Modulation of Tissue Inflammatory Response by Histamine Receptors in Scorpion Envenomation Pathogenesis: Involvement of H4 Receptor Amal Lamraoui, 1 Sonia Adi-Bessalem, 1 and Fatima Laraba-Djebari 1,2 Abstract—The inflammatory response caused by scorpion venoms is a key event in the pathogenesis of scorpion envenomation. This response was assessed in the cardiac, pulmonary, and gastric tissues of envenomed mice. The results reveal an increase of permeability in cardiac, pulmonary, and gastric vessels accompanied by an edema-forming, inflammatory cell infiltration, and imbalanced redox status. These effects are correlated with severe tissue alterations and concomitant increase of metabolic enzy- mes in sera. Pretreatment of mice with antagonists of H1, H2, or H4 receptors markedly alleviated these alterations in the heart and lungs. Nevertheless, the blockade of the H3 receptor slightly reduced these disorders. Histamine H2 and H4 receptors were the most pharmacological targets involved in the gastric oxidative inflammation. These findings could help to better understand the role of histamine in scorpion venom-induced inflammatory response and propose new therapy using as targets the H4 receptor in a- ddition to histamine H1 and H2 receptors to attenuate the induced inflammatory disorders encountered in scorpion envenoming. KEY WORDS: scorpion venom; inflammation response; oxidative stress; tissue injury; histamine receptors. INTRODUCTION Scorpion envenomation affected yearly many people in several tropical and subtropical countries leading to human fatalities [1, 2]. The scorpion venom neurotoxic compounds represent minor but most toxic fraction [3]. The high affinity binding of neurotoxins on voltage-gated sodium channels of excitable cells leads to the excessive release of neurotransmitters [1, 4], resulting in many path- ological disorders such as myocardiac damage, pulmonary edema, and respiratory failure [5–8]. The production and release of inflammatory mediators, such as cytokines, eicosanoids, and bioactive amines from activated immune cells, have been also reported in several cases of scorpion envenoming [6, 7, 9–17]. Moreover, increased vascular permeability is an early event in envenoming syndrome [18–20]. Because of its vasoactive properties, histamine could have a pivotal role in this process [21]. This bioactive amine is released from large variety of cell types mainly mast cells, basophiles, gastric enterochro- maffin-like cells, and histaminergic neurons, but also supplied by other cell varieties, for example, dendritic cells and lym- phocytes [22, 23]. Aside from its ability to increase vascular permeability, it promotes smooth muscle contraction, gastro- intestinal acid secretion, neurotransmission, inflam- matory cell recruitment, and activation of cytokine network, by stimulating different target cell receptors, H1, H2, H3, and H4 [24, 25]. Although histamine is well known for its in- volvement during scorpion envenomation pathogene- sis [26, 27], little is known about its role on inflam- matory response induced by scorpion venoms. There- fore, this study was designed to elucidate the effect of histamine through its receptors in the Aah venom- induced cardiac, pulmonary, and gastric acute inflam- mation by blocking its pharmacological targets. In- flammatory events were assessed by measuring vascu- lar permeability changes, edema formation, and some 1 Faculty of Biological Sciences, Laboratory Cellular and Molecular Biology, Department Cellular and Molecular Biology, USTHB, BP32, EL Alia, Bab Ezzouar, 16111 Algiers, Algeria 2 To whom correspondence should be addressed at Faculty of Biological Sciences, Laboratory Cellular and Molecular Biology, Department Cellular and Molecular Biology, USTHB, BP32, EL Alia, Bab Ezzouar, 16111 Algiers, Algeria. E-mail: [email protected]0360-3997/14/0000-0001/0 # 2014 Springer Science+Business Media New York Inflammation ( # 2014) DOI: 10.1007/s10753-014-9898-x
Transcript
Modulation of Tissue Inflammatory Response by HistamineReceptors in Scorpion Envenomation Pathogenesis:Involvement of H4 Receptor
Amal Lamraoui,1 Sonia Adi-Bessalem,1 and Fatima Laraba-Djebari1,2
Abstract—The inflammatory response caused by scorpion venoms is a key event in the pathogenesis ofscorpion envenomation. This response was assessed in the cardiac, pulmonary, and gastric tissues ofenvenomed mice. The results reveal an increase of permeability in cardiac, pulmonary, and gastricvessels accompanied by an edema-forming, inflammatory cell infiltration, and imbalanced redox status.These effects are correlated with severe tissue alterations and concomitant increase of metabolic enzy-mes in sera. Pretreatment of mice with antagonists of H1, H2, or H4 receptors markedly alleviated thesealterations in the heart and lungs. Nevertheless, the blockade of the H3 receptor slightly reduced thesedisorders. Histamine H2 and H4 receptors were the most pharmacological targets involved in the gastricoxidative inflammation. These findings could help to better understand the role of histamine in scorpionvenom-induced inflammatory response and propose new therapy using as targets the H4 receptor in a-ddition to histamine H1 and H2 receptors to attenuate the induced inflammatory disorders encounteredin scorpion envenoming.
Scorpion envenomation affected yearly many peoplein several tropical and subtropical countries leading tohuman fatalities [1, 2]. The scorpion venom neurotoxiccompounds represent minor but most toxic fraction [3].The high affinity binding of neurotoxins on voltage-gatedsodium channels of excitable cells leads to the excessiverelease of neurotransmitters [1, 4], resulting in many path-ological disorders such as myocardiac damage, pulmonaryedema, and respiratory failure [5–8].
The production and release of inflammatory mediators,such as cytokines, eicosanoids, and bioactive amines fromactivated immune cells, have been also reported in severalcases of scorpion envenoming [6, 7, 9–17]. Moreover,
increased vascular permeability is an early event inenvenoming syndrome [18–20]. Because of its vasoactiveproperties, histamine could have a pivotal role in this process[21]. This bioactive amine is released from large variety ofcell types mainly mast cells, basophiles, gastric enterochro-maffin-like cells, and histaminergic neurons, but also suppliedby other cell varieties, for example, dendritic cells and lym-phocytes [22, 23]. Aside from its ability to increase vascularpermeability, it promotes smooth muscle contraction, gastro-intestinal acid secretion, neurotransmission, inflam-matory cell recruitment, and activation of cytokinenetwork, by stimulating different target cell receptors,H1, H2, H3, and H4 [24, 25].
Although histamine is well known for its in-volvement during scorpion envenomation pathogene-sis [26, 27], little is known about its role on inflam-matory response induced by scorpion venoms. There-fore, this study was designed to elucidate the effect ofhistamine through its receptors in the Aah venom-induced cardiac, pulmonary, and gastric acute inflam-mation by blocking its pharmacological targets. In-flammatory events were assessed by measuring vascu-lar permeability changes, edema formation, and some
1 Faculty of Biological Sciences, Laboratory Cellular and MolecularBiology, Department Cellular and Molecular Biology, USTHB,BP32, EL Alia, Bab Ezzouar, 16111 Algiers, Algeria
2 To whom correspondence should be addressed at Faculty of BiologicalSciences, Laboratory Cellular and Molecular Biology, DepartmentCellular and Molecular Biology, USTHB, BP32, EL Alia, BabEzzouar, 16111 Algiers, Algeria. E-mail: [email protected]
0360-3997/14/0000-0001/0 # 2014 Springer Science+Business Media New York
markers of the influx of inflammatory cells to thetissues and the oxidative stress status. Furthermore,this study was supported by histological analysis andassessment of enzymes (CPK and LDH) activitiesreflecting tissue injuries.
MATERIALS AND METHODS
Materials
Venom
Crude venom of Androctonus australis hector (Aah)scorpion was provided by the Pasteur Institute and waskept at −20 °C until use (0.5 mg/kg) for injection.
Chemical Reagents
Chemical reagents used in this study were of analyticgrade.
Cetirizine dihydrochloride, cimetidine, and Betasercwere from Geo-Pharm (Algeria), Eurogenerics (Belgium),and Solvay Pharma (France), respectively. JNJ-7777120,o -d i an i s id ine hydroch lo r ide , N- (1 -nap thy l ) -ethylenediamine dihydrochloride, sulfanilamide, thiobar-bituric acid, trichloroacetic acid, acetic acid, andsulfosalicylic acid were purchased from Sigma-Aldrich(St Louis MO, USA). Evans Blue and formamide werefrom Merck (Darmstadt, F.R.G).
Animals
Animals were obtained from the animal breed-ing of Biological Sciences Faculty of the Universityof Sciences and Technology Houari Boumediene(Algeria) and kept under controlled conditions ofhumidity, lighting, and temperature throughout theexperiments with free access to standard rodent dietand tap water.
This study was conducted with N’MRI miceweighing 18 to 22 g. All animal experiments were inaccordance with the actual guidelines for the care of ani-mal’s laboratory.
Methods
In Vivo Protocol
Mice were divided into seven groups of 20animals each. Untreated mice (control groups) subcu-taneously received saline solution (NaCl 0.9 % (w/v))or dimethyl sulfoxide (DMSO 65 % (v/v in sterile
distilled water)). Mice of the third group receivedsubcutaneous injection of sublethal dose of Aah ven-om (0.5 mg/kg), whereas animals of groups 4, 5, and6 were pretreated intraperitoneally before envenom-ation with selective antagonist of histamine H1, H2,and H3 receptors: cetirizine (10 mg/kg; 1 h beforethe venom), cimetidine (15 mg/kg; 2 h before thevenom), and Betaserc (20 mg/kg; 1 h before thevenom), respectively. The seventh group includedmice pretreated with the selective histamine H4 re-ceptor antagonist, JNJ-7777120, which was givensubcutaneously at the dose of 10 mg/kg, 30 minbefore the experimental envenomation. All thesedrugs were dissolved in saline solution (NaCl 0.9 %(w/v)) except JNJ-7777120, which was dissolved inDMSO (65 % (v/v)). Control and envenomed micewere humanly sacrificed within 24 h after the injec-tion of saline, DMSO, or venom (Fig. 1). Blood andorgans were collected for further analysis.
Measurement of Vascular Permeability Changes
The extravasation of Evans blue from the vascularcompartment to the heart, lungs, and stomach was used asan indicator of cardiac, pulmonary, and gastric vascularpermeability increase because it has a high affinity forplasma proteins with which it binds in the first few secondsafter its injection [28].
Evans blue was administered intravenously intothe mice at a dose of 20 mg/kg (120 μl/20 g bodyweight) few seconds before the injection of the ven-om or the vehicles (saline or DMSO). Mice receivingthe Evans blue were sacrificed after 24 h. Organs(heart, lungs, and stomach) were recovered, weighed,and placed in 2 ml of formamide, an organic solventrequired for extraction of Evans blue. They wereincubated at 37 °C for 48 h. Absorbances were readat a wavelength of 620 nm. The concentration ofEvans blue was determined after extrapolation on astandard curve conducted with various concentrationsof Evans blue. The results were expressed as Evansblue concentration per microgram of tissue.
Organs-to-Body Weight Ratio
The heart, lungs, or stomach/body weight indexeswere calculated by dividing each organ mass per total bodyweight×100. This indicated cardiac, pulmonary, and gas-tric water content.
Lamraoui, Adi-Bessalem, and Laraba-Djebari
Evaluation of Inflammatory Cell Infiltration
Myeloperoxidase Activity (MPO) Assay. Myeloperoxidase(MPO) activity was measured as an index of neutrophilsequestration into the heart, lungs, and stomach. Theseorgans recovered from sacrificed mice were weighed andhomogenized (1/10 (w/v)) in physiological saline solution(NaCl 0.9 % (w/v)) using a polytron homogenizer (T25Utra-turax). They were then centrifuged at 2,486×g for20 min. The pellets underwent three cycles of freezing–thawing, 15 min each. They were then resuspended (1/8(w/v)) in saline solution (NaCl 0.9% (w/v). The supernatantswere collected by centrifugation at 2,486×g for 20 min.Phosphate buffer (1 ml at 50 mM; pH 6.6) containingortho-dianisidine (0.167 mg/ml) and H2O2 (30 %; v/v))was added to each supernatant (1 ml). Changes in opticaldensities were measured at 460 nm. Results were expressedas millimolar of H2O2 transformed per minute and per100 mg of tissue, using a molar extinction coefficient of11.3 mM−1 cm−1.
Eosinophil Peroxidase Activity Assay. Eosinophilsequestration and accumulation in the heart, lungs, andstomach were assessed using the measurement ofeosinophil peroxidase activity, which was performed asfollows: Organs (heart, lungs, and stomach) after animalswere sacrificed were recovered, weighed, andhomogenized (1/10 (w/v)) in physiological saline (NaCl0.9 % (w/v). They were centrifuged at 2,486×g for 20 min.Obtained supernatants were distributed in microplate
(50 μl/well), followed by an addition of 100 μl/well ofchromogenic substrate solution (20 mg OPD dissolved in10ml Tris–HCl 0.05M, pH 8, and 10μl of H2O2 at 30%v/v). The plate was then incubated for 1 h away from thelight. Absorbance readings were performed using amicroplate reader at 490 nm. The results were expressedas change in absorbance per 100 mg of tissue.
Oxidative Stress Marker Assays
Nitric oxide and malondialdehyde (MDA) levels, aswell as catalase activity, were measured in tissue superna-tants obtained as those used for EPO activity assay.
Nitric Oxide Content. Total nitric oxide levels weredetermined by measuring its stable products (nitrites)according to Sun et al. [29]. Briefly, 500 μl oftrichloroacetic acid was added to the same volume oftissue supernatant. This mixture was centrifuged at1,466×g for 10 min for protein precipitation. Obtainedsupernatants (500 μl) were placed in contact with 500 μlof Griess reagent. Absorbance was read at 543 nm after20 min of incubation in dark. Nitrite levels weredetermined after extrapolation on a standard curve ofKNO previously established. Results are expressed inmicromolat per gram of tissue.
Estimation of Lipid Peroxidation. Lipid peroxidationwas assessed by measuring MDA content according toOhkawa et al. [30]. It is based on the ability of this
Fig. 1. Experimental design. Cetirizine (10 mg/kg), cimetidine (15 mg/kg), and Betaserc (20 mg/kg) were administered by intraperitoneal injection. JNJ-7777120 (10 mg/kg) was given by subcutaneous route.
Involvement of H4 Receptor in Scorpion Envenomation Pathogenesis
marker to bind with two molecules of thiobarbituric acid(TBA) in acid medium and in hot temperature. To performthis assay, 150 μl of the tissue sample was mixed with anequal volume of trichloroacetic acid (35 % (v/v)) toprecipitate proteins. This mixture was incubated at 4 °Cfor 1 h and then centrifuged at 1,466×g for 10 min. Avolume of 200 μl of the obtained supernatant was added to100 μl of sodium dodecyl sulfate, 750 μl of acetic acid(20 %, pH 3.5), and 200 μl of distilled water. A volume of750 μl of thiobarbituric acid was then added to theprevious mixture. It was then heated for 1 h at 95 °C andthen cooled for 10 min at 4 °C. Absorbances weremeasured at 532 nm. Using a molar extinction coefficientof 1.56×105 M−1 cm−1, the results were calculated andexpressed as nanomolar of MDA content per 100 mg oftissue.
Determination of Catalase Activity. Measurement ofcatalase activity was carried out according to the Aebimethod [31]. Briefly, each sample (50 μl) was diluted(1/20) with phosphate buffer (50 mM, pH 7). Thereaction was thereafter initiated by 500 μl H2O2 (0.2 %).Kinetics of degradation of H2O2 was followed for 2 min at240 nm by spectrophotometer. The results were expressedin catalase unit (U) per 100 mg of tissue.
Histological Examination
Organs (heart, lung, and stomach) carefully excisedafter sacrifice of mice of each group were immediatelyimmersed in 10 % formalin for 48 h at room temperatureand subsequently processed for histological analysis. Sec-tions of 3 μm thick stained with hematoxylin and eosinwere analyzed using light microscopy (Motic Digital Mi-croscope PAL System).
Serum Enzyme Assays
Twenty-four hours after envenomation, blood sam-ples were collected in tubes without anticoagulant. Thesera were obtained by centrifugation at 1,398×g for10 min at room temperature. They were then stored imme-diately at 4 °C. Soon after that, serum enzymatic activityassays were carried out using CPK and LDH reagents(Spinreact) according to the manufacturer’s instructionsusing an automatic apparatus (Hitachi 902 ISE). The en-zyme activities were expressed in international units (IU/l).
Data Analysis
Results are expressed as mean ± standard error of themean (n=3). Differences between groups were analyzed
using Student’s t test. P values less than 0.05 were consid-ered significant.
RESULTS
Evaluation of the Role of Histamine Receptorsin the Induced Acute Cardiac, Pulmonary, and GastricEdema After Experimental Envenomation
The subcutaneous injection of Aah venom induced,24 h after its inoculation, a significant cardiac, pulmonary,and gastric edema as assessed by vascular permeabilityincrease in these organs and also by the increased mass tobody weight ratio (indexes of tissues water content) of theheart, lungs, and stomach (Fig. 2a, b). Results reveal thatthere are no significant differences between mice treatedwith saline (NaCl 0.9 %w/v) and those treated with DMSO(65 %v/v) in all experiments (data not shown). Therefore,in the following results, only the group receiving salinewill be represented.
The concentration of Evans blue dye extravasatedincreased significantly in the heart, lungs, and stomach(56.30±5.95, 58.28±1.67, and 44.30±4.52 per microgramof cardiac, pulmonary, and gastric tissues, respectively)compared with the basal respective concentrations of37.58±0.99, 11.87±1.37, and 10.15±2.14 per microgramof tissue. This was accompanied by an increment of theirfluid volume content indicated by the elevation of themasses-to-body weight ratio in the heart, lungs, and stom-ach (1.25±0.21, 0.90±0.01, and 2.03±0.28, respectively)compared with the respective control values of 0.56±0.03,0.76±0.05, and 0.99±0.06 (Fig. 2a, b).
Both H1 and H2 receptor antagonists (cetirizine andcimetidine, respectively) appeared to be effective in theprevention of the induced plasma extravasation of proteinsinto the cardiac (34.80±5.95 and 31.35±0.27 permicrogramof tissue) and pulmonary tissues (30.70±8.57 and 13.87±0.31 per microgram of tissue). In addition, these antagonistsreduced mass-to-body weight ratio compared with the ven-om group in the cardiac (0.57±0.1 and 0.68±0.05) andpulmonary tissues (0.78±0.07 and 0.83±0.005).
Although a slight decrease in the cardiac and pulmo-nary vascular permeability (47.28±5.73 and 47.26±4.60per microgram of tissue) and in the masses of these organsto body weight ratio (0.96±0.13 and 0.87±0.10) was ob-served in the presence of the histamine H3 receptor blocker(Betaserc), compared with the envenomed group. In thegastric tissue, the edema formation did not appear to bemediated by histamine H1 receptors (52.50±1.55 per
Lamraoui, Adi-Bessalem, and Laraba-Djebari
microgram tissue and 0.57±0.78) or the H3 receptors(62.88±9.29 per microgram tissue and 0.96±0.87) becauseit was still higher. However, the histamine H2 receptorantagonist seemed to be more efficient in the preventionof this event (31.19±2.14 per microgram tissue and 0.82±0.03). Given before Aah venom, JNJ-7777120, the hista-mineH4 receptor antagonist, significantly inhibited edema,as assessed by the decreased concentrations of the extrav-asated Evans blue dye in the heart (21.17±2.21 per micro-gram tissue), lungs (13.52±1.21 per microgram tissue),and the stomach (17.98±3 per microgram of tissue) andalso by the decrease in cardiac, pulmonary, and gastricmass-to-body weight ratio (0.68±0.121, 0.76±0.08, and1.09±0.05, respectively) (Fig. 2a, b).
Evaluation of Histamine Receptor Antagonist’s Effectson the Inflammatory Cell Infiltration and Activationin the Tissues Induced After ExperimentalEnvenomation
The recruitment of neutrophil and eosinophil cellsinto the studied tissues, 24 h after envenomation in theabsence or presence of histamine receptor antagonists, wasestimated by the measurement of activities of their enzymemarkers, myeloperoxidase, and eosinophil peroxidase, re-spectively. As results of venom administration, the MPOand EPO activities were significantly higher in the heart(0.096±0.007 mM/min/100 mg tissue and 1.03±0.10 ab-sorbance/100 mg tissue), lungs (0.260±0.009 mM/min/
Fig. 2. Effects of previous administration of cetirizine (10 mg/kg; i.p), cimetidine (15 mg/kg; i.p), Betaserc (20 mg/kg; i.p), or JNJ-7777120 (10 mg/kg; s.c)on the acute cardiac, pulmonary, and gastric edema induced by sublethal dose of Aah scorpion venom (0.5 mg/kg; s.c). a Quantification of Evans blue inheart, lungs and stomach. b Evaluation of cardiac, pulmonary, and gastric indexes. All data were presented as mean ± S.E.M. (n=3); *P<0.05; **P<0.01;***P<0.001; groups treated with venom versus controls (vehicle NaCl). #P<0.05; ##P<0.01; ###P<0.001; pretreated groups versus envenomed groups.
Involvement of H4 Receptor in Scorpion Envenomation Pathogenesis
100 mg tissue and 0.768±0.05 absorbance/100 mg tissue),and stomach (0.048±0.001 mM/min/100 mg tissue and0.396±0.05 absorbance/100 mg tissue) compared withthe cardiac (0.023±0.003 mM/min/100 mg tissue and0.414±0.02 absorbance/100 mg tissue), pulmonary(0.084±0.013 mM/min/100 mg tissue and 0.54±0.005absorbance/100 mg tissue), and gastric (0.0036±0.0001and 0.165±0.015 absorbance/100 mg tissue) activities ofuntreated mice (Fig. 3a–c). The pretreatment withcetirizine lowered the cardiac (33.3 %) and the pulmonary(57.7 %) MPO activity compared with mice receiving onlyvenom (Fig. 3a, b). In mice pretreated with cimetidine, thisactivity was reduced by 36.46 % in the heart and by 52.7 %in the lungs. The EPO activity was reduced in the presenceof cetirizine by about 44.7 and 33.94 % and also withcimetidine (37.81 and 50.7 %), compared with theenvenomed group of mice, respectively, in the cardiacand the pulmonary tissues (Fig. 3a, b). In the stomach,the activity of this enzyme was still higher in the presenceof cetirizine but decreased with cimetidine (24.7 %)(Fig. 3c). Neutrophil influx remained elevated in the pres-ence of Betaserc in the heart and the stomach, whereas areduction was observed in the lungs (36 %) (Fig. 3a–c).Eosinophil recruitment into the lungs and the stomach stillpersists even in the presence of this antagonist, but it isreduced in the heart (30 %). In addition, the previousadministration of JNJ-7777120, the histamine H4 receptorinhibitor, significantly decreased the peroxidase activities(MPO and EPO, respectively) in the heart (44.8 and 62 %),lungs (63 and 19.9 %), and stomach (60.41 and 48.6 %)(Fig. 3a–c), indicating that this receptor might play a crucialrole in inflammatory cell infiltration into all of these tissues.
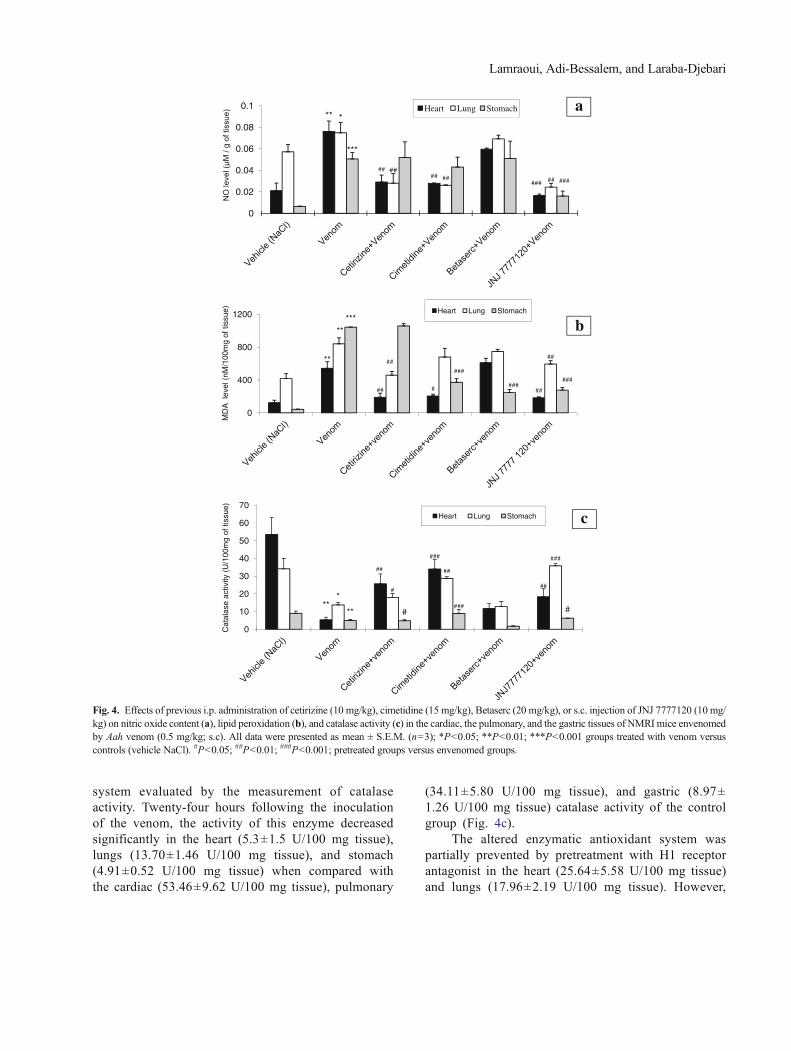
Effect of Aah Venom in the Absence or Presenceof Histamine Receptor Antagonists on Nitric OxideLevels
Nitric oxide (NO) levels were determined by measur-ing the rate of nitrites, the stable products of nitric oxide.Twenty-four hours after the subcutaneous injection of Aahvenom, levels of nitrites were increased in the heart (2.8-fold), lungs (0.3-fold), and stomach (7-fold) as comparedwith the respective controls (0.020±0.007, 0.057±0.007,and 0.0062±0.0004 μM/g tissue).
Pretreating animals with the blocker of the histamineH1 receptor significantly reduced Aah venom-induced in-crease of nitrite levels in the heart (61.84 %) and the lungs(63.51 %). In the stomach, these rates were not different
from the values expected in the envenomed group. Cimet-idine, the H2 receptor blocker, lowered the production ofnitrites by 64.47, 64.86, and 16 %, respectively, in theheart, lungs, and stomach compared with the envenomedgroup of mice. However, insignificant reductions wererecorded in the heart (0.060±0.001 vs. 0.076±0.009 μM/g tissue) and the lungs (0.069±0.003 vs. 0.074±0.009 μM/g tissue) of envenomed mice in the presence of the H3receptor antagonist. In addition, there was no reduction ofnitrite contents in the stomach, which were still higher(0.05±0.016 vs. 0.0505±0.006 μM/g tissue) in presenceof the H3 receptor blocker. Furthermore, H4 receptor an-tagonist pretreatment decreased nitrite levels in the heart(78.9 %), lungs (67 %), and stomach (68 %) when com-pared with animals receiving only the venom, indicatingthat this receptor might play a role in the nitrosative stressin these tissues (Fig. 4a).
Effects of Histamine Receptor Antagonists on LipidPeroxidation Induced After Envenomation
Malondialdehyde level was measured to estimate lip-id peroxidation. Following administration of venom, theMDA level increased markedly compared with that in thecontrols to reach values of 542.74±80.62, 840.12±72.73,and 1,043.51±2.09 nM/100 mg tissue, respectively, in thecardiac, pulmonary, and gastric tissues (Fig. 4b).
Cetirizine significantly suppressed the increase inMDA levels caused by the venom in the heart (190.10±49.34 nM/100 mg tissue) and lungs (458.12±45.84 nM/100 mg tissue). However, this pretreatment showed nosuch preventive effects in the stomach (1,058.25±26.44 nM/100 mg tissue). Cimetidine- and JNJ-7777120-pretreated mice showed a reduction of this marker of lipidperoxidation in the cardiac tissue (205.60±21.84 and182.95±12.51 nM/100 mg tissue) and also in the lungs(679.01±107.08 and 594.12±41.36 nM/100 mg tissue). Inthe stomach, MDAwas estimated to be 372.21±43.77 nM/100 mg tissue in the presence of cimetidine and 277.05±30.47 nM/100 mg tissue in the presence of JNJ-7777120.Betaserc administration reduced the lipid peroxidation inthe stomach (248.34±35 nM/100 mg tissue) but remainedhigher in the heart (609.87±54.51 nM/100 mg tissue) andin the lungs (746.40±26.87 nM/100 mg tissue) (Fig. 4b).
Effect of the Venom on the Enzymatic AntioxidantSystem in the Absence or Presence of HistamineReceptor Antagonists
Mice injected with Aah venom displayed a severealteration of the cardiac, pulmonary, and gastric antioxidant
Lamraoui, Adi-Bessalem, and Laraba-Djebari
Fig. 3. Effects of previous administration by i.p. route of cetirizine (10 mg/kg), cimetidine (15 mg/kg), Betaserc (20 mg/kg), or s.c. injection of JNJ 7777120(10 mg/kg) on MPO and EPO activities reflecting neutrophil and eosinophil cells infiltration into cardiac (a), pulmonary (b), and gastric (c) tissues of NMRImice envenomed by sublethal dose of Aah scorpion venom (0.5 mg/kg; s.c). All data were presented as mean ± S.E.M. (n=3); ***P<0.001 groups treatedwith venom versus controls (vehicle NaCl). #P<0.05; ##P<0.01; ###P<0.001; pretreated groups versus envenomed groups.
Involvement of H4 Receptor in Scorpion Envenomation Pathogenesis
system evaluated by the measurement of catalaseactivity. Twenty-four hours following the inoculationof the venom, the activity of this enzyme decreasedsignificantly in the heart (5.3±1.5 U/100 mg tissue),lungs (13.70±1.46 U/100 mg tissue), and stomach(4.91±0.52 U/100 mg tissue) when compared withthe cardiac (53.46±9.62 U/100 mg tissue), pulmonary
(34.11±5.80 U/100 mg tissue), and gastric (8.97±1.26 U/100 mg tissue) catalase activity of the controlgroup (Fig. 4c).
The altered enzymatic antioxidant system waspartially prevented by pretreatment with H1 receptorantagonist in the heart (25.64±5.58 U/100 mg tissue)and lungs (17.96±2.19 U/100 mg tissue). However,
**
####
###
*
#### ##
***
###
0
0.02
0.04
0.06
0.08
0.1 Heart Lung Stomach
**
## # ##
**
####
***
###
######
0
400
800
1200
MD
A l
evel
(nM
/100
mg
of ti
ssue
)
Heart Lung Stomach
**
##
###
##
*#
##
###
** #### #
0
10
20
30
40
50
60
70
Cat
alas
e ac
tivity
(U
/100
mg
of ti
ssue
)
Heart Lung Stomach
a
c
b
Fig. 4. Effects of previous i.p. administration of cetirizine (10 mg/kg), cimetidine (15 mg/kg), Betaserc (20 mg/kg), or s.c. injection of JNJ 7777120 (10 mg/kg) on nitric oxide content (a), lipid peroxidation (b), and catalase activity (c) in the cardiac, the pulmonary, and the gastric tissues of NMRI mice envenomedby Aah venom (0.5 mg/kg; s.c). All data were presented as mean ± S.E.M. (n=3); *P<0.05; **P<0.01; ***P<0.001 groups treated with venom versuscontrols (vehicle NaCl). #P<0.05; ##P<0.01; ###P<0.001; pretreated groups versus envenomed groups.
Lamraoui, Adi-Bessalem, and Laraba-Djebari
in the gastric tissue, catalase activity still compro-mised (4.79±0.67 U/100 mg of tissue). Moderateeffect on the activity of this enzyme in the heart(34±5.39 U/100 mg tissue), lungs (28.69±0.93 U/100 mg tissue), and stomach (8.86±2.37 U/100 mgtissue) was observed in the presence of cimetidine.Although insignificant increase of this activity wasobtained in the heart (11.64±2.84 U/100 mg tissue)and the lungs (12.84±2.74 U/100 mg tissue) of micepretreated with the H3 receptor antagonist, this pre-treatment aggravated the alteration of this antioxidantsystem in the stomach (1.71±0.27 U/100 mg tissue)(Fig. 4c).
In the heart, JNJ-7777120, the histamine H4 receptorantagonist, pretreatment increased catalase activity (18.34±4.51 U/100 mg tissue) without reaching the controlvalues. This was observed also in the stomach (6.24±0.07 U/100 mg tissue). H4 receptor blockade caused morepreventive effects in the lungs (35.79±1.18 U/100 mgtissue) (Fig. 4c).
Histological Examination
The histopathological analysis of the heart ofcontrol mice revealed normal cardiac tissue architec-ture consisting of parallel myocardial fibers (Fig. 5a).The administration of venom caused interstitial ede-ma, hemorrhage, and massive leukocyte infiltrationinto this tissue 24 h after its inoculation (Fig. 5band B′). The previous administration of the histamineH1 receptor antagonist reduced interstitial edema(Fig. 5c), whereas the pretreatment with cimetidineprevented hemorrhage and leukocyte infiltration intothe myocardium tissue and markedly reduced theinterstitial edema (Fig. 5d). On the other hand, thehistamine H3 receptor antagonist seemed to be noteffective against hemorrhage and inflammatory cellinfiltration (Fig. 5e). Interestingly, there were no ob-vious alterations in the cardiac tissue when animalswere pretreated with the histamine H4 receptor an-tagonist (JNJ-7777120) (Fig. 5f).
a
c d
e f
bN
H
Ie Li
H N
Ie
Ie
Li
Li
B’
H
B’
Fig. 5. Light microscopy of myocardium tissue of control and envenomed mice, in absence or in presence of histamine receptor antagonists, 24 h after en-venomation. Representative thin sections of 3 μm. a Control (vehicle NaCl). b Myocardium tissue of mice injected with subletal dose of A. australis hectorvenom (0.5 mg/kg; s.c). c–fMyocardium tissue of pretreated mice with cetirizine (10 mg/kg; i.p), cimetidine (15 mg/kg; i.p), Betaserc (20 mg/kg; i.p), and JNJ7777120 (10 mg/kg; s.c), respectively. N, nucleus. H, hemorrhage. Li, leukocyte infiltration. Ie, interstitial edema. Hematoxylin–eosin, magnification×400.
Involvement of H4 Receptor in Scorpion Envenomation Pathogenesis
The pulmonary parenchyma of control animals con-sists of alveoli separated by thin interalveolar septa(Fig. 6a). Microscopic examination of the lung tissuearchitecture of mice 24 h after envenomation re-vealed a disorganization of the pulmonary parenchy-ma characterized mainly by the thickening ofinteralveolar septa associated to hemorrhage, edema,and inflammatory cells in alveolar lumens (Fig. 6band B′). The previous administration of the histamineH1 receptor antagonist prevented all alterations in-duced by the venom in the pulmonary tissue includ-ing hemorrhage, edema, inflammatory cellular infil-tration, and a thinking of interalveolar septa(Fig. 6c), whereas few inflammatory cells infiltratingthe lung parenchyma were observed in the presenceof the histamine H2 receptor antagonist (Fig. 6d).The thickening of the interalveolar septa was stillobserved in the presence of Betaserc (Fig. 6e). Other-wise, a marked reduction of their thickness, absenceof edematous areas, and a few inflammatory cellinfiltration (data not shown) were observed in thepresence of JNJ-7777120 (Fig. 6f).
We further examined the gastric tissue, whichshowed a normal structure consisting of the mucosaand muscularis separated by a thin submucosa(Fig. 7a and A′). Envenomed mice showed severehistological changes characterized by a thick submu-cosa infiltrated with inflammatory cells as shown inFig. 7b, B′, and B″.
The thickening of the submucosa and the in-flammatory cell infiltration (less abundant than inthe gastric tissue of envenomed mice) were stillobserved in the presence of cetirizine (Fig. 7c andC′). There were no cellular inflammatory cells infil-trating the submucosa, and a marked reduction of itsthickness was observed in the gastric tissue ofpretreated mice with the histamine H2 receptorblocker (Fig. 7d). Moreover, the mice that receivedBetaserc before envenomation showed moderate en-largement of the submucosa with few cellular inflam-matory cell infiltration (Fig. 7e and E′), whereaspretreated animals with histamine H4 receptor antag-onist (JNJ-7777120) showed no obvious alterationsof this tissue structure (Fig. 7f).
a b
c d
e f
AE
H
A
Li
IS TIS
TIS
A
A
A
Li
B
Fig. 6. Light microscopy of pulmonary parenchyma of control and envenomed mice in absence or in presence of histamine receptor antagonists, 24 h afterenvenoming. Representative thin sections of 3 μm. a Control (vehicle NaCl). b Pulmonary tissues of mice injected with subletal dose of A. australis hectorvenom (10 μg/20 g; s.c). c–f Lung tissues of pretreated mice with cetirizine (10 mg/kg; i.p), cimetidine (15 mg/kg; i.p), Betaserc (20 mg/kg; i.p), or JNJ-7777120 (10 mg/kg; s.c), respectively. A, alveolar; E, edema;H, hemorrhage; IS, inter-alveolar septum; Li, leukocyte infiltration; TIS, thick inter-alveolar septum.Hematoxylin–eosin, magnification×400.
Lamraoui, Adi-Bessalem, and Laraba-Djebari
Effects of Histamine Receptor Blockers on CPK andLDH Activities
Serum activities of creatine kinase (CPK) and lactatedehydrogenase (LDH) of envenomed mice were signifi-cantly higher (2,514±79.92 and 1,917.67±87.37 IU/l, re-spectively) than those estimated in the control sera(1,608.33±38.18 and 1,691.66±95.17 IU/l, respectively).Pretreatment with histamine H1, H2, or H4 receptor antag-onists before venom injection resulted in a significantreduction in serum CPK activity (2,321.66±18.92,2,195.66±5.13, and 1,445.66±5.50 IU/l).
On the other hand, the blockade of these receptorsinduced lower levels of LDH activities (1,122±319, 1,241±207, and 1,593±40.41 IU/l, respectively) than those re-corded in envenomed animal sera. Nevertheless, the hista-mine H3 receptor antagonist exhibited no significant ef-fects on CPK (2,571.66±62.51 IU/l) or LDH (1,892.3±80.83 IU/l) activities when compared with the envenomedgroup of mice (Fig. 8a, b).
DISCUSSION
Scorpion envenoming syndrome is characterized byacute inflammatory disorders affecting human health andvariety of organs in experimental animals. The exact mech-anism of the triggering of this inflammatory response isstill not completely understood, and several related eventshave been suggested to be involved. Histamine, a well-known immuno-inflammatory modulator [32], could be anexcellent mediator of initiation and maintenance of inflam-matory response. Therefore, the current study emphasizedthe relevance of histamine through its four receptors in thisprocess.
This study showed that envenoming with Aah venomcaused an inflammatory reaction, characterized by an in-crease in the cardiac, pulmonary, and gastric vascular per-meability to serum proteins. This was indicated by theincrease in Evans blue extravasation from the vascularspace to these organs. Similar results observed in the lungs
a b
f
dc
Li
e
TSM
Li B’’B’
A’
D’
C’
E’ F’
M
M
M
M
M
M
TSM
TSM
MS
MS
MS
MS
MS
MS
Li
Fig. 7. Light microscopy of gastric tissue of control and envenomedmice, in absence or in presence of histamine receptor antagonists, 24 h after envenoming.Representative thin sections of 3 μm. aControl. bGastric tissue of mice injected with subletal dose of A. australis hector venom (0.5 mg/kg; s.c). c–fGastrictissue of pretreatedmicewith cetirizine (10mg/kg; i.p), cimetidine (15mg/kg; i.p), Betaserc (20mg/kg; i.p), or JNJ 7777 120 (10mg/kg; s.c), respectively.M,mucosa. SM, submucosa. TSM, thick submucosa.MS, muscularis. Li, leukocyte infiltration. Hematoxylin–eosin, (a–f) magnification×100. (A′, B′, B″, C′, D′,E′, and F′) magnification×400.
Involvement of H4 Receptor in Scorpion Envenomation Pathogenesis
of envenomed animals with Aah venom were reported [19,33]. Other studies reported that the venom of Tityusserrulatus venom is able to increase the alveolar–capillarypermeability [18, 20, 34]. In addition, vascular permeabil-ity was also increased in the paw of rats in response to thevenom of Buthus martensi Karsch [35]. In this study, wereport that the Aah venom induced also vasodilatation andincreased the permeability of the cardiac and gastric ves-sels. This event was accompanied by edema formation asassessed by the increased heart, lungs, and stomachmasses-to-body weight ratio and also by an increase incellular inflammatory infiltration, including neutrophiland eosinophil cells, indicated indirectly by the increaseof their enzymes markers (MPO and EPO) activities. Theseimmune cells represent important sources of reactive oxy-gen intermediates such as H2O2 derived from superoxideanion and nitrogen reactive species such as NO [36–38],which could partially explain the higher levels of NOassessed in these tissues after envenomation. Previousstudies reported the generation of NO and correlate its
production with the severity of envenomation [11, 13].The excessive release of NO and reactive oxygen speciesleads generally to severe imbalanced redox status withaltered or insufficient antioxidant systems [39], demon-strated here by the reduced catalase activity, which wasprobably compromised during this inflammatory reactionby excessive production of its own substrate (H2O2) [40].Reactive oxygen species and NO product interact and actas cytotoxic agents by cross-linking lipids, leading to lipidperoxidations and resulting in a loss of cell membranesintegrity [41]. Considered as final product of lipid peroxi-dation, MDAwas found to be higher after the experimentalenvenomation described here and also in the study de-scribed by Dousset and coworkers [15] and that by Adi-Bessalem and collaborators [42]. This was reported also inhuman envenoming cases [13], reflecting tissue injuries.This has led us to analyze organ structures. Indeed, Aahvenom-treated mice displayed cardiac and pulmonary his-topathological disorders marked by hemorrhages, edema,and infiltration of inflammatory cells. An important
Fig. 8. Effect of a subletal dose of A. australis hector venom (0.5 mg/kg; s.c) in absence or in presence of histamine H1 (cetirizine; 10 mg/kg; i.p), H2(cimetidine;15 mg/kg; i.p), H3 (Betaserc 20 mg/kg; i.p), or H4 (JNJ 7777120;10 mg/kg; i.p) receptor antagonists on the variation of CPK (a) and LDH (b)activities. All data were presented as mean ± S.E.M. (n=3); *P<0.05; ***P<0.001 groups treated with venom versus controls. #P<0.05; ##P<0.01;###P<0.001, pretreated groups versus envenomed groups.
Lamraoui, Adi-Bessalem, and Laraba-Djebari
observation in this study is that this venom induced dele-terious effects in the gastric tissue. The thickening of thesubmucosa and its infiltration by inflammatory cells werethe most noteworthy observations. The microscopic alter-ations in the heart and in the lungs were similar with manyprevious studies on this venom [7, 8, 43, 44] or on venomsof other scorpion species [6, 34, 45, 46]. The present is thefirst study demonstrating such alterations in the gastrictissue in response to scorpionic envenomation, althoughHeidarpour and collaborators [47] described that theHemiscorpius lepturus scorpion venom was able to inducetoxic effects in this organ. Nevertheless, they had notreported any detail demonstrating this observation. Theseanatomic alterations caused by Aah venom challenge faith-fully reflected in the serum indicated by the increased CPKand LDH activities, which are usually correlated with theintensity of tissue damages in scorpionic envenomationcases [2, 7, 48].
In our study, pretreatment of mice with the selectivehistamine H1 receptor antagonist, cetirizine, significantlyreduced the venom-induced cardiac and pulmonary plasmaextravasation and edema formation. These findings sup-port earlier studies, which suggest that H1 receptor has arole in promoting vascular permeability and edema induc-tion during scorpion envenomation [16, 26, 33]
Leukocyte infiltration and the concomitant generationof free radicals were also reversed in the presence ofcetirizine. A possible explanation could be related to thereduced vascular permeability, which precedes inflamma-tory cell infiltration, and also to the fact that histamine H1receptor stimulation activates the transcriptional factor NF-kappaB [49], which controls the expression of adhesionmolecules on the endothelium such as E selectins, VCAM-1, and ICAM-1 [50]. This molecule (ICAM-1) was foundto be activated during scorpion envenomation, leading toneutrophil and eosinophil recruitment [11, 51–54]. Thenuclear factor kB controls also the expression of the induc-ible NO synthase [55], explaining the reduction of NOproduction in the heart and lungs of cetirizine-pretreatedmice. Others [56] suggest that histamine acting on its H1receptor stimulates neutrophils to generate reactive oxygenspecies, ultimately leading to lipid peroxidation and cellu-lar damages [15, 42].
In contrast, Aah venom induced plasma extravasation,and edema was not inhibited in the stomach, indicating thatgastric oxidative inflammation is not mediated directly bythis receptor.
However, cimetidine, the histamine H2 receptor an-tagonist, appeared to be effective in the prevention of thedisorders occasioned by the venom in this organ.
Interesting results were found also in the heart and lungsin the presence of cimetidine. The histamine H2 receptorhas been previously involved in histamine-induced vaso-dilatation and permeability of the peripheral vessels [57,58]. Others suggest its involvement in edema formation[26, 59]. These data provided information that may supportour results, suggesting a role for H2 receptors in increasingcardiac, pulmonary, and gastric capillary permeability andthe subsequent edema induction after Aah envenomation.
Histamine H2 receptor has a crucial role in the stim-ulation of gastric acid secretion [24], but its involvement inimmunomodulation is not excluded. The presence of thisreceptor on dendritic cells [60] leads us to propose anotherexplanation. Indeed, histamine acting on H2 receptorsexpressed on these cells switches the immune responsetoward TH2 profile with Ig-E release [61]. This immuno-globulin might be induced by Ig-E-dependent mechanism,mast cell degranulation [62] involved in microvasculaturepermeability increase, and edema formation, secondary toamines and PLA2-derived metabolites release [26, 33]. Inaddition, these mediators have been recognized for theirrole in neutrophil attraction [63].
Eosinophil influx was also found to be reduced afterinhibition of H2 receptors. Histamine released mainly frommast cells and basophils, acting on H2 receptors, plays arelevant role as stimulatory mediator in increasing IL-5production by TH2 cells [64, 65]. This cytokine has animportant role in eosinophil activation [50, 54]. Thus, thiscould explain the reduction in the eosinophil recruitmentinto the tissues in the presence of this blocker.
Previous studies suggest the implication of the H2receptors in gastric lipid peroxidation [66], which indicatesthe ability of these receptors to promote cellular injuries.
Our results report that the administration of the hista-mine H3 receptor antagonist partially decreased the oxida-tive inflammation induced by the Aah venom in the heartand lungs, although it is well known that the stimulation ofthis receptor results in anti-inflammatory actions [67].Nevertheless, Raible and collaborators [68] showed thathistamine H3 receptors participated in the chemotaxis ofeosinophils to the inflammatory site. This seems to be veryinteresting and could explain the reduction of eosinophilinflux into the cardiac tissue in the presence of Betaserc.
In the gastric tissue, most inflammatory disorders(inflammatory cell infiltration, increase of NO levels, anddecrease of catalase activity) induced by the venom couldnot be prevented by blocking the histamine H3 receptors,suggesting that these events were not mediated by them.
Moreover, we further investigated the role of therecently identified histamine H4 receptor in the Aah
Involvement of H4 Receptor in Scorpion Envenomation Pathogenesis
venom-induced inflammatory disorders. Administration ofJNJ-7777120, the selective antagonist of this receptor,resulted in a marked prevention of the Aah venom-inducedcardiac, pulmonary, and gastric inflammatory disorders,which suggests an inflammatory role for this receptor inthe heart, the pulmonary tissue and also in the stomachduring Aah envenoming.
It has been previously reported that histamine medi-ates mast cell chemotaxis and mobilization of calcium inthese cells via H4 receptors [69]. Calcium influx is animportant event leading to mast cell degranulation [70],resulting in the release of various granule-stored mediatorswith vasoactive properties such as histamine, prostaglan-dins, and leukotrienes, found to be involved in the en-hancement of vessel permeability and the subsequent ede-ma formation in scorpion envenomation [26, 33].
In addition, histamine acting on this receptor contrib-utes to the chemotaxis of other immune cells expressing theH4 receptor on their surfaces including basophils anddendritic cells [71, 72], as well as eosinophils [73] andneutrophils [74].
Our research team demonstrates in the present studythe first evidence reporting the involvement of the hista-mine H4 receptor in the inflammatory response induced bythe scorpion venom.
Taken together, our results suggest the involvement ofthe histamine H1, H2, H4, and partially H3 receptors in theinflammatory response induced by Aah venom in the heartand lungs, whereas in the gastric tissue, this response wasmainly mediated through H2 and H4 receptors.
In conclusion, understanding of the molecular mech-anism of scorpionic envenomation pathogenesis is an es-sential step to develop efficient therapies to treat victimssuffering from scorpion stings. With our results as basis,histamine H4 receptor appears to be an interesting thera-peutic target to alleviate the inflammatory disorders causedby scorpion venom. Using histamine H4 receptor antago-nists, such as newer anti-inflammatory drugs, could pro-vide better treatment in scorpion envenomed cases.
REFERENCES
1. Ismail, M. 1995. Review article: the scorpion envenoming syndrome.Toxicon 33: 825–858.
2. Hammoudi-Triki, D., E. Ferquiel, A. Robbe-Vincent, C. Bon, V.Choumet, and F. Laraba-Djebari. 2004. Epidemiological data, clinicaladmission gradation and biological quantification by ELISA of scor-pion envenomations in Algeria: effect of immunotherapy. Royal So-ciety of Tropical Medicine and Hygiene 98: 240–250.
3. Delori, P., J.F. Van-Rietschoten, and H. Rochat. 1981. Scorpion venomand neurotoxins: an immunological study. Toxicon 19(3): 393–407.
4. Possani, L.D., B. Becerril, M. Delepierre, and J. Tytgat. 1999. Scor-pion toxins specific for Na+-channels. European Journal ofBiochemistry 264: 287–300.
5. Sofer, S., and M. Gueron. 1988. Respiratory failure in children fol-lowing envenomation by the scorpion Leiurus quinquestriatus: hemo-dynamics and neurological aspects. Toxicon 26: 931–939.
6. De-Matos, I.M., O.A. Rocha, R. Leite, and L. Freire-Maia. 1997.Lung edema induced by Tityus serrulatus scorpion venom in the rat.Comparative Biochemistry and Physiology 118C: 143–148.
7. Adi-Bessalem, S., D. Hammoudi-Triki, and F. Laraba-Djebari. 2008.Pathophysiological effects of Androctonus australis hector scorpionvenom: tissue damages and inflammatory response. Experimental andToxicologic Pathology 60(4–5): 373–380.
8. Sami-Merah, S., D. Hammoudi-Triki, M.F. Martin-Eauclaire, and F.Laraba-Djebari. 2008. Combination of two antibody fragmentsF(ab’)2/Fab: an alternative for scorpion envenoming treatment. Inter-national Immunopharmacology 8: 1386–1394.
9. D’Suze, G., S. Moncada, C. González, C. Sevcik, V. Aguilar, and A.Alagón. 2003. Relationship between plasmatic levels of various cyto-kines, tumour necrosis factor, enzymes, glucose and venom concen-tration following Tityus scorpion sting. Toxicon 41(3): 367–375.
10. Fukuhara, Y.D., M.L. Reis, R. Dellalibera-Joviliano, F.Q. Cunha, andE.A. Donadi. 2003. Increased plasma levels of IL-1beta, IL-6, IL-8,IL-10 and TNF-alpha in patients moderately or severely envenomedby Tityus serrulatus scorpion sting. Toxicon 41(1): 49–55.
11. Raouraoua-Boukhari, R., S. Sami-Merah, D. Hammoudi-Triki, and F.Laraba-Djebari. 2012. Immunomodulation of the inflammatory re-sponse induced by Androctonus australis hector neurotoxins: bio-marker interactions. Neuroimmunomodulation 19(2): 103–110.
12. Saidi, H., S. Adi-Bessalem, D. Hammoudi-Triki, and F. Laraba-Djebari. 2013. Effects of atropine and propranolol on lung inflamma-tion in experimental envenomation: comparison of two buthidaevenoms. Journal of Venomous Animals and Toxins including TropicalDiseases 19(8): 1–7.
13. Meki, A.R., and Z.M. Mohey El-Dean. 1998. Serum interleukin-1beta, interleukin-6, nitric oxide and alpha1-antitrypsin in scorpionenvenomed children. Toxicon 36(12): 1851–1859.
14. Petricevich, V.L., and C.F. Penã. 2002. The dynamics of cytokine dnitric oxide secretion in mice injected with Tityus serrulatus scorpionvenom. Mediators of Inflammation 11: 173–180.
15. Dousset, E., L. Carrega, J.G. Steinberg, O. Clot-Faybesse, B. Jouirou,N. Sauze, C. Devaux, Y. Autier, Y. Jammes, M.F. Martin-Eauclaire,and R. Guieu. 2005. Evidence that free radical generation occursduring scorpion envenomation. Comparative Biochemistry and Phys-iology, Part C 140: 221–226.
16. Nascimento Jr., E.B., K.A. Costa, C.M. Bertollo, A.C. Oliveira, L.T.Rocha, A.L. Souza, M.B. Glória, T. Moraes-Santos, and M.M.Coelho. 2005. Pharmacological investigation of the nociceptive re-sponse and edema induced by venom of the scorpion Tityus serrulatus.Toxicon 45(5): 585–593.
17. Pessini, A.C., A. Kanashiro, D.C. Malvar, R.R. Machado, D.M.Soares, M.J. Figueiredo, E. Kalapothakis, and G.E.P. Souza. 2008.Inflammatory mediators involved in the nociceptive andoedematogenic responses induced by Tityus serrulatus scorpion ven-om injected into rat paws. Toxicon 52: 729–736.
18. De-Matos, I.M., A. Talvani, O.O.A. Rocha, L. Freire-Maia, andM.M.Teixeira. 2001. Evidence for a role of mast cells in the lung edemainduced by Tityus serrulatus venom in rats. Toxicon 39: 863–867.
19. Adi-Bessalem, S., Sami-Merah, S., Mendil, A., Hamoudi-Triki, D.,Laraba-Djebari, F. 2010. Pharmacological assessment of inflammato-ry mediators after Androctonus australis hector envenomation: in-volvement of histamine H1 receptor. Meeting on Toxinology. 1-2.
20. Zuliani, J.P., T.A. Freitas, I.M. Conceicão, and F.H. Kwasniewski.2013. Tityus serrulatus venom increases vascular permeability in
Lamraoui, Adi-Bessalem, and Laraba-Djebari
selected airway tissues in a mast cell-independent way. Experimentaland Toxicologic Pathology 65: 229–234.
21. Buckley, I.K., and G.B. Ryan. 1969. Increased vascular permeability.The effect of histamine and serotonin on rat mesenteric blood vesselsin vivo. American Journal of Pathology 55(3): 329–347.
22. Ohuchi, Y., H. Ohtsu, E. Sakurai, K. Yanai, A. Ichikawa, Z. Radvany,et al. 1998. Induction of histidine decarboxylase in type2 T helperlymphocytes treated with anti-CD3 antibody. Inflammation Research47(1): 48–49.
23. Szeberenyi, J.B., E. Pallinger, M. Zsinko, Z. Pos, G. Rothe, E. Orso,et al. 2001. Inhibition of effects of endogenously synthesized hista-mine disturbs in vitro human dendritic cell differentiation. Immunol-ogy Letters 76: 175–182.
24. Hill, S.J., C.R. Ganellin, H. Timmermann, J.C. Schwartz, N.P.Shnakley, J.M. Young, W. Schunack, R. Levi, and H.L. Haas. 1997.International union of pharmacology XIII. Classification of histaminereceptors. Pharmacological Reviews 49: 253–278.
25. Parsons, M.E., and C.R. Ganellin. 2006. Histamine and its receptors.British Journal of Pharmacology 147(S1): 127–135.
26. Liu, T., Z.T. Bai, X.Y. Pang, Z.F. Chai, F. Jiang, and Y.H. Ji. 2007.Degranulation of mast cells and histamine release involved in rat pain-related behaviors and edema induced by scorpion Buthus martensiKarch venom. European Journal of Pharmacology 575: 46–56.
27. Dutta, A., and S.B. Deshpande. 2011. Indian red scorpion venom-induced augmentation of cardio-respiratory reflexes and pulmonaryedema involve the release of histamine. Toxicon 57: 193–198.
28. De Matos, I.M., D.G. Souza, D.G. Seabra, L. Freire-Maia, and M.M.Teixeira. 1999. Effects of tachykinin NK1 or PAF receptor blockadeon the lung injury induced by scorpion venom in rats. EuropeanJournal of Pharmacology 376: 293–300.
29. Sun, J., X. Zhang, M. Broderich, and H. Fein. 2003. Measurement ofnitric oxide production in biological systems by using Griess reactionassay. Sensors 3: 276–284.
30. Ohkawa, H., N. Ohishi, and K. Yagi. 1979. Assay for lipid peroxidesin animal tissues by thiobarbituric acid reaction. Analytical Biochem-istry 95: 351–358.
31. Aebi, H. 1984. Catalase in vitro.Methods in Enzymology 105: 121–126.32. Mac Glashan Jr., D. 2003. Histamine: a mediator of inflammation.
Journal of Allergy and Clinical Immunology 112(4): 53–59.33. Sami-Merah, S., Hammoudi-Triki, D., Adi-Bessalem, S., Mendil A.,
Martin-Eauclaire MF., Laraba-Djebari F. 2009. L’augmentation de lapermeabilité vasculaire serait-elle un facteur déclenchant de l’oedemepulmonaire induit par le venin du scorpion Androctonus australishector?: Toxine et Signalisation-Rencontre en Toxinologie Editionsde la SFET. 161-163.
34. Oliveira, F.N., M.R. Mortari, F.P. Carneiro, J.A. Guerrero-Vargas,D.M. Santos, A.M.C. Pimenta, and E.F. Schwartz. 2013. Anotherrecord of significant regional variation in toxicity of Tityus serrulatusvenom in Brazil: a step towards understanding the possible role ofsodium channel modulators. Toxicon 73: 33–46.
35. Chen, B., C. Wang, and Y. Ji. 2002. Asian scorpion BmK venominduces plasma extravasation and thermal hyperalgesia in the rat.Toxicon 40: 527–533.
36. Pincus, S.H., A.M. Di Napoli, and W.R. Schooley. 1982. Superoxideproduction by eosinophils: activation by histamine. Journal of Inves-tigative Dermatology 79: 53–57.
37. Elsner, J., S. Dichmann, and A. Kapp. 1995. Activation of the respira-tory burst in human eosinophils by chemotaxins requires intracellularcalcium fluxes. Journal of Investigative Dermatology 105: 231–236.
38. Defraigne, J.O., J. Pincemail, R. Larbuisson, F. Blaffart, and R. Limet.2000. Cytokine release and neutrophil activation are not prevented byheparin-coated circuits and aprotinin administration. Annals of Tho-racic Surgery 69: 1084–1091.
39. Kimura, H., T. Sawada, S. Oshima, K. Kozawa, T. Ishioka, and M.Kato. 2005. Toxicity and roles of reactive oxygen species. CurrentDrug Targets. Inflammation and Allergy 4: 489–495.
40. Lardinois, O.M.,M.M.Mestdagh, and P.G. Rouxhet. 1996. Reversibleinhibition and irreversible inactivation of catalase in presence ofhydrogen peroxide. Biochimica et Biophysica Acta 1295(2): 222–238.
41. Folkerts, G., J. Kloek, R.B. Muijsers, and F.P. Nijkamp. 2001. Reac-tive nitrogen and oxygen species in airway inflammation. EuropeanJournal of Pharmacology 429: 251–262.
42. Adi-Bessalem, S., A. Ladjal-Mendil, D. Hammoudi-Triki, and F.Laraba-Djebari. 2012. Immuno-inflammatory response after scorpionenvenomation: potential role of eïcosanoids and histamine H1-recep-tor. Toxicon 60: 95–248.
43. Bessalem, S., D. Hammoudi-Triki, and F. Laraba-Djebari. 2003. Effetde l’immunothérapie sur les modifications métaboliques ethistopathologiques après envenimation scorpionique expérimentale.Bulletin de la Societe de Pathologie Exotique 96: 297–302.
44. Boussag-Abib, L., and F. Laraba-Djebari. 2011. Enhanced immunesera and vaccine: safe approach to treat scorpion envenoming. Vaccine29: 8951–8959.
45. Corréa, M.M., S.V. Sampaio, R.A. Lopes, L.C. Mancuso, O.A.B.Cunha, J.J. Franco, et al. 1997. Biochemical and histopathologicalalterations induced in rats by Tityus serrulatus scorpion venom and itsmajor neurotoxin Tityustoxin-1. Toxicon 35: 1053–1067.
46. D’Suze, G., V. Salazar, P. Diaz, C. Sevcik, H. Azpurua, and N. Bracho.2004. Histopathological changes and inflammatory response inducedby Tityus discrepans scorpion venom in rams. Toxicon 44: 851–860.
47. Heidarpour, M., E. Ennaifer, H. Ahari, N. Srairi-Abid, L. Borchani, G.Khalili, H. Amini, A.A. Anvar, S. Boubaker, M. El-Ayeb, and D.Shahbazzadeh. 2012. Histopathological changes induced byHemiscorpius lepturus scorpion venom in mice. Toxicon 59: 373–378.
48. Daisley, H., D. Alexander, and P. Pitt-Miller. 1999. Acute myocarditisfollowing Tityus trinitatis envenoming: morphological and pathophys-iological characteristics. Toxicon 37: 159–165.
49. Bakker, R.A., S.B. Schoonus, M.J. Smit, H. Timmerman, and R.Leurs. 2001. Histamine H1-receptor activation of nuclear factor-k B:roles for Gbg- and Gaq/11-subunits in constitutive and agonist-medi-ated signaling. Molecular Pharmacology 60: 1133–1142.
50. Collins, P.D., S. Marleau, D.A. Griffiths-Johnson, P.J. Jose, and T.J.Willliams. 1995. Cooperation between interleukin-5 and the chemo-kine eotaxin to induce eosinophil accumulation in vivo. Journal ofExperimental Medicine 182: 1169–1174.
51. Shah, P.K., M. Lakhotia, M. Chittora, S. Mehta, and A. Purohit. 1989.Pulmonary infiltration with blood eosinophilia after scorpion sting.Chest 95: 691–692.
52. Voronov, E., R.N. Apte, and S. Sofer. 1999. The systemic inflamma-tory response syndrome related to the release of cytokines followingsevere envenomation. Journal of Venomous Animals and Toxins 5(1):19–23.
53. Borges, A., H.J.M. Op den Camp, and J.B. De Sanctis. 2011. Specificactivation of human neutrophils by scorpion venom: a flow cytometryassessment. Toxicology in Vitro 25(1): 358–367.
54. Adi-Bessalem, S., A. Mendil, D. Hammoudi-Triki, and F. Laraba-Djebari. 2012. Lung immunoreactivity and airway inflammation: theirassessment after scorpion envenomation. Inflammation 35(2): 501–508.
55. Kone, B.C., J. Schwöbel, P. Turner, M.G. Mohaupt, and C.B. Cangro.1995. Role of NF-kappa B in the regulation of inducible nitric oxidesynthase in anMTAL cell line. American Journal of Physiology 269(5Pt 2): 718–729.
56. Benbarek, H., A. Mouithys Mickalad, G. Deby-Dupont, C. Deby, S.Grulke, A. Nemmar, M. Lamy, and D. Serteyn. 1999. High concen-trations of histamine stimulate aquine polymorphonuclear neutrophisto produce reactive oxygen species. Inflammation Research 48(11):594–601.
57. Black, J.W., D.A. Owen, and M.E. Parsons. 1975. An analysis of thedepressor responses to histamine in the cat and dog: involvement ofbothH1- and H2-receptors. British Journal of Pharmacology 54: 319–324.
Involvement of H4 Receptor in Scorpion Envenomation Pathogenesis
58. Tamaoki, J., J. Nakata, K. Takeyama, A. Chiotani, and K. Konno.1997. Histamine H2 receptor mediated airway goblet cell secretionand its modulation by histamine-degrading enzymes. Journal of Al-lergy and Clinical Immunology 99: 233–238.
59. Yoshioka, T., W.W. Monafo, V.H. Ayvazian, F. Detz, and D. Flynn.1978. Cimetidine inhibits burn edema formation. The American Jour-nal of Surgery 136: 681–685.
60. Idzko, M., A. La Sala, D. Ferrari, E. Panther, Y. Herouy, S. Dichmann,et al. 2002. Expression and function of histamine receptors in humanmonocyte-derived dendritic cells. Journal of Allergy and ClinicalImmunology 109: 839–846.
61. Fontaine, C., and P. Demoly. 2006. Histamine and dendritic cells. RevueFrançaise d'Allergologie et d'Immunologie Clinique 46: 480–483.
62. Yamaguchi, M., C.S. Lantz, H.C. Oettgen, et al. 1997. IgE enhancesmouse mast cell Fc (Epsilon) RI expression in vitro and in vivo:evidence for a novel amplification mechanism in IgE-dependent reac-tions. Journal of Experimental Medicine 185: 663–672.
63. Gambero, A., E.C.T. Landucci, M.H. Toyama, S. Marangoni, J.R.Giglio, H.B. Nader, C.P. Dietrich, G. De Nucci, and E. Antunes.2002. Human neutrophil migration induced by secretory phospholi-pases A2: a role for cell surface glycosaminoglycans. BiochemicalPharmacology 63(1): 65–72.
64. Schmidt, J., S. Fleissner, I. Heimann-Weitschat, R. Lindstaedt, and I.Szelenyi. 1994. Histamine increases anti-CD3 induced IL-5 produc-tion of TH2-type Tcells via histamine H2-receptors.PubMed 42(3–4):81–85.
65. Krouwels, F.H., B.E.A. Hol, R. Lutter, et al. 1998. Histamine affectsinterleukin-4, interleukin-5, and interferon-gamma production by hu-man T cell clones from the airways and blood. American Journal ofRespiratory Cell and Molecular Biology 18: 721–730.
66. Ermolov, A.S, Pakhomova, G.V, Taeritneva, L.F, Matveev, S.B,Marchenko, V.V., Golikov, P.P. 1995. Effect of the histamine H2-receptor antagonist Zantac on lipid peroxidation an antioxidant systemin patients with gastroduodenal hemorrhage of ulcer etiology. PatolFiziol Eksp Ter. (1) : 23-25.
67. Cannon, K.E., R. Leurs, and L.B. Hough. 2007. Activation of periph-eral and spinal histamine H3 receptors inhibits formalin-induced in-flammation and nociception, respectively. Pharmacology, Biochemis-try, and Behavior 88: 122–129.
68. Raible, D.G., T. Lenahan, Y. Fayvilevich, et al. 1994. Pharmacologiccharacterization of a novel histamine receptor on human eosinophils.American Journal of Respiratory and Critical Care Medicine 149(6):1506–1511.
69. Hofstra, C.L., P.J. Desai, R.L. Thurmond, and W.P. Fung-Leung.2003. Histamine H4 receptor mediates chemotaxis and calcium mo-bilization of mast cells. The Journal of Pharmacology and Experi-mental Therapeutics 305: 1212–1221.
70. Gonzalez-Espinosa, C. 2010. Fyn kinase controls Fc_RI receptor-operated calcium entry necessary for full degranulation in mast cells.Biochemical and Biophysical Research Communications 391: 1714–1720.
71. Bäumer, W., S. Wendorff, R. Gutzmer, T. Werfel, D. Dijkstra, P.Chazot, H. Stark, and M. Kietzmann. 2008. Histamine H4 recep-tors modulate dendritic cell midration through skin-immunomodu-latory role of histamine. Allergy 63(10): 1387–1394. ISSN 1398-9995.
72. Shiraishi, Y., Takeda, K., Jia, Y., Domenico, J., Thurmond, R. L.,Karasuyama, H., Gelfand, E.W. 2011.Basophils contribute to allergicrhinitis through engagement of the histamine H4 receptor C34. Neu-trophils, mast cells, basophils and eosinophils in lung disease. pp.A4364
73. Ling, P., K. Ngo, S. Nguyen, R.L. Thurmond, J.P. Edwards, L.Karlsson, and W.P. Fung-Leung. 2004. Histamine H4 receptor medi-ates eosinophil chemotaxis with cell shape change and adhesionmolecule upregulation. British Journal of Pharmacology 142: 161–171.
74. Strakhova, M.I., C.A. Cuff, A.M. Manelli, et al. 2009. In vitro andin vivo characterization of A-940894: a potent histamine H4 receptorantagonist with anti-inflammatory properties. British Journal of Phar-macology 157: 44–54.