Gene, 46 (1986) 71-78 Elsevier 71 GENE 1715 (Recombinant DNA; homoser&-U-acetyltransferase; methionine biosynthesis; S-adenosyhnethionine; mRNA; yeast gene bank; transcriptional regulation) Ma~i~o Baromi, Susan Livian, Elnzo Martegani ami Liiia Alberghina * Dipartimento di Fisiologia e Biochimica Generali, Sezionedi Biochimica Comparata, Universitd diiUilano, Via Celoria 2620133 Milan (Italyl Tel. (02)2364218 (Received January 4th. 1986) (Revision received June 4th, 1986) (Accepted June 13th, 1986) SUMMARY The ME;TI;!gene of Saccharomyces cerevisiue, which codes for homoserine-0-acetyhransferase, a key enzyme in methionine biosynthesis, was isolated by complementation of a met2 mutant strain of S. cerevistie with a yeast gene bank. A 3.9-kb genomic fragment contains the entire gene, as demonstrated by genetic and molecular analysis of the integrative transformants. A ~lyad~ylat~ mRNA of 1700 nt is detected by Northern blot hybridization with a ME22 probe. The level of this mRNA decreases by addition of exogenous methionine or of S-adenosylmethionine, suggesting a transcriptional regulation, The level of specific mRNA and the enzyme activity found in transformants that bear the ME22 gene on a multicopy plasmid suggest that also a ~st-~~sc~ption~ regulatory mechanism may be operative in budding yeast. Studies done on the biosynthetic pathway for methionine in S. cerz?v&& have led to the ident& cation of a group of at least five enzymes whose * To whom correspondence and reprint request should be addressed. Abbreviations: aa, ammo acid(s); Ap, ampicillin; bp, base pair(s); EtdBr, ethidium bromide; HSTase, homoserine-U-ace- tyltransferase (EC 2.3.1.31); kb, 1000 bp; nt, nucIeotide(s); R, resistance; ‘, sensitivity; SAM, ~-adenosy~e~on~e; SDS, sodium dodecyl sulfate; YNB, yeast nitrogen base; Tc, tetra- cycline; wt, wiid type; [ 1, designates plasmid-carrier state. synthesis appears to be coordinated and controlled by the same regulatory system (Cherest et al., 1971), although the structural genes encoding for these enzymes are located on different chromosomes (Schiid and Mortimer, 1985). Biochemical and genetic studies on wt and mutant strains, in dii%rent growth conditions, have shown that the synthesis of these enzymes is repressed in presence of methionine and that the activated form of this aa, SAM, is also involved in the regulation (Cherest et al., 1973a,b). Recently it has been shown that the expression of two methionine biosynthetic genes, belonging to the above discussed group, MET3 (Cherest et al., 1985) 0378-I 1!9/86/$03.50 8 1986 Ekwier Science Publishers B.V. (Biomedical Division)
Ma~i~o Baromi, Susan Livian, Elnzo Martegani ami Liiia Alberghina *
Dipartimento di Fisiologia e Biochimica Generali, Sezione di Biochimica Comparata, Universitd diiUilano, Via Celoria 2620133 Milan (Italyl Tel. (02)2364218
(Received January 4th. 1986) (Revision received June 4th, 1986) (Accepted June 13th, 1986)
SUMMARY
The ME;TI;! gene of Saccharomyces cerevisiue, which codes for homoserine-0-acetyhransferase, a key enzyme in methionine biosynthesis, was isolated by complementation of a met2 mutant strain of S. cerevistie with a yeast gene bank. A 3.9-kb genomic fragment contains the entire gene, as demonstrated by genetic and molecular analysis of the integrative transformants. A ~lyad~ylat~ mRNA of 1700 nt is detected by Northern blot hybridization with a ME22 probe. The level of this mRNA decreases by addition of exogenous methionine or of S-adenosylmethionine, suggesting a transcriptional regulation, The level of specific mRNA and the enzyme activity found in transformants that bear the ME22 gene on a multicopy plasmid suggest that also a ~st-~~sc~ption~ regulatory mechanism may be operative in budding yeast.
Studies done on the biosynthetic pathway for methionine in S. cerz?v&& have led to the ident& cation of a group of at least five enzymes whose
* To whom correspondence and reprint request should be addressed.
synthesis appears to be coordinated and controlled by the same regulatory system (Cherest et al., 1971), although the structural genes encoding for these enzymes are located on different chromosomes (Schiid and Mortimer, 1985).
Biochemical and genetic studies on wt and mutant strains, in dii%rent growth conditions, have shown that the synthesis of these enzymes is repressed in presence of methionine and that the activated form of this aa, SAM, is also involved in the regulation (Cherest et al., 1973a,b).
Recently it has been shown that the expression of two methionine biosynthetic genes, belonging to the above discussed group, MET3 (Cherest et al., 1985)
and MET25 (Sangsoda et al., 1985) is subjected to a transcriptional control by exogenous methionine.
To study the mechanism of simultaneous control on the expression of these unlinked genes at a molecular level, we have isolated the S. cerevisiue ME72 gene, encoding for HSTase, which catalyzes the first step in methionine biosynthesis and seems to be an important control point of the pathway (de Robichon-Szuhnajster and Cherest, 1967).
Previous studies done on the level of HSTase activity in wt strain, measured either after growth in the presence of methionine or after addition of methionine or SAM to the crude extract, have led to the conclusion that synthesis of the enzyme is repressed by methionine, whereas its activity is inhi- bited by SAM (de Robichon-Szulmajster and Cherest, 1967).
In this paper we describe the molecular cloning, the physical characterization and a preliminary study of the expression of ME27 gene of S. cerevisiae. Our data strongly suggest that both a transcriptional and a post-transcriptional regulatory mechanism exists for HSTase synthesis in budding yeast.
MATERIALS AND METHODS
(a) strplias and media
The following haploid strains of S. cerevisiae were used: S288C (MATa SUC2, ga12, CUPl) obtained from Dr. S. Sora (Dept. of Genetics, University of Milan), X-4003-5B (MATa, udel, his& hp5, met2, u1u3,leu2, gall) obtained from Yeast Genetics Stock Center (Berkeley, CA, U.S.A.), X-4004-3A (MATa, lys5, uru3, mef2, trpl) obtained from Prof. E. Galli (Dept. of Genetics, University of Milan), GRF18 (MATa, leu2-3,2-112, hi.r3-11,3-15) obtained from Dr. P. Philippsen (Biozentrum, Basel).
E. coli HBlOl was used as host for plasmid maintenance, for gene bank and recombinant plas- mid construction and recovery of plasmid from yeast. For E. coli, LB medium was used (Maniatis et al., 1982). When necessary 50 pg Ap/ml or 15 pg Tc/ml were added.
Yeast cells were grown in Yeast Nitrogen Base (YNB) without aa (Difco, Detroit, U.S.A.), con- taining 2% glucose and 50 mg/l of the appropriate
supplements. Cells were grown in batch cultures in a shaking water bath at 30°C. Growth was monitored as increase in the cell number determined with a Coulter Counter ZBI (1OOqm orifice) (Pop010 et al., 1982).
Repressive conditions were obtained by adding to minimal medium 0.3 mM L-methionine or 0.15 mM SAM (Boehringer, Mannheim, F.R.G.). Presporu- lation and sporulation media were prepared as described by Simchen et al. (1972).
(b) Yeaat eeli-free extract and emyme assay
Yeast cell-free extracts were made as described by Cherest et al. (197 1) with some minor mod%cations. Cells from a loo-ml culture in exponential growth (about 8 x lo6 cells/ml) were collected and washed in 0.1 M potassium phosphate buffer (pH 7.5). They were then suspended in 0.5 ml of the same but&r and 1 g of glass beads (0.45 mm diameter) was ad&d. Cells were broken by vortexing for 1 min and then cooling in ice for 1 min, repeating this operation four times. Beads and cellular debris were removed by centrifugation and the resulting crude extract was used for enzymatic activity assay.
HSTase activity was assayed as described by Nagai and Kerr (1971) using the exchange reaction and [ “C]homoseaine (Amersham, U.K.).
O-Acetyl-homoserine was synthesized in our laboratory according to Nagai and Flavin (1971).
Protein concentration was measured with the mi- crobiuret method (Zamenhof, 1975), using bovine serum albumin (Sigma, St. Louis, MO, U.S.A.) as reference standard.
(c) PlasmidsandgenebtnL
E. coli plasmid pBR322 was provided by Prof. V. Sgaramella (University of Pa* Italy). The yeast shuttle plasmid pDB248’, which contains the LEU2 gene of S. cerevisiue and the origin of replication of 2~ plasmid (Beach et al., 1982), obtained from Dr. P. Nurse (Imperial Cancer Research Fund, London), was used to construct the gene bank. The yeast genomic bank was constructed accor&g to Beach et al. (1982). Total yeast DNA, extracted from strain S288C, was partiaBy d@&d with HindIII, then ligated with pDB24g’ l&tar&d with Hind111 and dephosphorylated with calf intestine
73
alkaline phosphatase. The ligation mix was used to transform E. coli HBlOl to Ap resistance obtaining
about 20 000 independent clones. About 50 y0 of the ApR clones was Tcs. Plasmid pURA-1 was obtained by selecting for Ura+,Leu+ transformants, atter transformation of strain X-4003 and selection in minimal plates lacking both leucine and uracil. In the Hind111 site of pDB248’ this plasmid bears a 1. 1-kb
fragment that contains the yeast URA3 gene (Bach et al., 1979).
(d) Isolation and manipulation of DNA
Preparation of plasmid DNA from E. coli cultures was performed as described by Maniatis et al. (1982). For rapid analysis the alkaline extraction procedure of Bimboim and Doly (1979) was used. Total yeast DNA was prepared according to Cryer et al. (1975). Plasmid transfer from S. cerevisiae to E. coli was performed as described by Crabeel et al. (1981). Restriction enzyme digestions, agarose gel electrophoresis, hybridization analysis, nick-trans- lation and S 1 mapping were performed according to standard procedures (Maniatis et al., 1982).
(e) Transformation
S. cerevkiae was transformed according to the method of Hinnen et al. (1978). Alternatively the alkaline LiCl method of Ito et al. (1983) was used. E. coli was transformed according to the procedure described by Maniatis et al. (1982).
(f) Preparation of yeast RNA
Yeast RNA was isolated according to Federoff et al. (1982). Poly(A)+ -enriched RNA was isolated by loading total yeast RNA on oligo(dT)-cellulose (Pharmacia, Uppsala, Sweden) according to Mania- tis et al. (1982).
RESULTS AND DISCUSSION
(a) Jiolation of a genomic sequence that comple- ments the met2 mutation
To isolate the plasmids containing the ME72 gene we transformed the yeast strain X-4003-5B with a
genomic library constructed in the pDB248’ vector and screened for transformed clones on minimal plates lacking both leucine and methionine. From about 40000 Leu + clones we obtained about 100 Leu’, Met + clones. Several of these clones were tested for stability of the transformed pheno- type. All the tested clones showed a simultanous loss of Leu’ and Met+ phenotypes after several generations in non-selective media.
From the same clones we recovered the plasmids by DNA extraction and transformation of E. coli
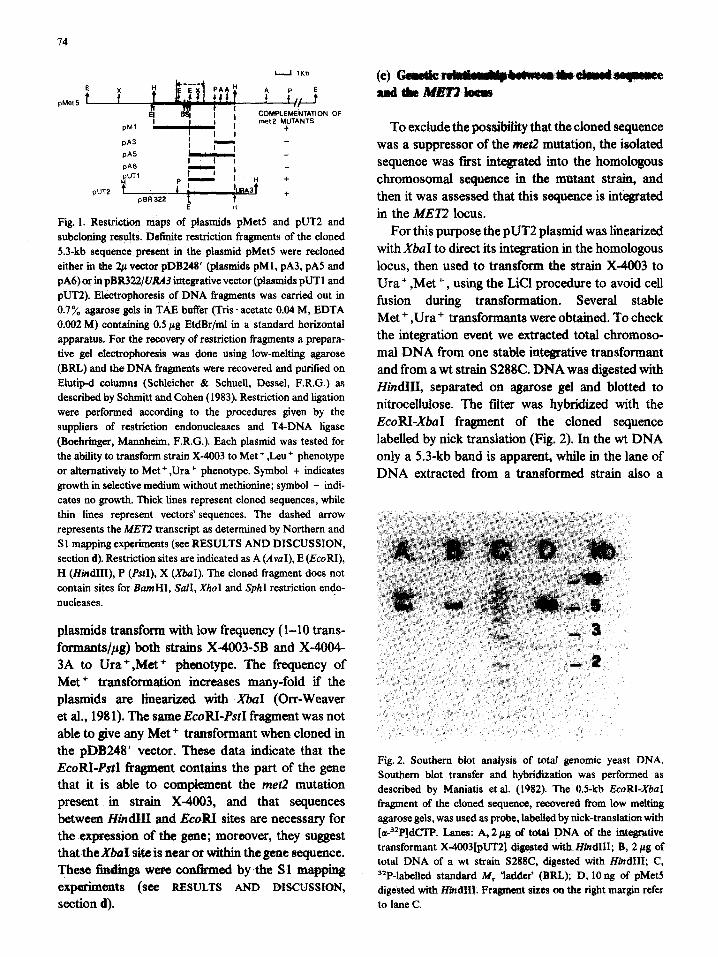
HBlOl to Ap resistance. All the clones contained a 17.7-kb plasmid that bears a 5.3-kb insert in the Hind111 site of pDB248’. One plasmid, called pMet-5 was further purified by CsCl-EtdBr gradients and characterized. pMet-5 maintains both ApR and TcR markers, indicating that the insertion of yeast DNA fragment into the Hind111 site, which lies in the promoter of the TcR gene, has not inacti- vated the expression of the TcR gene (Chevallier et al., 1980). This plasmid transforms with a high efficiency (more than 20 000 transformants/~g) strain X-4003 to Leu’ or Met + phenotypes, more- over, all the Leu+ transformants were also Met + and vice versa. The restriction map of the plasmid pMet-5 is shown in Fig. 1.
(b) Subcloning of the putative MET2 gene
The 5.3-kb Hind111 fragment present on pMet5 was subcloned both in the same pDB248’ vector and in integrative vectors.
Several digestion fragments were recovered by low-melting agarose gels and recloned in pDB248’ and the resulting plasmids were tested for their ability to transform strain X-4003 to Met + ,Leu+ pheno- type. Results are schematized in Fig. 1. The only fragment that gives a good complementation with the met2 mutation is the 3.9-kb HindIII-PsrI fragment which must contain the entire putative MET2 gene. Because the 1.6-kb HindHI-EcoRI fragment lacks restriction sites for all the enzymes we have tested, we aimed to localize the gene position via integrative transformation. For this purpose we constructed integrative vectors by cloning first the EcoRI-PstI and EcoRI-Hind111 fragments in pBR322, obtaining the plasmid pLl0 and pLl1 and then inserting in these plasmids the yeast URA3 gene in the Hind111 site obtaining plasmids pUT1 and pUT2. These two
74
PA3 I PA5
, ,I PA6
I I I
pUT1 I H P- I H +
PUT2 t ( I
pm322 t bA3 +
E H
Fig. 1. Restriction maps of plasmids pMet5 and pUT2 and subcloning results. Definite restriction fragments of the cloned 5.3-kb sequence present in the plasmid pMet5 were recloned either in the 2~ vector pDB248’ (plasmids pM1, pA3, pA5 and pA6) or in pBR322/VRA3 integrative vector (plasmids pUT1 and pUT2). Electrophoresis of DNA fragments was carried out in 0.7% agarose gels in TAE buffer (Tris . acetate 0.04 M, EDTA 0.002 M) containing 0.5 pg EtdBr/ml in a standard horizontal apparatus. For the recovery of restriction fragments a prepara- tive gel electrophoresis was done using low-melting agarose (BRL) and the DNA fragments were recovered and purified on Elutip-d columns (Schleicher & Schuell, Dessel, F.R.G.) as described by Schmitt and Cohen (1983). Restriction and ligation were performed according to the procedures given by the suppliers of restriction endonucleases and T4-DNA ligase (Boehringer, Mannheim, F.R.G.). Each plasmid was tested for the ability to transform strain X-4003 to Met + ,Leu + phenotype or alternatively to Met + ,Ura + phenotype. Symbol + indicates growth in selective medium without methionine; symbol - indi- cates no growth. Thick lines represent cloned sequences, while thin lines represent vectors’ sequences. The dashed arrow represents the ME72 transcript as determined by Northern and Sl mapping experiments (see RESULTS AND DISCUSSION, section d). Restriction sites are indicated as A (AvuI), E (EcoRI), H (HindIII), P (PsrI), X (XbaI). The cloned fragment does not contain sites for EarnHI, WI, XhoI and SphI restriction endo- nucleases.
plasmids transform with low frequency (l-10 trans- formants/& both strains X-4003-5B and X-4004- 3A to Ura+,Met+ phenotype. The frequency of Met + transformation increases many-fold if the plasmids are linearized with XbaI (Orr-Weaver et al., 1981). The same EcoBI-PstI fragment was not able to give any Met + transformant when cloned in the pDB248’ vector. These data indicate that the EcoBI-Psf fragment contains the part of the gene that it is able to complement the met2 mutation present in strain X-4003, and that sequences between HindI and EcoBI sites are necessary for the expression of the gene; moreover, they suggest that the Xbuf site is near or within the gene sequence. These tindings were contIrmed by the S.1 mapping eXp&?I’imatS (See RESULTS AND DISCUSSIGN,
section d).
To exclude the possibility that the cloned sequence was a suppressor of the met2 mutation, the isolated sequence was fnst integrated into the homologous chromosomal sequence in the mutant strain, and then it was assessed that this sequence is integrated in the MEi’2 locus.
For this purpose the pUT2 plasmid was linearized with XbuI to direct its integration in the homologous locus, then used to transform the strain X-4003 to Ura+ ,Met + , using the LiCl procedure to avoid cell fusion during transformation. Several stable Met + ,Ura + transformants were obtained. To check the integration event we extracted total chromoso- mal DNA from one stable integrative transformant and from a wt strain S288C. DNA was digested with HindIII, separated on agarose gel and blotted to nitrocellulose. The futer was hybridized with the EcoBI-XbuI fragment of the doned sequence labelled by nick translation (Fig. 2). In the wt DNA only a 5.3-kb band is apparent, while in the lane of DNA extracted from a transformed strain also a
Fig. 2. Southern blot analysis of total genomic yeast DNA. Southern blot transfer and hybridization was performed as described by Maniatis et al. (1982). The 0.5-kb EcoRI-XbaI fragment of the cloned sequence, recovered from low melting agarose gels, was used as probe, labelled by nick-translation with [a-32P]dCTF. Lanes: A, 2 pg of total DNA of the integrative transformant X_4003[pUT2] digested with HixdIII; B, 2 c(g of total DNA of a wt strain S288C, digested with HbrdIII; C, 32P-labelled standard M, ‘ladder’ (BRL); D, 10 ng of pMet5 digested with HindlII. Fragment sizes on the right margin refer to lane C.
75
7.3-kb band appears, as expected if the pUT2 sequences are integrated into the chromosome.
Some of the stable Met+ ,Ura + transformants
were crossed with strain GRF18 (rtra + met + ). Diploids were selected and then induced to sporu- late. A random spore analysis was performed to follow the resegregation of Met + and Ura + pheno- types. All the analysed spores (105) were Met + , while 22.8% were Ura-.
The fact that Met - spores have not been recover- ed, taken together with the subcloning results, indicates that the cloned sequence is integrated at ME72 locus. Moreover the site of integration is totally unlinked to the URA3 chromosomal locus.
(d) Northern blot experiments
The iUEZ? mRNA was identified both on total RNA and on poly(A) + -enriched RNA. Only a faint band of hybridization, that migrates just near the 17s rRNA species, can be detected on RNA blots per- formed with total RNA. Poly(A)+-enriched RNA extracted both from wt strain S288C and from X-4003 strain transformed with pMet5 was se- parated on denaturing gels, blotted to nitrocellulose and then hybridized with single-strand probes pre- pared by cloning the EcoRI-PstI fragment in the pSP64 and pSP65 vectors (Melton et al., 1984). A single mRNA species of about 1700 nt was detected with the pSP65 probe (Fig. 3), indicating that the direction of transcription was P&I +X&I + EcoRI as specified in Fig. 1. A very strong signal was detected for the transformant poly(A)+ RNA, indi- cating a many-fold increase of the MET2 mRNA level. The amount of ME22 mRNA molecules per cell in wt strain, under non-repressed conditions, was measured in comparison to URA3 mRNA, utilizing the slot-blot technique, and using probes prepared by nick transiation with the same specific activity (5 x lo7 cpm/pg), and was found to be slightly less than the U&f3 mRNA level, being in the order of 2-3 copies/cell (Bach et al., 1979).
Sl mapping experiments were performed on HindHI-%a1 and onXbaI-PstI fragments, annealed with excess of poly(A) + RNA. They indicate that the transcription starts about 400 bp on the right of the XbaI site and extends for about 1300 bp on the left of the sameXba1 site (results not shown) (see Fig. 1).
kb
3.4-
I.&
Fig. 3. Northern blot analysis of MET2 transcript. 2Opg of poly(A)+-enriched RNA, extracted from wt strain S288C (lanes A and C) or 10 ng extracted from a transformed strain X-4003[pMet5] (lanes B and D) exponentially growing in minimal medium, were hybridized to complementary single- strand probes. Poly(A) + -enriched RNA was denatured and sub- mitted to electrophoresis in agarose-formaldehyde gels (Maniatis et al., 1982). After electrophoresis, RNA was blotted to nitro- cellulose and hybridized. The probes were prepared by cloning in the two different orientations the 1.6-kb EcoRI-PstI fragment
of the MER sequence in the pSP65 (lanes A, B) and pSP64 (lanes C, D) vectors and by SP6 polymerase (Amersham, U.K.) transcription in presence of [LY-~*P]UTP (spec. act. 800 Ci/mmol) as described by the suppliers. The specific activity of RNA probes was about 2 x lo9 cpm/pg. The conditions ofthe hybridi- zation of RNA probes were as described by Melton et al. (1984).
A specific signal is obtained only with the pSP65 (MET2) probe. Ribosomal RNA of yeast and plamid pLl1 digested with PstI + EcoRI were used as M, standards (see left margin).
(e) Regulation of levels of MET2 mRNA
Using the cloned ME72 gene as a probe we measured the levels of ME22 mRNA both in wt and in a transformed strain during exponential growth in repressing and non-repressing conditions. Messen- ger RNA levels were quantitated with the slot-blot technique and the results are shown in Fig. 4 and in Table I. The HSTase activity present in cell-free ex- tracts was measured under the same conditions. In the wt strain we observed a reduction of both mRNA
Fig. 4. Slot-blot hybridization. Equal amount of RNA (15 pg), extracted both from a wt strain S288C (lanes A,B,C) and from a transformant strain, X-4003[pMet5] (lanes D,E,F), were loaded on nitrocellulose filters using a slot minifold apparatus (Schleicher & Schuell, Dessel, F.R.G.) as described by Oesch
et al. (1985). Filters were hybridized with a single stranded RNA probe obtained by transcription with SP6 polymerase of pSP65(MET2). The condition of hybridization and washings were as described by Melton et al. (1984). Lanes A and D, minimal medium; B and E, minimal medium + 0.15 mM SAM; C and F, minimal medium + 0.3 mM methionine.
levels and of enzyme activity in presence of 0.3 mM methionine, in agreement with a transcriptional regu- lation of the ME72 gene. A reduced mRNAleve1 was
also observed after addition of SAM, suggesting that this molecule can exert some regulatory function on ME72 gene expression. In a X4003[pMet5] strain we observed a ten-fold increase of the level of ME72
miWA, in agreement with a gene dosage effect corresponding to about ten plasmids per cell (see Table I, footnote d).
Also in the transformed strain we found a substan-
tial reduction of mRNAlev& in the presence of both methionine and SAM, indicating that in the MET2 gene cloned on the plasmid there sequences required for its repression are also present. An interesting difference with the findings observed with the other cloned genes of the methionine biosynthesis, like MET3 and ME725, was that the enzyme activity in transformed cells was only slightly higher than in wt cells. It does not increase in a parallel fashion with the ten-fold increment of the mRNA level, suggesting that a post-transcriptional modulation of the ME72
gene expression may also be operative.
(f) Conclusions
(1) The MET2 gene of S. cerevWe has been isolated from a yeast gene bank by fknctional com- plementation of the met2 mutation. A single specitic transcript of about 1700 nt was identified on Northern blots.
(2) Growth in the presence of methionine causes an 85% reduction of HSTase activity and a 70% reduction of the level of ME72 mRNA. These data strongly suggest that methionine represses the ME22 gene transcription, as reported for the MET3 and MET25 genes (Cherest et al., 1985; Sangsoda et al., 1985), and as suggested previously by Surdin-Kerjan
TABLE I
Expression of the cloned METZ gene in yeast
Strain Additions to Levels of minimal mediuma specific mRNA b
HSTase (specific activity”)
S288C None 1.0 0.98 L-Methionine 0.3 0.16 SAM 0.5 0.30
X4003[pMet5] d None L-Methionine SAM
9.0 1.95 3.4 ND” 3.4 0.76
* For, repression studies 0.3 mM L-methionine or 0.15 mM SAM were added to minimal medium. b Levels of specific mRNAs were measured either by a densitometric analysis of autoradiograms or by cutting and counting by liquid scintillation the nitrooelhdose lilters a&r a slot-blot hybridization performed as described in the legend of Fig. 4. Each vale is the mean of at least two ditkrent assays. Values are relative to those of strain S288C grown in minimal me&m and defmed as 1.00. c Expressed as nmol of acetyl-homoserine produced per min/mg of extracted protein. d The plasmid copy number per cell was about ten. Equal amounts of total DNA, extracted from a wt S288C strain and transformed strain X4003tpMet5], were completely digested with PstI, fractionated on agarose gel and blotted on nitrocellutose. The filter was then hybridized with a ME22 labelkd probe and autoradiographed. The plasmid copy number was determined by a densitometric analysis of the autoradiogram, taking the single-copy chromosomal MET2 gene as an internal standard. ’ ND. not determined.
77
and de Robichon-Szuhnajster (1975). Interestingly,
we have found also 50 % reduction of MER mRNA
levels in the presence of SAM. Previous works of de Robichon-Szuhnajster and Cherest (1967) have shown that SAM is able to inhibit enzyme activity, but there was no previous evidence for its involve- ment in the regulation of mRNA levels.
(3) The levels of mRNA in pMet54ransformed cells are about ten-fold higher than in wt or in X-4003 untransformed cells, and this relates well with the plasmid copy number. However, we have also found that in the same transformant strain the specific activity of HSTase is only slightly higher than in wt cells. These findings suggest that a post-tran- scriptional regulatory mechanism may be operative, otherwise we would have observed an increase of enzyme activity comparable with the mRNA level, as shown for MET3 and MElZ yeast gene products.
ACKNOWLEDGEMENTS
This work was supported by CNR grant, Progetto Finalizzato Ingegneria Genetica e Basi Molecolari delle Malattie Ereditarie CT 8501386.51.
REFERENCES
Bach, M.L., Lacroute, F. and Botstein, D.: Evidence for tran- scriptional regulation of orotidine-5’-phosphate decarboxy- lase in yeast by hybridization ofmRNA to the yeast structural gene cloned in Escherichiu coli. Proc. Natl. Acad. Sci. USA 76 (1979) 386-390.
Beach, D., Piper, M. and Nurse, P.: Construction of a Schizo-
saccharomyces porn& gene bank in a yeast bacterial shuttle vector and its use to isolate genes by complementation. Mol. Gen. Genet. 187 (1982) 326-329.
Bimboim, H.C. and Doly, J.: A rapid alkaline extraction proce- dure for screening recombinant plasmid DNA. Nucl. Acids Res. 7 (1979) 1513-1520.
Cherest, H., Surdin-Kejan, Y. and de Robichon-Szulmajster, H.: Methionine-mediated repression in Saccharomyces
cerevirioe: a pleiotropic regulatory system involvingmethionyl transfer ribonucleic acid and the product of gene eth2. J. Bacterial. 106 (1971) 758-772.
Cherest, H., Surdin-Kerjan, Y., Antoniewsky, J. and de Robichon-Szuhnajster, H.: S-Adenosyhnethionine mediated repression of methionine biosynthetic enzymes in Succharo-
myces cerevisiae. J. Bacterial. 114 (1973a) 928-933. Cherest, H., Surdin-Kerjan, Y., Antoniewsky, J. and de
Robichon-Szulmajster, H.: Effects of regulatory mutations
upon methionine biosynthesis in Sacchoromyces cere-
visiue:loci er&, eth3, ethZ0. J. Bacterial. 115 (1973b) 1084-1093.
Cherest, H., Thao, N.N. and Surdin-Kerjan, Y.: Transcriptional regulation of the MET3 gene of Saccharomyces cereviriae.
and translational expression of a chimeric bacterial-yeast plasmid in yeasts. Gene 11 (1980) 1 l-19.
Crabeel, M., Messenguy, F., Lacroute, F. and Glansdorff, N.: Cloning ARG3, the gene for omithine carbamoyltransferase from Saccharomyces cerevidae: expression in Escherichia coli
requires secondary mutations; production of plasmid beta- lactamase in yeast. Proc. Natl. Acad. Sci. USA 78 (1981) 5026-5030.
Cryer, D.R., Eccleshall, R. and Marmur, J.: Isolation of yeast DNA. In Prescott, D.M. (Ed.), Methods in Cell Biology. Academic Press, New York, 1975, Vol. 12, pp. 39-44.
de Robichon-Szulmajster, H. and Cherest, H.: Regulation of homoserine-O-transacetylase, first step in methionine biosynthesis in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 28 (1967) 256-262.
Federoff, H.J., Cohen, J.D., Eccleshall, T.R., Needleman, R.B., Buchferer, B.A., Giacalone, 3. and Mat-mm, J.: Isolation of a maltase structural gene from Saccharomyces cereviriae.
J. Bacterial. 149 (1982) 1064-1070. Hinnen, A,, Hicks, J.B. and Fink, G.R.: Transformation of yeast.
Proc. Acad. Natl. Sci. USA 75 (1978) 1929-1933. Ito, H., Fukuda, Y., Murata, K. and Kimura, A.: Transformation
of intact yeast cells treated with alkali cations. J. Bacterial.
A Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1982.
Melton, D.A., Krieg, P.A., Rebagliati, M.R., Maniatis, T., Zinn, K. and Green, M.R.: Efficient ‘in vitro’ synthesis of biologi- cally active RNA and RNA hybridization probes from plasmids containing a bacteriophage SP6 promoter. Nucl.
Acids Res. 18 (1984) 7035-7050. Nagai, S. and Flavin, M.: Synthesis of O-acetylhomoserine.
Methods Enzymol. 17B (1971) 423-424. Nagai, S. and Kerr, D.: Homoserine transacetylase. Methods
Enzymol. 17B (1971) 442-445.
Oesch, B., Westaway, D., Walchli, M., McKinley, M.P., Kent, S.B.H., Aebersold, M., Barry, R.A., Tempts, P., Teplow, D.B., Hood, L.E., Prusiner, S.B. and Weissmann, C.: A cellular gene encodes scrapie PrP 27-30 protein. Cell 40 (1985) 735-746.
Orr-Weaver, T.L., Szostak, J.W. and Rothstein, R.J.: Yeast transformation: a model system for the study of recombi- nation. Proc. Natl. Acad. Sci. USA 78 (1981) 6354-6358.
Popolo, L., Vanoni, M. and Alberghina, L.: Control of the yeast cell cycle by protein synthesis. Exp. Cell Res. 142 (1982) 69-78.
Sangspda, S., Cherest, H. and Surdin-Kerjan, Y.: The expression of the MET25 gene of Saccharomyces cerevisiae is regulated transcriptionally. Mol. Gen. Genet. 200 (1985) 407-414.
78
Schild, D. and Mortimer, RX.: A mapping method for Saccharo-
Schmitt, J.J. and Cohen, B.N.: Quantitative isolation of DNA restriction fragments from low-melting agarose by Elutip-d affinity chromatography. Anal. Biochem. 133 (1983) 462-464.
Sur&&erjan, Y. and de Robichon&ulmajster, PI.: Existence
of two levels of repression in the biosynthesis of methionine in Succhuromyces cwevkiue: efkct of lomo!kngin on enzyme synthesis. J. Bacterial. 122 (1975) 367-374.
Zamenhof, S.: Preparation and assay of deoxyribonucleic acid from animal tissue. Methods Enxymol. 3 (1957) 696-704.
Simchen, G., Pinon, R. and Salts, Y.: Sporulation in Saccharo-
myces cerevisiae: premeiotic DNA synthesis, readiness and commitment. Exp. Cell. Res. 75 (1972) 207-218.