ISSN 0564-3295 nl\{}n" Tropical Ecology 41(1): 47-60, 2000 @ International Society for Tropical Ecology Variations in leaf characteristics of two species of Miconia in the Brazilian cerrado under different light intensities ANDRÉA RODRIGUES MARQUES*, QUEILA SOUZA GARCIA*, JULIANA L. PASSOS REZENDE:I: & G WILSON FERNANDES** *Departamento de Botanica, **Ecologia Evolutiva de Herbívoros Tropicais/ DBG, Caixa Postal 486. ICB. Universidade Federal de Minas Gerais 30161-970, Belo Horizonte, Ma, Brazil Abstract: The relationships between severalleaf attributes (leaf area, mass, volume, den- sity, thickness. organic matter content. anatomy and leaf specific mass) in two sympatric shrub species.Miconia ibaguensis and M. stenostachya (Melastomataceae), in Brazilian celTado were explored. Leaf traits were assessed in two environments : a sun-exposed cerrado and in a shaded undestorey riparian forest. Area. mass, density and thickness ofthe leaves ofboth spe- cies varied significantly between the two habitats (t-tests. p < 0.05 alI). Variations in leafspe- cific mass were correlated with leaf mass. density. and thickness in both species (Pearson cor- relation, p < 0.001). Fully sun-exposed leaves were denser and thicker due to an increase in the length of the palisade parenchyma cells (M. ibaguensis: sun = 93 :t: 3 Jlm; shade = 61 :t: 2 Jlm; M. stenostachya: sun = 150 :t: 5 Jlm; shade = 92 :t: 4 Jlm). Miconia ibaguensis and M. stenostachya. presented an increase of 13% and 17% in stomatal density in the sumly habitat, respectively. Miconia ibaguensis also showed an increase of 57% in trichome density under high light conditions. These two species showed high leaf structure plasticity in relation to en- vironmental conditions. Resumen: Se estudiaron làs relaciones de varios atributos de Ias hojas (área, peso, volu- men, densidad, espesor, maten\ orgánica, anatomía y peso específicofoliar) de dos especiesar- bustivas simpátricas, Miconia ibaguensis y M. stenostachya (Melastomataceae), expuestas al sol y en Ia sombra soto de en el bosque deI cerrado brasileno. EI área, peso, densidad yespesor de Ias hojas de ambas especies variaron significantivamente entre 1osdos habitats (best t, p < 0.05). Las variaciones en e} peso específico foliar estuvieron correlacionadas con Ia masa, den- sidad y espesor de Ias hojas en ambas especies (Coeficiente de correlación de Pearson, p < 0.001). Las hojas expuestas completamente al sol fueron más densas y con mayor espesor de- bido al aumento de Ia extensión de Ias células deI parénquima en empalizada (M. ibltguensis: sol = 93:!: 3 pm; sombra = 61 :t 2 )lm;M. stenostach.ya: sol = 150:!: 5 pm; sombra = 92 :t 4 pm). M. ibaguensis y M. stenostachya presentaron un aumento en Ia densidad de estomas de 13(* y 17%, respectivamente; en el habitat con sol, M. ibaguensis presentó un aumento de 57% en Ia densidad de tricomas. Estas dos especies mostraron una alta plasticidad foIiar en relación a Ias condiciones ambientales. Resumo: Foram estudadas as relações entre os vários atributos foliares (área, massa, vol- ume, densidade, espessura, materia orgânica, anatomia e massa foliar específica) de duas espé- cies arbustivas simpátricas, Miconia ibaguensis e M, stenostach,Ya (Melastomataceae) no cer- rado brasileiro. As caracteristi~as foliares foram avaliadas em dois ambientes: cerrado exposto ao sol e à sombra do subbosque da mata ripária. A área, massa, densidade e espessura foliares das duas espécies variaram significativamente entre os dois habitats (teste t, p < 0.05), As variações na massa foliar (MFE) estavam correlacionadas com a massa, densidade e espessura Correspondence : Andréa R. Marques, Fax : 55-31-499-2673; E-mail: [email protected]
Transcript
ISSN 0564-3295
nl\{}n"Tropical Ecology 41(1): 47-60, 2000@ International Society for Tropical Ecology
Variations in leaf characteristics of two species of Miconia in the
Brazilian cerrado under different light intensities
ANDRÉA RODRIGUES MARQUES*, QUEILA SOUZA GARCIA*, JULIANA L. PASSOS REZENDE:I:& G WILSON FERNANDES**
*Departamento de Botanica, **Ecologia Evolutiva de Herbívoros Tropicais/ DBG, Caixa Postal486. ICB. Universidade Federal de Minas Gerais 30161-970, Belo Horizonte, Ma, Brazil
Abstract: The relationships between severalleaf attributes (leaf area, mass, volume, den-sity, thickness. organic matter content. anatomy and leaf specific mass) in two sympatric shrubspecies. Miconia ibaguensis and M. stenostachya (Melastomataceae), in Brazilian celTado wereexplored. Leaf traits were assessed in two environments : a sun-exposed cerrado and in ashaded undestorey riparian forest. Area. mass, density and thickness ofthe leaves ofboth spe-cies varied significantly between the two habitats (t-tests. p < 0.05 alI). Variations in leafspe-cific mass were correlated with leaf mass. density. and thickness in both species (Pearson cor-relation, p < 0.001). Fully sun-exposed leaves were denser and thicker due to an increase inthe length of the palisade parenchyma cells (M. ibaguensis: sun = 93 :t: 3 Jlm; shade = 61 :t: 2Jlm; M. stenostachya: sun = 150 :t: 5 Jlm; shade = 92 :t: 4 Jlm). Miconia ibaguensis and M.stenostachya. presented an increase of 13% and 17% in stomatal density in the sumly habitat,respectively. Miconia ibaguensis also showed an increase of 57% in trichome density underhigh light conditions. These two species showed high leaf structure plasticity in relation to en-vironmental conditions.
Resumen: Se estudiaron làs relaciones de varios atributos de Ias hojas (área, peso, volu-men, densidad, espesor, maten\ orgánica, anatomía y peso específico foliar) de dos especies ar-bustivas simpátricas, Miconia ibaguensis y M. stenostachya (Melastomataceae), expuestas alsol y en Ia sombra soto de en el bosque deI cerrado brasileno. EI área, peso, densidad yespesorde Ias hojas de ambas especies variaron significantivamente entre 1os dos habitats (best t, p <0.05). Las variaciones en e} peso específico foliar estuvieron correlacionadas con Ia masa, den-sidad y espesor de Ias hojas en ambas especies (Coeficiente de correlación de Pearson, p <0.001). Las hojas expuestas completamente al sol fueron más densas y con mayor espesor de-bido al aumento de Ia extensión de Ias células deI parénquima en empalizada (M. ibltguensis:sol = 93:!: 3 pm; sombra = 61 :t 2 )lm;M. stenostach.ya: sol = 150:!: 5 pm; sombra = 92 :t 4 pm).M. ibaguensis y M. stenostachya presentaron un aumento en Ia densidad de estomas de 13(* y17%, respectivamente; en el habitat con sol, M. ibaguensis presentó un aumento de 57% en Iadensidad de tricomas. Estas dos especies mostraron una alta plasticidad foIiar en relación aIas condiciones ambientales.
Resumo: Foram estudadas as relações entre os vários atributos foliares (área, massa, vol-ume, densidade, espessura, materia orgânica, anatomia e massa foliar específica) de duas espé-cies arbustivas simpátricas, Miconia ibaguensis e M, stenostach,Ya (Melastomataceae) no cer-rado brasileiro. As caracteristi~as foliares foram avaliadas em dois ambientes: cerrado expostoao sol e à sombra do subbosque da mata ripária. A área, massa, densidade e espessura foliaresdas duas espécies variaram significativamente entre os dois habitats (teste t, p < 0.05), Asvariações na massa foliar (MFE) estavam correlacionadas com a massa, densidade e espessura
foliares em ambas as espécies (Correlação de Pearson, p < 0.001). As folhas completamente ex-postas ao sol apresentaram-se mais densas e espessas devido ao aumento do comprimento dascélulas do parênquima paliçádico (M. ibaguensis: sol = 93 :i: 3 mm; sombra = 61 :i: 2 mm;.M.stenostachya : sol = 150 :i: 5 mm; sombra = 92 :i: 4 mm). As folhas de M. : ibaguensis e M.stenostachya apresentaram um aumento na densidade estomática de 13% e 17%, respecti-vamente, no habit'at ensolarado. A M. ibaguensis apresentou um aumento de 57% na densi-dade dos tricomas sob essa condições. Essas duas espécies mostraram uma alta plasticidade
Introduction temperature (Ku & Hunt 1973), altitude (Korner1989), and light intensity (Nobel. et al. 1975; Sims& Pearcy 1992; Upadhyaya & Furness 1994).These variations may involve changes in life form(Garnier & Laurent 1994), .leaf pubescence (Ehler-inger & Mooney 1978), leaf area (Jurik et al. 1979),leaf thickening (Turner 1994), and photosyntheticrate (Holmes & Cowling 1993; Romero-Aranda etal. 1997).
Tl)is study concerns two common sympatriccerrado shrubs species, Miconia ibaguensis(Bonpl.) Triana and Miconia stenostachya Schrank& Mart. ex DC. (Melastomataceae), that have beenfound growing together in sunny and shady habi-tats. Miconia ibaguensis is primarily from ripar-ian forest while M. stenostachya is from campo cer-rado (Martins et al. 1996). These species vary inarchitecture, which suggests differences in leafstructure. Few studies have examined naturalvariation in leaf anatomy in relation to light ac-climation. The aim of the present work was to de-termine the variations of leaf characteristics ofthese two shrubs in two different light intensityhabitats. These studies may elucidate aspects ofinter-and intra-specific adaptation to heterogene-ous light environments in Brazilian cerrado.
Methods
Cerrado is a type ofvegetation which predomi-nates on the Central Brazilian Plateau, similar tosavanna in physiognomy. It ranges from pur.egrassland (campo limpo) to nearly closed canopy ofmedium height trees overlying grass (cerrado,campo cerrado and campo sujo) (Lopes & Cox1977). There are physiognomic gradients based onthe amount and size of woody plant~. The woodycomponents of the Brazilian cerrad~ -are typicallysclerophyllous, resistant to annual fires, and seemto have access to the water table, therefore, re-maining photosynthetically active throughout theyear (Medina & Silva 1990). The differences inphysiognomic gradients of cerrado have been hy-pothesiz.ed as an evolutionary consequence of se-lection pressure imposed by the oligotrophic soils(Lopes & Cox 1977).
A common feature of the cerrado is the pres-ence of narrow riparian forests along rivers,washes, and creeks (Meguro et al. 1996). In theseriparian forests, light intensity is lower on shruband herb strata, and water and nutrient availabil-ity are higher (Eiten 1990). Some species arefound in cerrado and in riparian forest, but theyshow structural modifications. These features arethe basis for the study of leaf plasticity under dif-ferent environmental conditions in habitats sepa-rated only by a few meters.
Leaves of many plant species are known fortheir great growth plasticity varying in morphol-ogy and anatomy, as well as in physiology. For aparticular plant species phenotype may change inresponse to nutrient and/or moisture availability(Jurik et al. 1982; Witkowski & Lamont 1991),
Study site
The study was conducted in the Serra doCipó N ational Park, Minas Gerais, southeasternBrazil at the southern end of the EspinhaçoMountains (19°20' S, 43°44' W) at 800 m altitudeabove sea level (Fig. 1). The climate is tropical,with moist and warm summers, and with a de-
MARQUES ET AL 49
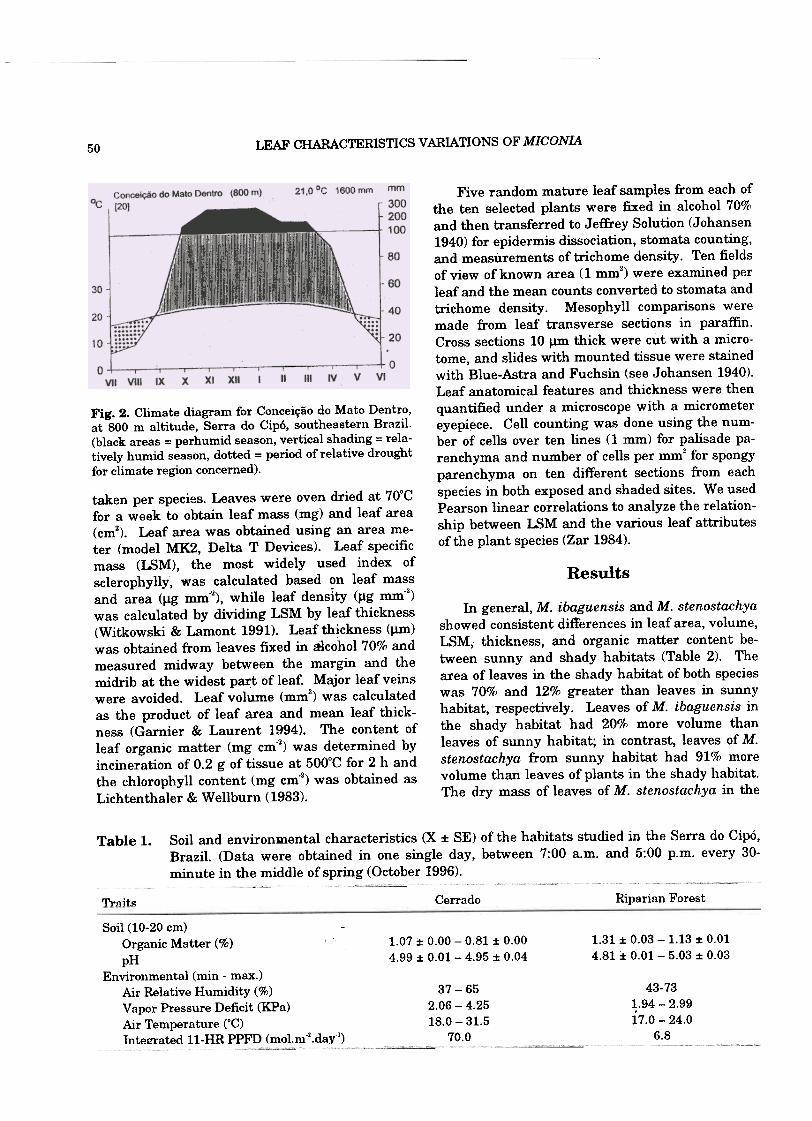
fined dry season (Fig. 2).averages 1600 mm.
The annual rainfall cumbent stem; both are components ofthe riparianforest understorey.
The habitats were 8lso characterized in relation tothe relative humidity (%), vapor pressure deficit(Kpa),and soil fertility (Table 1). AlI environmental data pre-sented were obtained, in one single day, between 7:00a.m. and 5:00 p.m. every 30-minute in the middle ofspring (October 1996). Relative humidity (%) and va.por pressure deficit (Kpa) of the air were obtainedwith a psychrometer (lCOTERM). The amount of soilorganic matter (SOM) was determined after the incin-eration of three soil samples at 500.C for two hours,the percentage of SOM being the difference betweenthe weights. The pH was determined by the methodofRaij & Quaggio (1983). We used the Student t-testto statistic8lly compare 8l1 the leaf traits under thetwo distinct environments (Zar 1984)..
Site characteristics
Habitats studied were selected by their dis-tinctiveness in light intensity .They were catego-rized by photosynthetic photon flux density(PPFD), measured with quantum sensors LI-189,as: (a) sunny habitat (integrated PPFD of 70 molm-2 day-l) and (b) shady habitat (integrated PPFDof 6.8 mol m-2 day-l). The sunny habitat is locatedin "campo cerradô" that consists of a continuousgrass cover with open or sparse shrubs or lowtrees, which cover less than a third of the surface.Miconiq ibaguensis and M. stenostachya are ap-proximately 1 m high in this habitat distributed indistinct groups. The shady habitat is located in ariparian forest, composed of medium tal1 (5 -7 m)arboreal forms with either closed or semi-open treecanopy. In this habitat M. ibaguensis is nearly 2.5m high, while M. stenostachya may have a pro-
'~~
c;.;'1..-~
Leaf attrib utes
In each site, five random mature leaf samples(5th node under shoot apex) of teri plants were
t\ ..
t...'-.., '
;.
~i 1
:\~;::,
~] Ce,rado[] 5tudy oreo
..,.
O 1000km..
1'-
ÇL2'
~
2:[o 6km~ -
..~3°30' W
Fig. I. National Park of Serra do Cipó, Minas Gerais, southeastern Brazil, where the studv site was located
',,' ..:.~:I
LEAF CHARACTERISTICS V ARIATIONS OF MICONIA50
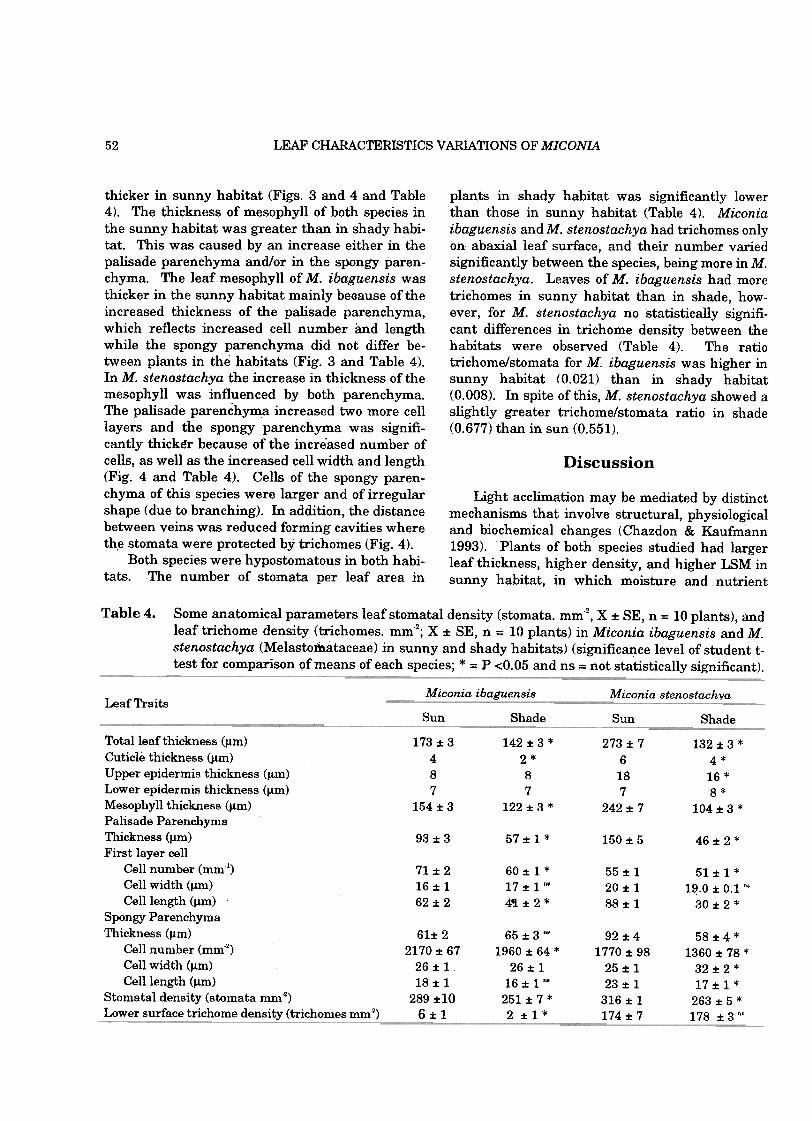
Five random mature leaf samples from each ofthe ten selected plants were fixed in alcohol 70%and then transferred to Jeffrey Solution (Johansen1940) for epidermis dissociation, stomata counting,and measuretnents of trichome density .Ten fieldsofview ofknown area (1 mm2) were examined perleaf and the mean counts converted to stomata andtrichome density. Mesophyn comparisons weremade from leaf transverse sections in paraffin.Cross sections 10 I.1m thick were cut with a micro-tome, and slides with mounted tissue were stainedwith Blue-Astra and Fuchsin (seeJohansen 1940).Leaf anatomical features and thickness were thenquantified under a microscope with a micrometereyepiece. Cen counting was done using the num-ber of cens over ten lines (1 mm) for palisade pa-renchyma and number of cens per mm2 for spongyparenchyma on ten different sections from eachspecies in both exposed and shaded sites. We usedPearson linear correlations to analyze the relation-ship between LSM and the various leaf attributesofthe plant species (Zar 1984).
Fig. 2. Climate diagram for Conceição do Mato Dentro,at 800 m altitude, Serra do Cipó, southeastern Brazil.(black areas = perhumid season, vertical shading = rela-tively humid season, dotted = period ofrelative drought
for climate region concerned).
taken per SpeCi5S. Leaves were oven dried at 70°Cfor a week to obtain leaf mass (mg) and leaf area(cm2). Leaf area was obtained using an area me-ter (model MK2, Delta T Devices). Leaf specificmass (LSM), the most widely used index ofsclerophylly, was calculated based on leaf massand area (~g mm.2), while leaf density (~g mm.3)was calculated by dividing LSM by leaf thickness(Witkowski & Lamont 1991). Leaf thickness (~m)was obtained from leaves fixed in álcohol 70% andmeasured midway between the margin and themidrib at the widest p~t of leaf. Major leaf veinswere avoided. Leaf volume (mm3) was calculatedas the product of leaf area and mean leaf thick-ness (Garnier & Laurent 1994). The content ofleaf organic matter (mg cm-2) was determined byincineration of 0.2 9 of tissue at 500°C for 2 h andthe chlorophy11 content (mg cm.2) was obtained asLichtenthaler & We11burn (1983).
Results
In general, M. ibaguensis and M. stenostachyashowed consistent differences in leaf area, volume,LSM; thickness, and organic matter content be-tween sunny and shady habitats (Table 2). Thearea of leaves in the shady habitat of both specieswas 70% and 12% greater than leaves in sunnyhabitat, respectively. Leaves of M. ibaguensis inthe shady habitat had 20% more volume thanleaves of sunny habitat; in contrast, leaves of M.stenostachya from sunny habitat had 91% morevolume than leaves of plants in the shady habitat.The dry mass of leaves of M. stenostachya in the
Soil and environmental characteristics (X :t SE) of the habitats studied in the Serra do Cipó,Brazil. (Data were obtained in one single day, between 7:00 a.m. and 5:00 p.m. every 30-
Air Relative Humidity (%)Vapor Pressure Deficit (KPa)Air Temperature ("C)Integrated 11-HR PPFD (mol.m2.dayl)
51MARQUESETAL
_..
The differences in LSM were inf1uenced bychanges in the thickness of cuticle, epidermis, pali-sade, and spongy parenchyma. All anatomicalparameters measured differed between sunny andshady habitats in both species. Structural devel-opment of leaf mesophyll in the sunny habitat wasgreater compared to shady habitat. The differ-ences in the development of the mesophyll wereprobably caused by an increase in the tissue thick-ness, in the number of cells per area, and in thecell size (Table 4).
Leaves of M. ibaguensis in the sun were 200/f,thicker than leaves of plants in the shady habitat,while leaves of M. stenostachya in the sunny habi-tat were 100% thicker than those in the shadyhabitat (Table 4). Although the epidermis did notdiffer in thickness for M. ibaguensis, the thicknessof leaf cuticle in the sunny habitat contributed toan increase in the leaf thickness in this species,and for M. stenostachya cuticle and epidermis were
sunny habitat was 92% greater than leaves in theshady habitat while M. ibaguensis showed no sta-tistical differences between the habitats. Plants insunny habitat had leaves with higher LSM anddensity compared to leaves of shady habitat(Table 2). The same pattern was observed in or-ganic matter content which was 77% greater in M.ibaguensis and 114% greater in M. stenostachya inthe sunny habitat than in the shady habitat.Likewise, leaves of both species in the sunny habi-tat were thicker than those in the shady habitat(Table 2). The chlorophyll content for both specieswas higher in shady than in sunny habitat(Table 2).
LSM did not correlate with leaf area, but corre-lated significantly with volume in leaves in theshady habitat of both species (Table 3). LikewiseLSM correlated significantly with leaf thicknessonly in the shady habitat for both species (M. iba-guensis r= 0.66; M. stenostachya r= 0.54).
Table 2. Leaf traits (X :t: SE) ofMiconia ibaguensis and M. stenostach.ya (Melastomataceae) in sunnyand shady habitats. (Test t, * = P <0.05 and NS = not statistical1y significant, LSM denotes
Linear (Pearson product moment) correlations between leaf specific mass (LSM) and variousleaf traits of Miconia ibaguensis and M, stenostachya (Melastomataceae) in sunny and shadyhabitats. (* = P <0.05 and ns = not statistically significant), different letters indicate signifi-cant differences between habitats (Test t. P <0,001),
thicker in sunny habitat (Figs. 3 and 4 and Table4). The thickness of mesophyll of both species inthe sunny habitat was greater than in shady habi-tat. This was caused by an increase either in thepalisade parenchyma and/or in the spongy paren-chyma. The leaf mesophyll of M. ibaguensis wasthicker in the sunny habitat mainly beoause of theincreased thickness of the palisade parenchyma,which reflects increased cell number and lengthwhile the spongy parenchyma did not differ be-tween plants in the habitats (Fig. 3 and Table 4).In M. stenostachya the increase in thickness of themesophyll was influenced by both parenchyma.The palisade parenchyma increased two more celllayers and the spongy parenchyma was signifi-cantly thickér because of the incre.ased number ofcells, as well as the increased cell width and length(Fig. 4 and Table 4). Cells of the spongy paren-chyma of this species were larger and of irregularshape (due to branching). In addition, the distancebetween veins was reduced forming cavities whereth.e stomata were protected by trichomes (Fig. 4).
Both species were hypostomatous in both habi-tats. The number of stomata per leaf area in
plants in shady habitat was significantly lowerthan those in sunny habitat (Table 4). Miconiaibaguensis and M. stenostachya had trichomes onlyon abaxial leaf surface, and their number variedsignificantly between the species, being more in M.stenostachya. Leaves of M. ibaguensis had moretrichomes in sunny habitat than in shade, how-ever, for M. stenostachya no statistically signifi-cant differences in trichome density between thehabitats were observed (Table 4). The ratiotrichome/stomata for M. ibaguensis was higher insunny habitat (0.021) than in shady habitat(0.008). In spite of this, M. stenostachya showed aslightly greater trichome/stomata ratio in shade(0.677) than in sun (0.551).
Discussion
Light acclimation may be mediated by distinctmechanisms that involve structural, physiologicaland biochemical changes ( Chazdon & Kaufmann1993). Plants of both species studied had largerleaf thickness, higher density, and higher LSM insunny habitat, in which moisture and nutrient
Table 4. Some anatomical parameters leaf stomatal density (stomata. mm-2, X f: SE, n = 10 plants), andleaftrichome density (trichomes. mm-2; Xf: SE, n = 10 plants) in Miconia ibaguensis and M.
stenostachya (Melasto1b.ataceae) in sunny and shady habitats) (significance leveI of student .t-test for comparison ofmeans ofeach species; * = P <0.05 and ns = not statistically significant).
Celllength (Jlm)Stomatal density (stomata mm2)Lower surface trichome density (trichomes mm2)
MARQUESETAL 53
1
I
5
BA
Fig. 3. A cross section of Miconia ibaguensis leaf in the sunny habitat (A) and in the shady habitat (B). 1, cuticle; 2,epiderlllis; 3, palisade parenchyma; 4, spongy parenchyma; 5, stolllata. Bar = 100 J1m. (x 250).
B
A
Fig. 4. A cross section of Miconia stenostachya leaf in the sunny habitat (A) anrl in ~he shady habitat (B). 1, cuticle; 2,epidermis; 3, palisade parenchyma; 4, spongy parenchyma; 5, stomata; 6, trichomes. Bar = 100 J1ln. (x 250).
However, for plants from understorey LSM waspositively correlated with leaf thickness. Con-trasts between the correlations of LSM and thick-ness have been reported in several studies (e.g.,Dijkstra & Lambers 1989; Garnier & Laurent1994; Kebede et al. 1994; van Arendonk & Poorter1994). On the other hand, LSM was positively cor-related with leaf mass and leaf density in bothspecies and habitats. Our data corroborate thefindings of Pammenter et al. (1986), Kebede et al.
availability were lower, in agreement with severalexperimental studies (e.g. , Boardman 1977; Sims& Pearcy 1992; Witkowski & Lamont 1991).Plants in the .understorey of the riparian foresthad thinner leaves resulting in greater area butlower density and LSM as reported by Corré(1983).
Our data sowed that there was no signiticantrelationship between LSM and leaf thickness ineither species studied in sun-exposed cerrado.
LEAF CHARACTERISTICS V ARIATIONS OF MICONIA54
Trichome and stomata density can also be in-fluenced by light. For M. ibaguensis, the ratiotrichome/stomata was highest in the sunny habi-tat. This suggests that the increase in tric.homedensity found in sunny habitat plays an importantrole in water regulation through an increase in theboundary layer and reduces the vapor pressuredifference between leaf surface and the microcli-mate just above thc leaf lamina, minimizing waterlosses (e.g., Ehleringer & Mooney 1978; Gay &Hurd ]975; Johnson 1975). Although the numberof trichomes of M. stenostachya was not statisti-cally different between the habitats, the stomata ofthis species in sunny habitat are in deep cavities,feature that may reduce water loss (Fahn & 'Cl.lt.-tler 1992). The greater number of stomata of M.stenostachya in sunny habitat could be related tothe CO2 diffusion rate because this species also hasthicker leaves. Romero-Aranda et al. ( 1997)showed that the increase in leaf thickness andlarge mesophyll cell volume increase the biophysi-cal resistancc to CO2 diffusion. Besides this,thicker leaves that increase the number of st.omataper unit area may favor leaf CO2 diffusion underhigh irradiance conditions (e.g., Mott et al. 1982;Mott & Michaelson 1991).
For M. ibaguensis, leaf anatomical variationswere less evident between habitats. Probably, M.ibaguensis invests in biochemical and physiologicalmechanisms that allow its survival in sunny habi-tat because the species primarily occurs in shadyhabitat (Martins et al. 1996). Miconiastenostachya, which is primarily a sun plant (Mar-tins et al. 1996), seems to invest more in leaf ana-tomical st.ructures in order to gain more protectionagainst. high irradiations. When it grows in ashady habitat it exhibits more conspicuous mor-pho]ogical changes in the leaves.
We can conclude that sympatric species mayshow variations in leaf anatomy, density, thick-ness and LSM that may be important ecologicaladaptations relative to light conditions, and thatthese features are very plastic structurally.
(1994) and Garnier & Laurent (1994) in which leafdensity played a major role in the determination ofLSM in the sunny habitat. The effects ofthicknessand/or leaf density on LSM have also been shownin the study of Witkowski & Lamont (1991), withshrub species growing in different environments.
Variation in leaf density can be brought aboutby several factors which pertain to chemical com-position and anatomy (Lambers & Poorter 1992).Anatomical characters leading to a high leaf den-sity are essentially those th.at result in a highamount of cell wall per unit volume. This occurswhen the proportion of tissues with thick walledcells is high relative to tissues with thin walledcells and/or the cells of one or several tissues aresmall (Garnier & Laurent 1994). The species stud-ied here showed increased mesophyll cell size andnumber which was reflected in mesophyll thick-ness and density, therefore, contributing to eitheran increase in the amount of the cellular walls orin the percentage of organic matter tsee alsoTurner et al. 1993).
Leaf thickness is influenced by variations inthe palisade tissue (number of layers and celllength), number oflayers and length ofmesophyll,epidermis, hypodermis, and indumentum, as wellas by increased cell size (Esau 1977; Fahn & Cut-tler 1992; Garnier & Laurent 1994; Sims & Pearcy1992; Thompson et al. 1992; Witkowski & Lamont1991). Our results showed an increase in thick-ness of cuticle and .epidermis of leaves in the sun-exposed cerrado in M. .stenostachya whereas, M.ibaguensis increased only in cuticle thickness.Chazdon & Kaufma:nn (1993) suggest that the in-crease in thickness of the upper epidermis mayincrease leaf reflectance. A thicker epidermallayer alters light-scattering processes withinmesophyll tissue or protect photosynthetic tissuesfrom excessive irradiance (Roth 1934). 1'hus, ourdata indicate that M. stenostachya may be moreprotected against light irradiation than M. iba-guensis.
The palisade parenchyma cells of M. ibaguen-sis and M. stenostachya were deeper in sun-exposed plants and the latter also showed morelayers. Thompson et al. (1992) stated that highlight irradiance induces the development of themesophyll in the palisade and spongy parenchymaproducing thicker leaves which have thinner aerialspaces. Conversely, leaves of shade plants havethinner blades and smaller and more denselyDacked leafcells (Nobel et aL 1975)-
Acknowledgements
We are very grateful to H. Ziegler, U. Liiettge,F. Scarano, E~nesto Medina and Augusto C.Franco for their critical reviews on this paper. .R.Goldemberg kindly identified the plant speciesstudied. Logistical support by the Parque Na-
1;1;MARQUES ET AL
cional da Serra do Cipó, and US Fish and WildlifeService for partially sponsoring a field course incerrado ecology (ECVMS/UFMG). This project waspartially supported by the CNPq (521772/95-8),and FAPEMIG (1950/95).
References
~
Boardman, N .K. 1977. Comparative photosynthesis ofsun and shade plants. Annual Review of Plant
Physiology 28: 335-377.Chazdon, R.L. & S. Kaufmann. 1993. Plasticity of leaf
anatomy oftwo rain forest shrubs in relation to pho-tosynthetic light acclimation. Functional Ecology 7:
385-394.Corré, W.J. 1983. Growth and morphogenesis of sun
and shade plants. I. The influence oflight intensity.Acta Botanical Neerling 32: 49-62.
Dijkstra, P. & H. Lamber.1989. Analysis ofspecificleafarea and photosynthesis oftwo inbred lines of Plan-tago major differing in relative growth rate. NewPhytologist 113: 283-290.
Ehlerillger, J.R. & H.A Mooney. 1978. Leafhairs: ef-fects on physiological activity and adaptative valuesto a desert shrub. Oecologia 37: 183-200.
Eiten, G. 1990. A vegetação do cerrado. pp. 9-65. In:M.N. Pinto (org.) Cerrado: Caracterização, ocupaçãoe perspectivas. Ed. UNB, Brasília.'
Esau, K. 1977. Anatomy ofSeed Plants. 2nd edn. Wiley,New York.
Fahn, A. & D.F. Cuttler. 1992. Xerophytes. Berlin : Ge-br11der Bomtraeger .
Garnier, E. & G. Laurent. 1994. Leafanatomy, specificmass and water content in cogeneric annual andperennial grass species. New Phytologist 128: 725-
736.Gay, AP. & G. Hurd. 1975. The influellce of light Oll
stomata1 density in tomato. New Phytologist 74: 37-46.Holmes, P.M. & R.M. Cowling. 1993. Effects of shade on
seedling growth, morphology and leaf photosynthe-sis in six subtropical thicket species from the east-ern Cape, South Africa. Forest Ecology all.d Man-agement 61: 199-220.
Johansen, D.A. 1940. Plant Microtechnique. Mc-Graw-Hil1, New York.
Johnson, H.B. 1975. Plant pubescence: an ecologicalperspective. The Botanical Review 41: 233-258.
Jurik, T.W., J.F. Chabot & B.F. Chabot. 1982. Effect oflight and nutrients on leaf size, CO2 exchange, alldanatomy in wild strawberry (Fragaria virginiana).Plant Ph'\1sioloi!'\1 70: 1044-1048.
Kebede, H., B. Martin, J. Nienhuis & G. King. 1994.Leaf anatomy of two Lycopersicon species with con-trasting gas exchange properties. Crop Scielwe 34:
i08-113.Korner, C.H. 1989. The nutritional status ofplants from
high altitudes -A worldwide comparison. Oecologia81: 379-391.
Ku, S.B. & L.A. Hunt. 1973. Effects of temperature onthe morphology and photosynthetic activity ofnewlymatured leaves of alfalfa. Canadian Journal of Bot-any 51: 1907-1916.
Lambers, H. & H. Poorter. 1992. Inherent variation ingrowth rate between higher plants : a search forphysiological causes and ecological consequences.Aduances in Ecological Research 23: 187-261.
Lichtenthaler, H.K. & A.R. Wellbunl. 1983. Determina-tion of total carotenoids and chlorophylls a and b ofleaf extracts in different solvents. Biochemical So-ciety Transactions 11: 591-592.
Lopes, AS. & F .R. Cox 1977. Cerrado vegetation in Bra-zil: an edaphic gradient. Agronomy Joumal 69:
828-831.Martins, AB., J. Semir, R. Goldenberg & E. Matins.
1996. O gênero Miconia Ruiz & Pav. (Melastomata-ceae) no Estado de São Paulo. Acta Botanica Bra-silica 10: 267-316. .
Medina, E. & J. Silva. 1990. The savannas ofNorthernSouth America: a steady state regulated by water-fire interactions on a background of low nutrientavailability. Joúrnal Biogeografic 17: 403-413.
Meguro, M., J .R. Pirani, R. Mello-Silva & A.M. Giulietti.1996. Estabelecimento de matas ripárias e capõesnos ecossistemas campestres da Cadeia do Espin-haço, Minas Gerais. Boletim de Botânica da Uni-uersidade de São Paulo 15: 1-12.
Mott, K.A., A.C. Gibson & J.W. O'Leary. 1982. Theadaptive significance of anphistomatic leaves.Plant, Cell and Enuironment 5: 455-460.
Mott, K.A. & O. Michaelson. 1991. Amphistomy as anadaptation to high light intensity inAmbrosia cordi-folia (Compositate). American Joumal of Botany78: 76-79.
Nobel, P.S., L.J. Zaragoza & W.K. Smith.1975. Relationbetween mesophyll surface area, photoSY11theticrate, and illumination leveI during development forleaves of Plectranthus paruiflorus Henckel. PlantPhysiology 55: 1067-1070.
Pammenter, N.W., P.M. Dr~nnan & V.R. Smith. 1986.Physiological and anatomical aspects of photosyn-thesis of two Agrostis species at a sub-Antarctic is-land. New Phytologist 102: 143-160.
Raij, B.V. & J.A. Quaggio. 1983. Métodos de análise desolo para fins de fertilidáde. Boletim. Técnico -81.Instituto AS!:ronômico de CamDinas. São Paulo.
56 LEAF CHARACTERISTICS V ARIATIONS OF MICONIA
Romero-Aranda, R., B.R. Bondada, J.P. Syvertsen &J.W. Grosser. 1997. Leaf characteristics and netgas exchange of diploid and autotetraploid citrus.A1l.nals of Botany 79: 153-160.
Roth, J. 1984. Stratification of Tropical Forests as Seenin Leaf Structure. W. Junk, The Hague.
Sims, D.A. & R.W. Pearcy. 1992.. Response ofanatomyand photosynthetic capacity in Alocasia macror-rhiza (Araceae) to a transfer from low to high light.American .Toumal of Botany 79: 449-455.
Thompson, W.A., P.E. Kriedeman & I.E. Craig. 1992.Photosynthetic response to light and nutrients insun-tolerant and shade-tolerant rainforest trees. I.Growth, leaf anatomy and nutrient content. Austra.lian Journal of Plant Physiology 19: 1-18.
Turner, J.M. 1994. A quantitative analysis ofleafformin woody plants from the world's major broad-leavedforest types. Joumal of Biogeography 24: 413-419.
Turner, J.M., N.F. Choong, H.T.W. Tan & P.W. Lucas.1993. How tough are sclerophylls ? Anna~s of Bot-any 71: 343-345. ,
Upadhyaya, M.K. & N.H. Furness. 1994. Influence oflight intensity and water stress on leat surfacecharacteristics ofCynoglossum officinale, Centaureaspp., and Tragopogon spp. Canadian Joul71.al ofBot.any 72: 1379-1386.
van Arendonk, J.J.C.M. & H. Poorter. 1994. Th:e ~hemj-cal composition and anatomical structu~e of leavesof grass species differing in relative gJ;owth rate.Plant, Cell and Environment 17: 963-970.
Witkowski, E.T..F., B.B. Lamont. 1991. Leaf specificmass confounds leaf density and thickness.Oecologica 88: 486-493.