Marc Derive Youcef Bouazza Corentine Alauzet Se ´bastien Gibot Myeloid-derived suppressor cells control microbial sepsis Received: 18 October 2011 Accepted: 28 February 2012 Published online: 3 May 2012 Ó Copyright jointly held by Springer and ESICM 2012 This article is discussed in the editorial available at: doi:10.1007/s00134-012-2575-3. Electronic supplementary material The online version of this article (doi:10.1007/s00134-012-2574-4) contains supplementary material, which is available to authorized users. M. Derive Á Y. Bouazza Á S. Gibot Groupe Choc, contrat Avenir INSERM, Faculte ´ de Me ´decine, Nancy Universite ´, Nancy, France C. Alauzet EA 4369, Laboratoire de Bacte ´riologie, Faculte ´ de Me ´decine, Nancy Universite ´, Nancy, France S. Gibot ( ) ) Service de Re ´animation Me ´dicale, Ho ˆpital Central, CHU Nancy, 29 boulevard du Mare ´chal de Lattre de Tassigny, 54035 Nancy Cedex, France e-mail: [email protected]Tel.: ?33-3-83852970 Fax: ?33-3-83858511 Abstract Purpose: To investigate the role of myeloid-derived suppres- sor cells (MDSCs) during sepsis in mice. MDSCs are a heterogeneous population of cells that expand during cancer, inflammation and infection. These cells, by their ability to sup- press T lymphocyte proliferation, regulate immune responses during various diseases. Their role during microbial infections is scarcely known. Methods: Septic shock was induced by caecal ligation and punc- ture in adult male BALB/c mice; sham-operated animals served as controls. Animals were killed under anaesthesia to harvest blood and organs. Results: Polymicrobial sep- sis induced a progressive accumulation of MDSCs in spleens that were found to be enlarged in surviving mice. MDSCs harvested at day 10 after the onset of infection were highly responsive to LPS in terms of cytokines secretion, NF-kB activation, ROS production and argi- nase I activity, whereas early- appearing (day 3) MDSCs poorly responded to this stimulus. By con- trast, both day 3 and day 10 MDSCs were able to inhibit T cell prolifera- tion. Adoptive transfer of day 10 MDSCs to septic mice attenuated peritoneal cytokine production, increased bacterial clearance and dramatically improved survival rate. Conclusion: These results provide new information on the role of MDSCs, suggesting a protective effect during sepsis. Pharmacologic agents known to promote the expan- sion of MDSCs should thus be further studied for sepsis treatment. Keywords Experimental septic shock Á CLP model Á Myeloid-derived suppressor cells Abbreviations AT Adoptive transfer CLP Caecal ligation and puncture GSK Glycogen synthase kinase IL Interleukin LPS Lipopolysaccharide MDSC Myeloid-derived suppressor cells MO-MDSC Macrophage-like MDSC PMN-MDSC Neutrophil-like MDSC ROS Reactive oxygen species TLR Toll-like receptor Intensive Care Med (2012) 38:1040–1049 DOI 10.1007/s00134-012-2574-4 EXPERIMENTAL
Transcript
Marc DeriveYoucef BouazzaCorentine AlauzetSebastien Gibot
Received: 18 October 2011Accepted: 28 February 2012Published online: 3 May 2012� Copyright jointly held by Springer andESICM 2012
This article is discussed in the editorialavailable at: doi:10.1007/s00134-012-2575-3.
Electronic supplementary materialThe online version of this article(doi:10.1007/s00134-012-2574-4) containssupplementary material, which is availableto authorized users.
M. Derive � Y. Bouazza � S. GibotGroupe Choc, contrat Avenir INSERM,Faculte de Medecine, Nancy Universite,Nancy, France
C. AlauzetEA 4369, Laboratoire de Bacteriologie,Faculte de Medecine, Nancy Universite,Nancy, France
S. Gibot ())Service de Reanimation Medicale,Hopital Central, CHU Nancy,29 boulevard du Marechal de Lattre deTassigny, 54035 Nancy Cedex, Francee-mail: [email protected].: ?33-3-83852970Fax: ?33-3-83858511
Abstract Purpose: To investigatethe role of myeloid-derived suppres-sor cells (MDSCs) during sepsis inmice. MDSCs are a heterogeneouspopulation of cells that expand duringcancer, inflammation and infection.These cells, by their ability to sup-press T lymphocyte proliferation,regulate immune responses duringvarious diseases. Their role duringmicrobial infections is scarcelyknown. Methods: Septic shock wasinduced by caecal ligation and punc-ture in adult male BALB/c mice;sham-operated animals served ascontrols. Animals were killed underanaesthesia to harvest blood andorgans. Results: Polymicrobial sep-sis induced a progressiveaccumulation of MDSCs in spleensthat were found to be enlarged insurviving mice. MDSCs harvested atday 10 after the onset of infectionwere highly responsive to LPS interms of cytokines secretion, NF-kBactivation, ROS production and argi-nase I activity, whereas early-appearing (day 3) MDSCs poorlyresponded to this stimulus. By con-trast, both day 3 and day 10 MDSCswere able to inhibit T cell prolifera-tion. Adoptive transfer of day 10MDSCs to septic mice attenuatedperitoneal cytokine production,
increased bacterial clearance anddramatically improved survival rate.Conclusion: These results providenew information on the role ofMDSCs, suggesting a protectiveeffect during sepsis. Pharmacologicagents known to promote the expan-sion of MDSCs should thus be furtherstudied for sepsis treatment.
Keywords Experimental septicshock � CLP model � Myeloid-derivedsuppressor cells
Septic shock, a complex clinical syndrome resulting froma harmful and damaging host response to infection, is theleading cause of mortality in intensive care units [1].Activation of innate immune cells is critical for appro-priate control of invading microorganisms, but theirhyper-responsiveness may lead to production of an excessof proinflammatory mediators that play a role in sepsis-associated multi-organ failure development [2]. A tightcontrol of innate immune response is therefore required.
Myeloid-derived suppressor cells (MDSCs) are a het-erogeneous population of cells able to suppress T cellresponses [3]. Although their definition remains debat-able, most investigators subdivide these cells into twodifferent subsets based on their expression of differentsurface markers (CD11b, Gr-1, Ly6G and Ly6C) [4, 5].‘Macrophage-like MDSCs’ (MO-MDSC) are defined asCD11b?Ly6GlowLy6Chigh and inhibit T cell proliferationthrough an iNOS mechanism. ‘Neutrophil-like MDSCs’(PMN-MDSCs) are CD11b?Ly6GhighLy6Clow cells andshow immunomodulating properties through arginase Iand interferon-c activities [6].
Besides their well-known role in tumour immuneevasion, a growing body of evidence supports a role ofMDSCs during other pathological disorders such astrauma [7], inflammation [8], burn [9] and autoimmunediseases [10]. The role of MDSC has been investigatedduring parasitic or viral infections [11–16]. All thesestudies showed immunosuppressive functions of MDSCs;however, these cells may nevertheless confer protectiveimmunity.
It was recently reported that MDSCs accumulate in thespleens of mice during polymicrobial sepsis and may havepotent host-protective anti-inflammatory functions [17].However, to what extent MDSCs modify the course of thedisease and if they critically contribute to immune regu-lation during sepsis remain unclear [18].
Here we confirm that MDSCs progressively accumu-late in spleens from septic mice. We observed that thesecells respond differently to TLR4 or IL6 stimulationdepending on the time of their appearance after the onsetof the septic insult. Importantly, adoptive transfer ofMDSCs protected septic mice from death by promotingmicrobial killing.
Materials and methods
Caecal ligation and puncture (CLP) polymicrobialsepsis model
Experiments were approved by our institutional animalcare and use committee. Male Balb/c mice (6–8 weeks)were anaesthetized with isoflurane. CLP was performed
as previously described [19]. After surgery, all mice wereinjected subcutaneously with 0.5 mL of 0.9 % NaClsolution for fluid resuscitation. Three, 7, 10 or 15 daysafter CLP, spleens were dissected under anaesthesia toallow the isolation of MDSCs. For adoptive transferexperiments, the animals were randomly grouped andtreated by intraperitoneal administration of 106 MDSCs orGr1- control cells in 250 lL 0.9 % NaCl solution 2 hafter the surgery, and then monitored for survival. Fiveadditional animals per group were killed under anaes-thesia at 24 h after CLP to allow the determination ofbacterial count, cytokine levels and organ samples. Peri-toneal lavage fluid was obtained and blood was collectedby cardiac puncture.
MDSCs were isolated from 70 lM filtered total spleniccells by magnetic separation of CD11b?/Gr1? cells (anti-mouse Gr1-APC from BDBiosciences, Le-pont-de-claix,France; MACS cell separator, CD11b microbeads andanti-APC microbeads from Miltenyi Biotech, Paris,France). MDSCs were then analysed for Ly6G and Ly6Cexpression by flow cytometry (anti-mouse Ly6C-FITCand anti-mouse Ly6G-PE from BDBiosciences) as well asfor TREM-1 expression (anti-mouse TREM-1, RnDSystems) and cultured in MDSCs medium (RPMI, 10 %inactivated SVF, 20 ng/mL GM-CSF, 1 mL sodiumpyruvate, 0.1 mM non-essential AA, 2 mM L-glutamine,100 U/mL penicillin, 100 mg/mL streptomycin, 10 mMHepes, 5 9 10-5 M 2-ME) with or without lipopolysac-charide (LPS) (100 ng/mL) or IL6 (10 ng/mL) atindicated times for further analysis. When indicated,splenic Gr1- cells were used as controls. In another set ofexperiments, MDSCs were similarly purified from bonemarrow of healthy mice and cultured with or without100 ng/mL LPS (Escherichia coli, O111:B4). Briefly,femurs were dissected and bone marrow flushed understerile conditions. Cells were first depleted in erythroidcells and progenitors by the use of Ter-119 microbeads(Miltenyi Biotech). Then magnetic selection of CD11b?/Gr1? was performed and the cells were cultured inMDSCs medium.
Cytokine concentration measurements
Cytokine concentration measurements in supernatants orplasma were done by ELISA (mouse Quantikine ELISAkits, R&D Systems, Lille, France) and cytokine panelassays (Proteome Profiler Mouse Cytokine Array Kit,Panel A, R&D Systems) according to the manufacturers’recommendations.
1041
ROS production and arginase activity quantification
FACS quantification of intracellular reactive oxygenspecies (ROS) production was assessed after 2 h stimu-lation by the use of DCFDA, a fluorogenic substrate(Fisher Scientific, Illkirch, France), according to manu-facturer’s instructions. Arginase activity was measuredafter 2 h stimulation with the QuantiChromTM arginaseassay kit (Gentaur, Paris, France).
Proliferation assays
MDSCs were co-incubated with carboxyfluoresceinsuccinimidyl ester (CFSE)-stained and phytohaemagglu-tinin (PHA)-stimulated lymphocytes (CFSE is partitionedequally among daughter cells with each division). Pro-liferation rate was then estimated by flow cytometry andratio comparison between proliferating/non-proliferatinglymphocytes with or without MDSCs on the fourthgeneration.
Protein phosphorylation analysis
Stimulated MDSCs were lysed in PhosphoSafe extractionreagent (Novagen, Merck Biosciences, Nottingham, UK)and centrifuged for 5 min at 16,0009g at 4 �C to collectthe supernatant. Protein concentration was determinedaccording to Bradford’s method (BCA Protein Assay Kit,Pierce, ThermoScientific, Brebieres, France). Lysateswere then analysed by Western blots (Criterion XT Bis–Tris Gel, 4–12 %, BioRad, Marnes-la-Coquette, France,and PVDF membrane, Millipore, Saint-Quentin en Yve-lines, France), which were revealed with anti-phospho-STAT3, anti-phospho-ERK1/2, anti-phospho-p38, anti-phospho-GSK-3a/b, or anti-socs3 and the correspondingsecondary antibody conjugated to horseradish peroxidase(Cell Signaling, Ozyme, Saint-Quentin en Yvelines,France) and SuperSignal West Femto Substrate (Pierce,ThermoScientific). Anti-STAT3, anti-ERK1/2, anti-p38and anti-GSK-3a/b (Cell Signaling) were used for nor-malization. Acquisition and quantitative signal densityanalyses were performed by a LAS-4000 imager (FSVT,Courbevoie, France) and Multi-Gauge software (Life-Science Fujifilm, France).
NF-jB activity measurement
MDSCs were collected after 20 min stimulation, nuclearextracts obtained by a nuclear extraction kit and NF-jBactivity measured with an NF-jB p50/p65 EZ-TFAtranscription factor assay (Colorimetric) (Millipore)according to the manufacturer’s instructions.
RT-qPCR
Total RNAs were extracted from isolated cells usingRNeasy Plus Mini Kit (Qiagen, Courtaboeuf, France) andquantified with NanoDrop (ThermoScientific) beforebeing retrotranscripted using the iScript cDNA synthesiskit (BioRad) and quantified by quantitative PCR usingprobes (Quantitect Primers, Qiagen) for mIL6, mIL10,mTNF-a and mActB. Alternatively, total RNAs wereretrotranscripted with RT2 First Strand Kit (SABio-sciences, Tebu-bio, Le Perray-en-Yvelines, France) forPCR arrays (mouse innate and adaptive immune respon-ses RT2 profiler PCR arrays, SABiosciences). All PCRswere performed in a MyiQ thermal cycler and quantifiedby iQ5 software (Qiagen). All thermal conditions werechosen following manufacturers’ instructions. PCR arrayresults were analysed using PCR array data analysissoftware (SABiosciences) and normalized with fivehousekeeping genes.
Preparation of Il10 knock-down MDSCs
MDSCs were transfected with IL10 shRNA (SureSilenc-ing shRNA plasmid, SABiosciences) using Lipofectamine(Life technologies, Courtaboeuf, France) accordinglywith manufacturer’s instructions. IL10 expression wassubsequently assessed by quantitative RT-PCR 24 h aftertransfection and before adoptive transfer to septic mice.Ninety per cent IL10 knock-down was achieved.
Bacterial count
Blood and crushed spleen were plated in serial log dilu-tions on tryptic soy supplemented with 5 % sheep bloodagar plates. After plating, tryptic soy agar plates wereincubated at 37 �C aerobically for 24 h and anaerobicallyfor 48 h. Results are expressed as CFU per millilitre ofblood or per gram of spleen.
Killing assays
Isolated MDSCs (106 cells) were co-incubated with 107
Escherichia coli ATCC 25922 (multiplicity of infection,10), with or without whole blood (cells/white blood cellseq.), in complete RPMI for 1 h, 37 �C, 5 % CO2 after ashort centrifugation to synchronize the reaction. Bacteriawithout cells or with 200 lL of whole blood were usedrespectively as negative and positive controls. After 1 hincubation, bacteria growth was stopped on ice andsupernatants were then immediately plated by serialdilutions and incubated overnight at 37 �C fornumeration.
1042
Statistical analysis
All data, unless indicated, are presented as mean ± SDand the statistical significance of differences betweengroups was analysed using Student’s t test or ANOVAand post hoc Dunnett’s test when appropriate. Kaplan–Meier survival curves were analysed using the logrank test. A p value less than 0.05 was deemedsignificant.
Results
Sepsis induces the recruitment of MDSCs whoseresponse to stimulation depends on the durationof infection
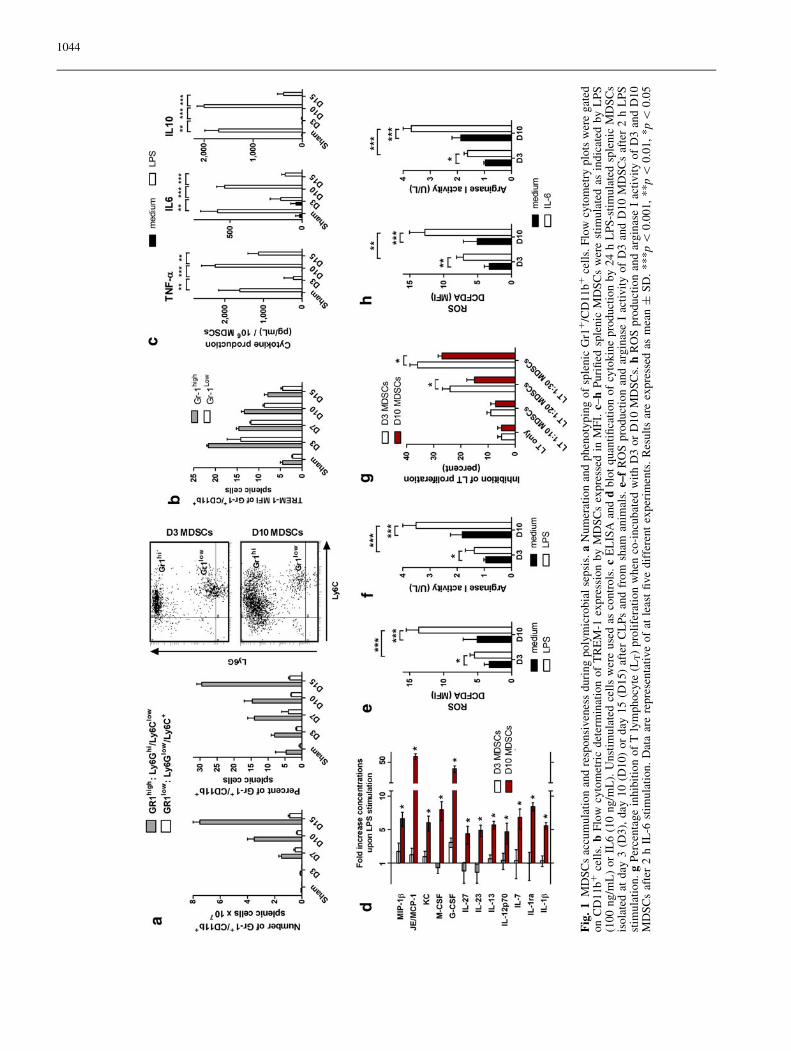
Polymicrobial sepsis induced a progressive accumulationof MDSCs, mainly PMN-MDSC (CD11b?Ly6Ghigh
Ly6Clow subset of cells) in spleens that were found to beenlarged in surviving mice (Fig. 1a). These cells alsoexpressed the triggering receptor expressed on myeloidcells-1, a myeloid lineage receptor [20], on their surface(Fig. 1b). To investigate how MDSCs respond to LPS, aTLR4 agonist, septic mice were killed 3, 10 or 15 daysafter the onset of sepsis and splenic MDSCs harvested.Although basal production of TNF-a, IL6 and IL10was similar between day 3, day 10, or day 15 MDSCs,late-onset cells were highly responsive to LPS whereasearly-onset or MDSCs obtained from sham (non-septic)animals poorly responded to this stimulus (Fig. 1c).These findings were extended to several other cytokines(Fig. 1d).
MDSCs are thought to mediate their activity througharginase I upregulation and ROS production [5]. Whereasday 10 MDSCs increased both their ROS production andarginase I activity after LPS stimulation, day 3 MDSCsremained poorly responsive (Fig. 1e, f). By contrast, bothday 3 and day 10 MDSCs were able to migrate through aMIP-2 or SDF1 gradient (not shown) and to inhibit T cellproliferation (Fig. 1g), though to a lesser extent for day 10cells.
Besides their activation through TLR4, MDSCs can beactivated through a different group of factors like cyto-kines [3, 21]. Interleukin-6 has been shown to play animportant role in MDSC expansion and activation [22].As sepsis is associated with elevated concentrations ofIL6, we tested the reactivity of MDSCs to this cytokine.Both early-onset (day 3) and late-onset MDSCs (day 10)responded to IL6 stimulation (arginase I activity and ROSproduction, Fig. 1h) in a similar way as they responded toLPS.
These data thus suggest that MDSCs respond differ-ently to TLR4 or IL6 stimulation depending on the timeof their appearance after the onset of the septic insult.
Effect of ex vivo LPS and IL-6 stimulationon intracellular signaling and gene expressionin MDSCs from septic mice
To further elucidate this discordant responsivenessbetween early- and late-accumulating MDSCs, LPS- andIL6-induced activation of three key intracellular proteinswas analysed. Stat3 activation has been shown to beessential for mobilization and tissue accumulation ofMDSCs [23], as well as for their suppressive activity [24].
LPS stimulation only induced Stat3 phosphorylation inday 3 MDSCs. Moreover, whereas IL6 induced an earlyand sustained Stat3 activation in day 3 MDSCs, day 10cells only showed a transient Stat3 phosphorylation(Fig. 2a). As expected, the expression of socs3 wasinversely correlated with the intensity of Stat3 phos-phorylation [25]. This suggests that upon inflammatorystimulation, day 3 MDSCs are more mobilizable andimportantly more immunosuppressive than day 10 cells,as also suggested by their higher capacity to inhibit T cellproliferation (Fig. 1g).
Day 3 MDSCs were found to basally express a highlevel of ERK1/2 activation, which may drive theirexpansion [24], whereas neither LPS nor IL6 was able toincrease this activation (Fig. 2b). By contrast, ERK1/2only phosphorylated in day 10 MDSCs upon LPS stim-ulation thus mediating the LPS-induced production ofcytokines. The same pattern of expression was observedfor p38, another MAPK.
Glycogen synthase kinase 3 (GSK3) plays a pivotalrole in regulating the production of pro- and anti-inflammatory cytokines. In innate immune cells, GSK3inactivation (phosphorylation) suppresses the productionof cytokines [26]. Here we found that while only a lowand constant level of GSK3 phosphorylation existed inday 3 MDSCs, GSK3 dephosphorylated upon LPS stim-ulation in day 10 cells thus possibly participating in thede-repression of cytokines production (Fig. 2c).
Finally, LPS-mediated NF-jB activation (Fig. 2d) wasimpaired in day 3 MDSCs as compared to day 10 cellsand this translated into a decreased induction of severalgenes involved in the inflammatory response, especiallyTNF, interleukins, and interferons (Fig. 2e–f and Sup-plementary Table).
Therefore MDSCs appearing early (day 3) after sepsisare prone to expansion but are not fully able to be acti-vated, whereas MDSCs arising late (day 10) are lessexpandable but are responsive to activation factors.
We next sought to investigate the role of MDSCs duringCLP-induced polymicrobial sepsis in mice. Adoptivelytransferred (AT) MDSCs persisted at least 24 h
1043
Fig
.1
MD
SC
sac
cum
ula
tio
nan
dre
spo
nsi
ven
ess
du
rin
gp
oly
mic
rob
ial
sep
sis.
aN
um
erat
ion
and
ph
eno
typ
ing
of
sple
nic
Gr1
?/C
D1
1b
?ce
lls.
Flo
wcy
tom
etry
plo
tsw
ere
gat
edo
nC
D1
1b
?ce
lls.
bF
low
cyto
met
ric
det
erm
inat
ion
of
TR
EM
-1ex
pre
ssio
nb
yM
DS
Cs
exp
ress
edin
MF
I.c–
hP
uri
fied
sple
nic
MD
SC
sw
ere
stim
ula
ted
asin
dic
ated
by
LP
S(1
00
ng
/mL
)o
rIL
6(1
0n
g/m
L).
Un
stim
ula
ted
cell
sw
ere
use
das
con
tro
ls.
cE
LIS
Aan
dd
blo
tquan
tifi
cati
on
of
cyto
kin
epro
duct
ion
by
24
hL
PS
-sti
mula
ted
sple
nic
MD
SC
sis
ola
ted
atd
ay3
(D3
),d
ay1
0(D
10
)o
rd
ay1
5(D
15
)af
ter
CL
Ps
and
fro
msh
aman
imal
s.e–
fR
OS
pro
du
ctio
nan
dar
gin
ase
Iac
tiv
ity
of
D3
and
D1
0M
DS
Cs
afte
r2
hL
PS
stim
ula
tio
n.
gP
erce
nta
ge
inh
ibit
ion
of
Tly
mp
ho
cyte
(LT)
pro
life
rati
on
when
co-i
ncu
bat
edw
ith
D3
or
D10
MD
SC
s.h
RO
Sp
rod
uct
ion
and
arg
inas
eI
acti
vit
yo
fD
3an
dD
10
MD
SC
saf
ter
2h
IL-6
stim
ula
tio
n.
Dat
aar
ere
pre
sen
tati
ve
of
atle
ast
fiv
ed
iffe
ren
tex
per
imen
ts.
Res
ult
sar
eex
pre
ssed
asm
ean
±S
D.
**
*p\
0.0
01
,*
*p\
0.0
1,
*p\
0.0
5
1044
Fig
.2
Eff
ect
of
exv
ivo
LP
San
dIL
-6st
imu
lati
on
on
intr
acel
lula
rsi
gn
alin
gan
dg
ene
exp
ress
ion
inM
DS
Cs
fro
mse
pti
cm
ice.
a–f
Sp
len
icM
DS
Cs
wer
est
imu
late
das
ind
icat
edb
yL
PS
(10
0n
g/m
L)
or
IL6
(10
ng
/mL
).U
nst
imu
late
dce
lls
wer
eu
sed
asco
ntr
ols
.a
–c
Wes
tern
blo
tan
aly
sis
of
tim
e-d
epen
den
tS
TA
T3
,S
OC
S3
,E
RK
-1/2
and
p3
8p
ho
sph
ory
lati
on
.c
Wes
tern
blo
tan
aly
sis
of
GS
K-3
a/b
ph
osp
ho
ryla
tio
naf
ter
30
min
LP
S.
dE
LIS
Aq
uan
tifi
cati
on
of
nu
clea
rN
F-j
Bsu
bu
nit
sp
50
and
p6
5.
eT
ime
cou
rse
of
TN
F-a
,IL
6an
dIL
10
exp
ress
ion
.f
Ex
pre
ssio
no
fse
lect
edin
flam
mat
ory
gen
esaf
ter
6h
LP
Sst
imu
lati
on
.D
ata
are
rep
rese
nta
tiv
eo
fat
leas
tfi
ve
dif
fere
nt
exp
erim
ents
exce
pt
for
fth
atsu
mm
ariz
esth
ree
dif
fere
nt
exp
erim
ents
.R
esu
lts
are
exp
ress
edas
mea
n±
SD
.*
**p\
0.0
01
,*
*p\
0.0
1,
*p\
0.0
5
1045
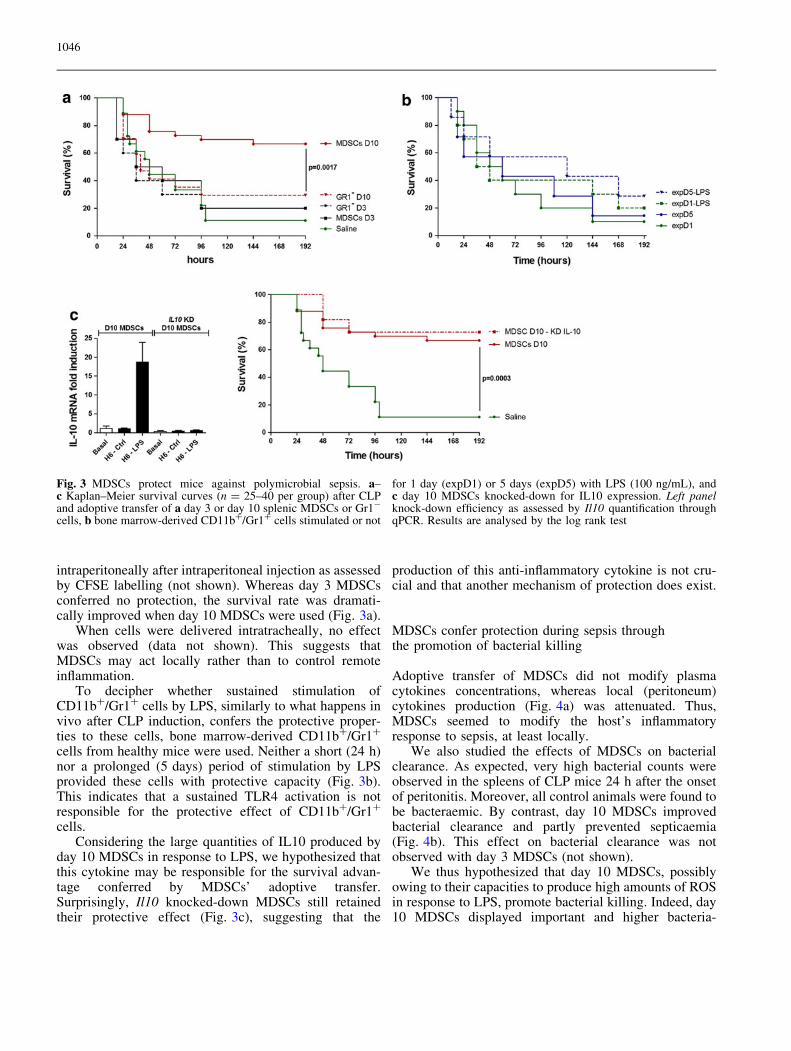
intraperitoneally after intraperitoneal injection as assessedby CFSE labelling (not shown). Whereas day 3 MDSCsconferred no protection, the survival rate was dramati-cally improved when day 10 MDSCs were used (Fig. 3a).
When cells were delivered intratracheally, no effectwas observed (data not shown). This suggests thatMDSCs may act locally rather than to control remoteinflammation.
To decipher whether sustained stimulation ofCD11b?/Gr1? cells by LPS, similarly to what happens invivo after CLP induction, confers the protective proper-ties to these cells, bone marrow-derived CD11b?/Gr1?
cells from healthy mice were used. Neither a short (24 h)nor a prolonged (5 days) period of stimulation by LPSprovided these cells with protective capacity (Fig. 3b).This indicates that a sustained TLR4 activation is notresponsible for the protective effect of CD11b?/Gr1?
cells.Considering the large quantities of IL10 produced by
day 10 MDSCs in response to LPS, we hypothesized thatthis cytokine may be responsible for the survival advan-tage conferred by MDSCs’ adoptive transfer.Surprisingly, Il10 knocked-down MDSCs still retainedtheir protective effect (Fig. 3c), suggesting that the
production of this anti-inflammatory cytokine is not cru-cial and that another mechanism of protection does exist.
MDSCs confer protection during sepsis throughthe promotion of bacterial killing
Adoptive transfer of MDSCs did not modify plasmacytokines concentrations, whereas local (peritoneum)cytokines production (Fig. 4a) was attenuated. Thus,MDSCs seemed to modify the host’s inflammatoryresponse to sepsis, at least locally.
We also studied the effects of MDSCs on bacterialclearance. As expected, very high bacterial counts wereobserved in the spleens of CLP mice 24 h after the onsetof peritonitis. Moreover, all control animals were found tobe bacteraemic. By contrast, day 10 MDSCs improvedbacterial clearance and partly prevented septicaemia(Fig. 4b). This effect on bacterial clearance was notobserved with day 3 MDSCs (not shown).
We thus hypothesized that day 10 MDSCs, possiblyowing to their capacities to produce high amounts of ROSin response to LPS, promote bacterial killing. Indeed, day10 MDSCs displayed important and higher bacteria-
Fig. 3 MDSCs protect mice against polymicrobial sepsis. a–c Kaplan–Meier survival curves (n = 25–40 per group) after CLPand adoptive transfer of a day 3 or day 10 splenic MDSCs or Gr1-
cells, b bone marrow-derived CD11b?/Gr1? cells stimulated or not
for 1 day (expD1) or 5 days (expD5) with LPS (100 ng/mL), andc day 10 MDSCs knocked-down for IL10 expression. Left panelknock-down efficiency as assessed by Il10 quantification throughqPCR. Results are analysed by the log rank test
1046
killing properties than day 3 MDSCs, alone or in com-bination with whole blood (Fig. 4c). Contribution of ROSproduction versus phagocytic activity was evaluated bypre-incubating MDSCs with 5 lg/mL of cytochalasin D(an inhibitor of phagocytosis): this treatment did notmodify the results significantly (not shown), thus sug-gesting that ROS production by day 10 MDSCs mayparticipate in the improvement of bacterial clearance.
Discussion
During microbial sepsis, neutrophils and macrophages arelargely responsible for the killing of invading microor-ganisms, but they may need the help of other immunecells.
MDSCs are a heterogeneous population of immaturemyeloid cells characterized by their ability to suppressantigen-specific T cell activation through depletion ofextracellular arginine, IL10 production, etc [3, 21].MDSCs accumulate during chronic or acute inflammatoryconditions like cancer [27], autoimmune diseases [10],trauma [7], burns [9] and sepsis [17]. Because of theirimmunosuppressive properties, MDSCs have been con-sidered to be deleterious, in promoting tumour growth,metastasis generation, and secondary immunosuppressionafter physical injury [3, 7, 28]. However, other importantproperties of these cells, such as increased ROS produc-tion and inflammatory cytokines secretion, as well as theircapacity to expand following virtually all inflammatorydisorders, suggest that MDSCs may be a physiologic andbeneficial component of the inflammatory response.Several published pieces of evidence indirectly supportthis hypothesis. Indeed, although MDSC expansion was
efficiently blocked during polymicrobial (CLP) sepsis inmice through the use of gemcitabine or Gr1 antibody,such an approach led to worse outcomes [18]. The sameconclusions were reached by Sander et al. [23] whoshowed that septic mice lacking gp130 and unable tosignal through IL-6 failed to expand their MDSC popu-lation and had markedly higher mortalities associatedwith increased inflammatory cytokine production. Takingthe opposite strategy, we found a net survival advantageconferred by the adoptive transfer of MDSCs, furthersupporting a beneficial role of these cells during sepsis.Nevertheless, to be fully effective in this model, weobserved that MDSCs needed to be stimulated for a longtime: administration of MDSCs harvested 3 days after theonset of infection did not show any effect sharply con-trasting with the protection conferred by MDSCs obtainedat day 10. We were unable to recapitulate the protectiveeffect of MDSCs when CD11b?/Gr1? cells were obtainedfrom bone marrow of healthy mice, even after 5-daystimulation by LPS. Indeed, we could not be sure thatthese bone marrow-derived cells really were MDSCs aswe lack specific surface markers to precisely differentiatethem from normal myeloid progenitors. Furthermore, a5-day period may have been too short, but unfortunatelywe were unable to cultivate these cells for a longer periodof time. This mechanism of defence is therefore notimmediate and may mainly be involved in the preventionof the secondary infections that frequently arise after aninitial septic insult [29]. Indeed, when MDSC expansionwas inhibited by gemcitabine, burned mice were renderedhighly susceptible to secondary Pseudomonas aeruginosainfections [30]. Supporting these data, Delano et al. [31]recently showed that mice were rendered extremely sen-sitive to P. aeruginosa pneumonia 3 days after peritonitis,whereas this susceptibility was lost by day 7 post CLP,
Fig. 4 MDSCs confer protection during sepsis through the promo-tion of bacterial killing. a Plasma and peritoneal lavage fluid (PLF)cytokines concentrations 24 h after CLP. b Bacterial counts inspleen and blood obtained 24 h after CLP and adoptive transfer.Results are expressed as CFU per ml of blood or per gram of
spleen. c Quantification of in vitro MDSCs bacterial killing activity,alone or in combination with whole blood (cells/white blood cellseq.). Results are expressed as mean ± SD. *p \ 0.05, **p \ 0.01,***p \ 0.001 compared with controls or between the indicatedgroups
1047
coinciding with repopulation of Gr1?/CD11b? cells. Herewe observed that MDSCs promote microbial killing bothin vivo (reduction of bacterial spreading) and in vitro(killing assays). These findings are in line with those ofPereira et al. [14] who showed that MDSCs were able tokill Leishmania major through a NO-dependent mecha-nism. Whether the properties of MDSCs are also modifieddepending on the duration of the stimulus during othernon-septic disorders remains to be determined.
Although encouraging, our experiments should nev-ertheless be viewed with caution as the phenotype ofMDSCs is not robust enough to allow for a preciseselection. It may thus have been possible that some other
cells rather than MDSCs have been adoptivelytransferred.
Contrasting with the accepted deleterious role ofMDSCs during cancer in promoting cell growth andmetastasis through their immunosuppressive properties,we showed here a protective role of these cells duringpolymicrobial sepsis. Pharmacologic agents known topromote the expansion of MDSCs during cancer such asgrowth factors, chemokines, etc. [3] should thus be furtherstudied for sepsis treatment.
Acknowledgments We are indebted to Chantal Montemont, Anne-Marie Carpentier and Kevin Patron for technical assistance.
References
1. Martin GS, Mannino DM, Eaton S, MossM (2003) The epidemiology of sepsis inthe United States from 1979 through2000. N Engl J Med 348:1546–1554
2. Hotchkiss RS, Karl IE (2003) Thepathophysiology and treatment ofsepsis. N Engl J Med 348:138–150
3. Gabrilovich DI, Nagaraj S (2009)Myeloid-derived suppressor cells asregulators of the immune system. NatRev Immunol 9:162–174
4. Movahedi K, Guilliams M, Van denBossche J, Van den Bergh R, GysemansC, Beschin A, De Baetselier P, VanGinderachter JA (2008) Identificationof discrete tumor-induced myeloid-derived suppressor cell subpopulationswith distinct T cell-suppressive activity.Blood 111:4233–4244
5. Youn JI, Nagaraj S, Collazo M,Gabrilovich DI (2008) Subsets ofmyeloid-derived suppressor cells intumor-bearing mice. J Immunol181:5791–5802
6. Peranzoni E, Zilio S, Marigo I, DolcettiL, Zanovello P, Mandruzzato S, BronteV (2010) Myeloid-derived suppressorcell heterogeneity and subset definition.Curr Opin Immunol 22:238–244
8. Bunt SK, Sinha P, Clements VK, LeipsJ, Ostrand-Rosenberg S (2006)Inflammation induces myeloid-derivedsuppressor cells that facilitate tumorprogression. J Immunol 176:284–290
9. Noel G, Wang Q, Schwemberger S,Hanson C, Giacalone N, Haar L, OgleCK (2011) Neutrophils, not monocyte/macrophages, are the major splenicsource of postburn IL-10. Shock36:149–155
10. Cripps JG, Gorham JD (2011) MDSC inautoimmunity. Int Immunopharmacol11:789–793
11. Goni O, Alcaide P, Fresno M (2002)Immunosuppression during acuteTrypanosoma cruzi infection:involvement of Ly6G(Gr1(?))CD11b(?)immature myeloidsuppressor cells. I. Int Immunol14:1125–1134
12. Voisin MB, Buzoni-Gatel D, Bout D,Velge-Roussel F (2004) Both expansionof regulatory GR1 ? CD11b ?myeloid cells and anergy of Tlymphocytes participate inhyporesponsiveness of the lung-associated immune system during acutetoxoplasmosis. Infect Immun72:5487–5492
13. Cuervo H, Guerrero NA, Carbajosa S,Beschin A, De Baetselier P, Girones N,Fresno M (2011) Myeloid-derivedsuppressor cells infiltrate the heart inacute Trypanosoma cruzi infection.J Immunol 187:2656–2665
15. Chen S, Akbar SM, Abe M, Hiasa Y,Onji M (2011) Immunosuppressivefunctions of hepatic myeloid-derivedsuppressor cells of normal mice and in amurine model of chronic hepatitis Bvirus. Clin Exp Immunol 166:134–142
16. Tacke R, Lee HC, Goh C, Courtney J,Polyak SJ, Rosen HR, Hahn YS (2011)Myeloid suppressor cells induced byhepatitis C virus suppress T cellresponses through the production ofreactive oxygen species. Hepatology.doi:10.1002/hep.24700
18. Cuenca AG, Delano MJ, Kelly-Scumpia KM, Moreno C, Scumpia PO,Laface DM, Heyworth PG, Efron PA,Moldawer LL (2011) A paradoxicalrole for myeloid derived suppressorcells in sepsis and trauma. Mol Med17:281–292
19. Gibot S, Kolopp-Sarda MN, Bene MC,Bollaert PE, Lozniewski A, Mory F,Levy B, Faure GC (2004) A solubleform of the triggering receptorexpressed on myeloid cells-1 modulatesthe inflammatory response in murinesepsis. J Exp Med 200:1419–1426
20. Ford JW, McVicar DW (2009) TREMand TREM-like receptors ininflammation and disease. Curr OpinImmunol 21:38–46
21. Youn JI, Gabrilovich DI (2010) Thebiology of myeloid-derived suppressorcells, the blessing and the curse ofmorphological and functionalheterogeneity. Eur J Immunol40:2969–2975
22. Bunt SK, Yang L, Sinha P, ClementsVK, Leips J, Ostrand-Rosenberg S(2007) Reduced inflammation in thetumor microenvironment delays theaccumulation of myeloid-derivedsuppressor cells and limits tumorprogression. Cancer Res67:10019–10026
23. Sander LE, Sackett SD, Dierssen U,Beraza N, Linke RP, Muller M, BlanderJM, Tacke F, Trautwein C (2010)Hepatic acute-phase proteins controlinnate immune responses duringinfection by promoting myeloid-derivedsuppressor cell function. J Exp Med207:1453–1464
24. Chalmin F, Ladoire S, Mignot G,Vincent J, Bruchard M, Remy-MartinJP, Boireau W, Rouleau A, Simon B,Lanneau D, De Thonel A, Multhoff G,Hamman A, Martin F, Chauffert B,Solary E, Zitvogel L, Garrido C, RyffelB, Borg C, Apetoh L, Rebe C,Ghiringhelli F (2010) Membrane-associated Hsp72 from tumor-derivedexosomes mediates STAT3-dependentimmunosuppressive function of mouseand human myeloid-derived suppressorcells. J Clin Invest 120:457–471
25. Yoshimura A, Ohishi HM, Aki D,Hanada T (2004) Regulation of TLRsignaling and inflammation by SOCSfamily proteins. J Leuk Biol75:422–427
26. Wang H, Brown J, Martin M (2011)Glycogen synthase kinase 3: a point ofconvergence for the host inflammatoryresponse. Cytokine 53:130–140
27. Ostrand-Rosenberg S, Sinha P (2009)Myeloid-derived suppressor cells,linking inflammation and cancer.J Immunol 182:4499–4506
29. Adib-Conquy M, Cavaillon JM (2009)Compensatory anti-inflammatoryresponse syndrome. Thromb Haemost101:36–47
30. Noel G, Wang Q, Osterburg A,Schwemberger S, James L, Haar L,Giacalone N, Thomas I, Ogle C (2010)A ribonucleotide reductase inhibitorreverses burn-induced inflammatorydefects. Shock 34:535–544
31. Delano MJ, Thayer T, Gabrilovich S,Kelly-Scumpia KM, Winfield RD,Scumpia PO, Cuenca AG, Warner E,Wallet SM, Wallet MA, O’Malley KA,Ramphal R, Clare-Salzer M, Efron PA,Mathews CE, Moldawer LL (2011)Sepsis induces early alterations ininnate immunity that impact mortalityto secondary infection. J Immunol186:195–202



















![[Ghiduri][Cancer]Acute Myeloid Leukemia](https://static.documents.pub/doc/80x56/55cf9686550346d0338c0f55/ghiduricanceracute-myeloid-leukemia.jpg)





