Monoamine oxidases A and B (amino:oxygen oxidoreductase (deaminating) (flavin-containing), EC 1.4.3.4) have been identified in the outer membranes of rat liver mitochondria by their covalent reaction with the inhibitor, [3H]pargyline. On analysis by polyacrylamide gel electrophoresis under denaturing conditions. Monoamine oxidase A was found to migrate more slowly that monoamine oxidase B. Proteins which correspond to monoamine oxidases A and B (as identified by the electrophoretic distribution of covalently bound [3H]pargyline) were excised from the gels. Subsequent analysis showed that both monoamine oxidase A and monoamine B had been highly purified by this procedure. Electrophoretic analysis of the peptides produced by limited proteolysis with bovine trypsin, a-chymotrypsin, Staphylococcus aureus V8 proteinase and cyanogen bromide indicate that monoamine oxidases A and B have different amino acid sequences.
Introduction
Monoamine oxidase (amino:oxygen oxidore- ductase (deaminating) (flavin-containing), EC 1.4.3.4) activity occurs as two distinct enzymatic activities. One activity, monoamine oxidase A, is exceedingly sensitive to the inhibitor clorgyline, and uses serotonin but not benzylamine as a sub- strate. The other activity is several orders of mag- nitude less sensitive-to clorgyline and oxidizes benzylamine but not serotonin [1-3]. While there is good evidence for the existence of the mono- amine oxidase A and B activities, the molecular basis for the activities is not so clear. Immunologi- cal studies indicate that antibodies against bovine or human monoamine oxidase B do not crossreact with the monoamine oxidase A from cattle or man, respectively [4,5]. While these studies suggest that the two activities are catalyzed by different proteins, immunochemical studies with proteins
* To whom correspondence should be addressed.
which have been solubilized from membranes are often difficult to interpret because the aggregate state of the proteins is usually not known. Other workers have concluded that the two monoamine oxidase activities are immunochemically similar [6,7]. In other experiments [8,9], mitochondrial monoamine oxidases A and B have been specifi- cally and covalently labeled using a radioactive form of the inhibitor, pargyline. Subsequent elec- trophoretic analysis under denaturing conditions showed that the labeled enzymes had different electrophoretic mobilities and presumably differ- ent molecular weights. Finally, after electro- phoretic analysis of proteolytic digests of mitochondria in which monoamine oxidases A and B had been labeled with radioactive pargyline, the distribution of the labeled fragments from the two monoamine oxidases appeared to be different [10,11]. While these last studies strongly indicate that the two monoamine oxidase activities are carried on different polypeptides, the possibility that the distributions of the fragments may be a
consequence of the influence of non-identical post translational modifications on the specificity of the proteolytic cleavage rather than differences in the amino acid sequences is not ruled out. In this connection, the amino acid sequence surrounding the covalently bound monoamine oxidase flavin, Ser-Gly-Gly-Cys(flavin)-Tyr, has been shown to be identical in both monoamine oxidase A and monoamine oxidase B from several species [12,13].
In our study the outer membranes of rat liver mitochondria have been used as a source for monoamine oxidases A and B. The two enzymes were identified by the electrophoretic distribution of [3H]pargyline which had been bound to the outer membranes during an incubation with this inhibitor. Monoamine oxidases A and B were ex- cised from the gels, and were subjected to limited proteolysis. The fragments produced from the two purified monoamine oxidases after treatment with various proteinases and cyanogen bromide were then analyzed by polyacrylamide gel electrophore- sis. The results of these experiments provide strong evidence that monoamine oxidase A and B en- zymatic activities are carried on two proteins with different amino acid sequences.
Materials and Methods
Chemicals. Acrylamide, bisacrylamide, am- monium persulfate and other reagents used in electrophoresis were purchased from Bio-Rad Laboratories. Trypsin and ~-chymotrypsin were products of the Sigma Chemical Company, while Staphylococcus aureus V-8 proteinase was bought from Miles Laboratories. These and other chem- icals purchased from various sources were all ob- tained in the highest possible purity. Animals were obtained from Charles River Breeding Labs.
Preparation of outer membranes. Outer mem- branes were prepared by a modification of the technique of Sottocasa et al. [14]. A typical pre- paration consisted of twelve male Wistar rats 250-300 g that were fasted overnight. The rats were killed by decapitation and the liver was per- fused with 20 ml of cold, isotonic saline. All subsequent operations were performed at 4°C. The livers were minced in a Harvard tissue press, ho- mogenized in 0.25 M sucrose and brought to a volume of 70 ml per liver. The homogenate was
centrifuged at 500 x g for 10 min and the resulting supernatant was again centrifuged at 7500 x g for 10 min to sediment the mitochondria. The mitochondria were resuspended in 0.25 M sucrose to 70 ml per liver and sedimented at 7500 x g for 10 min. This procedure was repeated a third time. The mitochondrial pellet was resuspended in 10 mM Tris phosphate (pH 7.5), 10 ml per liver, and allowed to stand on ice for 10 min. Sucrose was added to a final concentration of 0.40 M; ATP and MgSO 4 were added to 1.0 mM concentration and a final volume of 160 ml. After another 10 rain on ice, 20-ml aliquots were sonicated for 15 s using the large probe at the No. 6 setting of a Branson W185D sonifier. 10 ml of the mitochon- drial solution were then layered over 15 ml of 1.2 M sucrose and centrifuged at 78000 x g for 60 rain. The outer membranes which sedimented to the interface of the two sucrose solutions were collected with a Pasteur pipette and were washed with 100 ml 0.15 M NaC1/0.5 M Tris-HC1 (pH 7.5), and then collected by centrifugation at 78 000 × g for 60 min. These membranes were then re- suspended in 0.25 M sucrose to a volume of 0.25 ml per liver and were stored at - 2 0 ° C . Protein was assayed according to the method of Lowry et al. [15] and the protein concentration was typically 2 m g / m l .
Preparation and proteolytic analyses of mono- amine oxidases A and B. Outer membranes were dissolved by boiling in a buffer composed of 62.5 mM Tris-HC1 (pH 6.8), 2% SDS, 10% glycerol, 1.5 mM N-ethylmaleimide and 0.001% Bromphenol blue. About 3 mg of outer membrane proteins were separated according to the method of Laem- mli [16] using a 140 x 100 x 3 mm separating gel (10% monomer content) and a 140 × 5 x 3 mm stacking gel. After electrophoresis for about 20 h at 20 mA, the gels were briefly stained using 0.1% Coomassie brilliant blue in 7.5% acetic acid and those regions identified as monoamine oxidases A and B were cut out. The gel segments were then washed for 30 min in a solution of 125 mM Tris-HC1 (pH 6.8)/0.1% SDS/1 mM E D T A / 1 . 5 mM 2-mercaptoethanol and then frozen in the buffer solution. Accurate determination of protein concentrations under these conditions was very difficult and so the amount of monoamine oxidase B was calculated based on previous estimates [17]
to be 2.5% of the total outer membrane protein, and monoamine oxidase A was estimated to be present in half that amount based on the relative staining intensity of the two proteins.
Thawed gel segments were further analyzed by electrophoresis in gels with 7.5-15% monomer content by placing them in the wells of the stack- ing gel and overlaying the segments with 20% glycerol and 0.001% Bromphenol blue dissolved in the wash buffer described above. Peptide mapping was performed in this system as first described by Cleveland et al. [18]. Briefly, an additional overlay containing a proteolytic enzyme in 10% glycerol was added to the well and the sample was sub- jected to electrophoresis until the dye front reached the edge of the separating gel. Then the current was shut off for 30 min to permit proteolysis. After the products had been separated by electro- phoresis, the gels were stained using freshly pre- pared 0.1% Coomassie brilliant blue in 50% tri- chloroacetic acid and destained in 10% trichloro- acetic acid/10% methanol. Cyanogen bromide fragments of monoamine oxidases A and B were generated according to the procedure of Nikodem and Fesco [19]. After initial separation in a 10% gel, the slices were washed as above and placed in tubes. 225-250 mg cyanogen bromide were dis- solved in 100 #1 acetonitrile and 20 #1 of this solution were added to the tube containing the slice, followed by 200/~1 of 0.125 M Tris-HCl (pH 6.8) and 200 #1 0.6 M HC1. Controls contained all reagents except cyanogen bromide. The digestion was at 37°C for up to 1 h. The slices were then washed with 25 ml of 0.125 M Tris (pH 6.8)/0.1% SDS for 1 h with several changes of buffer. The slices were finally placed in the wash buffer which contained Bromphe~ol blue and mercaptoethanol and the fragments were separated by electrophore- sis in a 15% gel. The cyanogen bromide fragments were visualized by silver staining [20].
Other methods. Monoamine oxidases A and B were measured by using serotonin [2] a.nd benzyl- amine [4], respectively, as substrates. Fluorography was performed using EN3HANCE, manufactured by New England Nuclear Corp., according to the instructions supplied with the product. Dried gels were used to exposed Kodak X-Omat RP film for a period of several days.
Results
Monoamine oxidases A and B activities were measured in outer membranes prepared as de- scribed in Materials and Methods, and both activi- ties were present in rather high specific enzymatic activity. Monoamine oxidase A was quite sensitive to clorgyline but was not inhibited by pargyline unless the inhibitor concentration was greater than 1 -10 6 M. Monoamine oxidase B, on the other hand, could be completely inhibited by 1 • 10-7 M pargyline but was comparatively insensitive to clorgyline. It was known that pargyline is oxidized by monoamine oxidase and subsequently binds covalently, mole for mole, with the monoamine oxidase flavin [21,22]. Since the flavin cofactor is linked to the apoenzyme by a thioether bridge [23], it is possible to label specifically the monoamine
OM A AB B
l ' - 4 1
B
Fig. 1. About 100 t~g of outer membrane protein were in- cubated alone (AB) or with 10 6 M pargyline (A) or 10 -v M clorgyline (B). The outer membranes were collected by centrifu- gation at 100000× g for 30 min and resuspended in 0.1 ml 50 mM potassium phosphate (8.0). This mixture was transfered to a small polypropylene tube which contained 1 nmol of tritiated pargyline (New England Nuclear). The radioactive pargyline (normally in ethanol) had been dried under a stream of nitro- gen. The outer membranes and labeled pargyline were per- mitted to react for 60 min and then the sample was solubilized and subjected to SDS-polyacrylamide gel electrophoresis and fluorography (Materials and Methods). Another 100/~g sample of outer membranes (OM) was subjected to electrophoresis but then stained with Coomassie brilliant blue.
oxidase isoenzymes using a radioactive form of pargyline [8,17]. It can be seen in Fig. 1 that if outer membranes are incubated with a tritiated form of pargyline in a concentration that inhibits both monoamine oxidase A and monoamine oxidase B (1 .10 5 M) and then analyzed by SDS-polyacrylamide gel electrophoresis and fluo- rography, two prominent radioactive proteins are present. If monoamine oxidase B is completely inhibited by preincubation with 1 .10 -6 M un- labeled pargyline, only one band, monoamine oxidase A, is evident. If monoamine oxidase A is completely inhibited by preincubation with 10 7 M clorgyline, only monoamine oxidase B is de- tected. These results are similar to observations made by others [8-11] in crude mitochondrial fractions from various tissues. The outer mem- branes used in our experiments are highly purified organelles that are rich in both forms of mono- amine oxidase, and since the labeled bands corre- sponded to proteins that could be detected in
I.O
0 .5
~ 0 . 1
O
>
o
I I I I "?.5 I0.0 12.5 15.0
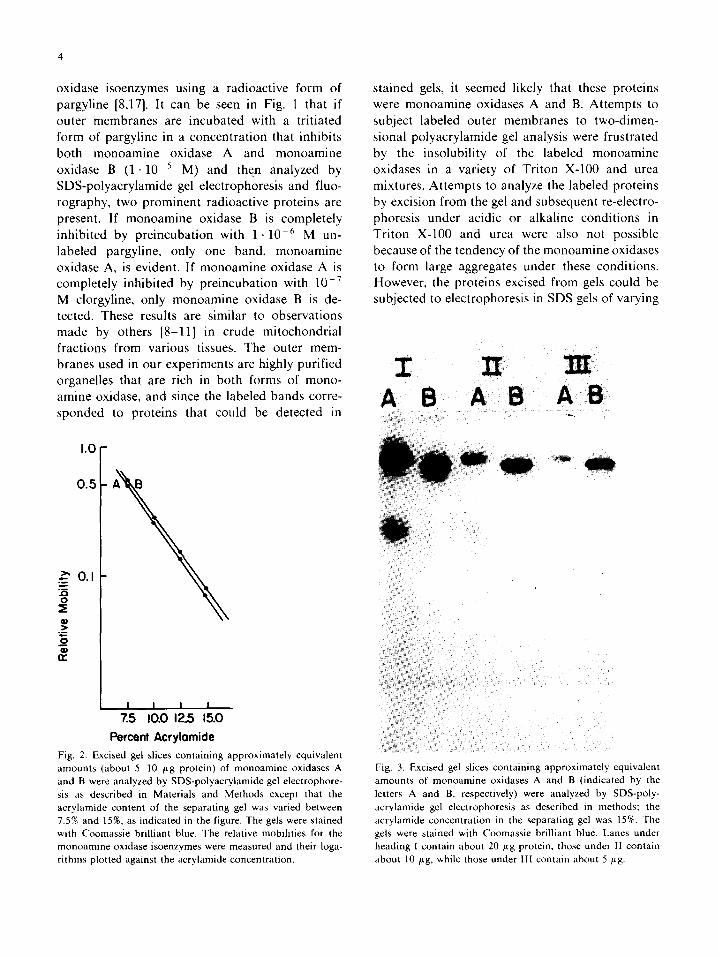
Percent Acrylomide Fig. 2. Excised gel slices containing approximately equivalent amounts (about 5-10 ~tg protein) of monoamine oxidases A and B were analyzed by SDS-polyacrylamide gel electrophore- sis as described in Materials and Methods except that the acrylamide content of the separating gel was varied between 7.5% and 15%, as indicated in the figure. The gels were stained with Coomassie brilliant blue. The relative mobilities for the monoamine oxidase isoenzymes were measured and their loga- rithms plotted against the acrylamide concentration.
stained gels, it seemed likely that these proteins were monoamine oxidases A and B. Attempts to subject labeled outer membranes to two-dimen- sional polyacrylamide gel analysis were frustrated by the insolubility of the labeled monoamine oxidases in a variety of Triton X-100 and urea mixtures. Attempts to analyze the labeled proteins by excision from the gel and subsequent re-electro- phoresis under acidic or alkaline conditions in Triton X-100 and urea were also not possible because of the tendency of the monoamine oxidases to form large aggregates under these conditions. However, the proteins excised from gels could be subjected to electrophoresis in SDS gels of varying
A B A B A B
Fig. 3. Excised gel slices containing approximately equivalent amounts of monoamine oxidases A and B (indicated by the letters A and B, respectively) were analyzed by SDS-poly- acrylamide gel electrophoresis as described in methods; the acrylamide concentration in the separating gel was 15%. The gels were stained with Coomassie brilliant blue. Lanes under heading I contain about 20 /xg protein, those under II contain about 10 ,ug, while those under llI contain about 5 btg.
acrylamide content and then stained with Coomas- sie brilliant blue. This type of analysis has been useful in resolving cytochrome oxidase subunits which comigrated when analyzed at one gel con- centration [24]. With the monoamine oxidases there was a linear relationship between the logarithm of the relative electrophoretic mobility and the acrylamide concentration and there was not evi- dence of band splitting at any of the acrylamide concentrations (Fig. 2). An experiment of this type in which a 15% acrylamide gel was used to analyze various amounts of monoamine oxidase A and B is shown in Fig. 3. In the studies with cytochrome oxidase, the gels with the highest acrylamide con- tent were most effective at resolving subunits, but the monoamine oxidase forms did not appear to be contaminated one with the other and except for a slight amount of a low molecular weight con- taminants in the monoamine oxidase A prepara- tion the two enzymes were homogeneous as judged by SDS-polyacrylamide gel electrophoresis.
A B A B A B A B
Fig. 4. Excised gel slices containing approx. 40 ,ttg of mono- amine oxidase A and B protein (indicated by the letters A and B, respectively) were used to derive peptide maps with the bacterial V-8 proteinase as described in Materials and Meth- ods. The amounts of proteinase that were used are as follows: 0.05 #g (I), 0.l ~g (II), 0.5 #g (III) and 1 btg (IV). The gels were stained with Coomassie brilliant blue.
I A B A B A B A B
Fig. 5. Excised gel slices containing approx. 40 ,ttg of mono- amine oxidase A and B protein (indicated by the letters A and B, respectively) were used to derive peptide maps as described in Materials and Methods. Lanes under the heading I were produced using 5 #g of a-chymotrypsin, those under II with 5 ,ag of trypsin and those under IV with 1250 mg cyanogen bromide per mg protein for 60 min. The two lanes under heading II! are controls for the cyanogen bromide experiment in which 40 ~g of each isoenzyme was incubated in the buffer system for the cyanogen bromide cleavage but without cyano- gen bromide.
Preparations of monoamine oxidases A and B similar to those depicted in Fig. 4 were then used to produce peptide maps according to the method of Cleveland et al. [18] using three proteinases: bovine trypsin, bovine a-chymotrypsin and S. a u r e u s V-8. Since under our conditions accurate estimates of the substrate concentration cannot be made, several concentrations of each proteinase were employed to rule out artifacts that might have been caused by differences in the ratios of the substrate and proteinase. After electrophoresis, the gels were stained so that the fragments could be compared. An example of this type of analysis is shown in Fig. 5. In this experiment the S. a u r e u s
V-8 proteinase was used in amounts ranging from 0.05 to 1 /~g against about 40 #g of each of monoamine oxidase A or monoamine oxidase B. The peptide maps from these two enzymes are very different, irrespective of the amount of pro- teinase (i.e. extent of proteolysis) used to generate the patterns. In fact, after the extensive proteolysis
seem with 1 ~g of the V-8 proteinase there appear to be very few fragments common to the two enzymes. A similar analysis of monoamine oxidases A and B was performed using a range of con- centrations of trypsin (1-25 t~g) and a- chymotrypsin (also 1-25 /tg) against about 40 #g of each isoenzyme. Representative patterns from these analyses are depicted in Fig. 5 and it can be seen that with both of these proteinases, as with the V-8 proteinase, the peptide maps from mono- amine oxidases A and B are very different. Finally, monoamine oxidases A and B which had been excised from gels were reacted for 60 min with a large excess of cyanogen bromide and then again subjected to SDS-gel electrophoresis; the patterns of fragments produced by this procedure are also shown in Fig. 5. Once again monoamine oxidases A and B have markedly different peptide maps.
Discussion
Immunologic evidence [4,5], electrophoretic estimates of molecular weight [8,9] and compari- sons of the electrophoretic distribution of pargy- line-labeled fragments after partial proteolysis [10,11] indicate that monoamine oxidase A and B are different proteins. Rat liver represents a con- venient source for both isoenzymes from a single tissue and species, and, furthermore, previous work [25,26] indicates that at least the B-isoenzyme is catalytically active as a monomer so that compari- sons between the enzymes should be less ambigu- ous than with monoamine oxidases that have been reported to contain several subunits. We have purified both of these isoenzymes from rat liver mitochondrial outer membranes by an electro- phoretic method. Our enzyme preparations appear to approach homogeneity as judged by SDS-poly- acrylamide gel electrophoresis; however, the in- solubility of the purified monoamine oxidases in non-ionic denaturants prevented us from using isoelectric focusing as an estimate of purity.
The purified enzymes were used to generate proteolytic fragments and these fragments were compared by electrophoresis. Others [10,11] have compared proteolytic fragments derived from monoamine oxidases A and B after the isoenzymes had been radiolabeled with tritiated pargyline; however, because the enzymes were labeled and
proteolytis was performed with mitochondria, it was only possible to compare radioactive (active site) fragments. Furthermore, because several [3H]pargyline fragments were obtained from each enzyme, it seemed that proteolysis was incomplete, and the apparent differences between the mono- amine oxidases may have been due more to the rate of proteolysis than to differences in primary structure. In our experiments, proteolytic digests of monoamine oxidases A and B were analyzed using stained gels so that a large number of frag- ments could be compared, and in each compari- son, not only are the results consistent with the work of others [7,10] that indicates that the two monoamine oxidases activities are catalyzed on different proteins, but our data also indicate that these monoamine oxidase isoenzymes differ con- siderably in primary structure. The markedly dif- ferent patterns persisted under a variety of condi- tions with three proteolytic enzymes and cyanogen bromide, and so it seems unlikely that the differ- ent maps are due to artifacts caused by different rates of proteolysis or resistance to enzymatic pro- teolysis as a consequence of different degrees of post-translational modification. The latter possibil- ity is ruled out by the cyanogen bromide maps. It is interesting that, although monoamine oxidases A and B have identical sequences of amino acids immediately adjacent to the covalently bound flavin [12], the two enzymes appear to have con- siderable differences in the primary structure of other regions of the molecule.
Acknowledgement
This work was supported by PHS Grant AM 19701, U.S. Government.