Proc. Nati. Acad. Sci. USA Vol. 89, pp. 3399-3403, April 1992 Neurobiology mRNA coding for neurotransmitter receptors in a human astrocytoma (Xenopus oocytes/acetylcholine/glutamate receptors/glioma) C. MATUTE*t, R. 0. ARELLANO*, B. CONDE-GUERRIt, AND R. MILEDI* *Laboratory of Cellular and Molecular Neurobiology, Department of Psychobiology, University of California, Irvine, CA 92717; and *Departamento de Ciencias Morfol6gicas, Universidad de Zaragoza, 50009-Zaragoza, Spain Contributed by R. Miledi, January 13, 1992 ABSTRACT Electrophysiological techniques and Xenopus oocytes were used to study the expression of neurotransmitter receptors encoded by mRNAs isolated from three human glioma cell lines. Oocytes injected with mRNAs from two glioblastoma cell lines did not show electrical responses to the various neurotransmitters tested. In contrast, oocytes in ected with mRNA from an astrocytoma cell line (R-1l1) acquired acetylcholine and glutamate receptors as well as a small number of N-methyl-D-aspartate (NMDA) receptors. Acetyl- choline elicited oscillatory Cl- currents that were abolished by muscarinic antagonists. The muscarnic receptors are coupled to the inositol phosphate-Ca2+ receptor-channel coupling sys- tem. Glutamate and its analogs kainate, quisqualate, and a-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid in- duced smooth currents. The non-NMDA responses were po- tently blocked by 6,7-dinitroquinoxaline-2,3-dione. Our results show that human astrocytoma cells contain mRNAs coding for functional acetylcholine and glutamate receptors that have properties similar to those of neurons. In contrast, human glioblastoma cells lacked those mRNAs. These differences might be useful for the development of new diagnostic and therapeutic procedures. Tumor cells of the brain, like other neoplastic cells, are known to release cytokines, growth-controlling factors, and other substances involved in cell regulation (1-3). It is, therefore, important to determine the pattern of receptors expressed in the membrane of tumor cells, as this may help us understand the way tumors develop and the way they modulate the immune system of the host. Such knowledge would also be very useful for the development of new diagnostic and therapeutic procedures. Electrophysiological studies have already shown that glial cells possess receptors to glutamate and other neurotrans- mitters (4-8). However, most of that work was done on murine cells in culture, and it is known that the procedures used to isolate the cells, as well as the culture process itself, can alter the types and properties of the receptors expressed (9, 10). Moreover, the receptors in murine cells might differ importantly from those present in human cells. It is thus necessary to study the human cells directly and, although human cells in vitro seem to undergo fewer genetic transfor- mations than murine cells, it would be preferable not to use cultured cells. Indeed, human brain tumors have been found to express some neurotransmitter receptors (12-15). How- ever, these studies used labeled ligand binding and autora- diography to characterize the receptors, and little informa- tion was gained about their functional properties. To avoid some of the problems mentioned above we decided to use a different approach to determine the presence of mRNAs coding for neurotransmitter receptors in human tumors borne in nude mice. To that effect, mRNA, isolated from the tumors, was injected into Xenopus oocytes, which have been shown to be a highly efficient system for express- ing various functional foreign receptors (16). The generation of a membrane current by a neurotransmitter, in an oocyte injected with tumor mRNA, signals the presence of the corresponding receptor mRNA in the tumor, and the ampli- tude of the current gives a relative measure of the amount of receptor mRNA in the sample (e.g., ref. 17). In this way we not only establish the presence of a neurotransmitter receptor mRNA in the tumor but we also gain some information on the functional characteristics of the receptor expressed. MATERIAL AND METHODS Establishment and Characterization of Tumor Cell Lines. Three human brain tumors were obtained at surgery. The three tumors (codes R-110, R-111, and R-162) were located in the parenquima of the parietal cerebral cortex: R-110 was removed from a 65-year-old female, R-111 from a 65-year-old male, and R-162 from a 68-year-old female. Part of the tumoral masses was used for histological examination. An- other part was dissociated and seeded into culture dishes in minimal essential medium supplemented with 10o fetal bo- vine serum, penicillin (100 units/ml), and streptomycin (100 ,ug/ml). After several passes in culture, proliferating cells were grown as solid tumors in nude mice. Standard immu- nofluorescence techniques were used to characterize the tumor cells. Briefly, cells were seeded on sterile poly(L- lysine)-coated coverslips overnight and then fixed with 95% ethanol/5% acetic acid for 15 min at 40C. Cells were incu- bated for 2 hr with rabbit antiserum to glial fibrillary acidic protein (GFAP), myelin basic protein (MBP), or vimentin, all diluted as suggested by the supplier, followed by several washes. Finally, cells were incubated with fluorescein iso- thiocyanate-labeled anti-rabbit antiserum at 1:100 for an additional hour, washed, and coverslipped. All antibody solutions and washings were made in sodium phosphate- buffered saline (145 mM NaCl/10 mM sodium phosphate, pH 7.4). Primary and secondary antibodies were purchased from Dakopatts (Glostrup, Denmark). RNA Extraction, Microinjection into Oocytes, and Electro- physiology. mRNA was extracted using the guanidinium/ phenol/chloroform method (18) followed by oligo(dT)- cellulose chromatography. A total of seven mRNA prepara- tions from three human glioma cell lines (codes R-110, R-111, and R-162) grown in nude mice was used in this study. Techniques for injection of mRNA and for electrophysiolog- Abbreviations: AcCho, acetylcholine; NMDA, N-methyl-D- aspartate; AMPA, a-amino-3-hydroxy-5-methyl-4-isoxazole propi- onic acid; CNQX, 6,7-dinitroquinoxaline-2,3-dione; GFAP, glial fibrillary acidic protein; MBP, myelin basic protein; I-V, current- voltage. tPresent address: Departamento de Neurociencias, Universidad del Pais Vasco, 48940-Leioa, Spain. 3399 The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Transcript
Proc. Nati. Acad. Sci. USAVol. 89, pp. 3399-3403, April 1992Neurobiology
mRNA coding for neurotransmitter receptors in ahuman astrocytoma
C. MATUTE*t, R. 0. ARELLANO*, B. CONDE-GUERRIt, AND R. MILEDI**Laboratory of Cellular and Molecular Neurobiology, Department of Psychobiology, University of California, Irvine, CA 92717; and *Departamento deCiencias Morfol6gicas, Universidad de Zaragoza, 50009-Zaragoza, Spain
Contributed by R. Miledi, January 13, 1992
ABSTRACT Electrophysiological techniques and Xenopusoocytes were used to study the expression of neurotransmitterreceptors encoded by mRNAs isolated from three humanglioma cell lines. Oocytes injected with mRNAs from twoglioblastoma cell lines did not show electrical responses to thevarious neurotransmitters tested. In contrast, oocytes in ectedwith mRNA from an astrocytoma cell line (R-1l1) acquiredacetylcholine and glutamate receptors as well as a smallnumber of N-methyl-D-aspartate (NMDA) receptors. Acetyl-choline elicited oscillatory Cl- currents that were abolished bymuscarinic antagonists. The muscarnic receptors are coupledto the inositol phosphate-Ca2+ receptor-channel coupling sys-tem. Glutamate and its analogs kainate, quisqualate, anda-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid in-duced smooth currents. The non-NMDA responses were po-tently blocked by 6,7-dinitroquinoxaline-2,3-dione. Our resultsshow that human astrocytoma cells contain mRNAs coding forfunctional acetylcholine and glutamate receptors that haveproperties similar to those of neurons. In contrast, humanglioblastoma cells lacked those mRNAs. These differencesmight be useful for the development of new diagnostic andtherapeutic procedures.
Tumor cells of the brain, like other neoplastic cells, areknown to release cytokines, growth-controlling factors, andother substances involved in cell regulation (1-3). It is,therefore, important to determine the pattern of receptorsexpressed in the membrane of tumor cells, as this may helpus understand the way tumors develop and the way theymodulate the immune system of the host. Such knowledgewould also be very useful for the development of newdiagnostic and therapeutic procedures.
Electrophysiological studies have already shown that glialcells possess receptors to glutamate and other neurotrans-mitters (4-8). However, most of that work was done onmurine cells in culture, and it is known that the proceduresused to isolate the cells, as well as the culture process itself,can alter the types and properties of the receptors expressed(9, 10). Moreover, the receptors in murine cells might differimportantly from those present in human cells. It is thusnecessary to study the human cells directly and, althoughhuman cells in vitro seem to undergo fewer genetic transfor-mations than murine cells, it would be preferable not to usecultured cells. Indeed, human brain tumors have been foundto express some neurotransmitter receptors (12-15). How-ever, these studies used labeled ligand binding and autora-diography to characterize the receptors, and little informa-tion was gained about their functional properties.To avoid some of the problems mentioned above we
decided to use a different approach to determine the presenceof mRNAs coding for neurotransmitter receptors in human
tumors borne in nude mice. To that effect, mRNA, isolatedfrom the tumors, was injected into Xenopus oocytes, whichhave been shown to be a highly efficient system for express-ing various functional foreign receptors (16). The generationof a membrane current by a neurotransmitter, in an oocyteinjected with tumor mRNA, signals the presence of thecorresponding receptor mRNA in the tumor, and the ampli-tude of the current gives a relative measure of the amount ofreceptor mRNA in the sample (e.g., ref. 17). In this way wenot only establish the presence ofa neurotransmitter receptormRNA in the tumor but we also gain some information on thefunctional characteristics of the receptor expressed.
MATERIAL AND METHODSEstablishment and Characterization of Tumor Cell Lines.
Three human brain tumors were obtained at surgery. Thethree tumors (codes R-110, R-111, and R-162) were located inthe parenquima of the parietal cerebral cortex: R-110 wasremoved from a 65-year-old female, R-111 from a 65-year-oldmale, and R-162 from a 68-year-old female. Part of thetumoral masses was used for histological examination. An-other part was dissociated and seeded into culture dishes inminimal essential medium supplemented with 10o fetal bo-vine serum, penicillin (100 units/ml), and streptomycin (100,ug/ml). After several passes in culture, proliferating cellswere grown as solid tumors in nude mice. Standard immu-nofluorescence techniques were used to characterize thetumor cells. Briefly, cells were seeded on sterile poly(L-lysine)-coated coverslips overnight and then fixed with 95%ethanol/5% acetic acid for 15 min at 40C. Cells were incu-bated for 2 hr with rabbit antiserum to glial fibrillary acidicprotein (GFAP), myelin basic protein (MBP), or vimentin, alldiluted as suggested by the supplier, followed by severalwashes. Finally, cells were incubated with fluorescein iso-thiocyanate-labeled anti-rabbit antiserum at 1:100 for anadditional hour, washed, and coverslipped. All antibodysolutions and washings were made in sodium phosphate-buffered saline (145 mM NaCl/10mM sodium phosphate, pH7.4). Primary and secondary antibodies were purchased fromDakopatts (Glostrup, Denmark).RNA Extraction, Microinjection into Oocytes, and Electro-
physiology. mRNA was extracted using the guanidinium/phenol/chloroform method (18) followed by oligo(dT)-cellulose chromatography. A total of seven mRNA prepara-tions from three human glioma cell lines (codes R-110, R-111,and R-162) grown in nude mice was used in this study.Techniques for injection of mRNA and for electrophysiolog-
Abbreviations: AcCho, acetylcholine; NMDA, N-methyl-D-aspartate; AMPA, a-amino-3-hydroxy-5-methyl-4-isoxazole propi-onic acid; CNQX, 6,7-dinitroquinoxaline-2,3-dione; GFAP, glialfibrillary acidic protein; MBP, myelin basic protein; I-V, current-voltage.tPresent address: Departamento de Neurociencias, Universidad delPais Vasco, 48940-Leioa, Spain.
3399
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
3400 Neurobiology: Matute et al.
ical recording from oocytes have been described (19-21).Briefly, Xenopus laevis oocytes were injected with about 50ng (1 jg/,lg) ofmRNA and kept in Barth's medium containing0.1 mg of gentamycin per ml at about 160C. Two days afterinjection, oocytes were treated with collagenase to removefollicular and other enveloping cells (20, 22). Four to 7 daysafter injection the oocytes were voltage-clamped using atwo-electrode voltage clamp, and agonist-induced membranecurrents were recorded at a clamp potential of -60 mV,unless otherwise stated. Ringer's solution (115 mM NaCl/2mM KCl/1.8 mM CaCl2/5 mM Hepes, pH 7.0) was contin-uously superfused over the oocyte and the drugs were appliedvia this perfusate. Aspartate (100 1LM) was applied togetherwith glycine (10 ,uM) to selectively activate N-methyl-D-aspartate (NMDA) receptors (16, 23). a-Amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA), quisqualate,and 6,7-dinitroquinoxaline-2,3-dione (CNQX) were pur-chased from Tocris Neuramin (Bristol, England). All otherdrugs and reagents were from Sigma.
RESULTSCellular Origin and Morphological Features of the Tumor
Cell Lines. Two tumors (R-110 and R-162) yielded cells thatappeared as human glioblastomas, according to establishedhistopathological criteria (24). Both types of cells had similarmorphological appearance and displayed comparable pat-terns ofimmunoreactivity to the antibodies used in this study.The majority of R-110 and R-162 cells was fibroblast-like,whereas the remaining 20-30%o of cells were flattened andpolygonal in shape. In contrast, R-111 cells, derived from atumor with features typical ofan anaplastic astrocytoma (24),were small (5-10 ,um) and formed a process-bearing, homo-geneous population with stellate shapes (Fig. 1A). The glialorigin of the three cell lines was further assessed usingantibodies to cytoskeletal components.Immunofluorescence showed that only a low proportion
(5-10%o) of R-110 and R-162 cells expressed GFAP, whereasin both cell lines an intense reactivity with anti-vimentinantibodies was observed in virtually all cells. Immunostain-ing with anti-MBP antibodies was also found in most R-110and R-162 cells, but the intensity of labeling was moderate tolow. In contrast, all R-111 cells were strongly reactive toanti-GFAP and vimentin antibodies (Fig. 1 B and C) but didnot express MBP. Thus, the presence ofGFAP in R-111 cellsconfirms this cell line as an astroglial tumor, whereas R-110and R-162 cells contain MBP, suggesting that they may haveoriginated from oligodendrocytes (25), though this is difficultto prove because glioblastoma multiforme is a primitive andhighly malignant form ofglioma whose cell origin is uncertain(26).
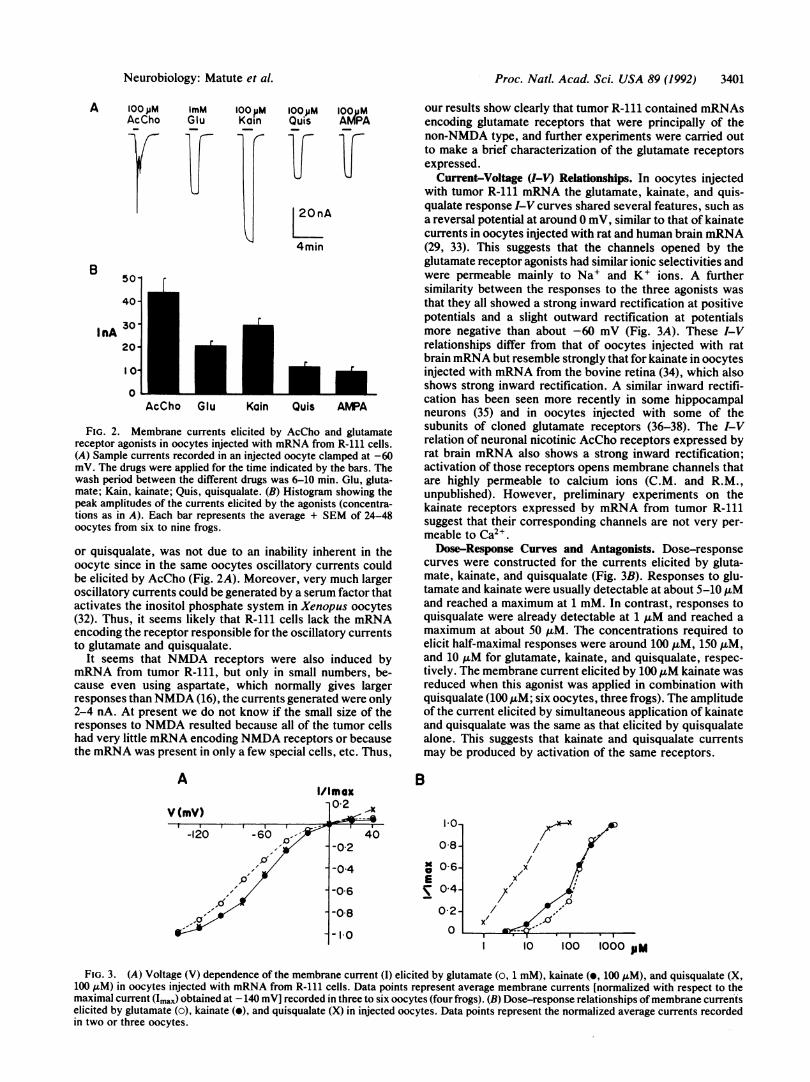
Expression of Neurotransmitter Receptors. Membrane cur-rent responses were recorded from 200 oocytes taken from>20 frogs. In all cases noninjected oocytes gave no signifi-cant responses to all neuroactive substances tested.No membrane currents were elicited by any of the neuro-
transmitters applied to oocytes injected with mRNAs ex-tracted from tumor R-110 or R-162 (38 oocytes, seven frogs).In contrast, oocytes from the same donor and other donorsinjected with mRNA from the R-111 tumor consistentlyshowed responses to acetylcholine (AcCho), glutamate, andkainate (Fig. 2). Glycine (1 mM) elicited small inward cur-rents (2-4 nA) that were difficult to discriminate from theslightly smaller currents observed in the control nonipjectedoocytes. Furthermore, -aminobutyric acid, P-alanine, andtaurine (all at 1 mM), serotonin (10 ,uM), substance P (200nM), or angiotensin II (1 ,LM) did not elicit significantcurrents. A few oocytes were also tested with norepinephrine(100 ,uM), dopamine (100 /LM), and somatostatin (100 nM)and again no significant responses were detected.
w4'~~~~ ~~~~~~~~~~~~~~A4 ,
.sw~~~~~~~~~~~~~~~~~~~1.,
FIG. 1. Morphological and immunofluorescence appearance ofastrocytoma R-111 cells in culture. (A) Phase-contrast. (B) Immu-nofluorescence labeling with antiserum to GFAP. (C) Immunoreac-tivity to vimentin antibodies. (Bar = 10 lum.)
It is known that activation of muscarinic AcCho receptors,either native or induced by brain mRNA, generates anoscillatory Cl- current due to stimulation of the inositolphosphate-Ca2+ receptor-channel coupling system (16, 20,27, 28). It appears that similar AcCho receptors were ex-pressed by the mRNA from tumor R-111 since AcChogenerated oscillatory Cl- currents that were blocked byatropine (10 nM) or pirenzepine (1 ,uM; see Fig. 4A) but werenot blocked by tubocurarine (5 ,uM). Therefore, the receptorsexpressed were muscarinic in nature, but their definite sub-type still needs to be determined.
Glutamate Receptors. In oocytes injected with rat brainmRNA kainate elicits a maintained smooth current, whereasglutamate and quisqualate elicit current responses that arecomprised of smooth and oscillatory components involvingthe inositol phosphate system (29-31). In contrast, in oocytesinjected with mRNA from tumor R-111 the responses to allthree agonists, as well as to AMPA, were smooth (Fig. 2A).The lack ofoscillatory currents, in the responses to glutamate
FIG. 2. Membrane currents elicited by AcCho and glutamatereceptor agonists in oocytes injected with mRNA from R-111 cells.(A) Sample currents recorded in an injected oocyte clamped at -60mV. The drugs were applied for the time indicated by the bars. Thewash period between the different drugs was 6-10 min. Glu, gluta-mate; Kain, kainate; Quis, quisqualate. (B) Histogram showing thepeak amplitudes of the currents elicited by the agonists (concentra-tions as in A). Each bar represents the average + SEM of 24-48oocytes from six to nine frogs.
or quisqualate, was not due to an inability inherent in theoocyte since in the same oocytes oscillatory currents couldbe elicited by AcCho (Fig. 2A). Moreover, very much largeroscillatory currents could be generated by a serum factor thatactivates the inositol phosphate system in Xenopus oocytes(32). Thus, it seems likely that R-111 cells lack the mRNAencoding the receptor responsible for the oscillatory currentsto glutamate and quisqualate.
It seems that NMDA receptors were also induced bymRNA from tumor R-111, but only in small numbers, be-cause even using aspartate, which normally gives largerresponses than NMDA (16), the currents generated were only2-4 nA. At present we do not know if the small size of theresponses to NMDA resulted because all of the tumor cellshad very little mRNA encoding NMDA receptors or becausethe mRNA was present in only a few special cells, etc. Thus,
A1/Imax
our results show clearly that tumor R-111 contained mRNAsencoding glutamate receptors that were principally of thenon-NMDA type, and further experiments were carried outto make a brief characterization of the glutamate receptorsexpressed.
Current-Voltage (I-V) Relationships. In oocytes injectedwith tumor R-111 mRNA the glutamate, kainate, and quis-qualate response I-V curves shared several features, such asa reversal potential at around 0 mV, similar to that of kainatecurrents in oocytes injected with rat and human brain mRNA(29, 33). This suggests that the channels opened by theglutamate receptor agonists had similar ionic selectivities andwere permeable mainly to Na' and K+ ions. A furthersimilarity between the responses to the three agonists wasthat they all showed a strong inward rectification at positivepotentials and a slight outward rectification at potentialsmore negative than about -60 mV (Fig. 3A). These I-Vrelationships differ from that of oocytes injected with ratbrain mRNA but resemble strongly that for kainate in oocytesinjected with mRNA from the bovine retina (34), which alsoshows strong inward rectification. A similar inward rectifi-cation has been seen more recently in some hippocampalneurons (35) and in oocytes injected with some of thesubunits of cloned glutamate receptors (36-38). The I-Vrelation of neuronal nicotinic AcCho receptors expressed byrat brain mRNA also shows a strong inward rectification;activation of those receptors opens membrane channels thatare highly permeable to calcium ions (C.M. and R.M.,unpublished). However, preliminary experiments on thekainate receptors expressed by mRNA from tumor R-111suggest that their corresponding channels are not very per-meable to Ca2+.Dose-Response Curves and Antagonists. Dose-response
curves were constructed for the currents elicited by gluta-mate, kainate, and quisqualate (Fig. 3B). Responses to glu-tamate and kainate were usually detectable at about 5-10 ,uMand reached a maximum at 1 mM. In contrast, responses toquisqualate were already detectable at 1 uM and reached amaximum at about 50 ,uM. The concentrations required toelicit half-maximal responses were around 100 ,uM, 150 ,uM,and 10 ,uM for glutamate, kainate, and quisqualate, respec-tively. The membrane current elicited by 100 ,uM kainate wasreduced when this agonist was applied in combination withquisqualate (100 1LM; six oocytes, three frogs). The amplitudeof the current elicited by simultaneous application of kainateand quisqualate was the same as that elicited by quisqualatealone. This suggests that kainate and quisqualate currentsmay be produced by activation of the same receptors.
B
-120 -60 40 10--0-2 08 /
-0 4 a 0 6- XE /X
,-'t -0-6 04_ /
-o ~ ~ ~ --2-0l 8 02 /Z<>~-I 10 100 1000 pM
FIG. 3. (A) Voltage (V) dependence of the membrane current (1) elicited by glutamate (o, 1 mM), kainate (o, 100 ,uM), and quisqualate (X,100 AM) in oocytes injected with mRNA from R-111 cells. Data points represent average membrane currents [normalized with respect to themaximal current (Umax) obtained at -140 mV] recorded in three to six oocytes (four frogs). (B) Dose-response relationships ofmembrane currentselicited by glutamate (o), kainate (o), and quisqualate (X) in injected oocytes. Data points represent the normalized average currents recordedin two or three oocytes.
Neurobiology: Matute et al.
Proc. NatL. Acad. Sci. USA 89 (1992)
A pz IPMAcCholOOpM -
1C
r.CNQX lpM 25pM
GlulmM_~ll-~
B ky. a. 500pMGlu ImM-l
DnA'5nA(D)nin D
Glu ImMD-APV 200pM
FIG. 4. Effects of antagonists on the re-sponses of oocytes expressing receptors fromR-111 astrocytoma cells. (A) Blockage of theresponse to AcCho by pirenzepine (pz, 1 IAM).(B) Kynurenic acid (ky. a., 500 AtM) partiallyblocked the 'membrane current elicited by glu-tamate (Glu). (C and D) The glutamate currentwas potently blocked by CNQX (IC50 = 1.6.uM;C) and was not affected by D-4-)-2-amino-5-phosphonovaleric acid (D-APV 200 IM; D).
A few glutamate receptor antagonists were examined fortheir effects on the currents generated by glutamate and theother agonists. The current elicited by glutamate (1 mM) wasreduced to 40% by 500 ,M kynurenic acid, a nonselectiveantagonist of glutamate receptors (four oocytes, two frogs;
Fig. 4B), and it was potently blocked by the non-NMDAreceptor antagonist CNQX (Fig. 4C), with a half-maximalinhibitory concentration (IC50) around 1.6 ,uM. D-(-)-2-Amino-5-phosphonovaleric acid, a presumed specific antag-onist forNMDA receptors, blocked the responses toNMDAbut at 200 ,uM had no effect on the responses to glutamate(Fig. 4D). In addition to blocking the response to glutamate,the antagonist CNQX inhibited also the kainate current, withan IC50 around 0.6 ,uM, and it fully blocked the AMPA (100,uM) responses at concentrations between 1 and 4 ,uM.
DISCUSSIONOur experiments show that Xenopus oocytes can be useful tostudy brain tumors and demonstrate that cells of a humanastrocytoma possess mRNAs encoding functional AcChoand glutamate receptors. It should be noted that the mem-brane currents elicited by the neurotransmitters in oocytesinjected with astrocytoma mRNA were rather small, whencompared with currents elicited in oocytes injected with ratbrain mRNA (e.g., ref. 16). Probably this results because ratneurons contain larger amounts ofthe corresponding mRNAsthan human astrocytoma cells, but it may also be that thedifferent "expressional potencies" of the mRNAs are partlya consequence of differences in the structure of rat andhuman receptor genes. Nevertheless, it is clear that thehuman astrocytoma R-111 cells contained mRNAs encodingneurotransmitter receptors, whereas the mRNAs from thetwo glioblastdtnas failed to express functional receptors inthe oocytes. Presumably this is because the glioblastomacells lacked the receptor mRNAs, although a less likelypossibility is that the mRNAs were present but somewhatdegraded, so that they failed to express functional receptors.The lack of expression of neurotransmitter receptors by
mRNAs isolated from the two glioblastomas, both of whichexpressed the oligodendroglial marker MBP, is in accord withreports that the membrane of rat oligodendrocytes lacksglutamate receptors (8-10). Similarly, the presence of gluta-mate receptor mRNAs in the human astrocytoma R-111 is inline with numerous reports that rat astrocytes possess glu-tamate receptors (4-10). Furthermore, the presence of anastrocytoma R-111 mRNA encoding muscarinic AcCho re-ceptors, which couple to the inositol phosphate system in theoocytes, also agrees with the finding of similar receptors inthe human astrocytoma cell line 1321 N1 (39-41). In spite ofthese important similarities some differences exist betweenour results and previous works. For example, it has beenreported that somatostatin, adrenergic, benzodiazepine, andsubstance P receptors are present in human gliomas (11-14).
However, the appropriate agonists failed to elicit membranecurrent responses in oocytes injected with the gliomamRNAs that we isolated. Moreover, there are also differ-ences in the glutamate receptors expressed. For instance, theastrocytoma R-111 mRNA induced the expression of someNMDA receptors, whereas these have not been detected inrat astrocytes (5-10), and the nonlinear I-V relationships ofcurrents generated by glutamate receptor agonists in theoocytes injected with the R-111 mRNA differ from the morelinear I-V relationships observed in rat astrocytes and 0-2Aprogenitor cells (4, 6, 9, 10). In addition, kynurenic acidblocked the responses to glutamate in oocytes injected withhuman R-111 mRNA, whereas in cultured rat astrocytes itfailed to block responses to glutamate (5) or blocked re-sponses to kainate (6). The cause of all of these differencesis not known, but some of them may be a consequence of thedifferent methods employed-e.g., cultured cells vs. tumorsborne in nude mice, etc. However, some genuine differencesprobably exist between human and rat glial cells and evenwithin different human gliomas. All of this will becomeclearer as more tumors, as well as the original human glialcells, are studied with electrophysiological and molecularbiological techniques. It will be particularly interesting todetermine ifthe presence, or absence, ofa particular receptormRNA is a consequence of the tumorigenic process.
In conclusion, our experiments show that mRNA isolatedfrom a human astrocytoma cell line expresses musearinicAcCho and glutamate receptors in Xenopus oocytes. Al-though the detailed molecular identity of these receptors stillremains to be elucidated, it seems clear that Xenopus oocytesare useful to characterize receptor functions and, in combi-nation with in vitro and in vivo studies, may help in thedevelopment of drugs affecting the rate of proliferation andsurvival of the tumor cells.
Th#s work was supported by Grant R01-NS23284 from the U.S.Public Health Service and Grant PM 88-100 from the SpanishDireccion General de Ciencia y Technologia. R.A. acknowledgessupport from The Pew Charitable Trusts.
1. Wrann, M., Bodmer, S., deMartin, R., Siepl, C., Hofer-Warbinck, R., Frei, K., Hofer, E. & Fontana, A. (1987) EMBOJ. 6, 1633-1636.
2. Siepl, C., Bodmer, S., Hofer, E., Wrann, M., Frei, K. &Fontana, A. (1988) Ann. N. Y. Acad. Sci 540, 437-438.
3. van Meir, E., Sawamura, Y., Diserens, A.-C., Hamou, M.-F.& deTribolet, N. (1990) Cancer Res. 50, 6683-6688.
4. Sontheimer, H., Kettenmann, H., Backus, K. H. & Schachner,M. (1988) Glia 1, 328-336.
5. Backus, K. H., Kettenmann, H. & Schachner, M. (1989) J.Neurosci. Res. 22, 274-282.
6. Usowicz, M. M., Gallo, V. & Cull-Candy, S. G. (1989) Nature(London) 339, 380-383.
7. Murphy, S. & Pearce, B. (1987) Neuroscience 22, 381-394.
3402 Neurobiology: Matute et al.
2 mrq
Proc. Natl. Acad. Sci. USA 89 (1992) 3403
8. Barres, B. A., Chun, L. L. Y. & Corey, D. P. (1990) Annu.Rev. Neurosci. 13, 441-474.
9. Barres, B. B., Koroshetz, W. J., Swartz, K. J., Chun,L. L. Y. & Corey, D. P. (1990) Neuron 4, 507-523.
10. Wyllie, D. J. A., Mathie, W. A., Symonds, C. J. & Cull-Candy, S. G. (1991) J. Physiol. 432, 235-258.
11. Lee, C.-M., Kum, W., Cockram, C. S., Teoh, R. & Young,J. D. (1989) Brain Res. 488, 328-331.
12. Reubi, J. C., Lang, W., Maurer, L. R., Koper, J. W. & Lam-berts, S. W. J. (1987) Cancer Res. 47, 5758-5764.
13. Ting, J., Sherman, P., Yokota, S., Moore, T. & Basta, P. (1988)Ann. N.Y. Acad. Sci. 540, 477-478.
14. Broaddus, W. C. & Bennett, J. P., Jr. (1990) Brain Res. 518,199-208.
15. Thomas, G. E., Szucs, M., Mamone, J. Y., Bem, W. T., Rush,M. D., Johnson, F. E. & Coscia, C. J. (1990) Life Sci. 46,1279-1286.
16. Miledi, R., Parker, I. & Sumikawa, K. (1989) in Fidia ResearchFoundation Neuroscience Award Lectures (Raven, NewYork), Vol. 3, pp. 57-90.
17. Carpenter, M. K., Parker, I. & Miledi, R. (1988) Proc. R. Soc.London Ser. B 234, 159-170.
18. Chomczynski, P. & Sacchi, N. (1987) Anal. Biochem. 162,156-159.
19. Miledi, R. (1982) Proc. R. Soc. London Ser. B 215, 491-494.20. Kusano, K., Miledi, R. & Stinnakre, J. (1982) J. Physiol.
(London) 328, 143-179.21. Miledi, R. & Sumikawa, K. (1982) Biomed. Res. 3, 390-399.22. Miledi, R. & Woodward, R. M. (1989) J. Physiol. (London) 416,
601-621.23. Verdoorn, T. A., Kleckner, N. W. & Dingledine, R. (1987)
Science 238, 1114-1116.24. Burger, P. C., Vogel, F. S., Green, S. B. & Strike, T. A. (1985)
Cancer 56, 1106-1111.25. Burger, P. C., Scheithauer, B. W. & Vogel, F. S. (1990) Sur-
gical Pathology of the Nervous System and Its Coverings(Churchill Livingston, New York), 3rd Ed.
26. Black, P. McL. (1991) N. Engl. J. Med. 324, 1555-1564.27. Parker, I., Gundersen, C. B. & Miledi, R. (1985) Neurosci. Res.
2, 491-496.28. Parker, I. & Miledi, R. (1986) Proc. R. Soc. London Ser. B 228,
307-315.29. Gundersen, C. B., Miledi, R. & Parker, I. (1984) Proc. R. Soc.
London Ser. B 221, 127-143.30. Sumikawa, K., Parker, I. & Miledi, R. (1984) Proc. Natl. Acad.
Sci. USA 81, 7994-7998.31. Hirono, C., Ito, I., Yamagishi, S. & Sugiyama, H. (1988)
Neurosci. Res. 6, 106-114.32. Tigyi, G., Dyer, D., Matute, C. & Miledi, R. (1990) Proc. Natl.
Acad. Sci. USA 87, 1521-1525.33. Gundersen, C. B., Miledi, R. & Parker, I. (1984) Nature
(London) 308, 421-424.34. Parker, I., Sumikawa, K. & Miledi, R. (1985) Proc. R. Soc.
London Ser. B 225, 99-106.35. Iino, M., Ozawa, S. & Tsuzuki, K. (1990) J. Physiol. (London)
424, 151-165.36. Boulter, J., Hollmann, M., O'Shea-Greenfield, A., Hartley,
M., Deneris, E., Maron, C. & Heinemann, S. (1990) Science249, 1033-1037.
37. Nakanishi, N., Schneider, N. A. & Axel, R. (1990) Neuron 5,569-581.
38. Lambolez, B., Curutchet, P., Stinnakre, J., Bregestovski, P.,Rossier, J. & Prado de Carvalho, L. (1991) Neurosci. Lett. 123,69-72.
39. Evans, T., McHardy, M. S., Tanner, L. I. & Harden, T. K.(1984) Mol. Pharmacol. 26, 395-404.
40. Brown, J. H., Goldstein, D. & Masters, S. B. (1985) Mol.Pharmacol. 27, 525-531.
41. Kunysz, E. A., Michel, A. D., Whiting, R. L. & Woods, K.(1989) Br. J. Pharmacol. 96, 271-278.