1 Vitreous Zonule Morphology and accommodative function of the vitreous zonule in human and monkey eyes Lütjen-Drecoll, Elke 1 ; Kaufman, Paul 2,4 ; Wasielewski, Rainer 1 ; Ting-Li Lin 3 ; Croft, Mary Ann 2 1 Institute of Anatomy II, University of Erlangen-Nuremberg, 91054 Erlangen, Germany; 2 Department of Ophthalmology and Visual Sciences, University of Wisconsin, Madison, WI 53792; 3 Biostatistics and Medical Informatics, University of Wisconsin, Madison, WI 53792; and 4 Wisconsin National Primate Research Center, University of Wisconsin, Madison, WI 53792 Running title: Vitreous Zonule Function in Human and Monkey Eyes To whom correspondence should be addressed: Mary Ann Croft, MS Department of Ophthalmology and Visual Sciences University of Wisconsin Clinical Sciences Center 600 Highland Avenue Madison, WI 53792-3220 Telephone: 608-263-7738 Fax: 608-263-0543 Email: [email protected]Word Count 4434 Support: This work was funded in part by NEI grants RO1 EY10213 & R21EY018370 to PLK, the Ocular Physiology Research & Education Foundation; and DFG DR 124/7 to ELD. We also acknowledge the Wisconsin National Primate Research Center, University of Wisconsin-Madison base grant # 5P51 RR 000167 and the Core Grant for Vision Research grant # P30 EY016665. Page 1 of 46 IOVS IOVS Papers in Press. Published on October 8, 2009 as Manuscript iovs.09-4008 Copyright 2009 by The Association for Research in Vision and Ophthalmology, Inc.
Transcript
1 Vitreous Zonule
Morphology and accommodative function of the vitreous zonule in human and monkey eyes Lütjen-Drecoll, Elke1; Kaufman, Paul2,4; Wasielewski, Rainer1; Ting-Li Lin3; Croft, Mary Ann2 1Institute of Anatomy II, University of Erlangen-Nuremberg, 91054 Erlangen, Germany; 2Department of Ophthalmology and Visual Sciences, University of Wisconsin, Madison, WI 53792; 3Biostatistics and Medical Informatics, University of Wisconsin, Madison, WI 53792; and 4Wisconsin National Primate Research Center, University of Wisconsin, Madison, WI 53792
Running title: Vitreous Zonule Function in Human and Monkey Eyes To whom correspondence should be addressed:
Mary Ann Croft, MS Department of Ophthalmology and Visual Sciences University of Wisconsin Clinical Sciences Center 600 Highland Avenue Madison, WI 53792-3220 Telephone: 608-263-7738 Fax: 608-263-0543 Email: [email protected]
Word Count 4434
Support:
This work was funded in part by NEI grants RO1 EY10213 & R21EY018370 to
PLK, the Ocular Physiology Research & Education Foundation; and DFG DR
124/7 to ELD. We also acknowledge the Wisconsin National Primate Research
Center, University of Wisconsin-Madison base grant # 5P51 RR 000167 and the
Core Grant for Vision Research grant # P30 EY016665.
Page 1 of 46 IOVS IOVS Papers in Press. Published on October 8, 2009 as Manuscript iovs.09-4008
Copyright 2009 by The Association for Research in Vision and Ophthalmology, Inc.
In the resting eye, the distance between the scleral spur and the posterior
insertion zone of the posterior vitreous zonule (Fig. 10 A, B) did not significantly
change with age (Fig. 10 C). During accommodation, it could clearly be seen that
the posterior insertion zone of the posterior vitreous zonule moved forward (see
Video Clip 1, Fig. 10 A, B). Thus the distance between the scleral spur and the
posterior insertion zone of the posterior zonule diminished with accommodation
(p<0.001) and did so more markedly in the young eyes (1.052 ± 0.067 mm, n=4)
than in the older eyes (0.403 ± 0.118 mm, n=4) (Table 1, Fig. 10 D).
Results of the regression analysis showed that accommodative amplitude
(p=0.0003; not shown) and accommodative forward movement of the posterior
insertion zone (p=0.017) declined significantly with age (Fig. 10 D). The
accommodative forward movement of the posterior insertion zone declined by
~60% in the older vs. the young eyes (p=0.001; Table 1). Accommodative
amplitude increased significantly with increased accommodative forward
movement of the vitreous zonule’s posterior insertion zone (Fig. 10 E).
Page 16 of 46IOVS
17 Vitreous Zonule
The results of the stepwise regression analysis showed that age better predicted
accommodative amplitude than did either the forward movement of the posterior
insertion zone or the cleft width.
Disruption of the Intermediate Vitreous Zonule in Live Monkeys
Morphology
SEM showed that the intermediate vitreous zonular bundles in the region of the
-chymotrypsin injection were absent (Fig. 11). In all 4 rhesus (aged 6-9 and 25
years) independent of phakic status, at rest the muscle in the enzyme-injected
temporal quadrant was positioned more anteriorly than the temporal quadrant in
the contralateral controls (Fig. 12). This was true to a lesser degree for the single
young cynomolgus. For all 5 monkeys considered together, the anterior
translation was 0.256 ± 0.079 mm (n=5; p=0.032; Table 2). The difference in
apical position between treated vs. control eyes became far less pronounced as
the distance from the injection site increased (Table 2). The muscle tended to be
thicker in the -chymotrypsin injected eyes than the contralateral control eyes,
but the difference was not significant.
Ultrasound biomicroscopy in live monkeys
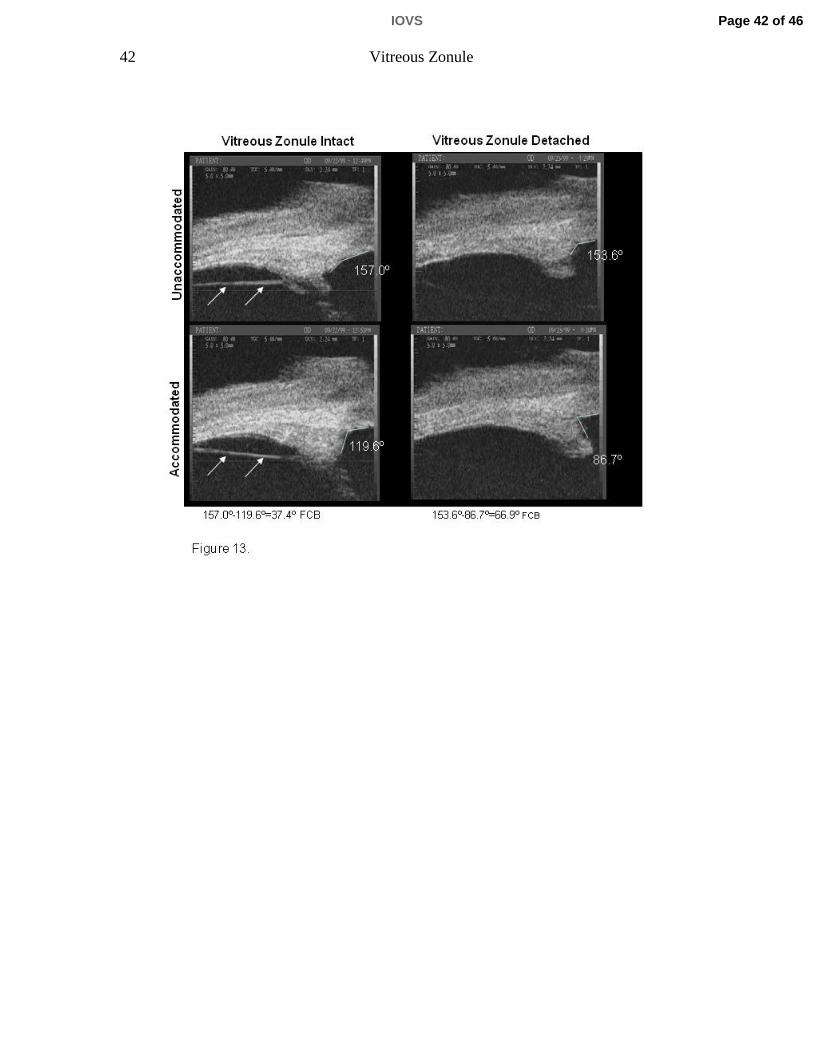
UBM imaging of the ciliary body in the temporal quadrant of 6 rhesus monkey
eyes (aged 6 to 26 years) before and after α-chymotrypsin lysis of the vitreous
zonule showed increased accommodative forward ciliary body movement post-
lysis vs. pre-lysis (p=0.044; Table 3, Fig. 13; see Video Clip 2). This was true for
Page 17 of 46 IOVS
18 Vitreous Zonule
phakic eyes, pseudophakic eyes with intact posterior capsule, and the one truly
aphakic eye without capsule. The latter eyes typically exhibited markedly
decreased accommodative ciliary body movement when the intermediate
vitreous zonule was intact. [21]
Discussion
This study describes the complexity of a vitreous zonular system that helps to
smoothly translate to the lens the driving forces for accommodation and
disaccommodation generated by ciliary muscle contraction and relaxation. Our
study also suggests how that structure allows and assists the very fine and rapid
refocusing and maintenance of focus at multiple distances required by primate
visual needs. [26]
In relation to the Helmholtz theory, [27] accommodation requires both centripetal
as well as forward ciliary body movement, and the age-related loss of
accommodation may be related to the loss of forward ciliary body movement. In
relation to the Coleman (catenary) theory of accommodation, which suggests that
accommodative shape changes in the lens are induced by vitreous pressure, [28-
30], age-related loss of ciliary body movement combined with increased lens size
[4, 31, 32] might alter the central catenary shape and thereby reduce
accommodative amplitude. We prefer to not go beyond what we have just stated
above, in order to avoid overspeculation with respect to the various theories of
Page 18 of 46IOVS
19 Vitreous Zonule
accommodation. We have opted to simply publish the findings and have provided
some discussion as to their meaning.
This study unveils the potential for novel mechanisms and suggests the need to
establish methods to include the vitreous membrane and vitreous zonule, along
with the anterior and posterior zonular complexes, in future finite element models
of the primate accommodation system. This manuscript also raises several
interesting questions about the future of anti-presbyopia surgery and treatment.
The presence of vitreous strands in living humans has been observed earlier by
Coleman [33] and in monkeys by Glasser et. al. [18] using ultrasound
biomicroscopy. However, neither the strands themselves nor their anatomic
relationships to other structures were definitively identified and characterized.
Indeed, some researchers have labeled the vitreous strands simply as anterior
hyaloid membrane. [20]
With respect to forward ciliary body movement, the angle measured includes the
inner aspect of the cornea and the anterior aspect of the muscle apex (not
including the ciliary processes) which is quite distinct and easily measured in the
iridectomized eye. Forward ciliary body movement as measured by ultrasound
biomicroscopy does not discern the individual contributions of the different areas
of the muscle (i.e., longitudinal, circular, reticular muscle portions) and what
occurs in the different internal anatomic regions of the muscle cannot be
Page 19 of 46 IOVS
20 Vitreous Zonule
precisely quantified. Nonetheless, the technique is a reliable indicator of overall
forward ciliary body/muscle movement (i.e., muscle apex) [3] [21] based on the
following: 1) the angle change reflects the translocation of the innermost aspect
of the muscle (apex); 2) the change in forward ciliary body movement as
measured by UBM following removal of the posterior restriction is corroborated
by the morphological findings in this paper and historically[1, 2, 5]; 3) the
qualitative examination of the dynamic video images (Video Clip 1 and 2) show
the overall muscle movement is forward and inward without isolated pockets
remaining static (with the exception of the outermost anterior longitudinal portion
of the muscle which is anchored/fixed anteriorly at the scleral spur). Therefore,
the overall forward movement of the muscle (including the muscle apex) could be
inferred from the forward ciliary body movement measured by UBM, and the
inward movement of the circular portion of the muscle could be inferred from
gonioscopically measured inward ciliary process movement. Comparisons of
forward vs. centripetal/inward ciliary body movement (measured by UBM and
goniovideography respectively) in living monkey eyes are made elsewhere and
are beyond the scope of this paper. [34]
There are only two regions in which the vitreous membrane is firmly attached to
structures of the accommodative system over the entire circumference: anteriorly
at the peripheral posterior lens capsule and posteriorly in the region of the ora
serrata. The intervening separation allows for the movement of the zonular
system needed for accommodation and disaccommodation. However, there are
Page 20 of 46IOVS
21 Vitreous Zonule
three other less-fixed fibrillar attachments, comprising the vitreous zonule, that
may stabilize and smooth the movement of the entire system and allow for finer
accommodative control and reduced stress on the ora serrata and the lens
capsule. This vitreous zonule, visualized by our improved SEM and UBM
techniques, is surprisingly robust and is intimately related to the anterior,
intermediate, and posterior vitreous membrane.
Our findings concerning the anterior vitreous zonule add new information to that
described previously for humans [9-14, 15 , 16, 17] and to goniovideography
observations previously reported for monkeys. [23] The earlier goniovideography
study included a video that showed the zonular attachments between the valleys
of the ciliary processes and Wieger’s ligament in vivo in the resting eye and
during accommodation. [23] The current SEM investigations indicate that, at most
places, the anterior vitreous membrane and posterior zonular tines of the anterior
zonular fork attaching to the posterior lens capsule are separated from each
other, presumably allowing unrestricted movement of the system. There are,
however, strands of anterior vitreous zonule that connect the zonular plexus with
the anterior vitreous membrane. This anterior vitreous zonule may help to
stabilize the anterior vitreous and thereby the position of the lens during
accommodation.
The cleft between the pars plana zonule of the ciliary body and the vitreous
membrane is bridged by bands of intermediate vitreous zonule fibers. The cleft
Page 21 of 46 IOVS
22 Vitreous Zonule
could provide a low-friction interface between the ciliary epithelium/pars plana
zonule and the vitreous membrane when the ciliary muscle moves anteriorly and
posteriorly during accommodation and disaccommodation, increasing the
efficiency of the system. The intermediate vitreous zonule may also stabilize the
vitreous membrane in its optimal contour, while dampening and smoothing the
muscle’s forward and inward movement.
The posterior vitreous zonule with its adjacent vitreous membrane, the pars
plana zonule, and the inner limiting membrane of the ciliary epithelium are joined
in an attachment zone of about 0.5 mm in monkeys and an astonishing 3-4 mm
in humans in anterior-posterior length, extending circumferentially around the eye
in the region of the ora serrata. This loose fibrillar attachment of the several
structures may also permit the forward-inward movement of the ciliary
muscle/zonular vitreous system in a dampened manner so as not to acutely
overstretch the peripheral retina and its attachment at the ora. This would
minimize the risk of tearing the peripheral retina upon accommodative effort. The
greater length and thickness of this zone in humans compared to monkeys may
reflect the need for protecting the retina over the much longer human lifespan.
With age, there is widening of the space between the ciliary body and vitreous
membrane and reduction of forward movement of the posterior insertion zone.
This could restrict forward movement of the zonular plexus upon accommodative
effort. However, our regression analysis showed that age itself was more
Page 22 of 46IOVS
23 Vitreous Zonule
important in predicting accommodative amplitude than was either the cleft width
or the forward movement of the posterior insertion zone. This is not surprising
given that there are likely other age-related changes occurring in the eye
concurrently that also affect accommodation.
Nonetheless, consistent with the hypothesis of age-related posterior restriction of
ciliary muscle/zonular movement, lysis of the posterior vitreous zonular system in
a few eyes increased the forward movement of the ciliary muscle upon central
electrical or peripheral pharmacological stimulation. While not a clinically
applicable technique, the findings and interpretations further support age-related
posterior restriction of ciliary muscle and zonular movement as both a
pathophysiologic factor and a potential therapeutic target in presbyopia.
Acknowledgements: The authors would like to thank Dr. Johannes Rohen for his immense help in preparing the specimens for SEM investigations, Elke Kretzschmar and Marco Gößwein for their technical assistance.
References 1. Lütjen-Drecoll, E., E. Tamm, and P.L. Kaufman, Age-related loss of
morphologic responses to pilocarpine in rhesus monkey ciliary muscle. Archives of Ophthalmology, 1988. 106: p. 1591-1598.
Page 23 of 46 IOVS
24 Vitreous Zonule
2. Tamm, E., M.A. Croft, W. Jungkunz, E. Lütjen-Drecoll, and P.L. Kaufman, Age-related loss of ciliary muscle mobility in the rhesus monkey: role of the choroid. Archives of Ophthalmology, 1992. 110: p. 871-876.
3. Croft, M., Glasser A, Heatley G, McDonald J, Ebbert T, Kaufman PL, The relationship between accommodative ciliary body and lens function in rhesus monkeys. I. Normal lens, zonule and ciliary process configuration in the iridectomized eye. Investigative Ophthalmology and Visual Science, 2006. 47(3): p. 1076-1086.
4. Strenk, S.A., J.L. Semmlow, I.M. Strenk, P. Munoz, J. Gronlund-Jacob, and J.K. DeMarco, Age-related changes in human ciliary muscle and lens: a magnetic resonance imaging study. Investigative Ophthalmology and Visual Science, 1999. 40: p. 1162-1169.
5. Poyer, J.F., P.L. Kaufman, and C. Flügel, Age does not affect contractile response of the isolated monkey ciliary muscle to muscarinic agonists. Current Eye Research, 1993. 12: p. 413-422.
6. Lütjen-Drecoll, E., E. Tamm, and P.L. Kaufman, Age changes in rhesus monkey ciliary muscle: Light and electron microscopy. Experimental Eye Research, 1988. 47: p. 885-899.
7. Gabelt, B.T., P.L. Kaufman, and J.R. Polansky, Ciliary muscle muscarinic binding sites, choline acetyltransferase and acetylcholinesterase in aging rhesus monkeys. Investigative Ophthalmology and Visual Science, 1990. 31: p. 2431-2436.
8. Tamm, E., E. Lütjen-Drecoll, W. Jungkunz, and J.W. Rohen, Posterior attachment of ciliary muscle in young, accommodating old, presbyopic monkeys. Investigative Ophthalmology and Visual Science, 1991. 32(5): p. 1678-1692.
9. Wieger, G., Ueber Canalis Petiti und ein Ligamentum hyaloideocapsulare. Diss. Inaug. Strassbourg, 1883.
10. Eisner, G., Biomicroscopy of the peripheral fundus. Heidelberg, Springerverlag, 1973.
11. Vail, D., The zonula of Zinn and ligament of Wieger. Transactions of the Ophthalmological Societies of the United Kingdom, 1957. 77: p. 441-499.
12. Eisner, G., Clinical examination of the vitreous. Transactions of the Ophthalmological Societies of the United Kingdom, 1975. 95: p. 360-363.
13. Streeten, B.W. and J.O. Pulaski, Posterior zonules and the lens extraction. Archives of Ophthalmology, 1977. 96: p. 132-138.
14. Bernal, A., J.M. Parel, and F. Manns, Evidence for posterior zonular attachment on the anterior hyaloid membrane. Investigative Ophthalmology and Visual Science, 2006. 47(11): p. 4708-4713.
15. Salzmann, M., Anatomie und Histologie des menschlichen. Augapfels. Wein, Franz Deuticke, 1912.
16. Eisner, G., Clinical anatomy of the vitreous. In: Tasman W, Jaeger EA,ed. /Duane's foundations of clinical ophthalmology. 2005, Philadelphia: Lippincott Williams & Wilkins. 34.
Page 24 of 46IOVS
25 Vitreous Zonule
17. Sebag, J., ed. Vitreous-from biochemistry to clinical relevance. In: Tasman W, Jaeger EA,ed. /Duane's foundations of clinical ophthalmology. 1998 ed. Vol. 1. 1998, Lippincott Williams & Wilkins: Philadelphia. 21.
18. Glasser, A., M.A. Croft, L. Brumback, and P.L. Kaufman, Ultrasound biomicroscopy of the aging rhesus monkey ciliary region. Optometry and Vision Science, 2001. 78: p. 417-424.
19. Rohen, J.W., Scanning electron microscopic studies of the zonular apparatus in human and monkey eyes. Investigative Ophthalmology and Visual Science, 1979. 18: p. 133-144.
20. Byrne, S.F. and R.L. Green, Ultrasound of the eye and orbit. Second Edition. ed. 2002, St. Louis, MO: Mosby Inc.
21. Wasilewski, R., J.P. McDonald, G. Heatley, E. Lütjen-Drecoll, P.L. Kaufman, and M.A. Croft, Surgical intervention and accommodative responses, II. Forward ciliary body accommodative movement is facilitated by zonular attachments to the lens capsule. Investigative Ophthalmology and Visual Science, 2008. 49(12): p. 5495-5502.
22. Ito, S. and M.J. Karnovsky, Formaldehyde-glutaraldehyde fixatives containing trinitro compounds. Journal of Cell Biology, 1968. 39: p. 168a.
23. Croft, M.A., J.P. McDonald, R.J. James, G.A. Heatley, T.L. Lin, E. Lütjen-Drecoll, and P.L. Kaufman, Surgical intervention and accommodative responses, I: Centripetal ciliary body, capsule, and lens movements in rhesus monkeys of various ages. Investigative Ophthalmology and Visual Science, 2008. 49(12): p. 5484-5494.
24. Crawford, K., E. Terasawa, and P.L. Kaufman, Reproducible stimulation of ciliary muscle contraction in the cynomolgus monkey via a permanent indwelling midbrain electrode. Brain Research, 1989. 503: p. 265-272.
25. Akaike, H., A new look at the statistical model identification. IEEE Transactions on Automatic Control, 1974. 19(6): p. 716-723.
26. May, C.A., N. Neuhuber, and E. Lütjen-Drecoll, Immunohistochemical Classification and Functional Morphology of Human Choroidal Ganglion Cells. Invest. Ophthalmol. Vis. Sci., 2004: p. 361-367.
27. von Helmholtz, H.H., Helmholtz's treatise on physiological optics, in Mechanism of accommodation., J.P.C. Southall, Editor. 1909, Dover Publications: New York. p. 143-172.
28. Coleman, D.J., Unified model for accommodative mechanism. American Journal of Ophthalmology, 1970. 69: p. 1063-1079.
29. Coleman, D.J., On the hydraulic suspension theory of accommodation. Transactions of the American Ophthalmological Society, 1986. 84: p. 846-868.
30. Coleman, D.J. and F. S.K., Presbyopia, accommodation, and the mature catenary. Ophthalmology, 2001. 108(9): p. 1544-51.
31. Wendt, M., Croft MA, McDonald J, Kaufman PL, Glasser A, Lens diameter and thickness as a function of age and pharmacologically stimulated accommodation in rhesus monkeys. Experimental Eye Research, 2008. 86: p. 746-752.
Page 25 of 46 IOVS
26 Vitreous Zonule
32. Rafferty, N.S., Structure, function, and pathology, in The ocular lens, H. Maisel, Editor. 1985, Marcel Dekker: New York. p. 1-60.
33. Coleman, D.J., R.H. Silverman, F.L. Lizzi, D.Z. Reinstein, M.J. Rondeau, H.O. Lloyd, and S.W. Daly, Ultrasonography of the Eye and Orbit. 2nd ed. 2006, Philadelphia: Lippincott Williams & Wilkins.
34. Croft, M.A., J.P. McDonald, N.V. Nadkarni, T.L. Lin, and P.L. Kaufman, Age-related changes in centripetal ciliary body movement relative to centripetal lens movement in monkeys. Experimental Eye Research, 2009. In Press.
Page 26 of 46IOVS
27 Vitreous Zonule
Figure legends Figure 1. Ultrasound Biomicroscopy (UBM) overview image (E-Tech) in a live
rhesus monkey shows a prominent straight line (arrow) extending from the pars
plicata region of the ciliary body to the ora serrata region, and separated from the
pars plana epithelium by a cleft, CP=ciliary processes, CB=ciliary body.
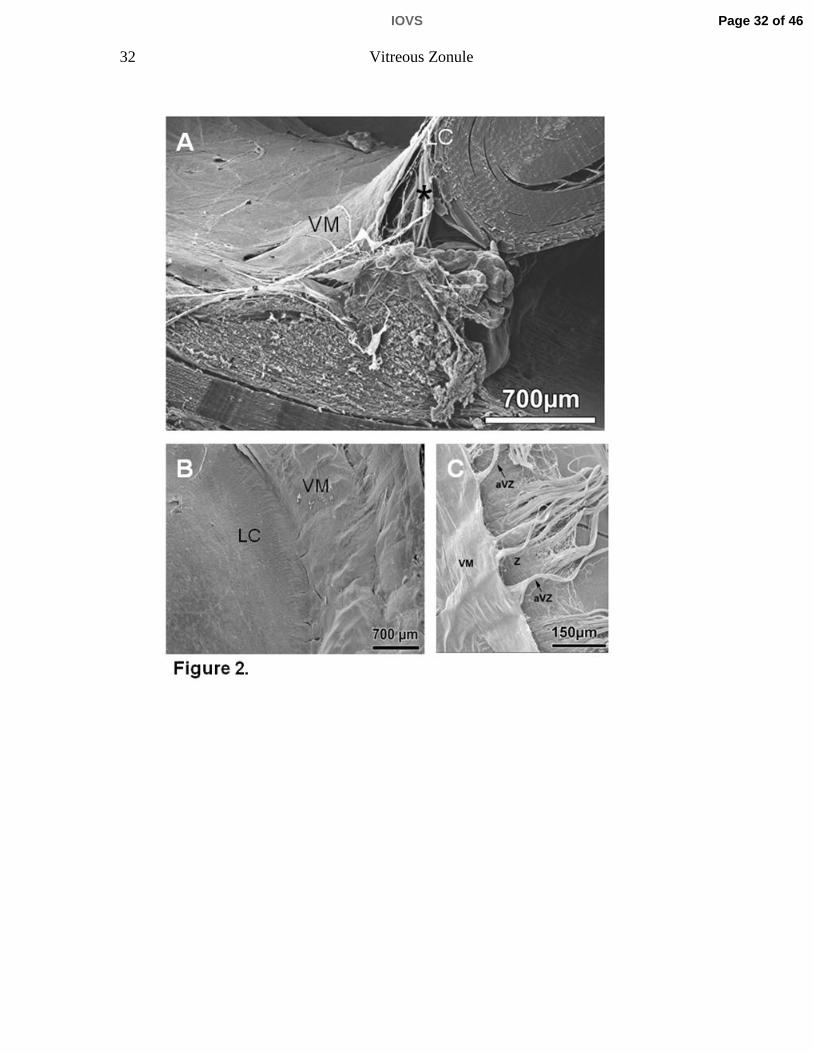
Figure 2. A 75-year-old human eye. A) Scanning electron micrograph of a sagittal
aspect of the anterior vitreous membrane (VM), the posterior tine of the zonular
fork (asterisk), and the posterior lens capsule (LC). B) Scanning electron
micrograph of the anterior VM and its attachment to the posterior LC. C) Careful
lifting of the VM reveals that the majority of the zonule (Z) inserts directly into the
posterior LC without attachment to the vitreous membrane that itself forms a
separate layer also directly inserting into the posterior LC. However, some
bundles of zonular fibers (anterior vitreous zonule, aVZ) do insert into the anterior
VM at spatial intervals of about 75 µm.
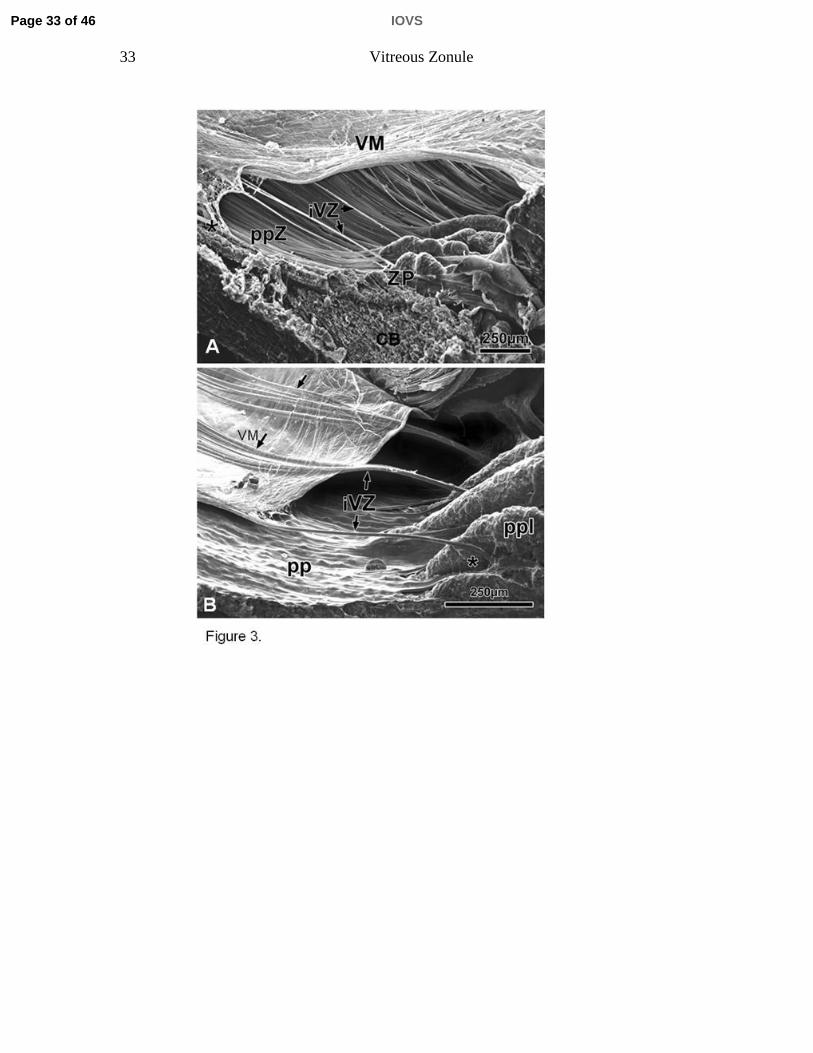
Figure 3. A) Scanning electron micrographs of a sagittal and oblique internal view
of the ciliary body (CB), the vitreous membrane (VM), the zonular plexus (ZP),
and the zonular connections bridging the cleft between the pars plana zonules
(ppZ) and VM (rhesus monkey, aged 8 years). The bridging bundles of zonular
fibers run from the region of the ZP in the valleys of the posterior pars plicata
towards the VM in the region of the ora serrata (asterisk). These zonular bundles
were termed intermediate vitreous zonule (iVZ). B) Scanning electron micrograph
showing the inner aspect of the posterior pars plicata (ppl), pars plana (pp), VM
and iVZ of a 10-year-old rhesus monkey. Anteriorly, the intermediate vitreous
zonular bundles split into a fork whose tines insert on both sides of the processes
in the valleys of the ppl (asterisk). Posteriorly, each main bundle splits into
several smaller bundles that merge with the VM (arrows).
Page 27 of 46 IOVS
28 Vitreous Zonule
Figure 4. A) Scanning electron micrographs of sagittal sections of the ora serrata
region (rhesus monkey, aged 6 years). The vitreous membrane (VM), the
posterior extensions of the intermediate vitreous zonule (iVZ) and the pars plana
zonule form an interconnected sponge-like structure (asterisk). B) After careful
elevation of the VM, the connections of the ppZ to the posterior VM become
visible (arrows).
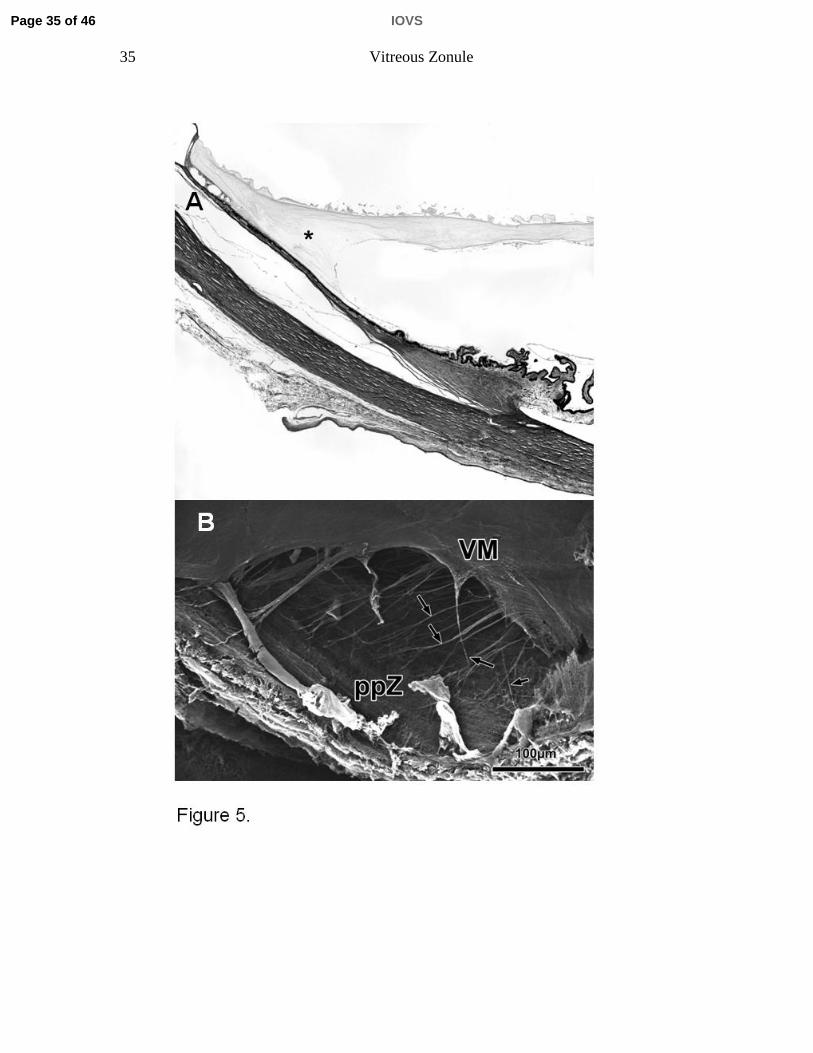
Figure 5. A) Histological sagittal section through a 100-year-old human eye
showing the posterior adherence of the vitreous membrane to the posterior pars
plana and ora region (asterisk). Note that this region in humans encompasses
nearly ½ the sagittal length between the scleral spur and ora serrata. B)
Scanning electron micrograph of the posterior vitreous zonule (sagittal aspect,
85-year-old human eye). Note the oblique-running fibrils between the pars plana
zonules (ppZ) and the vitreous membrane (VM); in contrast to the monkey (Fig. 4
B), these fibrils form a lattice-like structure (arrows).
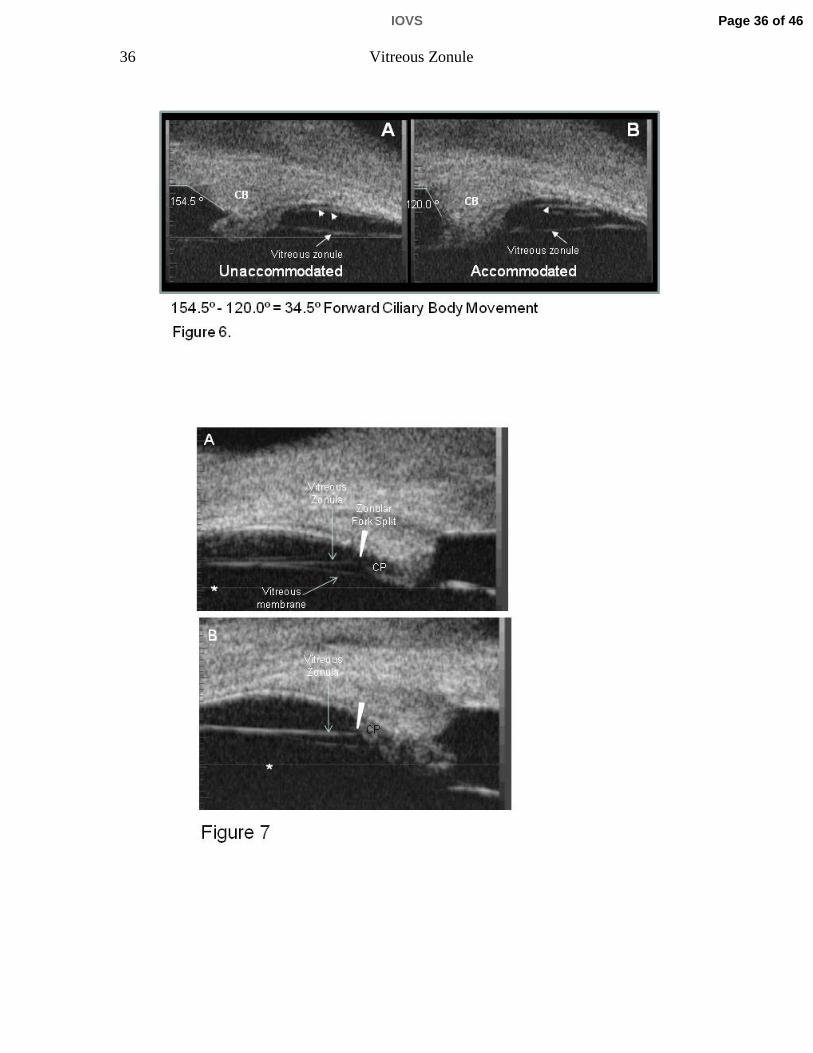
Figure 6 A, B. UBM images of unaccommodated (A) and accommodated (B)
ciliary muscle in live rhesus monkey, aged 25 years. In these somewhat oblique
sections, one can see the pars plana zonules immediately adjacent to the pars
plana epithelium (arrowheads). This is more apparent in the accommodated than
in the unaccommodated state. The straight line between the ciliary processes
and the ora serrata represents the vitreous zonule. The numbers represent the
angle between the anterior face of the ciliary body and the inner surface of the
peripheral cornea, as defined by the white lines. Narrowing of the ciliary body
(CB)-Cornea angle in the accommodated vs. the unaccommodated state was
used as a surrogate indicator of forward ciliary body movement.
Figure 7. Ultrasound biomicroscopy (UBM) images were obtained in a 15-year-
old rhesus monkey. Care was taken to assure that the vitreous zonule appeared
as a continuous prominent straight line parallel to the focus line (asterisk)
Page 28 of 46IOVS
29 Vitreous Zonule
imprinted by the instrument within all images, indicating that these were true
sagittal sections. A) Anteriorly, the vitreous membrane is separated from the
vitreous zonule. The vitreous zonule fork (seen by SEM, Fig. 3 B) occurs just
posterior to its insertion to the zonular plexus and, as the fibers split at the fork,
they course out of the UBM image plane (arrowhead). Thus the white line that
represents the vitreous zonule appears to discontinue (arrowhead) immediately
posterior to reaching the ciliary process region in this image. This “drop out” of
the vitreous zonule strand is not always apparent in UBM images, due to the
orientation of the UBM probe and the close proximity of the vitreous zonule fork
to the ciliary processes (CP), as in Panel B.
Figure 8. Ultrasound biomicroscopy (UBM) image (A) analogous to SEM section
(B), and also panel A of Fig. 3 reproduced for ease of comparison, from the same
8-year-old rhesus monkey. CB=ciliary body, pp=pars plana zonule, VZ=vitreous
Table 1. Data are mean ± s.e.m. (mm) accommodative forward movement of the intermediate posterior zonule’s posterior insertion zone during supramaximal central stimulation to induce accommodation in monkeys. The accommodative forward movement of the posterior insertion zone declined by ~60% in the older vs the
Table 1. Data are mean ± s.e.m. (mm) accommodative forward movement of the intermediate posterior zonule’s posterior insertion zone during supramaximal central stimulation to induce accommodation in monkeys. The accommodative forward movement of the posterior insertion zone declined by ~60% in the older vs the young monkey eyes. Young = 6 to 9, Older = 19 to 21 years old.
Page 43 of 46 IOVS
Table 2. Morphological Findings With and Without α-Chymotrypsin Treatment to Lyse the Vitreous Zonule in Monkey Eyes
A. Apical Position (mm) Muscle Width (mm)Age
Monkey (Years) Quadrant Temporal Inferior Superior Nasal Temporal Inferior Superior Nasaly ( ) p p p p1 cyno 3.50 Post Lysis 0.51 0.49 0.53 0.61 0.67 0.54 0.60 0.76
B Diff i A i l P iti ( ) Diff i M l Width ( )B. Differences in Apical Position (mm) Differences in Muscle Width (mm)
α-Chymotrypsin Treated minus Control Eye α-Chymotrypsin Treated minus Control EyeQuadrant Temporal Inferior Superior Nasal Temporal Inferior Superior Nasal
Table 2. Data are apical position and muscle width (A) and the differences in muscle apical position and muscle width between chymotrypsin treated and control monkey eyes (B) for the respective quadrants The eyes received Table 2. Data are apical position and muscle width (A) and the differences in muscle apical position and muscle width between ‐chymotrypsin‐treated and control monkey eyes (B) for the respective quadrants . The eyes received ‐chymotrypsin injection in the temporal quadrant to lyse the vitreous zonule (Post‐Lysis).The apical position of the muscle was farther anterior in ‐chymotrypsin injected eyes than in contralateral control eyes; the difference was most pronounced near the injection site (temporal quadrant; p=0.044, by two‐tailed paired t‐test). Overall the muscle widthtended to be thicker in the ‐chymotrypsin injected eyes than the contralateral control eyes but the change in width was not dependent upon proximity to injection site. NS=Not significant. Monkey #1 was a cynomolgus monkey and monkeys 2‐
Table 2. Data are apical position and muscle width (A) and the differences in muscle apical position and muscle width between ‐chymotrypsin‐treated and control monkey eyes (B) for the respective quadrants . The eyes received ‐chymotrypsin injection in the temporal quadrant to lyse the vitreous zonule (Post‐Lysis).The apical position of the muscle was farther anterior in ‐chymotrypsin injected eyes than in contralateral control eyes; the difference was most pronounced near the injection site (temporal quadrant; p=0.044, by two‐tailed paired t‐test). Overall the muscle widthtended to be thicker in the ‐chymotrypsin injected eyes than the contralateral control eyes but the change in width was not dependent upon proximity to injection site. NS=Not significant. Monkey #1 was a cynomolgus monkey and monkeys 2‐5 were rhesus monkeys. *Aphakic eye; **Pseudophakic eyes.
Page 45 of 46 IOVS
Table 3. Forward Ciliary Body Movement in Monkey Eyes as Measured by Ultrasound Biomicroscopy (degrees)Before and After α-Chymotrypsin
Post minus % FCB MovmentAge (Years) Monkey Pre α-Chymotrypsin Post α-Chymotrypsin Pre α-Chymotrypsin Increase
Table 3. Data are forward ciliary body (FCB) movement in six rhesus monkey eyes before and after ‐chymotrypsin injection. FCB was determined as the narrowing of the angle that is formed between the inner aspect of the cornea and the anterior aspect of the ciliary body in the accommodated versus the unaccommodated state measured in degrees (see Fi 13) Th i d h t i i j ti i th t l d t M t t k i th i
Table 3. Data are forward ciliary body (FCB) movement in six rhesus monkey eyes before and after ‐chymotrypsin injection. FCB was determined as the narrowing of the angle that is formed between the inner aspect of the cornea and the anterior aspect of the ciliary body in the accommodated versus the unaccommodated state measured in degrees (see Fig. 13). The eyes received ‐chymotrypsin injection in the temporal quadrant. Measurements were taken in the region of the injection site. FCB movement was significantly higher Post α‐chymotrypsin vs Pre (p= 0.044) by two‐tailed paired t‐test. Monkeys 1‐3 were stimulated to accommodate by central electrical stimulation while monkeys 4‐6 were stimulated to accommodate pharmacologically by carbachol iontophoresis. *Aphakic eye (see Materials and Methods); **Pseudophakic eyes).
Table 3. Data are forward ciliary body (FCB) movement in six rhesus monkey eyes before and after ‐chymotrypsin injection. FCB was determined as the narrowing of the angle that is formed between the inner aspect of the cornea and the anterior aspect of the ciliary body in the accommodated versus the unaccommodated state measured in degrees (see Fig. 13). The eyes received ‐chymotrypsin injection in the temporal quadrant. Measurements were taken in the region of the injection site. FCB movement was significantly higher Post α‐chymotrypsin vs Pre (p= 0.044) by two‐tailed paired t‐test. Monkeys 1‐3 were stimulated to accommodate by central electrical stimulation while monkeys 4‐6 were stimulated to accommodate pharmacologically by carbachol iontophoresis. *Aphakic eye (see Materials and Methods); **Pseudophakic eyes).