Molecular evidence supports hypervariability in Phytophthora colocasiae associated with leaf blight of taro Vishnu Sukumari Nath & Muthukrishnan Senthil & Vinayaka Mahabaleswar Hegde & Muthulekshmi Lajapathy Jeeva & Raj Shekhar Misra & Syamala Swayamvaran Veena & Mithun Raj Accepted: 23 January 2013 / Published online: 3 February 2013 # KNPV 2013 Abstract The oomycete Phytophthora colocasiae that causes taro leaf blight is the most devastating disease of taro and is widely distributed worldwide. Molecular and phenotypic techniques were employed for assessing and exploiting the genetic variability among four populations of P. colocasiae obtained from a fine spatial scale (multiple leaf blight lesions on single taro leaf). Phenotypic characters such as virulence, morphology and mating type showed no variation. ITS characterization revealed detectable polymorphism among isolates of P. colocasiae. The mean number of haplotypes (H), haplotype diversity (HD), nucleotide diversity (π), and nucleotide substi- tution rate (θ) among analyzed sequences were 6.75, 1.00, 0.069, and 0.088 respectively. High levels of inter and intra specific variation were detected by random amplified polymorphic DNA (RAPD) assays. Moderate genetic diversity (H =0.2651) was observed among populations of P. colocasiae. Analysis of molecular variance (AMOVA) confirmed that most of the genetic variability was confined to within a population (63.54 %). The coefficient of genetic dif- ferentiation among populations (G ST ) was 0.2007 and estimates of gene flow (Nm) among populations was 1.991 migrants per generation. Cluster analysis using UPGMA revealed that individuals from the same pop- ulation failed to cluster in one distinct group. The results of the study reveal considerable genetic diver- sity among and within populations of P. colocasiae obtained from fine spatial scale. The possible mecha- nisms and implications of this genetic variation are discussed. Keywords Single leaf . RAPD markers . P. colocasiae . Genetic diversity . Fine spatial scale . Multiple lesions Introduction Taro [Colocasia esculenta (L.) Schott] is an important tropical tuber crop, used as a staple food or subsistence food by millions of people in the developing countries in Asia, Africa and Central America (Sharma et al. 2008). The corms, leaves and petioles are used as a vegetable and considered as a rich source of carbohydrates, pro- teins, minerals and vitamins (Lakhanpaul et al. 2003). Taro corms and leaves are also credited to have medic- inal values and are used to reduce tuberculosis, ulcers, pulmonary congestion and fungal infection (Sharma et Eur J Plant Pathol (2013) 136:483–494 DOI 10.1007/s10658-013-0181-z Electronic supplementary material The online version of this article (doi:10.1007/s10658-013-0181-z) contains supplementary material, which is available to authorized users. V. S. Nath : M. Senthil : V. M. Hegde (*) : M. L. Jeeva : S. S. Veena : M. Raj Division of Crop Protection, Central Tuber Crops Research Institute, Thiruvananthapuram 695017, Kerala, India e-mail: [email protected]R. S. Misra Regional Centre of CTCRI, Dumduma HBC P.O, Bhubaneswar 751 019, Orissa, India
Transcript
Molecular evidence supports hypervariabilityin Phytophthora colocasiae associated with leafblight of taro
Accepted: 23 January 2013 /Published online: 3 February 2013# KNPV 2013
Abstract The oomycete Phytophthora colocasiaethat causes taro leaf blight is the most devastatingdisease of taro and is widely distributed worldwide.Molecular and phenotypic techniques were employedfor assessing and exploiting the genetic variabilityamong four populations of P. colocasiae obtainedfrom a fine spatial scale (multiple leaf blight lesionson single taro leaf). Phenotypic characters such asvirulence, morphology and mating type showed novariation. ITS characterization revealed detectablepolymorphism among isolates of P. colocasiae. Themean number of haplotypes (H), haplotype diversity(HD), nucleotide diversity (π), and nucleotide substi-tution rate (θ) among analyzed sequences were 6.75,1.00, 0.069, and 0.088 respectively. High levels ofinter and intra specific variation were detected byrandom amplified polymorphic DNA (RAPD) assays.Moderate genetic diversity (H=0.2651) was observed
among populations of P. colocasiae. Analysis ofmolecular variance (AMOVA) confirmed that mostof the genetic variability was confined to within apopulation (63.54 %). The coefficient of genetic dif-ferentiation among populations (GST) was 0.2007 andestimates of gene flow (Nm) among populations was1.991 migrants per generation. Cluster analysis usingUPGMA revealed that individuals from the same pop-ulation failed to cluster in one distinct group. Theresults of the study reveal considerable genetic diver-sity among and within populations of P. colocasiaeobtained from fine spatial scale. The possible mecha-nisms and implications of this genetic variation arediscussed.
Keywords Single leaf . RAPDmarkers .P. colocasiae .
Genetic diversity . Fine spatial scale .Multiple lesions
Introduction
Taro [Colocasia esculenta (L.) Schott] is an importanttropical tuber crop, used as a staple food or subsistencefood bymillions of people in the developing countries inAsia, Africa and Central America (Sharma et al. 2008).The corms, leaves and petioles are used as a vegetableand considered as a rich source of carbohydrates, pro-teins, minerals and vitamins (Lakhanpaul et al. 2003).Taro corms and leaves are also credited to have medic-inal values and are used to reduce tuberculosis, ulcers,pulmonary congestion and fungal infection (Sharma et
Electronic supplementary material The online version of thisarticle (doi:10.1007/s10658-013-0181-z) contains supplementarymaterial, which is available to authorized users.
V. S. Nath :M. Senthil :V. M. Hegde (*) :M. L. Jeeva :S. S. Veena :M. RajDivision of Crop Protection,Central Tuber Crops Research Institute,Thiruvananthapuram 695017, Kerala, Indiae-mail: [email protected]
R. S. MisraRegional Centre of CTCRI, Dumduma HBC P.O,Bhubaneswar 751 019, Orissa, India
al. 2008). Taro corms are utilized in various industriesfor the preparation of high fructose syrup and alcohols(Misra et al. 2008). These prospects make taro one of themost important tuber crops.
Leaf blight and corm rot caused by Phytophthoracolocasiae, a hemi-biotrophic oomycete plant patho-gen, is the most destructive disease of taro (Raciborski1900). The disease affects the leaves and petioles,resulting in extensive damage of the foliage. In addi-tion, it causes post harvest rot of stored corms. Thedisease is characterized by the formation of brownishwater-soaked circular lesions on young and matureleaves. With the advancement of the disease, lesionsenlarge and become irregular in shape and dark brownin colour with yellow margins. Rapid dissemination ofpathogen occurs through the sporangia and epidemicsare common during favourable conditions (intermit-tent rainy weather). Leaf blight has become the singlebiggest constraint for taro production in all taro-growing countries, including India, causing yield los-ses of up to 30 % (Jackson et al. 1980; Thankappan1985; Misra and Chowdhury 1997). In India, thisdisease is more prominent in northern and easternparts, which are potential areas of taro production. InSouth India, this disease appears occasionally but inserious proportions (Misra and Chowdhury 1997).
A good breeding strategy and appropriate deploymentof plant resistance requires a deep understanding of thepathogen diversity and the way in which virulenceevolves in the pathogen population. Variation among P.colocasiae isolates in phenotypic characters such asgrowth rate, colony morphology, metalaxyl resistanceand virulence were recognized in old populations(Misra et al. 2011). Significant genetic diversity in P.colocasiae isolates from distant geographical origins hasbeen previously described (Lebot et al. 2003; Mishra etal. 2010; Nath et al. 2012). However, it is likely thatgenetic diversity may exist in isolates collected fromdistant locations, since microbial populations dependon genetic variation to respond to novel environmentalchallenges (Chamnanpunt et al. 2001). Drawing defini-tive conclusions on the genetic diversity of the pathogenrequires comparing the isolates from a finer spatial scale.A considerable level of genetic variability was observedin Plasmodiophora brassicae isolates obtained from thesame field of crucifers (Manzanares-Dauleux et al.2001). Isolates of P. infestans obtained from individuallesions of potato leaves have previously been shown tobe different genotypes (Koh et al. 1994). Recently,
Sullivan et al. (2010) illustrated the dynamic nature ofPhytophthora nicotianae isolates collected from a singletobacco field in a 4-year cultivar rotation experiment. Todate, no studies have been reported concerning geneticvariation between isolates of P. colocasiae from finespatial scale.
The goals of the present study were: (1) to isolate P.colocasiae from multiple disease lesions present on asingle taro leaf, (2) to analyze the diversity of theseisolates through cultural and molecular methods, and(3) to assess how the genetic diversity is portionedamong and within isolates of P. colocasiae.
More detailed knowledge about the variability ofthis pathogen is essential for understanding the geneticstructure of the populations and also would be usefulin providing information for breeding programs, epi-demiological studies and disease management.
Materials and methods
Biological materials
Isolates ofP. colocasiae used in this study were obtainedfrom multiple leaf blight lesions present on a single taroleaf (Supplementary Fig. 1). Two-three months old taroleaves showing multiple lesions were collected on arandom basis for the same field (approx. 800 m2) ofthe Central Tuber Crops Research Institute (CTCRI),Thiruvananthapuram. The leaf samples were collectedat an early phase of the leaf blight season and lesions ofapproximately the same size was used for isolation. P.colocasiae isolates from each lesion were treated indi-vidually, and grouped into population according to theirsource leaves. A total of 41 isolates comprising fourpopulations were obtained and named as pop A(n=9), pop B (n=11), pop C (n=11) and pop D (n=10).All isolates were confirmed as P. colocasiae bycomparing the morphology and sporangial charac-teristics with authentic cultures maintained byCTCRI, Thiruvananthapuram, India.
Isolation of pathogen
For isolation, leaf tissue segments of 1–2 cm from leafblight infected area were excised. The segments weresterilized in 70 % ethanol for 1 min, followed by 1 %sodium hypochlorite for 2 min. The fragments werethen rinsed twice with sterile distilled water. Leaf
484 Eur J Plant Pathol (2013) 136:483–494
segments were dried on sterile Whatman filter paper ina laminar flow hood and placed onto Phytophthora-selective media (rye agar amended with 20 mg/l rifa-mycin, 200 mg/l vancomycin, 200 mg/l ampicillin,68 mg/l pentachloronitrobenzene and 50 mg/l 50 %benomyl). Segments were incubated in Petri dishes for4–5 days at 28 °C, and mycelia were then transferredonto the carrot agar medium (CA; 250 g/l carrot and20 g/l agar). Each isolate was stored at −20 °C in 50 %glycerol (long-term storage) and at 15 °C on carrotagar (CA) slants in the dark (short-term storage).
Colony morphology
Colony morphology was studied on potato dextroseagar (PDA; 250 g/l potato, 20 g/l dextrose and 20 g/lagar) and carrot agar (CA) medium. A 5-mm discobtained from the periphery of the colony, in areas ofactive growth, was placed at the centre of Petri dishescontaining PDA and CA medium. Plates were incubat-ed at 28 °C in the dark for 2 weeks. Following incuba-tion, morphology of P. colocasiae was characterizedbased on the colony texture. Three replicates were usedfor each isolate to confirm the characteristics at similarincubation conditions as mentioned above.
Pathogenicity assay
For the pathogenicity assay, a modified floating discmethod was used. Five leaf disks (5×5 cm) of taro (cvSree Kiran, susceptible to leaf blight) were floated insterile distilled water in 200-mm glass Petri plates andinoculated with a mycelial disc (5 mm) excised fromthe margins of actively growing cultures of P. coloca-siae. Leaf pieces with noncolonized agar plug servedas control treatments. Plates were covered with lidscontaining moistened filter paper to maintain highhumidity. After 4 days of incubation at 25 °C in thedark, the leaf disks were examined visually for diseasesymptoms.
Determination of mating type
The mating type of isolates was determined by pairingeach unknown isolate with the isolate of a known A1(98–111) and A2 (98-35a) mating type on carrot agar(CA) medium at 3 cm apart. After incubation at 28 °Cin darkness for 4 weeks, agar blocks were examinedmicroscopically. An isolate was designated to be
mating type A1 if oospores were present when pairedwith a known A2 tester and vice-versa. The soloculture of each isolate was examined for oosporeformation as a control. The positive control was across between two tester isolates of opposite matingtypes. The test was replicated two times.
Genomic DNA preparation
P. colocasiae isolates were grown in potato dextrosebroth medium (PDB; 250 g/lpotato, 20 g/l dextrose) at28 °C with 50 rpm. After 5–7 days mycelium washarvested and dried on sterile paper towels. DNAwasextracted using a Genomic DNA purification kit(Fermentas, EU) according to manufacturer’s instruc-tions. The nucleic acid obtained was dissolved in TEbuffer (100 μl; pH=8.0). The quality and integrity ofDNAwere assessed by agarose gel electrophoresis andstored at −20 °C until further use. The DNA concen-tration was adjusted to 20 ng/μl for amplification bythe polymerase chain reaction.
ITS characterization of isolates
Target DNAwas amplified using the universal primersITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) andITS4 (5′-TCCTCCGCTTATTGATATGC-3′) (Whiteet al. 1990). Each 25 μL of PCR reaction consistedof 50 ng of template DNA, 100 μM each deoxynu-cleotide triphosphate, 20 ng of each primer, 1.5 mMMgCl2, 1 x Taq buffer (10 mM Tris–HCl pH9.0,50 mM KCl, 0.01 % gelatin), 1 U of Taq DNApolymerase (Merck GeNei, India). Amplificationswere performed in an Agilent sure cycler 8800(Agilent Technologies, USA). The thermal cyclerwas programmed as follows: 2 min at 94 °C, 35 cyclesof 30 s at 94 °C, 1 min at 57.1 °C and 1 min 30 s at72 °C, and finally 8 min at 72 °C. Amplified productswere resolved on a 1.5 % agarose gel containing0.5 μgml−1 ethidium bromide and photograph wasscanned through the Gel Doc System (Alpha Imager,Alpha Innotech, USA). The PCR products wereextracted using a QIAquick Gel extraction kit(QIAGEN, Tokyo, Japan) and cloned into thepTZ57R/T vector (InsTAclone PCR cloning kit,Fermentas, EU). They were transformed into compe-tent DH5α cells and plated on Luria agar blue andwhite selective medium. The obtained white colonieswere randomly subjected to colony PCR in order to
Eur J Plant Pathol (2013) 136:483–494 485
confirm the presence of the insert and those foundpositive were sequenced using M13 forward and re-verse primers. Sequencing was carried out in the DNAfingerprinting wing of Rajiv Gandhi Centre forBiotechnology (RGCB), Thiruvananthapuram. Thenucleotide sequences obtained were processed to re-move vector sequence and low quality reads, trans-formed into consensus sequences with Geneious Prosoftware version 5.6. The resulted high qualitysequences were analyzed with BLASTn (NCBI) toconfirm the authenticity of isolates. The sequenceswere aligned using the computer package ClustalW(Thompson et al. 1994) and a phylogenetic study wascarried out with the programMEGA version 5 (Tamuraet al. 2011). Sequences were analyzed to determine therelationships between isolates by the neighbour-joining method (Saitou and Nei 1987) using theMaximum Composite Likelihood model. Bootstrapvalues were generated using 2000 replicates. Onlythose regions common to all the isolates were usedfor analysis (ITS1, 5.8S gene and ITS2).
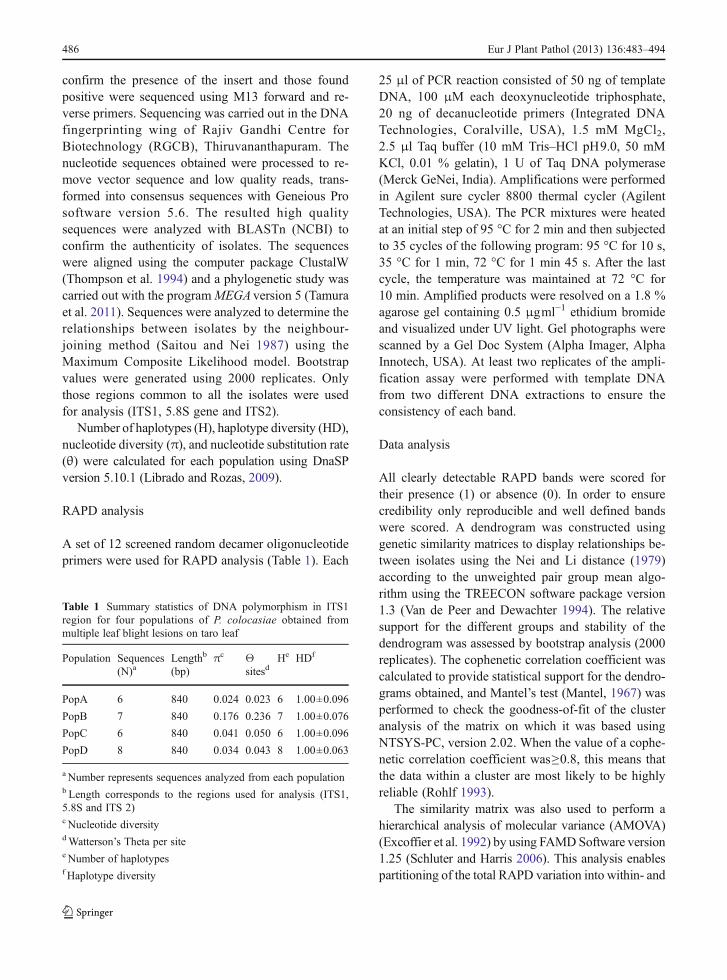
Number of haplotypes (H), haplotype diversity (HD),nucleotide diversity (π), and nucleotide substitution rate(θ) were calculated for each population using DnaSPversion 5.10.1 (Librado and Rozas, 2009).
RAPD analysis
A set of 12 screened random decamer oligonucleotideprimers were used for RAPD analysis (Table 1). Each
25 μl of PCR reaction consisted of 50 ng of templateDNA, 100 μM each deoxynucleotide triphosphate,20 ng of decanucleotide primers (Integrated DNATechnologies, Coralville, USA), 1.5 mM MgCl2,2.5 μl Taq buffer (10 mM Tris–HCl pH9.0, 50 mMKCl, 0.01 % gelatin), 1 U of Taq DNA polymerase(Merck GeNei, India). Amplifications were performedin Agilent sure cycler 8800 thermal cycler (AgilentTechnologies, USA). The PCR mixtures were heatedat an initial step of 95 °C for 2 min and then subjectedto 35 cycles of the following program: 95 °C for 10 s,35 °C for 1 min, 72 °C for 1 min 45 s. After the lastcycle, the temperature was maintained at 72 °C for10 min. Amplified products were resolved on a 1.8 %agarose gel containing 0.5 μgml−1 ethidium bromideand visualized under UV light. Gel photographs werescanned by a Gel Doc System (Alpha Imager, AlphaInnotech, USA). At least two replicates of the ampli-fication assay were performed with template DNAfrom two different DNA extractions to ensure theconsistency of each band.
Data analysis
All clearly detectable RAPD bands were scored fortheir presence (1) or absence (0). In order to ensurecredibility only reproducible and well defined bandswere scored. A dendrogram was constructed usinggenetic similarity matrices to display relationships be-tween isolates using the Nei and Li distance (1979)according to the unweighted pair group mean algo-rithm using the TREECON software package version1.3 (Van de Peer and Dewachter 1994). The relativesupport for the different groups and stability of thedendrogram was assessed by bootstrap analysis (2000replicates). The cophenetic correlation coefficient wascalculated to provide statistical support for the dendro-grams obtained, and Mantel’s test (Mantel, 1967) wasperformed to check the goodness-of-fit of the clusteranalysis of the matrix on which it was based usingNTSYS-PC, version 2.02. When the value of a cophe-netic correlation coefficient was≥0.8, this means thatthe data within a cluster are most likely to be highlyreliable (Rohlf 1993).
The similarity matrix was also used to perform ahierarchical analysis of molecular variance (AMOVA)(Excoffier et al. 1992) by using FAMDSoftware version1.25 (Schluter and Harris 2006). This analysis enablespartitioning of the total RAPD variation into within- and
Table 1 Summary statistics of DNA polymorphism in ITS1region for four populations of P. colocasiae obtained frommultiple leaf blight lesions on taro leaf
Population Sequences(N)a
Lengthb
(bp)πc Θ
sitesdHe HDf
PopA 6 840 0.024 0.023 6 1.00±0.096
PopB 7 840 0.176 0.236 7 1.00±0.076
PopC 6 840 0.041 0.050 6 1.00±0.096
PopD 8 840 0.034 0.043 8 1.00±0.063
a Number represents sequences analyzed from each populationb Length corresponds to the regions used for analysis (ITS1,5.8S and ITS 2)c Nucleotide diversitydWatterson’s Theta per sitee Number of haplotypesf Haplotype diversity
486 Eur J Plant Pathol (2013) 136:483–494
among-leaves, and provides a measure of inter-regiongenetic distances as the proportion of the total RAPDvariation residing between P. colocasiae of any twoleaves (called Phi statistics; Excoffier et al. 1992).
The percentage of polymorphic bands (PPB), theeffective number of alleles per locus (NE), the ob-served number of alleles per locus (NA), Shannon’sinformation index of diversity (I) (Lewontin, 1972),and Nei’s gene diversity (H) were calculated to esti-mate genetic variation. Population differentiation wasanalyzed for polymorphism between populations bythe gene differentiation coefficient (GST). The effec-tive number of migrants per generation, an indirectestimate of the gene flow between any pairwise pop-ulations, was estimated as: Nm=0.5 ((1-GST) /GST)(McDermott and McDonald, 1993). All calculationswere performed using POPGENE program Version1.32 (Yeh et al. 1997).
We evaluated the evidence for recombination byperforming linkage disequilibrium tests. The standard-ized index of association (rBarD) statistic (Agapowand Burt 2001) was used to estimate multilocus link-age disequilibrium (LD) in each population using thesoftware MULTILOCUS version 1.3 (Agapow andBurt 2001).
Results
Colony morphology
Isolates of P. colocasiae were analyzed for any mor-phological variation on PDA and CA plates. All theisolates tested had a similar growth pattern withinindividual media used. They depicted the white cot-tony appearance with concentric rings on the PDA andsparse uniform growth on CA plates. There was nodistinguishable morphological pattern for any isolate.This suggests that all the isolates shared a commongenetic base with respect to colony morphology.
Pathogenicity assay
All isolates were found to be virulent and reproducedtypical leaf blight symptoms. The isolates initiated le-sion development after 4 days of inoculation and theinoculated sites showed water soaked lesions at thebeginning which turned brown upon the progressionof the disease. There was no variation in lesion
development among the isolates upon visual inspection.This suggests that all the isolates were similarly aggres-sive and shared common pathogenicity characters.
Determination of mating type
All isolates of P. colocasiae tested were of A1 matingtype. Oospores were produced only when plates of eachisolate included the tester isolate of the A2 mating type.
ITS characterization of isolates
The ITS1, 5.8 S rRNA gene and ITS 2 region of a subsetof isolates was studied. All isolates were positivelyidentified as P. colocasiae by ITS amplification andsequencing. Amplification with ITS primers (ITS1 -ITS4) yielded ~850 bp product in all isolates studied.The ITS sequence analysis revealed 97–99% nucleotidesequence homology with each other and 95–99 % sim-ilarity among the isolates of P. colocasiae available inthe GenBank database (data not shown). Alignment ofsequences using Clustal W revealed considerable varia-tion in all the isolates examined. Variations ranged fromsingle base pair changes to multiple changes represent-ing deletions and insertions. More sequence variationwas evident in the ITS1 region with only a few shortregions showing complete homology across all isolatesexamined, whereas in ITS2 region the sequences weremore conserved, with many regions of complete homol-ogy interspersed with variable regions (SupplementaryFig. 2). The nucleotide polymorphism defined 12 ‘hap-lotypes’ and the representative sequences are depositedin Genbank under accession numbers JX134643-JX134654. The number of haplotypes, haplotype diver-sity and nucleotide diversity was relatively high in allthe sequences analyzed. The highest nucleotide diversi-ty was seen in population B while the remaining popu-lation shared similar values for various parametersanalyzed (Table 1). Phylogenetic tree clearly projectedthe variation among isolates of P. colocasiae. No isolatefrom the same leaf was grouped together indicating theyare genetically diverse (Fig. 1).
RAPD analysis
Distribution of amplification fragments was unique foreach primer. To ensure credibility in scoring, allmarkers were scored at least twice. All the bands weredistinct and reproducible. The 12 primer combinations
Eur J Plant Pathol (2013) 136:483–494 487
amplified 198 reproducible fragments ranging in sizefrom 200 to 1800 bp, of which 193 (97.9 %) werepolymorphic (Table 2). When fingerprints of theseisolates were compared, some bands common to themajority of isolates were observed, while others wereunique to one or few isolates. The highest number ofamplification products was obtained with the primerOPG19, while the lowest was with OPA 2; the averagenumber of bands among total 12 primers was 16.5.The number of polymorphic fragments detected by
each primer varied from 12 to 21, with an average of16. The highest number of polymorphic bands (21)was produced by the primer OPG 19, whereas theprimer OPA 2 generated the lowest number of poly-morphic bands (12).
Within population genetic diversity
Genetic diversity varied among populations withthe PPB values ranging from 54.04 % (Pop D)to 90.91 % (Pop A), with an average of 73.73 %.The average Nei’s gene diversity (H) was estimat-ed to be 0.265 within populations and 0.329 forthe pooled populations. The observed number ofalleles (NA) and the effective number of alleles(NE) and Nei’s gene diversity varied among pop-ulations. Among the four studied populations, pop-ulation D exhibited the lowest levels of diversity(PPB=54.04 %; H=0.2065; I=0.3052; NA=1.5404;NE=1.3555), whereas population A showed thehighest variability (PPB=90.91 %; H=0.3187; I=0.4743; NA=1.9091; NE=1.5565). The remainingpopulations of B and C exhibited diversity fromlow to high (Table 3).
Genetic structure of populations
Analysis of molecular variance (AMOVA) showedthat a high percentage of the total genetic diversity of
Fig. 1 ITS phylogeny of P. colocasiae isolates obtained frommultiple leaf blight lesions on the same taro leaf. Letters anddigits represent the population code and isolate number fromeach population. Numbers at node represents bootstrap values(2000 replicates)
Table 2 Attributes of the pri-mers used for RAPD amplifica-tion and number of bands perprimer
Primer Sequence 5′-3′ No. of bandsscored
No. ofpolymorphicbands
Mean no.of bands
Polymorphism (%)
OPT6 CAAGGGCAGA 15 15 4.3 100
OPT7 GGCAGGCTGT 20 20 6.5 100
OPT13 AGGACTGCCA 19 19 6.9 100
OPG10 AGGGCCGTCT 14 14 5.7 100
OPG 12 CAGCTCACGA 17 17 5.9 100
OPG 16 AGCGTCCTCC 16 16 4.3 100
OPG 17 ACGACCGACA 13 13 4.5 100
OPG 18 GGCTCATGTG 15 15 5.9 100
OPG19 GTCAGGGCAA 21 21 8.2 100
OPA2 TGCCGAGCTG 12 12 4.5 100
OPA10 GTGATCGCAG 16 16 6.9 100
OPA15 TTCCGAACCC 20 15 5.4 75
Total 198 193 69 97.9
Average 16.5 16.0 5.75 97.9
488 Eur J Plant Pathol (2013) 136:483–494
the four P. colocasiae populations in this study weredistributed on a small spatial scale with 63.54 % of thegenetic diversity distributed within populations andless yet significantly 36.45 % among populations(Table 4). The coefficient of genetic differentiationamong populations (GST) was 0.2007, which supportsthe AMOVA analysis indicating only limited geneticdiversity among populations and high diversity withinpopulations. The estimate of gene flow (Nm) amongpopulations was 1.991 migrants per generation,obtained from the GST value.
Cluster analysis
Nei’s genetic distances (D) between P. colocasiae iso-lates from four leaves ranged from 0.0866 to 0.9171,with an average of 0.5018. Similarly, the genetic identity(IN) between P. colocasiae isolates from leaf samplesvaried from 0.0699 to 0.9325. A UPGMA cluster anal-ysis of 41 individuals indicated that the samples from thesame population did not form a distinct group (Fig. 2).
P. colocasiae populations had comparatively lowrBarD values (0.1411, 0.0876, 0.0539 and 0.0837respectively) (Table 3) which suggests that recom-bination events are common in these populations.
The cophenetic correlation coefficient between thedendrogram and the original distance matrix of theRAPD profiles was significant, with a high correlationvalue (r=0.88, P=1).
Discussion
The present study was conducted to assess the diversityof P. colocasiae isolates obtained from a fine spatialscale such as multiple leaf blight lesions on a taro leaf.This study represents the first of its kind and providesnovel insights into P. colocasiae genetic diversity. Theresults of the study provide the first indication that thepopulations of P. colocasiae are highly heterogeneousand a substantial level of genotypic variability existsamong individual isolates of populations by providing
Table 3 Population genetic parameters for four populations of P.colocasiae based on RAPD data
Population code Polymorphic bands PPBa(%) NAb NE
c Hd Ie rBarDf
A 180 90.91 1.9091±0.2882 1.5565±0.3561 0.3187±0.1728 0.4743±0.2267 0.1411*
B 127 64.14 1.6414±0.4808 1.4393±0.4101 0.2459±0.2116 0.3605±0.2968 0.0876*
C 170 85.86 1.8586±0.3493 1.4815±0.3285 0.2894±0.1646 0.4385±0.2249 0.0539*
D 107 54.04 1.5404±0.4996 1.3555±0.3762 0.2065±0.2046 0.3052±0.2946 0.0837*
Average 146 73.73 1.7373±0.4044 1.4582±0.3677 0.2651±0.1884 0.3946±0.2607
Total 193 97.9 2.0000 1.5328±0.2490 0.3295±0.1139 0.5029±0.1388
a Percentage of polymorphic bands (PPB)b Observed number of alleles (NA)c Effective number of alleles (NE)d Nei’s gene diversity (H)e Shannon’s information index (I)f The standardized index of association (rBarD) statistic was estimated for each population (Agapow and Burt 2001). The significanceof rBarD was tested with 100 randomizations of the data by comparing the observed value to that expected under the null hypothesis ofrBarD=0. The null hypothesis of multilocus linkage equilibrium was rejected if P<0.010. * Significant at P<0.010
Table 4 AMOVA analysis of 41isolates of P. colocasiae fromfour populations
df degrees of freedom; SSD sumsof squared deviations
distinct banding patterns unique to isolate regardless ofthe phenotypic variation.
To ensure credibility to our study we started offwith basic cultural characterization i.e. by studyingthe morphology, pathogenicity and mating type ofthe isolates. However, no variation in morphology,pathogenicity and mating type was observed whenisolates were subjected to growth on PDA and CAplates, pathogenicity on detached taro leaf and matingtype determination on carrot agar. This suggests thatthey share a common genetic base with respect to thenormal growth and development. Uniformity in phe-notypic characters of P. colocasiae isolates obtainedfrom the same geographical region has been alreadyreported in old populations (Misra et al. 2011).
This is the first time that the sequence analysis ofthe ITS region with a population of P. colocasiae fromfine spatial scales has been conducted. Alignment ofthe ITS sequences revealed considerable variation in
the ITS1 region of all isolates studied. A similar ob-servation was reported by Cooke and Duncan (1997),who also observed more polymorphism in the ITS1region of Phytophthora species in a phylogeneticstudy using ITS1 and ITS 2 regions. The presence ofhigh levels of DNA polymorphism in ITS regionsprovides the indication that these pathogens arecontinuously evolving in nature. High haplotypicdiversities may be a consequence of heterozygousindividuals showing alleles with low frequencies(Cardenas et al. 2011). Mechanisms such as trans-locations, chromosome deletions and duplicationsare common in Phytophthora species (Goodwin1997) and this may be the case with P. colocasiaealso. However, a large area has not been sampledand sequencing of more number of isolates repre-senting diverse geographical regions is desirable toconfirm the overall population structure of P. colo-casiae with respect to ITS region.
Fig. 2 Dendrogram (UPGMA) of 41 isolates of P. colocasiae.The letter and digit represent the population code and isolatenumber from each population. The cophenetic correlation
coefficient between the dendrogram and the original distancematrix of the RAPD profiles was significant, with a high correla-tion value (r=0.88, P=1)
490 Eur J Plant Pathol (2013) 136:483–494
Molecular markers such as RAPD are usually used toassess total genetic variation that is not directly affectedby hosts, or environmental factors that influence theexpression of phenotypes (Cooke et al. 1996). RAPDhas been successfully used to estimate genetic diversityof several Phytophthora species (Chang et al. 1996;Meng et al. 1999; Mahuku et al. 2000; Wang et al.2003). The limitations of this technique are related todominance (Lynch and Milligan, 1994) and reproduc-ibility (Caetano-Anollés 1993). RAPD patterns are verysensitive to slight changes in amplification conditionsgiving problems of reproducibility and necessity forextensive standardization to obtain reproducible results(Penner et al. 1993). This study used the followingprocedures to overcome these limitations. The templateDNA concentration was kept uniform for all samplesand adequate replicates of the amplification assay wereperformed with template DNA from multiple DNAextractions. The DNA polymerase (Taq polymerase),buffer, and dNTPs used were from the same source.The present study shows that under identical amplifica-tion conditions, RAPD profiles for any particularprimer-template DNA concentration is highly reproduc-ible over a wide range of RAPD templates. Our resultsdemonstrated the utility of RAPD markers to assessgenetic diversity among isolates of P. colocasiae. Thehigh proportion of polymorphic loci found in the iso-lates reveals profound variability. High levels of geno-typic diversity such as those found in the present studyhave previously been described in P. colocasiae popu-lations from Southeast Asia and Pacific region (Lebotet al. 2003) as well as from India (Mishra et al. 2010;Nath et al. 2012). No correlation was observed betweenphenotypic characters and banding patterns defined byRAPD. This may be due to the preferential amplificationof a non- coding repetitive genome that is not directlylinked to pathogenic characteristics (Cooke et al. 1996).Genetic changes may occur in the pathogen duringasexual reproduction and these changes may or maynot affect the phenotype of the pathogen (Manzanares-Dauleux et al. 2001; Abu-El Samen et al. 2003). Theresults of the present study support the conclusion thatisolates of P. colocasiae have undergone some geneticchange that did not affect their phenotypic characters.
This study also demonstrated a high level of inter-population gene flow. The coefficient of populationdifferentiation was less but significant (GST=0.20)indicating that P. colocasiae isolates from differentpopulations are genetically diverse. The results of the
UPGMA cluster analysis of all samples revealed thatindividuals from the same population occasionallyfailed to cluster in one distinct group. This may bedue to the relatively high intrapopulation variancecompared to the interpopulation one as projected byAMOVA analysis.
The presence of low index of association (rBarD)values provides the first indication that variation seenin populations of P. colocasiae probably reflects theoutcome of mitotic recombination or mutations ratherthan sexual recombination; although a much largersample collection is needed for a definitive conclu-sion. Mitotic gene conversion was observed to occurat remarkably high frequencies in Phytophthora sojaedocumenting the potential for rapid generation of var-iation (Chamnanpunt et al. 2001). Lack of occurrenceof compatible mating type of P. colocasiae at a finespatial scale further supports this viewpoint (Lin andKo, 2008; Misra et al. 2011). However, evidence ofrecombination as inferred above must be further elu-cidated by the use of other markers. In general, muta-tion is the primary source of variation in oomycetes(Goodwin 1997). Rapid rates of mutation events havebeen already discovered in P. sojae (Drenth et al.1996) and occurrence of clonal lineages have beenidentified in P. infestans (Goodwin 1997). P. coloca-siae is known to rapidly reproduce asexually throughthe formation of large numbers of sporangia,which either germinate directly or differentiate intomotile zoospores under favourable condition (e.g.,intermittent rainy weather). These sporangia getdispersed in rain splashes to neighbouring hostsover short distances and thus possibly provide ameans for outsourcing events. Outsourcing canoccur when two strains are mixed in culture(Drenth et al. 1995, 1996) or infect the same plant(Fry and Goodwin 1995). Based on these observa-tions, it can be suggested that high levels of var-iability are most likely due to random mutation,recombination, exposure of the pathogen in diverseclimates, hosts, cropping patterns and movement ofthe pathogen within the country.
Many Phytophthora species have evolved in limit-ed geographic areas, and this is particularly true forthose species that are host specific (Goodwin 1997).Given the fact that P. colocasiae is host specific, thisobservation stands true for this species. Considerablegenetic diversity is reported in taro accessions acrossglobe (Lebot and Aradhya 1991; Lebot et al. 2000;
Eur J Plant Pathol (2013) 136:483–494 491
Mishra et al. 2010). This vast amount of host diversityimposes tremendous selection pressure on P. coloca-siae and may possibly be one of the reasons for theexistence/formation of new strains of the pathogen. Itis possible that different pathogen genotypes may haveevolved to selectively parasitize individual host range.High levels of diversity in a fungal population enhanceits ability to adapt to diverse conditions and overcomehost resistance (Chamnanpunt et al. 2001). In manycultivars, there is a differential degree of responseagainst the taro leaf blight disease caused by P. colo-casiae. This is mainly due to the genetic makeup oftaro which may favour the growth and spread of thepathogen or may resist and eliminate the spread of thepathogen. With a huge number of propagules andstrong selection imposed by host resistance genesand fungicides, a moderate mutation rate would besufficient to generate all of the observed variation.The knowledge about the genetic diversity of thepathogen has great potential in mitigating the leafblight of taro.
In conclusion, the current study raises numerousimplications on the present management strategiesadvocated for taro leaf blight. The degree of globalvariability present in populations of P. colocasiae(Lebot et al. 2003), and high level of genetic diversityobserved at a very small scale (multiple leaf blightlesions on a single taro leaf), support the assumptionthat mutation and recombination events play a majorrole in the life cycle of the pathogen. The finding thatextensive genetic differences occur in P. colocasiaepopulations should be an important consideration inchoosing breeding tactics to develop durable leafblight resistance. The high level of genetic diversityshows that the P. colocasiae populations could re-spond rapidly to selection exerted by newly introducedhost resistance genes or fungicides, underlining theimportance of relying on integrated disease manage-ment. Disease management programmes should focuson a local scale than on a regional level since it islikely that effective packages at one particular locationmay not prove so in other regions. Molecular markersthat are genotype-specific can be useful for studyinghow hosts, cultural practices and environment affectthe genetic structure of P. colocasiae populations. Weare currently working towards identifying regions atthe molecular level that are subjected to frequent mu-tation events in an attempt to explore the biology of P.colocasiae.
Acknowledgements The funding provided for conducting theresearch work by the Indian Council of Agricultural Research,New Delhi, is gratefully acknowledged. The authors are gratefulto Mr. U. Suresh Kumar, DNA examiner, DNA fingerprintingwing, Rajiv Gandhi Centre for Biotechnology, Thiruvanantha-puram for his assistance in DNA sequencing and analysis. Theauthors express their deep gratitude to the anonymous reviewersfor their valuable comments and helpful suggestions throughoutthe making of this manuscript.
References
Abu-El Samen, F. M., Secor, G. A., & Gudmestad, N. C. (2003).Genetic variation among asexual progeny of Phytophthorainfestans detected with RAPD and AFLP markers. PlantPathology, 52, 314–325.
Agapow, P. M., & Burt, A. (2001). Indices of multilocus linkagedisequilibrium. Molecular Ecology Notes, 1, 101–102.
Caetano-Anollés, G. (1993). Amplifying DNA with arbitraryoligonucleotide primers. Genome Research, 3, 85–94.
Cardenas, M., Grajales, A., Sierra, R., Rojas, A., Almario, A.G., Vargas, A., et al. (2011). Genetic diversity ofPhytophthora infestans in the Northern Andean region.BMC Genetics, 12, 23.
Chang, T. T., Yang, W. W., &Wang, W. Y. (1996). Use of randomamplified polymorphic DNA markers for the detection ofgenetic variation in Phytophthora cinnamomi in Taiwan.Botanical Bulletin of Academia Sinica, 37, 165–171.
Chamnanpunt, J., Shan, W. X., & Tyler, B. M. (2001). Highfrequency mitotic gene conversion in genetic hybrids of theOomycete Phytophthora sojae. Proceedings of NationalAcademy of Science USA, 98, 14530–14535.
Cooke, D. E. L., Kennedy, D. M., Guy, D. C., Russel, J.,Unkles, S. E., & Duncan, J. M. (1996). Relatedness ofgroupI species of Phytophthora as assessed by randomlyamplified polymorphic DNA (RAPD) and sequences ofribosomal DNA. Mycological Research, 100, 297–303.
Cooke, D. E. L., & Duncan, J. M. (1997). Phylogenetic analysisof Phytophthora species based on ITS1 and ITS2 sequen-ces of the ribosomal RNA gene repeat. MycologicalResearch, 101, 667–677.
Drenth, A., Janssen, E. M., & Govers, F. (1995). Formation andsurvival of oospores of Phytophthora infestans under nat-ural conditions. Plant Pathology, 44, 86–94.
Drenth, A., Whisson, S. C., Maclean, D. J., Irwin, J. A. G., Obst, N.R., & Ryley, M. J. (1996). The evolution of races ofPhytophthora sojae in Australia.Phytopathology, 86, 163–169.
Excoffier, L., Smouse, P. E., & Quattro, J. M. (1992). Analysisof molecular variance inferred from metric distancesamong DNA haplotypes: application to human mitochon-drial DNA restriction data. Genetics, 131, 479–491.
Fry, W. E., & Goodwin, S. B. (1995). Recent migrations ofPhytophthora infestans. In L. J. Dowley, E. Bannon, L.R. Cooke, T. Keane, & E. O’Sullivan (Eds.), Phytophthorainfestans 150 (pp. 89–95). Dublin: Boole Press.
Goodwin, S. B. (1997). The population genetics ofPhytophthora. Phytopathology, 97, 462–473.
Jackson, G. V. H., Gollifer, D. E., & Newhook, F. J. (1980).Studies on the taro leaf blight fungus Phytophthora
492 Eur J Plant Pathol (2013) 136:483–494
colocasiae in the Solomon Islands: control by fungicidesand spacing. Annals of Applied Biology, 96, 1–10.
Koh, Y. J., Goodwin, S. B., Dyer, A. T., Cohen, B. A.,Ogoshi, A., Sato, N., et al. (1994). Migrations anddisplacements of Phytophthora infestans in EastAsian countries. Phytopathology, 84, 922–927.
Lakhanpaul, S., Velayudhan, K. C., & Bhat, K. V. (2003).Analysis of genetic diversity in Indian C. esculenta(Colocasia esculenta (L.)Schott) using random amplifiedpolymorphicDNA (RAPD) markers. Genetic Resourcesand Crop Evolution, 50, 603–609.
Lebot, V., & Aradhya, K. M. (1991). Isozyme variation in taro(Colocasia esculenta (L.)Schott) from Asia and Oceania.Euphytica, 56, 55–66.
Lebot, V., Hartati, S., Hue, N. T., Viet, N. V., Nghia, N. H., &Okpul, T. (2000). Genetic variation in taro (Colocasiaesculenta) in SouthEast Asia and Oceania. TwelfthSymposium of the ISTRC. Potential of root crops for foodand industrial resources. Sept.10–16, 2000. Tsukuba,Japan, pp. 524–533.
Lebot, V., Herail, C., Gunua, T., Pardales, J., Prana, M.,Thongjiem, M., et al. (2003). Isozyme and RAPD variationamong Phytophthora colocasiae isolates from South EastAsia and the Pacific. Plant Pathology, 52, 303–313.
Lewontin, R. C. (1972). The apportionment of human diversity.Evolutionary Biology, 6, 381–398.
Librado, P., & Rozas, J. (2009). Dnaspv5: A software for com-prehensive analysis of DNA polymorphism data.Bioinformatics, 25, 1451–1452.
Lin, J. M., & Ko, H. W. (2008). Occurrence of isolates ofPhytophthora colocasiae in Taiwan with homothallicbehavior and its significance. Mycologia, 100(5),727–734.
Lynch, M., & Milligan, B. G. (1994). Analysis of populationgenetic structure with RAPD markers. Molecular Ecology,3, 91–99.
McDermott, J. M., & McDonald, B. A. (1993). Gene flow inplant pathosystems. Annual Review of Phytopathology, 31,353–373.
Mahuku, G., Peters, R. D., Platt, H. W., & Daay, F. (2000).Random amplified polymorphic DNA analysis ofPhytophthora infestans isolates collected in Canada during1994–96. Plant Pathology, 49, 252–260.
Mantel, N. (1967). The detection of disease clustering andgeneralized regression approach. Cancer Research, 27,209–220.
Manzanares-Dauleux, M. J., Divaret, I., Baron, F., & Thomas, G.(2001). Assessment of biological and molecular variabilitybetween and within field isolates of Plasmodiophora brassi-cae. Plant Pathology, 50, 165–173.
Meng, X. Q., Soemaker, R. C., & Yang, X. B. (1999). Analysisof pathogenic i ty and genet ic var ia t ion amongPhytophthora sojae isolates using RAPD. MycologicalResearch, 103, 173–179.
Mishra, A. K., Sharma, K., & Misra, R. S. (2010). Isozymeand PCR-based genotyping of epidemic Phytophthoracolocasiae associated with taro leaf blight. Archives ofPhytopathology and Plant Protection, 43(14), 1367–1380.
Misra, R. S., Sharma, K., & Mishra, A. K. (2008). Phytophthoraleaf blight of Taro (Colocasia esculenta)—a review. The
Asian and Australasian Journal of Plant Science andBiotechnology, 2, 55–63.
Misra, R. S., Mishra, A. K., Sharma, K., Jeeva, M. L., & Hegde,V. (2011). Characterisation of Phytophthora colocasiaeisolates associated with leaf blight of taro in India.Archives of Phytopathology and Plant Protection, 44(6),581–591.
Misra, R. S., & Chowdhury, S. R. (1997). Phytophthora leafblight disease in taro. Technical Bulletin Series 21,C.T.C.R.I. (ICAR), Trivandrum.
Nath, V. S., Sankar, M. S., Hegde, V. M., Jeeva, M. L., Misra, R.S., Veena, S. S., et al. (2012). Analysis of genetic diversityin Phytophthora colocasiae using RAPD markers. TheAsian and Australasian Journal of Plant Science andBiotechnology, 6(1), 38–43.
Nei, M., & Li, W. H. (1979). Mathematical modelfor studyinggenetic variation in terms of restriction endonucleases.Proceedings of National Academy of Science, USA, 76,5269–5273.
Penner, G. A., Bush, A., Wise, R., Kim, W., Domier, L., Kasha,K., et al. (1993). Reproducibility of random amplifiedpolymorphic DNA (RAPD) analysis among laboratories.Genome Research, 12, 341–345.
Raciborski, M. (1900). Parasitische Algen und Pilze, Java’s(Java’s Parasitic Algae and Fungi). I. Batavia. (Cited inWaterhouse1970a under P. colocasiae).
Rohlf, F. J. (1993). Contributions to morphometrics:Relative warp analysis and an example of its applica-tion to mosquito wings. In Marcus et al. (Eds.),Museo Nacional de Ciencias Naturales (pp. 131–159). Madrid
Saitou, N., & Nei, M. (1987). The neighbor-joining method: Anew method for reconstructing phylogenetic trees.Molecular Biology and Evolution, 4, 406–425.
Schluter, P. M., & Harris, S. A. (2006). Analysis of multilocusfingerprinting data sets containing missing data. MolecularEcology Notes, 6, 569–572.
Sharma, K., Mishra, A. K., & Misra, R. S. (2008). The geneticstructure of C. esculenta: a comparison of RAPD andisozyme markers. Plant Biotechnology Reports, 2, 191–198.
Sullivan, M. J., Parks, E. J., Cubeta, M. A., Gallup, C. A.,Melton, T. A., Moyer, J. W., et al. (2010). An assessmentof the genetic diversity in a field population ofPhytophthora nicotianae with a changing race structure.Plant Disease, 94(4), 455–460.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei,M., & Kumar, S. (2011). MEGA5: molecular evolu-tionary genetics analysis using maximum likelihood,evolutionary distance, and maximum parsimony meth-ods. Molecular Biology and Evolution, 28(10), 2731–2739.
Thankappan, M. (1985). Leaf blight of taro-a review. Journal ofRoot Crops, 11, 223–236.
Thompson, J. D., Higgins, D. G., & Gibson, T. J. (1994).Clustal-W—Improving the sensitivity of progressive mul-tiple sequence alignment through sequence weighting,position-specific gap penalties and weight matrix choice.Nucleic Acids Research, 22, 4673–4680.
Van de Peer, Y., & Dewachter, R. (1994). Treecon for Windows-A software package for the construction and drawing of
Eur J Plant Pathol (2013) 136:483–494 493
evolutionary trees for the Microsoft Windows environ-ment. Computer Applications in the Biosciences, 10,569–570.
Wang, H. B., Wang, X. M., & Zhu, Z. D. (2003). Analysis ofgenetic diversity of Phytophthora sojae isolates in Chinausing RAPD. Mycosystema, 22(2), 219–227.
White, T. J., Bruns, T., Lee, S., & Taylor, J. (1990).Amplification and direct sequencing of fungal ribosomal
RNA genes for phylogenetics. In M. A. Innes, D. H.Gefland, J. Sninsky, & T. J. White (Eds.), PCR protocols:A guide to methods and applications (pp. 315–322).SanDiego: Academic.
Yeh, F. C., Boyle, T., Yang, R., Ye, Z., & Xiyan, J. M. (1997).Microsoft window-based freeware for population geneticanalysis (POPGENE version 1.31). Edmonton: Universityof Alberta and Centre for International Forestry Research.