&p.1:Abstract Colorectal epithelium is composed of polar-ised absorptive enterocytes, mucus-producing gobletcells and enteroendocrine cells. All these cell lineagesare thought to arise from multipotential stem cells locat-ed near the base of the crypt, but the mechanisms whichcontrol differentiation and commitment of cells to a par-ticular lineage are poorly understood. We have used thehuman rectal adenocarcinoma cell line, HRA-19, to in-vestigate the regulation of expression of lineage-specificmarkers. HRA-19 cells have multipotential characteris-tics, forming absorptive, mucous and endocrine cellswhen grown as xenografts. However, HRA-19 cellsgrown in vitro in culture medium containing 10% foetalcalf serum show negligible expression of the differentiat-ed phenotypes observed in vivo. These findings initiallysuggested that the absence of positive stimuli from extra-cellular matrix, stromal cells and/or soluble factors pres-ent in vivo resulted in the lack of differentiation in vitro.The subsequent demonstration of a marked inhibitory ef-fect of foetal calf serum on differentiation provided analternative explanation for the differences between in vi-vo and in vitro differentiation. In addition, the inhibitionof differentiation differed widely between batches of foe-tal calf serum and limited the usefulness of the systemfor studying the regulation of differentiation. This manu-script describes the development of chemically definedculture conditions (Dulbecco’s Eagles medium supple-mented with insulin, transferrin and ascorbic acid) whichreproducibly induced the multilineage differentiation ofHRA-19 cells into absorptive, mucous and endocrinecells. Morphological characteristics and the expressionof lineage-specific markers, as determined by immuno-cytochemistry, identified absorptive, goblet and endo-crine cells in HRA-19 monolayers grown in this serum-free medium. Differentiation of cloned HRA-19 cells in-
to the three cell lineages proceeds in the absence of stro-mal cells and without exogenous extracellular matrix, al-though these factors may subsequently be shown to mod-ulate the rate of cell differentiation. These chemicallydefined culture conditions will facilitate the study of dif-ferentiation in the HRA-19 cell line in the absence of thecomplex mixture of growth factors, hormones and differ-entiation inhibitory factor(s) present in foetal calf serum.&bdy:
Introduction
The large intestine is lined with a single-layered epitheli-um with numerous invaginations or crypts. This epitheli-um is composed of absorptive, mucous and endocrinecells, all of which are thought to arise from commonstem cells located near the base of the crypt [4, 7, 41].Stem cell descendents undergo several rounds of cell di-vision and differentiate into three lineages as they mi-grate up the crypt towards the surface epithelium. Themechanisms which regulate proliferation of stem cellsand subsequent allocation of cells to a particular cell lin-eage are poorly understood. However, mesenchymalcells [16, 46] and extracellular matrix molecules (ECM)[17], acting via cell surface receptors [43, 49] have beenproposed as regulators of these processes.
We have used the human rectal adenocarcinoma cellline, HRA-19 [23], to investigate the regulation of ex-pression of lineage-specific markers. Cloned HRA-19cells have multipotential characteristics forming absorp-tive, mucous and endocrine cells when grown as xeno-grafts [24]. Although endocrine differentiation is a com-mon feature of colorectal carcinomas [48], only two oth-er colorectal cell lines have been shown capable of endo-crine differentiation. The NCI-H716 cell line [35] growsin vitro as floating aggregates with sporadic endocrinedifferentiation (<1%) while greater than 50% of NCI-H716 cells in xenografts have an endocrine phenotype[9]. In addition, the Caco-2 cell line, which has beenwidely used as an in vitro model of enterocytic differen-tiation [53] forms xenografts containing goblet cells, en-
K. Henderson · S.C. Kirkland (✉)Imperial Cancer Research Fund Histopathology Unit,Department of Histopathology,Royal Postgraduate Medical School,Hammersmith Hospital, DuCane Road, London W12 ONN, UK&/fn-block:
Multilineage differentiation of cloned HRA-19 cells in serum-free medium:A model of human colorectal epithelial differentiation
&misc:Accepted in revised form: 25 March 1996
terocytes, Paneth cells and endocrine cells [10]. HoweverCaco-2 cells cannot be induced to form endocrine cellsin vitro under a wide variety of conditions [10]. Likewisethe HRA-19 cells grown in vitro in 10% foetal calf se-rum (FCS) showed negligible expression of lineagemarkers [24]. Endocrine cells in xenografts of bothCaco-2 and HRA-19 were predominantly located adja-cent to the basement membrane. This lead to the sugges-tion that stromal cells and/or extracellular matrix mole-cules are critical factors in the induction of endocrinedifferentiation. However the subsequent demonstrationof differentiation-inhibitory activity in FCS provided analternative explanation for the absence of differentiatedphenotypes from HRA-19 monolayers [25]. Althoughdifferentiation of HRA-19 cells could be induced byswitching cells from 10% to 1% FCS [25], the amount ofdifferentiation differed widely between batches of foetalcalf serum and limited the usefulness of the system forstudying the regulation of differentiation. The variabilityand known differentiation-inhibitory activity of FCS sug-gested that serum-free conditions would be more appro-priate for the study of differentiation of HRA-19 cells invitro.
This manuscript describes the development of chemi-cally defined serum-free conditions which support thedifferentiation of cloned HRA-19 cells into both endo-crine and mucous phenotypes in monolayer culture. Un-der the serum-free conditions the differentiation reper-toire of HRA-19 cells has been further investigated byscreening for absorptive lineage markers. Previous workhas shown that HRA-19 cells grown in 10% FCS havethe morphological features of poorly differentiated ab-sorptive cells with sparse disorganised microvilli [23].However microvilli are more abundant and organised onthe absorptive cells seen in xenografts [24], demonstrat-ing a potential for absorptive differentiation given appro-priate conditions. In this study, absorptive cell differenti-ation has been investigated on the basis of the expressionof villin, a component of the intestinal brush border cyto-skeleton [47]. Brush border assembly was further analy-sed using transmission and scanning electron microsco-py.
Methods
HRA 19 cell line
The HRA 19 cell line was established from a primary human rec-tal adenocarcinoma [22]. Cloning of the cell line has previouslybeen described [24]. Experiments in this study were performedwith a cloned clone of the cell line designated HRA 19a1.1. be-tween passages 20–50. The cells were free from Mycoplasmacon-tamination.
Cell culture
Glass coverslips.&p.2:Confluent HRA-19 cells were trypsinised fromT25 flasks and seeded in Dulbecco’s Minimal Eagles Medium(DMEM; Gibco, Paisley) containing sodium pyruvate (110 mg/l),kanamycin (100 mg/l) plus 10% foetal calf serum (FCS; Gibco,
Paisley), onto 22-mm×22-mm untreated glass coverslips in 6-wellplates.One confluent T25 flask provided enough cells for 12 wells.Cells were allowed to settle for 72 h and then fed every 48 h witheither DMEM (as above) plus 1%–10% FCS or serum-free medi-um. Cells became confluent approximately 15 days after plating.All experiments were carried out in triplicate.
Permanox chambers.&p.2:Confluent HRA-19 cells were trypsinisedfrom T25 flasks and seeded into 8-chamber Permanox multislides(Nunc; Gibco, Paisley), each chamber being seeded with cellsequivalent to 1µg DNA. Cell numbers were estimated using afluorimetric DNA assay [42] because it was not possible to obtaina single cell suspension to enable direct cell counting. Cells wereeither seeded in 10% FCS and transferred to serum-free mediumor seeded directly in serum free medium after trypsinisation.
Serum free media was developed by testing combinations ofcomponents in DMEM for their ability to support proliferation anddifferentiation of HRA-19 cells. The compounds tested were insu-lin (2 µg/ml), transferrin (2µg/ml), ascorbic acid (10µg/ml), hy-drocortisone (10 ng/ml), all Sigma tissue culture products (Sigma,Poole) and 1% trace elements (Gibco, Paisley).
At the end of the experimental period, cells were washed withPBS and then either fixed in ethanol for 10 min at room tempera-ture for immunocytochemistry or frozen at –20°C for the DNA as-say.
Immunocytochemistry
Immunocytochemistry was performed as previously described [24,25] except that slides were developed in a diaminobenzidine(DAB)/nickel solution: 50 ml 0.1M acetate buffer (pH 6) contain-ing DAB (25 mg), nickel ammonium sulphate (1.25 g), βD glucose(100 mg), ammonium chloride (20 mg) and glucose oxidase(50 µl)) which yielded a black reaction product. Cells were lightlycounterstained with haematoxylin. The monoclonal antibodiesused in this study were LK2H10 (Boehringer Mannheim, Lewes)to chromogranin A within the granules of endocrine cells (1:100)[32], PR4D4(MAb to colonic mucus; 1:10) [45] and a monoclo-nal antibody to villin (ID2C3; Serotec, Oxford; 1:500)[47]. Thesecondary antibody was a peroxidase-conjugated rabbit antibodyto mouse immunoglobulins (Dako, High Wycombe).
For localisation of mucous and endocrine cells within the samemonolayer, cells were stained sequentially with two mouse mono-clonals [31]. Cells were firstly stained for mucus antigens usingthe PR4D4 antibody. To prevent the secondary antibody (rabbitanti-mouse immunoglobulins) binding subsequent mouse antibod-ies, any free arms on this secondary antibody are blocked by add-ing a non-specific mouse antibody which would not bind to thecells. This was mouse anti-smooth muscle actin (Sigma, Poole;1:16,000) applied for 30 min. The slides were washed three timesin PBS then incubated in rabbit anti-mouse IgG F(ab) fragments(Stratech Scientific, Luton; 1:50) for 30 min. These F(ab) frag-ments block any sites that the second secondary antibody mightrecognise. F(ab) fragments are used as they have only one bindingsite and therefore cannot subsequently bind the second primaryantibody. Following a further three washes with PBS, endocrinecells staining was performed with the chromogranin monoclonalas usual.
Western blotting
Adherent cells on glass coverslips were rinsed in phosphate buf-fered saline (PBS), scraped into 500µl lysis buffer (50 mM Trisbuffer, pH 6.8, containing 2% sodium dodecyl sulfate, 5% β-mer-captoethanol and 10% glycerol) and immediately boiled for10 min then placed on ice. The solubilised extracts were centri-fuged (13,000 rpm, 5 min, 4°C)and the supernatants subjected tothe Bradford protein assay (Biorad Laboratories, Hemel Hemp-stead). Lysates were stored at –70°C until needed. On thawing,1 µl 0.002% bromophenol blue in lysis buffer was added to sam-
260
ples which were then respun and subjected to sodium dodecyl sul-fate polyacrylamide gel electrophoresis (SDS/PAGE) using 7.5%resolving gels according to Laemmli [26]. Proteins were trans-ferred to nitrocellulose (Amersham International, Amersham) inTris-glycine buffer containing 20% methanol for 1 h at 100 V con-stant voltage. Villin was identified by immunostaining with MAbID2C3 (Serotec, Oxford).
Electron microscopy
Electron microscopy was performed as previously described [24].
Results
Differentiation in foetal calf serum
Previous experiments had shown negligible differentia-tion of HRA-19 cells in 10% FCS. However cells grownin 1% FCS reproducibly differentiated into endocrineand mucous cells. In this study the ability of threebatches of FCS to support endocrine and mucous differ-entiation by HRA-19 cells was tested.The total numberof cells expressing endocrine and mucous phenotypeswas up to 8-fold different in monolayers grown in 1%FCS from different batches. In addition the ratio of en-docrine to mucous cells was different for each batch ofFCS. No correlation was seen between the number ofdifferentiated cells and total cell number, which wasalso very variable between batches of FCS. This lack of
261
Fig. 1A–D Time course analysis of the differentiation and prolif-eration of HRA-19 cells grown on glass coverslips in either 1%foetal calf serum (FCS; open bars) or serum-free medium (IT-AHE; solid bars). A Total endocrine cells per coverslip. B Totalmucous cells per coverslip. C Total absorptive cells per coverslip.D Total DNA per coverslip. Mean ±S.D. Serum-free media gaveconsistently higher differentiation rates than 1% FCS for mucousand endocrine cells (P<0.01) and absorptive (P<0.05) cells&/fig.c:
Fig. 2A–C Effect of Dulbecco’s Eagles medium supplementedwith different additives on endocrine (A) and mucous (B) differen-tiation and proliferation (C) of HRA-19 cells. Cells were stainedon day 17. Mean ±S.D. The rate of endocrine and mucous differ-entiation was not statistically different in ITAHE and ITA. A as-corbic acid (10µg/ml), T transferrin (2µg/ml), I insulin (2µg/ml),H hydrocortisone (10ng/ml), E trace elements (1%)&/fig.c:
reproducibility coupled with the known differentiation-inhibitory activity of FCS led us to try to develop de-fined serum-free conditions compatible with cell differ-entiation.
Differentiation in serum-free medium
HRA-19 cells were grown in serum-free medium com-prising Dulbecco’s Eagles medium (DMEM) supple-mented with sodium pyruvate, kanamycin, insulin(2 µg/ml), transferrin (2 µg/ml), ascorbic acid(10 µg/ml), hydrocortisone (10 ng/ml) and 1% trace ele-ments (ITAHE). If cells were seeded onto glass cover-slips in this serum-free medium, attachment was verypoor, therefore cells were seeded in medium containing10% FCS and then transferred to serum-free conditionswhen they had attached to the coverslips. Cells express-ing lineage-specific markers, chromogranin A (endocrinecells), colonic mucus (goblet cells) and villin (absorptivecells) were counted in monolayers grown in 1% FCS andserum-free (SF) medium. Monolayers grown in SF medi-um contained increased numbers of differentiated pheno-types. Growth of cells in serum-free medium induced in-creases in endocrine (7-fold; Fig. 1A), mucous (10-fold;Fig. 1B) and absorptive (2-fold) cells (Fig. 1C) com-pared to equivalent monolayers grown in 1% FCS. Dif-ferentiated cells were declining by day 25, probably as aresult of declining cell viability. Although SF mediumhad a marked effect on differentiation, total cell numberwas similar whether cells were grown in SF medium or1% FCS (Fig. 1D).
262
Fig. 3A, B Endocrine (A) and mucous (B) differentiation in Dul-becco’s modified Eagles medium (DMEM) alone or DMEM sup-plemented with either serum (1%FCS) or different combinationsof insulin (I), transferrin (T) and ascorbic acid (A). Cells stainedon day 13. Mean ±S.D. All supplements except IA and 1% FCSgave significantly higher rates of differentiation than control medi-um (P<0.05)&/fig.c:
Fig. 4A–C Phase contrast mi-crographs of HRA-19 cellsgrown in 10% FCS (A) and se-rum free medium (B, C). Arrowindicates glandular organisa-tion. Scale bar, 50µm&/fig.c:
263
The increase in density of differentiated phenotypesin SF medium removed the need to scan such largemonolayer areas (484 mm2). Instead of 22 mm2 cover-slips, cells were now seeded into 8-chamber Permanoxmultislides (81-mm2/chamber). This system has severaladvantages over the coverslip method in that all cellsplated in the chamber are subsequently assessed for dif-ferentiation, whereas coverslip analysis relies on even at-
tachment across the area of the coverslip and the sur-rounding well for reproducibility. In addition, the cham-bers require much smaller quantities of reagents.
Using the multislide assay, we investigated whetherall the components present in our complete SF medium(ITAHE) were necessary for HRA-19 differentiation.This was achieved by removing one or more componentsof the SF medium and measuring both endocrine andmucous differentiation. Ascorbic acid (10µg/ml) andtransferrin (2µg/ml) were particularly effective whentested in isolation, and their combination together withinsulin (2 µg/ml) gave differentiated cell numbers equalto those in complete medium (ITAHE; Fig. 2). Removalof any of these three components from the ITA combina-tion resulted in reduced cell differentiation (Fig. 3),therefore further experiments were performed with thiscombination of culture additives.
Our initial experiments with glass coverslips had indi-cated poor attachment in SF medium, however these ex-periments were repeated using ITA medium and Per-manox slides. Under these conditions cells attached well,and this enabled the investigation of cell differentiation
Fig. 5A, B Localization of villin expression by HRA-19 cells af-ter 4 days (A) and 8 days (B) of growth in serum free medium. A4 days. Speckled apical membrane staining on cells in the centreof colonies. Cells at the edge of colonies are negative. B 8 days.Cells with intense positive staining are scattered in the monolayer.Scale bar, 20µm&/fig.c:
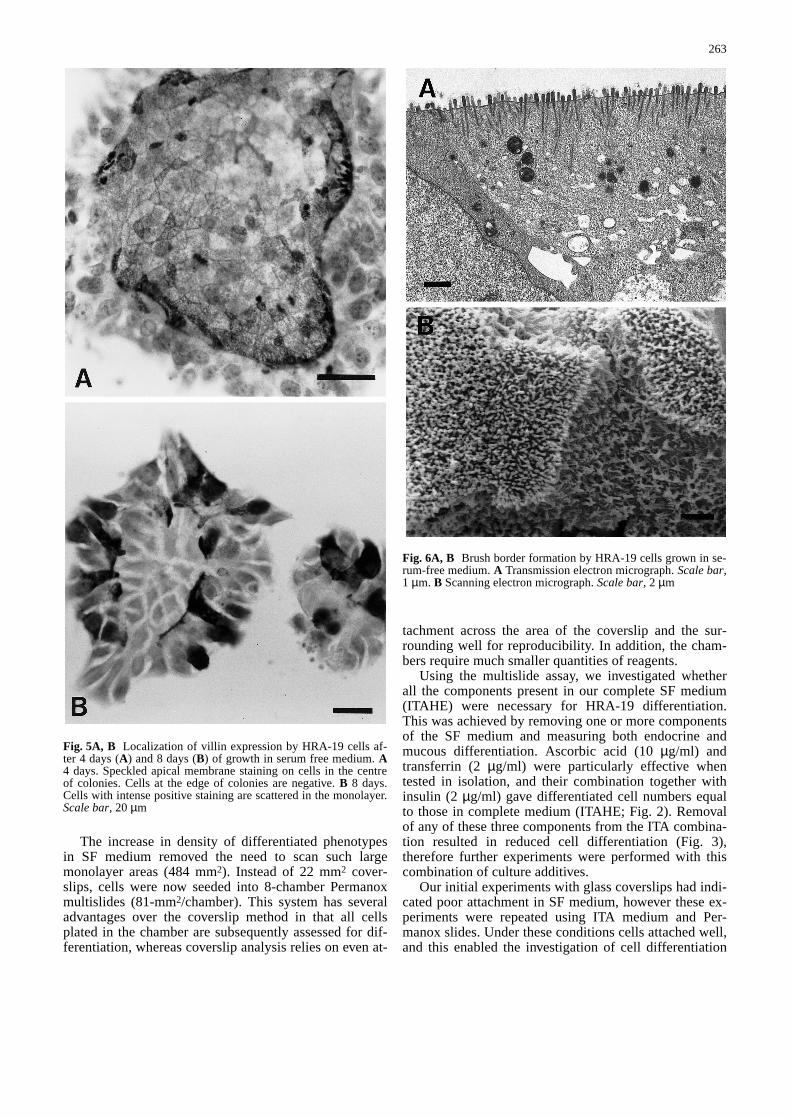
Fig. 6A, B Brush border formation by HRA-19 cells grown in se-rum-free medium. A Transmission electron micrograph. Scale bar,1 µm. B Scanning electron micrograph. Scale bar, 2 µm&/fig.c:
in the absence of factors which may have applied duringthe FCS preincubation. HRA-19 cells seeded and grownin SF medium contained similar numbers of cells toequivalent cultures seeded in FCS (unpublished observa-tions). Endocrine cells, which were the first differentiat-ed cell type to be detected in HRA-19 monolayers, werenow present in sufficient numbers 3 or 4 days after plat-ing, considerably reducing the time taken for the differ-entiation assay. On day 6 after plating in SF medium,HRA-19 monolayers contain endocrine, mucous and ab-sorptive cells.
Morphology of differentiating monolayers
HRA-19a1.1. cells grown in FCS are well spread withdistinct pale nuclei (Fig. 4A). Under serum-free condi-tions, cells are more cuboidal with less intercellularspaces (Fig. 4B,C). Palisading of cells at the edge of col-
onies is frequently seen in serum-free medium (Fig. 4B)with occasional formation of glandular structures with acentral lumen (Fig. 4C).
Absorptive cell differentiation
Immunocytochemical studies of villin expression by dif-ferentiating monolayers showed a change in pattern ofexpression with time. In the first few days after initiationof the culture, villin expression was weak but widespreadthroughout the monolayer except for cells at the edge ofcolonies (Fig. 5A). At this stage, staining was predomi-nantly cytoplasmic although weak speckled cell surfacestaining and cell border staining was apparent in someareas. With time, the pattern of villin staining was mark-edly changed with the appearance of scattered cells withintense cell surface staining (Fig. 5B). This staining wastaken to represent the assembly of a brush border by asubpopulation of cells, and these were the only cells in-cluded for quantitation of absorptive cells.
HRA-19 monolayers were investigated with transmis-sion and scanning electron microscopy in order to corre-late patterns of villin staining with brush border forma-tion. Electron microscopy confirmed the presence ofcells with many microvilli (Fig. 6A,B). Scanning elec-tron microscopy also showed that the central staining ofcell colonies with the villin antibody correlated well withthe presence of microvilli on cells within the colonies(Fig. 7A,B). Sufficient villin was present in the serum-free cultures to enable its identification by western blotanalysis (Fig. 8).
Cell lineage distribution
Cell staining for lineage-specific markers was more in-tense in SF medium than equivalent cells in serum-con-taining medium. In addition, differentiation was nowfound in preconfluent monolayers, whereas growth ofcells in 1% FCS required the cells to be confluent beforedifferentiated phenotypes could be detected. Cells ex-pressing lineage-specific markers were found scatteredin patches within the monolayer (Fig. 9). These areas
264
Fig. 7 Scanning electron micrographs of HRA-19 cells grown inserum-free medium (day 5). A Low power to show the differentsurface characteristic of cells in the centre of the colony (thick ar-row) compared the “edge” cells (thin arrow). Scale bar, 20 µm. BHigher magnification of the same colony showing “edge” cells(arrow) with few microvilli while cells in the centre (star) haveabundant microvilli. Scale bar, 5 µm&/fig.c:
Fig. 8A, B Western blot of vil-lin expression in HRA-19 cellsgrown in serum-free medium.Lane A, standards (kDa). LaneB, HRA-19 cell extract probedwith a MAb to villin (Bandcorresponds to 93 kDa)&/fig.c:
contained all cell lineages, i.e., absorptive, mucous andendocrine, interspersed with undifferentiated cells. Uni-form patches of cells composed of one cell lineage onlywere never observed in HRA-19 monolayers, instead dif-ferentiated phenotypes were found in the same areasometimes adjacent to each other (Fig. 10).
Discussion
HRA-19 colorectal carcinoma cells have multipotentialcharacteristics with cloned cells able to differentiate intoabsorptive, mucous and endocrine cells in vivo [24]. Al-though such differentiation was rarely observed in HRA-19 monolayers grown in 10% FCS, reduction of FCSconcentration to 1% induced the expression of lineage-specific markers in postconfluent monolayers [25]. How-ever the amount of differentiation observed was highlydependent on the individual batch of FCS. In addition,
the continued use of serum in this system would havecomplicated the interpretation of experiments intended toidentify factors involved in regulating differentiation. Toovercome these problems, a chemically defined mediumcontaining insulin, transferrin and ascorbic acid was de-veloped. Proliferation of HRA-19 cells in this SF medi-um was similar to 1% FCS although differentiation wassignificantly higher in serum-free cultures. The removalof the inhibitory activity in FCS was probably an impor-tant factor in the increased differentiation, although me-dium additives were also clearly required to induce dif-ferentiation. The importance of transferrin and insulinfor cell survival and growth has been well documentedfor many cell lines [2]. Ascorbic acid is involved inmany cellular functions [34] and has previously beenshown to stimulate cell proliferation in human colorectaltumour cells in serum free medium [19, 33]. Other stud-ies have described the development of chemically de-fined conditions for the growth of intestinal epithelialcells. Foetal rat intestinal epithelial cells proliferate rap-idly in SF medium, but proliferation is inhibited by ei-ther FCS or horse serum [14]. The proliferation of coloncarcinoma cells in SF medium has been shown to be ei-ther increased [33], equivalent [20] or decreased [44]when compared to serum-containing medium. These dis-parate findings can probably be attributed, at least inpart, to the use of SF medium with differing compositionin these various investigations and also to the widely dif-fering growth requirements of colorectal carcinoma celllines depending on their cellular differentiation [19]. Al-though HRA-19 cell proliferation was similar in 1% FCSor SF medium, their differentiation rate was significantlyincreased under serum-free conditions, with differentiat-ed cells now apparent in preconfluent monolayers. In-creased differentiation following the removal of serumhas also been reported in the WiDr cell line, where mu-cin synthesis is enhanced when cells are transferred to
265
Fig. 10 Scanning electron micrograph of HRA-19 cells grown inserum-free medium. Goblet cell (arrow) is seen adjacent to a welldifferentiated absorptive cell on the leftand an undifferentiatedcell with sparse microvilli on the right. Scale bar, 2 µm&/fig.c:
Fig. 9A, B Double immunocytochemistry on the same area of anHRA-19 monolayer grown in serum-free medium. Monolayerstained sequentially with anti-mucus antibody (PR4D4) (A) andthen with chromogranin antibody (B). A Mucous cells (arrow). BMucous cells (arrow) and endocrine cells (arrowheads). Scalebar, 50µm&/fig.c:
SF medium [44], although brush border enzyme expres-sion in Caco-2 cells is similar whether cells are grownwith serum or under chemically defined conditions [20].In addition to changes in differentiation, transfer of cellsto serum-free conditions can lead to changes in morphol-ogy. HC84S cells grown on collagen in SF mediumformed three-dimensional gland-like structures, unlikethe monolayers formed by the same cells in serum-con-taining medium [33]. These gland-like structures areseen in monolayers of HRA-19 cells when grown onplastic in SF medium. Therefore the behaviour of colo-rectal carcinoma cells when switched to SF medium isvery variable depending on the characteristics of individ-ual cell lines. Our previous studies have centred on endo-crine and mucous differentiation of HRA-19 cells [25],but the differentiation potential of the HRA-19 cells wasfurther investigated in the present study using absorptivelineage markers. Absorptive cell differentiation wasquantitated in the SF system from the cellular expressionof villin, a component of the intestinal brush border cyto-skeleton [3], and the presence of a brush border, usingboth transmission and scanning electron microscopy. Inintestinal mucosa, villin expression at the apical cell sur-face correlates with the presence of a mature brush bor-der [47], however villin is also expressed in the cyto-plasm of undifferentiated cells of the adult intestinal mu-cosa [47] and colonic carcinoma cell lines [5]. Analysisof enterocytes along the crypt-villous axis shows that un-differentiated cells have some staining near the apicalsurface while mature enterocytes display strong apicalstaining [47]. Experiments using in vitro models of en-terocytic differentiation give similar results. HT29-18, aclone from the HT29 cell line, undergoes enterocytic dif-ferentiation in the absence of glucose to yield cells withwell-developed brush borders [12]. Immunocytochemi-cal analysis of villin expression during this enterocyticdifferentiation shows that villin is localised to the brushborder area of the differentiated cells while cells grownunder non-differentiating conditions exhibit diffusestaining [12]. The results with the HRA-19 monolayersare very similar with undifferentiated cells displaying adiffuse mainly cytoplasmic staining pattern as expectedfor cells of intestinal origin. Some weak apical stainingand staining at cell borders was also observed in theseundifferentiated monolayers. This weak cell staining atcell borders probably correlates with the previously de-scribed weak lateral membrane staining for villin de-scribed in human colonic epithelium [50]. The pattern ofvillin staining in undifferentiated cultures and early se-rum-free cultures was characteristically restricted to cellsoccupying the centre of colonies in pre-confluent mono-layers, while most cells on the edge of colonies remainednegative. The differential staining pattern correlates withthe increased polarisation and cuboidal morphologycharacteristic of “central” cells compared with the flat-tened “edge” cells, and thus probably reflects the estab-lishment of cell polarity and formation of microvilli atthe apical surface. The pattern of staining in differentiat-ed HRA-19 monolayers was markedly different, with the
appearance of cells with intense cell surface staining,correlating with our ultrastructural observations showingscattered cells with a well-developed brush border. Ab-sorptive differentiation in HRA-19 monolayers was mea-sured by counting only those strongly staining cellswhich were readily distinguishable from the weaklystained background. Therefore monolayers grown underserum-free conditions contain undifferentiated, absorp-tive, mucous and endocrine cells. This cell line providesa model of colorectal differentiation which will enablequantitative analysis of differentiation along three lin-eages.
Several other human colorectal carcinoma cell lines,most notably HT-29 and Caco-2, have been used as invitro models of colorectal epithelial differentiation.Caco-2 cells spontaneously undergo enterocytic differen-tiation in postconfluent cultures [40], forming monolay-ers of polarised cells with tight junctions and an apicalbrush border. Postconfluent Caco-2 monolayers alsocontain “domes” indicating an ability for transepithelialion transport, which is also present in other colorectalcell lines such as T84 and HCA-7. Polarised monolayersof Caco-2 [15], T84 [11] and HCA-7[8] have been usedto investigate the mechanisms of colonic epithelial iontransport. Caco-2 cells have also been widely used in theinvestigation of brush border enzyme expression [6], re-cent studies demonstrating a role for triiodothyronine incontrolling sucrase expression by Caco-2 cells grown inSF medium [21]. Caco-2 clones have been describedwhich display apical villin expression, morphologicalhomogeneity and stability of brush border expression[36]. These C2BBe clones have been used as an in vitromodel of intestinal brush border assembly [37]. Differen-tiation in Caco-2 cells in vitro is principally enterocyticalthough some endocrine differentiation has been report-ed in xenografts [10]. However other cell lines such asHT-29 or LIM1863 [51] can show bidirectional differen-tiation in vitro into enterocytes and goblet cells. The HT-29 cell line, established by Fogh [13] has been widelyused in studies of colorectal differentiation. Under stan-dard culture conditions, HT-29 cells are principally un-differentiated, growing as a multilayer of unpolarisedcells [39], although a few mucous cells are present [1,27]. Rare cells which bind either mucus antibody PR5D5or columnar absorptive cell antibody PR 1A3 are presentin “undifferentiated” monolayers [45]. However, unlikethe HRA-19 cell line, PR4D4 positive cells were not ob-served in HT-29 monolayers. Although HT-29 monolay-ers are mostly undifferentiated under standard cultureconditions, differentiated populations (mucus and ab-sorptive) have been isolated from the HT-29 cell line fol-lowing growth in glucose-free medium [52], exposure tosodium butyrate [1], methotrexate [27] or 5-fluorouracil[28]. It has been suggested that these differentiated pop-ulations arise from pre-existing differentiated cells [27]which would need to be more adaptable to the inducing“stress” conditions. The finding that differentiated cellsare more resistant to 5-fluorouracil and methotrexatesupports such an hypothesis [29]. Clones of HT-29 cells
266
can display absorptive [18] or mucous [18, 38] differen-tiation. Such clones have been useful in the investigationof mucin gene expression [30]. These HT-29 clones de-rived from the heterogeneous parent line are character-ised by widespread homogeneous expression of the dif-ferentiated phenotypes of a single cell lineage [18]. Incontrast, it has not been possible to isolate populationsfrom the HRA-19 cell line which express characteristicsof a single lineage. Instead, HRA-19 clones regeneratethe heterogeneity of the parent cell line even after repeat-ed cloning, giving rise to monolayers containing absorp-tive, mucous and endocrine cells. Individual cell lineagesare found interspersed with other cell types in patchesthroughout the monolayer, suggesting that following acommitment to differentiate, all cell types are producedby precursor cells. Therefore HRA-19 cells resemble themultipotential stem cells of the crypt, which are respon-sible for self maintenance and generation of all differen-tiated lineages within colorectal epithelium. AlthoughHRA-19 cells have a differentiation repertoire whichmirrors colorectal epithelial stem cells, the ratio of celllineages in short-term monolayers differs from the in vi-vo situation. In normal colorectal epithelium endocrinecells represent only approximately 1% of differentiatedcells. However in short-term HRA-19 monolayers, endo-crine cells are present first and in greater numbers thaneither mucous or absorptive cells. Unlike the steady-statesituation found in normal colorectal crypts, HRA-19monolayers represent a dynamic system with increasingnumbers of differentiated cells with time. Our resultssuggest that the endocrine differentiation pathway is anearly event in the multilineage differentiation and thatthis pathway may diverge from the absorptive/mucouspathway. The neuropeptide products from these endo-crine cells may influence subsequent differentiationalong the mucous/absorptive pathway. In addition, thecell lineage ratios will almost certainly depend on cul-ture conditions and will be investigated in the presenceof matrix molecules and mesenchymal cells.
In summary, these studies have further characterisedthe HRA-19 model of colorectal epithelial differentiationby identifying the potential of HRA-19 cells for entero-cytic differentiation as well as the previously describedendocrine and mucous differentiation. Serum-free condi-tions have also been developed which induce a more rap-id and reproducible differentiation than seen with previ-ous experiments using FCS. This multilineage differenti-ation proceeds in the absence of stromal cells and with-out the addition of extracellular matrix molecules, al-though such factors may subsequently be shown to mod-ulate the ratio or absolute numbers of differentiated celltypes. HRA-19 cells will provide a useful model systemto investigate the sequence of events which follow induc-tion of differentiation in colorectal epithelial cells. Suchexperiments can now be performed in the absence of thecomplex mixture of growth factors, hormones and differ-entiation inhibitory factor(s) present in FCS
&p.2:Acknowledgements We are grateful to Carole Upton, ICRF Elec-tron Microscopy Unit, for transmission electron microscopy and toTim Ryder and Margaret Mobberley, Queen Charlottes Hospital,for scanning electron microscopy. This work was supported by theImperial Cancer Research Fund.
References
1. Augeron C, Laboisse CL (1984) Emergence of permanentlydifferentiated cell clones in a human colonic cancer cell line inculture after treatment with sodium butyrate. Cancer Res 44:3961–3969
2. Barnes D, Sato G (1980) Methods for growth of cultured cellsin serum free medium. Anal Biochem 102: 255–270
3. Bretscher A, Weber K (1980) Villin is a major protein of themicrovillous cytoskeleton which binds both G and F actin in acalcium-dependent manner. Cell 20: 839–847
4. Chang WWL, Leblond CP (1971) A unitarian theory of the or-igin of the three populations of epithelial cells in the mouselarge intestine. Anat Rec 169: 293
5. Chantret I, Barbat A, Dussaulx E, Brattain MG, ZweibaumA(1988) Epithelial polarity, villin expression and enterocyticdifferentiation of cultured human colon carcinoma cells: Asurvey of twenty cell lines. Cancer Res 48: 1936–1942
6. Chantret I, Rodolosse A, Barbat A, Dussaulx E, Brot-LarocheE, Zweibaum A, Rousset M (1994) Differential expression ofsucrase-isomaltase in clones isolated from early and late pas-sages of the cell line Caco-2: evidence for glucose-dependentnegative regulation. J Cell Sci 107: 213–225
7. Cheng H, Leblond CP(1974) Origin, differentiation and re-newal of the four main epithelial types in the mouse small in-testine. V. Unitarian theory of the origin of the four epithelialcell types. Am J Anat 141: 537–567
8. Cuthbert AW, Egleme C, Greenwood H, Hickman ME, Kirk-land SC, MacVinish LJ (1987) Calcium and cyclic AMP de-pendent chloride secretion in human colonic epithelium. Br JPharmacol 91: 503–515
9. De Bruine AP, Dinjens WNM, Pijls MMJ, Van der LindenEPM, Rousch MJM, Moerkerk PT, De Goeij AFPM, BosmanFT(1992) NCI-H716 cells as a model for endocrine differenti-ation in colorectal cancer. Virchows Arch. B Cell Pathol 62:311–320
10. De Bruine AP, De Vries JE, Dinjens WNM, Moerkerk PT, Vander Linden EPM, Pijls MMJ, Kate JT, Bosman FT(1993) Hu-man Caco-2 cells transfected with c-Ha-Ras as a model for en-docrine differentiation in the large intestine. Differentiation53: 51–60
11. Dharmsathaphorn K, McRoberts JA, Mandel KG, Tisdale LD,Masui H (1984) A human colonic tumor cell line that main-tains vectorial electrolyte transport. Am J Physiol 246: G204–G208
12. Dudouet B, Robine S, Huet C, Sahuquillo-Merino C, Blair L,Coudrier E, Louvard D (1987) Changes in villin synthesis andsubcellular distribution during intestinal differentiation ofHT29-18 clones. J Cell Biol 105: 359–369
13. Fogh J, Trempe G (1975) New human tumor cell lines. In:Fogh J (ed) Human tumor cells in vitro. Plenum, New York,pp 115–141
14. Fukamachi H (1992) Proliferation and differentiation of fetalrat intestinal epithelial cells in primary serum-free culture. JCell Sci 103: 511–519
15. Grasset E, Pinto M, Dussaulx E, Zweibaum A, Desjeux JF(1984) Epithelial properties of human colonic carcinoma cellline Caco-2: electrical parameters. Am J Physiol 247: C260–C267
16. Haffen K, Kedinger M, Simon-Assmann P (1987) Mesen-chyme dependent differentiation of epithelial progenitor cellsin the gut. J Ped Gastroenterol Nutr 6: 14–23
17. Hahn U, Stallmach A, Hahn EG, Riecken EO (1990) Base-ment membrane components are potent promoters of rat intes-
267
tinal epithelial cell differentiation in vitro. Gastroenterology98: 322–335
18. Huet C, Sahuquillo-Merino C, Coudrier E, Louvard D(1987)Absorptive and mucus-secreting subclones isolated from amultipotent intestinal cell line (HT-29) provide new models forcell polarity and terminal differentiation. J Cell Biol 105:345–357
19. Huschtscha L, Rozengurt E,Bodmer W (1991) Growth factorrequirements of human colorectal tumour cells: Relations tocellular differentiation. Eur J Cancer 27: 1680–1684
20. Jumarie C, Malo C (1991) Caco-2 cells cultured in serum freemedium as a model for the study of enterocytic differentiationin vitro. J Cell Physiol 149: 24–33
21. Jumarie C, Herring-Gillam FE, Beaulieu JF, Malo C (1996)Triiodothyronine stimulates the expression of sucrase-isomal-tase in Caco-2 cells cultured in serum free medium. Exp CellRes 222: 319–325
22. Kirkland SC, Bailey IG (1986) Establishment and character-isation of six human colorectal adenocarcinoma cell lines. Br JCancer 53: 779–785
23. Kirkland SC(1986)Endocrine differentiation by a human rectaladenocarcinoma cell line (HRA-19). Differentiation 33:148–155
24. Kirkland SC (1988) Clonal origin of columnar, mucous andendocrine cell lineages in human colorectal epithelium. Can-cer 61: 1359–1363
25. Kirkland SC, Henderson K (1994) Endocrine and mucous dif-ferentiation by a cloned human rectal adenocarcinoma cell line(HRA19) in vitro: inhibition by TGF-β1. J Cell Sci 107:1041–1046
26. Laemmli UK (1970) Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227: 680–684
27. Lesuffleur T, Barbat A, Dussaulx E, Zweibaum A (1990)Growth adaptation to methotrexate of HT-29 human colon car-cinoma cells is associated with their ability to differentiate intocolumnar absorptive and mucus-secreting cells. Cancer Res50:6334–6343
28. Lesuffleur T, Kornowski A, Luccioni C, Muleris M, Barbat A,Beaumatin J, Dussaulx E, Dutrillaux B, Zweibaum A (1991)Adaptation to 5-fluorouracil of the heterogeneous human co-lon tumor cell line HT-29 results in the selection of cells com-mitted to differentiation. Int J Cancer 49: 721–730
29. Lesuffleur T, Kornowski A, Augeron C, Dussaulx E, Barbat A,Laboisse C, Zweibaum A (1991) Increased growth adaptabili-ty to 5-fluorouracil and methotrexate of HT-29 subpopulationsselected for their commitment to differentiation. Int J Cancer49: 731–737
30. Lesuffleur T, Porchet N, Aubert JP, Swallow D, Gum JR, KimYS, Real FX, Zweibaum A (1993) Differential expression ofthe human mucin genes MUC1 to MUC5 in relation to growthand differentiation of different mucus-secreting HT-29 cellsubpopulations. J Cell Sci 106: 771–783
31. Lewis Carl SA, Gillette-Ferguson I, Ferguson DG (1993) Anindirect immunofluorescence procedure for staining the samecryosection with two mouse monoclonal primary antibodies. JHistochem Cytochem 41: 1273–1278
32. Lloyd RV, Wilson BS (1983) Specific endocrine tissue markerdefined by a monoclonal antibody. Science 222: 628–630
33. Murakami H, Masui H (1980). Hormonal control of humancolon carcinoma cell growth in serum-free medium. Proc NatlAcad Sci USA 77: 3464–3468
34. Padh H (1990). Cellular functions of ascorbic acid. Cell Biol68: 1166–1173
35. Park JG, Oie HK, Sugarbaker PH, Henslee JG, Chen TR,Johnson BE and Gazdar A (1987) Characteristics of cell linesestablished from human colorectal carcinoma. Cancer Res 47:6710–6718
36. Peterson MD, Mooseker MS (1992) Characterization of the en-terocyte-like brush border cytoskeleton of the C2BBe clones ofthe human intestinal cell line, Caco-2. J Cell Sci 102: 581–600
37. Peterson MD, Mooseker MS (1993) An in vitro model for theanalysis of intestinal brush border assembly. Ultrastructuralanalysis of cell contact-induced brush border assembly inCaco-2BBe. J Cell Sci 105: 445–460
38. Phillips TE, Huet C, Bilbo PR, Podolsky DK, Louvard D,Neutra MR (1988) Human intestinal goblet cells in monolayerculture: characterisation of a mucus-secreting subclone de-rived from the HT29 colon adenocarcinoma cell line. Gastro-enterology 94: 1390–1403
39. Pinto M, Appay M-D, Simon-Assmann P, Chevalier G, Draco-poli N, Fogh J, Zweibaum A (1982). Enterocytic differentia-tion of cultured human colon cancer cells by replacement ofglucose by galactose in the medium. Biol Cell 44: 193–196
40. Pinto M, Robine-Leon S, Appay M, Kedinger M, Triadou N,Dussaulx E, Lacroix B, Simon-Assman P, Haffen K, Fogh J,Zweibaum A (1983) Enterocyte-like differentiation and polar-ization of the human colon carcinoma cell line Caco-2 in cul-ture. Biol Cell 47: 323–330
41. Ponder BAJ, Schmidt GH, Wilkinson MM, Wood MJ, MonkM, Reid A (1985) Derivation of mouse intestinal crypts fromsingle progenitor cells. Nature 313: 689–691
42. Rao J, Otto WR (1992) Fluorimetric DNA assay for cellgrowth estimation. Anal Biochem 207: 186–192
43. Rao M, Manishen WJ, Maheshwari Y, Sykes DE, SiyanovaEY, Tyner AL, Weiser MM (1994) Laminin receptor expres-sion in rat intestine and liver during development and differen-tiation. Gastroenterology 107: 764–772
44. Real FX, Egea G, Franci C, Schussler MH, Xu M, Welt S(1991) Mucin production by colon cancer cells cultured in se-rum-free medium. Int J Cancer 49: 787–795
45. Richman PI, Bodmer WF (1987) Monoclonal antibodies to hu-man colorectal epithelium: Markers for differentiation and tu-mour characterisation. Int J Cancer 39: 317–328
46. Richman PI, Bodmer WF (1988) Control of differentiation inhuman colorectal carcinoma cell lines: epithelial-mesenchy-mal interactions. J Pathol 156: 197–211
47. Robine S, Huet C, Moll R, Sahuquillo-Merino C, Coudrier E,Zweibaum A, Louvard D (1985) Can villin be used to identifymalignant and undifferentiated normal digestive epithelialcells? Proc Natl Acad Sci USA 82:8488–8492
48. Smith DM, Haggitt RC (1984). The prevalence and prognosticsignificance of argyrophil cells in colorectal carcinomas. Am JSurg Pathol 8: 123–128
49. Simon-Assmann P, Leberquier C, Molto N, Uezato T, Buoz-iges F, Kedinger M (1994). Adhesive properties and integrinexpression profiles of two colonic cancer populations differingby their spreading on laminin. J Cell Sci 107: 577–587
50. West AB, Isaac CA, Carboni JM, Morrow JS, MoosekerMS,Barwick KW (1988) Localization of villin, a cytoskeletalprotein specific to microvilli, in human ileum and colon and incolonic neoplasms. Gastroenterology 94: 343–352
51. Whitehead RH, Jones JK, Gabriel A, Lukies RE (1987) A newcolon carcinoma cell line (LIM1863) that grows as organoidswith spontaneous differentiation into crypt-like structure invitro. Cancer Res 47: 2683–2689
52. Zweibaum A, Pinto M, Chevalier G, Dussaulx E, Triadou N,Lacroix B, Haffen K, Brun JL, Rousset M (1985) Enterocyticdifferentiation of a subpopulation of the human colon tumorcell line HT-29 selected for growth in sugar free medium andits inhibition by glucose. J Cell Physiol 122: 21–29
53. Zweibaum A, Laburthe M, Grasset E, Louvard D (1991) Useof cultured cell lines in studies of intestinal cell differentiationand function. In: Frizzell R, Fields H (eds) Handbook of phys-iology: the gastrointestinal system IV, Alan Liss, New York,Chap 7, pp 223-255