• By 1974, mRNA from a variety of eukaryotic species and viruses were found to be methylated
• A significant amount of this methylation was clustered at the 5’-end of mRNA
• This methylation cluster formed a structure we call a cap
15-3
Cap Structure
• Early study used viral mRNA as they are easier to purify and investigate
• The -phosphate of a nucleoside triphosphate remains only in the first nucleotide in an RNA– Cap is at the 5’-terminus of RNA– The cap is made of 7-methylguanosine, m7G– Linkage is a triphosphate– Charge on the cap area is near -5
15-4
Reovirus Cap Structure• The m7G contributes
a positive charge• Triphosphate linkage
contributes 3 negative charges
• Phosphodiester bond contributes 1 negative charge
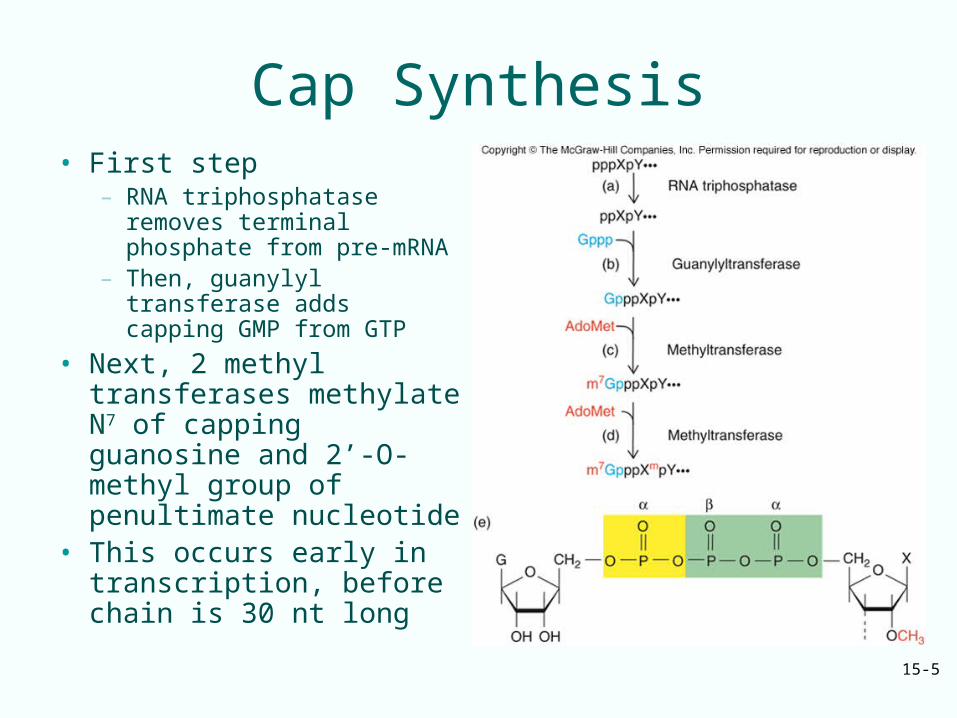
– RNA triphosphatase removes terminal phosphate from pre-mRNA
– Then, guanylyl transferase adds capping GMP from GTP
• Next, 2 methyl transferases methylate N7 of capping guanosine and 2’-O-methyl group of penultimate nucleotide
• This occurs early in transcription, before chain is 30 nt long
15-6
Functions of Caps
Caps serve at least four functions:– Protect mRNAs from degradation– Enhance translatability of mRNAs– Transport of mRNAs out of nucleus – Efficiency of splicing mRNAs
15-7
15.2 Polyadenylation
• The process of adding poly(A) to RNA is called polyadenylation
• A long chain of AMP residues is called poly (A)
• Heterogeneous nuclear mRNA is a precursor to mRNA
15-8
Poly(A)
• Most eukaryotic mRNAs and their precursors have a chain of AMP residues about 250 nt long at their 3’-ends
• Poly(A) is added posttranscriptionally by poly(A) polymerase
15-9
Functions of Poly(A)
• Poly(A) enhances both the lifetime and translatability of mRNA
• Relative importance of these two effects seems to vary from one system to another
• In rabbit reticulocyte extracts, poly(A) seems to enhance translatability by helping to recruit mRNA to polysomes
15-10
Basic Mechanism of Polyadenylation
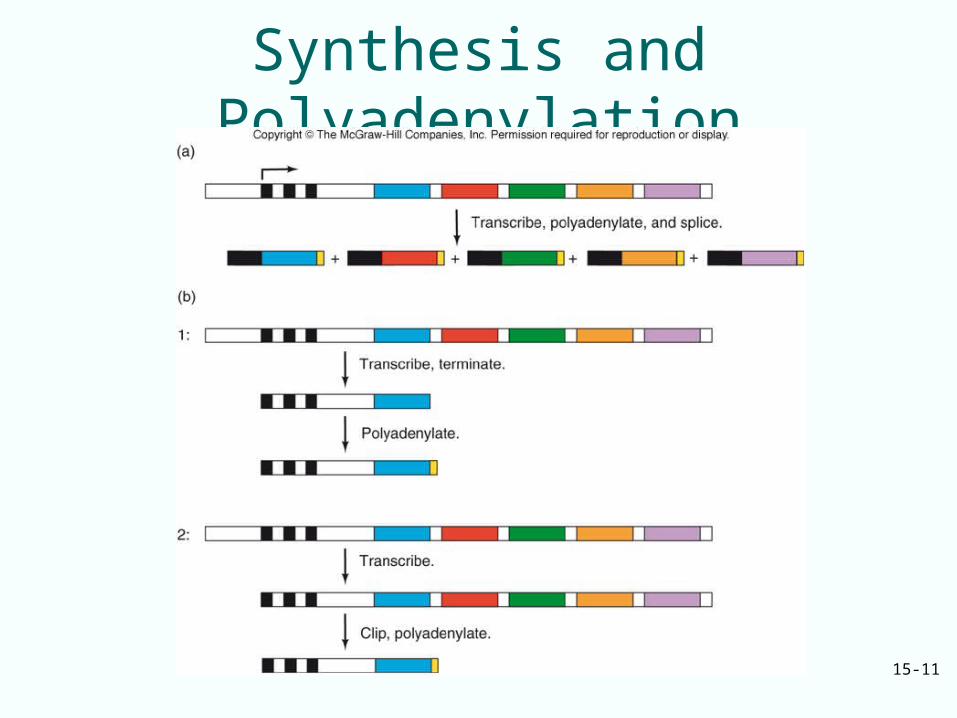
• Transcription of eukaryotic genes extends beyond the polyadenylation site
• The transcript is: – Cleaved– Polyadenylated at 3’-
end created by cleavage
15-11
Synthesis and Polyadenylation
15-12
Polyadenylation Signals• An efficient mammalian polyadenylation signal
consists of:– AAUAAA motif about 20 nt upstream of a
polyadenylation site in a pre-mRNA– Followed 23 or 24 bp later by GU-rich motif– Followed immediately by a U-rich motif
• Variations on this theme occur in nature– Results in variation in efficiency of polyadenylation– Plant polyadenylation signals usually contain
AAUAAA motif– More variation exists in plant than in animal motif– Yeast polyadenylation signals are even more different
15-13
Cleavage of Pre-mRNA
• Polyadenylation involves both:– Pre-mRNA cleavage– Polyadenylation at the cleavage site
• Cleavage in mammals requires several proteins– CPSF – cleavage and polyadenylation specificity
Initiation of Polyadenylation• Short RNAs mimic a newly created mRNA 3’-
end can be polyadenylated• Optimal signal for initiation of such
polyadenylation of a cleaved substrate is AAUAAA followed by at least 8 nt
• When poly(A) reaches about 10 nt in length, further polyadenylation becomes independent of AAUAAA signal and depends on the poly(A) itself
• 2 proteins participate in the initiation process– Poly(A)polymerase– CPSF binds to the AAUAAA motif
15-15
Elongation of Poly(A)• Elongation of poly(A) in mammals requires
a specificity factor called poly(A)-binding protein II (PAB II)
• This protein – Binds to a preinitiated oligo(A)– Aids poly(A) polymerase in elongating poly(A)
to 250 nt or more
• PAB II acts independently of AAUAAA motif– Depends only on poly(A)– Activity enhanced by CPSF
15-16
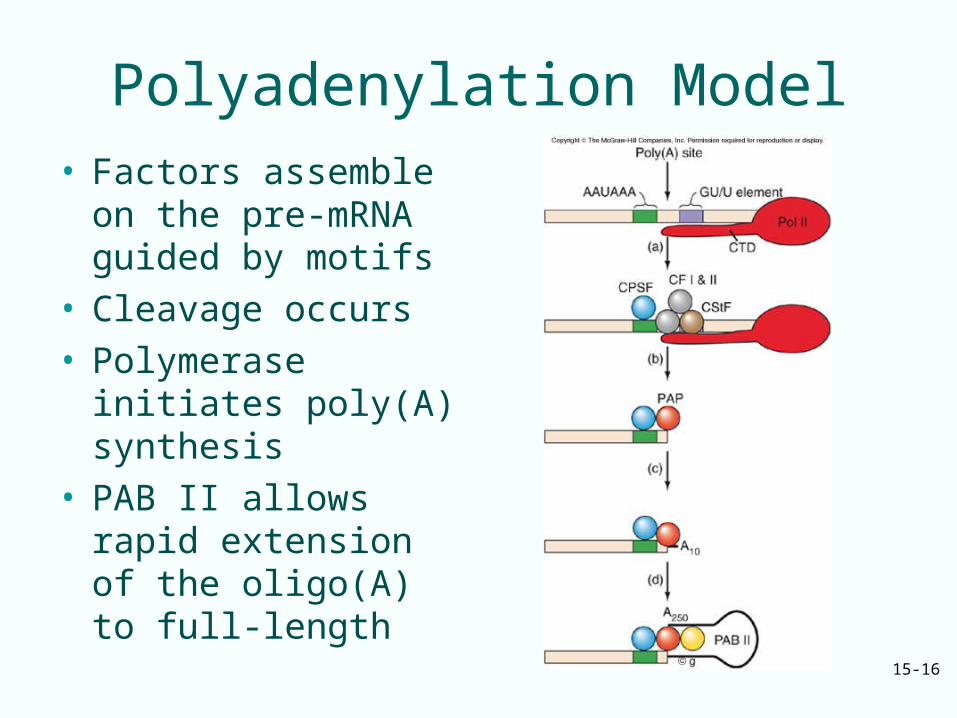
Polyadenylation Model• Factors assemble on
the pre-mRNA guided by motifs
• Cleavage occurs• Polymerase initiates
poly(A) synthesis• PAB II allows rapid
extension of the oligo(A) to full-length
15-17
Poly(A) Polymerase• Cloning and sequencing cDNAs encoding
calf thymus poly(A) polymerase reveal a mixture of 5 cDNAs derived from alternative splicing and alternative polyadenylation
• Structures of the enzymes predicted from the longest sequence includes: – RNA-binding domain– Polymerase module– 2 nuclear localization signals– Ser/Thr-rich region – this is dispensable for
activity in vitro
15-18
Turnover of Poly(A)
• Poly(A) turns over in the cytoplasm
• RNases tear it down
• Poly(A) polymerase builds it back up
• When poly(A) is gone mRNA is slated for destruction
15-19
Cytoplasmic Polyadenylation
• Cytoplasmic polyadenylation is most easily studied using Xenopus oocyte maturation
• Maturation-specific polyadenylation of Xenopus maternal mRNAs in the cytoplasm depends on 2 sequence motifs:– AAUAAA motif near the end of mRNA – Upstream motif called the cytoplasmic
polyadenylation element (CPE)• UUUUUAU• Or closely related sequence
15-20
15.3 Coordination of mRNA Processing Events
• After reviewing capping, polyadenylation and splicing, it is clear that these processes are related
• Cap can be essential for splicing, but only for splicing the first intron
• Poly(A) can also be essential, but only for splicing out the last intron
15-21
Effect of Cap on Splicing• Removal of the first intron fro model pre-mRNAs
in vitro is dependent on the cap• This effect may be mediated by a cap-binding
complex involved in spliceosome formation
15-22
Effect of Poly(A) on Splicing
• Polyadenylation of model substrates in vitro is required for active removal of the intron closest to the poly(A)
• Splicing any other introns out of these substrates occurs at a normal rate even without polyadenylation
15-23
mRNA-Processing Occurs During Transcription
All three of the mRNA-processing events take place during transcription
– Splicing begins when transcription is still underway
– Capping • When nascent mRNA is about 30 nt long• When 5’-end of RNA first emerges from
polymerase
– Polyadenylation occurs when the still-growing mRNA is cut at the polyadenylation site
15-24
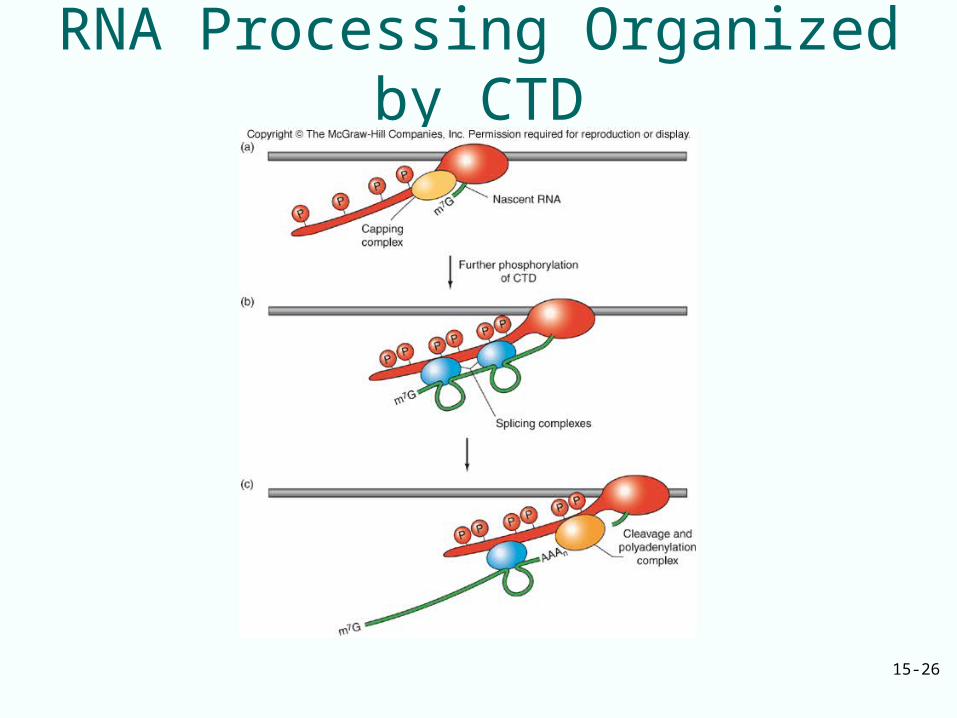
Binding of CTD of Rpb1 to mRNA-Processing Proteins
• The CTD of Rpb1 subunit of RNA polymerase II is involved in all three types of processing
• Capping, polyadenylating, and splicing enzymes bind directly to the CTD which serves as a platform for all three activities
15-25
CTD Phosphorylation
• Phosphorylation state of the CTD of Rpb1 in transcription complexes in yeast changes as transcription progresses– Transcription complexes close to the promoter
contain phosphorylated Ser-5– Complexes farther from the promoter contain
phosphorylated Ser-2
• Spectrum of proteins associated with the CTD also changes– Capping guanylyl transferase is present early when
the complex is close to promoter, not later– Polyadenylation factor Hrp1 is present in transcription
complexes near and remote from promoter
15-26
RNA Processing Organized by CTD
15-27
Coupling Transcription Termination with End Processing• An intact polyadenylation site and active
factors that cleave at the polyadenylation site are required for transcription termination
• Active factors that polyadenylate a cleaved pre-mRNA
15-28
Mechanism of Termination
• Termination of transcription by RNA polymerase II occurs in 2 steps:– Transcript experiences a cotranscriptional
cleavage (CoTC) within termination region downstream of the polyadenylation site
• This occurs before cleavage and polyadenylation at the poly(A) site
• It is independent of that process
– Cleavage and polyadenylation occur at the poly(A) site
• Signals polymerase to dissociate from template
15-29
Termination Signal
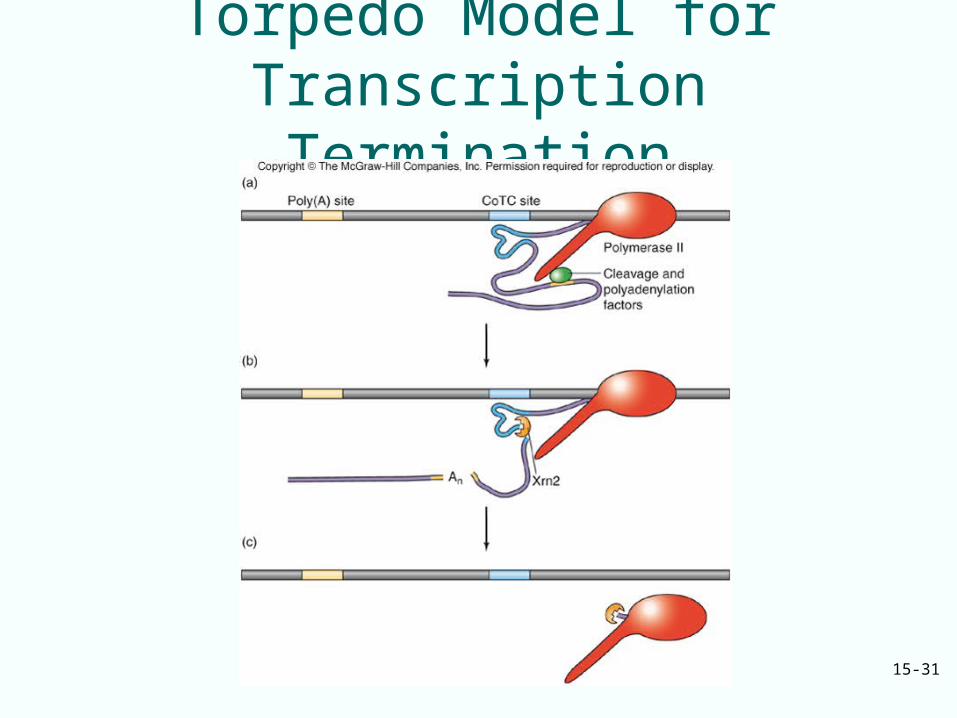
• CoTC element downstream of the polyadenylation site in the human b-globin mRNA is a ribozyme that cleaves itself– This generates a free RNA 5’-end– This cleavage is required for normal
transcription termination– It provides an entry site for Xrn2, a 5’3’
exonuclease that loads onto the RNA and chases RNA polymerase by degrading the RNA
15-30
Xrn2, Exonuclease
• Xrn2 terminates transcription like a “torpedo”
• There is a similar torpedo mechanism in yeast where cleavage at poly(A) site provides entry for the 5’3’ exonuclease Rat1
• Rat1 degrades the RNA until it catches the polymerase and terminates transcription
15-31
Torpedo Model for Transcription Termination
15-32
Role of Polyadenylation in mRNA Transport
• Polyadenylation is required for efficient transport of mRNAs from their point of origin in the nucleus to the cytoplasm