Accepted Manuscript Molecular phylogenetic reconstructions identify East Asia as the cradle for the evolution of the cosmopolitan genus Myotis (Mammalia, Chiroptera) Manuel Ruedi, Benoît Stadelmann, Yann Gager, Emmanuel J.P. Douzery, Charles M. Francis, Liang-Kong Lin, Antonio Guillén-Servent, Alice Cibois PII: S1055-7903(13)00333-3 DOI: http://dx.doi.org/10.1016/j.ympev.2013.08.011 Reference: YMPEV 4692 To appear in: Molecular Phylogenetics and Evolution Please cite this article as: Ruedi, M., Stadelmann, B., Gager, Y., Douzery, E.J.P., Francis, C.M., Lin, L-K., Guillén-Servent, A., Cibois, A., Molecular phylogenetic reconstructions identify East Asia as the cradle for the evolution of the cosmopolitan genus Myotis (Mammalia, Chiroptera), Molecular Phylogenetics and Evolution (2013), doi: http://dx.doi.org/10.1016/j.ympev.2013.08.011 This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Transcript
Accepted Manuscript
Molecular phylogenetic reconstructions identify East Asia as the cradle for the
evolution of the cosmopolitan genus Myotis (Mammalia, Chiroptera)
Manuel Ruedi, Benoît Stadelmann, Yann Gager, Emmanuel J.P. Douzery,
Charles M. Francis, Liang-Kong Lin, Antonio Guillén-Servent, Alice Cibois
3) identify eastern Asia as the cradle of Myotis evolution. There is some uncertainty whether 647
the Oriental region alone or in combination with the Eastern Palaearctic (as shown on Fig. 3) 648
is the origin of early Myotinae, but alternative bioregions are clearly less likely. In addition, 649
13 of the 21 intercontinental range expansions originated from the Oriental region (inset of 650
Fig. 3) suggesting that this area is a center of origin for the radiation of Myotis species. The 651
deepest lineage identified in molecular reconstructions, the latirostris lineage, that is sister to 652
all remaining modern Myotis (Fig. 1 and Lack et al., 2010), is also endemic to this region, 653
further supporting an ancient, East Asian origin for this radiation. Finally, the Myotis species 654
diversity in the Oriental region (inset of Fig. 1) is already the highest in the world based on 655
currently recognized taxa, and is likely strongly underestimated (Francis et al., 2010), again 656
pointing to this area as a cradle of evolution for the genus. 657
658
4.6 Conclusions 659
660
Reconstruction of the biogeographic origin of Myotis (Fig. 3) suggests that the East 661
Asian region is the center of origin for the radiation of modern Myotis and is thus probably 662
also the cradle for the entire Myotinae. The Indo-Malayan, Indo-Chinese, Sino-Himalayan, 663
21
and East Asiatic floras and faunas (Corbet and Hill, 1992) meet in this region and engendered 664
the highest biodiversity hotspots on earth (Myers et al., 2000). This geographical area, which 665
has been subjected to gradual uplifting of the Tibetan plateau (the trans-Himalayan range) 666
during the last several million years, includes several high mountain ranges exhibiting sharp 667
topographical complexity, and shows climates ranging from tropical to arctic. Such high 668
environmental and topographic complexity could have promoted speciation. 669
Considering the potential vagility of bats, and their current worldwide distribution, the 670
overall number of transcontinental migrations in the Myotis radiation is relatively low (Fig. 3). 671
This illustrates the relative inability of Myotis species to cross some physical barriers. In turn, 672
Myotis species can occupy a variety of ecological niches, and it is not exceptional to find up 673
to 12 species coexisting in sympatry. This and the high ecological and morphological 674
diversity found within each Myotis assemblage at a worldwide scale support the central 675
impact of biogeography on the Myotis evolutionary history. Climatic and vegetation changes 676
since the Middle Miocene might also have favored speciation during the evolution of Myotis. 677
Recent molecular surveys of Myotis taxa from different parts of the world identified 678
that several species currently diagnosed by morphological characters certainly contain species 679
complexes: M. muricola, M. siligorensis, M. horsfieldii (Francis et al., 2010), M. montivagus 680
(Görföl et al., 2013; Volleth and Heller, 2012), M. nigricans (Larsen et al., 2012a), and even 681
European species such as M. nattereri (Salicini et al., 2011). Many more cryptic species are 682
present in the Oriental region, as suggested by the presence of numerous unnamed taxa in our 683
reconstructions (Table S1). Here, phylogenetic tools based on the comparison of polymorphic 684
mitochondrial and nuclear molecular markers will be of central interest to identify cryptic 685
species, refine the Myotis systematics, and help choosing among priorities for biological 686
conservation purposes. Unfortunately, this region of high biodiversity is also facing high rates 687
of habitat destruction, and is in urgent need of protection to maintain this evolutionary 688
diversity (Sodhi et al., 2004). 689
690
Acknowledgements 691
We are deeply indebted to those individuals and institutions who generously donated 692
tissues for genetic analyses. These samples provide the foundation of such broad taxonomic 693
surveys and would not be possible without their help. These individuals are: P. Benda 694
(Natural History Museum, Prague), L. R. Heaney and W. Stanley (Field Museum of Natural 695
History, Chicago, USA), G. Jones (University of Bristol, UK), K. Kawai (Institute of Low 696
Temperature Science, Hokkaido University, Sapporo, Japan), T. Kingston (Boston 697
University, Boston, USA), S. Kruskop and A. V. Borisenko (Zoological Museum of Moscow 698
22
State University), T. H. Kunz (Boston University, Boston, USA), J. Ma (Beijing, China), V. 699
Matveev (Moscow Lomonosov State University, Russia), late O. von Helversen, and F. 700
Mayer (University of Erlangen, Germany). This research was supported by grants from the 701
Swiss National Funds for Scientific research to M. Ruedi and A. Cibois (3100A0-105588), 702
and from Bat Conservation International to B. Stadelmann. This publication is contribution No 703
2013-XXX of the Institut des Sciences de l‟Evolution de Montpellier (UMR 5554 – CNRS - 704
IRD). 705
706
References 707
Abe, H., Ishii, N., Itoo, T., Kaneko, Y., Maeda, K., Miura, S., Yoneda, M., 2005. A guide to 708 the mammals of Japan. Tokai University Press, Kanagawa, Japan. 709 Alfaro, M.E., Zoller, S., Lutzoni, F., 2003. Bayes or bootstrap? A simulation study comparing 710 the performance of Bayesian Markov Chain Monte Carlo sampling and bootstrapping in 711 assessing phylogenetic confidence. Mol. Biol. Evol. 20, 255-266. 712 Bates, P.J.J., Hendrichsen, D.K., Walston, J.L., Hayes, B., 1999. A review of the mouse-eared 713 bats (Chiroptera: Vespertilionidae: Myotis) from Vietnam with significant new records. Acta 714 Chiropterol. 1, 47-74. 715 Bickham, J.W., Patton, J.C., Schlitter, D.A., Rautenbach, I.L., Honeycutt, R.L., 2004. 716 Molecular phylogenetics, karyotypic diversity, and partition of the genus Myotis (Chiroptera: 717 Vespertilionidae). Mol. Phylogenet. Evol. 33, 333-338. 718 Bininda-Emonds, O.R.P., Vazquez, D.P., Manne, L.L., 2000. The calculus of biodiversity: 719 integrating phylogeny and conservation. Trends Ecol. Evol. 15, 92-94. 720 Borisenko, A.V., Kruskop, S.V., Ivanova, N.V., 2008. A new mouse-eared bat (Mammalia: 721 Chiroptera: Vespertilionidae) from Vietnam. Russian J. Theriol. 7, 57-69. 722 Butler, P.M., 1984. Macrocelidea, Insectivora and Chiroptera from the Miocene of East 723 Africa. Paleovertebrata 14, 117-200. 724 Castella, V., Ruedi, M., Excoffier, L., 2001. Contrasted patterns of mitochondrial and nuclear 725 structure among nursery colonies of the bat Myotis myotis. J. Evolution. Biol. 14, 708-720. 726 Castella, V., Ruedi, M., Excoffier, L., Ibáñez, C., Arlettaz, R., Hausser, J., 2000. Is the 727 Gibraltar Strait a barrier to gene flow for the bat Myotis myotis (Chiroptera: 728 Vespertilionidae)? Mol. Ecol. 9, 1761-1772. 729 Cooper, S.J.B., Day, P.R., Reardon, T.B., Schulz, M., 2001. Assessment of species 730 boundaries in Australian Myotis (Chiroptera : Vespertilionidae) using mitochondrial DNA. J. 731 Mammal. 82, 328-338. 732 Corbet, G.B., Hill, J.E., 1992. The mammals of the Indomalayan region: a systematic review. 733 Oxford University Press, Oxford, UK. 734 Datzmann, T., Dolch, D., Batsaikhan, N., Kiefer, A., Helbig-Bonitz, M., phel, U., Stubbe, M., 735 Mayer, F., 2012. Cryptic diversity in Mongolian vespertilionid bats (Vespertilionidae, 736 Chiroptera, Mammalia). Results of the Mongolian-German biological expeditions since 1962, 737 No. 299. Acta Chiropterol. 14, 243-264. 738 Davalos, L.M., Cirranello, A.L., Geisler, J.H., Simmons, N.B., 2012. Understanding 739 phylogenetic incongruence: lessons from phyllostomid bats. Biol. Rev. 87, 991-1024. 740 Douady, C., Delsuc, F., Boucher, Y., Doolittle, W., Douzery, E., 2003. Comparison of 741 Bayesian and maximum likelihood bootstrap measures of phylogenetic reliability. Mol. Biol. 742 Evol. 20, 248-254. 743 Drummond, A.J., Ho, S.Y.W., Phillips, M.J., Rambaut, A., 2006. Relaxed phylogenetics and 744 dating with confidence. Plos Biol. 4, 699-710. 745
23
Drummond, A.J., Rambaut, A., 2007. BEAST: Bayesian evolutionary analysis by sampling 746 trees. Bmc Evolutionary Biology 7, 214. 747 Evin, A., Baylac, M., Ruedi, M., Mucedda, M., Pons, J.M., 2008. Taxonomy, skull diversity 748 and evolution in a species complex of Myotis (Chirotpera: Vespertilionidae): a geometric 749 morphometric appraisal. Biol. J. Linn. Soc. 95, 529-538. 750 Fenton, M.B., Bogdanowicz, W., 2002. Relationships between external morphology and 751 foraging behaviour: bats in the genus Myotis. Can J. Zool. 80, 1004-1013. 752 Findley, J.S., 1972. Phenetic relationships among bats of the genus Myotis. Syt. Zool. 21, 31-753 52. 754 Francis, C., Borisenko, A., Ivanova, N., Eger, J., Lim, B., Guillen-Servent, A., Kruskop, S., 755 Mackie, I., Hebert, P., 2010. The role of DNA barcodes in understanding and conservation of 756 mammal diversity in Southeast Asia. Plos One 5, e12575. 757 García-Mudarra, J.L., Ibañez, C., Juste, J., 2009. The Straits of Gibraltar: barrier or bridge to 758 Ibero-Moroccan bat diversity? Biol. J. Linn. Soc. 96, 434-450. 759 Görföl, T., Estók, P., Csorba, G., 2013. The subspecies of Myotis montivagus - taxonomic 760 revision and species limits (Mammalia: Chiroptera: Vespertilionidae). Acta Zool. Hung. 59, 761 41-59. 762 Gunnell, G.F., Eiting, T.P., Simons, E.L., 2012. African Vespertilionoidea (Chiroptera) and 763 the antiquity of Myotinae. In: Gunnell, G.F., Simmons, N.B. (Eds.), Evolutionary history of 764 bats. Fossils, molecules and morphology. Cambridge University Press, New York, pp. 252-765 266. 766 Gunnell, G.F., Simmons, N.B., 2005. Fossil evidence and the origin of bats. J. Mamm. Evol. 767 12, 209-246. 768 Han, N.-J., Zhang, J.-S., Reardon, T., Lin, L.-K., Zhang, J.-P., Zhang, S.-Y., 2010. 769 Revalidation of Myotis taiwanensis Ärnbäck-Christie-Linde 1908 and its molecular 770 relationship with M. adversus (Horsfield 1824) (Vespertilionidae, Chiroptera). Acta 771 Chiropterol. 12, 449-456. 772 Happold, M., Happold, D.C.D., 2013. The Mammals of Africa: Hedgehogs, Shrews and Bats. 773 Bloomsbury Publishing, London, UK. 774 Helversen, v.O., Heller, K.-G., Mayer, F., Nemeth, A., Volleth, M., Gombkötö, P., 2001. 775 Cryptic mammalian species: a new species of whiskered bat (Myotis alcathoe n. sp.) in 776 Europe. Naturwissenschaften 88, 217-223. 777 Hill, J.E., Topál, G., 1973. The affinities of Pipistrellus ridleyi Thomas 1898 and Glischropus 778 rosseti Oey, 1951 (Chiroptera: Vespertilionidae). Bull.br.Mus.nat.Hist.Zool. 24, 447-454. 779 Hoofer, S.R., Reeder, S.A., Hansen, E.W., Van den Bussche, R.A., 2003. Molecular 780 phylogenetics and taxonomic review of noctilionoid and vespertilionoid bats (Chiroptera: 781 Yangochiroptera). J. Mammal. 84, 809-821. 782 Hoofer, S.R., Van den Bussche, R.A., 2003. Molecular phylogenetics of the chiropteran 783 family Vespertilionidae. Acta Chiropterol. 5 (supplement), 1-63. 784 Horácek, I., 2001. On the early history of vespertilionid bats in Europe: the Lower Miocene 785 record from the Bohemian Massif. Lynx 32, 123-154. 786 Horácek, I., Hanák, V., Gaisler, J. (Eds.), 2000. Bats of the Palearctic region: a taxonomic 787 and biogeographic review. Platan publishing house, Warsaw, Poland. 788 Ibáñez, C., García-Mudarra, J.L., Ruedi, M., Stadelmann, B., Juste, J., 2006. The Iberian 789 contribution to cryptic diversity in European bats. Acta Chiropterol. 8, 277-297. 790 Janis, C.M., 1993. Tertiary mammal evolution in the context of changing climates, vegetation, 791 and tectonic events. Annu. Rev. Ecol. Syst. 24, 467-500. 792 Jiang, T.L., Sun, K.P., Chou, C.H., Zhang, Z.Z., Feng, J., 2010. First record of Myotis flavus 793 (Chiroptera: Vespertilionidae) from mainland China and a reassessment of its taxonomic 794 status. Zootaxa 2414, 41-51. 795 Jones, G., Parsons, S., Zhang, S.Y., Stadelmann, B., Benda, P., Ruedi, M., 2006. Echolocation 796 calls, wing shape, diet and phylogenetic diagnosis of the endemic Chinese bat Myotis 797 pequinius. Acta Chiropterol. 8, 451-463. 798
24
Kawai, K., Nikaido, M., Harada, M., Matsumura, S., Lin, L.K., Wu, Y., Hasegawa, M., 799 Okada, N., 2002. Intra- and interfamily relationships of Vespertilionidae inferred by various 800 molecular markers including SINE insertion data. J. Mol. Evol. 55, 284-301. 801 Kawai, K., Nikaido, M., Harada, M., Matsumura, S., Lin, L.K., Wu, Y., Hasegawa, M., 802 Okada, N., 2003. The status of the Japanese and East Asian bats of the genus Myotis 803 (Vespertilionidae) based on mitochondrial sequences. Mol. Phylogenet. Evol. 28, 297-307. 804 Kitchener, D.J., Cooper, N., Maryanto, I., 1995. The Myotis adversus (Chiroptera: 805 Vespertilionidae) species complex in Eastern Indonesia, Australia, Papua New Guinea and the 806 Solomon Islands. Rec. West. Aust. Mus. 17, 191-212. 807 Koopman, K.F., 1993. Order Chiroptera. In: Wilson, D.E., Reeder, D.M. (Eds.), Mammal 808 species of the World. A taxonomic and geographic reference. Smithsonian Institution Press, 809 Washington, pp. 137-242. 810 Koopman, K.F., 1994. Chiroptera: Systematics. In: Niethammer, J., Schliemann, H., Starck, 811 D. (Eds.), Handbuch der Zoologie. de Gruyter, Berlin, Germany, pp. 100-109. 812 Kruskop, S.V., Borisenko, A.V., Ivanova, N.V., Lim, B.K., Eger, J.L., 2012. Genetic diversity 813 of northeastern Palaearctic bats as revealed by DNA barcodes. Acta Chiropterol. 14, 1-14. 814 La Val, R.K., 1973. A revision of the Neotropical bats of the genus Myotis. Natural History 815 Museum of Los Angeles County, Sciences Bulletin 15, 1-54. 816 Lack, J.B., Roehrs, Z.P., Stanley Jr, C.E., Ruedi, M., Van den Bussche, R.A., 2010. Molecular 817 phylogenetics of Myotis indicate familial-level divergence for the genus Cistugo (Chiroptera). 818 J. Mammal. 91, 976-992. 819 Lack, J.B., Van Den Bussche, R.A., 2010. Identifying the confounding factors in resolving 820 phylogenetic relationships in Vespertilionidae. J. Mammal. 91, 1435-1448. 821 Larsen, R.J., Knapp, M.C., Genoways, H.H., Khan, F.A.A., Larsen, P.A., Wilson, D.E., 822 Baker, R.J., 2012a. Genetic diversity of Neotropical Myotis (Chiroptera: Vespertilionidae) 823 with an emphasis on South American species. Plos One 7. 824 Larsen, R.J., Larsen, P.A., Genoways, H.H., Catzeflis, F.M., Geluso, K., Kwiecinski, G.G., 825 Pedersen, S.C., Simal, F., Baker, R.J., 2012b. Evolutionary history of Caribbean species of 826 Myotis, with evidence of a third Lesser Antillean endemic. Mamm. Biol. 77, 124-134. 827 Matveev, V.A., Kruskop, S.V., Kramerov, D.A., 2005. Revalidation of Myotis petax Hollister, 828 1912 and its new status in connection with M. daubentonii (Kuhl, 1817) (Vespertilionidae, 829 Chiroptera). Acta Chiropterol. 7, 23-37. 830 Mein, P., Ginsburg, L., 1997. Les mammifères du gisement miocène inférieur de Li Mae 831 Long, Thaïlande: systématique, biostratigraphie et paléoenvironnement. Geodiversitas 19, 832 783-844. 833 Menu, H., Hand, S., Sigé, B., 2002. Oldest Australian vespertilionid (Microchiroptera) from 834 the early Miocene of Riversleigh, Queensland. Archeringa 26, 319-331. 835 Menu, H., Sigé, B., 1971. Nyctalodontie et myotodontie, importants caractères de grades 836 évolutifs chez les chiroptères entomophages. Comptes Rendus de l'Académie des Sciences de 837 Paris 272, 1735-1738. 838 Miller, S.A., Dykes, D.D., Polesky, H.F., 1988. A simple salting procedure for extracting 839 DNA from human nucleated cells. Nucleid Acids Res. 16, 215. 840 Murphy, W.J., Eizirik, E., O'Brien, S.J., Madsen, O., Scally, M., Douady, C.J., Teeling, E., 841 Ryder, O.A., Stanhope, M.J., de Jong, W.W., Springer, M.S., 2001. Resolution of the early 842 placental mammal radiation using Bayesian phylogenetics. Science 294, 2348-2351. 843 Myers, N., Mittermeier, R.A., Mittermeier, C.G., da Fonseca, G.A.B., Kent, J., 2000. 844 Biodiversity hotspots for conservation priorities. Nature 403, 853-858. 845 Nylander, J.A.A., 2004. MrModeltest v. 2.3. Program distributed by the author. . Uppsala 846 University, Sweden 847 Paradis, E., 1998. Detecting shifts in diversification rates without fossils. Am. Nat. 15, 176-848 187. 849 Purvis, A., Agapow, P.-M., Gittleman, J.L., Mace, G.M., 2000. Nonrandom extinction and the 850 loss of evolutionary history. Science 288, 328-330. 851
25
Rambaut, A., Drummond, A.J., 2009. Tracer v1.5. Available from 852 http://beast.bio.ed.ac.uk/Tracer 853 Rautenbach, I.L., Bronner, G.N., Schlitter, D.A., 1993. Karyotypic data and attendant 854 systematic implications for the bats of southern Africa. Koedoe 36, 87-104. 855 Ree, R.H., Smith, S.A., 2008. Maximum likelihood inference of geographic range evolution 856 by dispersal, local extinction, and cladogenesis. Syst. Biol. 57, 4-14. 857 Ronquist, F., Huelsenbeck, J.P., 2003. MRBAYES 3: Bayesian phylogenetic inference under 858 mixed models. Bioinformatics 19, 1572-1574. 859 Ruedi, M., Biswas, J., Csorba, G., 2012. Bats from the wet: two new species of Tube-nosed 860 bats (Chiroptera: Vespertilionidae) from Meghalaya, India. Rev. suisse Zool. 119, 111-135. 861 Ruedi, M., Mayer, F., 2001. Molecular systematics of bats of the genus Myotis 862 (Vespertilionidae) suggests deterministic ecomorphological convergences. Mol. Phylogenet. 863 Evol. 21, 436-448. 864 Salicini, I., Ibáñez, C., Juste, J., 2011. Multilocus phylogeny and species delimitation within 865 the Natterer‟s bat species complex in the Western Palearctic. Mol. Phylogenet. Evol. 61, 888-866 898. 867 Sanderson, M.J., Donoghue, M.J., 1996. Reconstructing shifts in diversification rates on 868 phylogenetic trees. Trends Ecol. Evol. 11, 15-20. 869 Sechrest, W., Brooks, T.M., da Fonseca, G.A., Konstant, W.R., Mittermeier, R.A., Purvis, A., 870 Rylands, A.B., Gittleman, J.L., 2002. Hotspots and the conservation of evolutionary history. 871 Proc. Natl. Acad. Sci. USA 99, 2067-2071. 872 Simmons, N.B., 2005. Order Chiroptera. In: Wilson, D.E., Reeder, D.M. (Eds.), Mammal 873 species of the World. A taxonomic and geographic reference. Johns Hopkins University Press, 874 Washington, pp. 312-529. 875 Sodhi, N.S., Koh, L.P., Brook, B.W., Ng, P.K.L., 2004. Southeast Asian biodiversity: an 876 impending disaster. Trends Ecol. Evol. 19, 654-660. 877 Stadelmann, B., Herrera, G., Arroyo-Cabrales, J., Ruedi, M., 2004a. Molecular systematics of 878 the piscivorous bat Myotis (Pizonyx) vivesi. J. Mammal. 85, 133-139. 879 Stadelmann, B., Jacobs, D., Schoeman, C., Ruedi, M., 2004b. Phylogeny of African Myotis 880 bats (Chiroptera, Vespertilionidae) inferred from cytochrome b sequences. Acta Chiropterol. 881 6, 177-192. 882 Stadelmann, B., Kunz, T.H., Lin, L.K., Ruedi, M., 2007. Molecular phylogeny of New World 883 Myotis (Chiroptera, Vespertilionidae) inferred from mitochondrial and nuclear DNA genes. 884 Mol. Phylogenet. Evol. 43, 32-48. 885 Stamatakis, A., 2006. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses 886 with thousands of taxa and mixed models. Bioinformatics 22, 2688-2690. 887 Tate, G.H., 1941. A review of the genus Myotis (Chiroptera) of Eurasia, with special 888 reference to species occurring in the East Indies. B. Am. Mus. Nat. Hist. 78, 537-565. 889 Teeling, E.C., Springer, M.S., Madsen, O., Bates, P., O'Brien, S.J., Murphy, W.J., 2005. A 890 molecular phylogeny for bats illuminates biogeography and the fossil record. Science 307, 891 580-584. 892 Tiunov, M.P., Kruskop, S.V., Feng, J., 2011. A new mouse-eared bat (Mammalia: Chiroptera, 893 Vespertilionidae) from South China. Acta Chiropterol. 13, 271-278. 894 Topál, G., 1970. On the systematic status of Pipistrellus annectans Dobson, 1871 and Myotis 895 primula Thomas, 1920 (Mammalia). Annals hist.-nat. Mus. nat. hung. 62, 373-379. 896 Topál, G., 1983. New and rare fossil mouse-eared bats from the Middle Pliocene of Hungary 897 (Mammalia, Chiroptera). Fragm. Min. Palaeont. 11, 43-54. 898 Tsytsulina, K., 2004. On the taxonomical status of Myotis abei Yoshikura, 1944 (Chiroptera, 899 Vespertilionidae). Zool. Sci. 21, 963-966. 900 Tsytsulina, K., Dick, M.H., Maeda, K., Masuda, R., 2012. Systematics and phylogeography of 901 the steppe whiskered bat Myotis aurascens Kuzyakin, 1935 (Chiroptera, Vespertilionidae). 902 Russian J. Theriol. 11, 1-20. 903
26
Volleth, M., Heller, K.G., 2012. Varations on a theme: Karyotype comparison in Eurasian 904 Myotis species and implications for phylogeny. Vespertilio 16, 329-350. 905 Volleth, M., Tidemann, C.R., 1991. The origin of the Australian Vespertilioninae bats, as 906 indicated by chromosomal studies. Z. Saugetierkd. 56, 321-330. 907 Weyeneth, N., Goodman, S.M., Ruedi, M., 2011. Do diversification models of Madagascar‟s 908 biota explain the population structure of the endemic bat Myotis goudoti (Chiroptera: 909 Vespertilionidae)? J. Biogeogr. 38, 44-54. 910 Zhang, Z.Z., Tan, X.Y., Sun, K.P., Liu, S., Xu, L.J., Feng, J., 2009. Molecular systematics of 911 the Chinese Myotis (Chiroptera, Vespertilionidae) inferred from cytochrome-b sequences. 912 Mammalia 73, 323-330. 913 Ziegler, R., 2003. Bats (Chiroptera, Mammalia) from Middle Miocene karstic fissure fillings 914 of Petersbuch near Eichstätt, Southern Franconian Alb (Bavaria). Geobios 36, 447-490. 915 916 917
918
27
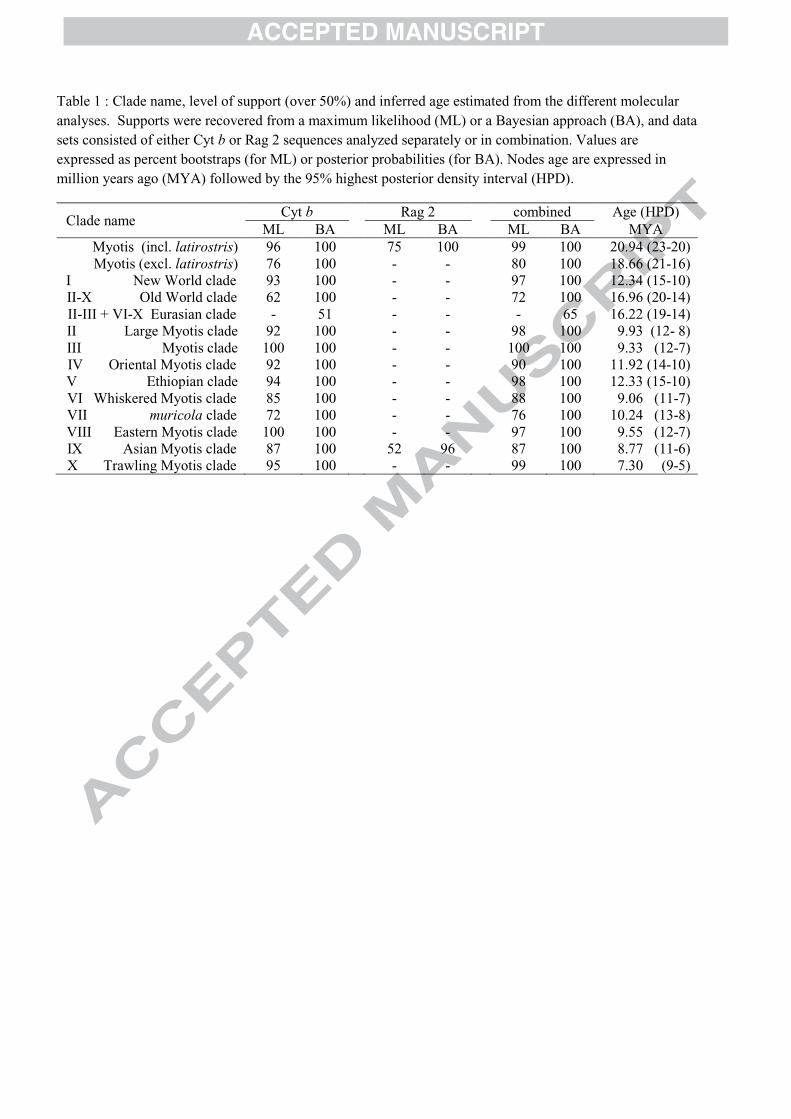
Table 1 Clade name, level of support (over 50%) and inferred age estimated from the 919
different molecular analyses. Supports were recovered from a maximum likelihood (ML) or a 920
Bayesian approach (BA), and data sets consisted of either Cyt b or Rag 2 sequences analyzed 921
separately or in combination. Values are expressed as percent bootstraps (for ML) or posterior 922
probabilities (for BA). Node age is expressed in million years ago (MYA) followed by the 923
95% highest posterior density interval (HPD). 924
925
Table S1 Origin and biogeographic assignation of the specimens analyzed for the Cyt b and 926
Rag 2 genes. Species names not listed in Simmons (2005) are marked with a star (*) and their 927
taxonomy discussed in the material and methods section. The symbol ø denotes information 928
unavailable. Vouchers (if any) are deposited in the following institutions or belong to personal 929
collections: Instituto Politécnico Nacional in Mexico (CDR), Estacion Biologica de Doñana 930
(EBD), Frieder Mayer (FM), Field Museum of Natural History in Chicago (FMNH), Gareth 931
Jones (GJ), Jean-François Maillard, DIREN, Martinique (JFM), Kuniko Kawai (KK), Kishi 932
Maeda (KM), Natural History Museum of Geneva (MHNG), National Natural History 933
Museum of Paris (MNHN), Manuel Ruedi (MR), Museum of Vertebrate Zoology at Berkeley 934
(MVZ), National Museum Prague (NMP), Osaka City University Graduate School of 935
Medicine (OCUMS), Royal Ontario Museum (ROM), Sumiko Matsumura (SM), 936
Senckenberg Museum of Frankfurt (SMF), T.H. Kunz (THK), Tigga Kingston (TiK), 937
Museum of Texas Tech University (TK), Transvaal Museum, South Africa (TM), University 938
of Alaska Museum (UAM), UKM (Universiti Kebangsaan Malaysia), Zoological Museum of 939
Moscow State University (ZMMU), Z. Zhang (ZZ). 940
941
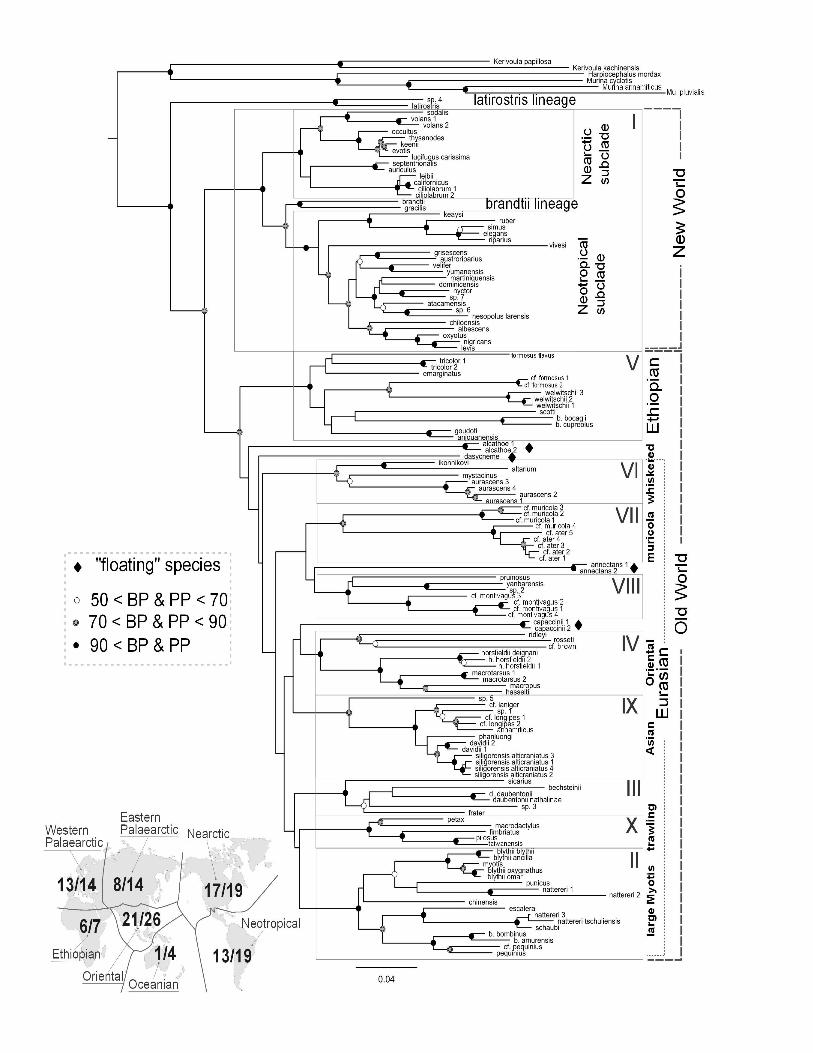
Figure 1 Maximum likelihood tree for 135 sequences of Myotis and six outgroup species 942
based on analyses of the combined mitochondrial and nuclear data set (1140 bp of Cyt b and 943
1148 bp of Rag 2). Nodal support is represented as standard bootstrap value (BP) obtained 944
with RAxML or posterior probabilities (PP) obtained with MrBayes. Empty circles represent 945
28
support values comprised between 50 and 70 %, grey circles values between 71 and 90% and 946
filled circles values above 90% in both ML and BA reconstructions. Species with uncertain 947
phylogenetic position (“floating species”) are indicated with a filled diamond. The various 948
clades discussed in the text are highlighted by boxes or broken lines. The inset represents the 949
geographic distribution of the 103 Myotis species listed in Simmons (2005) (right value) 950
versus the numbers sampled for the molecular reconstructions (left values) in the different 951
biogeographic regions of the world. 952
953
Figure 2 Chronogram of Myotis taxa based on Bayesian dating analysis using BEAST and the 954
combined mitochondrial and nuclear DNA gene dataset. Mean divergence values (expressed 955
as million year ago, MYA) are given at each node and horizontal bars represent the 95% 956
highest posterior density ranges. Clade names correspond to those given in Fig. 1. 957
958
Figure 3 Biogeographic evolution of ancestral areas of Myotis species reconstructed in the 959
chronogram of Fig. 2. The most likely ancestral areas determined with Lagrange under a 960
model with dispersals limited to adjacent bioregions are given in boxes at each node. The 961
inset map illustrates the seven biogeographic regions included for this study – A: the Western 962
Palaearctic, B: the Eastern Palaearctic, C: the Oriental, D: the Oceanian, E: the Ethiopian, F: 963
the Nearctic and G: the Neotropical region. Inferred range expansion events are highlighted 964
by black boxes (on the chronogram) or by arrows (on the world map), and vicariance events 965
by filled diamonds. The bottom scale represents time (in million years before present, MYA) 966
and the approximate subdivision of paleontological epochs. 967
968
Figure S1 Majority rule (50%) consensus Bayesian tree with posterior probability values 969
obtained with the analysis of 135 mitochondrial Cyt b sequences of Myotis taxa. Six outgroup 970
taxa from the Murininae and Kerivoulinae were used to root the tree. 971
972
29
Figure S2 Majority rule (50%) consensus Bayesian tree with posterior probability values 973
obtained with the analysis of 122 nuclear Rag 2 sequences of Myotis taxa. Six outgroup taxa 974
from the Murininae and Kerivoulinae were used to root the tree. 975
976
Figure S3 Occlusal views of right dentaries showing the nyctalodont lower molars (where the 977
postcristid links the hypoconulid) of Myotis latirostris (A.) and M. siligorensis alticraniatus 978
(B.), and the myotodont molars (where the postcristid links the entoconid) of Cistugo seabrae 979
(C.). See Menu and Sigé (1971) for a definition of lower molar configurations. 980
981
Table 1 : Clade name, level of support (over 50%) and inferred age estimated from the different molecular analyses. Supports were recovered from a maximum likelihood (ML) or a Bayesian approach (BA), and data sets consisted of either Cyt b or Rag 2 sequences analyzed separately or in combination. Values are expressed as percent bootstraps (for ML) or posterior probabilities (for BA). Nodes age are expressed in million years ago (MYA) followed by the 95% highest posterior density interval (HPD).
Clade name Cyt b Rag 2 combined Age (HPD)
ML BA ML BA ML BA MYA Myotis (incl. latirostris) 96 100 75 100 99 100 20.94 (23-20)
Myotis (excl. latirostris) 76 100 - - 80 100 18.66 (21-16) I New World clade 93 100 - - 97 100 12.34 (15-10) II-X Old World clade 62 100 - - 72 100 16.96 (20-14) II-III + VI-X Eurasian clade - 51 - - - 65 16.22 (19-14) II Large Myotis clade 92 100 - - 98 100 9.93 (12- 8) III Myotis clade 100 100 - - 100 100 9.33 (12-7) IV Oriental Myotis clade 92 100 - - 90 100 11.92 (14-10) V Ethiopian clade 94 100 - - 98 100 12.33 (15-10) VI Whiskered Myotis clade 85 100 - - 88 100 9.06 (11-7) VII muricola clade 72 100 - - 76 100 10.24 (13-8) VIII Eastern Myotis clade 100 100 - - 97 100 9.55 (12-7) IX Asian Myotis clade 87 100 52 96 87 100 8.77 (11-6) X Trawling Myotis clade 95 100 - - 99 100 7.30 (9-5)
50 < BP & PP < 70
70 < BP & PP < 90
90 < BP & PP
"floating" species
blythii omari
annamiticus
taiwanensis
cf. montivagus 4
cf. montivagus 2cf. montivagus 1
cf. montivagus 3
h. horsfieldii 2horsfieldii deignani
h. horsfieldii 1
cf. muricola 4
Murina annamiticus
latirostris lineage
Ne
w W
orld
Eth
iop
ian
wh
iskere
dm
uri
co
laO
rie
nta
lA
sia
nla
rge
My
oti
s
I
V
VI
VII
VIII
IV
IX
III
X
II
Eura
sia
n
Mu. pluvialis
daubentonii nathalinae
Nearc
tic
subcla
de
Neotr
op
ical
subcla
de
brandtii lineage
Old
World
pilosus
tra
wli
ng
cf. browni
cf. formosus 2cf. formosus 1
formosus flavus
WesternPalaearctic
EasternPalaearctic
Ethiopian
Oriental
13/14
1/4
6/7 21/26
8/14 17/19
13/19
Nearctic
0.0 MYA5.010.015.020.025.0
nesopolus
tricolor 2tricolor 1
brandtii
23.1
26.2
20.9
18.7
12.3
7.8
16.4
12.3
12.3
17.0
12.3
16.2
15.5
15.2
12.9
10.2
9.1
12.1
14.8 13.4
12.68.8
4.5
3.1
14.2
12.5
13.1
7.811.9
9.8
9.9
8.1
9.4
0.6
0.3
1.6
1.3
3.2
4.44.9
4.2
1.4
2.8
2.9
4.0
5.64.1
2.34.4
0.58
0.35
0.55
1.1
1.4
1.4
0.8
0.6
0.6
0.5
1.2
0.7
pilosus
b. oxygnathusb. omari
b. blythiib. ancilla
b. amurensis
n. tschuliensis
d. nathalinae
taiwanensis
s. alticraniatus 3s. alticraniatus 2
s. alticraniatus 1s. alticraniatus 4
lucifugus
I
V
VI
VII
VIII
IX
IV
X
III
II
cf. montivagus 4
formosus flavus
cf. formosus 1cf. formosus 2
6.6
6.0
4.46.01.0
2.7
1.1
8.5
10.9
9.4
6.43.8
8.5
6.8
6.0
5.58.1
9.3
8.2
11.3
11.6 9.53.1
1.7
7.8
5.9
2.5
2.6
2.9
2.4
9.6
8.8
5.5
4.9 2.2
3.4
3.0
2.2
8.6 2.0
6.34.9
7.3
6.0
5.11.7
9.38.4
6.8
7.1
1.0
8.98.2
6.42.7
1.34.4
5.94.3
3.0
1.8
1.9
1.0
0.9
1.1
1.5
1.9
1.71.1
1.6
0.9
0.5
1.3
0.8
brandtii
pilosus
Miocene Pliocene Plei
s. alticraniatus
FF
cf.muricola1-3cf.muricola 4
Harpiocephalus
l. carissima
n. larensis
cf. browni
taiwanensis
b. ancillab. blythii
n. tschuliensis
Oligocen.
I
V
VI
VII
VIII
IX
IV
X
III
II
f. flavus
cf. formosus
C
C
C
CCC
C
C
C
C
C
C
C
C
CC
CC
C
C
C C
C
CC
CC
C
C
C C
C
C
CC
C
C
C
F
F
F
F
F
F
F
FF
F
F
F
FF
F
F
GG
G
G
G
G
G
G
G
G
G
GG
G
GG
G
B
B
B
B
BB
BB
B
E
E
E
E
E
BF
A
FG
FG
FG
CE
CA
E
E
A
AC
A
AC
AC
A A
C
BC
A
C
AC
A
C
AC
A
B
AB
A
ABA
F
A
AA A
CB
D
A
C
vicariance (8x)
expansions (21x)
0.0 MYA5.010.015.020.025.0
AB
E ECC CE
C AC
A ABC BC
A AB
C BC
C BC
C CB
C CD
B BF
B BA
F FG
C CA
C AC
C AC
C AC
A AC
A AB
C BC
C BC
Ethiopian
clade
latirostris lineage
New World Myotis
Nearctic
subclade
brandtii lineage
Neotropical
subclade
Eurasian
clade
25 15 10 5 MYA 20 0
Old World Myotis
BC
C
B => BF
C => CE
C => CA
B
C
Nearly complete phylogeny of Myotis distributed worldwide
New World versus Old World clade
Likelihood models of range evolution identify Eastern Asia for their origins
Ancestral area reconstructions suggest 21 intercontinental dispersals during 21 MYA of
evolution
Discrepancies in molecular dating and interpretation of fossil record due to homoplasies