Page 1
Indian Journal of Experimental Biology
Vol. 56, June 2018, pp. 373-384
Molecular characterization of microsymbionts
associated with root nodules of Crotalaria burhia Buch.-Ham. ex Benth.,
a native keystone legume species from Thar Desert of India
Indu Singh Sankhla1,2
, Raju Ram Meghwal1, Sunil Choudhary
1,3, Sonam Rathi
1, Nisha Tak
1,
Alkesh Tak1 & Hukam Singh Gehlot
1*
1BNF and Microbial Genomics Laboratory, Department of Botany, Jai Narain Vyas University, Jodhpur-342 033, Rajasthan, India 2Department of Botany, University of Rajasthan, Jaipur-302 004, Rajasthan, India 3ICFRE-Arid Forest Research Institute (AFRI), Jodhpur-302 005, Rajasthan, India
Received 19 April 2016; revised 14 July 2017
Establishment of legume-rhizobia symbiosis has ample agronomic and ecological significance. Characterization of native
rhizobia could enhance our understanding of their natural distribution and co-evolution. The Great Indian Thar Desert is an
ecologically significant unique habitat with its flora and fauna. Crotalaria spp. is an economically important legume widely
distributed in the Thar Desert and can be considered its one of the bioresources, particularly for biological nitrogen fixation
with their symbiotic rhizobia. Here, we examined the legume Crotalaria burhia Buch.-Ham. ex Benth. in search of potential
novel rhizobial species. Out of 72 root nodule bacterial (RNB) strains isolated from C. burhia, 51 rhizobia-like strains were
examined for genetic diversity based on ARDRA and RAPD patterns. BLASTn sequence similarity results based on 16S
rRNA gene of selective thirteen strains representing four ARDRA types revealed that they were related to genera Ensifer,
Rhizobium and Bradyrhizobium. In 16S rRNA gene phylogeny, five (CB5, CB17, CB36, CB44, CB56) strains were closer
to Ensifer kostiensis, three (CB6, CB12, CB32) to E. terangae and CB11 showed similarity with E. kostiensis and E. saheli.
Strain CB4 was similar to Bradyrhizobium yuanmingense and three (CB29, CB31, CB46) strains were closer to species of
Rhizobium (R. etli, R. sullae and R. borbori respectively). Symbiotic (nodA and nifH) genes phylogeny of Ensifer sp. CB56
was incongruent and showed close similarity with E. fredii whereas sym gene phylogeny of Bradyrhizobium sp. CB4 was
congruent with 16S rRNA gene phylogeny. In Rhizobium strains sym genes could not be amplified and they failed to
nodulate host. Our study suggests that C. burhia is nodulated by diverse strains of Ensifer and Bradyrhizobium in alkaline
soil of Thar Desert and these strains effectively cross-nodulated crop Vigna radiata.
Keywords: ARDRA, Bradyrhizobium, Burhia Rattlepod, Ensifer, nifH, Nitrogen fixation, nodA, Nodulation, Root nodule
bacterial (RNB) strains
Legume-rhizobia symbiosis known for biological
nitrogen fixation have long term agronomical and
ecological significance and is one of the alternative to
nitrogen fertilizers. Rhizobia are Gram negative,
aerobic, soil bacteria with ability to mostly form N
fixing nodules on roots of legumes1. Most of the
legume-rhizobia symbiosis studies have been carried
out on either agricultural crops (soybean, mungbean,
pea and chickpea) or pasture legumes (clover and
alfalfa). From last two decades such studies have been
extended to wild/native legumes from tropical and
semi-tropical climatic zones that resulted in
discovering several novel species and genera belonging
to class alphaproteobacteria and betaproteobacteria2.
The genus Crotalaria L. is the third largest in
Papilionoideae subfamily of Leguminosae having
more than 700 species3. It is distributed in tropical and
sub-tropical regions of the world with the majority of
species (543) native to tropical Africa and
Madagascar. In India, it is the largest legume taxa
comprising 93 species of which 27 are endemic4.
Many species of Crotalaria have great economic
importance in terms of insecticides, fibres, silage and
green manure having agronomic values4,5
. Crotalaria
burhia Buch.-Ham. ex Benth., (Burhia Rattlepod)
locally called ‘Shinio’, is a perennial bushy legume
widely distributed on sandy areas throughout the Thar
Desert of India and adjoining Pakistan. The Indian
Thar Desert is characterized by erratic precipitation,
high irradiation, high temperature and saline tracts6.
Overall, the area is alkaline and the soil has been
________
*Correspondence:
Phone: +91 94141 24939 (Mob.)
E-mail: [email protected]
Page 2
INDIAN J EXP BIOL, JUNE 2018
374
classified as ‘desert soil’ poor in nitrogen and available
phosphorus. Therefore the plants, animals, and microbes
growing in Thar Desert are continuously under biotic
and abiotic stresses. Gehlot et al.7 suggested some
traditional and modern scientific approaches for
characterization, conservation and sustainable utilization
of bioresources of the Thar Desert. Nodulation in
number of wild/native legumes of Thar Desert have
been reported as well as root nodule microsymbionts of
several legumes have been characterized at molecular
and genomic level recently8-19
.
The C. burhia is one of the keystone species of
Thar Desert and has a great potential in terms of
restoration of soil fertility through nitrogen fixation
and also in prevention of desertification through its
soil binding and sand dune stabilizing properties,
hence it have great ecological importance.
Crotalaria has been reported to be nodulating by
Bradyrhizobium strains20
. However, Crotalaria
podocarpa is nodulated by a novel rhizobia
Methylobacterium nodulans21
. Furthermore, Liu et al.22
isolated Rhizobium and Bradyrhizobium strains from
nodules of C. pallida in China, while Rocha23
isolated
Mesorhizobium sp. from root nodules of
C. spectabilis. On the basis of Amplified r-DNA
Restriction Analysis (ARDRA) Singha et al.24
identified
three groups of rhizobial strains (Rhizobium,
Bradyrhizobium and Mesorhizobium) from C. pallida in
Assam, India. Sankhla et al.12
reported that
C. medicagenia is nodulated by Ensifer strains in sandy
areas of Indian Thar Desert. In this study, we
investigated genetic diversity of root nodule bacteria
(RNB) associated with C. burhia and characterized its
rhizobia at molecular level including symbiotic genes.
Materials and Methods
Nodule sampling and isolation of rhizobia
Root nodules of C. burhia were collected from 25
sites in five districts of Western Rajasthan (Table 1).
The nodules were collected during the monsoon and
Table 1 — Nodulation status of Crotalaria burhia growing at different sampling sites throughout Thar Desert and
origin of root nodule bacterial (RNB) strains
District Soil sampling site Geographical coordinates pH OC
(%)
Total N
(%)
P
(kg/ha)
Avg. no. of
nodules per plant
Purified root nodule
bacterial strains
Jodhpur Amritlal stadium 26°20'25.40"N/73° 3'3.09"E 8.2 0.16 0.0078 9.2 08 CB1, CB2, CB45, CB46r,
CB47, CB48
JNVU New
Campus
26°14'49.85"N/73°
1'18.65"E
8.2 0.18 0.0091 12.4 09 CB5e, CB7, CB27, CB28,
CB29r, CB30
Kailana 26°18'5.83"N/72°58'29.38"E 8.2 0.10 0.0052 8.2 09 CB8, CB55
Osian 26°44'26.57"N/72°53'49.26"E 8.9 0.14 0.0071 18.4 14 CB38, CB39, CB40, CB42, CB44e
Pratapnagar 26°17'29.28"N/73° 0'6.36"E 8.8 0.23 0.0048 16.3 09 CB49, CB50, CB51
Shergarh 26°19'10.58"N/72°18'1.42"E 8.7 0.19 0.0112 9.2 09 CB67, CB68
Nagaur Alai 27°19'24.91"N/73°35'6.99"E 8.2 0.24 0.0092 6.4 08 CB10, CB61
Baghnada 27° 8'42.94"N/73°48'32.42"E 8.2 0.26 0.0111 11.2 11 CB6e, CB15, CB16
Deh 27°18'30.40"N/73°54'53.51"E 8.4 0.28 0.0069 8.2 10 CB3, CB4b
Harima 27°15'26.84"N/73°51'39.74"E 8.5 0.29 0.0071 7.8 14 CB21, CB22, CB23
Inana 27° 8'2.63"N/73°49'39.20"E 8.1 0.22 0.0114 10.8 12 CB18, CB19, CB20
Tausar 27° 6'36.85"N/73°46'30.60"E 8.4 0.23 0.0121 9.8 10 CB24, CB25, CB26
Barmer Barmer 25°45'10.09"N/71°26'5.31"E 8.2 0.14 0.0079 10.2 08 CB33, CB43
Bhuka 25°39'54.66"N/72° 0'54.03"E 8.8 0.13 0.0095 9.2 09 CB31r, CB32e, CB34
Chohtan 25°29'44.82"N/71° 5'6.90"E 8.1 0.16 0.0048 11.2 05 CB35, CB36e, CB37
Nimdee 26° 4'2.97"N/71°19'48.46"E 8.6 0.18 0.0081 11.9 07 CB9, CB11e, CB12e
Jaisalmer Chandhan 26°59'18.13"N/71°18'13.62"E 8.8 0.17 0.0117 6.4 07 CB52, CB41
Jaisalmer 26°53'49.55"N/70°56'31.18"E 8.3 0.19 0.0050 6.9 07 CB63, CB64
Kuldhera 26°52'28.36"N/70°46'34.28"E 8.6 0.19 0.0094 5.5 09 CB56e, CB57
Longewala 27°30'8.57"N/70° 7'28.76"E 8.5 0.29 0.0098 7.1 07 CB65, CB66
Pokaran 26°58'25.49"N/71°54'6.92"E 8.7 0.22 0.0069 5.1 05 CB71, CB72
Sum 26°50'31.14"N/70°32'53.19"E 8.1 0.11 0.0069 8.7 09 CB58, CB62, CB69, CB70
Bikaner Bikaner 28° 1'49.04"N/73°15'30.63"E 7.9 0.21 0.0047 10.2 09 CB53, CB54
Deshnok 27°47'20.09"N/73°20'22.46"E 8.7 0.21 0.0117 11.9 10 CB13, CB14, CB17e
Nokha 27°34'11.22"N/73°27'36.13"E 9.0 0.29 0.0092 11.2 08 CB59, CB60
[CB, Crotalaria burhia; Strains identified as eEnsifer sp., bBradyrhizobium sp. and rRhizobium sp. on the basis of 16S rRNA gene sequences]
Page 3
Sankhla et al.: CHARACTERIZATION OF MICROSYMBIONTS ASSOCIATED WITH CROTALARIA ROOT NODULES
375
post-monsoon season (July-October) in the years
2011, 2012 and 2013. The Flora of the Indian Desert6
and regional Botanical Survey of India were referred
for identification of C. burhia plants in the field.
Whole plant were excavated with intact root system
and then thoroughly washed with tap water. The
nodulation status of excavated plants was recorded
and 4-5 root nodules with roots were kept in moist
soil and brought to the laboratory for preservation of
nodules and isolation of rhizobia. The viable seeds of
C. burhia were also collected to perform host
authentication experiments. Root nodules of C. burhia
were surface sterilized and bacterial strains were
isolated and purified according to standard procedure
as described by Vincent25
and Somasegaran and
Hoben26
. All bacterial strains were cultured on Congo
Red-Yeast Extract Mannitol Agar (CR-YEMA) plates
and incubated at 28ºC. The CR-YEM agar plates were
checked regularly to record colony characteristics of
isolates and to confirm their purity.
Soil sampling and analysis
Rhizosphere soil of C. burhia was collected from
each sampling sites at the time of nodule sampling for
chemical analysis. Chemical properties of the
collected soil such as pH (aqueous extract), total
nitrogen (N), organic carbon (OC) and available
phosphorus (P) were determined by using various
standard methods described in Gehlot et al.8.
Molecular characterization
Isolation of genomic DNA
Purified bacterial strains were grown in YEM broth
and were used for isolation of genomic DNA using
method described by Cheng and Jiang27
. Bacterial
cells were washed with TE (Tris-EDTA) and STE
(Sodium chloride-Tris-EDTA) buffer, and lysed
directly by adding phenol. The supernatant was
treated with chloroform to remove traces of phenol.
The supernatant (containing DNA) was used as
template for amplification of 16S rRNA and
symbiotic (nodA and nifH) genes through thermo
cycler (BioRad T100).
Amplification of 16S rRNA gene
Table 2 enlists various primers used in this study
for molecular characterization. Nearly full length 16S
rRNA gene of selected bacterial strains was amplified
using universal primers (18F and 1492R)28
. Each
reaction was carried out in a final volume of 20 μL
containing: 1 μL of template DNA (90 ng/μL), 0.6 U
of Taq DNA polymerase, 1.2 μM of each of the
primers, 2 mM MgCl2, 150 μM of each dNTP and 1X
PCR buffer. The PCR temperature profiles were as
follows: initial denaturation at 94ºC for 5 min
followed by 35 cycles of 94ºC for 30 s, 53ºC for 30 s,
72ºC for 60 s and a final extension at 72ºC for 5 min.
Amplified PCR products along with 500 bp marker
were run on 0.89% (w/v) agarose gel, prepared in 1X
TAE buffer and pre-stained with ethidium bromide.
The bands were visualized under BIO-RAD Gel Doc
System (Bio Rad SR+, USA Inc.). The amplified PCR
products were quantified using Denovix (USA) DS11
spectrophotometer (nanodrop).
Molecular fingerprinting
Genetic variability among bacterial strains was
studied using following DNA fingerprinting patterns:
ARDRA (Amplified r-DNA Restriction Analysis) pattern
The restriction endonucleases MspI (Genei
Bangalore) was used to digest the amplified PCR
products (16S rRNA gene). The reactions were carried
out in a final volume of 20 μL containing 2 μL of 10X
buffer, 0.25 μL of MspI enzyme (10U/μL), 10 μL
aliquots of PCR products and 7.75 μL of nuclease free
water. The reaction mixture was incubated at 37ºC for
overnight. Digested PCR products along with 100 bp
DNA ladder (Genei Bangalore) were run on 2.0%
agarose gel at 80 V for 1-2 h for separation of restricted
fragments and were visualized by staining with ethidium
bromide using BIO-RAD Gel Doc System.
RAPD (Random Amplification of Polymorphic DNA) pattern
RPOI primer (nif gene directed primer widely used
for study of genetic diversity) was used to randomly
amplify genomic DNA as described by Richardson et al.29
.
The reactions were carried out in a final volume of
Table 2 — List of primers used in this study for
molecular characterization
Primer Oligonucleotide sequence (5'→ 3')a Reference
RPOI AATTTTCAAGCGTCGTGCCA 30
18F AGAGTTTGATCCTGGCTCAG 29
1492R CTACGGCTACCTTGTTACG 29
800F GTAGTCCACGCCGTAAACGA 31
820R CATCGTTTACGGCGTGGACT 31
nodA1 TGCRGTGGAARNTRNNCTGGGAAA 32
nodA2 GGNCCGTCRTCRAAWGTCARGTA 32
nifHF TACGGNAARGGSGGNATCGGCAA 33
nifHI AGCATGTCYTCSAGYTCNTCCA 33
[aA, C, G, T =standard nucleotides; N=A, C, G or T; R=A or G;
Y= C or T; S=G or C and W=A, T]
Page 4
INDIAN J EXP BIOL, JUNE 2018
376
20 μL containing: 1.5 μL of template DNA (90 ng/μL),
1U of Taq DNA polymerase, 3 μM of RPOI primer,
3.75 mM MgCl2, 150 μM of each dNTP and 1X PCR
buffer. The thermal cycling condition were as
follows: initial denaturation at 94ºC for 5 min
followed by 5 cycles at 94ºC for 30 s, 50ºC for 60 s,
72ºC for 90 s and then 30 cycles at 94ºC for 30 s,
55ºC for 25 s and 72ºC for 90 s and a final extension
at 72ºC for 5 min. The amplified DNA fragments
were resolved on 2.0% agarose gel at 80 V for 1-2 h
along with DNA ladder of 100 bp and generated
fingerprints were visualized under BIO-RAD Gel Doc
System. Sequencing of 16S rRNA gene
On the basis of ARDRA and RAPD patterns, 13
isolates were selected for sequencing of their 16S
rRNA gene. Amplified sample in adequate quantity
(50 μL having 20-30 ng/μL DNA) were sent to
Xcleris Genomics Labs Ltd., Ahmedabad for
sequencing. Universal external (18F and 1492R)28
and
internal (800F and 820R)30
primers were used to
obtain nearly full length (1.5 kb) nucleotides sequence
of 16S rRNA gene. The sequencing was done on
Applied Biosystems platform using Big Dye version
3.1 terminator and 5X buffer. Gene Tool Lite 1.0
(2000) software (Doubletwist, Inc., Oakland, CA,
USA) was used for analysis of all raw sequences.
Sequence similarity searches for nucleotide
sequences were performed at the National Centre for
Biotechnology Information (NCBI) server using basic
local alignment search tool (BLAST). After
completion of analysis, sequences were submitted to
NCBI database using Sequin.
Nodulation test
Selective strains were evaluated for their ability to
form nodules on their original host (C. burhia) as well
as crop legume Vigna radiata. Seeds of C. burhia
were scarified and surface sterilized by treatment with
95% (v/v) ethanol for 2 min and 0.1% HgCl2 for 3 min
followed by several washes with sterile distilled water
and then allowed to germinate in petri plates,
containing sterilized moist filter paper. Three to four
germinated seedlings were planted aseptically into
each plastic pot containing washed and sterilized
sand. Each seedling was inoculated with 1 mL
(109cells/mL) of bacterial strains at exponential
growth phase. Un-inoculated seedlings either supplied
with mineral nitrogen (as 0.1% KNO3 in nutrient
solution) or grown without nitrogen served as N+ and
N- controls, respectively. The entire experimental set
up of inoculated and controls plants were designed in
triplicates. Seedlings were grown under natural
sunlight and temperature (28±2ºC day and night) in
glass house. Similar procedure was applied for
V. radiata except scarification of seeds. Plants were
harvested and checked for nodulation after 6-8 weeks
of inoculation as described by Somasegaran and
Hoben26
. Rhizobia were re-isolated and purified from
root nodules of inoculated plants and compared with
the parental strains on the basis of colony morphology
and genetic fingerprints (RAPD pattern) using RPOI
primer29
.
Amplification and sequencing of symbiotic genes (nodA and nifH)
In selected isolates nodA and nifH genes were
amplified. Primers nodA1 and nodA231
were used for
PCR amplification of an internal fragment of size
650 bp of nodA gene (codes for N-acyl transferase
nodulation protein). The reactions were carried out in
a final volume of 20 μL containing: 1 μL of template
DNA, 0.6U of Taq DNA polymerase, 1 μM of each of
the primers, 3.12 mM MgCl2, 150 μM of each dNTP,
and 1X PCR buffer. PCR cycling conditions were as
follows: initial denaturation at 94ºC for 5 min, 5
cycles at 94ºC for 30 s, 55ºC for 30 s and 72ºC for
60 s followed by 30 cycles at 94ºC for 30 s, 62ºC for
45 s, 72ºC for 90 s and a final extension at 72ºC for
7 min. Amplification of nifH region (750 bp codes for
Fe protein of nitrogenase enzyme) was carried out
using primers nifHF and nifHI32
and the reaction
mixture was prepared same as for nodA. PCR cycling
conditions were as follows: initial denaturation at
94ºC for 5 min followed by 25 cycles at 94ºC for 30 s,
57ºC for 30 s, 72ºC for 30 s and a final extension at
72ºC for 7 min. PCR products along with DNA ladder
of 100 bp (Genei Bangalore) were run on 1.0%
agarose gel at 80 V for 1 h to confirm the
amplification of targeted gene. The amplified
products were sequenced with the corresponding
primers using an Applied Biosystems sequencer.
Phylogenetic analysis
Nucleotide sequences of RNB isolated from C. burhia
in this study were submitted in the GenBank database.
The accession numbers have been specified in the
corresponding phylograms. The multiple sequence
alignment program CLUSTALW33
was used to align
the nucleotide sequences obtained in present study
(16S rRNA and symbiotic genes sequences), together
with related sequences of type strains retrieved from
Page 5
Sankhla et al.: CHARACTERIZATION OF MICROSYMBIONTS ASSOCIATED WITH CROTALARIA ROOT NODULES
377
the NCBI database. The phylogenetic trees were
generated using MEGA 6 software34
with maximum
likelihood method based on a GTR+G+I model.
A bootstrap for 1000 replicates was performed to
obtain the confidence values for the tree topologies.
Results
Nodulation status and soil analysis
Soil samples and nodules of C. burhia were
collected from all the sampling sites in five districts of
Western Rajasthan (Table 1). The C. burhia plants
growing in natural habitat is shown in Fig. 1A-C; with
flower (Fig. 1D); collected seeds (Fig. 1E) and root
nodules (Fig. 1F). The nodulation in C. burhia was
observed at all sampling sites, although the average
number of nodules per plant varied from site to site.
The highest average number of nodules per plant was
recorded fourteen in Osian (Jodhpur) and Harima
(Nagaur) sampling sites (semi-arid) and lowest was
five in Chohtan (Barmer) and Pokaran (Jaisalmer),
both arid to hyper arid sites (Table 1). Initially,
nodules were globular but became elongated,
indeterminate and branched on maturity (Fig. 1F).
The morphology and internal structure of C. burhia
root nodules have been described in our previous
study8. Soil characteristics such as total N, P, organic
carbon and pH of various sampling sites were not
significantly different as descried earlier8. The pH of
soils in all sampling sites in Thar Desert was alkaline
and ranged from pH 7.9 to 9 (Table 1); however, soil
texture, annual rainfall (100-500 mm), mean daily
temperature in summer (25-35ºC) and winter (7-15ºC)
were found varied.
DNA fingerprinting (ARDRA and RAPD) and sequencing of
16S rRNA gene
A single band of approx. 1.5 kb of 16S rRNA gene
was amplified in all strains. The 51 rhizobial strains
formed four groups on the basis of the ARDRA
(Table 3). The largest ARDRA group-I had 42 strains
and group II had five strains which were identified as
species of Ensifer and Bradyrhizobium respectively,
on the basis of 16S rRNA gene sequencing and
BLASTn results. The group III and IV each
containing 2 strains were identified as species of
Rhizobium.
All the isolates showed considerable variation in
RAPD pattern and were distributed in 14 groups. The
largest ARDRA group-I that comprised of 42 strains
Fig. 1 — (A) The Crotalaria burhia plants growing in natural
habitat during rainy season; (B) in dry season; (C) closer view; (D)
a flower; (E) collected seeds; and (F) indeterminate root nodules.
Table 3 — Grouping of selective Crotalaria burhia-RNB strains
based on genetic fingerprinting
RPOI genetic
groups
Strains ARDRA genetic
groups
I CB7, CB8, CB11e, CB14, CB15,
CB17e, CB27, CB28, CB30,
CB40, CB42, CB72
I (42 strains)
II CB1, CB2, CB3, CB5e, CB44e
III CB6e, CB36e, CB59
IV CB10, CB60, CB61, CB62,
CB63, CB66
V CB54, CB55, CB56e, CB68
VI CB57, CB58, CB64, CB67,
CB69
VII CB32e,CB70, CB71
VIII CB52, CB53
IX CB12e
X CB20
XI CB4b, CB9, CB43, CB49, CB65 II (5 strains)
XII CB29r III (2 strains)
XIII CB46r
XIV CB31r, CB51 IV (2 strains)
[Strains identified as eEnsifer sp., bBradyrhizobium sp. and rRhizobium sp. on the basis of 16S rRNA gene sequences]
Page 6
INDIAN J EXP BIOL, JUNE 2018
378
(including nine sequenced Ensifer strains) was further
resolved into 10 RAPD groups (Table 3). This
suggests that RPOI based fingerprinting gives better
resolution of genetic diversity when compared with
ARDRA. Strains CB4, CB9, CB43, CB49 and CB65
showed similar banding patterns in both ARDRA and
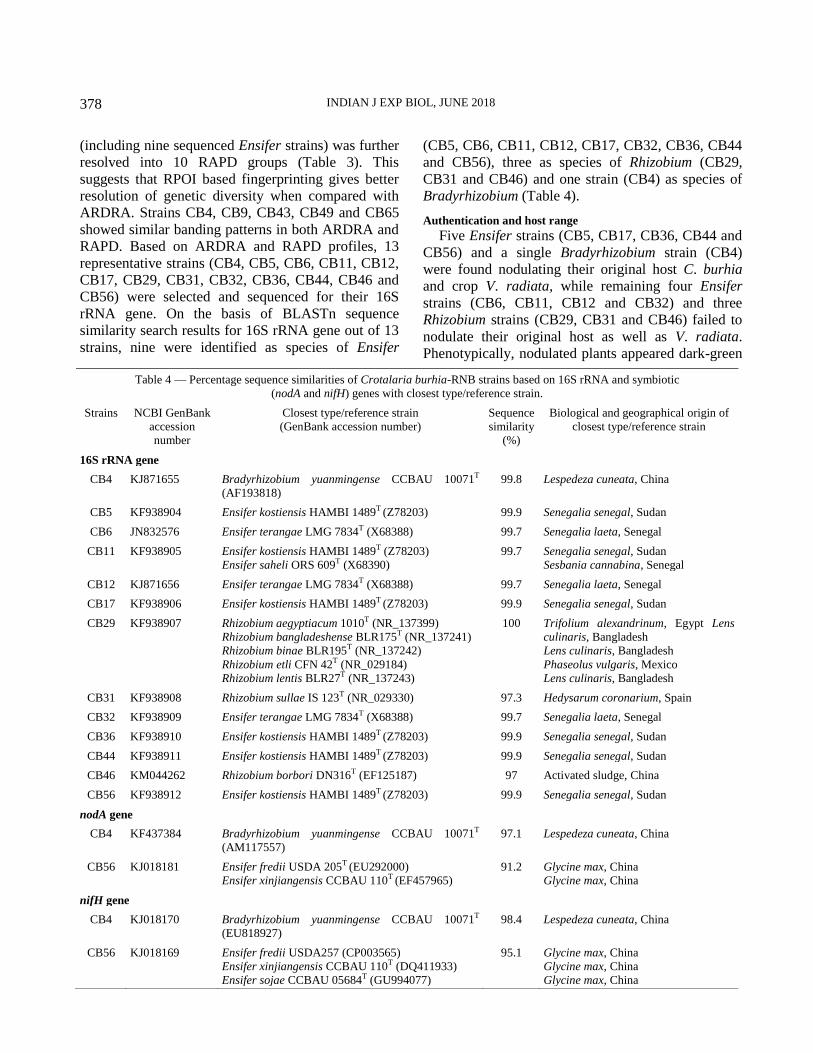
RAPD. Based on ARDRA and RAPD profiles, 13
representative strains (CB4, CB5, CB6, CB11, CB12,
CB17, CB29, CB31, CB32, CB36, CB44, CB46 and
CB56) were selected and sequenced for their 16S
rRNA gene. On the basis of BLASTn sequence
similarity search results for 16S rRNA gene out of 13
strains, nine were identified as species of Ensifer
(CB5, CB6, CB11, CB12, CB17, CB32, CB36, CB44
and CB56), three as species of Rhizobium (CB29,
CB31 and CB46) and one strain (CB4) as species of
Bradyrhizobium (Table 4).
Authentication and host range
Five Ensifer strains (CB5, CB17, CB36, CB44 and
CB56) and a single Bradyrhizobium strain (CB4)
were found nodulating their original host C. burhia
and crop V. radiata, while remaining four Ensifer
strains (CB6, CB11, CB12 and CB32) and three
Rhizobium strains (CB29, CB31 and CB46) failed to
nodulate their original host as well as V. radiata.
Phenotypically, nodulated plants appeared dark-green
Table 4 — Percentage sequence similarities of Crotalaria burhia-RNB strains based on 16S rRNA and symbiotic
(nodA and nifH) genes with closest type/reference strain.
Strains NCBI GenBank
accession
number
Closest type/reference strain
(GenBank accession number)
Sequence
similarity
(%)
Biological and geographical origin of
closest type/reference strain
16S rRNA gene
CB4 KJ871655 Bradyrhizobium yuanmingense CCBAU 10071T
(AF193818)
99.8 Lespedeza cuneata, China
CB5 KF938904 Ensifer kostiensis HAMBI 1489T (Z78203) 99.9 Senegalia senegal, Sudan
CB6 JN832576 Ensifer terangae LMG 7834T (X68388) 99.7 Senegalia laeta, Senegal
CB11 KF938905 Ensifer kostiensis HAMBI 1489T (Z78203)
Ensifer saheli ORS 609T (X68390)
99.7 Senegalia senegal, Sudan
Sesbania cannabina, Senegal
CB12 KJ871656 Ensifer terangae LMG 7834T (X68388) 99.7 Senegalia laeta, Senegal
CB17 KF938906 Ensifer kostiensis HAMBI 1489T (Z78203) 99.9 Senegalia senegal, Sudan
CB29 KF938907 Rhizobium aegyptiacum 1010T (NR_137399)
Rhizobium bangladeshense BLR175T (NR_137241)
Rhizobium binae BLR195T (NR_137242)
Rhizobium etli CFN 42T (NR_029184)
Rhizobium lentis BLR27T (NR_137243)
100 Trifolium alexandrinum, Egypt Lens
culinaris, Bangladesh
Lens culinaris, Bangladesh
Phaseolus vulgaris, Mexico
Lens culinaris, Bangladesh
CB31 KF938908 Rhizobium sullae IS 123T (NR_029330) 97.3 Hedysarum coronarium, Spain
CB32 KF938909 Ensifer terangae LMG 7834T (X68388) 99.7 Senegalia laeta, Senegal
CB36 KF938910 Ensifer kostiensis HAMBI 1489T (Z78203) 99.9 Senegalia senegal, Sudan
CB44 KF938911 Ensifer kostiensis HAMBI 1489T (Z78203) 99.9 Senegalia senegal, Sudan
CB46 KM044262 Rhizobium borbori DN316T (EF125187) 97 Activated sludge, China
CB56 KF938912 Ensifer kostiensis HAMBI 1489T (Z78203) 99.9 Senegalia senegal, Sudan
nodA gene
CB4 KF437384 Bradyrhizobium yuanmingense CCBAU 10071T
(AM117557)
97.1 Lespedeza cuneata, China
CB56 KJ018181 Ensifer fredii USDA 205T (EU292000)
Ensifer xinjiangensis CCBAU 110T (EF457965)
91.2 Glycine max, China
Glycine max, China
nifH gene
CB4 KJ018170 Bradyrhizobium yuanmingense CCBAU 10071T
(EU818927)
98.4 Lespedeza cuneata, China
CB56 KJ018169 Ensifer fredii USDA257 (CP003565)
Ensifer xinjiangensis CCBAU 110T (DQ411933)
Ensifer sojae CCBAU 05684T (GU994077)
95.1 Glycine max, China
Glycine max, China
Glycine max, China
Page 7
Sankhla et al.: CHARACTERIZATION OF MICROSYMBIONTS ASSOCIATED WITH CROTALARIA ROOT NODULES
379
in comparison to control and non-nodulated plants.
After recording the nodulation status 2-3 re-isolates
were successfully isolated and purified from
excavated root nodules of each host (C. burhia and
V. radiata) plant. The colony characteristics and
RAPD patterns (using RPOI primer) of re-isolates
were exactly similar to corresponding inoculated
parental strains (Fig. 2). The results of host
authentication, suggest that C. burhia is nodulated by
both, species of Ensifer and Bradyrhizobium. Sequencing of symbiotic genes
Symbiotic (nodA and nifH) genes of selective
nodulating isolates CB56 (Ensifer strain) and CB4
(Bradyrhizobium strain) were successfully amplified
and sequenced. The symbiotic genes could not be
amplified in the case of non-nodulating Rhizobium
(CB29, CB31 and CB46) and Ensifer (CB6, CB11,
CB12 and CB32) strains, although various primer sets
were used (Table 2).
Phylogenetic studies
16S rRNA gene phylogeny
In phylogenetic analyses, the Ensifer strains
formed three 16S rRNA types clustering into separate
clades and a single lineage (Fig. 3). The first clade
(16S rRNA type I) comprised by five nodulating
strains (CB5, CB17, CB36, CB44 and CB56) showing
100% sequence similarities with Ensifer sp. TF7,
TW10, RA9 and PC2 isolated from native legumes of
the Thar Desert as well as with Ensifer sp. K15
(LK936546) and Vr38 (LN851899) isolated from
chickpea and mung bean nodules, respectively, from
Pakistan. These five strains were very close to
E. kostiensis HAMBI 1489T (99.9% sequence
similarities) as well as to E. saheli LMG 7837T
(=ORS 609T) with 99.7% sequence similarities. The
strain CB11 alone formed a separate lineage (16S
rRNA type II) and showed 99.7% sequence similarity
with type strains E. kostiensis HAMBI 1489T and
E. saheli LMG 7837T. The second clade (16S rRNA
type III) comprised of 3 strains (CB6, CB12 and
CB32) showing 99.7% sequence similarities to
Ensifer sp. E60 (isolated from acacias in Algeria), and
type strain E. terangae LMG 7834T (Fig. 3 and Table 4).
The phylogenetic analyses of Bradyrhizobium strain
CB4 revealed that it had 100% sequence similarity to
strains PRNB-26, M11 and GX5 isolated from
Pongamia pinnata (India), V. radiata (Nepal) and
V. radiata (China), respectively and was adjacent to
type strain B. yuanmingense CCBAU 10071T
(99.8%). In addition, the strain CB4 also showed
99.9% sequence similarity with B. yuanmingense
TF17 isolated from root nodules of Tephrosia
falciformis from Thar Desert (Fig. 3). The three non-
nodulating Rhizobium strains (CB29, CB31 and
Fig. 2 — The RPOI-DNA fingerprinting gel images of nodulating
parental (P) Crotalaria burhia (CB)-Ensifer strains compared
with re-isolates from host Crotalaria burhia (R1) and Vigna
radiata (R2 and R3).
Fig. 3 — Phylogenetic tree constructed using 16S rRNA gene
sequences of Ensifer and Bradyrhizobium strains isolated from the
Indian native legume Crotalaria burhia together with those of
type strains and close relatives. [The tree was built using a
Maximum Likelihood (ML) method and bootstrap values
calculated for 1000 replications are indicated at internodes. The
scale bar indicates 2% substitutions per site. Accession numbers
from GenBank are in parenthesis. (Abbreviations: B, Bradyrhizobium;
CB, Crotalaria burhia; E, Ensifer; JNVU, Jai Narain Vyas
University and T indicates type strain)]
Page 8
INDIAN J EXP BIOL, JUNE 2018
380
CB46) formed three distinct types (Fig. 4). The strain
CB29 showed 100% similarity with following ty
pe strains: R. aegyptiacum 1010T, R. bangladeshense
BLR175T, R. binae BLR195
T, R. etli CFN 42
T and
R. lentis BLR27T. Strain CB31 showed 97.3%
similarity with R. sullae IS 123T and strain CB46 was
close to R. borbori DN316
T with 97% similarity.
Symbiotic gene phylogeny
The symbiotic genes of two strains CB56 and
CB4 each representing group of other similar
nodulating strains of Ensifer and Bradyrhizobium,
respectively, were successfully amplified and
sequenced to study their phylogeny. Our results
showed that the phylogeny of nodA gene of CB4
was congruent with its species phylogeny (close to
B. yuanmingense based on 16S rRNA gene). While
such congruence was not seen in the case of strain
CB56 (close to E. kostiensis based on 16S rRNA
gene). The nodA gene of CB56 showed 91.2%
sequence similarity with E. xinjiangensis CCBAU
110T and E. fredii USDA 205
T forming a distinct
lineage (Fig. 5 and Table 4). The nodA gene of
Bradyrhizobium strain CB4 clustered together with
NGB-SR15 isolated from soybean root nodules
from Egypt and showed 97.1% sequence similarity
to B. yuanmingense CCBAU 10071T.
The phylogeny of nifH gene was consistent with
that of nodA gene. Like nodA gene phylogeny, the
nifH gene of Ensifer sp. CB56 showed highest
sequence similarity (95.1%) with E. fredii USDA
257, E. sojae CCBAU 05684T and E. xinjiangensis
CCBAU 110T
(Fig. 6A and Table 4). Like 16S
rRNA and nodA phylogeny, the nifH gene of
Bradyrhizobium sp. CB4 was adjacent to B.
yuanmingense CCBAU 10071T and showed 98.4%
sequence similarity. The strain CB4 also showed
99.4% sequence similarity with strains such as
TF17, D1 and SR42 isolated form T. falciformis,
Glycine max and V. radiata, respectively
(Fig. 6B).
Fig. 4 — Phylogenetic tree constructed using 16S rRNA gene
sequences of non-nodulating Rhizobium strains isolated from
the Indian native legume Crotalaria burhia together with
those of type strains. [The tree was built using a Maximum
Likelihood (ML) method and bootstrap values calculated for
1000 replications are indicated at internodes. The scale bar
indicates 1% substitutions per site. Accession numbers from
GenBank are in parenthesis. (Abbreviations: CB, Crotalaria
burhia; JNVU, Jai Narain Vyas University; NR, NCBI
Reference sequence; R, Rhizobium and T indicates type
strain)]
Fig. 5 — Phylogenetic tree constructed using nodA gene
sequences of Ensifer and Bradyrhizobium strains isolated
from the Indian native legume Crotalaria burhia together
with those of type strains. [The tree was built using a
Maximum Likelihood (ML) method and bootstrap values
calculated for 1000 replications are indicated at internodes.
The scale bar indicates 10% substitutions per site. Accession
numbers from GenBank are in parenthesis. (Abbreviations:
B, Bradyrhizobium; CB, Crotalaria burhia; E, Ensifer;
JNVU, Jai Narain Vyas University and T indicates type
strain)]
Page 9
Sankhla et al.: CHARACTERIZATION OF MICROSYMBIONTS ASSOCIATED WITH CROTALARIA ROOT NODULES
381
Discussion
Like other native legumes, such as Acacia and
Prosopis, the C. burhia is also well adapted and
nodulates in the nutritionally poor alkaline soil of
Thar Desert of India. Analysis of soil from the various
sampling sites in present investigation supports our
previous report8 and further confirms that soil in Thar
Desert is alkaline throughout and poor in total N and
available P. Although the soil is not significantly
differing in nutritional characters at various sites but
the sampling sites in the different districts belongs to
semi-arid and hyper arid regions and varied in
average annual rainfall and mean temperature in
summer and winter. The texture is also different and
purely sandy in Barmer, Bikaner and Jaisalmer
districts as compared to other sites. The field survey
showed that C. burhia is widely distributed
throughout Thar Desert in rain-fed open area and on
marginal lands that indicates its well adaptation to
harsh climatic conditions of Thar Desert. The plants
of C. burhia were found nodulating at all sampling
sites although the number of nodules per plant varied
from site to site. The average number of nodules per
plant was poor at the sampling sites of arid and hyper-
arid regions (Barmer and Jaisalmer), where soil was
more sandy, poor in texture as compared to other
sampling sites (semi-arid areas). Our results are in
accordance with the previous studies35
that edapho-
climatic conditions such as rainfall, salinity, pH and
temperature may affect the number of effective root
nodule bacteria (RNB) in the soil that establish
functional nitrogen-fixing symbiosis and therefore
number of nodules per plant varied.
Molecular techniques such as ARDRA and RAPD
have been extensively used in rhizobial ecology for
genetic grouping and identification of strains. In the
present investigation, our results on the basis of DNA
fingerprinting indicates that fast growing Ensifer
strains are genetically more diverse than the slow
growing Bradyrhizobium strains. The more
occupancy of Ensifer strains compared to other
rhizobial (Bradyrhizobium and Rhizobium) strains in root
nodules of C. burhia indicates that Ensifer species are
well adapted to alkaline soils of Thar Desert and is
dominant microsymbiont of C. burhia and other native
legumes in the Thar Desert8,9,12,13,15,16,17,19
. There are
several reports8,9,12,13,15-19,
from different legumes
suggesting that alkaline soil and arid conditions
favours more incidences of Ensifer species in root
nodules rather than slow-growing species of
Bradyrhizobium. Genomic studies done by Tian
et al.36
also strengthen such assumptions and reported
that several gene clusters involved in osmoregulation
and adaptation to alkaline pH are present in the
species of Ensifer as compared to Bradyrhizobium.
The nine Ensifer strains sequenced in the present
study were classified into three types on the basis of
phylogenetic analysis of 16S rRNA gene, which were
mainly close to one of the three type strains
E. kostiensis HAMBI 1489T (isolated from root
nodules of Senegalia senegal, Sudan)37
, E. saheli
ORS609T (isolated from Sesbania cannabina,
Senegal)38
and E. terangae LMG 7834T (isolated from
Senegalia laeta, Senegal)38
; and also have close
similarity with Ensifer strains isolated from other
native legumes (Tephrosia spp., Rhynchosia aurea
Fig. 6 — Phylogenetic tree constructed using nifH gene sequences
of Ensifer (A); and Bradyrhizobium (B) strains isolated from the
Indian native legume Crotalaria burhia together with those of
type/reference strains. [The trees were built using a Maximum
Likelihood (ML) method and bootstrap values calculated for 1000
replications are indicated at internodes. The scale bar indicates
2% substitutions per site. Accession numbers from GenBank are
in parenthesis. (Abbreviations: B, Bradyrhizobium; CB,
Crotalaria burhia; E, Ensifer; JNVU, Jai Narain Vyas University
and T indicates type strain)]
Page 10
INDIAN J EXP BIOL, JUNE 2018
382
and Prosopis cineraria) of the Thar Desert8,13
as well
as with Ensifer strains from Pakistan, which indicate
their Asiatic origin. On the other hand, strain CB4
showed similarity with Bradyrhizobium sp. isolated
from Tephrosia species from the semi-arid regions of
western Rajasthan8. Strain CB4 also showed close
similarity with B. yuanmingense CCBAU 10071T
(isolated from the root nodules of Lespedeza cuneata
from China)39
but formed a separate lineage. In
addition, B. yuanmingense type of strains has also
been isolated in India from the root nodules of
soybean from different agricultural-ecological-
climatic regions of India40
. The first report of isolation
of B. yuanmingense is from wild legume L. cuneata
from China39
and the same is the case in our present
study, and the previous reports from our group where
such strains have been isolated from wild native
legumes (C. burhia, Tephrosia spp. and Vachellia
leucophloea)8,16
of Thar Desert where no soybean
cultivation history is found. This suggests that
B. yuanmingense is important unexplored microbial
resource of India that has to be further studied for its
wide host range. The occurrence of non-nodulating
Rhizobium strains in the present investigation is
interesting as these non-nodulating strains showed
close similarity with few nodulating Rhizobium
strains, such as Phaseolus vulgaris microsymbiont
(R. etli) from Mexico; R. aegyptiacum (isolated from
root nodules of Trifolium alexandrinum, Egypt) and
three type strains (R. bangladeshense, R. binae and
R. lentis) isolated from the root nodules of Lens
culinaris, Bangladesh. A single strain CB46 was close
to non-nodulating R. borbori isolated from activated
sludge, China.
The result of nodulation test suggests that C.
burhia is nodulated by both Ensifer and
Bradyrhizobium strains but not by species of
Rhizobium. Interestingly CB-Ensifer and CB-
Bradyrhizobium strains are nodulating crop legume V.
radiata. This is the first report on molecular
characterization of N fixing microsymbiont of C.
burhia, an important native legume and a good soil
binder of arid regions of the Thar Desert. The
isolation of non-nodulating Rhizobium from root
nodules of native legumes in our case is similar to
other researchers who isolated non-nodulating
Rhizobium strains from a wide range of plant taxa.
The symbiotic genes could not be amplified in non-
nodulating Rhizobium strains. There could be a
possibility that these Rhizobium strains were
originally nodulating, but may have lost the ability to
nodulate or the related nod genes during the
subculturing. Surprisingly, some strains of Ensifer
(CB6, CB11, CB12 and CB32) belonging to 16S
rRNA type II and III failed to nodulate their host
C. burhia. This lack of nodulation efficiency can be
explained by assumptions, such as (i) these strains of
Ensifer may have opportunistically entered along with
other nodulating rhizobia; and (ii) these strains might
have lost their symbiotic traits during symbiosis or
once purified onto artificial YEMA media41
.
Similar to other studies32,42
, the phylogeny based on
symbiotic genes (nodA and nifH) of Ensifer sp. CB56
was incongruent with its 16S rRNA gene phylogeny.
Strain CB56 (close to E. kostiensis HAMBI 1489T in
16S rRNA gene phylogeny) was closer to E. fredii
USDA 205T43
and E. xinjiangensis CCBAU 110T44
in
its symbiotic gene phylogeny. Such incongruence
suggests that the sym genes in it have been acquired
from other symbiotic Ensifer species/strains present in
the soil through horizontal gene transfer (HGT). In
contrary, Bradyrhizobium sp. CB4 was close to
B. yuanmingense CCBAU 10071T in both 16S rRNA
and symbiotic gene phylogenies.
Conclusion
Crotalaria burhia is nodulated by both, the fast
growing species of Ensifer that has close similarity
with the Old World Ensifer strains (E. kostiensis,
E. saheli and E. terangae) and the slow growing
species of Bradyrhizobium close to B. yuanmingense.
The Ensifer strains in the present study are genetically
diverse as compared to Bradyrhizobium strains and
both type of strains cross-nodulated crop V. radiata.
The genetic diversity and incongruence between
species (16S rRNA) and symbiotic gene phylogeny is
the result of HGT due to stressful conditions
prevailing in Thar Desert of India. Few strains in the
present study are phylogenetically divergent from the
existing type strains which indicate the presence of
potential novel species of rhizobia in the desert
region. Occurrence of non-nodulating Ensifer and
Rhizobium strains suggests that root nodules harbours
both symbiotic as well as opportunistic bacteria.
Furthermore polyphasic approach including multi
locus sequence analysis (MLSA) of conserved
protein-coding housekeeping genes, DNA-DNA
hybridization, BIOLOG, FAME analysis and whole
genome sequencing will be required for the
description of a formal new species of rhizobia
Page 11
Sankhla et al.: CHARACTERIZATION OF MICROSYMBIONTS ASSOCIATED WITH CROTALARIA ROOT NODULES
383
nodulating C. burhia in the Thar Desert of India.
Host-range of these effective N fixing strains needs to
be studied for using them as inoculums in future.
Such basic studies on identification,
characterization, screening of broad host range and
efficient native desert rhizobia will be useful in
preparing consortium of agriculturally important
rhizobial strains and will also form a platform for
more detailed genomic studies to understand the
mechanism of adaptability to changing environment
and spread of aridity.
Acknowledgement
Author Indu S. Sankhla acknowledges the
University Grants Commission (UGC), New Delhi,
for junior and senior research fellowships. The work
was also supported by grants from UGC-SAPII-CAS-
I, UGC-BSR Start-Up-Grant (F.30-16/2014) and the
Department of Biotechnology, Govt. of India
(BT/PR11461/AGR/21/270/2008).
References
1 Sprent J, Legume nodulation: A Global Perspective, (John
Wiley & Sons, Inc), 2009, 65.
2 Peix A, Ramírez-Bahena MH, Velázquez E & Bedmar EJ,
Bacterial associations with legumes. Crit Rev Plant Sci, 34
(2015) 17.
3 Lewis GP, Schrire B, Mackinder B & Lock M, Legumes of
the world, (Kew Royal Botanic Gardens, Kew) 2005.
4 Ansari AA, Crotalaria L. in India, (Bishen Singh Mahendra
Pal Singh, Dehra Dun), 2008, 52.
5 Ranjith M, Bajya DR & Manoharan T, Field study on
repellent efficacy of Crotalaria burhia Buch.-Ham. ex
Benth. and Anacardium occidentale L. against Odontotermes
obesus (Rambur). Indian J Nat Prod Resour, 6 (2015) 288.
6 Bhandari MM, Flora of the Indian Desert, (MPS Report,
Jodhpur), 1990, 103.
7 Gehlot HS, Tak N, Dagla HR & Davis TD, Indigenous and
modern scientific strategies for characterization, conservation
and sustainable utilization of bio-resources of the Indian Thar
Desert. J Arid Land Stud, 24 (2014) 5.
8 Gehlot HS, Panwar D, Tak N, Tak A, Sankhla IS, Poonar N,
Parihar R, Shekhawat NS, Kumar M, Tiwari R, Ardley J,
James EK & Sprent JI, Nodulation of legumes from the Thar
desert of India and molecular characterization of their
rhizobia. Plant Soil, 357 (2012) 227.
9 Gehlot HS, Tak N, Kaushik M, Mitra S, Chen WM, Poweleit N,
Panwar D, Poonar N, Parihar R, Tak A, Sankhla IS, Ojha A,
Rao SR, Simon MF, dos Reis Junior FB, Perigolo N,
Tripathi AK, Sprent JI, Young JPW, James EK &
Gyaneshwar P, An invasive Mimosa in India does not adopt
the symbionts of its native relatives. Ann Bot, 112 (2013) 179.
10 Tak N, Gehlot HS, Kaushik M, Choudhary S, Tiwari R,
Tian R, Hill Y, Bräu L, Goodwin L, Han J, Liolios K,
Huntemann M, Palaniappan K, Pati A, Mavromatis K,
Ivanova N, Markowitz V, Woyke T, Kyrpides N & Reeve W,
Genome sequence of Ensifer sp. TW10; a Tephrosia
wallichii (Biyani) microsymbiont native to the Indian Thar
Desert. Stand Genomic Sci, 9 (2013) 304.
11 Panwar D, Tak N & Gehlot HS, Nodulated Native Legumes
in an Arid Environment of Indian Thar Desert. In: MH
Fulekar & RK Kale (eds.) Recent Trends in Plant Sciences
(IK International Publishing House Pvt. Ltd. New Delhi, India), 2014, 284.
12 Sankhla IS, Meghwal RR, Tak N, Tak A & Gehlot HS,
Phenotypic and molecular characterization of
microsymbionts associated with Crotalaria medicagenia: a
native legume of the Indian Thar Desert. Plant Archives, 15 (2015) 1003.
13 Tak N, Awasthi E, Bissa G, Meghwal RR, James EK,
Sprent JS & Gehlot HS, Multi locus sequence analysis and
symbiotic characterization of novel Ensifer strains nodulating
Tephrosia spp. in the Indian Thar Desert. Syst Appl Microbio, 39 (2016) 534.
14 Gehlot HS, Ardley J, Tak N, Tian R, Poonar N,
Meghwal RR, Rathi S, Tiwari R, Adnawani W, Seshadri R &
Reddy TB, High-quality permanent draft genome sequence
of Ensifer sp. PC2, isolated from a nitrogen-fixing root
nodule of the legume tree (Khejri) native to the Thar Desert of India. Stand Genomic Sci, 11 (2016) 43.
15 Sankhla IS, Tak N, Meghwal RR, Choudhary S, Tak A,
Rathi S, Sprent JI, James EK & Gehlot HS, Molecular
characterization of nitrogen fixing microsymbionts from root
nodules of Vachellia (Acacia) jacquemontii, a native legume from the Thar Desert of India. Plant Soil, 410 (2017) 21.
16 Choudhary S, Meghwal RR, Sankhla IS, Tak N & Gehlot HS,
Molecular characterization and phylogeny of novel diverse
nitrogen fixing microsymbionts associated with Vachellia
(Acacia) leucophloea in arid and semiarid regions of Rajasthan. Indian Forester, 143 (2017) 266.
17 Rathi S, Gaur S, Tak N, Tak A & Gehlot HS, Genetically
diverse root nodule bacteria associated with Alysicarpus
vaginalis from alkaline soil of Rajasthan, India. Plant
Archives, 17 (2017) 495.
18 Le Quéré A, Tak N, Gehlot HS, Lavire C, Meyer T, Chapulliot D,
Rathi S, Sakrouhi I, Rocha G, Rohmer M & Severac D,
Genomic characterization of Ensifer aridi, a proposed new
species of nitrogen-fixing rhizobium recovered from Asian,
African and American deserts. BMC Genomics, 18 (2017) 85.
19 Choudhary S, Tak N & Gehlot HS, Phylogeny and genetic
diversity assessment of Ensifer strains nodulating Senegalia
(Acacia) senegal (L.) Britton. in arid regions of Western Rajasthan, India. Microbiology, 87 (2018) 127.
20 Samba RT, de Lajudie P, Gillis M, Neyra M, Spencer-Baretto
MM & Dreyfus B, Diversity of rhizobia nodulating Crotalaria
spp. from Senegal. Symbiosis, 27 (1999) 259.
21 Jourand P, Giraud E, Bena G, Sy A, Willems A, Gillis M,
Dreyfus B & de Lajudie P, Methylobacterium nodulans sp.
nov., for a group of aerobic, facultatively methylotrophic,
legume root-nodule forming and nitrogen-fixing bacteria. Int
J Syst Evol Microbiol, 54 (2004) 2269.
22 Liu XY, Wang ET, Li Y & Chen WX, Diverse bacteria
isolated from root nodules of Trifolium, Crotalaria and
Mimosa grown in the subtropical regions of China. Arch Microbiol, 188 (2007) 1.
23 Rocha AL, Isolation and characterization of bacterial symbionts
from Crotalaria spectabilis grown on trichloroethene
Page 12
INDIAN J EXP BIOL, JUNE 2018
384
contaminated soil, Dissertation, Missouri University of Science
and Technology, Missouri, United States, 2011.
24 Singha B, Mazumder PB & Pandey P, Characterization of
plant growth promoting rhizobia from root nodule of
Crotalaria pallida grown in Assam. Indian J Biotechnol, 15
(2016) 210.
25 Vincent JM, A manual for the practical study of root-nodule
bacteria, (Blackwell Scientific Publications, Oxford), 1970.
26 Somasegaran P & Hoben HJ, The handbook for Rhizobia:
methods in legume Rhizobia technology, (Springer Verlag,
New York), 1994.
27 Cheng HR & Jiang N, Extremely rapid extraction of DNA
from bacteria and yeasts. Biotechnol lett, 28 (2006) 55.
28 Weisburg WG, Barns SM, Pelletier DA & Lane DJ, 16S
ribosomal DNA amplification for phylogenetic study. J
Bacteriol, 173 (1991) 697.
29 Richardson AE, Viccars LA, Watson JM & Gibson AH,
Differentiation of Rhizobium strains using the polymerase
chain reaction with random and directed primers. Soil Biol
Biochem, 27 (1995) 515.
30 Yanagi M & Yamasato K, Phylogenetic analysis of the
family Rhizobiaceae and related bacteria by sequencing of
16S rRNA gene using PCR and DNA sequencer. FEMS
Microbiol Lett, 107 (1993) 115.
31 Haukka K, Lindstrom K & Young JPW, Three phylogenetic
groups and nodA and nifH genes in Sinorhizobium and
Mesorhizobium isolates from leguminous trees growing in
Africa and Latin America. Appl Environ Microbiol, 64
(1998) 419.
32 Laguerre G, Nour SM, Macheret V, Sanjuan J, Drouin P &
Amarger N, Classification of rhizobia based on nodC and
nifH gene analysis reveals a close phylogenetic relationship
among Phaseolus vulgaris symbionts. Microbiology, 147
(2001) 981.
33 Thompson JD, Higgins DG & Gibson TJ, CLUSTAL W:
improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, position-specific gap
penalties and weight matrix choice. Nucleic Acids Res, 22
(1994) 4673.
34 Tamura K, Stecher G, Peterson D, Filipski A & Kumar S,
MEGA6: Molecular Evolutionary Genetics Analysis Version
6.0. Mol Biol Evol, 30 (2013) 2725.
35 Zahran HH, Rhizobium–legume symbiosis and nitrogen
fixation under severe conditions and in an arid climate.
Microbiol Mol Biol Rev, 63 (1999) 968.
36 Tian CF, Zhou YJ, Zhang YM, Li QQ, Zhang YZ, Li DF,
Wang S, Wang J, Gilbert LB, Li YR & Chen WX,
Comparative genomics of rhizobia nodulating soybean
suggests extensive recruitment of lineage-specific genes in
adaptations. Proc Natl Acad Sci, 109 (2012) 8629.
37 Nick G, de Lajudie P, Eardly B, Soumalainen S, Paulin L,
Zhang X, Gillis M & Lindstrom K, Sinorhizobium arboris
sp. nov., and Sinorhizobium kostiense sp. nov., isolated from
leguminous trees in Sudan and Kenya. Int J Syst Bacteriol,
49 (1999) 1359.
38 de Lajudie P, Willems A, Pot B, Dewettinck D, Maestrojuan G,
Neyra M, Collins MD, Dreyfus B, Kersters K & Gillis M,
Polyphasic taxonomy of Rhizobia: Emendation of the genus
Sinorhizobium and description of Sinorhizobium meliloti
comb. nov., Sinorhizobium saheli sp. nov., and
Sinorhizobium teranga sp. nov. Int J Syst Bacteriol, 44
(1994) 715.
39 Yao ZY, Kan FL, Wang ET, Wei GH & Chen WX,
Characterization of rhizobia that nodulate legume species of the
genus Lespedeza and description of Bradyrhizobium
yuanmingense sp. nov. Int J Syst Evol Microbiol, 52
(2002) 2219.
40 Appunu C, N’Zoue A & Laguerre G, Genetic diversity of
native bradyrhizobia isolated from soybeans (Glycine max L.) in
different agricultural-ecological-climatic regions of India.
Appl Environ Microbiol, 74 (2008) 5991.
41 Brom S, Garcia-de los Santos A, Stepkowsky T, Flores M,
Davila G, Romero D & Palacios R, Different plasmids of
Rhizobium leguminosarum bv. phaseoli are required for
optimal symbiotic performance. J Bacteriol, 174 (1992)
5183.
42 Vinuesa P, Silva C, Werner D & Martinez-Romero E, Population
genetics and phylogenetic inference in bacterial molecular
systematics: the roles of migration and recombination
in Bradyrhizobium species cohesion and delineation. Mol
Phylogenet Evol, 34 (2005) 29.
43 Chen WX, Yan GH & Li JL, Numerical taxonomic study of
fast-growing soybean rhizobia and a proposal that Rhizobium
fredii be assigned to Sinorhizobium gen. nov. Int J Syst
Bacteriol, 38 (1988) 392.
44 Peng GX, Tan ZY, Wang ET, Reinhold-Hurek B, Chen WF
& Chen WX, Identification of isolates from soybean nodules
in Xinjiang Region as Sinorhizobium xinjiangense and
genetic differentiation of S. xinjiangense from Sinorhizobium
fredii. Int J Syst Evol Microbiol, 52 (2002) 457.