(Received 18 February 2002; accepted in revised form 27 March 2002)
Abstract. Elongation of the primer 32pdA(pdA)8pA proceeds by the reaction of the 5′-phosphori-midazolides of adenosine and uridine in the presence of montmorillonite clay. Daily addition ofthe activated nucleotides for up to 14 days results in the formation of 40–50 mers using the 5′-phosphorimidazolide of adenosine (ImpA) and 25–30 mers using the 5′-phosphorimidazolide ofuridine (ImpU). The limitation on the lengths of the chains formed is not due to the inhibitorsformed since the same chain lengths were formed using 2–3 times the amount of montmorillonitecatalyst. The shorter oligomers formed by the addition of U monomers is not due to its greater rateof decomposition since it was found that both the A and the U adducts decompose at about thesame rates. Alkaline phosphatase hydrolysis studies revealed that some of the oligomers are cappedat the 5′-end to form, with ImpA, Ap32pdA(pdA)8pA(pA)n. The extent of capping depends on thereaction time and the purine or pyrimidine base in the activated mononucleotide. Hydrolysis withribonuclease T2 followed by alkaline phosphatase determined the sites of the 3′, 5′- and 2′, 5′-phosphodiester bonding to the primer. The potential significance of the mineral catalyzed formationof 50 mer oligonucleotides to the origin of life based on RNA (the RNA world scenario) is discussed.
Keywords: mineral catalysis, montmorillonite, oligoadenylates, prebiotic synthesis, RNA, RNAelongation
1. Introduction
In the RNA world scenario RNA was the essential biopolymer in the first lifeon Earth because of its ability to store genetic information and catalyze reac-tions (Crick, 1968; Orgel, 1968; Cech et al., 1981; Guerrier-Takada, 1983; Gilbert,1986). In vitro evolution studies have demonstrated the broad catalytic activity ofRNA (Joyce et al., 1998) and the x-ray structure determination of the ribosomeestablished the central role of RNA catalysis in the biosynthesis of the peptidebond of proteins (Ban et al., 2000; Muth et al., 2000). While others propose thatthe RNA in the first life evolved from simpler biopolymers (Orgel, 1998), I aminvestigating the direct formation of RNA from RNA monomers as part of a studyof the role of catalysis in prebiotic synthesis (Ferris, 1993).
The synthesis of 6–14 mer oligonucleotides by the montmorillonite-catalyzedreaction of activated 5′-mononucleotides in pH 8 aqueous solution at 25 ◦C has
Figure 1. Activated monomers a. N = A, ImpA. b. N = U, ImpU.
been reported (Figure 1; Ferris, 1993). The reaction occurs with the 5′-activatedphosphates of adenosine, guanosine, uridine, cytidine and inosine (Ferris andErtem, 1993; Prabahar et al., 1997; Ding et al., 1996; Ertem and Ferris, 1997;Kawamura and Ferris, 1999).
Chain lengths of 6–14 mers are unlikely to have been sufficiently long for in-formation storage or catalysis in the first life on Earth (Joyce and Orgel, 1999;Szostok and Ellington, 1993). The initial report on the preparation of longer oli-gonucleotides (40–50 mers) by the addition of adenylic acid units to a 10 merprimer was described (Ferris et al., 1996). In the present report the elongation ofthe same primer with uridylic acid units is described. Results on the regioselectivityof phosphodiester bond formation and the extent of formation of 5′-caps, usingregiospecific hydrolytic reactions, is presented for both RNAs.
2. Experimental Section
The montmorillonite Volclay, a gift from the American Colloid Company, was con-verted to a homoionic Na+-montmorillonite by the titration (Banin, 1973; Banin etal., 1985). Mononucleotides were purchased from Sigma and converted to the 5′-phosphorimidazolides (Joyce et al., 1984). The 10 mer used as primer (dpA(pdA)8-pA) was synthesized on an Applied Biosystems 391 DNA synthesizer or purchasedfrom Operon. The 5′-terminal 32P-labeling of the primers was performed using[32P]ATP (Amersham) using T-4 polynucleotide kinase (New England Biolabs).Gel electrophoresis (Owl Scientific) using an EC-500 power supply (ApparatusCorporation) was performed on a 20% acrylamide-bisacrylamide denaturing gel on16 × 28 cm. plates. Autoradiography was performed using Kodak X-OMAT filmand the extent of 32P-labeling was measured on a Packard Tricarb 2100TR counter.Centrifugation was performed on an Eppendorf 5415 centrifuge and solutions wereconcentrated to dryness using an Eppendorf Concentrator model 5301. Reagentgrade chemicals were purchased.
MONTMORILLONITE CATALYSIS OF 30–50 MER OLIGONUCLEOTIDES 313
2.1. GEL ELECTROPHORESIS
Approximately equal counts of radioactive material were loaded into the wells ofthe electrophoresis gel together with xylene cyanole and bromphenol blue and thegel was run for ∼2.5 hr at 1000 volts. The gel was then transferred to Saran Wrapand autoradiographed overnight at –20 ◦C and the film was then developed. The10 mer, a 32P-labeled primer, 32pdA(pdA)8pA was assigned a relative migrationdistance of 10. Addition of a nucleotide to the 3′-end of the primer, an increaseof one nucleoside and one negative change, decreases the migration distance oneunit from 10 to 11. The band moving more slowly on the gel was assigned thehigher number since it has the higher molecular weight. The 5′-capped 10 mer,Ap32pdA(pdA)8pA, formed by the addition of one nucleoside with no changein the number of negative charges, migrated more slowly by about 1.5 units at11.5 units. From these data it was determined that the addition of one nucleosidedecreases the extent of migration by 1.5 units while the addition of one negativecharge increases the extent of migration by 0.5 unit. The extent of elongation ofthe oligomers was estimated using the above data. In general, it was not possibleto resolve the longer capped and not capped oligomers by gel electrophoresis so aband at about 15.5–16 may reflect the presence of both an uncapped 14 mer anda capped 12 mer. The areas of some of the bands were determined on a PowerMacintosh G3 computer using public domain NIH Image program, version 1.61(developed at the U.S. National Institutes of Health and available on the Internet athttp://rsb.iinfo.nih.gov/nih-image/).
2.2. ENZYMATIC HYDROLYSES
Alkaline Phosphatase (APH). 5′-terminal phosphate groups were cleaved with theenzyme (0.1–0.001 units) in the standard buffer (10 mM tris, 1 mM MgCl2, 1 mMZnCl2, pH 8.3) on incubation for 15 min at 37 ◦C, 30 min at 55 ◦C, followed byinactivation of the enzyme by addition of EDTA and heating for 10 min at 75 ◦C.Gel electrophoresis of the hydrolysis products revealed the capped oligomers.
Ribonuclease T2 (RNase T2), The oligomers were incubated with ribonucleaseT2 (RNase T2) (Sigma) (0.1–0.01 units) in ammonium acetate (pH 4.3) for 2 hr at37 ◦C and then 4–6 µL of 0.1 M ammonium hydroxide was added to bring the pH to9 to inactivate the enzyme. RNase T2 hydrolysis monitored by gel electrophoresisreveals the number of 2′, 5′-links to the primer before a 3′, 5′-link is formed. Thisanalysis is complicated by the presence of both capped and not capped oligomersas well as the possibility for the capped oligomers to elongate from both ends of theprimer. The subsequent APH hydrolysis reveals the capped hydrolysis products.
314 JAMES P. FERRIS
2.3. CONVERSION OF 32pdA(pdA)8pA TO Ap32pdA(pdA)8pA (CHU AND
ORGEL, 1984)
To 10 µL of a solution of 32pdA(pdA)8pA (93,000 counts) in water was added10 µL of 1 M MgCl2, 20 µL of 0.5 M HEPES buffer (pH 7) and 10 µL of 1 MImpA. The mixture was heated at 55 ◦C for 6 hr, desalted on a Nensorb (NewEngland Nuclear) column and the oligomer fraction was concentrated to drynessin the Eppendorf spin concentrator. The product was shown by electrophoresisto consist of a mixture of approximately 25% unreacted 32pdA(pdA)8pA, 50%Ap32pdA(pdA)8pA and 25% Ap32pdA(pdA)8pApA. Treatment with APH removedthe band assigned to 32pdA(pdA)8pA but not the bands assigned to Ap32pdA(pdA)8-pA and Ap32pdA(pdA)8pApA.
2.4. ELONGATION OF 32pdA(pdA)8pA
The reactions were first performed in 1.7 mL microcentrifuge tubes. To 2 mg Na+-montmorillonite in the microcentrifuge tube was added 32pdA(pdA)8pA (∼140,000counts) in 10 µL of water and 20 µL of a mixture of 0.4 M NaCl, 0.15 M MgCl2and 0.2 M HEPES (pH 8) (2x BE). Then 10 µL of 0.06 M ImpA was added andthe mixture was vortexed and allowed to stand at room temperature for 24 hr.The tube was then centrifuged at 14,000 rpm and the supernatant removed witha pipette. The montmorillonite was washed by resuspending the clay-oligomertwice in 40 µL of BE by stirring with a sterile, plastic inoculation loop (CopanDiagnostics), vortexing and centrifuging at 14,000 rpm for 6 min. The elongationwas initiated by the addition of 20 µL of 2X BE, 10 µL of water and 10 µL of0.06 M ImpA and suspending the clay in the solution by stirring with an inoculationloop and vortexing the reaction mixture for ∼1 min. The reaction was then washedas described after the first 24 hr of reaction. At the end of the desired number ofcycles the reaction solution was centrifuged and washed as described previouslyand to the clay-oligonucleotide mixture was added 40 µL of 0.1 M sodium pyro-phosphate (pH 9) and the clay was stirred to a slurry with the inoculation loop andthen vortexed for about 1 min. The tube was then centrifuged at 14,000 rpm for15 min or longer to give 30–40 µL of supernatant. The wash was performed twomore times and the eluates were combined and desalted using a Nensorb columnby the procedure supplied by the manufacturer. Recently it was found that the useof Pall filter centrifuge tubes containing a filter insert tube were more effective forthe generation of longer oligomers. The filter was first washed with ∼400 µL ofdistilled water and spun dry at 14,000 rpm and then 2 mg of clay was weighedinto the filter tube insert. The liquid reagents were added as described above andthe mixture was stirred to a slurry with an inoculation loop. The filter tube wasplaced in the accompanying microcentrifuge tube, was vortexed and placed in aclosed container which contained sufficient water to maintain the humidity andhence minimize the evaporation of the water from the reaction solution. The same
MONTMORILLONITE CATALYSIS OF 30–50 MER OLIGONUCLEOTIDES 315
procedure described for the reaction in the microcentrifuge tube was used for thesubsequent steps in the reactions performed in the filter tubes.
3. Results
Investigation of the effect of ImpA concentration on the extent of elongation of32pdA(pdA)8pA revealed that the optimum concentration was about 15 mM where6 monomer units were added to the primer in 24 hr. Use of 1.7 and 0.17 mMresulted in the addition of four and one monomer units, respectively. A 170 mMconcentration of ImpA resulted mainly in the formation of the capped primer,Ap32pdA(pdA)8pA and a small amount of 32pdA(pdA)8pApA. About 60% of theprimer was eluted from the montmorillonite with this concentration of ImpA ascompared to 10% when the lower concentrations were used. The inhibition ofelongation is due to displacement of the 32pdA(pdA)8pA from the montmorilloniteclay by the high ImpA concentration.
A brief investigation of the optimal amount of montmorillonite was undertakenusing 4 mg instead of the usual 2 mg in microcentrifuge tubes. A slower rate ofgrowth of oligomers was observed using the 4 mg of clay. In experiments to bedescribed later, the primers were elongated to the same chain lengths with eitheramount of clay but it just took longer with the larger amount of clay. Increasing theamount of clay to 10 mg and decreasing the solution volume to 25 µL resulted inthe more efficient binding of the primer to the clay surface but less elongation ofthe primer was observed than with 2 mg of clay and 40 µL volumes. These datasuggest that with a larger amount of clay the primer is spread over a larger surfacearea on the clay so there is less chance for ImpA to be bound proximate to the3′-end of the primer where it can react.
3.1. ELONGATION OF 32pdA(pdA)8pA WITH ImpA
The conditions determined above were used to explore the length of oligomersformed in a 12 day elongation reaction using ImpA in microcentrifuge tubes. Thechain length increased by about 27 mers after 6 days but no further elongationwas observed after 12 days (Figure 2). The principal process in the 6–11 day timeperiod was the reaction of the ImpA with the primer to add an Ap group to the5′-end of the oligomers as shown by the increase in intensity of an electrophoresisband due to Ap32pdA(pdA)8pA. That band in the gel for the elongation reactionhad the same migration distance as an authentic sample (Chu and Orgel, 1984).
It was observed during the elongation study that the amount of clay presentdecreased by about 25% at the end of 6 days of reaction but did not decrease furtherafter a 12 day reaction period. A possible explanation for the absence of elongationafter 6 days reaction time was the loss of some catalytic clay during the washingand pipetting procedures used in the experiments. That the decrease in the elong-ation was due in part to the loss of the clay catalyst was shown by the elongation
316 JAMES P. FERRIS
Figure 2. Gel electrophoresis of the elongation of 32pdA(pdA)8pA with ImpA in microcentrifugetubes. Lane 1, 32pdA(pdA)8pA; lanes 2–6 elongation in the presence of montmorillonite; lane 7,elongation in the absence of montmorillonite.
MONTMORILLONITE CATALYSIS OF 30–50 MER OLIGONUCLEOTIDES 317
of the oligomers formed after 4 days using fresh samples of montmorillonite. Thechain lengths of oligomers formed in the presence of fresh clay to which ImpA andImpU were added for an additional 4 days were longer than those formed for in a8 day reaction using the same clay sample (Figure 3).
The loss of clay was eliminated by performing the elongation reactions in the0.45 micron insert in Gelman spin tubes. Preliminary tests confirmed that the clayparticles did not pass through the filter when a clay slurry was centrifuged at 14,000rpm and that the primer did not bind to the nylon filter. The elongation procedureused for the spin tubes was essentially the same as that for the microcentrifugetubes with the exception that the elongation reaction occurred in the filter tube in-sert and the clay phase was separated by the liquid phase by centrifugation with theclay remaining in the filter tube. The spin tubes were placed in a closed container,which had sufficient water to maintain a constant level of humidity to minimize theevaporative loss of water from the spin tubes. Elution of the oligomers from the clayusing 0.1 M Na4P2O7 required 15–40 min centrifugation. Use of the filter tubesmade it possible to add about 40 monomer units to the primer, 32pdA(pdA)8pA,to give 40–50 mers after 14 days reaction (Ferris et al., 1996). This elongation to50 mers was repeated using Pall Filtron tubes with a 0.45 micron filters since theGelman spin tubes are no longer available (Figure 4). The longer oligomers ob-tained using spin tubes (Figure 4) versus microcentrifuge tubes (Figure 2) suggeststhat the shorter chain lengths using microcentrifuge tubes reflect losses of the claycatalyst during the removal of the supernatant by pipetting.
There is an upper limit of elongation to 40–50 mers using the spin tubes (Fig-ure 4). The use of 4 instead of the usual 2 mg of Na+-montmorillonite resultedin less elongation after a 3, 6, and 9 day reactions (lanes 1–6) but comparablechain lengths were observed after a total of 12 days reaction (lanes 7, 8). The sameelongation limit was attained in other studies (data not shown) by adding fresh clayto the reaction after 3 days and after 3 and 6 days and carrying out the elongationreactions for 12 days. Since the oligonucleotide elongation with ImpA did notproceed past the 40–50 mers using more clay or with the addition of fresh clay, thelimit on the extent of elongation is probably not due to an inhibitor that is formedin the course of the elongation reaction. The addition of fresh clay would have atleast partially overcome the effect of inhibitors formed in the reaction mixture.
3.2. ELONGATION OF 32pdA(pdA)8pA WITH ImpU
It was only possible to add about 20 U monomer units to 32pdA(pdA)8pA in a6 day elongation time using spin tubes and no further elongation was observed onextended elongation to 10 days (Figure 5). Decomposition of the oligomers appearsto be the main reaction in the 8–10 day time period.
318 JAMES P. FERRIS
Figure 3. Role of fresh montmorillonite on elongation of 32pdA(pdA)8pA in microcentrifuge tubes.Lane 1, 32pdA(pdA)8pA; lane 2, elongation with ImpA for 2 days; lane 3, elongation for 8 days; lane4, elongated for 4 days and product eluted from montmorillonite and bound to fresh montmorilloniteand elongated for 4 days. Lane 5, 32pdA(pdA)8pA; lane 6, elongated for 4 days with ImpU; lane 7,elongated for 8 days; lane 8, elongated for 4 days, eluted from montmorillonite and bound to freshmontmorillonite and elongated 4 days.
MONTMORILLONITE CATALYSIS OF 30–50 MER OLIGONUCLEOTIDES 319
Figure 4. Gel electrophoresis of the elongation products of 32pdA(pdA)8pA using ImpA with 2 and4 mg of montmorillonite catalyst in Pall filter spin tubes.
3.3. DECOMPOSITION STUDIES
An investigation of the stability of the oligomers formed by the addition of U andA monomer units to 32pdA(pdA)8pA was performed to determine the relative ratesof decomposition. Oligomers containing up to ∼20 mers, formed by 2 elongationcycles of ImpA and ImpU to the primer in microcentrifuge tubes, were bound toNa+-montmorillonite for 5 and 10 days and the products were analyzed by gelelectrophoresis (Figures 6 and 7). Visual observation of the gels suggested thatboth oligomers decomposed at the same rate when bound to clay. In the case ofthe decomposition of the primer with A units added, the decomposition appearedto be greater with clay present than with it absent (Figure 6, compare lanes 3and 5). Semi-quantitative analysis of the extent of decomposition was obtainedby excising the ‘10 mer; and the band moving faster than the ‘10 mer’ band (‘9mer’) individually from the gel. The remaining bands on the gel were extracted
320 JAMES P. FERRIS
Figure 5. Gel electrophoresis of elongation products of 32pdA(pdA)8pA with ImpU using spin tubes.Lane 6 32pdA(pdA)8pA.
MONTMORILLONITE CATALYSIS OF 30–50 MER OLIGONUCLEOTIDES 321
Figure 6. Monitoring the decomposition of oligomers elongated with ImpA. Lane1, 32pdA(pdA)8pA; lane 2, elongation of 32pdA(pdA)8pA with ImpA for 2 days; lane 3, 2day elongation product dissolved in buffer-electrolyte, in the absence of montmorillonite, for 10days; lane 4, oligomers formed in 2 days bound to the montmorillonite stand for 5 days in thebuffer-salt reaction mixture; lane 5, oligomers formed in 2 days bound to the montmorillonite for 10days in buffer-electrolyte reaction mixture.
322 JAMES P. FERRIS
Figure 7. Monitoring the decomposition of oligomers elongated with ImpU. Lane1, 32pdA(pdA)8pA; lane 2, elongation of 32pdA(pdA)8pA with ImpU for 2 days; lane 3,oligomers formed in 2 days bound to the montmorillonite for 5 days in buffer-electrolyte reactionmixture; lane 4, oligomers formed in 2 days bound to the montmorillonite for 10 days inbuffer-electrolyte reaction mixture.
MONTMORILLONITE CATALYSIS OF 30–50 MER OLIGONUCLEOTIDES 323
TABLE I
Decomposition of Adenylate and Uridylate Elongation Products ofof 32pdA(pdA)8pA on Montmorillonitea
Adenylate Uridylate
Time 32P (%) Change (%) 32P (%) Change (%)
Initial –
9 mer 4.3 – 4.5 –
10 mer 9.5 5.6 –
5 Days
9 mer 9.1 4.8 8.2 3.7
10 mer 12.3 2.8 8.2 2.6
10 Days
9 mer 7.3 3.0 9.6 5.1
10 mer 12.8 3.3 9.4 3.8
a The extent of hydrolysis of the adenylate and uridylate adductsof the primer when bound to montmorillonite was determined fromthe gels shown in Figures 6 and 7, respectively. The fastest mov-ing band (9 mer) and next fastest moving band (10 mer) lanes 2,4 and 5 in Figure 6, were each eluted from the gel. The structuresof the 9 mer and 10 mer are assumed to be 32pdA(pdA)8pAp andAp32pdA(pdA)8pAp, respectively. The initial counts in from lane 2were subtracted from the counts for the 9 mer and 10 mer in lanes 4and 5. The percentage of 9 mer ands 10 mer was calculated fromthe total counts in lanes 4 and 5. The same procedure was usedto calculate the percent decomposition of the uridylate oligomersshown in Figure 7 using lanes 2, 3 and 4.
together and each of the three eluates was counted. These data were used to de-termine the percent decomposition of the oligomers. Structures 32pdA(pdA)8pApand Np32pdA(pdA)8pAp (N = A or U) were assigned to the 9 mer and 10 mer,respectively on the basis of their migration distances relative to that of the primer.The 9 – and 10 – mer bands increase at about 3–5% for both the A and the Uaddition products when bound (Table I). The absence of a significant difference inthe rates between the oligomers of A and U indicates that the shorter chain lengthsformed in the Na+-montmorillonite catalyzed addition of ImpU is not due to themore rapid decomposition of the U oligomers.
3.4. CHARACTERIZATION OF ELONGATION PRODUCTS
The oligomers were characterized by hydrolysis with alkaline phosphatase (APH)and ribonuclease T2 (RNase T2) hydrolysis. APH was used to differentiate those
324 JAMES P. FERRIS
oligomers in which the 5′-terminal phosphate group was capped by the additionof an 5′-Ap grouping to give Ap32pdA(pdA)8pAp(N)n and those which are notcapped. Those oligomers that are not capped disappear from the gel after hydrolysis(Figure 8). The primer (lane 6) does not give a band while the bands in lanes 3 and5 reveal those oligomers with 5′-caps. The gel shows that a substantial number ofthe oligomers formed by reaction with ImpA are capped (lanes 3 and 5 for 2 and8 days, respectively). The extent of capping after 2 and 8 days was estimated byexcising lanes 2 and 3 and 4 and 5 and measuring their radioactivity counts. Fromthe ratios of 3:2 and 5:4 it was determined that 34 and 63% of the oligomers werecapped after 2 and 8 days of elongation.
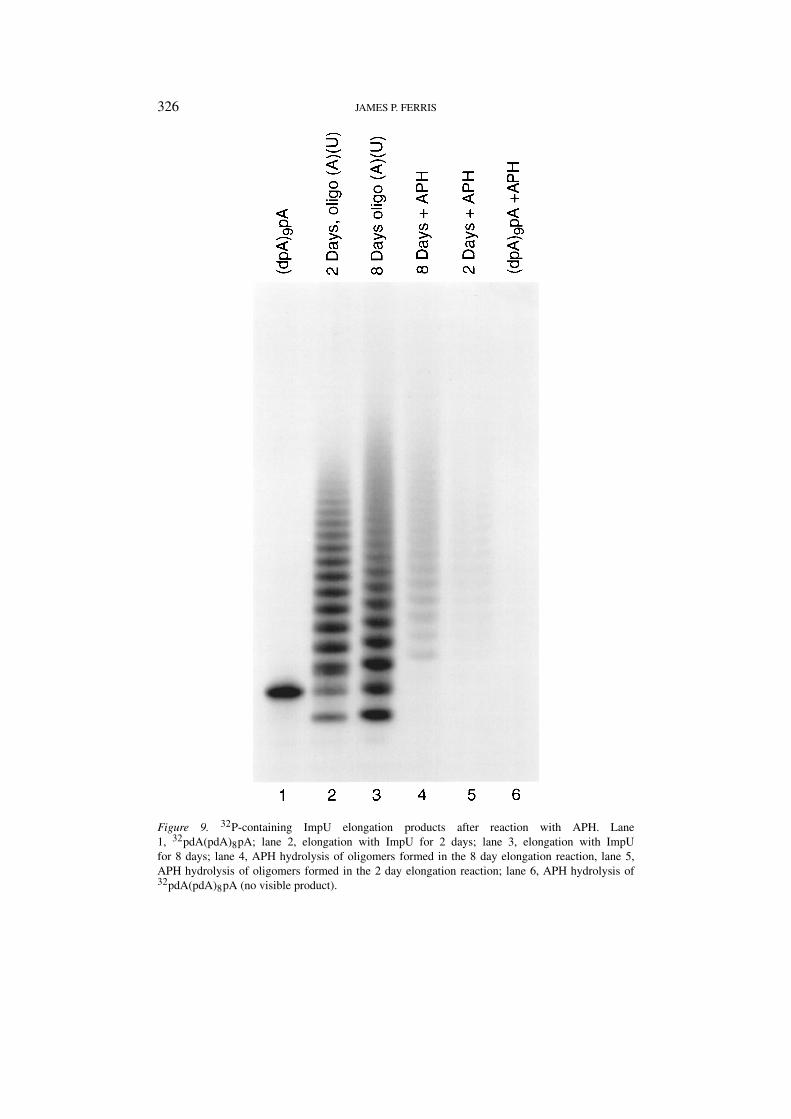
The same study was performed using the elongation products obtained withImpU (Figure 9). Here the bands remaining from the APH hydrolysis productswere much fainter and it was determined that the percent of capped oligomers after2 and 8 days of elongation was 14 and 24 %, respectively.
The types of phosphodiester bonds formed between the primer and oligomersbound to it were determined by RNaseT2 hydrolysis and the subsequent APH hy-drolysis of the oligomers formed after 2 elongation cycles. The hydrolysis productsthat contain P-32 were separated by gel electrophoresis (Figures 10 and 11). Areaction scheme consistent with the electrophoretic data of the hydrolysis productsof the ImpA addition reaction is given in Figure 12. The fastest moving band inlane 3 of Figure 10 was shown to be Ap32pdA(pdA)8pA by comparison with anauthentic sample. The band in lane 2 of Figure 10, migrating faster than the primer(lane 1), was assigned to the 3′-phosphorylated primer structure formed by thehydrolysis of the 3′, 5′-linked bond between the primer and the nucleotide boundto it at the 3′-position. Similar reasoning was used to deduce the structures of theother products in each band in the gels from the hydrolysis products.
The percentages of some of the bonds formed between the primer and themonomers was obtained by measurement of the areas of the bands in the RNaseT2 and APH hydrolyses hydrolysis products in the autoradiograms using the NIHImage program. The data derived from these measurements (Table II) for the higherpercent of 3′, 5′-phosphodiester bond formation is consistent with previous ob-servations with short oligoadenylates where it was observed that the reaction ofadenosine nucleotides yield a greater proportion of 3′, 5′-phosphodiester bondsthan do uridine nucleotides (Ferris and Ertem, 1993; Ding et al., 1996; Ertem andFerris, 2000).
4. Discussion and Conclusions
The addition products of the Na+-montmorillonite-catalyzed reaction of ImpA tothe 10 mer primer yields products with chain lengths as long as 40–50 mers. Oli-gomers as long as 30 mers are formed from the addition of ImpU to the sameprimer. Oligomers that contain 5′-caps are present, with a greater number of these
MONTMORILLONITE CATALYSIS OF 30–50 MER OLIGONUCLEOTIDES 325
Figure 8. 32P-containing ImpA elongation products after reaction with APH. Lane1, 32pdA(pdA)8pA; lane 2, elongation with ImpA for 2 days; lane 3, APH hydrolysis ofoligomers formed in the 2 days elongation reaction, lane 4, elongation with ImpA for 8 days; lane5, APH hydrolysis of oligomers formed in the 8 day elongation reaction; lane 6, APH hydrolysis of32pdA(pdA)8pA (no visible product).
326 JAMES P. FERRIS
Figure 9. 32P-containing ImpU elongation products after reaction with APH. Lane1, 32pdA(pdA)8pA; lane 2, elongation with ImpU for 2 days; lane 3, elongation with ImpUfor 8 days; lane 4, APH hydrolysis of oligomers formed in the 8 day elongation reaction, lane 5,APH hydrolysis of oligomers formed in the 2 day elongation reaction; lane 6, APH hydrolysis of32pdA(pdA)8pA (no visible product).
MONTMORILLONITE CATALYSIS OF 30–50 MER OLIGONUCLEOTIDES 327
Figure 10. Reaction of ImpA elongation products with RNase T2 with the subsequent hydrolysiswith APH. Lane 1, elongation with ImpA for 2 days; lane 2, hydrolysis of the elongation product withRNase T2 lane 3, APH hydrolysis of the RNase T2hydrolysis products; lane 4, elongation with ImpAfor 8 days; lane 6, APH hydrolysis of the RNase T2 hydrolysis products; lane 7, 32pdA(pdA)8pA.
328 JAMES P. FERRIS
Figure 11. Reaction of ImpU elongation products with RNase T2 with the subsequent hydrolysiswith APH. Lane 1, 32pdA(pdA)8pA; lane 2 elongation with ImpU for 2 days; lane 3, hydrolysis ofthe elongation product with RNase T2; lane 4, APH hydrolysis of the RNase T2 hydrolysis products;lane 5, RNase T2 hydrolysis of the 8 day elongation products; lane 6, elongation with ImpU for 8days; lane 7, APH hydrolysis of the RNase T2 hydrolysis products.
MONTMORILLONITE CATALYSIS OF 30–50 MER OLIGONUCLEOTIDES 329
Figure 12. Structures of products of enzymatic hydrolysis of the ImpA elongation products after twoelongation cycles. Capped oligomers can elongate from either end of the primer but for simplicitythe isomers shown only have the nucleotides added to the 3′-end of the primer. Predicted migrationdistances based on 32pdA(pdA)8pA = 10 and Ap32pdA(pdA)8pAp ∼11.5 as standards. Two addi-tional negative charges from either standard results in migration one unit faster and one nucleosideadditional results in migration ∼1.5 units slower. It is not possible to resolve ∼0.5 unit differencesso the bands may consist of mixtures. For example in Figure 10 lane 2, band 1, migration distance =9; band 2, migration distance = 9.5–10; band 3, migration distance = 10.5–11; band 4, 11.5–12.
TABLE II
Oligomer binding to primer after two feeding cyclesa,b
ImpA Addition ImpU
Addition
(%) (%)
3′, 5′-phosphodiester bonds 37 33
Capped with one 2′, 5′-phosphodiester bond 15 7
Capped with two 2′, 5′-phosphodiester bonds 6 3
a Data derived from percentages in Figures 10 and 11 and the experimentaldetermination of the percent capped oligomers for ImpA of 34% and ImpUof 14%.b Bonds directly to the primer before a 3, 5′-phosphodiester bond occurs inthe oligonucleotide chain.
330 JAMES P. FERRIS
caps on the oligomers formed from ImpA than from ImpU. It was possible todetermine the extent of 3′, 5′-and 2′, 5′-phosphodiester bonding to the primer but itwas not possible to determine the nature of the links in the oligomer chain that arenot bound to the primer. Extrapolation from studies on the formation of oligomersdirectly from ImpA and ImpU would predict that the oligomers formed from ImpAwould have a greater number of 3′, 5′-links and those from ImpU more 2′, 5′-links((Ferris and Ertem, 1993; Ding et al., 1996).
It was not possible to extend the elongation products formed from ImpA muchbeyond the 50 mers and those from ImpU beyond the 30 mers. This limit does notappear to be due to the inhibition of catalytic sites on the montmorillonite becausethe addition of fresh clay or the use of greater amounts of clay did not result in theformation of longer oligomers. A likely explanation is that 2′, 5′-links on the endof the growing chain add another nucleotide more slowly than the corresponding3′, 5′-phosphodiester bond (Ferris and Ertem, 1993). Since each elongation stepforms both 2′, 5′- and 3′, 5′-phosphodiester bonds chain elongation will proceedless effectively as the proportion of newly formed phosphodiester bonds decreases.
The slower rate of elongation of 2′, 5′-linkages may also explain why the ImpUelongation products are shorter than those formed from ImpA. Since ImpU tends toform more 2′, 5′-linkages (Ding et al., 1996), its elongation products will terminatesooner than those formed by ImpA addition to the primer.
The efficient elongation of short oligomers by their reaction with activatedmonomers on montmorillonite clay highlights the potential significance of min-eral catalysis to the origins of life (Ferris, 1993, 1985). While it is not known ifmontmorillonite catalysis initiated the RNA world, the mineral catalysis observedin this study and previous studies (Ferris et al., 1996) illustrates the potential role ofmineral catalysis for the formation of the biopolymers of the first life in an aqueousenvironment.
This route to oligonucleotides provides a prebiotic route to the RNA world viamineral catalysis. A mineral that binds and catalyzes the synthesis of biopolymersdownstream from a source of activated monomers could have served as the site forthe first living system. The RNAs formed on and bound to the surface of the min-eral could have interacted with each other to catalyze the ligation and replicationof RNA. The integrity of this simple system would have been maintained by thebinding of the longer RNAs to the mineral surface.
The generation of RNAs with chain lengths greater that 40 mers would havebeen long enough to initiate the first life on Earth (Joyce and Orgel, 1999; Szostakand Ellington, 1993). These oligomers would have been sufficiently long for replic-ation without loss of the core information of the RNA. The RNA would have alsobeen sufficiently long enough to fold into a three dimensional structure capable ofbinding and catalyzing the reactions of other RNA molecules.
MONTMORILLONITE CATALYSIS OF 30–50 MER OLIGONUCLEOTIDES 331
Acknowledgement
This research was initiated during a sabbatical leave at the Salk Institute and wascompleted at RPI. Dr. Leslie Orgel provided laboratory facilities and many helpfuland stimulating discussions. I thank Aubrey Hill, Jr. and Dr. Barbara Chu at Salkfor their expert guidance and help with many of the experimental procedures usedin this research. Derrick Meinhold and Charmi Miller at RPI measured the gelband areas in Figures 9 and 10 using NIH Image. The research at Salk was fundedby the NASA NSCORT/Exobiology Grant NAG5-4546 and NASA grant NAGW-2881 while the research at RPI was funded by National Science Foundation GrantCHE-0076281 and the NY Center for Studies on the Origins of Life funded byNASA Grant NAG5-7598 and NASA Exobiology grant NAG5-9607.
References
Ban, N., Nissen, P., Hansen, J., Moore, P. B. and Seitz, T. A.: 2000, The Complete Atomic Structureof the Large Ribosomal Subunit At 2.4 A Resolution, Science 289, 905–930.
Banin, A.: 1973, US Patent 3,725,528.Banin, A., Lawless, J. G., Mazzurco, J., Church, F. M., Margulies, L. and Orenberg, J. B.: 1985, Ph
Profile of the Adsorption of Nucleotides onto Montmorillonite, Origins Life Evol. Biosphere 15,89–101.
Cech, T. R., Zaug, A. J. and Grabowski, P. J.: 1981, In Vitro Splicing of the Ribosomal RNA Pre-cusor of Tetrahymena: Involvement of a Guanosine Nucleotide in the Excision of the InterveningSequence, Cell 27, 487–496.
Chu, B. C. F. and Orgel, L. E.: 1984, Preparation of Ligation Intermediates and Related Polynuc-leotide Pyrophosphates, Biochim. Biophys. Acta 782, 103–105.
Crick, F. H. C.: 1968, The Origin of the Genetic Code, J. Mol. Biol. 38, 367–379.Ding, Z. P., Kawamura, K. and Ferris, J. P.: 1996, Oligomerization of Uridine Phosphorimidazolides
on Montmorillonite: A Model for the Prebiotic Synthesis of RNA on Minerals, Origins Life Evol.Biosphere 26, 151–171.
Ertem, G. and Ferris, J. P.: 2000, Sequence- and Regio- Selectivity in the Montmorillonite-CatalyzedSynthesis of RNA, Origins Life Evol. Biosphere 30, 411–422.
Ertem, G. and Ferris, J. P.: 1997, Template-Directed Synthesis Using the Heterogeneous TemplatesProduced by Montmorillonite Catalysis. A Possible Bridge Between the Prebiotic and RNAWorlds, J. Am. Chem. Soc. 119, 7197–7201.
Ferris, J. P.: 1993, Catalysis and Prebiotic RNA Synthesis, Origins Life Evol. Biosphere 23, 307–315.Ferris, J. P. and Ertem, G.: 1993, Montmorillonite Catalysis of RNA Oligomer Formation in Aqueous
Solution. A Model for the Prebiotic Formation of RNA, J. Am. Chem. Soc. 115, 12270–12275.Ferris, J. P., Hill, A. R., Jr., Liu, R. and Orgel, L. E.: 1996, Synthesis of Long Prebiotic Oligomers
on Mineral Surfaces, Nature 381, 59–61.Ferris, J. P.: 1985, Minerals and Life’s Origins, Origins Life Evol. Biosphere 15, 75–76.Gilbert, W: 1986, The RNA World, Gilbert, Nature 319, 618.Guerrier-Takada, C., Gardiner, K., Marsh, T., Pace, N. and Altman, S.: 1983, The RNA Moiety of
Ribonuclease P is the Catalytic Subunit of the Enzyme, Cell 35, 849–857.Joyce, G. F. and Orgel, L. E.: 1999, Prospects for Understanding the Origin of the RNA World, in
The RNA World: The Nature of Modern RNA Suggests a Prebiotic RNA; Second Ed.; Atkins, J.F., Ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, New York, pp. 49–77.
332 JAMES P. FERRIS
Joyce, G. F.: 1998, Nucleic Acid Enzymes: Playing With a Fuller Deck, Proc. Natl. Acad. Sci. U.S.A.95, 5845–5847.
Joyce, G. F., Inoue, T. and Orgel, L. E.: 1984, Non-Enzymatic Template-Directed Synthesis on RNARandom Copolymers: Poly(C,U) Templates, J. Mol. Biol. 176, 279–306
Kawamura, K. and Ferris, J. P.: 1999, Clay Catalysis of Oligonucleotide Formation: Kinetics of theReaction of the 5′-Phosphorimidazolides of Nucleotides With the Non-Basic Heterocycles Uraciland Hypoxanthine, Origins Life Evol. Biosphere 29, 563–591.
Muth, G. W., Ortoleva-Donnelly, L. and Strobel, S. A.: 2000, A Single Adenosine With A Neutralpka in the Ribosomal Peptidyl Transferase Reaction Center, Science 289, 947–950.
Orgel, L. E.: 1968, Evolution of the Genetic Apparatus, J. Mol. Biol. 38, 381–393.Orgel, L. E.: 1998, The Origin of Life – A Review of Facts and Speculation, TIBS 23, 491–495.Prabahar, K. J. and Ferris, J. P.: 1997, Adenine Derivatives As Phosphate-Activating Groups for
the Regioselective formation of 3′, 5′-Linked Oligoadenylates On Montmorillonite: PossiblePhosphate-Activating Groups for the Prebiotic Synthesis of RNA, J. Am. Chem. Soc. 119,4330–4337.
Szostak, J. W. and Ellington, A. D.: 1993, In Vitro Selection of Functional RNA Sequences in TheRNA World. The Nature of Modern RNA Suggests A Prebiotic RNA World; Atkins, J. F., Ed.; ColdSpring Harbor Laboratory Press: Cold Spring Harbor, New York, 1993; Vol. Monograph 24.