MODES OF AMMONIA TRANSPORT ACROSS THE GILLEPITHELIUM OF THE MARINE TELEOST FISH OPSANUS
BETA
BY DAVID H. EVANS, KANE J. MORE AND STEVEN L. ROBBINS
Department of Zoology, University of Florida, Gainesville, FL 32611, USA
Accepted 6 February 1989
Summary
The modes of ammonia transport across the gill epithelium of the marine teleostOpsanus beta were investigated using an isolated, perfused head preparation.During perfusion for approximately 2h there was no macro- or microscopicdamage to the exterior surface of the gill filaments or lamellae, no swelling oflamellae, the structural leak was only 0-3%, the afferent perfusion pressure wasconstant, and the transepithelial electrical potential (TEP) approximated thatmeasured in vivo. The rate of ammonia efflux was similar to that measured in vivo,was constant for at least 80min, and approached zero when the perfusate wasnominally ammonia-free. Addition of ouabain or bumetanide to the perfusateindicated that basolateral Na+/NH4
+ exchange, but not Na+/NH4+/2CPcotransport, was important in transbranchial ammonia transport. No evidence wasfound for apical Na+/NH4+ exchange. Specific increases in perfusate NH3 werenearly 100 times as effective as increases in NH4+ concentrations in stimulatingammonia efflux. Alteration of TEPs could not account for any of these effects.Calculated relative permeabilities to NH3 and NH44" indicate that the toadfish gillis relatively permeable to NH4"1" compared to the shark gill and other 'tight'epithelia, consistent with the general conclusion that marine teleost gills arerelatively cation-'leaky'. Computation of the relative roles of the three delimitedpathways for ammonia efflux indicate that 22% is via basolateral Na+/NH4+
exchange, 57% via non-ionic diffusion of NH3 and 21% via ionic diffusion ofNH4"1". Comparison with other published data suggests that the relative roles ofnon-ionic and ionic diffusion of ammonia across epithelia may be correlated withrelative ionic permeabilities.
Introduction
Ammonia, which is the dominant product of amino acid catabolism in fishes, isexcreted predominantly across the gill epithelium. Evidence has been presentedfor the presence of various potential transport pathways in this epithelium,including basolateral and apical Na+/NH4+ exchange, basolateral Na+/NH4+/2C1~ cotransport, paracellular and/or transcellular diffusion of NH3, and paracel-
pey words: toadfish, Opsanus beta, gill, ammonia.
340 D . H . E V A N S , K. J. MORE AND S. L. ROBBINS
lular diffusion of NH4"1" (see Evans & Cameron, 1986, for a recent review). Wehave recently utilized the isolated, perfused head of the dogfish pup (Squalusacanthias) to quantify the role of these pathways in ammonia transport across thebranchial epithelium of an elasmobranch (Evans & More, 1988). The currentstudy used the isolated, perfused head of the Gulf toadfish {Opsanus beta) toexamine the role of each of these pathways in a teleost because the gills of marineteleosts have been found to have a much higher cationic permeability than those ofmarine elasmobranchs (Evans, 1979) and, therefore, might be expected to excretea larger percentage of ammonia as NH4+ (Evans & Cameron, 1986). In fact,preliminary studies of this species have suggested that ionic diffusion of NH4"1" maybe quite substantial (Goldstein etal 1982) and that basolateral Na+/NH4+
exchange may also play a role (Claiborne et al. 1982).
Materials and methodsCollection and maintenance
Sexually mature specimens of Opsanus beta were collected by a commercialshrimp fisherman in the Gulf of Mexico, near Cedar Key, Florida throughout theyear. Individuals utilized in the present study were significantly smaller (about20-40g) than those used in previous studies (about 150-250g: Oduleye etal.1982). Animals were maintained at room temperature (21-24°C) in glass aquariawith charcoal/cotton nitration, and were not fed for at least 24 h beforeexperimentation. Atlantic sea water (32-34 %o) was collected from the runningseawater system at Marineland, Florida.
Preparation of the perfused head
Perfusion of the isolated head followed essentially the procedure of Payan &Matty (1975) as modified by Claiborne & Evans (1980). Fish were injected with1000 units of sodium heparin 20-30 min before anesthesia with 0-01 g P 1 MS-222,and weighed to the nearest 0-1 g. The pericardial cavity was exposed, the heartligated between the atrium and ventricle, and the bulbus arteriosus/ventral aortacannulated with about 3 mm of PE 50 tubing attached to a perfusion line composedof PE90 tubing joined to Gilson Tygon tubing (i.d. =0-76mm). The body wasimmediately removed from the head behind the pectoral fin, and perfusioninitiated, so that the branchial vasculature was ischemic for only 2-5 min. Theswimbladder and gut were removed distal to ligatures, but the liver was usually notligated before its removal since preliminary experiments indicated that ligation ofhepatic vessels sometimes produced some postbranchial swelling. The mouth wasthen sewn shut around an irrigation input of PE 160 tubing attached to Tygontubing (i.d. =4-25 mm). Finally, the dorsal edge of the opercular opening wassewn shut to ensure filling of the buccal cavity before the irrigate left the gillchamber. The head was fitted with a plastic collar using stainless-steel surgicalstaples and held inverted in a translucent plastic cylinder by a rubber dam(proximal end of a small balloon). Cannulation of the dorsal aorta is not possibU
Toadfish gill ammonia transport 341
in this preparation so efferent outflow of perfusate was collected as the totaleffluent from the cut end of the head. Perfusion was driven by a Gilson Minipuls 2pump at constant flow (about 600^1min~1) which produced afferent pressures of2-67-4-66 kPa, in the range of that measured in larger animals in vivo (J. B.Gaiborne & D. H Evans, unpublished results). Perfusion pressures weremonitored, via a T-connection to the perfusion line, by a Gould Statham pressuretransducer and recorded on a Gilson Duograph to the nearest 0-13 kPa. Theperfusion flows, as well as the pressure transducer, were calibrated before eachexperiment. All pressures were corrected for internal perfusion-line resistance,which was measured before each experiment. Perfusion pulsations produced bythe Minipuls 2 pump were damped by a small cylinder in the perfusion line whichwas partially filled with perfusate (Windkessel). Irrigation flow was set at aboutlOmlmin"1 (about 40 ml 100 g~1min~1), with postbranchial irrigate drainingdirectly out of the opercular opening, through a hole in the bottom of theperfusion chamber, and into a beaker through a small funnel. The irrigate (100 mlof sea water) was bubbled with air and recycled via a Cole-Parmer Masterflexpump. Perfusate was bubbled with 1% CO2 in air provided from commercial,premixed gas cylinders. The composition of the Ringer's solution is presented inTable 1. Unless otherwise stated, perfusates contained 10~7moll~1 epinephrine,which produced spontaneous and continuous ventilation in most preparations.Ventilatory rates were not quantified, but approximated the relatively low ratesobserved in vivo for this bottom-dwelling species.
Heads were perfused for about 30 min after preparation to allow for completeremoval of blood from the branchial and cephalic vasculature and to achieve stable
Table 1. Compositions of experimental solutions
Solute
NaClNaHCO3
NaH2PO4
Sodium benzenesulfonateKC1KHCO3
Potassium benzenesulfonateCaCl2
CaSO4
Choline chlorideMgSO4
MgCl2.6H2OHeparinPolyvinylpyrrolidoneGlucose
RS
14814-92-7
2-6
1-26
1-24
2500i.u.r1
3 %5
-NaASW
7-52-5
11
50030-327
-C1ASW
2-7
499-5
10
10
56-8
* All solute concentrations are in mmol 1 l .RS, Ringer's solution; —NaASW, sodium-free artificial sea water; —QASW, chloride-free
artificial sea water.
342 D . H . E V A N S , K. J. MORE AND S. L. ROBBINS
afferent perfusion pressures. Thus, time zero in most experiments with thispreparation was at least 45min after initial perfusion of the head.
Measurements of structural and functional integrity of the perfused head
To assess the integrity of this preparation, and therefore its usefulness for aninvestigation of the modes of ammonia transport across the branchial epithelium,we measured the surface micromorphology of gill tissue, the actual structural leakof perfusate at the end of approximately 2h of perfusion, and the transepithelialelectrical potentials across the head.
To examine gill surface morphology, gill arches were removed from intact orperfused heads (after at least 2 h of perfusion) and fixed for at least 24 h in 2 %glutaraldehyde in 0-1 moll"1 cacodylate-HCl buffer (pH7-4) adjusted to theproper osmolarity with sucrose. Fixed tissues were rinsed in distilled water,dehydrated through a graded ethanol series, critical-point dried in liquid CO2,mounted on a specimen stub, and coated with a gold-palladium alloy. Mountedtissues were then viewed and photographed at a specified magnification on aHitachi model S-415A scanning electron microscope set at an acceleration voltageof 15 kV.
The measurement of actual structural leaks was monitored as previouslydescribed (Evans & More, 1988). At the end of each experimental measurement ofammonia effluxes, the heads were perfused for a final 20min with Ringer'ssolution containing 0-018% (2-3xlO~4moll"1) erioglaucine (Acid Blue 9, mol-ecular weight 783 Da; Sigma Chemical Co.). The amount of dye appearing in theirrigate during the efflux period was computed by comparing the absorbance (at632 nm; Beckman model 34 spectrophotometer) of a sample of irrigate with astandard curve prepared by adding various amounts of dye to 100 ml of sea water.
The method described by Claiborne & Evans (1984) was used for measuring theTEP across the perfused head of O. beta. PE 100, 3 % agar-Ringer-filled bridgesconnected the irrigate and the perfusate in the open peritoneal cavity to calomelelectrodes immersed in saturated KC1 solutions. The electrical PD between theelectrodes (perfusate relative to irrigate) was measured on a Keithley model 616digital electrometer. To provide good electrical contact on both sides of thebranchial epithelium, the irrigate outflow from the perfusion chamber was blockedfor a short period to allow the irrigation solution to fill the chamber to the levels ofthe gills. Both perfusion and irrigation pumps were then turned off for a fewseconds, since stable TEPs could not be recorded during fluid flow. Asymmetriesbetween the two bridge tips (sum of calomel electrode differentials and tippotentials in two different salt media) were monitored by placing the bridge tips inbeakers filled with samples of irrigate or perfusate, connected via anotheragar-Ringer bridge. The asymmetries were usually less than 1 mV, stable over thecourse of the experiment, and always subtracted from the TEPs measured acrossthe gills. TEPs were measured when the head was irrigated with sea water andafter the irrigation solution had been changed to either Na+- or CP-free artificialseawater solutions (Table 1). To ensure removal of external Na+ or Cl~ in thesa
Toadfish gill ammonia transport 343
determinations, the heads were irrigated for a few minutes with the appropriateion-free solution before the perfusion chamber was partially filled with thepostbranchial irrigate for the TEP measurement. Standard protocol was tomeasure TEPs sequentially (for approximately 1 min): in SW, Na+-free SW, SW,Cl~-free SW and finally SW. In parallel experiments, the same recording systemwas used to measure the TEPs across intact O. beta, with a PE10, agar-Ringerbridge inserted through a 19 gauge needle into the peritoneal cavity and the otherbridge in the external, seawater or ion-substituted bath. Animals were anesthe-tized (0-01% MS-222 initially, 0-005% during TEP measurement) for theseexperiments and transfers to Na+- or Cr-free solutions were made with a smallhandnet. TEPs across the perfused heads were also monitored in a separate seriesof experiments. These mimicked the protocols outlined below for the ammoniaefflux experiments in order to determine any changes in the TEP produced bythese protocols which might have had an effect on the diffusion of NH4
+ across theperfused head.
Measurement of modes of ammonia transport
Essentially the same protocol as described previously (Evans & More, 1988) forthe perfused head of Squalus acanthias was used in the present study. Unlessindicated, the perfusate in all experiments contained 300-400 ;umol I"1 NH4C1.Initial time-control experiments involved the determination of the consistency ofammonia efflux over four 20-min efflux periods. All other efflux experimentsinvolved an initial 20-min control efflux period, followed by another one or two20-min efflux periods. 10-min wash periods, during which the perfusate andirrigate were changed, separated all efflux periods. The importance of branchialcell deamination in total ammonia efflux was determined by perfusing the headwith ammonia-free, polyvinylpyrrolidone (PVP)-free Ringer's solution, followedby a second efflux period with ammonia added to the perfusate. The putative roleof basolateral Na+/NH4+ exchange in ammonia efflux was measured by followinga control efflux period with a second period in which the head was perfused withRinger containing 10~4 mol I"1 ouabain. The potential involvement of basolateralNa+/NH4+/2C1~ cotransport was monitored in another series of experiments byadding 5xlO~ 5 moir 1 bumetanide to the perfusate during a second efflux period.The potential role of an apical Na+/NH4
+ exchange was examined by adding10~3moir1 amiloride (see Evans & More, 1988) to the irrigate during a secondefflux period. To investigate possible amiloride involvement with basolateralNa+/NH4+ exchange, ouabain was added to the perfusate during a third effluxperiod in these experiments to inhibit basolateral Na+/K+-activated ATPase. Toinvestigate this interaction further, in another series of experiments, amiloride wasadded to the irrigate during a third efflux period, with ouabain in the perfusateduring the second and third efflux periods. Finally, to quantify the relativepermeabilities to NH3 and NH4"1", control efflux periods were followed byabout 10-fold increases in either the perfusate NH3 or NH4"1" concentrations,^cording to the method described previously (Goldstein etal. 1982; Evans &
344 D . H . E V A N S , K. J. M O R E AND S. L. ROBBINS
More, 1988). (NH3 diffusion obviously will be down partial pressure, rather thanconcentration, gradients; however, for comparative purposes we have equatedconcentration with partial pressure, assuming constant solubility of the gas in theexperimental solutions.) In all protocols the ammonia concentration in perfusatesor irrigates from each experimental period was determined by the phenyl-hypo-chlorite method of Solorzano (1969). Similarly, the pH of perfusate and irrigate ineach experiment was determined on a Radiometer PHM84 pH meter, with theelectrode calibrated with buffer in saline solution to approximate more closely theionic concentration of the experimental solutions. The actual NH3 or NH4"1"concentrations of experimental solutions were calculated by the Henderson-Hasselbalch equation, using a pK of 9-35 according to the equations of Cameron &Heisler (1983).
Adrenalin was manufactured by ESI Pharmaceuticals, amiloride and ouabainwere obtained from Sigma Chemical Co., and bumetanide was supplied by LeoPharmaceutical Products.
Experimental results are given as means ± S . E . (N) and statistical differencesbetween experimental means were determined by Student's f-test (using paireddata when appropriate). Calculations of slopes for NH3-stimulated and NH4"1"-stimulated ammonia fluxes, and corrections for changes in the NH3 gradient in thelatter experiments were performed using Multiplan (Microsoft) on an Apple Hecomputer.
Results
Structural integrity of the perfused toadfish head
Photomicrographs of gill filaments, lamellae and interlamellar filamentalsurface cells selected from non-perfused and perfused heads of O. beta are shownin Fig. 1. It is clear that perfusion of the head for over 2h with teleost Ringercontaining 3 % PVP and 10~7moll~1 epinephrine did not result in any noticeablestructural damage.
The erioglaucine leak measured in 35 experiments was only 0-32 ± 0-07 % in a20-min period after over 3h of perfusion of the head preparation, indicatingsubstantial structural integrity, even after this prolonged period of perfusion.
The TEP measured across the toadfish head in sea water was significantly morenegative (about 5 mV) than that monitored in the intact fish in the present study(Fig. 2). Replacement of the irrigate Na+ or Cl~ produced hyperpolarization(about 5mV) or depolarization (about 10mV), respectively, of the same magni-tude as that seen in intact animals, suggesting similar relative Na+ and Cl~permeabilities in vitro and in vivo. When the irrigate was returned to sea waterafter ionic substitutions in the perfused head, the TEP returned to initial levels,indicating that the substitutions themselves did not result in any significantalterations in the relative ionic permeabilities. In the intact animal, the TEP didnot return to seawater levels after Cl~ substitution.
Toadfish gill ammonia transport 345
•W//////////K
Fig. 1. Scanning electron micrographs of gill tissue from Opsanus beta. Filaments andlamellae from (A) non-perfused (scale bar, 400 /an) and (B) isolated, perfused heads(scale bar, 300/an). Individual filament with associated lamellae from (C) non-perfused and (D) perfused heads, respectively (scale bar, 60/an). (E,F) Enlargementsof interlamellar region of filament from C and D, respectively (scale bar, 12 fjm).
346 D. H. EVANS, K. J. MORE AND S. L. ROBBINS
1cu
i
nca
l
u
1 e J
CD
'5.u
ca
H
— I J
-10
-5-
0-
- 5
-10-
-15-
/ / \
^ * • ' ' * * / '
sw -NaSW SW -C1SW sw
Fig. 2. Transepithelial electrical potentials (TEPs) measured across the isolated,perfused head of Opsanus beta and the intact, anesthetized fish. TEPs are expressed asperfusate or blood relative to external bath. —NaSW and — C1SW denote Na+-free andCl~-free artificial sea water (SW), respectively (formulations in Table 1). Dashed line,intact fish (N= 12); solid line, isolated, perfused heads (N = 8).
Baseline, control efflux of ammonia
The perfused toadfish head maintained relatively consistent total ammoniaefflux for periods up to 80min (about 2h total perfusion) (Table 2). During thethird, 20-min period the efflux appeared to decline, but it returned to control levelsduring the fourth period, so we assume that the decline was not physiologicallyimportant. The afferent perfusion pressure also remained consistent during the80min, indicating that the hemodynamics of the branchial vasculature had notdeteriorated during that period.
The role of branchial cell ammoniogenesis
When the toadfish head was perfused with ammonia-free Ringer's solution, the
Table 2. Total ammonia effluxes across the toadfish gill and afferent perfusionpressures during successive 20-min periods
0-20 min 20-40 min 40-60 min 60-80 min
EffluxPressure
15-4 ±2-73-92 ±0-19
12-3±2-2t3-87±0-2f
8-9 ±1-7*3-84±0-21t
ll-2±l-9t3-78±0-2t
* Significantly different (P = 0-01) from period 1, but not significantly different from period 2or period 4.
tNot significantly different from period 1.Values are means ± S.E.M., TV = 8.
Fluxes are measured in /anol 100g~' h"1; pressure in kPa.
Toadfish gill ammonia transport 347
No ammoniain perfusate
300-400/OToll"ammonia in perfusate
Fig. 3. The role of perfusate ammonia in transepithelial ammonia efflux from theisolated, perfused head of Opsanus beta. Each line connects points from a singleperfused head before and after addition of ammonia to the perfusate.
total ammonia efflux was essentially zero. Addition of 300-400 ^anol 1 1 NH4CI tothe perfusate stimulated the efflux substantially (Fig. 3), indicating that, under theconditions of these experiments, branchial cell ammoniogenesis played a vanish-ingly small role in the excretion of ammonia.
The role of basolateral Na+/NH4+ exchange
When 10~4moll~1 ouabain was added to the perfusate, after a control 20-minefflux period, the total efflux of ammonia was reduced by 22% (17-8 ± 1-9/umollOOg"^" 1 vs 22-8±2-1 /xmol 100g^n""1; P<0-01; N=ll). In these exper-iments, the afferent perfusion pressure increased slightly, but significantly(3-33 ± 0-13 vs 4-12 ± 0-13 kPa; P< 0-001) when the perfusate contained ouabain.
The role of basolateral Na+/NH4+/2CI~ cotransport
The addition of 5xlO~5molP1 bumetanide to the perfusate, after a control20-min efflux period, did not significantly affect the total efflux of ammonia(38-1 ± 4-2 vs 35-9 ± 3-2/xmol 100g"1 h"1; N = 6). The afferent perfusion pressurealso was not altered in these experiments (3-19 ± 0-67 kPa in both cases).
When 10~3moll~
The role of apical Na+/NH4+ exchange
I"1 amiloride was added to the irrigate, after a control 20-minefflux period, the ammonia efflux declined from 28-8 ±6-2 to 17-4 ± 4-4 ^mol100g"1 h"1 (P = 0-01; N = 5), suggesting that apical Na+/NH4+ exchange plays ameasurable role in ammonia transport across the toadfish gill. However, when10^4moll~1 ouabain was added to the perfusate during a third period (with|miloride still present in the irrigate) the ammonia efflux did not change
348 D. H. EVANS, K. J. MORE AND S. L. ROBBINS
(16-1 ± 2-6/zmol 100 g *h \ P>0-2) . In these experiments, addition of neitheramiloride nor ouabain had a significant effect on the afferent perfusion pressure(data not shown). When another series of heads was perfused with ouabain in theperfusate during an initial flux period, and amiloride was then added to the irrigateduring the second period (ouabain still present), there was no significant change inthe ammonia efflux (13-8 ±2-2 vs 9-2 ± 2-3/zmoll00g-1ir1; P > 0 - l ; N = 9).Addition of amiloride to the irrigate did not alter afferent perfusion pressures(data not shown), but addition of ouabain increased the pressures slightly(3-72 ± 0-27 vs 4-52 ± 0-27kPa, P < 0-01).
The role of non-ionic diffusion of NH3
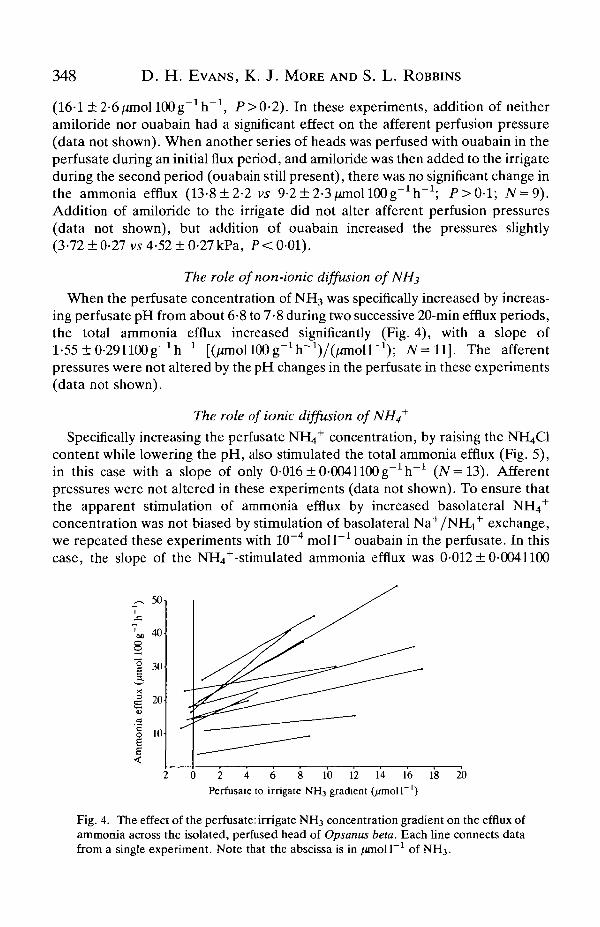
When the perfusate concentration of NH3 was specifically increased by increas-ing perfusate pH from about 6-8 to 7-8 during two successive 20-min efflux periods,the total ammonia efflux increased significantly (Fig. 4), with a slope ofl-55±0-291100g-1h"1 [( /miollOOg^h-VOanoir1) ; AT =11]. The afferentpressures were not altered by the pH changes in the perfusate in these experiments(data not shown).
The role of ionic diffusion of NH4+
Specifically increasing the perfusate NH4+ concentration, by raising the NH4CIcontent while lowering the pH, also stimulated the total ammonia efflux (Fig. 5),in this case with a slope of only 0-016±0-0041100g^h"1 (A/=13). Afferentpressures were not altered in these experiments (data not shown). To ensure thatthe apparent stimulation of ammonia efflux by increased basolateral NH4"1"concentration was not biased by stimulation of basolateral Na+/NH4+ exchange,we repeated these experiments with 10~4 moll"1 ouabain in the perfusate. In thiscase, the slope of the NH4+-stimulated ammonia efflux was 0-012 ±0-0041100
Fig. 4. The effect of the perfusate: irrigate NH3 concentration gradient on the efflux ofammonia across the isolated, perfused head of Opsanus beta. Each line connects datafrom a single experiment. Note that the abscissa is in /anoll"1 of NH3.
Toadfish gill ammonia transport
100
90
^ 80
349
Perfusate to irrigate NR,+ gradient (mmoll ')
Fig. 5. The effect of the perfusate: irrigate NH4"1" concentration gradient on the effluxof ammonia across the isolated, perfused head of Opsanus beta. Each line connectsdata from a single experiment. Note that the abscissa is in mmolF1 of NH4"1".
g ^ h " 1 (N = 26), not significantly different (P>0-4) from the previous exper-iment, indicating that basolateral Na+/NH4+ exchange was not stimulated byraising perfusate NH4+ concentrations. In these experiments, the afferentpressures were increased (P<0-01) from 3-33 ±0-13 to 3-86±0-27kPa by theaddition of ouabain to the perfusate.
The effect of changes in the transepithelial electrical potentials
Measurement of the TEP in a separate series of experiments, which combinedthe perfusate and irrigate manipulations outlined above, demonstrated that thealterations in the electrical potentials across the gills were minor (<2 mV) in allcases (data not shown).
Discussion
Structural integrity of the perfused toadfish head preparation
Perfusion of the toadfish head for periods exceeding 2 h did not produce anyobvious gill surface abnormalities (Fig. 1), in contrast to the obvious edemaproduced in the eel {Anguilla australis) holobranch (Ellis & Smith, 1983) or therainbow trout (Salmo gairdneri) head (Part et al. 1982) perfused for much shorterPeriods with adrenalin-free Ringer's solution.
350 D . H . EVANS, K. J . M O R E AND S. L. ROBBINS
The extremely small leak (0-3%) in this preparation is even less than thatdescribed for the perfused dogfish shark pup head preparation (0-7%; Evans &More, 1988), and demonstrates directly that this preparation is structurally sound.If we assume that the actual gill permeability to erioglaucine is zero (taking theaverage perfusate ammonia concentration as approximately 400//moll"1, theperfusion flow as650|zlmin~1, and an animal mass of 30 g), an erioglaucine leak of0-3% for 20min would produce an apparent ammonia efflux of only0-15 /imol 100g^h" 1 , approximately 1 % of that measured in the ammoniaexperiments described below.
Afferent perfusion pressures, assumed to be proportional to branchial andcephalic resistance, were stable in this preparation for at least 80min (Table 2),indicating that, like the perfused dogfish pup head (Evans & More, 1988), thetoadfish head is hemodynamically patent for the entire length of the experimentsdescribed here. The afferent pressures monitored in the present experiments werevery similar to those measured, under control conditions, (2-79 and 4-12 kPa) in anearlier series of experiments on the toadfish perfused head (Claiborne et al. 1982).
The transepithelial electrical potential (—5 mV) measured across the toadfishhead irrigated with sea water is significantly more negative than that measuredacross intact, anesthetized fish in a parallel series of experiments (Fig. 2),suggesting relatively higher C P conductances in vitro. However, it is quite close tothe seawater TEPs measured in the same species in other in vivo studies ( - 8 mV:Evans & Cooper, 1975; — 6mV: Kormanik & Evans, 1979; -9mV: Kormanik &Evans, 1982). Irrigation with Na+-free artificial seawater solution hyperpolarizedthe TEP by some 5 mV in both the perfused head and intact fish. Slightly higherhyperpolarizations (about 7 mV) were observed in previous, in vivo studies (Evans& Cooper, 1975; Kormanik & Evans, 1982). Replacement of the irrigate C P withbenzenesulfonate depolarized the TEP across the perfused head and intact fish bysome 10 mV, significantly less than the degree of depolarization (20 mV) caused byC P substitution in an earlier in vivo study (Kormanik & Evans, 1979). It isimportant to note that we observed that the TEPs were more stable and morerapidly attained after ionic substitutions using the perfused head than in the intactfish in the present experiments. For example, after C P substitution and transferback into sea water, the TEP across the perfused head approached that measuredpreviously in sea water, whereas that across the intact fish was extremely variable,slow to stabilize (data not shown) and 7 mV depolarized from the initial seawaterTEP (Fig. 2). One might suggest, therefore, that placement of the internal bridgein the postbranchial perfusate of the perfused head provides a more accuratemeasurement of the true TEP than implantation of a bridge in the muscle mass orperitoneal cavity of an intact, anesthetized fish. Most importantly, the present datademonstrate that reproducible TEPs, of the same order as in vivo TEPs, can bemeasured across the perfused head of O. beta. Therefore, changes in passive ionicfluxes produced by alteration of electrochemical gradients can be monitored andfactored out in any subsequent studies of ion transport by this preparation.
Taken together, these data show clearly that the perfused head of O.
Toadfish gill ammonia transport 351
maintains structural, hemodynamic and electrical integrity, even after prolonged(1-2 h) perfusion, when compared with the intact fish.
Baseline, control efflux of ammonia
The control efflux of ammonia from the perfused head in these experiments(Table 2) was somewhat below the 26//moll00g~1hT1 described for the intact,normocapnic toadfish in one earlier experiment (Evans, 1982), but equal to thatfound in a similar study (15 junol 100 g"1 h"1; Evans, 1977) in hypercapnic toadfish(ll^moll00g~1h~1; Evans, 1982) and in an earlier study of ammonia transportacross the perfused toadfish head (16/«noll00g~1h~1; Claiborne et al. 1982). It isclear, therefore, that the preparation maintains near in vivo rates of ammoniatransport. In addition, the relative consistency over the course of the currentexperiments (Table 2) supports, again, the patency of this preparation.
The role of branchial cell ammoniogenesis
We found that ammoniogenesis by the branchial epithelium plays no measure-able role in the ammonia excretion by the perfused toadfish head (Fig. 3). Thequantitative importance of branchial cell ammoniogenesis in the total gillextrusion of ammonia by fishes has rarely been investigated. Pequin (1962) foundthat extraction of blood ammonia could account for all the ammonia excretionacross the intact carp (Cyprinus carpio) and Goldstein et al. (1964) found similarresults with the marine sculpin Myoxocephalus scorpius. However, Payan (1978)demonstrated that some 5 % of the ammonia excretion by the perfused trout headcould be accounted for by branchial cell production, and Cameron & Heisler(1983) calculated similar levels using arteriovenous differences in intact trout.
The role of basolateral Na+/NH4+ exchange and Na+/NH4
+/2Cr cotransport
The observation that addition of ouabain to the perfusate reduced the rate ofammonia efflux by 22 % corroborated our earlier study (Claiborne et al. 1982) ofO. beta, which demonstrated that ammonia efflux from the perfused head wassensitive to perfusate ouabain and K+ concentrations, and suggests again thatbasolateral Na+/NH4+ exchange plays a measureable role in transporting am-monia across the toadfish branchial epithelium. An alternative explanation ofthese results is that inhibition of basolateral Na+/K+-activated ATPase byouabain indirectly inhibits a basolateral Na+/K+/2C1~ cotransporter. Thisbumetanide-sensitive carrier has been shown to be sensitive to NH44" (Kinne et al.1986; O'Grady et al. 1987) and to play a role in N H / reabsorption in the thickascending limb of the rabbit nephron (Garvin et al. 1988; Good, 1988). Our earlierstudy of ammonia transport across the dogfish pup gill epithelium demonstratedthat a small, but measureable, ammonia efflux took place via a bumetanide-sensitive system (Evans & More, 1988). However, in the present experiments,bumetanide addition to the perfusate did not inhibit ammonia efflux (see Results),consistent with the conclusion that the ouabain effect was directly on basolateral
4+ exchange.
352 D. H. EVANS, K. J. MORE AND S. L. ROBBINS
T50_cToo8 40"og~ 30X
•2 20o
E< 10-
10 20 30 40 50Afferent perfusion pressure (kPa)
60
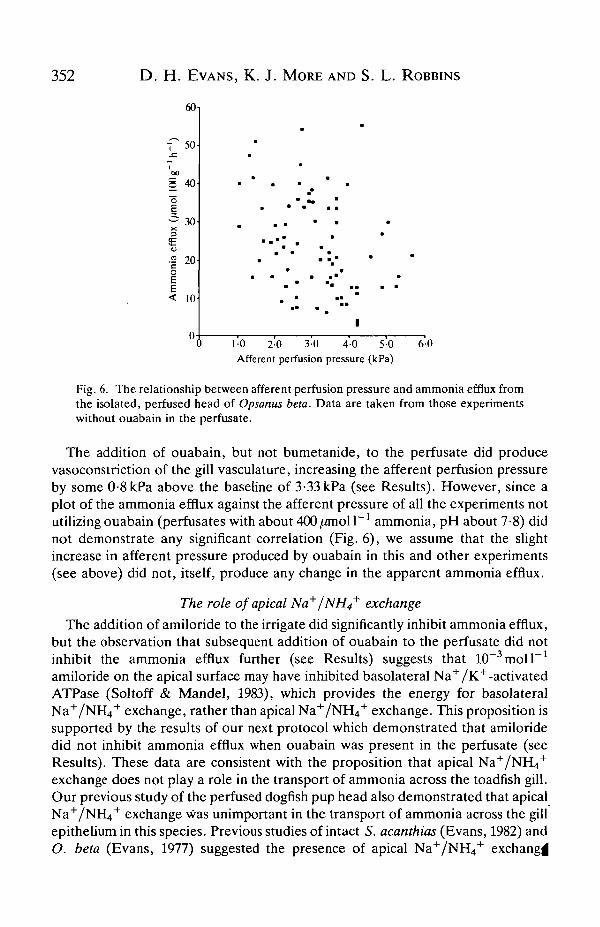
Fig. 6. The relationship between afferent perfusion pressure and ammonia efflux fromthe isolated, perfused head of Opsanus beta. Data are taken from those experimentswithout ouabain in the perfusate.
The addition of ouabain, but not bumetanide, to the perfusate did producevasoconstriction of the gill vasculature, increasing the afferent perfusion pressureby some 0-8kPa above the baseline of 3-33 kPa (see Results). However, since aplot of the ammonia efflux against the afferent pressure of all the experiments notutilizing ouabain (perfusates with about 400//moll"1 ammonia, pH about 7-8) didnot demonstrate any significant correlation (Fig. 6), we assume that the slightincrease in afferent pressure produced by ouabain in this and other experiments(see above) did not, itself, produce any change in the apparent ammonia efflux.
The role of apical Na+/ NH4+ exchange
The addition of amiloride to the irrigate did significantly inhibit ammonia efflux,but the observation that subsequent addition of ouabain to the perfusate did notinhibit the ammonia efflux further (see Results) suggests that 10~3molP1
amiloride on the apical surface may have inhibited basolateral Na+/K+-activatedATPase (Soltoff & Mandel, 1983), which provides the energy for basolateralNa+/NH4+ exchange, rather than apical Na+/NH4+ exchange. This proposition issupported by the results of our next protocol which demonstrated that amiloridedid not inhibit ammonia efflux when ouabain was present in the perfusate (seeResults). These data are consistent with the proposition that apical Na+/NH4+
exchange does not play a role in the transport of ammonia across the toadfish gill.Our previous study of the perfused dogfish pup head also demonstrated that apicalNa+/NH4+ exchange was unimportant in the transport of ammonia across the gillepithelium in this species. Previous studies of intact 5. acanthias (Evans, 1982) andO. beta (Evans, 1977) suggested the presence of apical Na+/NH4+ exchange
Toadfish gill ammonia transport 353
because, in both species, removal of external Na+ reduced ammonia efflux.However, this could have been due to cessation of apical Na + /H + exchange (inboth species the H + efflux was reduced to at least zero), which would have affectedthe NH3 gradient, and thereby the efflux of ammonia (Evans & Cameron, 1986).In addition, removal of external Na+ could have affected paracellular NH4"1"conductance (Zadunaisky, 1984; Evans & Cameron, 1986).
The roles of non-ionic diffusion of NH3 and ionic diffusion of NH4+
The data in Figs 4 and 5 demonstrate that the ammonia efflux across the gillepithelium of the toadfish is stimulated by an increase in either the NH3 or theNFLi"1" concentration of the perfusate. The stimulation by both NFLj"1" and NH3 issomewhat at odds with earlier data (Goldstein et al. 1982) which appeared todemonstrate that only changes in perfusate NFLi"1" stimulated ammonia effluxacross O. beta gills, suggesting a zero permeability to NH3 However, in the earlierstudy, specific NH3 gradients were not monitored, and it is unlikely that anyepithelium is totally impermeable to NH3. Our finding that ammonia can traversethe toadfish gill epithelium by ionic diffusion of NH4
+ confirms the earlier study(Goldstein et al. 1982), as well as our study of the dogfish pup gill (Evans & More,1988), a recent study of the intact, marine teleost, Myoxocephalus octadecimspino-sus (Claiborne & Evans, 1988) and studies of the turtle bladder (Schwartz &Tripolone, 1983) and the rabbit renal proximal straight tubule (Garvin et al. 1987).
The slopes in Figs 4 and 5 indicate that the toadfish gill's permeability to NH3 isnearly 100 times (97:1) the NH4
+ permeability. In our previous study wedetermined that the relative permeabilities were 1100:1 in the gill of S. acanthias(Evans & More, 1988). Using a gill surface area of 2-04 cm2 g"1 (taken from dataon a congeneric, O. tau, of similar mass; Hughes & Gray, 1972), we can calculatethe apparent NH3 and NH4"1" permeabilities of the toadfish gill epithelium andcompare them with those published for other epithelia (Table 3). It is clear that thetoadfish gill maintains relatively high permeabilities to both NH3 and NH4"1", in therange of those described for the 'leaky' proximal tubule of the mammaliannephron, but much greater than those described for either turtle bladder or sharkgill, both considered to be relatively 'tight' epithelia (Erlij & Martinez-Palomo,1978; Evans, 1979). In particular, the relatively high permeability to NH4"1" of the
Table 3. Apparent NH3 and NH4+ permeabilities of epithelial tissues
Tissue
Turtle bladderTurtle bladderRabbit proximal
straight tubuleShark pup gillToadfish gill
All permeabilities are in
NH3
26072
16000
4202150
N T 6 c m s " 1 .
NH4"1"
4.9
4-5450
0-422
Reference
Schwartz & Tripolone (1983)Arruda et al. (1984)Garvin et al. (1987)
Evans & More (1988)Present study
354 D . H . E V A N S , K. J. MORE AND S. L. ROBBINS
toadfish gill correlates with the relatively high cationic permeability found in thegills of a variety of marine teleost fishes (Evans, 1979). In fact, the Na+ efflux fromthe toadfish gill is relatively low compared with that from the gills of other marineteleosts (Evans, 1979), suggesting that NH4"1" diffusion may be even moresubstantial across the gills of other marine teleosts. Since the gills of fresh- orbrackish-water teleosts appear to have substantially lower ionic permeabilities(Evans, 1979), one could suggest that diffusion of NH4"1" might also be reduced inthese species. In fact, our preliminary studies on perfused heads of O. betaacclimated to 5 % sea water have demonstrated that the slope of the NH3-stimulated ammonia efflux is 3-67 ±0-691100g^h"1 (TV =6) and the slope of theNH4"1"-stimulated ammonia efflux (with 10~4moll~1 ouabain in the perfusate toblock putative basolateral exchanges) is 0-003 ±0-0011100 g ^ h " 1 (N=6),suggesting that NH3 permeability significantly increased, while NH4"1" permeabilityfell, concomitant with acclimation to reduced salinity. In fact, the ratio ofpermeabilities was 1200:1, close to that described for the dogfish shark pup (Evans& More, 1988), and the cationic permeability of elasmobranchs is of the sameorder as that of freshwater teleosts (Evans, 1979).
These data support the proposition that the gill epithelium of a marine teleost issignificantly more permeable to NH4"1" than the gill of either a marine elasmo-branch or the same teleost species acclimated to extremely low salinities. Thisincreased permeability to the ionic form of ammonia may be correlated with therelatively higher cationic permeability of teleosts in sea water, compared withmarine elasmobranchs or teleosts in reduced salinities (Evans, 1979).
Relative roles of various transport pathways
Since separate measurements of the transepithelial electrical potential (duringmanipulations of the perfusate and/or irrigate as outlined above) indicated thatchanges in the TEP were minimal (see Results), we can utilize the data justdescribed to calculate the relative importance of various putative pathways forammonia transport across the toadfish gill epithelium. Assuming an ammonia pKof 9-35 for teleost Ringer's solution at 24°C (Cameron & Heisler, 1983) and the pHof the perfusate (7-8), the concentration ratio of the two species of ammonia is0-028:1-0 (NH3:NH4+). Since the actual relative efflux is the product of therelative permeability and the relative concentration, the actual ratio of the effluxesof NH3: NH4
+ across the gill is 2-7:1, despite the nearly 100-fold difference in theirrespective permeances across the epithelium. Given that 22% of the totalammonia efflux is ouabain-sensitive, 78% has to be via diffusion of NH3 andNH4"1". Fig. 7 summarizes these calculations and demonstrates that, despite thefact that over 50% of the total ammonia efflux is via non-ionic diffusion, ionicdiffusion of NH4"1" and basolateral Na+/NH4+ exchange play substantial roles inthe transport of ammonia across the gill epithelium of the toadfish. In contrast, ourearlier study of ammonia transport by the dogfish shark pup gill epithelium (Evans& More, 1988) demonstrated that basolateral Na+/NI-Li+ exchange did not play ameasureable role, basolateral Na + /NH4 + /2Cr transport could account for 17 °/4
Toadfish gill ammonia transport 355
Serosal Mucosal
Ouabain »-(22%l
Bumetanide Amilonde
Fig. 7. Summary of the relative roles played by various putative modes for transport ofNH3 and NH4"1" across the isolated, perfused head of the marine teleost fish Opsanusbeta. Redrawn and modified from Evans & Cameron (1986).
of the efflux, and diffusion of NH3 was 12 times greater than NH4"1" diffusion.These are the only direct, quantitative measurements of the role of variousputative ammonia transport pathways across the fish gill epithelium. However, it isapparent that the importance of various pathways may vary with species, and thatthe relative roles of NH3 and NH4"1" diffusion may be correlated with the relativeionic permeabilities of the gill epithelium (Evans, 1979).
This research was supported by NSF PCM 8302621 to DHE. Figures were drawnby Daryl Harrison.
ReferencesARUDDA, J. A. L., DYTKO, G. & WITHERS, L. (1984). Ammonia transport by the turtle urinary
bladder. Am. J. Physiol. 246, F635-F647.CAMERON, J. N. & HEISLER, N. (1983). Studies of ammonia in the rainbow trout: physico-
chemical parameters, acid-base behaviour and respiratory clearance. J. exp. Biol. 105,107-125.
CLAIBORNE, J. B. & EVANS, D. H. (1980). The isolated, perfused head of the marine teleost fish,Myoxocephalus octodecimspinosus: Hemodynamic effects of epinephrine. J. comp. Physiol.138, 79-85.
CLAJBORNE, J. B. & EVANS, D. H. (1984). Transepithelial potential measurements in theisolated, perfused head of a marine teleost. /. exp. Zool. 230, 321-324.
CLAIBORNE, J. B. & EVANS, D. H. (1988). Ammonia and acid-base balance during highammonia exposure in a marine teleost {Myoxocephalus octodecimspinosus). J. exp. Biol. 140,89-105.
CLAIBORNE, J. B., EVANS, D. H. & GOLDSTEIN, L. (1982). Fish branchial Na+/NFLi+ exchange isvia basolateral Na,K-activated ATPase. /. exp. Biol. 96, 431-434.
ELLIS, A. G. & SMITH, D. G. (1983). Edema formation and impaired O2 transfer in Ringer-perfused gills of the eel, Anguilla anguilla. J. exp. Zool. 227, 371-380.
|ERLU, D. & MARTINEZ-PALOMO, A. (1978). Role of tight junctions in epithelial function. In
356 D. H. EVANS, K. J. MORE AND S. L. ROBBINS
Membrane Transport in Biology, vol. Ill, Transport Across Multimembrane Systems (ed.G. Giebisch, D. C. Tosteson & H. H. Ussing), pp. 27-53. Berlin: Springer-Verlag.
EVANS, D. H. (1977). Further evidence for Na+/NH4+ exchange in marine teleost fish. J. exp.Biol. 70, 213-220.
EVANS, D. H. (1979). Fish. In Comparative Physiology of Osmoregulation in Animals (ed.G. M. O. Maloiy), pp. 305-390. New York: Academic Press.
EVANS, D. H. (1982). Mechanisms of acid secretion by two marine fishes: the teleost, Opsanusbeta, and the elasmobranch, Squalus acanthias. J. exp. Biol. 97, 289-299.
EVANS, D. H. & CAMERON, J. N. (1986). Gill ammonia transport. J. exp. Zool. 239, 17-23.EVANS, D. H. & COOPER, K. (1975). The presence of Na-Na and Na-K exchange in sodium
excretion by three species of fish. Nature, Lond. 259, 241-242.EVANS, D. H. & MORE, K. (1988). Modes of ammonia transport across the gill epithelium of the
dogfish pup (Squalus acanthias). J. exp. Biol. 138, 375-397.GARVIN, J. L., BURG, M. B. & KNEPPER, M. A. (1987). NH3 and NH,"1" transport by rabbit renal
proximal straight tubules. Am. J. Physiol. 252, F232-F239.GARVIN, J. L., BURG, M. B. & KNEPPER, M. A. (1988). Active NH4"1" absorption by the thick
ascending limb. Am. J. Physiol. 255, F57-F65.GOLDSTEIN, L., CLAIBORNE, J. B. & EVANS, D. H. (1982). Ammonia excretion by the gills of two
marine teleost fish: The importance of NFL,+ permeance. /. exp. Zool. 291, 395-397.GOLDSTEIN, L., FORSTER, R. P. & FANELLI, G. M., JR (1964). Gill blood flow and ammonia
excretion in the marine teleost, Myoxocephalus scorpius. Comp. Biochem. Physiol. 12,489-499.
GOOD, D. W. (1988). Active absorption of NFL(+ by rat medullary thick ascending limb:inhibition by potassium. Am. J. Physiol. 255, F78-F87.
HUGHES, G. M. & GRAY, I. E. (1972). Dimensions and ultrastructure of toadfish gills. Biol. Bull.mar. biol. Lab., Woods Hole 143, 150-161.
KINNE, R., KOENIG, B., HANNAFIN, J., KINNE-SAFFRAN, J. E., SCOTT, D. M. & ZIEROLD, K.(1986). The use of membrane vesicles to study the NaCl/KCl cotransporter involved in activetransepithelial chloride transport. Pfliigers Arch, ges. Physiol. 405 (Suppl. 1), S101-S105.
KORMANIK, G. A. & EVANS, D. H. (1979). HCO3-stimulated Cl efflux in the gulf toadfishacclimated to sea water. /. exp. Zool 208, 13-16.
KORMANIK, G. A. & EVANS, D. H. (1982). The relation of Na and Cl extrusion in Opsanus beta,the gulf toadfish, acclimated to seawater. J. exp. Zool. 224, 187-194.
ODULEYE, S. O. & EVANS, D. H. (1982). The isolated, perfused head of the toadfish, Opsanusbeta. II. Effects of vasoactive drugs on unidirectional water flux. /. comp. Physiol. 149,115-120.
O'GRADY, S. M., PALFREY, H. C. & FIELD, M. (1987). Characteristics and functions of Na-K-Clco-transport in epithelial tissues. Am. J. Physiol. 253, C177-C192.
PART, P., TUURALA, H. & SOIVIO, A. (1982). Oxygen transfer, gill resistance and structuralchanges in rainbow trout (Salmo gairdneri, Richardson) gills perfused with vasoactive agents.Comp. Biochem. Physiol. 71C, 7-13.
PAYAN, P. (1978). A study of the Na+/NFl4+ exchange across the gill of the perfused head of thetrout (Salmo gairdneri). J. comp. Physiol. 124, 181-188.
PAYAN, P. & MATTY, A. J. (1975). The characteristics of ammonia excretion by a perfusedisolated head of a trout (Salmo gairdneri): Effect of temperature and CO2-free Ringer.J. comp. Physiol. 96, 167-184.
PEQUIN, L. (1962). Les teneursen azote ammoniacal du sangchez la Carpe (Cyprinus carpio L.).C.R. hibd. Seanc. acad. ScL, Paris 255, 1795-1797.
SCHWARTZ, J. H. & TRIPOLONE, M. (1983). Characteristics of NH "1" and NH3 transport acrossthe isolated turtle urinary bladder. Am. J. Physiol. 245, F210-F216.
SOLORZANO, L. (1969). Determination of ammonia in natural waters by the phenylhypochloritemethod. Limnol. Oceanogr. 14, 799-801.
SOLTOFF, S. P. & MANDEL, L. J. (1983). Amiloride directly inibits the Na,K-ATPase activity ofrabbit kidney proximal tubules. Science 220, 957-959.
ZADUNAISKY, J. A. (1984). The chloride cell: The active transport of chloride and theparacellular upathways. In Fish Physiology, vol. XB (ed. W. S. Hoar & D. J. Randall),pp. 129-176. Orlando: Academic Press.