
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
y I96o COGHLAN, DENTON, GODING and WRIGHT: Control of Aldosterone Secretion 95
tion of the Pulmonary Artery5 reported that experimental right-sided
ailure caused a six-fold increase in aldo-secretion. In this situation, right atrial
e is increased whereas with constriction ofracic vena cava it is decreased, an observa-ich has to be considered in the light of theof Bartterl37 and Farrell et al.138 thatng the right atrium reduces aldosteroneon.
Consideration of Matters Relevantriments on the Control of Aldosterone
as already been shown that the adrenalds by aldosterone secretion to concurrent
of Na+ and increase of K; concentrationenvironment. A considerable number ofents has been reported dealing with then of humoral stimulation of aldosteroneon. At this stage it can be said that theof the Na+ depleted adrenally-insufficient4,15 and the blood of the dog with I.V.C.22 when used for cross circulation either inival preparationl4 or the acute experiment22the adrenal over the period of a few
of the experiment.tions which arise are, (I) whether activationby excitation or by release of inhibition,nce arises the humoral agent and what
be its nature, (3) by what mechanism is theof the humoral agent governed.ion or Release of Inhibitione tissues are inactive unless excited whereasare spontaneously active, e.g. sheep'sheart muscle. Inactivity in the lat-
ues is due to inhibitory influences andarises by release of inhibition. It is
ically possible that the adrenal is under thece of an inhibitory material acting whenimal is in normal Na+ and body fluidI. The relative inactivity of in vitros makes this improbable, but aldosteronetion in these circumstances is high com-with cortisol.85 That total removal of theanial contents, including the hypophysis,ot activate aldosterone secretion in sheep inNa+ balance points in the same direction.the present we may accept the propositiondosterone secretion arises from excitationadrenal.
ature of the Excitationusual connotation of excitation is thatcommences at some relatively constant
) period after application of the excitatorycontinues for the duration of application
EDGAR NORMAL SODIUM BALANCE100 86/5/59
40PAROTIDSALIVARY 2ol)
NGYK*RATIO IO
PAROTIDSALIVARYSECRETION0 , 4
RATE O "' --,ml./minr
SPINO-MEDULLARYARY DECEREBRATION
AND HYPOPHYSECTOMY4 6 0 12 14 16 -re A 22
FIG. 13.-Edgar-normal Na+ balance. The effect onparotid salivary Nai'K+ ratio of spinomedullarydecerebration and hypophysectomy. The parotidsecretion rate is shown also.
ELMER DECEREBRATION NORMAL SOoIUM BALANCE29-30/4/S9
PAROTID 4
SALIVA //
RATIO
PAROTID 10SAU'A 0
SECRETION *).. W^. - -- -
RATE ISeNoaNC03
m!~ SPWQO-MEDULLARY
RY AND HPOHYSECTOMY4 6 6 c0 1'2 4 56i 202i2 4
HOURS
FIG. I4.-Elmer-normal Na+ balance. The effect onparotid salivary Na+/K+ ratio of spinomedullarydecerebration and hypophysectomy. The parotidsecretion rate is shown also. Approximately 5hours before the decerebration, the sheep wasgiven 150 m. equiv. of NaHCOs to ensure thatNa+ balance was normal.
(failure to do so is due to either fatigue or accom-modation), and ceases at some fairly constantperiod after cessation of the application. There isa usual assumption, based on experience and itsstatement, e.g. in Muller's law, that the reactingtissue will have highest sensitivity to one parti-cular environmental change (chemical in thisinstance), and that this substance arises locally orfrom specialized tissue which is usually groupedin an organ.Removal of the site of origin should result in
cessation of activation, with the appropriate timerelations. Figs. 13 and 14 depict the results of
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
96 POSTGRADUATE MIEDICAL JOURNAL February I14an experiment which indicate several considera-tions of importance in the analysis of thesemechanisms. Sheep in normal Na+ balanceshowed a variable reduction of parotid Na-/K-ratio associated with anaesthesia and preparativesurgery. Two hours after removal of the in-tracranial contents this Na K- ratio began to riseand five hours later was at the top of the normalrange. If the experiment had terminated then theresult would indicate that the source of the excita-tion of the adrenal had been removed, but two tofour hours later, without consistent trend in anyphysiological function being measured, the parotidNa+/K~ ratio began to fall again and in six hourshad reached the previous low level. Presumablya series of changes followed each other but it seemslikely that the adrenals were stimulated as a resultof some change not involving activity of theencephalon or hypophysis. The last six hours ofthis experiment produced a result which makesclear that a simple interpretation at the 13 hoursmark is not satisfactory.The result contrasts with some of the findings
in our series of experiments on Na + depleted sheep,15where de-encephalization and hypophysectomydid not cause the low salivary Na+iK+t ratio torise. That the continued low level was due toactivity of the adrenals was demonstrated by theappropriate rise after bilateral adrenalectomy.Other experiments demonstrated that this con-tinued activity was not reversed by replacement ofNa+ to produce plasma Na+ levels much in excessof normal. Indeed, these experiments, taken inconjunction with the failure of local increase of theNa+ concentration in the adrenal artery duringNa+ deficiency to stop aldosterone secretion andBartter's curious results showing failure ofinhibition of the adrenal after caval deconstrictionin vagotomized dogs, raise the possibility that aspecial mechanism is required to stop the adrenalactivity once it has been started, or that the acuteexperimental procedures have started up a newmechanism activating the adrenal. This latter isclearly a possibility in experiments which producepulmonary oedema and other dire results asrecorded by Davis et al.98 after cervical vagotomy.The importance of temporary consequences of
procedures in assessing the significance of short-term experiments applies also to more physiologicalprocedures than those dealt with above. If to asheep depleted of 700 mEq. of Na+ 6o ml. of4 M NaCl240omEq. Na+ are given intravenouslyduring 40 min., the result is likely to be that after3 to 4 hr. the Na+/K+ ratio is nearly normal, anindication that the adrenal activity receded tonormal. But after 7 to 9 hr. the ratio may bereduced again-even to the original level-indicating reactivation of the adrenal, though there
had been only 50 mEq. Na+ loss in this periApparently the rapid injection produced chanat appropriate points which stopped the adreactivity, but then perhaps redistribution of tNa+ occurred and brought about reactivatiInterpretation at the peak might have suggesthat the Na+ depletion was completely correcbut interpretation when stability was reacclearly indicated that replacement was not coplete and that there were most interesting phphenomena.Having these general points under view, so
principal experiments concerned with informatabout the sources of activating substances willconsidered.
The Pituitary Gland(a) Hypophysectomy. Data on the chronic effe
of hypophysectomy indicate some reductionaldosterone in adrenal vein blood of dogs26 whichconsistent with the results of Davis et al.'40 aSinger and Stack-Dunne94 for rats. An importfeature of the experiments of Davis et al. on dowas that hypophysectomy was carried out sixeight weeks before caval constriction, which cauthe usual rise of aldosterone excretion. Llauradhad similar results for urinary aldosterone ecretion in man. In acute experiments on dogs wichronic ascites due to caval constriction hphysectomy caused a large fall (76 to 97 per ceof aldosterone in adrenal vein blood during t2-hr. period of observation.l42 In discussingfinding by Farrell et al.26 that hypophysectoreduced aldosterone by no more than 40 to 50cent., Davis et al. attributed the difference to tfact that in Farrell's experiments there wasadditional factor of blood loss stimulatingadrenal, it being implicit that blood loss did nact via the pituitary. A number of reportspanhypopituitarism in man143-145 indicate variaurinary levels of aldosterone for patients not unNa+ depletion, and even more variable responsesthis, being approximately normal in somealmost absent in others.'46 Daily and Ganonrecorded normal response to Na+ restrictionhypophysectomized dogs. McDonald (unplished)148 has shown that hypophysectomized srespond to sodium depletion by normal fallsalivary Na+/K- ratio. Knobil and Greefound that hypophysectomized monkeys toleraNa+ restriction as well as intact monkeys,they did no specific studies on aldosterone.
ACTHThere can be little doubt that ACTH does
control the level of aldosterone productiHernando et al.44 refer to similar diurnal excretrhythms for aldosterone and other cortical p
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
Fi(;. 15.--Normal sheep brain with pineal gland itt situ.
·ir- :-"p.: .ai?p.ip.ca;·· ri
ii··;v *·I:aa;'; ·i:": .f':.: :i··.·'i·
t·
..:·..
a
Ir.·.
* ilEci :k.s :·..1; .r ·il. ··
(:··
.
;;:·-
.*, "'· ··'·q:;im::i·.Prrl:
*, *; :;k·-·..Fi;2.··R ·I
% aY.. .i.BL1-
bbIl'k. ?a.3111EE.EL .' asl-.
.c.C.,,
i: ·;·r·*
FIG. I6.-Delilah-complete pinealectomv.
ts and Davis'4" finds corticosterone and aldo-one varying approximately concomitantly afterphysectomy, but there are many instances ofependent variations.:4.i 1. 1, 88 Davis's'l'Its also show some non-correspondence ofterone and corticosterone. As previouslytioned,'4 excessive doses of cortisol do not
vent normal salivary response to Na- depletion.lar results came from Farrell, Banks andetsky,'50 who showed that prolonged cortisoneage suppressed adrenal secretion of cortisol
and corticosterone, but had little effect on aldo-sterone.
There are, however, reports which indicate thatACTH may play some part in aldosterone pro-duction. Hernando et al.'44 came to the conclusionthat ACTH may enhance the secretion of aldo-sterone, but not initiate it. This is consistent withthe results of NMcDonald and Reich152 on aldo-sterone output from the adrenal of Na--deficientsheep. Singer and Stack-Dunne94 got similarresults in rats, and de GratffW45 had similar reports
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
FIG. 17.-Freud--serial section of diencephalon and mid brain of sheep Removal of pineal and a sectionof the roof of the aqueduct of Svlvius.
for humans. Some of the differences betweencontradictory results may be due to contamina-tion of ACTH preparations with aldosteronestimulating material, but the variation in the assaymethod is also probably important.
There are no reports clearly indicating thatgrowth hormone or MSH have an effect on aldo-sterone secretion.
Pineal GlandFarrell'l' has presented data which suggest that
the pineal gland may be the source of a humoralsubstance regulating aldosterone secretion. Withthis in view complete pinealectomy was done ontwo sheep with parotid fistulae. Fig. i5 shows anormal brain with the pineal gland in situ immedi-atelv anterior to the superior colliculi. Fig. i6shows the same area at the post-mortem examina-tion of Delilah. The pineal has been completelyremoved, a finding confirmed by microscopy ofserial sections. Fig. 17 shows the posterior di-encephalon and midbrain of Freud cut in coronalsections. The pineal has been removed and therehas been destruction of an area along the roof of theaqueduct. Histological examination is proceeding.Three weeks after operation Delilah was tested andresponded normally to Na-- deficiency. After threedays' depletion the animal was permitted to drink600 mMol. of NaHCO3 in 2 1. of water and thesalivary Na--'K` ratio rose from o.9 to 28 overthe next 4 hr. (Fig. I8). With Freud, as shownabove, the lesion was much more extensive, anda significant feature of the post-operative conditionwas gross hvperextension of the neck. The sheep
+ 30 DELILAH SOIUM DEPLETEDFREUD 1/10/59
+25 DELLAH Na/K, RATIO*---,. K+CONCENTRATION
FREUD NK RIO
+20 K+CONCENTRATION.ORRECTEDPAROTID
SALIVARY
NaK*+ I5sRATIO
-120 mequli/l+10 CORRECTED DRANK
PAROTID8O SALIVARY I
POTASSIUM NHC3n+I 2I20
o40
+40
s-S.
2 4 6 6 ' OHOURS
Fi(;. i8.-Delilah-pinealeztoniiz ed and Na- depleteFreud-pinealectomized and Na- depleted. 'heffect on parotid salivary Na- K- ratio (correctfor secretion rate variation) and salivary K- concentration (corrected for secretion rate variation)voluntary intake of 6o: m. equiv. of NaHCOdissolved in 2 1. of H.,O. In each instance thsheep drank all the solution immediately it waoffered.
required considerable attention during the sereweeks it was observed, and this included perioof hand feeding. Because it contintiallv held i
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
l9ar60I96((C()(;HIAN, DENTON, (;()DING; and \VRI(;HT: ('ontr, l (of .Aldostcr,re Secretion 99
ad in this position accurate assessment of saliv-aas not possible. However, the salivary
Na+!,'' ratio did decrease w-hen the daily Napplement w\-as withheld and, as shown- on Fig. IS,e ratio rose at approximately the usual timehen it Nwas permitted to drink 6oo m:\I. ofaHC()O solution. Cardiovascular deteriorationlnth Na- deficiency wvas greater than normal.
Gross postural disturbanice interfering Nwith thevoluntary intake was observ-ed by Andersson
rsonal communication) in the course of hisperiments examining Gilbert'si:3 assertion thate subcommissural organ has a secretory roleportant in water and electrolyte metabolism.It seems clear therefore that the pineal com-nent of material extracted by Farrell et atl."S
which increased aldosterone production by theadrenals in his preparation is not essential forrnormal aldosterone responses to Na' deficiency.Nor does it appear that the decrease in aldosteroeic'production resulting from pinealectomy in siInilaripreparations'54 gives a cogent reason for regarding[this structure as having other than its Cartesianfunction.In the light of earlier consideration of the
possibility of functional integration of aldosteronecontrol and the neural mechanisms determiningithe selective appetite for Na- exhibited by de-
1ficient sheep,'4 15 a salt appetite study w-as con-ducted on Delilah. The animal demonstratedthe usual capacity of sheep, losing approximately0o mEq. Na'-day from a parotid fistula, to adjusta+ intake to loss despite variation in the con-ntration of the NaHCO, solution (Fig. i9).xtractsThere have been a number of papers which hav-eported an enhancement of aldosterone secretiony infusion of extracts. Almost every part of theiencephalon has been extracted'l°l 31, 14The -widespread areas from which activity hasen demonstrated raise the question as tocificity of the material used, and indicate theed for careful consideration of the criteria forrmone characterization set out in the intro-
uction. Formal study of the response of therenal to pharmacologically active materials isuired also. There are suggcstive aspects of
e studies but, to date, no specific tissue has beenentified as the source of a substance causinghysiological excitation of aldosterone secretion.
~ervous illechanisnms Concerned zwith Release ofHumoral AgentThe adrenal response to Na- depletion has beenown to be proportional to the absolute Naficit and irrespective of raised or loweredasma Na- contriv-ed by! varying the relative body
30 DE LI LAH Oct.1959 (Pineolectomized)Parotid ,
Salivary 20Na?K'/Ratio 10
2-69 I. 247 i. 2-74 I. 2-68 I.m.equiv. Average saliva+400 volume 'day
Cumulative
Sodium+200
Balance
-200
m.equiv. av. 470 av.460 av.400 av.470600
VoluntarySodium300Intake
m.equiv,t600
Contn. of300
NaHCO3Solution
5 10 15DAYS
FIG. I9.-Delilah. Study of selective appetite for Na .The effect of variation of the concentration ofNaHCO3 solution offered on the voluntary intakeof NaHCO. The salivary Na ,K+ ratio and theaverage daily loss of saliva in each period is shownalso. The cumulative Na'- balance as calculatedat the end of each 24-hour period is shown. WVaterwas available continuously and the animal ate allof its constant ration of food each day throughoutthe experiment.
wvater content. There has been no systematicinvestigation of receptors concerned with thisreaction. The response to bleeding is stated byHolzbauer et al.'3~ to require intact splanchnicnerves as receptors. Mills, Casper and Bartter95indicated that intact vagus nerves were essentialfor cessation but not for initiation of the responseto caval constriction. Davis et al.98 have con-firmed this latter proposition but did not releasethe caval constriction of any dogs followingvagotomy. They did not confirm Bartter's'00other proposition that the carotid sinus innervationplays a part in initiating aldosterone production.Rauschkolb and Farrell30 report that cervicalspinal transection, vagotomy, and section of thecervical sympathetic did not reduce the aldosteroneproduction of dogs.
Removal of part of the nervous system has beenwvidely practised. Farrell et al. indicated that
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
100 POSTGRADUATrE AIEDICAL JOURNAL Febrluar-y 9g6Feruri9decapitation or mid-collicular decerebration, w-ithnoradrenaline support, greatly reduced the secre-tion of aldosterone in their dogs, whereas decortica-tion and spinal transection did not.
In less extensiv-e lesions, Farrell et al./1°'0. 13raise the possibility that there is an area in the mid-brain crucial to aldosterone secretion but methodo-logical aspects of the evidence wNarrant caution.
Davis et al.'36 studied the effects of chroniclesions on the aldosterone excretion associated w-ithcaval constriction. He found that only thoselesions involving the median eminence causedreduction of aldosterone secretion but the lesionsdid not cover the area most effective according toFarrell, nor do Farrell's lesions involve the medianeminence.
In this laboratory we have investigated the effectof removing some parts of the nervous system onthe parotid salivary Na- K*- response in Na-depleted and normal sheep. Earlier experimentsinvolving de-encephalization and hypophysectomvin sheep in normal -Na balance w-ere mentioned.It has not yet been possible to keep animals w\ithde-encephalizatioin alive long einough to seex-hether they respond to Na- depletion by changeof parotid 'Na K-- ratio. It has been found thatdecerebration at the level of the anterior border ofthe superior colliculus does not cause a change illthe parotid Na- K of depleted sheep, but suchanimals respond to intravenous correction of Na-deficiency. UWhen the sections passed through theinferior colliculus or Xwere prepontine or spino-medullary, and there was anterior removalassociated w-ith hypophysectomy, in the majoritythere was little or no effect on the low parotidNa- 'K- and those animals did not respondnormally to intravenous sodium replacement.In some animals there w-as a definite rise ofNai''KK: upon spino-medullary deencephalization,but in one instance this later reverted to theoriginal level. That the low ratio \was due toadrenal activity was shown formally in three suchexperiments by the ratio rising to io by 3 to 9 hr.after bilateral adrenalectomv. The results ofthe 3 I experiments in this series to date tentativelysuggest a significance in relation to Na- controlof the region between the anterior border of thesuperior colliculus and the rostral border of thepons but much more convincing evidence, in-cluding survival experiments, is required.
It seems clear that there is a nervous level oforganization required for reversal of response toNa depletion in excess of that required to keepthe reaction continuing despite Na- replacement.The hypophysis is not essential to keep the reactioingoing.The present state of knoNwledge in regard to
regulation of aldosteronie secretion is thus far froill
complete. Progress w-ill depend upon clerecognition of the several sorts of circumstanc\-hich can elicit the adrenal response. It mabe necessary to overcome the great difficultiinherent in demonstration by survival experimein order to substantiate as physiologicall) valisuggestive evidence on aldosterone control arisinfrom acute experimental preparations.SUMMARYThe elucidation of complex clinical conditio
involving disorder of aldosterone secretion requires identification by animal experiment of thmain mechanisms 'which control aldosteronsecretion. The introductory section of this papeis concerned with the methodological basis osuch animal experiments. The principle oapproach adopted in the authors' laboratory habeen that of step by step surgical preparation oan animal so that the function of anatomicalldeep seated, and otherwise inaccessible, structures can be studied in the conscious confidenanimal. It is important that the process omeasurement of a physiological function shoulnot itself introduce uncontrolled or undesirevariables. The baseline levels are found in thconscious confident animal. A sheep w-ithpermanent unilateral parotid fistula and w-ith thleft adrenal gland transplanted to a combinecarotid artery-jugular vein skin loop in the necrepresents a Xvery satisfactory preparation forexperiments on aldosterone control. Using theKliman and Peterson double isotope derivativeassay for steroids, it is possible to have an accuratelrecord, at frequent intervals, of adrenal secretionrates without sampling causing significant bloodloss, and concurrent measurement of parotidsalivary Na-- 'K- ratio gives a continuous recordof peripheral blood aldosterone concentrationw-hich, because of the short half-life of aldo-sterone, rapidly reflects changes of adrenal secre-tion rate. Difficulties in the interpretation ofacute experiments involving surgical proceduresiunder anaesthesia are discussed. The criteria for!categorization of an extract which is active uponlan effector organ as a hormone are considered.Some of the principal chemical and biological
methods for measuring aldosterone are described!in the second section of the paper. In the thirdsection, effects of variation of Na- and body fluidbalance on aldosterone secretion are discussed.This includes a summary of experiments in thislaboratory demonstrating that local concurrentreduction of Na-- and increase of the K- concen-trations in adrenal arterial blood causes increasedaldosterone secretion. Some aspects of the evi-dence that there is a humoral agent other thanACTH Nwhich stimulates aldosterone secretion-
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
_ 1I960 COGHLAN, DENTON, GODING and WRIGHT: Control of Aldosterone Secretion Ixocussed also. The final section of the papery concerned with experiments on the
nce of the central nervous system on aldo-secretion. Decerebration experiments in
rs' laboratory indicate central nervousnce on aldosterone secretion and suggest anant localization in the midbrain, but the
ts emphasize the difficulties of interpretationriments involving acute nervous ablations.
val experimental study of pinealectomizeds did not provide evidence of this structurea role in the adrenal response to changes
a+ balance.REFERENCES
UGUST, J. T., NELSON, D. H., and THORN, G. W.(i958), .C.din. Invest., 37, I549.NN, J. W. (I955), J. Lab. clini. Med., 45, 3.NN, J. W., and LOUIS, L. H. (1956), Ann. intern. Med.,
44, .OSS, F and LICHTLEN, P. (g958), 'Aldosterone,'
edited byA. F. Muller and C. M. Connor, p. 39. London:Churchill.AVIS, J. O., PECHET, M. M., BALL, W. C., Jnr., andGOODKIND, M. J. (1957), ..clin. Inm*st., 36, 689..AVIS, J. O., HOWELL, D. S., and SOUT1HWORTH,J. L. (I953), Circulation Res., X, 260.
MAN, W. H. (1957), Biochem. Y., 66, 17.:YRES, P. J., BARLOW J., GARROD, O., KELLIE,A E., TAIT, S. A. S., TAIT, J. F., and WALKER, G.(I958), 'Aldosterone' edited by A. F. Muller and C. M.O'Connor, p. 73. London: Churchill.ICK, S., LARAGH, J. H., and LIEVERMAN, S. (I958),
Trans. Ass. Amer. Phys., 71, 225.RNE:Y, E. B. (I947), Proc. roy. Soc. B., 13S, 25.NTON, D. A., and McDONALD, I. R. (1957), . Physiol.,138. 44.KETT, M. F., O'CONNOR, W. J., and VERNEY, E. B.
(1942), 'Quart. . exp. Physiol., 31, 333.rARLING, E. H., and VERNEY, E. B. (1925), Proc. roy.Soc: B., I99, I.NTON, D. A., GODING, J. R., and WRIGHT, R. D.
'(1959), Brit. mred. Y., iH, 447 and 522..ENTON, D. A., GODING, J. R., and WRIGHT, R. D.(959), ' Progress in Clinical Endocrinology' (in the press).New York: Grune & Stratton.ENTON, D. A. (I957), 7. Physiol., 135, 227.DING, J. R., and DENTON, D. A. (I959), Aust. Y. exp.
Biol. med. Sci., 37, 2I 1.ENTON, D. A. (956), .7. Physiol., 131, 13.:NTON, D. A. (1957), Quart.7. exp. Plysiol., 42, 72.cDONALD, I. R., GODING J. R., and WRIGHT, R. D.(1958), Aust. 2. exp. Biol. mcd. Sci., 36, 83.LIMAN, B., and PETERSON, R. E. (x958), Fed. Proc.,
17, 255.ANKOPOULIS, N. A., DAVIS J. O., KLIMAN, B., andPETERSON, R. E. (I959), i.cin Invcst., 38, 1278.ERSON, R. E. (I959), Rec. Progr. Hormone Res., IS, 231.E, D. M., and NELSON, D. H. (1954), Surg. Forwnm,
5, 568.'VER, R G., and EIK-NES, K. B. (I959), 7. appl.
Physiol., x4, 665.ARRELL, G. L., RAUSCHKOLB, E. W., and ROYCE,P. C. (1955), Amer. Y. Physiol., 182, 269.ARRELL, G. L., ROSNAGLE, S and RAUSCHKOLB,E. W. (I956), Circulation Res., 4, 66.USCHKOLB, E. W., and FARRELL, G. L. (I956),
Endocrinology, 59, 526.ARRELL, G. L. (I958), Physiol. Rev., 38, 709.ARRELL G. L FLEMING, R. B., RAUSCHKOLB,E. W., YATSU, i. M., McCALLY, M., and ANDERSON,C. H. (I958). Endocrinology, 6a, so6.ARRELL, G. L. (i99s), Ibid., 65, 29.ARELL, G. L. (1959), bid., 6s, 239.ILL, J S.S. (843), 'A System of Logic,' p. Sos. London:Longmans.ORN, G. W., ROSS, E. J CRABBE, J., and VAN'T
HOFF, W. (1957), Brit. med. i., i, 955.
35. EBERLEIN, W. R., and BONGIOVANNI, A. M. (s955),Arch. Biochem., 59, 90.
36. DEANE, H. W., SHAW, J. H., and GREEP, R. 0. (x948),Endocrinology, 43, 133.
37. LUETSCHER, J. A., Jnr. (1956), Advanc. intern. Med.,8, s55.
38. GROSS, F. (Is59), Schweiz. med. Wschr., 89, I.39. GAUNT, R., RENZI, A. A., and CHART, J. J. (I955),
J. clin. Endocr., IS, 621.40. AUGUST, J. T., NELSQN, D. H., and THORN, G. W.
(I958), New Engl. 7. Med., 259, 917 and 967.41. SIMPSON, S. A., TAIT, J. F., WETTSTEIN, A., NEHER,
R., VON EUW, J., and REICHSTEIN, T. (I953), Experi-entia (Basel), 9, 333.
42. SIMPSON, S. A., TAIT, J. F WETTSTEIN, A., NEHER,R., VON EUW, J., SCHINDLER, O., and REICH-STEIN, T. (I954), Htelt. chim. Acta, 37, xI63.
43. ULICK, S., and LIEBERMAN, S. (x957), J. Amer. chem.Soc., 79, 6567.
44. KAGAWA, C. M., CELLA, J. A., and VAN ARMAN, C. G.(i957), Science, 126, IOI5.
45. LIDDLE, G. W. (I957), Ibid.;' 26, ioi6.46. DASGUPTA, D., and -GIROUD,i C.. J. P. (I959), Endo-
cr,nok,oy, 65, Soo.47. SINGER, B. (g959), Ibid., 65, gi2.48. RENOLD, A. E., CRABBE, J., ROSS, E. J., HERNANDO-
AVENDANO, L., NELSON, D. H., and THORN, G. W.(I956), Y. clin. Invest., 35, 731.
49. RENOLD, A., CRABBE, J., HERNANDO-AVENDANO,L., NELSON, D. H., ROSS, E. J., EMERSON, K., andTHORN, G. W. (I957), New Engl. J. Med., 256, x6.
50. JENKINS J S., MEAKIN, J. W., and NELSON, D. H.(1959), ndrocrinology, 64, 572.
5I. MACH R. S., FABRE, J., DUCKERT, A., BORTH, R,and bUCOMMUN, P. (I954), Snhweiz. med. Wschr.,84 407.
s2. SPEIRS, R. S., SIMPSON, S. A., and TAIT, J, F. '(954),Endocrinology, 55, 233.53. GROSS, F., and GYSEL, H. (I954), Acta endocr. (Kbh.),
IS, 199.54. WOODBURY, D. M., and KOCH, A. (I957), Proc. Soc.
cxp. Biol. N.Y., 94, 720.55. CONN, J. W. (I949), Arch. intern. Med., 83, 4Ix56. FRAWLEY, T. F., and THORN, G. W. (Ig9I, Proc. 2nd
ACTH Conference, I, I15 London: Churchill.57. GODING, J. R., and DENTON, D. A. (1057), Aust..7. exp.Biol. med. Sci., 35, 301.O8. ROSS, E. J., CRABBE, J., RENOLD, A. E., EMERSON,
K., Jnr., and THORN, G. W, (I958), Amer..7. Mea.,2S, 278.59. SWINGLE, W. W., DA VANZO, J. P., GLE.NISTER, D.,CROSSFIELD H. C., and WAGLE, G. (I959), Anmer. .7
Physiol., s96, 283.60. NEHER, R., MEYSTRE, Ch., and WETTSTEIN, A. (I959),Helv. chim. Acta, 42, 132.6i. BLIZZARD, R. M., LIDDLE, G. W., MIGCEOTN, E. J., and
WILKINS, L. (t959), .7 clin. Invest., 38, I442.62. NEHER, R., DESAULLES, P., VISCHER, E., WIELAND,P., and WETTSTEIN, A. (I958), Helv. chim. Acta, 41, x667.'63. DESAULLES, P. (x959), Experimnta, IS, 30o.63a. DESAULLES, P. (1958), 'Aldosterone,' edited by A. F.
Muller and C. M. O'Conndi, p. 29. London: Churchill.64. AYRES, P. J., GARROD, O., PEARLMAN, W. H., TAIT,J. F., TAIT, S. A. S., and WALKER, G. (x957), Cibaoudation Cooquia on Endocrinology, II, 309. London:
Churchill.65. BUSH, I. (1952), Biochem. .7, 5o, 370.66. ZAFFARONI, A., and BURTON, R. B. (i95I), .. biol.
Chem., 193, 749.67. GORNALL, A. G., GWILLIAM, C., and HALL, A. E. D.
(1956), Rev. canad. Biol., IS, 252.68. NOWACZYNSKI, W., KOIW, E., and GENEST, J. (x957),Can. .. Biochem., 35, 425.69. DYRENFURTH, I., and VENNING, E. H. (I959), Endo-
crinology, 64, 648.70. PETERSON, R. E. (x958), 'Proc. Symposium on Advances
in Tracer Applications of Tritium,' p. I6. New York:New England Nuclear Corp.
71. BUZZANO, G., and DINGMAN, J. F. (I959), Fed. Proc.,i8, x88.
72. SINGER, B., and WENER, J. (I953), Amer. Heart.7, 45, 79573. LUETSCHER, J. A., Jnr., and JOHNSON, B. B. (I953),Amer. .7. Med., 5S, 417.74. LUETSCHER, J. A., Jnr., and JOHNSON, B. B. (x954),.7. clin. Invest., 33, 276.
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F
102 POSTGRADUATE MEDICAL JOURNAL February il475. LUETSCHER, J. A., Jnr., and JOHNSON, B. B. (I954),
Ibid., 33, I441.76. ORTI, E., RALLI, E. P., LAKEN, B., and DUMM, M. E.
(I957), Amer. 7. Physiol., 19I, 323.77. LLAURADO, J. G. (1956), Endocrinology, 58, 390.78. SIMPSON, S. A., and TAIT, J. F. (1952), Ibid., 50, ISo.79. BUSH, I. E. (I957), Ciba Foundation Colloquia on Endo-
crinology, Ix, 263.8o. DAUGHADAY, W. H. (1958), 7. clin. Invest., 37, 511.8I. SANDBERG, A. A., SLAUNWHITE, W. R., Jnr., and
ANTONIADES, H. N. (I957), Rec. Progr. Hormone Res.,13, 209.
82. AYRES, P. J., GOULD, R. P., SIMPSON, S. A., andTAIT, J. F. (I956), Biochem. 37., 63, 19 P.
83. WETTSTEIN, A., KAHNT, F. W., and NEHER, R. (I955),Ciba Foundation CoUoquia on Endocrinology, 8, 170.
84. ROSEMBERG, E., ROSENFELD, G., UNGAR, F., andDORFMAN, R. (I956), Endocrinology, 58, 708.
85. GIROUD, C. J. P., STACHENKO, J., and VENNING,E. H. (I956), Proc. Soc. exp. Biol., N.Y., 92, 855.
86. MULROW, P. J., SHMAGRANOFF, G., LIEBERMAN, A.,SHADE, C., and LEUTSCHER, J. A., Jnr. (I959), Endo-crinology, 64, 631.
87. GIROUD, C. J. P., STACHEN-KO, J., and PILETTA, P.(I958), 'Aldosterone,' edited by A. F. Muller and C. M.O'Connor, p. 56. London: Churchill.
88. LANE, N., and DE BODO, R. C. (1952), Amer. 7. Physiol.,I68, x.
89. DE BODO, R. C. (I959), Recent Progr. Hormone Res., 15, 302.0o. FELDBERG, W. (1941), 3. Physiol., 99, I04.
9I. VOGT, MARTHE (i944), Ibid., 103, 317.92. VOGT, MARTHE (igS5), Ibid., 113, I129.93. FARRELL, G. L., ROYCE, P. C., RAUSCHKOLB, E. W.,
and HIRSCHMANN, H. (I954), Proc. Soc. exp. Biol.(N.Y.), 87, I4I.
q94- SINGER, BERTHA, and STACK-DUNNE, M. P. (I955),Y. Endocr., 2a, I30.
-*95. MILLS, I. H., CASPER, A. G. T., and BARTTER, F. C.(I958), Science, 128, 1140.
.96. HILTON, J. G., WEAVER, D. C., MUELHEIMS, G.,GLAVIANO, V. V., and WEGRIA, R. (1958), Amer. 3.Physiol., 192, 525.
97. DAVIS, J. O., KLIMAN, B., YANKOPOULIS, N. A., andPETERSON, R. E. (I958), 3. clin. Invest., 37, 1783.
98. DAVIS, J. O., YANKOPOULIS, N. A., and HOLMAN, J.(I959), Amer. 3. Physiol., 197, 207.
99. DAVIS, J. O., BAKER, R. C., YANKOPOULIS, N. A.,KLIMAN, B., and PETERSON, R. E. (I959), Ibid.,197, 380.
Ioo. BARTTER, F. C., MILLS, I. H., BIGLIERI, E. G., andDELEA, C. (I959), Rec. Progr. Hormone Res., rS, 311.
IoI. FARRELL, G. (I959), Ibid., IS, 275.t02. NEWMAN, A. E., REDGATE, E. S., and FARRELL, G.
(I958), Endocrinology, 63, 723.103. MASONI, A. (1957), -Acta med. scand., 159, 225.t04. LEVY, S. E., and BLALOCK, A. (I939), Ann. Surg., IO9, 84.so5. SiMPSON, S. A., and TAIT, J. F. (1955), Ciba Foundation
Colloquia on Endocrinology, 8, 204.,o6. LANGLEY, L. L., BEALL, W. A., and SMITH, J. A.
(I959), Amer. 3. Physiol., 197, 565.o07. GODING, J. R., and COGHLAN, J. P. (to be published).io8. BARGER, A. C., BERLIN, R. D., and TULENKO, J. F.
(I958), Endocrinology, 62, 804.109. McDONALD, I. R., and REICH, M. (I959), 3. Physiol.,
147, 33.xIo. ROSNAGLE, R. S., and FARRELL, (;. L. (1956), Amer. 7.
Physiol., 187, 7.II. CRABBE J., REDDY, W. J., ROSS, E. J., and THORN,
G. W. (1958 ),. clin. Endocr., 18, II47.112. MILLS, I. H (I959), Rec. Progr. Hormone Res., s5, 261-264.113. McDONALD, I. R., and DENTON, D. A. (1956), Nature,
177, 1035.114. GORNALL, A. G. (I959), Rec. Progr. Hormone Res., 15, 338.Z15. VAN BUCHEM, F. S. P., DOORENBOS, H., and ELINGS,
H. S. (1956), Lancet, 271, 335.iI6. CHALMERS, T. M., FITZGERALD, M. G., JAMES,
A. H., and SCARBOROUGH, H. (I956), Ibid., 270, 127.
117. WARFIELD, J. M., FOX, C. L., and MERSHEIW. L. (I95I), Ann. Surg., I34, 626.
Ix8. MILNE, M. D., and MEUHRCKE, R. C. (1957), Brit.Bull., I3, I 5.
II9. NEWMAN, A., REDGATE, E. S., and FARRELL, G(1958), Endocrinology, 63, 804.
120. DUNCAN, L. E., Jnr., LIDDLE, G. W., and BARF. C. (I956), J. dlin. Invest., 35, 1299.
X2I. LIDDLE, G. W., DUNCAN, L. E., Jnr., and BARF. C. (I956), Amer. Y. Med., 21, 380.
122. DENTON, D. A. (1957), Y. Physiol., I40, 129.123. LARAGH, J. H., and STOERK, H. C. (I957), J. clin. I
36, 383.124. JOHNSON, B. B., LIEBERMAN, A. J., and MULR
P. J. (I957), Iid., 36, 757.125. BARTTER, F. C., LIDDLE, G. W., DUNCAN, L. E.,
BARBER, JOAN, and DELEA, CATHERINE (!9Ibid., 35, 1306.
126. VENNING, E. H., DYRENFURTH, I., GIROUD, C. J.and BECK, J. C. (X957), Canad. med. Ass. Y., 77, 773.
127. FALBRIARD, A., MULLER, A. F., NEHER, R.,MACH, R. S. (I955), Schweiz. med. Wschr., 85, 1218.
I28. MULLER, A. F., and MACH, R. S. (1957), Reunion d'Ecrinologie (Geneva), 4, 25.
129. GRAY, J. S. (Igpo), 'Pulmonary Ventilation and its Phylogical Regulation.' Springfield, Illinois: Thomas.
130. FARRELL, G. L., RAUSCHKOLB, E. W., FLEMIR. B., and YATSU, F. M. (1957), Proc. 39th Mtg. ESoc. (N.Y.), p. 32.
I31. NICHOLS, G., and NICHOLS, N. (I956), Met5, 439.
132. RICHTER, C. P. (I958), in 'L'instinct dans le compment des animaux et de l'homme.' Paris: Masson.
133. CHESTER JONES, I., PHILLIPS, J. G., and HOLW. N. (I958),' Comparative Endocrinology,' p. 582.York: Wiley & Sons.
I34. CHESTER JONES, I. (I957), 'The Adrenal CoLondon: Cambridge University Press.
135. BALL, W. C., and DAVIS, J. 0. (I957), Amer. J. Phy191, 339.
136. DAVIS, J. O., and BALL, W. C. (1958), Ibid., 192, 538.137. BARTTER, F. C. (i959), in Rec. Progr. Hormone
IS, 305.138. McCALLY, M., ANDERSON, C. H., and FA
G. L. (I958), Proc. 40th Mtg. Endocrine Soc. (San Frp. II9.
139. HOLZBAUER, M., and VOGT, M. (I959), Proc. pSoc., May 23, 19S9, p. I3.
140. DAVIS, J. O., BAHN, R. C., GOODKIND, M. J.,BALL, W. C., Jnr. (I957), Amer. 3. Physiol., 191, 329.
141. LLAURADO, J. G. (I957), Metabolism, 6, 556.142. DAVIS, J. O., BAHN, R. C., YANKOPOULIS, N.
KLIMAN, B., and PETERSON, R. E. (I959), AmPhysiol., 197, 380.
143. LUETSCHER, J. A., Jnr., and AXELRAD, B. J. (iJ. clin. Endocr., 14, io86.
144. HERNANDO, L., CRABBE, J., ROSS, E. J., REDW. J., RENOLD, A. E., NELSON, D. H., and THG. W. (1957), Metabolism, 6, 5x8.
145. DE GRAEFF, J. (1958), 'Aldosterone,' edited byMuller and C. M. O'Connor, p. 130. London: Ch
146. MULLER, A. F., MANNING, E. L., and RIONDEL, A.(I958), Ibid., p. I I I.
147. DAILY, W. J. R., and GANONG, W. F. (1958), Ecrinology, 62, 442.
I48. McDONALD, I. R., in the press.149. KNOBIL, E., and GREEP, R. 0. (I958), Ibid., 62, 6i.I5o. FARRELL G. L., BANKS, R. C., and KOLETSKY
(1956), Ibid., 58, 104.ISI. LAMSON, E. T., ELMADJIAN, F., HOPE, J. M., PINC
G., and JORJORIAN, D. (1956), J. clin. Endocr., x6,I52. McDONALD, I. R., and REICH, M. (I959), J. Phy
147, 33-I53. GILBERT, G. J. (1956), Anat. Rec., 126, 253.154. FARRELL, G., KOLETSKY, S., and LAPHAM, L.
(I959), Fed. Proc., 18, 44.I55. NEWMAN, A. E., REDGATE, E. S., and FARRELL, G
(x959), Ibid., 18, I I3.I56. DAVIS, J. O., BAKER, R. C., and BALL, W. C., Jnr. (i
Amer. J. Physiol., 197, 387.
by copyright. on F
ebruary 1, 2022 by guest. Protected
http://pmj.bm
j.com/
Postgrad M
ed J: first published as 10.1136/pgmj.36.412.76 on 1 F