Multiscale climate change impacts on plant diversity in the Atacama Desert
Francisca P. Díaz1,2 | Claudio Latorre3,4 | Gabriela Carrasco‐Puga1,2 | Jamie R. Wood5 | Janet M. Wilmshurst5,6 | Daniela C. Soto1,2 | Theresa L. Cole5,7 | Rodrigo A. Gutiérrez1,2
AbstractComprehendingecologicaldynamicsrequiresnotonlyknowledgeofmoderncom‐munities but also detailed reconstructions of ecosystem history. Ancient DNA(aDNA)metabarcodingallowsbiodiversityresponsestomajorclimaticchangetobeexploredatdifferentspatialandtemporalscales.WeextractedaDNApreservedinfossilrodentmiddenstoreconstructlateQuaternaryvegetationdynamicsinthehy‐peraridAtacamaDesert. By comparing our paleo‐informedmillennial recordwithcontemporaryobservationsofinterannualvariationsindiversity,weshowlocalplantcommunitiesbehavedifferentiallyatdifferenttimescales.Intheinterannual(yearstodecades) time frame, only annual herbaceous expand and contract their distribu‐tional ranges (emerging from persistent seed banks) in response to precipitation,whereasperennialsdistributionappearstobeextraordinarilyresilient.Incontrast,atlongertimescales(thousandsofyears)manyperennialspeciesweredisplacedupto1,000mdownslopeduringpluvialevents.Givenongoingandfuturenaturalandan‐thropogenically inducedclimatechange,our resultsnotonlyprovidebaselines forvegetationintheAtacamaDesert,butalsohelptoinformhowtheseandotherhighmountainplantcommunitiesmayrespondtofluctuationsofclimateinthefuture.
Climate has a major role in the distribution of ecosystems andspecies range on Earth. The last few decades have revealed
overwhelming evidence that climate is changing in the world (Pachauri etal., 2014; Root etal., 2003). How climate change isaffectingspecies,biodiversityandecosystemstructure,functionandservices,bothnowandinthefuture,arequestionsofutmost
importance (Root etal., 2003). Moreover, how existing adapta‐tionscouldpotentiallybufferplantsandotherspeciesfromongo‐ingandfutureclimatechangeremainsunresolved.Understandingthe impact of climate change is particularly relevant in arid re‐gions, where ecosystems are often vulnerable to natural or an‐thropogenic ecosystem changes in water availability (Diaz,Grosjean, &Graumlich, 2003).Moreover, plants in such regionsdisplayremarkableadaptationstosuchdemandingenvironmentalconditions,andlocalcommunitiescanbesurprisinglydiverseanduniqueprovidingrichgeneticresourcesofglobalinterest.
On the western slopes of the Andes, the ecosystems of theAtacamaDesert (whichrangefromhyperarid tohighAndean)canbefoundacrossanelevationalrangespanningmorethan6,000mwhich encompasses pronounced precipitation and temperaturegradients over relatively short distances (~300km). Elevationalgradientsare important tools forunderstandingclimaticandevenevolutionary controls on biodiversity (Arroyo, Squeo, Armesto,&Villagran, 1988; Lenoir,Gégout,Marquet,DeRuffray,&Brisse,2008;Whittaker,Willis,&Field,2001).TheAtacamaisthusanex‐cellent model with which to study biodiversity dynamics and their climatic controls. On interannual timescales and during the late Holocene, Atacama climate variability is drivenmainly by ElNiñoSouthernOscillation(ENSO)(Vuille,Bradley,&Keimig,2000).Afterlarge precipitation events, annual blooms are frequent, increasingplantrichnessandcover,andsoilmicrobialactivity(Schulze‐Makuchetal.,2018).AncientDNA(aDNA)ofplantpathogensalsoincreasedin abundance and diversity in the Atacama during wetter events in thepast (Woodetal.,2018).Onmuch longer (millennial) times‐cales,theAtacamahasexperiencedmajorhydroclimatevariations.Airtemperaturesstartedtowarmfromglacialtointerglacialcondi‐tionsat~19,000calyearbpcoupledwithac.4°Cincreaseinregionalseasurfacetemperature (SST) (Kaiser,Schefuss,Lamy,Mohtadi,&Hebbeln,2008)alongwitha6°Cincreaseinterrestrialtropicaleco‐systems (Thompson,2000).Moistureorprecipitationchanges fol‐lowed,withaperiodofincreasedprecipitation(byaround500mm),originallydescribedastheCentralAndeanPluvialEvent (CAPE) intwodistinctintervals(CAPEI:15,900–13,800andCAPEII:12,700–9,700calyearbp) (Quadeetal.,2008).Here,wefollowtherecom‐mendationofarecentrevisionofdifferentpaleoclimaterecords(DePorras,Maldonado,DePol‐Holz,Latorre,&Betancourt,2017)fromthe central‐southern Atacama Desert, which defines widespreadwetconditionsfrom~17,500–13,800(CAPEI)and12,700–8,500calyear bp(CAPEII).
As each plant species currently inhabits a preferred tempera‐ture and precipitation range (determined mainly by elevation inthis environment), the exact identificationof the plants that livedin thedesert throughdifferent timeperiods canprovide informa‐tionaboutpastclimatechange(Díaz,Latorre,Maldonado,Quade,&Betancourt,2012;Latorre,Betancourt,Rylander,&Quade,2002).DNAmetabarcodingisamoleculartechniqueforidentifyingspeciesusingmolecularmarkers(barcodes)obtainedfromasamplecontain‐ingDNAofmorethanoneorganism(Hebert,Cywinska,&Ball,2003;Taberlet,Coissac,Pompanon,Brochmann,&Willerslev,2012)and
combinedwithaDNAcanbeusedtoreconstructpastdiversityfromancient or fossil samples like lake sediments andother substrates(Alsosetal.,2016;Murrayetal.,2012;Willerslevetal.,2003).
TherecoveryofaDNAorpaleoenvironmentalDNAfromavari‐etyoforganismsisatechniquethathasbeengreatlyimprovedduringthe lastdecade (andnowoften termedpaleogenomics) (Rawlenceetal., 2014;Willerslev&Cooper, 2005). The highest success rateforaDNAextractionhasbeenattainedwithorganismsfromfrozensites(Gould,León,Buffen,&Thompson,2010)andgenerallyfromdry or cold environments, rather than hot or humid sites (Grealyetal., 2016; Haouchar etal., 2014; Poinar etal., 1998; Rawlenceetal.,2014).TheAtacamaDesert,duetoitshyperaridity,thereforeoffers a potentially perfect environment to preserve aDNA (Kuchetal.,2002),butthehighdiurnaltemperaturesandstrongradiationcould also limit this.
Pastvariationsinclimateandvegetationcanbereconstructedbyanalyzing the contents of fossilmiddens of herbivorous rock‐dwell‐ing rodents (Betancourt & Saavedra, 2002; Pearson & Betancourt,2002),which are found in deserts around theworld (Betancourt&Davis,1984;Gil‐Romera,Scott,Marais,&Brook,2007;Murrayetal.,2012).Middensareamalgamationsofplant,animal,andotherorganicremains, embedded in a matrix of crystallized urine (Betancourt &Saavedra,2002).TheycanbepreservedfortensofmillenniainthearidAtacama(withtheoldestsamplesdatingto>50,000yearsbp),offeringexceptional spatiotemporal and taxonomic resolution.Middens pro‐videauniquesourceofancientmaterialpreservedinidealconditions,astheyareusuallyfoundinrockcreviceswhichoffersomeprotectionagainsttheeffectsofhighdiurnaltemperaturesanddirectradiation.
SequenceinformationfromaDNAcanbeusedtoidentifytaxathatareoftennot resolvedbymorphological analyses (plantmac‐rofossils, cuticles, or pollen) and has the potential of providing acomplementaryviewofbiodiversityovertimeascomparedtomoretraditionalmethodologies(Rawlenceetal.,2014;Woodetal.,2012).Although aDNA from one 11,700‐year‐old fossil rodent middenfromtheAtacamadeserthaspreviouslybeenusedtoreconstructpast rodent distributions (Kuch etal., 2002), we present the firstapplicationofDNAmetabarcodingandnext‐generationsequencingforentireplantcommunities,fromatimeseriesofmiddensspanning27,590–200yearsbeforepresent.
Ourgoal istounderstandhowdifferentmagnitudesofclimatechangeaffectplantcommunities in theAndeanAtacama.To inte‐grate spatial and temporal changes in plant biodiversity from an‐nual tomillennial timescales,wecompareaDNAdatafromrodentmiddens with contemporary plant biodiversity information overthecourseofeightconsecutiveyearsacrossanaltitudinalgradient(from2,500to4,500ma.s.l.)withstrongprecipitationdifferences(from10to300mm/year).Plantcommunitycompositionshouldbeaffectedinproportiontothemagnitudeofclimatechange,butthechallengebehindthishypothesis is todefine thesensitivityof thesystem.Forinstance,howhasvegetationrespondedtothelastde‐cadeofclimatechangeincontrasttomuchlongertimescaleswithmuch greater magnitudes of climate change (i.e., across the LastGlacial–interglacialtransition)(Nolanetal.,2018).
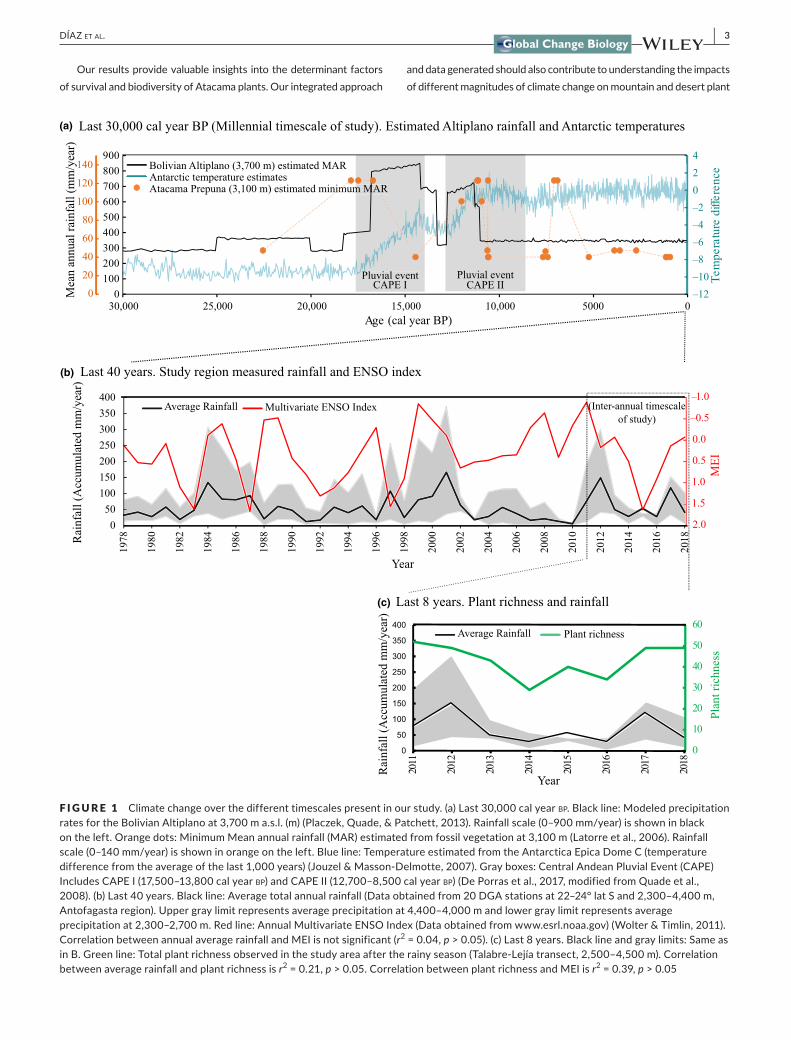
F I G U R E 1 Climatechangeoverthedifferenttimescalespresentinourstudy.(a)Last30,000calyearbp.Blackline:ModeledprecipitationratesfortheBolivianAltiplanoat3,700ma.s.l.(m)(Placzek,Quade,&Patchett,2013).Rainfallscale(0–900mm/year)isshowninblackontheleft.Orangedots:MinimumMeanannualrainfall(MAR)estimatedfromfossilvegetationat3,100m(Latorreetal.,2006).Rainfallscale(0–140mm/year)isshowninorangeontheleft.Blueline:TemperatureestimatedfromtheAntarcticaEpicaDomeC(temperaturedifferencefromtheaverageofthelast1,000years)(Jouzel&Masson‐Delmotte,2007).Grayboxes:CentralAndeanPluvialEvent(CAPE)IncludesCAPEI(17,500–13,800calyearbp)andCAPEII(12,700–8,500calyearbp)(DePorrasetal.,2017,modifiedfromQuadeetal.,2008).(b)Last40years.Blackline:Averagetotalannualrainfall(Dataobtainedfrom20DGAstationsat22–24°latSand2,300–4,400m,Antofagastaregion).Uppergraylimitrepresentsaverageprecipitationat4,400–4,000mandlowergraylimitrepresentsaverageprecipitationat2,300–2,700m.Redline:AnnualMultivariateENSOIndex(Dataobtainedfromwww.esrl.noaa.gov)(Wolter&Timlin,2011).CorrelationbetweenannualaveragerainfallandMEIisnotsignificant(r2=0.04,p>0.05).(c)Last8years.Blacklineandgraylimits:SameasinB.Greenline:Totalplantrichnessobservedinthestudyareaaftertherainyseason(Talabre‐Lejíatransect,2,500–4,500m).Correlationbetweenaveragerainfallandplantrichnessisr2=0.21,p>0.05.CorrelationbetweenplantrichnessandMEIisr2=0.39,p>0.05
–1.0
–0.5
0.0
0.5
1.0
1.5
2.0050
100150200250300350400
1978
1980
1982
1984
1986
1988
1990
1992
1994
1996
1998
2000
2002
2004
2006
2008
2010
2012
2014
2016
2018
0
10
20
30
40
50
60
0
50
100
150
200
250
300
350
400
2011
2012
2013
2014
2015
2016
2017
2018
Rai
nfal
l (A
ccum
ulat
ed m
m/y
ear)
Plan
t ric
hnes
s
(Inter-annual timescale of study)
(a) Last 30,000 cal year BP (Millennial timescale of study). Estimated Altiplano rainfall and Antarctic temperatures
(b) Last 40 years. Study region measured rainfall and ENSO index
0
40
100
140
60
80
20
120Bolivian Altiplano (3,700 m) estimated MARAntarctic temperature estimatesAtacama Prepuna (3,100 m) estimated minimum MAR
InnorthernChile's regionofAntofagasta lie thedriestareas in theworldasitcorrespondstothemaximumpenetrationoftheAtacamaDesert into the interior from the Pacific coast. Seasonal timing ofprecipitation determines a single season productivity pulse in theAtacama(aftersummer,duringMarchtoApril).Meanannualrainfall,obtainedfrom20DirecciónGeneraldeAguas,Chile(DGA)weatherstations locatedat22–24° latSand2,300–4,400m is54mm/year(Figure1b). The elevation gradient and their close interactionwithincreasing rainfall and decreasing temperatures (among other fac‐torssuchasgrazingorhumanimpacts)giverisetodistinctvegeta‐tion beltswhich can be consistently recognized across the region,based on overall plant physiognomy (Figure2a). However, species
Present relationships between environmental variables andplant distributions were established with vegetation surveys per‐formedduring the firstweekofAprilovereight consecutiveyears
F I G U R E 2 VegetationbeltsintheAndeanAtacamaDesertandcomparisonacrossdifferenttimescalesshowingmajorelevationalrangesofplanttaxainthestudysites.(a)PhotographsofvegetationbeltsintheAndeanAtacama.(b)Boxplotsshowtheinterannualvariabilityofrangedistributionsofannualandperennialtaxaobservedduringthe8yearsofvegetationsurveys.Elevationchangeisshownasannualvariationsalongtheupperandlowerlimitsofdistributionsforeachspecies,comparedtotheaveragepresent‐dayelevation(averagedoverthe8yearsofthestudy).Outliersrepresentspecieswithlowcoverage(onlyfoundinsomeyears).(c)Long‐termrangechangescomparedtopresent‐dayranges(metersbeloworabovetheircurrentdistribution).ZerohereisdefinedasthemaximumdistributionrangeestimatedfromA.Forinstance,ifaspecieswasfoundina10,000calyearbpmiddencollectedat3,000mandthatspeciesgrowstodaybetween4,000–4,200m,itwillappearasapointat−1,000m(theactualelevationalchange).Asmostmiddenswerecollectedalongthelowerlimitofthevegetation,theselimitstendtodescendinthepast,whichgivesagoodestimateoftheirpastlowerlimitofdistribution
(c) (b)
(a)
500
1000
1500
–1,000
–1,500
–500
–1,000
–1,500
–500
0
500
1000
1500
Years before present (cal year BP)2,5000 5,000 7,500 10,000 12,500 15,000 17,500 20,000 22,500 25,000 27,500
| 5DÍAZ et Al.
(2011–2018)andfollowingevery100mofelevation(22sites)acrossan elevation gradient from approximately 2,500 to 4,500m a.s.l.(Talabre‐Lejíatransect,TLT)(seeDíaz,Frugone,Gutiérrez,&Latorre,2016formoredetails).Atotalof61plantspecieswerecollectedandfrozentoconstructalocalDNAreferencedatabase(Atacama data‐base)acrossthisgradient(TableS1).Plantspeciesrichnesswasmea‐sured using two 250 m2plotsateachofthe22sitesoveralltheyearsof the study (Figure1c). Species observed permanence (i.e., howmanyyearsthesamespeciesappearedateachsite)wascalculatedforeachsiteandalongeachspeciesdistributionrange(minandmaxaltitude) aswell as for the averageupper and lower limit of distri‐butionofeachplantspecies in the last8years’surveys.Additionaldata regardingperennialspeciesdistribution (usingasimilarcollec‐tionmethod)wereavailablefromMay1998alongthesametransect(Latorre, 2002). Simple linearmodels were used for assessing anypossiblecorrelationswithPearson'scorrelationcoefficient,analyseswereperformedintherProgrammingLanguage(RCoreTeam,2018).
2.2 | Barcodes database
Using local DNA barcode databases can significantly increasethe resolutionorcapacity to identify taxa fromacomplexsample(Taberletetal.,2007).Toimproveresolutionandsubsequentanaly‐sisofaDNAreconstruction,weconstructedalocalbarcodeAtacama database using the species collectedduring the plant surveys andsomeadditionalspeciesthatwecollectedpreviouslyfromloweral‐titudesites(TableS1).Theseplantswereidentified,andspecimenswere deposited at the Herbarium at Universidad de Concepción,Chile (“Herbariode laUniversidaddeConcepciónCONC”;specieslistandcodesinTableS1).WeextractedDNAfromthesespecimensusingthePureLinkPlanttotalDNApurificationkit(Invitrogen)fol‐lowing themanufacturer's protocol.We amplified the chloroplasttrnLregionusingprimerscanddasdescribedbyTaberlet(Taberletetal.,2007).Amplificationwasperformedin25μl containing 0.4 μl of Phusion High‐Fidelity DNA Polymerase (Thermo Scientific,EEUU);8.0μlofenzymebuffer;0.2μlofeach10mMdNTP(ThermoScientific,EEUU);2.0μl fromeach10mMprimerand4.0μlofex‐tracted DNA (between 10 and 50 ng/μl). Following an activationstepof30s at95°C for theenzyme, thePCRmixtureunderwent25cyclesof5sat95°C,30sat55°Cand5minat72°Cona2720ThermalCycler(AppliedBiosystems,EEUU).
WeobtainedtrnLsequencesorbarcodesfromthose61speciesto construct the Atacama database (GenBank accession numbersMH115328–MH115388). The resolution of the selected barcode
2.3 | Fossil rodent middens collections and radiocarbon dating
All middens (N:26) were previously collected in Cerros de Aiquina(Antofagastaregion,northernChile,from2,414to3,380ma.s.l.)andradiocarbondatedbyC.Latorreandcollaborators(Latorre,Betancourt,Rylander,Quade,&Matthei,2003)(seeTableS2fordetails).Middensaredepositedepisodically andmostlydiscontinuously, neverthelessouragesdistributioncoversthedifferentclimaticeventsdescribedinFigure1a.Aftercollection,middenswereprocessedtoidentifymac‐rofossilsandpollen(resultspublishedinDePorrasetal.,2017;Latorreetal.,2003,comparisonsinFigure3)andanunprocessedsubsampleofeachmiddenwasretainedasavoucherspecimen(usedheretoex‐tractaDNA).FigureS1showsphotographsandaworkflowsummaryfortheaDNArecoveryandanalyses.Acceleratormassspectrometryradiocarbondatingwasperformedatdifferent facilities, seedetailsinTableS2.RadiocarbondatesforthemiddenswerecalibratedusingShCal13(Hoggetal.,2013)inOxCalv.4.3.2(Ramsey,1995)andme‐dianageswereusedforanalyses(TableS1).Twoofthemiddensweredeterminedtobemodern(after14Cdating)andtheothersarefrom49,600to20014C year bp.However, in thisstudyweconsideronlysamplesfrom27,590to200calyearbp,becausewecouldnotamplifyplantDNAfromtheoldersample(seeSection3.2inResults).
2.4 | Midden subsampling, aDNA extraction and amplification
Torecover thedamagedaDNAandavoidmodernDNAcontamina‐tion,subsampling,extractionandamplificationwereperformedatthespecializedaDNAlaboratoryatManaakiWhenua‐LandcareResearchLong‐TermEcologyLaboratory,Lincoln,NewZealand(FigureS1).First,weisolatedaninnerpieceofmaterialfrom24fossilrodentmiddens,following the protocol for subsampling late Quaternary coprolites(detailedinWood&Wilmshurst,2016).Thesubsamplingprocedurewascarriedoutinasterilizedsubsamplinghoodandconsistedof:ir‐radiatingthesamplewithUVC,removingtheoutersurface,irradiat‐ingagain,bisectingthesampleandtakingasampleforaDNAanalysisfromtheinteriorofonehalf.Toolsandthesamplingboxweresteri‐lizedbetweensamplesusingbleach,ethanol,andUVC.
F I G U R E 3 Numberoftaxaidentifiedfromthesamefossilrodentmiddens(CerrosdeAiquinaorCDAsites)usingdifferenttechniques.Macrofossildatawereobtainedfrom(Latorreetal.,2003)andpollendatawereobtainedfrom(DePorrasetal.,2017)
SubsamplingwasundertakeninsidethephysicallyisolatedaDNAlaboratorywhereportionsweighing5gwererehydratedinH2Ofor24hr.Tocheckforpossiblecontamination(Cooper&Poinar,2000;Rawlence etal., 2014;Wood&Wilmshurst, 2016),we carried outextractionblanksalongsideactualtemplatesamples.TheaDNAex‐tractionwasperformedusingthePowerMaxsoilDNAisolationkit(MoBio).Fromtwomiddens,wedidextractionreplicatestodetectheterogeneity inside themiddenand insamplesize.Sincesuitablemetabarcodes foraDNAstudies shouldbeno longer than300bp,but typicallyaround~130bpforplants (Rawlenceetal.,2014),weamplifiedasmallersectionoftrnL(163–228bp),usingprimerscandh(includingtheP6loop)describedin(Taberletetal.,2007).Assomeinconsistent results betweenPCRsof the same samplehavebeenshown in amplifications from environmental or complex samples(Ficetolaetal.,2015),wedidtwoindependentamplificationsusingthesameprimersforeachsample.Wemodifiedtheprimersaddingalinkersequence.Toavoidincorrectdetectionofclustersofourlownucleotidediversitylibrariesduringnext‐generationsequencing,weusedamixedsetoffourdifferentlengthprimerswiththesametar‐get(trnLc‐h),butwith0,1,2,or3nucleotidespacersbetweenthetrnLprimersandlinkersequence.TheaDNAPCRprotocolforonereaction (25 μl)was:BSA2.5μl,Buffer10×2.5μl,MgSO4 1 μl,dNTP0.2 μl,Fprimer0.5μl,Rprimer0.5μl,TaqPlatinumHiFi0.25,H2O 15.3 μl and DNA 2 μl(PCRprotocol:94°3:00then50–55cycles[94°0:30;55°0:30;68°0:45]and68°10:00).WealsofollowedaShrimpDNAaseprotocol (0.25μl:DNAasediluted10times,15minat37°and15minat65°)toremovepossiblecontaminantDNAfrommastermixbeforeaddingtheaDNA.ExtractioncontrolsandPCRnegativecontrols were included in all PCR reactions. We checked the ex‐tractionandamplificationsuccessbyvisualizingtheampliconsinanagarosegelelectrophoresis.DNAamplificationandalldownstreamprocedures (quantification,purification,andsequencing)wereper‐formedinlaboratoriesphysicallyseparatedfromtheaDNAlabora‐tory.Wesequencedbothamplificationreplicatesfromeachsample.Tocontrolforpossiblecontamination,wealsosequencedthreeex‐tractionblanks,whichweredoneinparallelwiththeextractedsam‐ples.AsweusedPCRtoamplify,itwasnotpossibletoinvestigatethequalityofDNApreservationbecausetheendsofDNAmoleculesarenotrevealed.ButwefollowothercriteriaofauthenticityforaDNA(Paaboetal.,2004)suchasthementionedextractioncontrols,PCRcontrolsandrepeatedamplificationsfromallextracts.
2.5 | Libraries preparation and sequencing
Topreparethelibraries,wefollowedtheprotocol“16SMetagenomicSequencing Library Preparation” (Illumina, 2013), adapted to ourmarker trnL (c‐h). We did a PCR clean‐up using magnetic beads(AgencourtAMPureXP),cleaningbothsizeextremes.Weranasec‐ondamplificationusing Illuminaadaptorswith the Illumina indices(NexteraXT)andweranasecondClean‐upusingthesamemethod.WequantifiedthelibrariesusingtheIlluminaLibraryQuantificationKit, Universal qPCR Mix (KAPA Biosystems) and real‐time PCRand validate them through capillary electrophoresis. Finally, we
Togeneratethedatabasesandanalyzethesequencingresults,weusedtheOBIToolspackage (Boyeretal.,2016).Weprocessedthedata, constructing a local Atacama database with our sequencesobtainedfromthevegetationsurveysandaglobalDDBJdatabaseperforminganinsilicoPCRtoDDBJ(Mashimaetal.,2016)(mirrorofGenBank)withourprimersusingecoPCRtool(Ficetolaetal.,2010).TheaDNAsequenceswereprocessedpairingforwardandreversereads,removing“joined”pairs.Then,wedereplicatedthereads,andfiltered thembycountnumber (more than5 reads) andby length(between60and240bp).Finally,we identifiedand removedpos‐siblePCRerrorsanddidthetaxonomicassignation(topscoretaxon,morethan95%ofidentity)tobothdatabasesusingNCBItaxonomydatabaseasreference(Federhen,2011).TheAtacama database was usedtoidentifyinthefirstplace.TheDDBJdatabasewasusedtoidentify missing taxa, higher taxonomic levels and contaminants(TableS4).Threelow‐qualitysamples(withmorethan10%ofcon‐taminants)werenotincludedinposterioranalyses.Weselected29(outof73)ofthemostinformativeplanttaxatoconstructtheresultsdiagraminFigure4(mostlyidentifiedtospeciesorgenuslevelandrepresentativeofAtacamavegetationbelts).
Consideringthatourmiddenseriesarenotcontinuouslydepos‐ited,andhavearelativelylowsamplesize,wewerenotabletodotime‐seriesstatisticalanalyses.Resultsarepresentedastaxaabun‐dancechartsandtaxaelevationdisplacementcomparedtothepres‐ent‐dayranges(metersbeloworabovetheircurrentdistribution).TocomparePCRandextractionreplicates,wecalculatedaBray–Curtisdissimilarity index, and then compared groups using an ANOVAfollowedbyaTukeymeandifferencestest(FigureS2).AllanalysesandchartswereperformedintheRProgrammingLanguage(RCoreTeam,2018),usingpackagerioja(Juggins,2007).
2.7 | Data availability statement
Sequencedata(localplantbarcodes)thatsupportthefindingsofthisstudy have been deposited inGenBankwith the accession codesMH115328–MH115388. Unprocessed reads from each middensample (.fastq) of this study are available from the correspondingauthoruponreasonablerequest.
3 | RESULTS
3.1 | Perennial plant species in high Andean Atacama exhibit resilience to short‐term climate changes
How vegetation adjusts to short‐term and interannual environ‐mentalchangesisrelevantforthedevelopmentofcomprehensive
diversitystudiesinaglobalchangescenario(Figure1).Thespeciesaccumulationcurvefromourstudyareashowsthatittookatleast6yearsofsurveystofindthebaselineofplantbiodiversity(>90%ofobservedspeciesandcurvesaturation).Asexpected,precipitationseemstobethemaindriverof interannualproductivityandplantspecies richness (Figure1c). Although other factors, such as tim‐ingofprecipitationandtemperaturechanges,werealsoimportantforgerminationofsomespecies.Forinstance,weobservedanim‐portantannualbloomofLupinus(reachingalmost50%ofsoilcoverin somesites)onlyduringone “LaNiña”year in2012 (Figure1b).Thisspecieswasnotregisteredinotheryearswithcomparablepre‐cipitation (2017)or another “LaNiña”year in2018, indicatinganinteractionofenvironmentalfactorsisrequiredformajorchangesin Lupinusgerminationrates.TheseresultsunderscoreasurprisingpersistenceofLupinusseedbanks,andalsotheimportanceofsus‐tainedeffortsovertimetounderstandbiodiversity,particularlyinextreme environments.
distribution during the 8years of surveys. In addition, perennialplantsshowedthesamedistributioninthelastyearsascomparedtoa survey from1998 (Villagránetal.,1998). Incontrast, annualherbaceous species contracted and expanded their distributionrangeeachyear(probablygerminatingfromapersistentsoilseedbank),accordingtointerannualweatherconditions(Figures1cand2b) and their permanence at each sitewas generally low (on av‐erageeachannualwasseeninthreeoutofthe8years).Theout‐comeofoursurveysunderscoresasurprisingpersistenceofseedbanksforsomeannuals (perhapsfordecades,andnot justyears)inahyperariddesert.Theseresultsindicatedistributionofperen‐nialplants isresilienttoshort‐term(annualanddecadal)environ‐mentalchangesandannualplantsshowedmoreflexibilityintheirobserveddistributionaccordingtoenvironmentalparameters,butprobablyalsoasaproductofaresilientsoilseedbank.Thesere‐sultsalsosuggestthattoproducesignificantcommunitychangesinperennialspecies,themagnitudeofclimatechangeshouldbemoreextensive in time or magnitude than what we have observed in the last two decades.
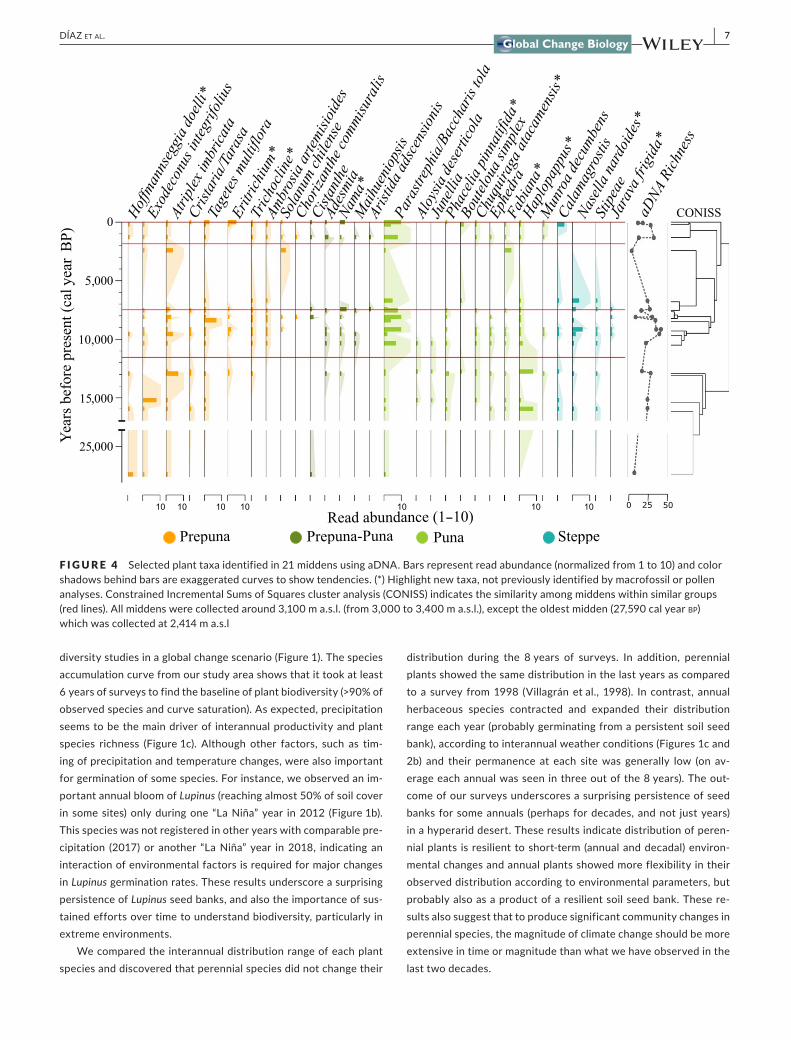
F I G U R E 4 Selectedplanttaxaidentifiedin21middensusingaDNA.Barsrepresentreadabundance(normalizedfrom1to10)andcolorshadowsbehindbarsareexaggeratedcurvestoshowtendencies.(*)Highlightnewtaxa,notpreviouslyidentifiedbymacrofossilorpollenanalyses.ConstrainedIncrementalSumsofSquaresclusteranalysis(CONISS)indicatesthesimilarityamongmiddenswithinsimilargroups(redlines).Allmiddenswerecollectedaround3,100ma.s.l.(from3,000to3,400ma.s.l.),excepttheoldestmidden(27,590calyearbp)whichwascollectedat2,414ma.s.l
10101010101010
8 | DÍAZ et Al.
3.2 | Using DNA barcodes increases taxonomic resolution and identification of plant species in samples as old as 27,590 years
Tounderstanddiversity changesover longerperiodsof time thanispossibletoobservebystandardfieldsurveys,weobtainedhigh‐quality aDNA, amplified and sequenced the trnL barcode in fossilsamples (dated200–27,590calyearbp)andfrommodernmiddensusing Illumina technology (see Figure S1 formidden photographsand a workflow summary of aDNA recovery and analyses). Fivelow‐quality samples were not included in posterior analyses (seeMaterialsandMethodsfordetails).Forinstance,theoldestmidden(49,60014C year bp)had92%of reads identifiedascontamination(assigned as Pottiaceae), and due to the absence of Pottiaceae inthe study region (and itspresence in thenegativecontrol)wedidnotincludethissampleinthefinalanalyses.Forothersamples,ex‐traction and amplification success were independent of age. Weobtainedameanof115,000readspermidden/samplefor21mid‐dens (seedetails inTableS2).Wecompared replicates anddiffer‐ent samplesusing theBray–Curtisdissimilarity index (BCI) (FigureS2).ExtractionreplicatesfromtwomiddensrevealsimilaridentifiedtaxawithameanBCIof0.3.PCRreplicatesforallsamplesyieldedsimilar results (mean BCI of 0.3) and were significantly differentcompared to independent samples (meanBCI:0.8).Therefore,weaveraged and combined PCR replicates for posterior analysis.WesequencedthreePCRnegativecontrols,obtaininglessthan70reads(mostly Asteraceae, Pottiaceae, Pooideae), except for one samplefromwhich3,000readswereobtained(mostlyTriticeaetribe),andremovedthesetaxafromposterioranalyses.Ofthesequences,94%weretaxonomicallyassignedusingbothplantdatabases.Ofthetotalassignedreads,99%wereclassifiedasAtacamataxaand1%ascon‐taminants (mainly fromvegetationoutside the laboratoryor food,seeTableS4).Weidentifiedatotalof73Atacamaplanttaxa(25spe‐cies,46genera,and26families)(TableS3)(Figures3and4).
We compared our aDNA record with pollen (De Porras etal.,2017) andmacrofossil (Latorreetal., 2003) reconstructionsprevi‐ouslyanalyzedinthesesamemiddens(Figure3).Thetotalnumber
of taxa was highest for aDNA (n=73), followed by macrofossils(n=63)andfinallypollen(n=24).Atotalof29plantfamilieswereidentifiedbyallthreemethods,butninewereidentifiedexclusivelybyaDNA(Figure3).Usingpaleogenomics,wewereabletoidentify14 species that were not previously recorded in either pollen ormacrofossil records.WebelievetheuseofaDNAandbarcodese‐quencingprovidesandaccurate,complementaryand,insomecases,morecompleteassessmentofbiodiversityovertimeinthesefossilsamplescomparedwithwhatcanbeobtainedwithtraditionalexper‐imentalapproachesalone.
3.3 | Major ecosystem responses to climate change in the past
To assess biodiversity in Atacama plant communities over the last27,590calyearbp,weusedourbarcodesequencedatafromaDNAandcomparedspeciesoccurrencetocurrentplantdistributionsandplantassemblagesfrommodernmiddensforwhichweusedthesameDNAbarcodes.Figure4showsselectedplanttaxa(29of73)andtheirap‐pearancesovertime.Asplanttolerancedefinesthealtitudeatwhichwe finda species,wepresent the results in relationship toexistingvegetationbelts (i.e.,Prepuna,PunaorSteppe,Figures2aand5) tofacilitateinterpretations.Wecomparedshort‐termdistributionrangeofeachplantspecieswith long‐termpastdistributions (Figure2b,c).DuringtheLastGlacial,at27,590calyearbp richness is very low and justafewPrepunaspecieswereidentified.Themostdramaticchangesoccurred from 17,000 to 7,000cal year bp, when Puna and Steppeplantspeciesoccurredalmost1,000mlowerthantoday(Figure2b).Nevertheless,Prepunaspecieswerealsopresentduringthatperiod,generating mixed and novel plant communities (Figure5b). Duringthelast7,000calyearbpplantspeciesrichnessislowanddominatedbyPrepunaspecies,exceptforthe200calyearbp midden which has morePunaelements,indicativeofmorehumidconditionsthantoday.
Atmillennialtimescales,ourresultsshowthat largemagnitudeclimatechanges(inlengthorintensity)triggeredbothperennialsandannualstomigratedownslopewherewaterlimitationandtempera‐tures are extreme today.
F I G U R E 5 Present‐daydistributionvs.majorpastvegetationchangeintheAtacamaDesert.(a)Currentvegetationbeltsat23°24′lat.S,AtacamaDesert,Chile.Aselevationandprecipitationincrease,temperaturedecreases.Whiteregionsindicateabsenceofplants.(b)Pastdownslopeplantmovementsduringpluvialeventsprovokedamixingofdifferentvegetationbeltsor“nonanalogcommunities”from2,500to3,500m.Thequestionmarkabove4,000mrepresentsourlackofmiddensfromhighaltitudes(allmiddensitesarefrom2,414to3,380m,seeTableS2fordetails)
(a) Present day (late-Holocene) (b) Pluvial events (late-Pleistocene and early Holocene)
Relative Distance0 km 25 km Relative Distance0 km 25 km
Elev
atio
n
Elev
atio
n
| 9DÍAZ et Al.
4 | DISCUSSION
Long‐termpaleoecologicalstudieslinkedtoshort‐termbiodiversitystudiesarefundamentaltounderstandingvegetationresponsestoongoing (anthropogenicallydriven) climatechange. InAndeanaridecosystems, we are severely data‐limited on both of these time‐scales. Fossil rodent middens are unique archives of biodiversityandanimportantsourceofinformationfromthepast.SequencingaDNAfrommiddensisnowpossible(Kuchetal.,2002;Murrayetal.,2012),andthisapproachoffersgreatopportunitiestoaddressques‐tionsofchangesinbiodiversityoverlongperiodsoftime.Thisareaofresearch,attheintersectionofgenomicsandpaleobotany,shouldbecomeanimportantareaofdiscoveriestounderstandvegetationresponsestoclimatechange.
4.1 | An aDNA record from fossil rodent middens
Weuse radiocarbondated fossil rodentmiddens andplant aDNAmetabarcoding to describe plant community changes up to27,590yearsbeforepresent.Onlyonepreviousstudy (Kuchetal.,2002)hassuccessfullyextractedDNAfromanAtacamarodentmid‐denbutusedSangersequencingfromindividualclones.WecompareouraDNArecordwithpollenandmacrofossilsreconstructions,andconcludethatdifferentexperimentalapproacheshavestrengthsandweaknessesandshouldthereforebeconsideredascomplementary(Figure3)(Parduccietal.,2015;Woodetal.,2012,2013).Thenum‐berofgenerawassimilarbetweenaDNAandmacrofossils,andbothrecordsarecomplementaryintermsofidentification.Morethanhalfof thegenerawere identified in justonetypeof record (aDNAormacrofossils).Plantmacrofossilsstillprovidethehighestnumberofidentifiedspecies(37),butaDNAwasabletoidentify14newspecies(includingsomekeyindicatorsspeciesfromtheSteppebelt,seetaxamarkedwithanasteriskinFigure4)notfoundasmacrofossilsorpol‐len(Figure3).Thecombineduseofpollenandmacrofossilsformid‐denanalysesistime‐consumingandinvolvesahighlevelofexpertisetoidentifylocalflorasfromthefragmentaryplantremainsfoundinmiddens. Incontrast,aDNArequiresspecialized facilities forDNAextractionfromfossilsamplesandPCRamplification,butifDNAiswell‐preserved taxonomic resolutiononly requiresa localbarcodedatabase.The fact thatouraDNAanalyseswereable to replicateandoftenexpandonthetotaldiversityindicatesthistechnologyasan importantcomplement to standardmiddenanalysis.Moreover,ourstudyprovidesareferencedatabaseforfuturestudiesofthesespecies.
Aswithotherproxies,theuseofaDNAtoreconstructbiodiver‐sityislimitedbytheresolutionofthemolecularmarkersorbarcodesobtained frommodern floras. ThedevelopmentofDNAbarcodesfromlocalflorasisafundamentalsteptoimprovetheresolutionofanyaDNArecord.Theserecordscanalsobefurtherbiasedbytheuniquetaphonomicenvironmentandageofthemidden.Therefore,more than establishing comparisonswith the entiremodern plantcommunities we use specific plants as key indicators of past cli‐mate change. DNA originates from different sources, including
pollenfrombothlocalandregionalsources(DePorras,Maldonado,Zamora‐Allendes,&Latorre,2015),particularlyinthecaseofwind‐pollinatedspecies.Sincetheephemeralappearanceofsomespeciesand the limitationsmentionedbefore for identifyingspeciesusingDNAbarcodes,somelocalflorafromthemiddencollectionsitesdonotappearintheDNAfrommodernmiddens,butalsoDNArevealspeciesthatwedidnotobserveinthefield.Nevertheless,DNAfrommodern middens shows that the reconstructed communities are similartopresent‐daycommunities,indicatingthatregionalsourcesofDNAlikelycontributeverylittletotherecord.Middensaccumu‐latedoverafewyearsoradecadeandcouldreflectawidertemporalframethanaone‐timepointsurvey.
4.2 | Integrating multiscale climate change with biodiversity dynamics
Plantcommunitiesandwhathasbeendefinedasvegetationbeltsaretemporaryagglomerationsofspeciesconstantlyrespondingtoclimatic,biologic,andanthropogenicfactors.Moreover, thosecur‐rentassemblagesareephemeralatthemillennialtimescalealthoughtheirpersistenceat shorter timescales is still unknown.Withpro‐nounced rainfall seasonality and interannual variability, thedesertvegetation of the Atacama can be challenging to study regardingshort‐termecologicalresponses.Annualsexpandandcontracttheirdistributionalranges inresponsetoprecipitationintheshortterm(yearstodecades)andtheirpermanenceateachsiteisgenerallylow.Thesespeciescanrespondquickly,andtheseedbanksareabundantandpersistentenoughtobridgeacrossscarceprecipitationevents.Such strategies allow plants to remain sensitive to environmentalchange of relatively little magnitude, such as ENSO variations. Incontrast, the elevational distribution of perennials did not changeover the entire sampling period (2011–2018) and did not changeevenwhenweincludedasurveyfrom1998(Villagránetal.,1998).Thisabsenceofchangeisunmistakableasareductioninmeanan‐nual precipitation (Sarricolea,Meseguer Ruiz, & Romero‐Aravena,2017)andanincreaseinmeanannualtemperature(Bennett,New,Marino,&Sillero‐Zubiri,2016)hasoccurredover the lastdecadesinAtacamaDesert.Thissuggests thatneitherENSOvariationnorrecentclimatechangehasbeenofsufficientmagnitudeoverthelast20yearstoinduceanysignificantelevationalmigrationinperenni‐alsorplantcommunitychanges.Thisresiliencecouldbeexplainedby theuniqueadaptations tovariablearidandhyperaridAtacamaconditionsorcouldbetheexpressionofanextinctiondebtor“ex‐tinctionlag,”ashasbeendescribedinotherhigh‐altitudeplantcom‐munities(Alexanderetal.,2018;Dullingeretal.,2012).
Incontrast,over longer timescales (thousandsofyears)stress‐resistantperennialherbaceousandshrubs(e.g.,Arroyoetal.,1988)appear to respond to larger or sustained climate change, such asexperiencedbyplantsduringtheLastGlacialperiodorthepluvialeventsatthePleistocene‐Holocenetransition (e.g.,CAPEIandII).Thebiggestchangescomparedtothepresentoccurredfrom17,000to 7,000cal bp, when plant species were found up to a 1,000mbelowtheircurrentdistribution(Figure2c),likelydrivenbyincreases
10 | DÍAZ et Al.
inprecipitationduringmajorpluvialevents(e.g.,Latorreetal.,2002,2003).ThevegetationidentifiedusingaDNArevealssomeimport‐ant differences between CAPE I and CAPE II. Diverse perennialshrubsandfewSteppegrassesarepartofCAPEIassemblages,asfoundinourmiddensdatedfrom16,000to15,000calyearBP.Incontrast, middens dated between 10,500 and 8,500cal year BP(CAPEII)showdominanceofParastrephia/Baccharis (both high ele‐vationPunashrubs)andSteppegrasses.ThesedifferencesbetweenthetwowetphasesofCAPEshowthatCAPEIIwaspossiblywetterandlikelywarmerthanCAPEI.
SuchmovementswerenotlimitedtothePleistocene,however,andourdatashowthatafewtaxadescended~800moverthelast3,000calbp,indicatingthatplantdistributionsrespondedtoshortertermclimatevariationsduringthelateHoloceneandoncentennialtimescalesduringtheHistoricPluvialEventattheendoftheLittleIceAge(c.1,400–1,900CE,seeLima,Christie,Santoro,&Latorre,2016; Mujica etal., 2015). Since plant diversity and productivityhave strong bottom‐up effects (Scherber etal., 2010), major pastplantchangeswouldbefeltthroughouttheentireAndeanAtacamaDesert ecosystem (Marquet etal., 1998). As interannual and sea‐sonal climate variability have strong effects on short‐lived herbi‐vores,suchasrodents(Meserve&Glanz,1978),climatevariabilityonlengthiertimescalesshouldimpactlong‐livedherbivoresandcar‐nivoressuchasfoxesandsouthAmericancamelids(Marquetetal.,1998).
Althoughtotalspeciesrichnessincreasedduringpluvialevents,vegetationbeltswerenotcompletelydisplaceddownslope.Instead,Prepuna communities were enriched by species moving in fromhigher elevations (Puna and Steppe) (Figure5b), generatingmixednovelplantcommunitiesor“nonanalogcommunities”(e.g.,Williams& Jackson, 2007). Temperature changes, although important inexplaining the presence of extreme xerophytes found at higherelevationswhere theygrowtoday (Latorre,Betancourt,&Arroyo,2006)weremostlikelynotimportantinexplainingthesedownslopeshifts. Indeed, colder temperatures during the Pleistocene (~6°colder;Thompson,2000)wouldhave forcedmanySteppespeciesdownslopefromtheAltiplano(whereitwouldhavebeentoocold;Mujica etal., 2015) but the presence of such species inwhat aretodayPrepunaenvironments,requiresincreasesinprecipitationasthecurrentenvironmentwouldhavebeentoodrytosupportthem(Latorreetal.,2006).Indeed,aspreviouslyshowninplantmacrofos‐silrecords,aDNAfloraduringtheverycoldtemperaturesoftheLastGlacialMaximumweremostlycomposedofjustafewPrepunaspe‐cies(Figure4).Clearly,thehigherelevationbelt(Steppe)exhibitsthelargestelevationalshifts.Inpart,this isduetowherethemiddenswerecollected(inlowerelevationbelts)butanothercausecouldre‐sideinthefactthathighAndeanvegetationrequiresmorefrequentand largeramountsofwater (rainfall)which inturnmakes itmuchmoresensitivetopastand(mostlikely)futureclimatechange.
Vegetation belts at our study sites (and in general for theAtacamaDeserthighlands) arehighlydynamic and their composi‐tionappears tobemostlydrivenbypast climatechangealthoughtheextentofhumanimpacts(bothhistoricalandinprehistory)has
notbeenfullyaddressedintheseecosystemsandremainsanopenquestion.PreviousstudiesintheAtacamashowhowrecentclimatefluctuationsoverthepast600yearsappeartobedrivingchangesinrichness (Mujicaetal.,2015)andthatevenhumanpopulationsre‐spondtothesesamechanges(Limaetal.,2016).Here,weevaluatedtheeffectofrecentclimatechange(overthelasttwodecades)andobservedthat theperennialplantdistributionshavechangedverylittlefrom1998topresent.AlthoughpasthumanoccupationsintheAtacamapeakedaround1,000yearago(Gayo,Latorre,&Santoro,2015),thehyperaridnatureofthedesertpreservespastlandscapeuse.Thus,humanpopulationdensityaroundoursiteswaslikelyverylowinthepastandisevenlowertoday,thusminimizingimpactonthenatural landscapewherewechoseoursites.Furthermore,oursites are close to a small road, but away fromanyevidenthumanperturbationsuchassettlements,majorminingcompanies,farmingterraces, fences, or garbage. The onlymajor human impact todaycouldbeattributedtodomesticatedllamasroamingthelandscape,buttheirpopulationdensityislowandthepressuretheseherdscur‐rentlyexertcouldbeofthesamemagnitudeastheherbivorouspres‐suresexertedbywildmegafauna(includingguanacos)inthepast.
Howvegetationadjuststoshiftingenvironmentalconditions,andthespeciesrichnessandcommunitydiversitypatternsthatcanarisefromsuchrestructuring,isrelevantforthedevelopmentofmorecom‐prehensivediversitystudiesandmodels(Whittakeretal.,2001).Forexample,regionalclimatemodelsfornorthernChilepredictincreasesin temperature (4°Chigher than today) anddecreases inprecipita‐tion (10%–30% lower than today)over thenext century (Minvielle&Garreaud, 2011; Thibeault, Seth,&Wang, 2012). SuchpotentialfutureclimatechangeissimilarinmagnitudetothatwhichoccurredduringtheLastGlacial–interglacialtransitionandoveratimescaleofseveralthousandsofyears.Inafast‐changingworld,Atacamaplantcommunitieswillmostlikelyundergoprofoundchangesasperenni‐alsaswellasannualsadjustaccordingly.Suchchangeswouldseeanupwardshiftof theupperboundaryof theabsolutedesertaspre‐cipitationdecreases,togetherwithaninvasionofhigh‐altitudecoldenvironmentsastemperatureincreases.Along‐termperspectiveandwidespreadintegrationofpaleoecologyandconservationbiologywillneedtobeadoptedifthevibrantAndeanAtacamaecosystemsaretobeconservedforthefuture(Barnoskyetal.,2017).Moreover,ourre‐sultsprovideimportantelementstoconsidermodelsofbiodiversityto climate change in other desert or high mountain ecosystems.
ACKNOWLEDG EMENTS
ThisresearchwasfundedbyFONDECYTPostdoctorado#3150616toF.P.D.,MillenniumInstituteforIntegrativeBiology(iBio)IniciativaCientífica Milenio‐MINECON, Fondo de Desarrollo de AreasPrioritarias (FONDAP)Center forGenomeRegulation (15090007)and EvoNet grant (DE‐FOA‐0001207) from Department ofEnergy,UnitedStatesofAmerica.C.L.acknowledgesfundingfromCONICYT'sAportedeFinanciamientoBasal (AFB‐170008). J.R.W.and J.M.W. acknowledge Strategic Science Investment Fundingfor Crown Research Institutes from theNew ZealandMinistry of
| 11DÍAZ et Al.
Business, Innovation and Employment's Science and InnovationGroup, and Manaaki Whenua – Landcare Research for hostingF.P.D.'svisittotheLandcareResearchLong‐TermEcologyLaboratorywhere the aDNA extractions were undertaken. We thank AliciaMarticorenaandRosaMonterofortheirhelpintheHerbariodelaUniversidad de Concepción (CONC). We appreciate helpful com‐mentsandconstructivecriticismsfromthereviewersandeditorofourmanuscript.Wewouldalsoliketoacknowledgethoughtfulcom‐mentsandsuggestionsbyDr.JulioBetancourtthatgreatlyhelpedinimprovingourmanuscript.
ORCID
Francisca P. Díaz https://orcid.org/0000‐0002‐1100‐7801
Jamie R. Wood http://orcid.org/0000‐0001‐8008‐6083
Janet M. Wilmshurst http://orcid.org/0000‐0002‐4474‐8569
Daniela C. Soto http://orcid.org/0000‐0002‐6292‐655X
Theresa L. Cole http://orcid.org/0000‐0002‐0197‐286X
Rodrigo A. Gutiérrez http://orcid.org/0000‐0002‐5961‐5005
R E FE R E N C E S
Alexander,J.M.,Chalmandrier,L.,Lenoir,J.,Burgess,T.I.,Essl,F.,Haider,S., … Pellissier, L. (2018). Lags in the response of mountain plantcommunities to climate change. Global Change Biology,24,563–579.https://doi.org/10.1111/gcb.13976
Alsos,I.G.,Sjögren,P.,Edwards,M.E.,Landvik,J.Y.,Gielly,L.,Forwick,M., … Pedersen, M. W. (2016). Sedimentary ancient DNA fromLakeSkartjørna,Svalbard:AssessingtheresilienceofarcticfloratoHolocene climate change. The Holocene, 26, 627–642. https://doi.org/10.1177/0959683615612563
Arroyo,M.T.K.,Squeo,F.A.,Armesto,J.J.,&Villagran,C.(1988).EffectsofaridityonplantdiversityinthenorthernChileanAndes–Resultsofanaturalexperiment.Annals of the Missouri Botanical Garden,75,55–78.https://doi.org/10.2307/2399466
Barnosky,A.D.,Hadly,E.A.,Gonzalez,P.,Head,J.,Polly,P.D.,Lawing,A.M.,…Zhang,Z.(2017).Mergingpaleobiologywithconservationbiology toguide the futureof terrestrialecosystems.Science,355,eaah4787.https://doi.org/10.1126/science.aah4787
Bennett,M.,New,M.,Marino,J.,&Sillero‐Zubiri,C.(2016).Climatecom‐plexityintheCentralAndes:Astudycaseonempirically‐basedlocalvariationsintheDryPuna.Journal of Arid Environments,128,40–49.https://doi.org/10.1016/j.jaridenv.2016.01.004
Betancourt,J.L.,&Saavedra,B.(2002).Rodentmiddens,anewmethodfor Quaternary research in arid zones of South America. Revista Chilena De Historia Natural,75,527–546.
Boyer, F.,Mercier,C.,Bonin,A., LeBras,Y., Taberlet,P.,&Coissac,E.(2016). obitools: A unix‐inspired software package for DNA me‐tabarcoding. Molecular Ecology Resources,16, 176–182. https://doi.org/10.1111/1755‐0998.12428
Cooper, A., & Poinar, H. N. (2000). Ancient DNA: Do it right ornot at all. Science, 289, 1139–1139. https://doi.org/10.1126/science.289.5482.1139b
De Porras, M. E., Maldonado, A., De Pol‐Holz, R., Latorre, C., &Betancourt, J. L. (2017). LateQuaternary environmental dynamicsin the Atacama Desert reconstructed from rodent midden pollenrecords. Journal of Quaternary Science, 32, 665–684. https://doi.org/10.1002/jqs.2980
De Porras, M. E., Maldonado, A., Zamora‐Allendes, A., & Latorre, C.(2015).CalibratingthepollensignalinmodernrodentmiddensfromnorthernChiletoimprovetheinterpretationofthelateQuaternarymidden record. Quaternary Research, 84, 301–311. https://doi.org/10.1016/j.yqres.2015.10.004
Díaz,F.P.,Frugone,M.,Gutiérrez,R.A.,&Latorre,C.(2016).Nitrogency‐clinginanextremehyperaridenvironmentinferredfromδ 15 N anal‐ysesofplants,soilsandherbivorediet.Scientific reports,6,22226.
Díaz, F. P., Latorre, C., Maldonado, A., Quade, J., & Betancourt, J. L.(2012). Rodent middens reveal episodic, long‐distance plant col‐onizations across the hyperarid Atacama Desert over the last34,000 years. Journal of Biogeography, 39, 510–525. https://doi.org/10.1111/j.1365‐2699.2011.02617.x
Dullinger,S.,Gattringer,A.,Thuiller,W.,Moser,D.,Zimmermann,N.E.,Guisan, A., …Hülber, K. (2012). Extinction debt of high‐mountainplants under twenty‐first‐century climate change. Nature Climate Change,2,619.https://doi.org/10.1038/nclimate1514
Federhen, S. (2011). The NCBI taxonomy database. Nucleic Acids Research,40,D136–D143.
Ficetola, G. F., Coissac, E., Zundel, S., Riaz, T., Shehzad,W., Bessière,J., … Pompanon, F. (2010). An in silico approach for the eval‐uation of DNA barcodes. BMC Genomics, 11, 434. https://doi.org/10.1186/1471‐2164‐11‐434
Ficetola, G. F., Pansu, J., Bonin, A., Coissac, E., Giguet‐Covex, C., DeBarba,M.,…Taberlet,P. (2015).Replication levels, falsepresencesand the estimationof thepresence/absence fromeDNAmetabar‐coding data. Molecular Ecology Resources,15, 543–556.https://doi.org/10.1111/1755‐0998.12338
Gayo,E.M.,Latorre,C.,&Santoro,C.M.(2015).Timingofoccupationand regional settlement patterns revealed by time‐series analysesof an archaeological radiocarbon database for the South‐CentralAndes (16–25°S). Quaternary International, 356, 4–14. https://doi.org/10.1016/j.quaint.2014.09.076
Gil‐Romera,G.,Scott,L.,Marais,E.,&Brook,G.A.(2007).LateHoloceneenvironmental change in the northwestern Namib Desert margin: New fossil pollen evidence from hyrax middens. Palaeogeography Palaeoclimatology Palaeoecology,249,1–17.https://doi.org/10.1016/j.palaeo.2007.01.002
Gould,B.A.,León,B.,Buffen,A.M.,&Thompson,L.G.(2010).Evidenceofahigh‐Andean,mid‐Holoceneplantcommunity:AnancientDNAanalysisof glaciallypreserved remains.American Journal of Botany,97,1579–1584.https://doi.org/10.3732/ajb.1000058
Grealy, A.,Douglass, K.,Haile, J., Bruwer, C.,Gough, C., &Bunce,M.(2016). Tropical ancient DNA from bulk archaeological fish bonerevealsthesubsistencepracticesofahistoriccoastalcommunityinsouthwestMadagascar.Journal of Archaeological Science,75,82–88.https://doi.org/10.1016/j.jas.2016.10.001
Haouchar, D., Haile, J., Mcdowell, M. C., Murray, D. C.,White, N. E.,Allcock, R. J. N., … Bunce, M. (2014). Thorough assessment ofDNApreservation fromfossilboneandsedimentsexcavated froma late Pleistocene‐Holocene cave deposit on Kangaroo Island,SouthAustralia.Quaternary Science Reviews,84,56–64.https://doi.org/10.1016/j.quascirev.2013.11.007
Hebert, P. D., Cywinska, A., & Ball, S. L. (2003). Biological identifica‐tions through DNA barcodes. Proceedings of the Royal Society of London B: Biological Sciences,270,313–321.https://doi.org/10.1098/rspb.2002.2218
Hogg,A.G.,Hua,Q.,Blackwell,P.G.,Niu,M.,Buck,C.E.,Guilderson,T.P.,…Zimmerman,S.R.H.(2013).SHCal13SouthernHemispherecalibration, 0–50,000 years cal BP. Radiocarbon, 55, 1889–1903.https://doi.org/10.2458/azu_js_rc.55.16783
Jackson, S. T., &Blois, J. L. (2015). Community ecology in a changingenvironment:PerspectivesfromtheQuaternary.Proceedings of the National Academy of Sciences of the United States of America, 112,4915–4921.https://doi.org/10.1073/pnas.1403664111
Juggins,S. (2007).C2 Version 1.5 User guide. Software for ecological and palaeoecological data analysis and visualisation. Newcastle uponTyne, UK: Newcastle University. https://doi.org/10.1371/journal.pone.0040025
Kaiser, J., Schefuss, E., Lamy, F., Mohtadi, M., & Hebbeln, D. (2008).Glacial to Holocene changes in sea surface temperature andcoastal vegetation in north centralChile:High versus low latitudeforcing. Quaternary Science Reviews, 27, 2064–2075. https://doi.org/10.1016/j.quascirev.2008.08.025
Latorre, C., Betancourt, J. L., Rylander, K. A., & Quade, J. (2002).Vegetation invasions into absolute desert: A 45,000 yr rodentmidden record from the Calama‐Salar de Atacama basins, north‐ern Chile (lat 22–24°S).Geological Society of America Bulletin, 114,349–366. https://doi.org/10.1130/0016‐7606(2002)114<0349:VIIADA>2.0.CO;2
Latorre, C., Betancourt, J. L., Rylander, K. A., Quade, J., & Matthei,O. (2003). A vegetation history from the arid prepuna of north‐ern Chile (22–23°S) over the last 13,500 years. Palaeogeography, Palaeoclimatology, Palaeoecology, 194, 223–246. https://doi.org/10.1016/S0031‐0182(03)00279‐7
Lima,M.,Christie,D.A.,Santoro,M.C.,&Latorre,C. (2016).Coupledsocio‐environmental changes triggered indigenous aymara de‐population of the semiarid Andes of Tarapacá‐Chile during the
late 19th‐20th centuries. PLoS ONE, 11, e0160580. https://doi.org/10.1371/journal.pone.0160580
Marquet,P.A.,Bozinovic,F.,Bradshaw,G.A.,Cornelius,C.,Gonzalez,H.,Gutierrez,J.R.,…Jaksic,F.M.(1998).EcosystemsoftheAtacamaDesert and adjacent Andean area in northern Chile. Revista Chilena De Historia Natural,71,593–617.
Meserve,P.L.,&Glanz,W.E.(1978).Geographicalecologyofsmallmam‐mals in the northernChilean arid zone. Journal of Biogeography,5,135–148.https://doi.org/10.2307/3038168
Minvielle,M.,&Garreaud,R.D.(2011).Projectingrainfallchangesoverthe South American Altiplano. Journal of Climate, 24, 4577–4583.https://doi.org/10.1175/JCLI‐D‐11‐00051.1
Mujica,M.I.,Latorre,C.,Maldonado,A.,González‐Silvestre,L.,Pinto,R.,Pol‐Holz,R.,&Santoro,C.M.(2015).LateQuaternaryclimatechange,relictpopulationsandpresent‐dayrefugiainthenorthernAtacamaDesert:AcasestudyfromQuebradaLaHiguera (18°S).Journal of Biogeography, 42, 76–88. https://doi.org/10.1111/jbi.12383
Murray, D. C., Pearson, S. G., Fullagar, R., Chase, B. M., Houston, J.,Atchison, J., … Bunce, M. (2012). High‐throughput sequencing ofancient plant and mammal DNA preserved in herbivore middens.Quaternary Science Reviews,58,135–145.https://doi.org/10.1016/j.quascirev.2012.10.021
Paabo,S.,Poinar,H.,Serre,D.,Jaenicke‐Despres,V.,Hebler,J.,Rohland,N., … Hofreiter, M. (2004). Genetic analyses from ancient DNA.Annual Review of Genetics,38,645–679.https://doi.org/10.1146/an‐nurev.genet.37.110801.143214
Pachauri,R.K.,Allen,M.R.,Barros,V.R.,Broome,J.,Cramer,W.,Christ,R.,…vanYpserle,J.P.(2014).Climate change 2014: Synthesis report. Contribution of Working Groups I, II and III to the fifth assessment re‐port of the Intergovernmental Panel on Climate Change. Geneva,Switzerland:IPCC.
Parducci, L., Väliranta,M., Salonen, J. S., Ronkainen, T.,Matetovici, I.,Fontana, S. L., … Suyama, Y. (2015). Proxy comparison in ancientpeat sediments: Pollen, macrofossil and plant DNA. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences,370,20130382.
Pearson,S.,&Betancourt,J.L.(2002).Understandingaridenvironmentsusing fossil rodentmiddens. Journal of Arid Environments,50, 499–511.https://doi.org/10.1006/jare.2001.0901
Placzek,C.,Quade,J.,&Patchett,P.(2013).A130kareconstructionofrainfallontheBolivianAltiplano.Earth and Planetary Science Letters,363,97–108.https://doi.org/10.1016/j.epsl.2012.12.017
RCoreTeam(2018).R: A language and environment for statistical comput‐ing.Vienna,Austria:RFoundationforStatisticalComputing.
Ramsey, C. B. (1995). Radiocarbon calibration and analysis of stratig‐raphy: TheOxCal program.Radiocarbon,37, 425–430. https://doi.org/10.1017/S0033822200030903
Rawlence,N.J.,Lowe,D.J.,Wood,J.R.,Young,J.M.,Churchman,G.J.,Huang,Y.T.,&Cooper,A.(2014).UsingpalaeoenvironmentalDNAtoreconstructpastenvironments:Progressandprospects.Journal of Quaternary Science,29,610–626.https://doi.org/10.1002/jqs.2740
Root, T. L., Price, J. T.,Hall, K. R., Schneider, S.H., Rosenzweig,C.,&Pounds,J.A.(2003).Fingerprintsofglobalwarmingonwildanimalsandplants.Nature,421,57.https://doi.org/10.1038/nature01333
Sarricolea, P., Meseguer Ruiz, O., & Romero‐Aravena, H. (2017).TendenciasdelaprecipitaciónenelnortegrandedeChileysurel‐ación con las proyecciones de cambio climático. Diálogo andino,41‐50.
Schulze‐Makuch, D., Wagner, D., Kounaves, S. P., Mangelsdorf, K.,Devine,K.G.,deVera, J.P.,…Zamorano,P. (2018).Transitorymi‐crobialhabitat inthehyperaridAtacamaDesert.Proceedings of the National Academy of Sciences of the United States of America, 115,2670–2675.https://doi.org/10.1073/pnas.1714341115
Taberlet,P.,Coissac,E.,Pompanon,F.,Brochmann,C.,&Willerslev,E.(2012). Towards next‐generation biodiversity assessment usingDNA metabarcoding. Molecular Ecology,21,2045–2050.https://doi.org/10.1111/j.1365‐294X.2012.05470.x
Thibeault,J.,Seth,A.,&Wang,G. (2012).Mechanismsofsummertimeprecipitation variability in the Bolivian Altiplano: Present and fu‐ture. International Journal of Climatology,32,2033–2041.https://doi.org/10.1002/joc.2424
Thompson, L. G. (2000). Ice core evidence for climate change in theTropics:Implicationsforourfuture.Quaternary Science Reviews,19,19–35.https://doi.org/10.1016/S0277‐3791(99)00052‐9
Vuille,M.,Bradley,R.S.,&Keimig,F. (2000). Interannual climatevari‐ability in theCentralAndesand its relation to tropicalPacific andAtlantic forcing. Journal of Geophysical Research‐Atmospheres, 105,12447–12460.https://doi.org/10.1029/2000JD900134
Whittaker, R. J., Willis, K. J., & Field, R. (2001). Scale and spe‐cies richness: Towards a general, hierarchical theory of spe‐cies diversity. Journal of Biogeography, 28, 453–470. https://doi.org/10.1046/j.1365‐2699.2001.00563.x
Willerslev, E., & Cooper, A. (2005). Ancient DNA. Proceedings of the Royal Society of London B: Biological Sciences,272,3–16.https://doi.org/10.1098/rspb.2004.2813
Willerslev, E., Hansen, A. J., Binladen, J., Brand, T. B., Gilbert, M. T.,Shapiro,B.,…Cooper,A. (2003).Diverseplantandanimalgeneticrecords from Holocene and Pleistocene sediments. Science, 300,791–795.https://doi.org/10.1126/science.1084114
Williams, J. W., & Jackson, S. T. (2007). Novel climates, no‐analogcommunities, and ecological surprises. Frontiers in Ecology and the Environment,5,475–482.https://doi.org/10.1890/070037
Wolter, K., & Timlin, M. S. (2011). El Niño/Southern Oscillation be‐havioursince1871asdiagnosedinanextendedmultivariateENSOindex(MEI.ext).International Journal of Climatology,31,1074–1087.https://doi.org/10.1002/joc.2336
Wood, J. R., Díaz, F. P., Latorre, C., Wilmshurst, J. M., Burge, O. R.,& Gutiérrez, R. A. (2018). Plant pathogen responses to LatePleistocene and Holocene climate change in the central AtacamaDesert,Chile.Scientific Reports,8,17208.https://doi.org/10.1038/s41598‐018‐35299‐2
Wood,J.R.,Wilmshurst,J.M.,Richardson,S.J.,Rawlence,N.J.,Wagstaff,S. J.,Worthy, T.H., & Cooper, A. (2013). Resolving lost herbivorecommunity structure using coprolites of four sympatric moa spe‐cies(Aves:Dinornithiformes).Proceedings of the National Academy of Sciences of the United States of America,110,16910–16915.
Additional supporting information may be found online in theSupportingInformationsectionattheendofthearticle.
How to cite this article:DíazFP,LatorreC,Carrasco‐PugaG,etal.Multiscaleclimatechangeimpactsonplantdiversityinthe Atacama Desert. Glob Change Biol. 2019;00:1–13. https://doi.org/10.1111/gcb.14583