Page 1
Molecular cloning and characterization of a nonsymbiotichemoglobin gene (GLB1) from Malus hupehensis Rehd.with heterologous expression in tomato
Xingzheng Shi • Xinliang Wang • Futian Peng •
Yu Zhao
Received: 18 September 2011 / Accepted: 16 April 2012 / Published online: 25 April 2012
� Springer Science+Business Media B.V. 2012
Abstract Nonsymbiotic hemoglobins (nsHbs) are
involved in a variety of cellular processes in plants. Previous
studies indicate that nsHb expression improves plant toler-
ance during waterlogging and hypoxia. In the present work,
the nsHb class-1 coding sequence was cloned from Malus
hupehensis Rehd. var. pinyiensis Jiang and subsequently
named MhGLB1. The results elucidated the expressed
characteristics and physiological effects of MhGLB1. The
full-length cDNA contained a 477 bp open reading frame
encoding a protein with a molecular mass of 17.8 KDa with
158 amino acids. Quantitative real-time PCR analysis
showed that MhGLB1 expresses in roots, stems and leaves
growing under normal and nitrate-induced conditions.
Hypoxic stress induced accumulation of MhGLB1 within
12 h, and abscisic acid significantly induced expression of
MhGLB1 in roots. The photosynthetic, transpiration and
stomatal conductance rates of transgenic MhGLB1 tomato
plants decreased more slowly than that of wild-type plants
under waterlogging treatment. These results indicated that
the MhGLB1 gene has an important role in hypoxia.
Keywords Hypoxic stress � Malus hupehensis Rehd �MhGLB1 � Nonsymbiotic hemoglobin
Introduction
Hemoglobins (Hbs) were originally described in animals as
cells that facilitate oxygen transport in blood. Plant Hbs
were first isolated from root nodules of symbiotic nitrogen
(N) -fixing plant species, where they were expected to play
a role in binding and transport of molecular oxygen.
However, since the discovery of expressed hemoglobin
genes in non-nodulating plant species, the research for non-
symbiotic functions of plant Hbs has been ongoing. Three
distinct types of Hbs have been characterized in plants:
symbiotic, nonsymbiotic and truncated. Symbiotic hemo-
globins (sHbs) are found primarily in plant nodules and
their function is to regulate the oxygen supply to N-fixing
bacteria for symbiotic N fixation [1]. Nonsymbiotic
hemoglobin (NsHbs) are not involved in symbiotic N fix-
ation, but have an affinity for oxygen and may serve a
physiological function. These forms of hemoglobin are
believed to exist throughout the plant kingdom. The role of
truncated Hbs remains unclear. However, the most recent
research suggested that truncated Hbs share some charac-
teristics with nsHbs [2].
The nsHbs are divided into class-1 and class-2 Hbs.
Class-1 (nsHb-1s) has a very high affinity for oxygen and is
most likely an evolutionary precursor to symbiotic Hbs [3].
NsHb-1s proteins are relatively conserved across the plant
kingdom, suggesting that members of this class might have
important physiological functions. Class-2 Hbs (nsHb-2s)
have a lower affinity for oxygen and are similar to the sHbs
[4]. Previous studies indicate that nsHb-1s gene expression
is induced by nitrate (NO3), nitrite (NO2), nitric oxide (NO)
[5] and hypoxia [4].
Recently, class-1 nsHb genes have been isolated from
monocots and dicots, including Arabidopsis, Trema,
Parasponia, Solanum, barley, rice, tomato, cotton and
maize [6]. In Arabidopsis, non-symbiotic hemoglobin
controls bolting by scavenging NO, which serves as a floral
transition signal molecule [7]. In barley, only one nsHb
gene exists with strong expression in plant roots grown
X. Shi � X. Wang � F. Peng (&) � Y. Zhao
State Key Laboratory of Corp Biology/College of Horticulture
Science and Engineering, Shandong Agricultural University,
Tai’an 271018, Shandong, People’s Republic of China
e-mail: [email protected]
123
Mol Biol Rep (2012) 39:8075–8082
DOI 10.1007/s11033-012-1654-4
Page 2
under hypoxic stress [8]. ZmHb mRNA levels in maize
seedlings are induced by high-salt and osmotic stress in
addition to hypoxic stress [9]. Similarly, two copies of
class-1 nsHb genes, OsGLB1a and OsGLB1b, were detec-
ted in rice with 94 % identity at the nucleotide level. Both
genes were strongly induced in roots by the addition of
NO3, NO2 or NO [5].
NsHb-1s genes have been identified that exhibit physi-
ological functions. For example, it has been shown that
nsHb proteins modulate cellular nitric oxide (NO) levels in
plants and play an important role in abiotic tolerance and
other cellular processes [4]. In the current study, we cloned
a pingyitiancha (Malus hupehensis Rehd. var. pinyiensis
Jiang PYTC) nsHb-1s gene MhGLB1. Quantitative real-
time PCR assays revealed the expression of MhGLB1 was
induced by NO3, hypoxic stress and abscisic acid (ABA).
Furthermore, over-expression of MhGLB1 in transgenic
tomato plants improved plant tolerance to waterlogging.
Materials and methods
Plant material and growth conditions
PYTC seeds were germinated on vermiculite. Seedlings
were grown in vermiculite irrigated with distilled water for
4 weeks in a green house at 24 �C under fluorescent light
and a photoperiod of 16 h (day)/8 h (night). The 4 weeks
old seedlings were transferred to a growth chamber pro-
viding the same conditions as a greenhouse. However the
seedlings were grown hydroponically in aerated Hoa-
gland’s nutrient solution containing 0.8 mM NO3 for
1 week to generate N-deprived seedlings. Following
treatment, the seedlings were removed from the treatment
solution and dried with paper towels. Whole leaves, stems
and roots were excised, poured into liquid N and stored at
-80 �C.
Search for candidate MhGLB1 expressed sequence tags
(ESTs)
The amino acid sequences of each GLB1 in model plants
were used to search the apple (Malus) EST database with
tBLASTn (http://www.ncbi.nlm.nih.gov/BLAST/).
RNA isolation and cDNA synthesis
Total RNA was extracted from 0.1 g of tissue from
2 weeks old seedlings with TRIzol (Invitrogen Inc., USA)
reagent then treated with RNase-free DNaseI at room
temperature for 15 min in reaction buffer containing
20 mM Tris–HCl (pH 8.4), 2 mM MgCl2, and 50 mM
KCl. DNaseI was inactivated by adding EDTA (2.5 mM
final concentration) and heated to 65 �C for 10 min. The
quality and quantity of total RNA were measured using
both electrophoresis and optical absorbency. Complemen-
tary DNAs were synthesized with the SuperScriptIII Kit
(Invitrogen Inc, USA) following the manufacturer’s
instructions.
Polymerase chain reaction (PCR) amplification, plant
expression vector construction and tomato
transformation
Primers were designed for PCR to amplify the PYTC nsHb
gene using sequences at the start and stop codons of Ma-
lus 9 domestica nsHb cDNA (GenBank accession number
AY224132). The primer sequences were sp1 (sense 50-GC
GGATCCATGGAAGGCAAAGTTTTC-30) and sp2 (anti-
sense 50-GCGAGCTCCTAATTAAGGGGAGGCTTCAT-
30). The restriction sites for BamHI and SacI are under-
lined, respectively. Total PYTC cDNA (*0.1 lg) was
used as a template for PCR amplification. PCR reactions
were conducted in the following reaction mixture: 0.4 lM
of each sense and antisense primer, 100 lM of 10 9 PCR
buffer and 2 mM MgCl2 in a final volume of 25 ll. PCR
amplification was carried out for 30 cycles at an annealing
temperature of 55.2 �C using a Mycycler thermacycler
(Bio-Rad, CA, USA). PCR products were detected in a 1 %
agarose gel after staining with ethidium bromide, isolated
from the gel using the GeneClean kit (Sangon, Shanghai,
China), and cloned into the pMD18-T cloning vector
(TaKaRa) following the manufacturer’s instructions,
sequenced to confirm identity with known GLB genes.
Following excision with the restriction enzymes BamHI
and SacI, the products were inserted into the plant
expression vector PBI121 (Clontech, Palo Alto, CA)
behind the 35S cauliflower mosaic virus (CaMV 35S)
promoter. The binary vector containing the GLB1 cDNA
construct was introduced into Agrobacterium tumefaciens
LBA4404 [10], which was subsequently used for tomato
transformation (Lycopersicon esculentum wild type cv.
Sy12f) [11]. Regenerated shoots with kanamycin resistance
were selected in MS medium [12] supplemented with
50 mg L-1 kanamycin, 3 mg L-1 6-benzyladenine (6-BA)
and 0.2 mg l-1 indoleacetic acid (IAA). Shoots were roo-
ted in 1/2MS medium supplemented with 0.3 mg l-1
indoleacetic acid (IAA). The REDExtract-N-Amp plant
PCR Kit (Sigma-Aldrich, Poole, Dorset, UK) was used to
confirm the kanamycin-resistant plantlets (T0) with sp1 and
sp2 for the transgenic tomato. The PCR-positive plantlets
were transplanted into soil. After 4 weeks of growth, the
plants were collected for quantitative real-time PCR anal-
ysis. The transgenic lines were selected by T1 seedling
segregation analysis following germination on kanamycin
medium. The kanamycin-resistant T1 seedlings were grown
8076 Mol Biol Rep (2012) 39:8075–8082
123
Page 3
to maturity to collect T2 seeds. The transgenic lines were
selected by T2 seedling segregation analysis following
germination on kanamycin medium. Total RNA was pre-
pared from leaves of 4 week old T2 plants for quantitative
real-time PCR analysis. The methods of RNA isolation and
cDNA synthesis have been described in the previous par-
agraph. T1 plants were also self-pollinated to obtain
homozygous plants, and T2 transgenic plants from T1
independent lines were analyzed.
Wild-type (WT) and transgenic tomato plants were
grown in the greenhouse under natural light supplemented
by sodium vapor lamps with a photoperiod of 14/10 h
(light/dark). Average day/night temperatures were
approximately 28/18 �C. The transgenic tomato plants
were transplanted into plastic pots and placed on plastic
trays in the greenhouse at 25 �C and a 14/10 h light/dark
cycle. Simulated flooding treatments were applied by
adding tap water to the plastic trays. The tap water for
waterlogging was equilibrated in the growing chamber at
25 �C for 24 h and the water level was maintained at the
pot soil surface. The waterlogging treatments were con-
ducted in both the transgenic lines and the controls. The
portable photosynthesis system CI-340 was used to mea-
sure the photosynthetic rate, stomatal conductance and
transpiration rate of the transgenic and control plants.
Quantitative real-time PCR
We examined the response of MhGLB1 in the presence of
NO3, hypoxia and ABA. Plants were N-deprived for 48 h
for the NO3 experiments, and subsequently transferred to
fresh medium supplemented with 10 mM KNO3 and
allowed to grow under the same conditions for 2, 6 and 8 h.
Seedlings grown in Hoagland’s nutrient solution containing
10 mM KCl served as controls. For ABA experiments,
seedlings were pretreated with 200 lM the NO scavenger
2-(4-carboxypheny)-4, 4, 5, 5,-tetramethilimidazoline-1-
oxyl-3-oxide (c-PTIO) and distilled water for 4 h, then
treated with 100 lM ABA for 4 h. The NO Detection Kit
(nitrate reductase) from Nanjing Jiancheng Bioengineering
Institute measured NO content. Seedlings were grown in
distilled water as controls. Seedlings were grown under
anaerobic conditions for 12 and 24 h for the hypoxia
experiments. Aerated seedlings served as controls. At
harvest, plants were separated into roots, stems and leaves.
Specific gene primers were designed from the MhGLB1
cDNA sequence and analyzed using the Primer 5.0 soft-
ware (PE Applied Biosystems) following the manufac-
turer’s guidelines. The r18s gene was used as the internal
constitutively expressed control (house-keeping gene).
Primers (reverse-R and forward-F) and [50] 6-FAM, [30]TAMRA-labeled probes (-P) listed in Table 1 were syn-
thesized by Invitrogen and used at a 200 nM final
concentration. PCR reactions were performed in a 25 ll
final volume according to the manufacturer’s protocol and
three PCR replicates were assayed for each sample. Real-
time quantitative PCR was conducted on a FTC2000
machine. PCR cycles were as follows: 1 cycle of 2 min at
50 �C; 10 min at 95 �C; followed by 40 cycles each of 15 s
at 95 �C, and 1 min at 60 �C. Control reactions were per-
formed without a template. Relative gene expression was
calculated according to the relative method (DCT) using
r18s as a constitutively expressed gene. Mean values of
2-DCT (DCT = CT(gene of interest) - CT(r18s)) were calcu-
lated from three independent experiments. PCR efficiencies
of MhGLB1, LeGLB1, Mhr18S and Ler18S were 98.12,
99.01, 98.30 and 98.57 %, respectively. In order to ensure
the accuracy of the experiment, intraassay variation was
\1.20 % and interassay variation \1.10 %.
Results
Full length MhGLB1 isolation and sequence analysis
We designed PCR primers from Malus cDNA to charac-
terize nsHb named MhGLB1. The full-length 477 bp PYTC
cDNA template, encoding a protein of 158 amino acids and
a mass of 17.8 KDa was amplified. The deduced nsHb
amino acid sequence exhibited a respective 95.57, 80.12,
82.82 and 79.50 % identity to pear, alfalfa, cotton and
soybean, and lower sequence identities of 65.76, 67.65,
71.17 and 71.69 % to orange, rice, wheat and maize,
Table 1 Primer and [50] 6-FAM, [30] TAMRA-labelled probe
sequences used in real-time quantitative PCR assays of genes
MhGLB1 and r18S. Accession numbers are given in parenthesis
Primer Sequence
MhGLB1 (EF690362)
GLB1-F: 50-CGCATTGTTGGAAACCATAAAG-30
GLB1-R: 50-TCATAAGCTTCTCCCCATGCA-30
GLB1-P: 50-AGGCCTTACCGGAAATGTGGTCA-30
Mhr18S (DQ341382)
Mhr18 s-F: 50-AAACGGCTACCACATCCA-30
Mhr18 s-R: 50-CACCAGACTTGCCCTCCA-30
Mhr18 s-P: 50-AGCAGGCGCGCAAATTACC-30
LeGLB1(AY026343)
LeGLB1-F: 50-GGTGTGGTTGATGAGCACTTTGA-30
LeGLB1-R: 50-AGGCCTCTCCCCATGCATTCT-30
LeGLB1-P: 50-CAAAATATGCCTTGTTGGAGA-30
Ler18S (X51576)
Ler18s-F: 50-GCCCGGGTAATCTTTGAAAT-30
Ler18s-R: 50-AGTAAGCGCGAGTCATCAGC-30
Ler18s-P: 50-CGGATCATTCAATCGGTAGG-30
Mol Biol Rep (2012) 39:8075–8082 8077
123
Page 4
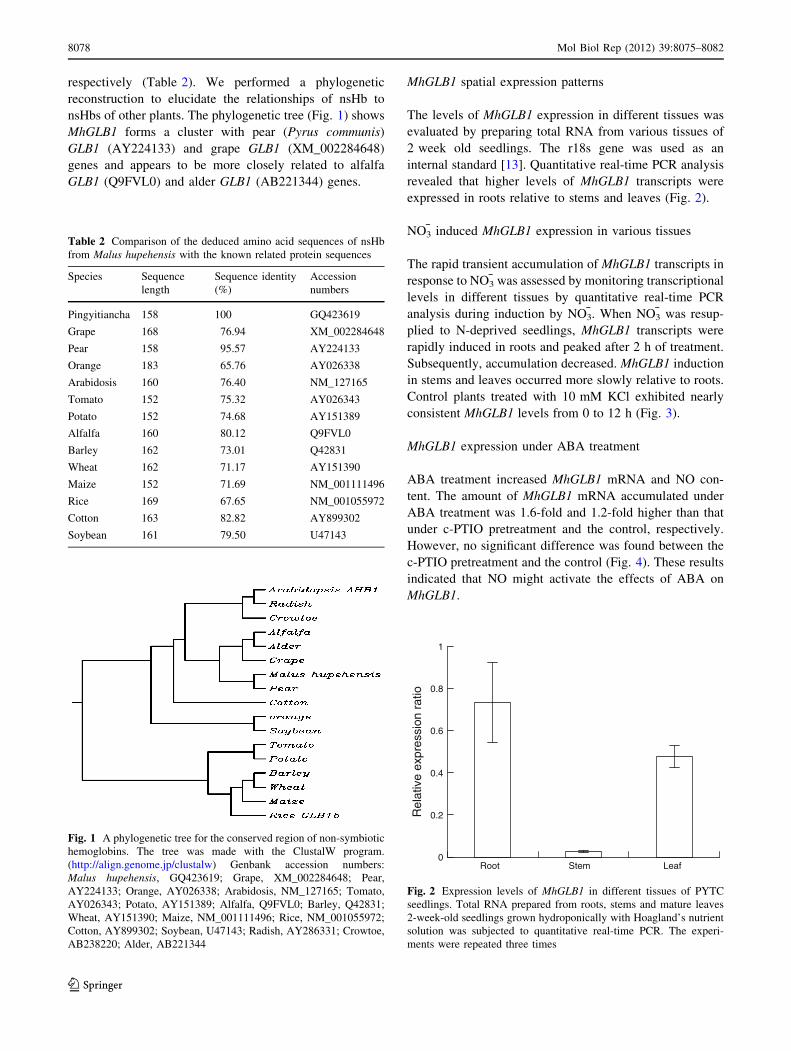
respectively (Table 2). We performed a phylogenetic
reconstruction to elucidate the relationships of nsHb to
nsHbs of other plants. The phylogenetic tree (Fig. 1) shows
MhGLB1 forms a cluster with pear (Pyrus communis)
GLB1 (AY224133) and grape GLB1 (XM_002284648)
genes and appears to be more closely related to alfalfa
GLB1 (Q9FVL0) and alder GLB1 (AB221344) genes.
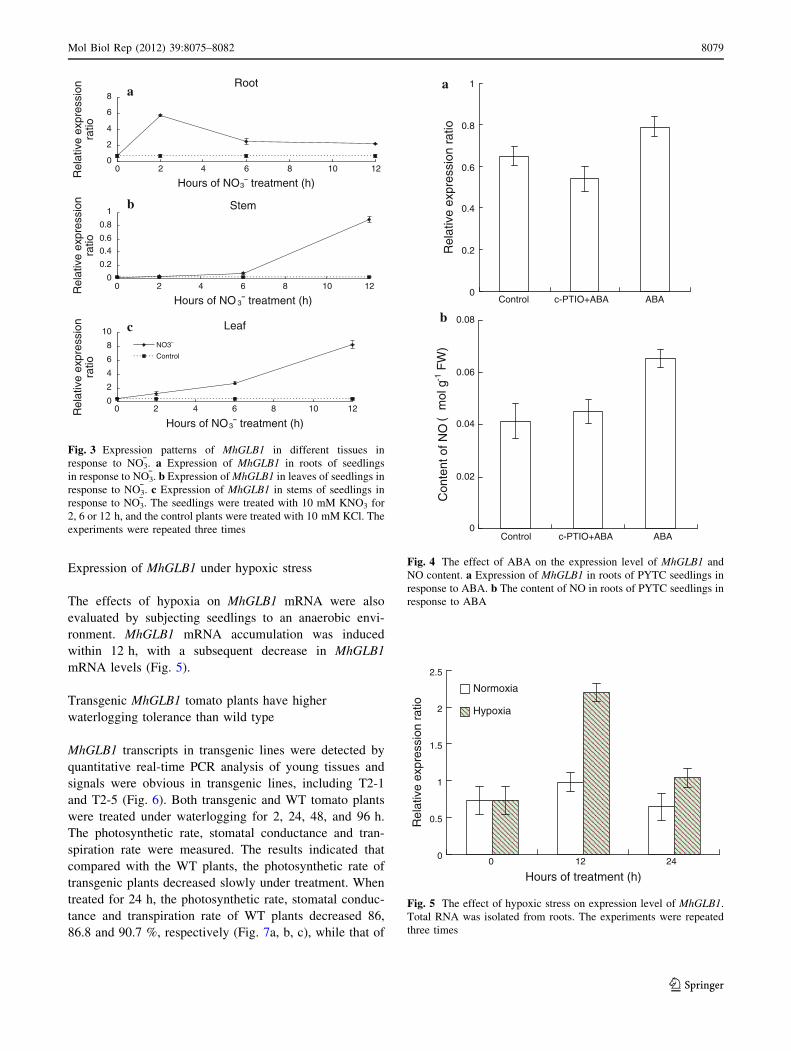
MhGLB1 spatial expression patterns
The levels of MhGLB1 expression in different tissues was
evaluated by preparing total RNA from various tissues of
2 week old seedlings. The r18s gene was used as an
internal standard [13]. Quantitative real-time PCR analysis
revealed that higher levels of MhGLB1 transcripts were
expressed in roots relative to stems and leaves (Fig. 2).
NO3 induced MhGLB1 expression in various tissues
The rapid transient accumulation of MhGLB1 transcripts in
response to NO3 was assessed by monitoring transcriptional
levels in different tissues by quantitative real-time PCR
analysis during induction by NO3. When NO3 was resup-
plied to N-deprived seedlings, MhGLB1 transcripts were
rapidly induced in roots and peaked after 2 h of treatment.
Subsequently, accumulation decreased. MhGLB1 induction
in stems and leaves occurred more slowly relative to roots.
Control plants treated with 10 mM KCl exhibited nearly
consistent MhGLB1 levels from 0 to 12 h (Fig. 3).
MhGLB1 expression under ABA treatment
ABA treatment increased MhGLB1 mRNA and NO con-
tent. The amount of MhGLB1 mRNA accumulated under
ABA treatment was 1.6-fold and 1.2-fold higher than that
under c-PTIO pretreatment and the control, respectively.
However, no significant difference was found between the
c-PTIO pretreatment and the control (Fig. 4). These results
indicated that NO might activate the effects of ABA on
MhGLB1.
Table 2 Comparison of the deduced amino acid sequences of nsHb
from Malus hupehensis with the known related protein sequences
Species Sequence
length
Sequence identity
(%)
Accession
numbers
Pingyitiancha 158 100 GQ423619
Grape 168 76.94 XM_002284648
Pear 158 95.57 AY224133
Orange 183 65.76 AY026338
Arabidosis 160 76.40 NM_127165
Tomato 152 75.32 AY026343
Potato 152 74.68 AY151389
Alfalfa 160 80.12 Q9FVL0
Barley 162 73.01 Q42831
Wheat 162 71.17 AY151390
Maize 152 71.69 NM_001111496
Rice 169 67.65 NM_001055972
Cotton 163 82.82 AY899302
Soybean 161 79.50 U47143
Fig. 1 A phylogenetic tree for the conserved region of non-symbiotic
hemoglobins. The tree was made with the ClustalW program.
(http://align.genome.jp/clustalw) Genbank accession numbers:
Malus hupehensis, GQ423619; Grape, XM_002284648; Pear,
AY224133; Orange, AY026338; Arabidosis, NM_127165; Tomato,
AY026343; Potato, AY151389; Alfalfa, Q9FVL0; Barley, Q42831;
Wheat, AY151390; Maize, NM_001111496; Rice, NM_001055972;
Cotton, AY899302; Soybean, U47143; Radish, AY286331; Crowtoe,
AB238220; Alder, AB221344
0
0.2
0.4
0.6
0.8
1
Root Stem Leaf
Rel
ativ
e ex
pres
sion
rat
io
Fig. 2 Expression levels of MhGLB1 in different tissues of PYTC
seedlings. Total RNA prepared from roots, stems and mature leaves
2-week-old seedlings grown hydroponically with Hoagland’s nutrient
solution was subjected to quantitative real-time PCR. The experi-
ments were repeated three times
8078 Mol Biol Rep (2012) 39:8075–8082
123
Page 5
Expression of MhGLB1 under hypoxic stress
The effects of hypoxia on MhGLB1 mRNA were also
evaluated by subjecting seedlings to an anaerobic envi-
ronment. MhGLB1 mRNA accumulation was induced
within 12 h, with a subsequent decrease in MhGLB1
mRNA levels (Fig. 5).
Transgenic MhGLB1 tomato plants have higher
waterlogging tolerance than wild type
MhGLB1 transcripts in transgenic lines were detected by
quantitative real-time PCR analysis of young tissues and
signals were obvious in transgenic lines, including T2-1
and T2-5 (Fig. 6). Both transgenic and WT tomato plants
were treated under waterlogging for 2, 24, 48, and 96 h.
The photosynthetic rate, stomatal conductance and tran-
spiration rate were measured. The results indicated that
compared with the WT plants, the photosynthetic rate of
transgenic plants decreased slowly under treatment. When
treated for 24 h, the photosynthetic rate, stomatal conduc-
tance and transpiration rate of WT plants decreased 86,
86.8 and 90.7 %, respectively (Fig. 7a, b, c), while that of
Stem
0
0.2
0.4
0.6
0.8
1
Hours of NO 3¯ treatment (h)
b
Root
0
2
4
6
8
Hours of NO3¯ treatment (h)
a
Leaf
0
2
4
6
8
10
0 2 4 6 8 10 12
0 2 4 6 8 10 12
0 2 4 6 8 10 12
Hours of NO3¯ treatment (h)
NO3¯
Control
c
Rel
ativ
e ex
pres
sion
ratio
Rel
ativ
e ex
pres
sion
ratio
Rel
ativ
e ex
pres
sion
ratio
Fig. 3 Expression patterns of MhGLB1 in different tissues in
response to NO3. a Expression of MhGLB1 in roots of seedlings
in response to NO3. b Expression of MhGLB1 in leaves of seedlings in
response to NO3. c Expression of MhGLB1 in stems of seedlings in
response to NO3. The seedlings were treated with 10 mM KNO3 for
2, 6 or 12 h, and the control plants were treated with 10 mM KCl. The
experiments were repeated three times
b
0
0.02
0.04
0.06
0.08
Con
tent
of N
O(µ
mol
g-1
FW
)
a
0
0.2
0.4
0.6
0.8
1
Control c-PTIO+ABA ABA
Control c-PTIO+ABA ABA
Rel
ativ
e ex
pres
sion
rat
io
Fig. 4 The effect of ABA on the expression level of MhGLB1 and
NO content. a Expression of MhGLB1 in roots of PYTC seedlings in
response to ABA. b The content of NO in roots of PYTC seedlings in
response to ABA
0
0.5
1
1.5
2
2.5
0 12 24
Hours of treatment (h)
Rel
ativ
e ex
pres
sion
rat
io
Normoxia
Hypoxia
Fig. 5 The effect of hypoxic stress on expression level of MhGLB1.
Total RNA was isolated from roots. The experiments were repeated
three times
Mol Biol Rep (2012) 39:8075–8082 8079
123
Page 6
transgenic plants decreased 40.1, 72.5 and 78.7 %,
respectively (Fig. 7a, b, c).
Discussion
Previously, sHbs had only been identified in animals.
However, Hbs have been characterized from all kingdoms
of life and known to function as oxygen carriers, oxygen
sensors, and in oxygen storage; in NO detoxification, and
in peroxidase activity. NsHbs encoding genes in plants
have been isolated, but few GLB1s in woody plants have
been detected. PYTC is an important rootstock for apple
production in China and exhibits many valuable properties.
To obtain an increased understanding of MhGLB1
expression, we identified and isolated nsHbs cDNA, called
MhGLB1, from young roots of PYTC. The phylogenetic
tree showed MhGLB1 has nearest homologous relationship
with pear GLB1. It will be as the PYTC nsHb-1s hemo-
globin encoding gene.
Our study detected MhGLB1 mRNA in roots, leaves and
stems, with the highest gene expression in root tissue
(Fig. 2). In Arabidopsis, the nsHb-1s are active in germi-
nating seedlings and can be induced by hypoxia and by
increased sucrose addition [14]. Barley shows nsHb-1s
expressed in the aleurone layers and roots subjected to low
oxygen stress [8], and expression is not regulated directly
by oxygen availability, but by ATP or some consequence
of ATP action [15]. Under normal growth conditions in
rice, nsHb-1s and -2s are expressed in leaves, however only
nsHb-1s are expressed in roots [16]. Our results and pre-
vious research together suggest that this gene is expressed
in different tissues (i.e. roots, stems and leaves), but
exhibits tissue-specific expression in other plant species.
In angiosperms, NO3 is not only a nutrient but also
serves as a signal for regulating gene expression, such as
glutamine synthetase (GLN) and asparagine synthetase
(ASN) [17]. Rapid induction of MhGLB1 transcripts in
roots within 2 h following addition of NO3 and down-
regulation after 6 h (Fig. 3a), suggests NO3 sensitivity to
the regulatory mechanisms of MhGLB1 expression. How-
ever, decreased MhGLB1 induction and subsequent accu-
mulation was observed in stems and leaves relative to root
tissues (Fig. 3b, c). We hypothesize a signal transition
process from roots to stems to leaves operates throughout
induction. Two nsHb-1s genes, ORYsaGLB1a and ORY-
saGLB1b, were strongly induced by NO3. NO3 began to
induce accumulation of both ORYsaGLB1a and ORY-
saGLB1b transcripts after 1–2 h of treatment, with amounts
peaking at 4–8 h, followed by a decrease in accumulation.
In this study, our results together with previous studies
0.000
0.050
0.100
0.150
0.200
0.250
0.300
0.350
WT T2-1 T2-5
Rel
ativ
e ex
pres
sion
rat
io
MhGLB1
LeGLB1
Fig. 6 Quantitative real-time PCR analysis expression of MhGLB1 in
transgenic tomato plants. LeGLB1, the GLB1 gene of tomato.
Quantitative real-time PCR analysis was performed using total
RNA from the young leaves of the plants. The experiments were
repeated three times
c
0
1
2
3
4
Hours of treatment (h)
Tra
nspi
ratio
n ra
te
(mm
ol m
-2 s
-1) T2-1
T2-5
WT
0
02468
101214161820
Hours of treatment (h)
Pho
tosy
nthe
tic R
ate
µ mol
CO
2 m
-2 s
-1
a
b
0
20
40
60
80
100
0 20 40 60 80 100
20 40 60 80 100
0 20 40 60 80 100
Hours of treatment (h)
Sto
mat
al c
ondu
ctan
ce
(mm
ol H
2O m
-2 s
-1)
-2
Fig. 7 Changes in leaf photosynthesis rates, stomatal conductance
and transpiration rate in transgenic tomato plants and WT under
waterlogging treatment. a Change of leaf photosynthesis rate in
transgenic tomato and WT plants. b Change of stomatal conductance
in transgenic tomato and WT plants. c Change of transpiration rate in
transgenic tomato and WT plants
8080 Mol Biol Rep (2012) 39:8075–8082
123
Page 7
suggest that GLB1 gene could respond to NO3 signals at the
transcriptional level in some species.
In this study, MhGLB1 mRNA accumulated under ABA
treatment and was higher than the c-PTIO pretreatment and
control. Significant differences were not detected between
c-PTIO pretreatment and the control. These results indi-
cated that the effects of ABA on MhGLB1 maybe activated
by NO (Fig. 4).
The accumulation of MhGLB1 mRNA was detected
within 12 h, and subsequently declined under hypoxic
stress. GLB1 mRNA gene expression is induced in isolated
barley aleurone layers exposed to anaerobic conditions, and
in the roots of flood-stressed barley plants [8]. These results
indicated that MhGLB1 gene could respond to hypoxic
stress, and it maybe play an important role in protecting
plants from hypoxic stress.
Overexpression of a candidate gene in a model plant
species, such as Arabidopsis and tomato, may provide
important information for understanding gene function. In
this research, we introduced MhGLB1 under the control of
the 35S CaMV promoter into tomato plants. Two inde-
pendent kanamycin-resistant transformants were identified
by quantitative real-time PCR.
In the present study, compared with wild-type tomato,
MhGLB1 overexpression enhanced tomato tolerance under
waterlogging. The photosynthetic rate of transgenic plants
decreased slowly, whereas the wild-type photosynthetic
rate declined significantly (Fig. 7a). Stomatal conductance
and transpiration rate for wild-type plants declined by a
large margin, and a slower decrease was observed in
transgenic plants (Fig. 7b, c). Overexpression of a GLB1
protects Arabidopsis thaliana from the effects of severe
hypoxia [18]. Increased tolerance of transgenic plants
under waterlogging was supported by hemoglobin but
regulated by NO levels. Previous studies have demon-
strated that NO is formed during hypoxia in alfalfa root
cultures and the levels of NO detected were inversely
related to the levels of nsHb-1s expression in tissues [4].
During the last 2 years, an increasing number of reports
have implicated nsHbs as the key enzymatic system for NO
scavenging in plants, indicating that the function of
hemoglobin may well be to protect against nitrosative
stress and to modulate NO signaling functions [19]. In
transgenic alfalfa root cultures, extracts from lines over-
expressing hemoglobin had approximately twice the NO
conversion rate of either control or antisense lines under
normoxic conditions. Only the control line showed a sig-
nificant increase in the rate of NO degradation when placed
under anaerobic conditions [20]. Nonsymbiotic hemoglo-
bin AHB1 was shown to scavenge NO through production
of S-nitrosohemoglobin and reduce NO emission under
hypoxic stress in Arabidopsis thaliana [21]. Our data in
conjunction with other studies has led us to conclude that
the MhGLB1 gene has an important role in hypoxia. Future
studies should obtain the overexpression or antisense
MhGLB1 gene in PYTC plants to confirm these results.
This approach will precisely clarify the roles of the
MhGLB1 gene and serve to elucidate the potential appli-
cation of MhGLB1 in enhancing plant stress tolerance.
Acknowledgments This work was supported by the Earmarked
Fund for Modern Agro-industry Technology Research System.
References
1. Appleby C (1992) The origin and functions of haemoglobin in
plants. Sci Prog 76:365–398
2. Watts RA, Hunt PW, Hvitved AN, Hargrove MS, Peacock WJ,
Dennis ES (2001) A hemoglobin from plants homologous to
truncated hemoglobins of microorganisms. Proc Natl Acad Sci
USA 98:10119–10124. doi:10.1073/pnas.191349198
3. Duff SMG, Wittenberg JB, Hill RD (1997) Expression, purifi-
cation, and properties of recombinant barley (Hordeum sp.)
hemoglobin: optical spectra and reactions with gaseous ligands.
J Biol Chem 272:16746–16752. doi:10.1074/jbc.272.27.16746
4. Dordas C, Hasinoff BB, Igamberdiev AU, Manac’h N, Rivoal J,
Hill RD (2003) Expression of a stress-induced hemoglobin affects
NO levels produced by alfalfa root cultures under hypoxic stress.
Plant J 35:763–770. doi:10.1046/j.1365-313X.2003.01846.x
5. Ohwaki Y, Kawagishi-Kobayashi M, Wakasa K, Fujihara S,
Yoneyama T (2005) Induction of class-1 non-symbiotic hemo-
globin genes by nitrate, nitrite and nitric oxide in cultured rice
cells. Plant Cell Physiol 46:324–331. doi:10.1093/pcp/pci030
6. Hebelstrup KH, Igamberdiev AU, Hill RD (2007) Metabolic
effects of hemoglobin gene expression in plants. Gene 398:
86–93. doi:10.1016/j.gene.2007.01.039
7. Hebelstrup KH, Jensen EØ (2008) Expression of NO scavenging
hemoglobin is involved in the timing of bolting in Arabidopsis
thaliana. Planta 227:917–927. doi:10.1007/s00425-007-0667-z
8. Taylor ER, Nie XZ, MacGregor AW, Hill RD (1994) A cereal
haemoglobin gene is expressed in seed and root tissues under
anaerobic conditions. Plant Mol Biol 24:853–862. doi:10.1007/
BF00014440
9. Zhao L, Gu RL, Gao P, Wang GY (2008) A nonsymbiotic
hemoglobin gene from maize, ZmHb, is involved in response to
submergence, high-salt and osmotic stresses. Plant Cell Tiss
Organ Cult 95:227–237. doi:10.1007/s11240-008-9436-3
10. Hoekma A, Hirsch PR, Hooykass PJJ, Schilperoort R (1983) A
binary plant vector strategy based on separation of the vri- and
T-region of Agrobacterium tumefaciens Ti plasmid. Nature
303:179–180
11. McCormac AC, Cherry JR, Hershey HP, Vierstra RD, Smith H
(1991) Photoresponses of transgenic tobacco plants expressing an
oat phytochrome gene. Planta 185:162–170. doi:10.1007/BF001
94057
12. Murashig T, Skoog F (1962) A revised medium for rapid growth
and bioassays with tobacco tissues. Plant Physiol 15:473–497.
doi:10.1111/j.1399-3054.1962.tb08052.x
13. Peng J, Peng FT, Zhu CF, Wei SC (2008) Molecular cloning of a
putative gene encoding isopentenyltransferase from pingyitiancha
(Malus hupehensis) and characterization of its response to nitrate.
Tree Physiol 28:899–904
14. Hunt PW, Watts RA, Trevaskis B, Llewelyn DJ, Burnell J,
Dennis ES, Peacock WJ (2001) Expression and evolution of
functionally distinct haemoglobin genes in plants. Plant Mol Biol
47:677–692. doi:10.1023/A:1012440926982
Mol Biol Rep (2012) 39:8075–8082 8081
123
Page 8
15. Nie X, Hill RD (1997) Mitochondrial respiration and hemoglobin
gene expression in barley aleurone tissue. Plant Physiol
114:835–840. doi:10.1104/pp.114.3.835
16. Arredondo-Peter R, Hargrove MS, Sarath G, Moran JF, Lohrman
J, Olson JS, Klucas RV (1997) Rice hemoglobins: gene cloning,
analysis and oxygen-binding kinetics of a recombinant protein
synthesized in Escherichia coli. Plant Physiol 115:1259–1266
17. Hsieh MH, Lam HM, Frank J, van de Loo FJ, Coruzzi G (1998) A
PII-like protein in Arabidopsis: Putative role in nitrogen sensing.
Proc Natl Acad Sci USA 95:13965–13970. doi:10.1073/pnas.95.
23.13965
18. Hunt PW, Klok EJ, Trevaskis B, Watts RA, Ellis MH, Peaeock
WJ, Dennis ES (2002) Increased level of hemoglobin 1 in
enhances survival of hypoxic stress and promotes early growth in
Arabidopsis thaliana. Proc Natl Acad Sci USA 99:17197–17202.
doi:10.1073/pnas.212648799
19. Perazzolli M, Romero-Puertas MC, Delledonne M (2006) Mod-
ulation of nitric oxide bioactivity by plant hemoglobins. J Exp
Bot 57:479–488. doi:10.1093/jxb/erj051
20. Igamberdiev AU, Seregelyes C, Manach N, Hill RD (2004)
NADH-dependent metabolism of nitric oxide in alfalfa root
cultures expressing barley hemoglobin. Planta 219:95–102. doi:
10.1007/s00425-003-1192-3
21. Perazzolli M, Dominici P, Romero-puertas MC, Zago E, Zeier J,
Sonoda M, Lamb C, Delledonne M (2004) Arabidopsis non-
symbiotic hemoglobin Ahb1 modulates nitric oxide bioactivity.
Plant Cell 16:2785–2794. doi:10.1105/tpc.104.025379B
8082 Mol Biol Rep (2012) 39:8075–8082
123