Page 1
Challenge of human synovialis with rheumatoidlymphocytes within diffusion chambers
Item Type text; Thesis-Reproduction (electronic)
Authors McNutt, Neil Scott, 1940-
Publisher The University of Arizona.
Rights Copyright © is held by the author. Digital access to this materialis made possible by the University Libraries, University of Arizona.Further transmission, reproduction or presentation (such aspublic display or performance) of protected items is prohibitedexcept with permission of the author.
Download date 06/06/2018 05:07:01
Link to Item http://hdl.handle.net/10150/551575
Page 2
Neil. Scott McNutt
A Thesis Submitted to the Faculty of the
DEPARTMENT OF AGRICULTURAL' BIOCHEMISTRY
In P artia l Fulfillment of the Requirements
For the Degree of
MASTER OF .SCIENCE
In the Graduate College
■ THE UNIVERSITY OF ARIZONA
1962
Page 3
STATEMENT BY AUTHOR
This thesis has been submitted in partial fulfillment of req u ire ments for an advanced degree at The University of Arizona and is deposited in The University Library to be made available to borrowers under ru les of the L ibrary.
Brief quotations from this thesis are allowable without special perm ission, provided that accurate acknowledgment of source is made. Requests for perm ission for extended quotation from or reproduction of this m anuscript in whole or in part may be granted by the head of the major department or the Dean of the Graduate College when in their judgment the proposed use of the m aterial is in the in terests of scho larship. In all other instances, however, perm ission must be obtained from the author.
SIGNED:
APPROVAL BY THESIS DIRECTOR
This thesis has been approved on the date shown below:
C.A.L. Stephens^/Tr., M.
ii
Page 4
\ : ACE^OWLEDGMENTS ' :
Grateful thanks are due Associate P rofessor Alice B„ Stanfield
and Charles A. L. Stephens, F.A.C.P. for the ir untiring
efforts in thought, encouragement, and editing of this worki Without
their aid this study would never have been carried out.
", >' Acknowledgment is especially given to Dr. A. R. Kem m erer '
and the United States Public Health Service for financial backing and
direction for this .investigation* The author is also deeply indebted to
the Southwestern Clinic and Research Institute, Inc. for financial and
technical assistance. A mention? here has been well earned by M rs .
Julia P. Bailey for her help in preparation of slides and photographs.
Of course,,. without the enthusiastic cooperation of the volunteer
donors of cells and tissues, any study of this nature is impossible.
Page 5
TABLE OF CONTENTS'
List of Tables'..........................
List of Figures.........................
'■' EiliAPTER' . ;;
x'T-F-; INTRODUCTION.....
n EXPERIMENTAL PROCEDURES
M : ■ RESULTS. ^ ............
1"F" DISCUSSION, o o ... o o . ...ooo.ooo. ... . .. . . o .
IZ" ■ iSii i ............... . 0 0 . . . . . . . . . . . . .
. ' LITERATURE C IT E D ...............
Page 6
v y v ; - ;:v ' i i s T0 F.mBLES - ■ : v - ; ' ■■'■ •Table : : y . - y - ' / r :v' ■ ;:>: V,. ; ; Page;
■ :: Composition of Earle’s Balanced Salt. Solution....... ' '24 ’, ’"
, 2 o ' Sample iE%peirinxental iPIan® <> < > » e ® o # » o ® o » e . ®» e ® ® # o o » ? > » # <> <> o o « » ® » o 25
. 3.;: Composition of Blended Bat D iet.. , » . , . . . . . . - > . . . . . . . . 2 5
4 . „ Sources of Synovial T i s s u e S o " « V p o . i , . . . . . . o . . o . ? . . . O . o . . . < , = , , 0 . .30
5 „ Sources of Blood Lymphocytes, „ » . . . » . 3 1
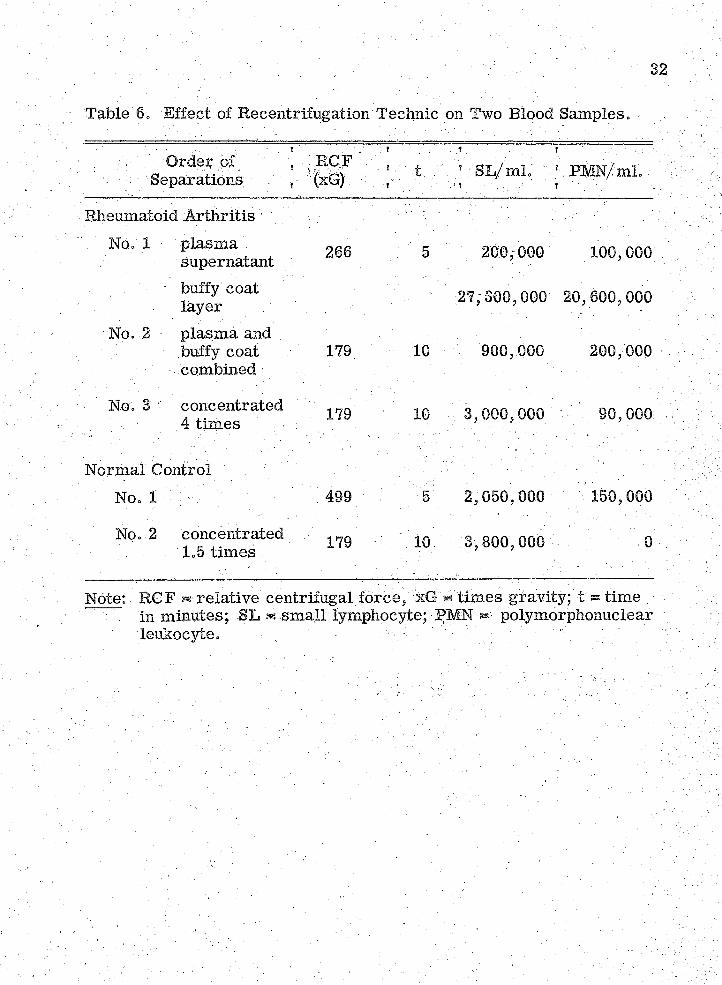
6„ Effect of Recentrifucation Technic on Two BloodSamples , p o , p , , m , * , o P , , o o o o p m p , , p o @ p o o o o .o o p p p p o » o o o o p p p p o p p e 'o e e o o a e o 32
7p Lymphocyte Inocula per Cham ber. . . a . . a a a a » a . a . o , . a a « » : . . o . 33
8 a Destruction of Lymphocytes Shown by F ilter,S45C9,nrXn3tlOn, a a a a a o a o a p a a a a p a a o a a a a a a a a a a a a a a a a a a a a a a a p a a a o a a a o a a a a a , 3 4 .
' ::9. . 'Destruction of Tissue Fragm ents Shown by Paraffin • y •5eCtions a a a a a a a a a a a a a a. a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a ■ ■ ‘3 5
1 0 a ■ Effect of Double Implantation on Chamber N ecrosis., 3 6 ■
I ! . . Challenge N ecrosis - -Lymphocytes and Tissue y.; '0/ Ond )S r e la a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a a 5 1
Page 7
LIST OF FIGURES
' Figure v . ' ./. Page
'''lV : ■ Rlieumatoid Syiiovialis cultured alone. . . . . . . . . . . . . . . . . . .. ... . 62./
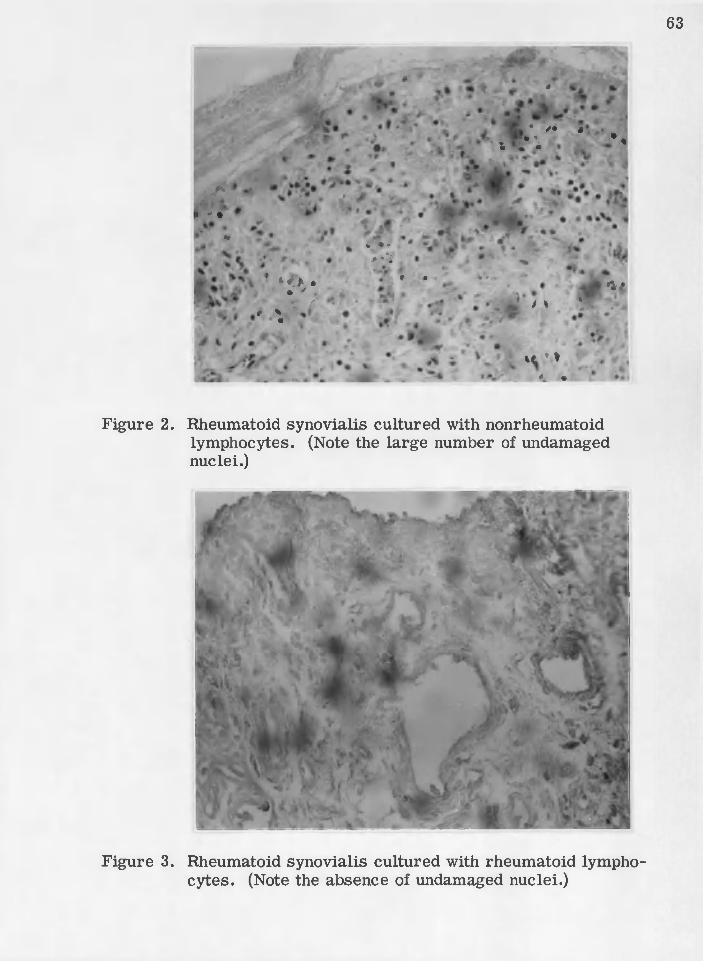
2 ; lElheumatoid synovialis cultured with nonrheumatpid' . ■ l ^ ^ m ^ ^ I l O C ^ ^ t O S ^ e o ''o O' e <) o o- o d e . o o- o e-'e o o .e o 0 o o -o e O'Oi o o o » - e e o e o o 0 - 0 o o e tt o o » o o o t> e o o o o o ' 6 6 .
■ ■ 3 o / ■'Rheumatoid synovialis cultured with .rheumatoid :. lymphocyte 8 0 * * -# # * * » » e * " * <>.0 • e - * < • • • «* o * ♦ ♦ o » 6 6
4 o Nohrheumatoid synovialis cultured a l o n e . . . . . . » 64
5. Nonrheumatoid synovialis cultured with nonrheuma-■ ■1 to id ' lymphocyte s .. o.... . . . . . . 0 o o o.. 0 , 0 o o... o... o. o. o. o o. * o. o o..... 6 5
6 0 Nonrheumatoid synovialis cultured with rheumatoid' lymphocytes, o o o o o o o o o o o o o o o o o o . o o o o o o . o o o . o o o o o o o o o o o o o o . 0 0 0 0 0 0 0 0 0 0 . 6 5
7. Rheumatoid lymphocytes cultured alone.. 0 . 0 .p....o....... 6 6
8 . Rheumatoid lymphocytes cultured near rheumatoids y n o v i a l i a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ............................ 67
9. Rheumatoid lymphocytes cultured near nonrheuma-- toid synovialis.... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 67
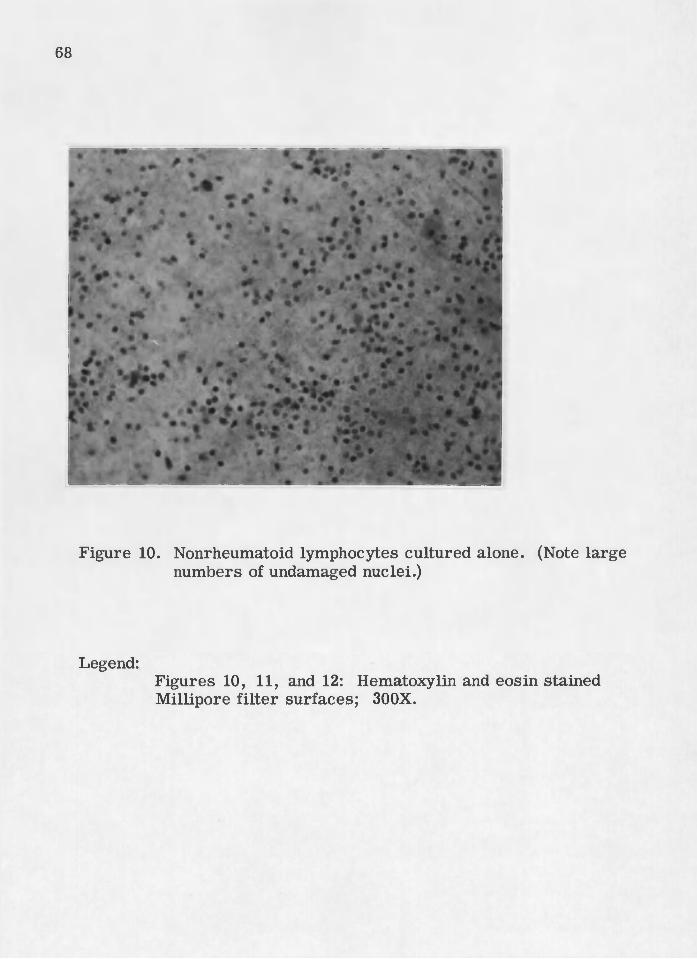
10. . Nonrheumatoid lymphocytes cultured alone.. . . . . . . . . . . . . . 6 8
11. Nonrheumatoid lymphocytes cultured near rheum a-, 'toicl synovialis. . . . . . . . . . . . . . . . . o o . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 69
12. Nonrheumatoid lymphocytes cultured near nonrheumatoid. synovialis o . . . . . 6 9
VI
Page 8
; v. ■chajpte'e r i
' INTRODUCTION ;- ; v. • - 1: : .
Rheumatoid a rth ritis is a "constitutional d isease which mani- ‘
fests itself as a profound system ic disturbance ahd by chronic, deform- •
ing, progressive polyarthritis. It has no known single cause and no
known single specific cure." It was so described by Holbrook and ; 1
mu (i). ■ V - - ' > v ^
The anatomical structure which shows very early manifestations
of the disease is the synovialis, the membrane lining the joint capsule (2).
The synovialis and capsule, being attached to the articu lar cartilage :
surfaces, form a seal for the joint cavity, which contains a viscous,
lubricating fluid—the synovial fluid. Normal synovialis is a m oderately -
cellular connective tissue which contains a capillary network near the : ■
joint cavity. The relatively smooth surface of the synovialis may be
thrown into folds either tem porarily due to the position of the joint or
permanently due to the form ation of villi which project into the joint .
cavity. The increase in size.' and number of norm al v illi is directly
proportionate to age. Normal villi consist of a collagenous fiber network
Page 9
2
containing blood vessels and lym phatics, fibrocytes, macrophages,
fat cells, and a few leukocytes (3, 4). v ; , . ' •
, Rheumatoid a rth ritis may exhibit periods of g reat activity and
periods of natural rem ission (5, 10) - In the active disease, the syno-
vialis develops many folds and, enlarged villi, which protrude into the
joint cavity. The outer portion of these villi is formed by a dense p ro
liferation of surface synovialis cells. The central portion of a villus
is highly vascularized and consists mainly of large, medium, and sm all
lymphocytes, both diffusely distributed and present in follicles. Also,
plasm a cells are diffusely distributed throughout the cores of the
The main antibody forming organs, lymph nodes and spleen,
are composed mainly of lymphocytes arid, plasma .cells. The., p resen ce :
of these cells in the cores of the v illi suggests that an immunologic
reaction is occurring in the synovialis. The plasm a ce ll has been
associated with the formation and secretion of the serum gamma glob
ulins, which eontairi the Serum antibodies (7, 8). The specific role of
the lymphocyte in immunologic reactions is as yet undefined (9). It is
known,, however, that in certain immunologic reactions, the lymphocyte
is necessary and plays a central role (33).
■ A histological picture sim ila r to that of the rheumatoid synovialis
is encountered in studies of tissue transplantation from one Vertebrate
animal to another. Of utmost importance are genetic differences not
Page 10
the specific type of tissue transferred . If tissues are tran sferred from
one identical twin to the other, an isograft, the relationship between
the recipient and the graft is the same as that occurring in 'an individual
with tissues tran sferred from one place to another on his own body, an
autograft. If a graft is made between two members of the same species
who are not identical twins, the recipient will destroy an initial homo- ,
graft by the f irs t-s e t homograft reaction. Rejection will not occur if
the recipient has been made tolerant due to prenatal or neonatal contact
with nucleated cells from the same graft donor (11, 12) or has been
made unresponsive by various s tre s so rs (11). h
. The f irs t-s e t homograft reaction is an invasion of the graft by
hematogenous mononuclear cells. This occurs after vascular flow in
the graft has been re-established from the host granulation tissue in
the graft bed. The invasion form s perivascular cuffs of proliferatihg
mononuclear cells with associated destruction. When the graft is of
skin, destruction is usually described in term s of the readily observed
changes in the graft epithelium: vacuolation of epithelial cell cyto
plasm containing invading m ononuclears,: sometimes associated with v
pycnotic epithelial nuclei. Often epiderm al regeneration processes
compete with the invasion-destruction process, resulting in excessive
epiderm al thickening. P lasm a cell nests occur in the inflammatory
infiltrate but make up no m ore than five percent of the invading cells (13).
Connective tissue damage is histologically identified with loss of ■
Page 11
nuclear staining or pycnosis and fragmentation of nuclei-(M). However,
in skin graft rejection the collagen fib rils rem ain unaffected (13).
The involvement of hematogenous mononuclear c ells (mainly
lymphocytes), plasm a cell nests, and marked thickening of the epithe
lium in the f irs t-s e t homdgraft reaction suggest the histological picture
seen in rheumatoid synovial tissues. Despite this s im ila rity ,. there are
a number of marked differences. In rheumatoid synoyialis, the follic- : ;
ular arrangem ents of the lymphocytes and other reticulo-endothelial
cells a re independent of adjoining blood vessels and therefore are not
perivascular cuffs of hematogenous mononuclear cells (15). Since this
fact was not derived from a study of the initial lesion of rheumatoid
a rth ritis , it does not exclude the possibility that this follicular arrange
ment la ter replaced the perivascular cuffs of hematogenous mononuclear
cells. ■, \
Another difference should be noted concerning the plasm a cell
involvement, although the picture is less clear in rheumatoid. a rth ritis ; '
than in the homograft reactions. In the homograft reaction pMsma cells N.
appear to be stim ulated to develop by antigens released in the invasion-
destruction process. This conelusioh is based on the following observa
tions: plasm a cells' are not with the invasive cells initially; piasm a cells
’ never com prise m o re than fiVe percent of the invading cells; plasm a : • .d }
cells may .be formed in 'areas not in immediate, contact-with th e .g ra ft (IS) '
The serum gamma globulins--hemagglutihins: (17) and leukoagglutinins (18) —
Page 12
produced by these plasm a cells are immunologically specific for the
graft donOr (19). In the rheumatoid synovialis, however^ the plasm a
cell nests have been associated with a high m olecular weight gamma
globulin--the rheumatoid factor. Two facts suggest that the rheum a
toid factor might represen t a reaction to abnormal gamma globulins.
F irs t, rheumatoid factor has a sedimentation coefficient of 22S. This
22S complex has been separated into two components—one 19S and one
S,. The 19S component is a. member of the macroglobulin class and
has the characteristic reactivity to gamma globulin shown by the other
mem bers of this class. The 7S component does not exhibit specificity
(20, 21, 22). Secondly, the rheumatoid factor reac ts with human and
, certain other species' gamma globulins which have been modified
either by being in an antibody-antigen complex, or by being adsorbed
onto a la rger particle such as latex (23). That the rheumatoid factor
does hot play the significant role in this d isease is suggested by: the
rheumatoid factor is p resent in only ten percent of the cases of child-
: - hood rheumatoid a rtb ritis (2%); the incidence of rheumatoid a rth ritis is
higher among patients with agammaglobuliriemia than among the general
population (25); patients with lupus erythem atosus, syphilis, and
sarcoidosis often give a positive rheumatoid factor te s t (24); tra n s
fusions of rheumatoid plasm a into nOnarthritic patients maintained a
high tite r of rheumatoid factor for s ix weeks without producing
Page 13
manifestations of the disease ( 2 6 ) Here, the sim ilarity to homograft
hypersensitivity re tu rns. Serum antibodies are not the important fac
to rs in homograft rejection because they are not able by them selves to
destroy tissue homografts (31, 32, 33) but, can destroy these same
cells in suspension (34), In homograft rejection, as contrasted to
rheumatoid a rth ritis , a difference is suggested by homograft tolerance
in cases of agammaglubulinemia (22) since in this condition there is a
higher than norm al incidence of rheumatoid a rth ritis (8). Medawar has
suggested that the few reported cases of homograft tolerance in
agammaglobulinemia represen t two manifestations of an underlying
phenomenon (28).
A number of studies have indicated that the lymphocyte is the
cell responsible for the rejection of the homograft. Passive transfer
of hypersensitivity to homografts by means of lymphocytes has suc
ceeded but by means of serum has failed (29, 30). The work of Algire,
Weaver, and Prehn has firm ly established that lymphocytes, or less
specifically lymphoid cells,, are the active agents of the destruction of
tissue homografts (31, 32, 33), .
The initial lesion of the synbvialis in rheumatoid a rth ritis may
involve the same meehahismms homograft rejection. An essential link
in drawing such a comparison would be the demonstration that the
lymphocyte is the agent of chronic destruction of the rheumatoid syno-
vialis, ... ■ . ■ , ^ ; " ' ' . :
Page 14
1 Si nce rheumatoid a rth ritis is a Human disease which has yet to ■
. ;;-be reproduced , in anim als, the available technics' for such a dem onstra
tion of destructive agents are lim ited. Tissue culture of human cells
■ represents one of the most widely used technics by which such destrue- ,
tive agents might be demonstrated.
Six frequently quoted investigators have attempted to demon
stra te in tissue culture that the lymphocyte is the agent of destruction ,
in the homograft reaction, an otherwise w ell-established principle.
Trowell used organ culture of lymph node fragments in contact with
fragm ents of u re te r. The target u re te r epithelium m igrated over the
sensitized lymph node but resulted only in tissue fusion, with no lympho
cytic infiltration or specific destruction (35). The two main questions
about Trow ell's experiments concern the degree of cellu lar contact and
the use of the completely synthetic T8 medium. Scothorne and Nagy
also used T8 medium for the organ culture of lymph node from a skin
graft donor in contact with regional lymph node from the homologous
Skin g raft recipient. They failed to observe any cell destruction in
sensitized system s that did not occur in controls (36), Again, cellular
contact and the use of protein-free T8 medium are factors to be con-
sidered, Medawar cultured fragm ents of sensitized lymph node and
spleen with skin from the sensitizing graft donor and found the lymphoid
cells did no damage to the mitotic or niigratory activity of the cultured;
Page 15
epiderm is (37). The cellular contact ‘may have been adequate since
some lymph nodes were completely perm eated by epiderm al cells.
However, m ost of his experiments used spleen which plays a minor
role in orthotopic skin graft hypersensitivity (36). When using lymph
node only one experiment survived beyond four days. Weaver, A lgire,
and Frehn also combined spleen and epiderm al cells in tissue culture
and sim ilarly found no clear destruction of the epiderm al cells (33). In
contrast to these four failures, two of the six investigators reported
cytolytic action by lymphocytes. Govaerts reported that living lympho
cytes from graft recipients produced specific cytotoxic lesions in renal
cell culture from corresponding donors. He found that complement
increased the cytolytic properties of the lymphocytes (38) . RoSenau
and Moon added sensitized homologous lymphocytes to tissue cultures
of mouse fibrocytes and reported clustering of lymphocytes about the
fibrocytes, which exhibited marked cytopathogenic. changes. They
observed that addition of complement and protein enrichm ent to their
completely synthetic, medium were without effect on this phenomenon (39).
W eaver, A lgire, and Frehn had reported failure in the use Of
in yitro technic in demonstrating the cytolytic properties of sensitized
lymphocytes in contact with their target cells. They did repdrt success
when this, system was enclosed'within- a diffusion..chamber implanted in
an animal (33). Their success was repeated by Amos , (61). ■ . ■
Page 16
A diffusion chamber cdnsists; essentially of a plastic tube with
M illipore filte rs sealed on both ends. The pore size of; the filte rs may
be regulated within very fine lim its. Usually, the pore size is sm all
enough to prevent cellular exchange between the chamber contents and
the host animal, but is large enough to allow the perfusion of host
in tercellu lar fluid through the chamber. A construction so simple has
many variations to fit the needs of the u se r. Algire, Weaver, and Pretm
have laid the foundation for the use of the diffusion chamber both in
general and specifically in the study of the homograft reaction (31, 32,
33, 40, 41, 42, 43, 44). Previously, the same principles had been
applied in the use Of collodion bags (45, 46) . Since its introduction, the
diffusion chamber has been used to delineate the effects of cellular
contact in contrast to humoral factors (33, 47, 52). It has been used
to study the effects of the inflammatory environment on the morpholog
ical and behavioral characteristics of lymphoid cells (48, 56, 57, 58)
and the effect of nutritional factors on any such morphological change
(49, 50). The morphological differentiation of human mononuclear
leukocytes into fibroeytes in diffusion chambers has been studied in
individuals with rheumatoid a rth ritis (50, 51) and without this disease
(48, 50). No qualitative growth or. differentiation differences were
found between norm al volunteers and persons with rheumatoid a r th r it is .
It was found that cortisone prevented the differentiation into fibro-
cytes (50). Cells and tissues in diffusion cham bers have been shown
Page 17
by the atioye reports to maiirtain fo^ ability. In addition, the '
viability is assured both by studies made on endocrine transplants in
'diifusion: chambers as therapy for endocrinological d isorders (53) and
' particularly by immunologicaliy com petent'cells within diffusion, t
chambers participating in the immunologic reactions of the h o stan i-
■ m ai:(54','55). : ■ v/ ; V V';
It is the purpose of the experim ental portion of this study to ;
compare the homograft reaction with the joint inflammation in rheum a
toid a rth ritis . The approach taken involyes the technic of challenge
studies within diffusion cham bers implanted subcutaneously in ra ts .
Using this technic in the study of homograft reaction, Weaver, A lgire,
and Prehn found that combination or challenge of sensitized lympho
cytes with their specific target, of sensitizing cells resulted in faster
and g rea ter destruction of the contents of the chamber than combina
tions of these same target cells with nonsensitized lymphocytes. This
pattern was not affected by the species of the host animal (33).
. The heterograft nature of the chamber contents in relation to
the host did not affect the growth characteristics for a period of fifteen
days (44) or the immunologic reactiv ity of the chamber contents for at
least six days (33) i Therefore, this technic represented a method for
the detection of sensitized lymphocytes apd their particu lar target cells .
This investigation recognized the above method as applicable to the ,
: study ofthe human .disease^ rheumatoid' arthritis. Thus, .rheumatoid:;: : -: pi
Page 18
: : , : ^ ^ \ f ; : : \ ; : ; . ^y ::; :,;Y ' - / ; : , , ; : - : ' ' : ; % : . .11;
and; nonrheumatoid synovial tissues were combined in diffusion .ctiaih-
bers with blood lymphocytes from patients with ahd without the d isease»:
Histological c r ite ria were employed during micrbscopic examination of
the chamber contents to determine the extent of cellular interaction.
Page 19
CHAPTER ;i l '
' EXPERIMENTAL PROCEDURE :
On the basis of resu lts obtained from six pilot experim ents, the
following procedure of choice was adopted„ ■ The diffusion chamber
technic employed was eclectically chosen from that of A lgire et aL (44)
Weaver et aL (33) , and Najarian and Feldman (54) „ After the title of
each subsection, an estim ate is included of the time required for one
person to process th irty cham bers. Manipulation tim e is extremely
c ritical, since the degree of necrosis of the cells not only increases
with tim e but also is the crite rion of damage due to cellu lar interaction
• P reassem bly of diffusion \chamber components (5-6 hours)
Each diffusion chamber consisted of one Plexiglas ring mounted
between two M illipore f ilte rs with MF cement, formulation No. 1 (Milli
pore F ilte r Corporation, Bedford, M assachusetts),,}
Plexiglas tubing was machined to ,produce rings: one-half inch
outside diam eter, three-eighths inch inside diam eter, one-eighth- inch
in height. In the side of the rings, a ope -sixteenth inch access opening
was drilled.
. . 'Y:: :
Page 20
Type HA Millipord filte rs (diameter 13 pore size 0.45
m icrons) were used. The ME cement was best applied with a glass tube
drawn put to a fine point in a flame and fire polished to produce a sm all
: behd.'"' ' ' ' . , ' . •
Millipore filte rs were placed severa l inches apart on a sheet of
alumihum foil. A-ring was held with forceps while ME cement was
- applied tp thetop-rim . '■ lt':was inverted on a filter and held firm ly until.
:' The,procedure was repeated;in attaching the ■' .
second filte r to the opposite rim of the ring. Each diffusion chamber .
was. labelled opposite the access opening in the Plexiglas ring by sea l-
■ ing pn a typewritten number with clear cement.
Sterilization of the prefabricated diffusion chambers was
accomplished by filling and c over ing them for one -half hour with 1:750
aqueous Zephiran chloride (Winthrop L aboratories, New York). Fol-
lowing chamber sterilization, all manipulations were ca rried out under
standard s te rile conditions. The chambers were removed from the
Zephiran chloride, rinsed three tim es with E arle ’s balanced salt solu
tion (BSS, Table 1), and stored until final assemblage in 100 ml. of
BSS plus 10, 000 units of penicillin-streptom ycin m ixture (Microbio- :
logical. A ssociates, Bethesda, Maryland).
: The closure for the side opening in the cham ber was a sm all
Plexiglas cone ground from rod using an ordinary pencil sharpener.
Page 21
The point was then firm ly inserted in the opening and cut to allow
approximately one -sixteenth of an inch for handling purposes. These
plastic stoppers were stored in 70% ethanol until needed.
•: ' • P reparation of human tissue . - dand lymphocyte specimens (5-9 hours)
: ,' Synoviaitissue specimen&'were obtained at the tim e.of open .-.d:
'su rg e ry ,■ transported within'One. hour to the labora to ry ln :BSSv/ith ; ;’V;
added penicillin-streptom ycin mixture (100 u n its /m l.), and incubated
at 37° C. O rdinarily, 6-9 hours elapsed before use in final chamber .
assembly, : ' ' '."i : : :
* Blood samples were obtained by venipuncture and the lympho
cytes were separated by a modified Jago (60) technic. Twenty ml. were
divided between two ste rile 15 ml. conical centrifuge tubes, each con
taining 0.05 ml. of isotonic heparin solution, 1,000 units per ml.
(Liquaemin sodium, Organon Inc., West Orange, New Jersey). The
sam ples w ere mixed very carefully to avoid the presence of bubbles in
the centrifuge tubes. If bubbles were form ed, they were aseptically
drawn off with sm all bore straight Pasteur pipets. The tubes containing
blood from patients with active rheumatoid a rth ritis were centrifuged
at 1,100 r.p .m . (266 tim es gravity, xG) for five minutes. The blood
tubes from norm al volunteers were centrifuged at 1,500 r.p .m . (499 x<3)
for five minutes. The decrease in relative centrifugal force used when
Page 22
centrifuging rheumatoid Mood sam ples compensated in p art for the
increased sedimentation ra tes characteristic of active rheumatoid
a rth ritis . With s te rile straight Pasteur.p ipets, the supernatant plasm a
and "buffy. coat” were removed and placed.in a clean, s te rile 15 mb
centrifuge tube. The cell suspension was well mixed, avoiding' bubbles,
and recentrifuged at 900 r.p .m . (179 xG) for ten minutes. The super
natant plasm a from this second centrifugation was removed down to
the top of the sedimented red blood cells, which inevitably contamin
ated the "buffy coat” in the f irs t separation. This recentrifugation
increased the purity of the lymphocyte suspension. Total white blood
cell and differential counts were made on the suspensions by standard
hemocytometer technic. In order to equalize the lymphocyte concen
trations in the normal and rheumatoid suspensions, often it was neces
sary: to Centrifuge again at 900 r.p .m . for ten minutes, to remove
carefully and aseptically a certain amount of Clear plasm a from the top
of the supernatant, and then to resuspend the sedimented lymphocytes
by gentle aspiration in the remaining plasm a volume, Immediate use
followed equalization of the lymphocyte concentrations in the normal
and rheumatoid suspensions. Approximately 2-4 hours were required
for venipunctures, centrifugations, and cell Counts,
Page 23
: : - : " v f ' ' ; \ y: ' ' ; ' ' ' ' : ' ; ' : . r ■
:: ". ■ Final assembly and sealing v .,, , , -/ v. ; of diffusion cham bers . . :■■ -V’ (2-3 hours)
Small s trip s of synovialis, approximately orie-sixteenth of an
inch wide and one-eighth of an inch long, ' were dissected on a glass -
plate with fractured razo r blades. A typical experim ental plan
required nine such strip s of each tissue (Table 2). The se ries listed
in the squares of Table 2 were the identifying numbers on the randomly
selected diffusion cham bers. Nine deep P e tri dishes were labelled for
each treatm ent and three randomly selected chambers were placed in
each dish. The diffusion chambers were balanced on the ir sides,
access openings up. Using a sm all pointed m etal probe, the appropriate
tissue slices were carefully introduced through the side opening into
the cham bers. After gentle aspiration, 0.2 m l. of the appropriate
lymphocyte suspension was dispensed from a tuberculin syringe to fill
the designated.cham bers. The lymphocyte suspensions in autologous
plasm a readily displaced any BSS inside the chamber. In control
cham bers, 0.2 ml. of BSS was substituted.
The opening in the Plexiglas ring was dried with a heated m etal
probe and a drop of MF cement was applied. Immediately one of the
dry conical Plexiglas Stoppers was inserted firm ly into the cement^ '
filled aperture. The P e tr i dish lid was left a jar for one-half hour to
allow the cement to thoroughly dry. To insure a good seal, additional
MF cement was applied and again allowed- to d ry . P he completed - , :
Page 24
chambers were stored together in 100 ml. of BSS at 37° C. until time
of implantation in the ra ts .
Subcutaneous implantationof cham bers in ra ts (2-3 hours)
Male Holtzman rats (Holtzmari Co., Houston, Texas) weighing
125 to 375 gms. were used for implantation; all ra ts in a single exper-■ . ■ ' . , ■ i. . ' ■ ■ ‘ 1 ' -*, '... ■ , -
imental run were the sam e weight and age. The ra ts were anesthetized
with ether. An incision approximately one inch in length was made
through the skin in the scapular area. On each side of the incision, a
subcutaneous tunnel was opened by spreading the connective tissue with
as little traum a to the blood vessels as possible. One randomly selected
chamber was inserted in each tunnel to a. depth of approximately one
inch. The incision was securely sutured with cotton thread. Penicillin-
streptom ycin m ixture, 1,000 units, was injected into the operation
a rea after closure of the incision. The: ra ts were placed in separate •
. cages and ra t diet (Table 3) .and water were available adZlibitum.''
. ■ ■■■ Removal of chambers /1 ; from the ra ts - (2-3 hours) >
Six days following implantation, the ra ts were, anesthetized with :
ether. The positions of the chambers were located by ca re M manipu-
Tation o f the skin of the operation area . The stitches w ere cut: and;the ry .
orig inal incision reopened. The host ra ts reacted to the presence; of /.V ;
Page 25
the diffusion chambers by the foreign-body reaction, which produced
fibrous connective tissue capsules around the cham bers. By carefully
spreading-away the connective tissue organization with the points of
sc is so rs , the edges of the diffusion cham bers were freed of the cap
sules. Exercising care to avoid puncture of the M illipore filte rs , the
chambers were removed with large blunt forceps and stored immedi
ately in 10% form alin at 37p G. for approximately eight hours.
Processing and opening ' of sealed cham bers . (5-8 hours)
: The host ra t connective tissue and blood cells were removed
from the outside of each chamber by abrasion with Kimwipes (Kimberly-
Clark Corp., Neenah, Wisconsin). The wipes were used alternately -
dry and wet with 10% •formalin. The- chambers' were opened with a
.scalpel by cutting through the filte rs a t the-inner edge of the. Plexiglas
rings. : : : . . v: /:'' : i ' - ' ' v'-V' '-
Lbosely attached protein m aterial and cells were rinsed away
very gently with., a few drops of.10%.formalin,.dispensed from a wash. ;
" b o ttle .. V/ith .fine-pointed forceps, the 'tissue'fragm ents were-carefully
removed from the M illipore filte rs and stored in labelled test tubes of
• 10% form alin until.tim e of sectioning. :
. • . The M illipore filters, were strung on; a cotton sewing, thread; with
a fine sewing needle, in se rted first"through;the abraded surface. ...An, V
Page 26
V ■ ■ ■■ ^ . ; ,'' '
accurate ch a rt was kept of the order of the filte rs on the string. .They-;
were spaced approximately one-fourth inch apart by glass beads. The
string of filters, was stored in 10% form alin until tim e of staining.
. . V Paraffin sectioning of cultured 'fragm ents of synovialis ' (16-24 hours)
The fragm ehts of synovialis were each tied in a separate gauze
bag, strung on a length of sewing thread, and identified by paraffined
paper labels between each bag. The fragm ents were transferred from
■one bath of dehydrating1 agent to another aCGbrding to- the folio wing ' ' -
schedule: 80% ethanol (one-half hour) 95% ethanol (one-half hour), a
Second 95% ethanol (one-half hour), n-butanol (one hour), a second
n-butanol (one hour), a large bath of paraffin at the melting point (one .
hour), and another. sim ilar paraffin bath (one -half hour). The use of
n-butanol in place of absolute ethanol: was adopted to give softer m ore
pliable fragm ents, easie r to section. One labelled bag at a time was
removed from the string and cut with warm Scissors. With warmed
forceps, the gauze was opened and the tissue fragm ent was gently
picked up and embedded inside a previously prepared mold of warm
paraffin. The label was inserted in the edge of the mold and the paraffin
was allowed to harden. , :
The paraffin blocks were placed in an ice bath. The ice-cold
blocks were sectioned on a microtome to produce slices 4 microns ;V
Page 27
thick. These slices were floated on the surface of a warm water bath
and straightened by pulling on the paraffin adjacent to the fragment.
The sections were transferred to clean m icroscope slides and allowed
to dry. : . ' •. ' - - . - . ■ ' ; " : '
Staining of paraffin sections ; ' and M illipore filte rs " ' (2 hours) ''
Hematoxylin and eosin stain was used throughout. Paraffin
sections for one experimental run were stained simultaneously. The
same was true for the M illipore filte rs . . '
To remove the paraffin from the sections, the slides were
placed in a rack and dipped through two baths of xylene for several
minutes each. They were then hydrated by dipping for several minutes
in n-butanol, two baths of 95% ethanol, and finally in w ater. The se c
tions were im m ersed for three minutes in H arris Alum Hematoxylin,
transferred to tap water (one minute), to tap water made basic with a
few drops of ammonia (one minute), and then to 0.5% eosin (three m in
utes). They were dehydrated by dipping quickly through two baths of
95% ethanol, and then into two baths of n-butanol and two baths of xylene
for three minutes each. The slides were removed one by one from the
final xylene bath, a drop of Permount mounting medium (Fisher Scien
tific Co., Fair Lawn, New Jersey) placed on each, - and a clean cover-
.slip positioned over the sections. Once the Perm ount had dried several
hours, the slides were ready for microscopic examination.
Page 28
The procedure for staining the M illipore filte rs was more
sim ple„ The string of filte rs needed only to be rinsed in two baths of
water to remove most of the form alin before transfer into hematoxylin.
The time in hematoxylin was decreased to one minute. The time in
eosin was decreased to two seconds. The filte rs were dehydrated and
mounted on slides with Permount. In the xylene, the M illipore filte rs
became .transparent and remained' so when permanently mounted. ,
M icroscopic examination was possible after several hours drying tim e.
" • ■ . . . Examination of slides (24-36.hours)
All examinations were made without knowledge of the identity
of the slides. Descriptions were w ritten for each filte r and for each
slice of the sectioned fragm ents. The degree of necrosis was estim ated
on the basis of a plus system . Undamaged, tissue sections and filte rs
were represented by a rating of less than one plus. Almost totally
necrotic f ilte rs or tissues were designated four plus. The c r ite ria for
necrosis in filte r examinations were two in number: (1) the absence
of added lymphocytes, and (2) the presence of debris from nuclear
fragmentation. The c r ite r ia for necrosis in paraffin section examina
tions were three: (1) the presence of eosinophilic nuclei, (2) the p res
ence of hypochromic.nuclei, and (3) a qualitative reduction in number
of normally stained tissue cell nuclei Over comparable areas in the
controls. Any disruption of the fib rilla r s tructu re of the presence of
Page 29
holes around lymphocytes were considered as possible artefacts. Upon
completion, all descriptions were sorted and compiled with reference
to the original experimental plan chart.
D epartures from the — --------OToeedtire of choice
There were six pilot studies, RI (Rat Implantation) 1 - 6, and
four experimental runs, R1 7-10.
Variations were presen t in the chamber sterilization procedure
with Zephiran chloride. In RI 7, three washes of BSS plus penicillin-
■ streptom ycin m ixture (100 units/m l.) were used. In RI 8 and 9, each ; '
chamber was washed with one ml. of anhydrous ether, dispensed from
a syringe into the side opening. Before the filte rs dried, the chambers
were dropped into ste rile BSS to prevent filte r contraction and the
resultant splitting due to excess drying.
It Was also necessary to modify the cementing procedure. ;
Instead of MF cement, a thick preparation of 4% Plexiglas in dichloro-
methane/'Was utilized. in Rl 7 and RI 8. This cement was la ter aban
doned since it was necessary to use large amounts to eliminate host
invasion of the implanted cham bers. In RI 8, an extra modification
Occurred when the side opening Was not cemented before receiving, the
Plexiglas stopper.
Page 30
AvailaMlity of synovial tissues im posed.aii unavoidable varia
tion- In RX 7, synovialis from a nonrheumatoid patient was not obtained-
In RI 9, 10, and 11, rheumatoid synovialis was not available -
Finally, in RI 7, ten hours instead of 2-3 hours elapsed before
the rheumatoid lymphocytes were used to challenge the tis su e .
Page 31
Table 1. Composition of, E a rle ’s Balanced Salt Solution (59)7
Component , . 7 Amount per lite r
' \ NaCl : ■. ' ' : v,';.; ; 6.80 gms, '
. ; 7KCL ■ 1 , - ■ , : ./ ' :::0. 40 ■:n: : 7 '■
MgSG4 .. 7 HgO '-',.7 " / ' 0.20 ' " - '
■ . ' NaHgPCy . HgQ : -; . 7 . . Cl. 1/1 : 7 '
, • -■ ; N a H C o 3 . ■ 7 ; ; - : 2 , 2 0 \ ^ 7
, Glucose : //. : . 7 : . 1 .00\ ■l' ^
, (triple distilled) to volume: • z ■ ; ■ - ; ■ ■■ ■. • ? : . ' .7.7' " . -7- : :
7, phenol red (0,5%) ; • 7 ' \ . - 0.15 mL . .. - -
Page 32
Table-2o' Sample Experimental Plan.
Tissue-v -
1 ;; . - ; ■ Lymphoeytes’ Rheumatoid
: ; 1 a rth ritis.' Normal '
' ’ controls ' Blank . ;. •
Rheumatoid a rth ritis . 10, 2 3 ,2 5 . ; ; - 7, 15, .26 - 1, 4, 11
O ther' d isorders : . ; 13r 18, -27 6, 17, 20':. ■•5,;9, 19
Blank;' ' ■ : 2,; 12, 24- 16, 21, 22 ; '3, 8, 14
Note: Numbers are labels on cham bers.
Table- 3v Composition; of .Blended S a t Diet.- ^
Compbhent . '■ : : :f ; ■
' ■ ’■ t - v ■'. . ’ i
: . . ■ % by weight- - - ■
Ground whole wheat . ,. - 58.0 „ : ;Dried skini milk . .. . ' 29.0Meat scraps 7.0Wesson oil • 4 . 4 i - . ' ;V i-
SaltCod liver oil ’■ , . 0.5.; - ’ '
Page 33
CHAPTER M
■ : : . • RESULTS': • . ■ ; ' y / = ':
Specimens of synovialis were obtained from two patients with
rheumatoid a rth ritis and four patients with other orthopedic d isorders
(Table 4)/. : \
The rheumatoid blood sam ples were from volunteers with an
established diagnosis of active rheumatoid a rth ritis , either chronic or
acute, Nonrheumatoid lymphocytes w ere separated from the blood of
individuals with no known joint d isorder or autoimmune disease (Table
The recentrifugation technic for lymphocyte separation increased
the purity of the lymphocyte suspensions. The effect of this procedure
on two blood sam ples is shown by the data in Table 0. Since the lympho
cytes .for different experim ental runs were from different indiyiduals,
it was necessary to equalize the concentrations at different levels.
Therefore, the inocula varied over a wide range (Table 7). Sim ilarly,
the polymorphonuclear leukocyte contamination varied widely.
In challenge chambers containing rheumatoid lymphocytes,
destruction of lymphocytes averaged twice that seen in contro ls-- •
Page 34
rheumatoid lymphocytes alone or the sam e tissues challenged with non-
rheumatoid lymphocytes (Table 8) „
The tissues within chambers containing rheumatoid lymphocytes
exhibited an average of three to four tim es more necrosis than tissue
challenged with equal numbers of nonrheumatoid lymphocytes (Table '9)Vv';:;
-An average of RI 10 mid 11 show: control# tissues 0.3, tissues chal-
lenged with nohrheumatoid lymphocytes 1.0, tissues contacted by rheu
matoid lymphocytes 1.7. This indicates thaty for RI 10 and 11, nonrheu-
matoid lymphocytes caused alm ost as much damage Us rheumatoid lymph
ocytes. However, in RE 7, 8, and 9, this was not true since challenge
with nonrheumatoid lymphocytes produced the same average tissue
necrosis as culture of synovialis alone.
The difference between necrosis in challenges and controls was
not g reat either for the lymphocytes in E l 10 or for the tissues in RI 10
’ and 11., The ' relatively-low lymphocyte: inpcula used in .these two ra t ■ \
implantations might partially explain the ir difference from; E l 7, 8, and
9 (Table 7). Polymorphonuclear leukocyte contamination of the inocula
imposed no conclusive stim ulatofy or inhibitory effect oh observed
necrosis. .
Photographs included.demonstrate the types of changes observed.
Figures 1, 2, and 3 display the changes observed when rheumatoid
synovialis (Figure 1) was challenged with nonrheumatoid lymphocytes .
Page 35
' : ■ ' ■ 28
(Figure 2) and with rheumatoid lymphocytes (Figure 3} „ Note the
decreased number of undamaged nuclei in Figure. 3. Sim ilarly, this
was observed for challenge of nonrheumatoid synovialis (Figures 4, 5, \
and 6). \ . . - . : - -
On the filte r surfaces, a decreased number of rheumatoid lymph
ocytes wa^ also an observed consequence of challenge (Figures 7, 8,
and 9). No such destruction was significant for nonrheumatpid lympho
cytes (Figures 10, 11, and 12) „
Various possibilities were tested for interaction between the two
chambers "a” and !!b?' implanted subcutaneously in a single ra t (Table
10). In El 7, where chambers ”an and r'bn were identical, the lympho
cyte necrosis resu lts were identical, although the tissues showed some
degree of variation. In RI 8 and 11, the condition of the challenged
lymphocytes or tissue in chamber "a" did not affect the condition of the
chamber "bn contents. In RI 9, four of the chambers ”ipin were identical
and their necrotic indices were more sim ila r to each other than to their
corresponding chamber nafrm atest For El 10, the f i r s t three combina
tions listed indicate the necrosis due to challenge with rheumatoid lymph
ocytes in chambers ,fa” did not influence the necrosis in chambers nbn,
which contained the same tissue challenged with nonrheuihatoid lympho-
cytes. Therefore, on the basis of the data presented in TabM lO, no
necrotic interaction between doubly-implanted cham bers occurred.
Page 36
Variations in technic caused only incidental differences in
resu lts . For example, in RI 8 the ex tra modification of not cementing
the side access opening before inserting the Plexiglas stopper produced
40% loss of chambers due to host invasion. No bacterial contamination
was encountered due to variation of the sterilization method. ,
Page 37
Table 4, Sources of Synovial Tissues,
Experiment ’ Sex' t T t
l : . ' A g e '? • t
Diagnosis,. "ErythrQcyte .
7 * v. Sedimentation ' RateXrnm,/hr,)-
, v M S r eoperatiye ', • - ‘ Medication
El 7 V ■ 21 ••• - Rheumatoid, a rth ritis
"7 7' Gold - . • -v .
■ F 63 \ Rheumatoidarth ritis
;94"di''vr Steroids
E l 8 m ; 27 ' Degenerativearth ritis
No steroids
.. M 18 B ilateralbunions
No steroids
- E l 10 '■ ■ M 3 7 : Degenerativearth ritis
No steroids "
y W 11 F / ■ 51 Splayfoot 7 No steroids
Page 38
Table 5„,: Sources; of"Bloed :fcymphdc3rt:es..
Experiment
t -i : S e x 'r -
. t• r';v:'4ge;
4-Steroid •iJT'; " "-pcnR < ' -• x: ' ■ •; WBC ;
, (per cu.’ mm.)
Rheumatoid A rthritisRI "7 F \ ' 86 : ; No ' 88 7,400 ; :R I8 ' M . ' ; '56 NO 49 9,500RI 9 M ■v 24 No 52 ; 9,800RI 10 : 6 3 ' No ■■ 41 10,000
. RI 11 M . '5 2 ' Yes 45 8,000
Normal Controls
RI 7 F : .24 No 18 8,800
' RI 8 ' ; , H ; : : 21 ; ■ No 2 ' 7,000VRI 9 F ' 3 3 -. No ; 1? 6,600
RI 10 ; Vm . 40\ No ' : o; . 6, 600
. RI 11 ; ; m . ; ■ :3 ; 4 q 3 No ' 0. ;• ' ' . 6 , 600';
Note; ESR ss Erythrocyte sedimentation ra te . / ; WBG @ Wtiite'blboti^c ' ,
Page 39
32
Table @ = Effect of Recentrifugation Technic on Two Blood Samples.
Separations; t
Iv ;. ... . 1 . . . ■ ■:'.R m N /m i. /
Rheumatoid A rthritisNo. 1 plasm a
supernatant : 266 : 5 200,000 100,000
buffy coat layer . ' 27/300,000 20,600,000
No. 2 plasm a and buffy coat combined
179 10 900,000 200,000
■ ' No. 3; v concentrated 4 tim es \ ■ V 179 ' IQ ; ;; 3,000, 000. ■ 90,000
Normal Control '' No. 1 : : : 499 ' , 5 2,050,000 / 150,000
' No. 2 concentrated 1.5 tim es 179 10 3,800,000
Note: RGF ^ relative centrifugal fo rce / xG « tim es gravity; t ® tim e .in miiiutes; -SL - sm all lymphocyte; PMN polymorphonuclear
. leukocyte. . : ■ / - • : .
Page 40
Table 7, Leukocyte Inocula per Chamber .
Experiment ' Rheumatoid A rthritis .. ' Normal Controls :, r . . No. of.SL * . No. Of PMN -' No. of SL ' - No., of PMN: -
' RI '7 • - ■ ••■'z; 250,000;; : 200,000 2;i5O;OO0 V ' 95,000 ' - -
,m 8 : ■ 1, 200,000 : ■■ ■' 0 - ' ■ ' ■ V930, 00d\'f ; :;-v - 18,000 - '
:66o, ooo ; . c"O5B.0OO ■;y: •
; ; ■ Rl 10 000,000:; 1 8 , 0 0 0 v.:; ■ 770,000 ,
RI 11 'iPyOOO: hv-;■ •
/ \ 770,000 . ; ' ' V'v
Note: SL « sm all lymphocyte; PMN polymorphonuclear leukocyte.
03W
Page 41
34
Table 8. Destruction of Lymphocytes Shown by F ilte r Examination,
Experimentt
. ? .. . , Estim ated N ecrosisf
, L*' 4- t* 'L *4vt , i + t^ :L4-,t': '
m 7 . ' 1. \ 4-4" 4-4-4- :y'<:4.y:2 44- v 4-4-4- ' -<. 4- -3 - 4 - 4 - , 4-4-4-
R1 8 1 4- 4-4- 4-4- < 4- ; . / i - :;v--;;:_ 4-4-2 , :>-■++: 4-4-4- i i . . 4- 4-
3 : .. i - i . 4-4-4-4- ■ " i . .■ i. ’ - I , i -RIB. 1 •• 4-4- 4-4-4- 4- 4- - 4- 4-
2 •4-4- 4-4--K4- - 4- 4- 4- 4-
3 • 4i4-4-4- : 1 'i '
4 i ' 4-4-4-4-RI 10 1 <.+ 4- ' < 4- < 4-
2 > 4- - 4-4- : : 4- 4- ' .
,3 . la ; . 4-4-4- ‘ - r - . 4- 4-
m i i . 1- K < 4- - 4-4--H < 4- : ■-'<■ +'2 .. 4- 4-4-tf < 4- ■ 4-
3 : • . ' i ' - 44-4-4- i • 4- '
Average l: 1,4 ■: 2,6 3.2 0.4. 1,3
Note: Rheumatoid: - lymphocytes, t* r synovialis; nonrheumatoid:L - lymphocytes, t - synovialis; i ss host invasion; LA s laboratory accident. ' ' ■ -
Page 42
Table 9. Destruction of Tissue Fragm ents Shown by Paraffin Sections,
i Estim ated Necrosh i r ; - - '' ? . v".;'"'."'
' r & v i
? ■- ;
RI 7 - ‘ '**+*: 2- '; :: W ' ; i . . ■***": .
.’ s ' i-" ft*"M 8; I,. '; : '- ,V:; \ . i - . •< 4- i'
-i-f-; : " : ' ; i i i i ; .+4-+ ■
■ i D i ■ i r, i;;': . y'ii- i . ' _ i■ , . " t t :
RE: 9 \ : ' .;¥■ i . : ; < + . , "< 4-. ' " . 4- . - '
. < 4- '
; 4" : ; ; :.4-4-4-'- 4-44-
RE 10 ' i i - < + : " . <" + • .
’ 2. ' : ; - ^ 4- -. + •.
3 ;;';i "V: ;:4-;- : "■+■ +4- '■ : v, + t * . ;< : 4-. :. 4-4- - - -
/ - < 4- - ; ' :
;-3 .4-4-:'"':. r : :f+ /y . ' .
A verage': ; : i 0*8 :: 0.7; 0 :0 -:
Note: Rheumatoid: lymphocytes ? t* - synovialis; nonrheumatoid:: L- - lymphocytes, \;t': - synovialis; 1. ^ .hpst invasion; DA /«' labor a- v tory accident, ' ...........................
Page 43
36
Table 10./' Effect of Double Implantation on Chamber N ecrosis.
. ■ : C h a m b e r . .c o m b i n a t i o n s r
-E y m p h o .c y te '■ ■ n e c r o s i s ;
. T" 1 '<
' ' - T i s s u e / ;-;v n e c r o s i s
■ ■ ' / - a Z';.: t • a - ' " b ;
R 1 .7 ; L * ■ < r :. f f . - ; / ' .
t * ; W - h M ' :" ; / / f f f ’ ' / ' f f f ',' f f f f : ' f f f ; / ' - ,
' L • . ' f ■' -■
' ; ' : • L -h ' t • l f t ■: ' ;'< ; + . f f f - .v , , f , , ' ■ : v’
R I 8 . L * > t - " : ~ Z T f f f f < 4 ■
. L * ' + t* /
L + t t* f - f
R i l l - L*' + t L * ■ f f f f
L +. t - L * ; : j < f .4-''
' . L ^ t L : ; . f . < f
H I 9 L L * ' f t ' : f f ■
: . L * - ::L * ' ' f t / : ' f f ; ■,++++'■'
• L -f t ' L * - f t - " ' . ' f f . ++++ 4- '' , ’ . 4-4"4- - ;
L +■■ t . m ;f tV :;;;': ’ , ++*f+ ; < K ' - 4- "
■/ / ';; l > t " ^ 4" . 'C 4*
H I 10 L * -f t ' :/ L > t : •; : : f f ; ,< 4- ' -< 4* ; :
L * -f t L f t ' 'O f f f - ' f • ■ 44*4* ' , ; 4
' ' L * ' - f t L f t ' '• f ■; 4 ; ■' 4 4 •
t ' . ; t . \ v < f : " < f c
Note: Rheumatoid: L*- lymphocytes, t* - synovialis; nohrheumatoid: L - lymphocytes, t - synovialis; i ® host invasion.
Page 44
CHAPTER IV
DISCUSSION ; - .
This study has dem onstrated that rheumatoid lymphocytes are
agents of destruction to synovialis within diffusion cham bers. There
are three possible explanations for this resu lt. One, the destruction
involves an immunologic mechanism presen t within the human body.
Two, rheumatoid lymphocytes might be m ore readily lysed than non-
rheumatoid by toxic synovial tissue products. In turn, the products of
lymphocytic lysis might have damaged the tissue. 'Three, rheumatoid
lymphocytes might have a g rea ter metabolic requirem ent than nonrheu-
matofd for some nutrient also required for tissue survival but present
only in low concentrations in the chamber fluid. The la tte r two explana
tions represen t possible, artefacts produced by the s tre s s of the diffusion
chamber environment.
The f irs t and m ost plausible explanation is that necrosis within
the chambers is the resu lt of combination of sensitized lymphocytes
with their specific antigen-containing target tissue. Thus, it would be
implied that the. rheumatoid lymphocytes are cells sensitized against
or causing destruction of synovial tissue. Also, it would be indicated '
' - V:-: ' ; ;37 • v ■
Page 45
. tfaat in rheumatoid a rth ritis , the blood ca rrie s lymphocytes with such,
destructive potential. The foundation for this explanation lies in th ree
points of analogy to the homograft reaction. F irs t, rheumatoid a rth ritis
is thought to be a possible immunologic disorder; the homograft reaction
is known to be an immunologic mechanism. Second, the histologic p ic
tu re seen in rheumatoid synovialiS is sim ilar to that observed during
a firs t-S et homograft rejection. Third, if rheumatoid lymphocytes are
sensitized lymphocytes,; the observed destruction could have been p re
dicted since hom ograft-sensitized lymphocytes will destroy their geueti-
cally:-specific ta rget tissue within diffusion chambers (33). Therefore,
it is quite plausible that within both diffusion chambers and the human
body, rheum atoid lymphocytes destroy synovial tissue by some type of
immunologic mechanism; thus, rheumatoid a rth ritis can be concluded to
be an autoimmune disease. .
However, it is plausible on the basis of this experimental study
that rheumatoid lymphocytes are m ore easily lysed than nonrheumatoid
by toxic synovial tissue pr oducts... if so , the r esult of th is type of destruc
tion of rheumatoid lymphocytes might be a complete poisoning of the '
fluid of the chamber by term inal re lease of toxic substances:. These
toxins would in turn produce nonspecific destruction before the fluid
exchange between the host and the chamber would be capable of removing
them; Two observations, if present, would support this second explana
tion: (1) if rheumatoid lymphocytes in control cham bers showed m ore
Page 46
autolysis than sim ilar nonrheumatoid controls; and (2) if tissue necrosis
seemed independent of d irect contact with lymphocytes.
Lysis of rheumatbid lymphocytes averaged th ree tim es that of ;■
nonrheumatoid lymphocyte controls (Table 7). However, in RI 7, the
rheumatoid lymphocytes had been incubated for a longer tim e before
Use than the nonrheumatoid. By elimination of both RI 7 lymphocyte
controls from consideration of Table 7, the average control lympho
cyte necrosis becomes 1.2 for rheumatoid, 0.8 for nonrheumatoid. The
subjective plus system is not sufficiently accurate to base a qualitative
difference on these figures. The conclusion must be made that rheum a
toid lymphocytes in them selves are only very slightly less stable than
nonrheumatoid. Other investigators using diffusion chamber technic
have not reported any difference in stability, growth, or differentiation
characteristics of rheumatoid lymphocytes as compared to other types :
(50, 51). While these two facts do not invalidate the hypothesis that
rheumatoid lymphocytes are more easily lysed than nonrheumatoid by
synovial tissue products, they certainly detract from any argument
based on a difference in stability. -
The-second hypothetical observation which would support the
possibility of nonspecific destruction is: if tissue necrosis seemed
independent of d irect contact with lymphocytes. It was impossible to
determine the mechanism of the destruction by examination of the slides
prepared from the cultured synovialis fragm ents. There was no
Page 47
indication that ce ll-to -cell contact was required to produce destruction
because no great degree of invasion of cells by lymphocytes occurred.
For this point an explanation might lie in the work with the
homograft reaction (33), Where destruction within the chambers was
described.' It;was stated that contact between lymphocytes and the
target cell was necessary for initial destruction; A consequence of
contact destruction of the target cell was subsequent death of the adjoin
ing lymphocytes. Destruction of both cell types f irs t occurred at the
surface of the tissue fragment. The longer the cham bers remained
implanted the m ore degeneration occurred in the target tissue beyond
the ,rzone of contact’5 with homOgraft-sensitized lymphocytes. Weaver
e t al. (33) explained this type of destruction on the .basis of the possible
re lease of a diffusible toxin following the initial immunologic surface
necrosiso Connective tissue fibrocytes have cytoplasmic boundaries :
Which a re very hard to ;descry^:;-Therefore, :it is possible that m icro
scopic examination might have^failetf to • re v ea l, the' ■mechanism of initial /
destruction of synovialis. - Thus, the obvious consequence of challenge
might have been necrosis resulting from the term inal re lease of diffus
ible toxin. The toxins might have caused tissue cell necrosis and in
addition caused the fragmentation of adjoining lymphocytes.
. Also, it is plausible on the basis of this study that the tissue
destruction Without invasion by lymphocytes is the re su lt of an artefact.
A g reater metabolic requirem ent in rheumatoid lymphocytes than
Page 48
nonrheumatoid may exist for some nutrient, also required for tissue
survival but p resen t only in low concentrations in the diffusion chamber
fluid. This would resu lt in starvation of the immobile solid piece of
the synovialis by the mobile lymphocytes. This tissue destruction with
out lymphocyte invasion would in tu rn cause partial lymphocyte destruc
tion due to the term inal release of toxins. With the data presented in
th is study, this possibility cannot be eliminated. Despite the lack of
reported differences in stability, growth, or differentiation characte r
istics between rheumatoid and other types of lymphocytes (50, 51), tbe
yreasobihgism ot'm ^ ' -
The tissue destruction within chambers does not appear to be
produced by the contact immunologic mechanism reported by Weaver
e t a h ;(S3). It is reasonable to postulate an immunologic mechanism
sim ilar to homograft reactivity in which destruction is present without
cell-to^celi contact. The work of Amos and Wakefield (61 > 62, 63), ;
Gabourel (64), and Algire (65), Najarian- and Feldman (54), and a theory
Of delayed hypersensitivity published by Karush and Eisen (66) suggest
the destruction within diffusion cham bers may be the resu lt of the p ro
duction of a high-affinity humoral antibody in low concentrations. ;
In order to proceed, an im portant question requires answering.
. Are peripheral blood lymphocytes, used for challenge in this study,
capable of prbducing some humoral type of antibody response? Humoral
antibody is usually considered to be formed only by plasm a cells.
Page 49
Fishman (68) initiated an antibody response in tissue cultures of ra t
lymph node cells as early as five days without the formation of plasm a
,cells . He detected low 'Concentrations of a diffusible type of antibody ;
by a very sensitive m ethod--neutralization of infectiyity of bacterio
phage T2 in E. coli. With rabbit thoracic duct lymphocytes within dif
fusion cham bers, a prim ary antibody response to a protein antigen was
obtained (55): Lymph nodes may contribute lymphocytes to the p eri- . :
pheral blood by way of the thoracic duct. Lymphocytes from the c ircu
lating blood make up a large portion of the hematogenous mononuclear
cells responsible for homograft rejection (13).. Finally, in man, tra n s
fer of peripheral blood leukocytes can readily transfer tuberculin sensi
tivity (67), Therefore, the peripheral blood lymphocytes obtained in
buffy coat are possibly immunologically competent c e lls , capable of - ..
some type .of antibody response. Sueh antibody production might be y
effective in a diffusion chamber but extrem ely hard to detect in vitro
since the diffusion chamber has been shown to provide better nutrition
for antibody producing cells than Eagle’s , Puck’s, P a rk e r’s 199, or
T row eirs tissue culture media (68). , :
An analysis of the data presented in Tables 8 and 9 may help to
• determine whetiier the destruction is tissue specific or lymphocyte
' specific. That is , by analyzing Tables 8 and 9 together, one might be
able to determ m e which cell was the f irs t type to die within the challenge
cham bers. The assumption would be that the cells which died f irs t would
Page 50
show the greatest difference in necrosis over the: controls. Thus, if
in challenge chambers tissue necrosis was three plus over the control
tissu e’s one plus and the rheumatoid lymphocyte, necrosis, was only one
plus g rea ter than the lymphocyte Control’s one plus-, then it can be
said that the destructive mechanism was tissue specific. In Table 11,
the data indicate that for challenges with rheumatoid lymphocytes, the
necrosis is in fact g rea ter for the tissue than for the lymphocytes
except in HI 11. The data show that for challenges with nonrheumatoid
lymphocytes, the necrosis is not g rea ter for the lymphocytes than for
the tissue to an extent approaching s ta tis tica l significance (p@ 0.2 -
0.1)o Not enough data exist to draw any definite conclusion about the
challenge of rheumatoid tissues. Here, the situation is complicated by
■ the presence of large numbers of autologous rheumatoid lymphocytes
in the tissue itse lf. To take exception to R I 11 would not be unreason
able since the control rheumatoid lymphocytes were agglutinated but
definitely present. Furtherm ore, a six tim es lower inocula of lympho
cytes was used compared to RI 9 and 10. The need for m ore experi
mental work is indicated.
Certainly the argument for some type of immunologic mechanism
is strengthened by the following facts: (1) in challenges with nonrheu
matoid lymphocytes, the necrosis is definitely not tissue specific; and
(2) in challenges with rheumatoid lymphocytes, 75% of the. experim ental
data indicate that the necrosis is tissue specific. Competition for some
Page 51
necessary nutrient might also produce tissue specific destruction. This
type of destruction would not be produced by lysis of lyraphocytes.'
With homograft reac tiv ity / four explanations are possible for
the absence of cytotoxicity in vitro (33, 35, 36, 37) but presence in
diffusion cham bers (33, 61) i F irs t, the degree of cellu lar contact has
been indicated as important in vitro (38, 39) but relatively unimportant ,
for challenges within diffusion chambers (33) „, Secondly, the fact that
tissue culture provides Subpptimal nutrition to antibody-producing cells
has been considered a deciding factor by some w orkers (33, 68), Third,
a large vblume of fluid medium is required to maintain cells, in vitro as
compared to the diffusion chamber. This has been suggested as being
deleterious to a challenge effect because of dilution of any diffusible
toxin or antibody produced by the lymphoid cells. However, in diffusion
cham bers, the low cellular growth ra te requires so sm all a volume of
nutrient fluid that a diffusible toxin or antibody might reach sufficient V,
concentrations to produce a clearly observable effect (33), Fourth,
complement, a complex mixture of plasm a globulins known to enter
into some immunologic reactions, has been implicated as the principal
difference between in v itro and diffusion chamber challenges (33),
Complement form s 0;5% of the total serum proteins and its
participation in immunologic reactions Is not antigen specific. When
complement is removed from the human body, its activity rapidly d is- ,
appears within 24 hours at room tem perature. It is required for lysis ..
Page 52
of bacteria or erythrocytes by antibodies (69). Complement is necessary
for lysis by tuberculoprotein of leukocytes from tuberculin-sensitive
individuals or norm al leukocytes if in the presence of an undefined glob*-
ulin factor from tuberculous plasm a (70). Complement is required for
th e ly s is of .mammalian cells by heteroantibody, i.e ., an tisera from
animals of a different; species injected with such cells (71). In the papers
reporting cytotoxic activity of lymphocytes in tissue culture, one reported
, that complement increased the effect (38), the other stated complement
was without effect (39). The form er also reported that complement was
required, for the lysis of cells by heteroantibddy but isoantibody (from ■ '
homologous serum) was without cytolytic effect Oven with the addition of
complement. ' .
Pertinent to any discussion of the influenCe of complement in
tissue culture cytolysis are the comments of Pulvertaft et al.(72)„ The
principal te s t for m easuring the effects of complement is the loss of
cytolysis when the challenging fluid has previously been heated to 56° C.
for 20 minutes and its Subsequent return when guinea-pig complement
is added (69, 72). Pulvertaft cited the finding bf heat-labile antibodies
a lso destroyed a t 56°; C.> namely: . reagins, autoantibody in a case of
hepatic c irrh o sis , and autoantibody in HasMmoto se ra . He also stated:
that his thyroid cell cultures were not Susceptible to damage by Hashi-
moto se ra if the cultures were four to six days old (72). These additional
Page 53
facts show the complexity of evaluation necessary in any tissue culture
report of failure in demonstrating cytotoxicity.
Complement has been shown to be an important factor for the
prpduction of cytptoxie effects by isoantibody on cells within diffusion
chambers (61 > .62, 63} and on cells in vitro (73)„ The significance of
these reports for the present investigation is that complement may be
the factor necessary to show the cytolytic action of antibody, if such is
produced against synovialis by rheumatoid lymphocytes. This possibil
ity is strongly supported by the work of Amos (61). His system involved
diffusion cham bers within the peritoneal cavity of m ice. Substances
were added to the peritoneal fluid; subsequently the host serum , the
peritoneal fluid, and f%e:'diffusion cham bers were - sampled as a function
of time... It was'found that the M illipore filte rs of the chamber acted as
simple b a rr ie rs tb diffusion. The ra te of entry and the equilibrium con
centration of substances in the chamber were dependent upon the com
position of the peritoneal fluid. This was shown not only for oxygen
tension changes but also for entry of antibody and com plem ent.: As an
example, when antibody was injected into the peritoneal fluid, equili
brium with the blood was established within thirty minutes;' but in this
time only sm all quantities had penetrated into the chamber. < Approxi
mately one hour was required for the chamber to attain equilibrium with
the peritoneal fluid. Antibody passed very rapidly from the peritoneal
cavity into the blood, but very slowly in the reverse direction. It was
Page 54
then determined that the .complement concentration within the chamber
was approximately one-tenth that found in the serum . A fter determ in
ation of the norm al growth ra te of mouse ascites tumor cells within
diffusion cham bers, isoantibody was injected into the peritoneal cavity,
resulting in a depression of the growth ra te . Further depression was
produced only by the addition of complement to the peritoneal fluid.
Thus, it was determined that complement was a: controlling factor in .
the cytotoxicity of isoantibody to a homograft.
Some emphasis is now placed on the role of antibody in at least
the second-set homograft rejection, the reaction which is possibly
analogous to rheumatoid arth ritic inflanamation . In the work of Algire
(65) ,< Amos and Wakefield (61, 62, 63), and Gabourel (64) 5 antibody has
been implicated as being functional in Some types of homograft rej ec - :
tion. These investigators enclosed within diffusion cham bers the same
type of homologous cells which had previously been injected into the
host up to four tim es, once weekly, to ' 'hyperimmunize'? the animats
Antibody to the chamber contents was found to penetrdte the chamber
in sufficient amounts to cause either lysis or growth inhibition of the
cells. Cellular contact of host with chamber contents was not necessary.
Two facto rs should be mentioned. The chamber contents were always
cells without a supporting strom a. Gabourel (64) sta tes that no Signif
icant growth inhibition of mouse L-fibroblasts, occurred within the
cham bers, while reactions analogous to f irs t- and second-set homograft
Page 55
rejections were pcctirring outside in the host. However, this does not
rule out the possibility that the diffusion of any antibody from the '
serum , to the peritoneal fluid, and then into the chamber was insuf
ficient to be beyond the adaptability of the cells. :
In addition, the work of Naj arian and Feldman (54) indicates
that a diffusible factor can be responsible for accelerated rejection as
early as the second-set homograft reaction. Their sensitized lympho
cytes were within the diffusion chamber! constantly releasing into the
blood of nohimmuhized hosts some necessary diffusible factor. It
accelerated the rejection of skin grafts placed outside the cham bers.
T his; experimental resu lt certainly supports the other information con- :
cerning the role of a diffusible substance in homograft rejection.
Also, the slight but s till six tim es g rea ter necrosis of nonrheu-
matoid tissue challenged with nonrheumatoid lymphocytes when com
pared to honrheumatoid tissue controls, should be explainable in te rm s
of this system . This necrosis in a homologous but rionsensitized sy s
tem is also reported by; Weaver e ta l.! (33). For homograft challenge
system s within cham bers, it was Stated that, compared to sensitized
lymphocytes, the homologous but nonsensitized lymphocytes caused
slower and leSs destruction of the target ce lls . It is possible that here
the lymphocytes slowly became sensitized in response to homologous
tissue antigens. They might have produced some diffusible-antibody !-
which was responsible for tissue destruction without ce ll-to -cell contact
Page 56
- . - ' ■ V ; ■ 49
From examination of the sm all amount of data in Table 11, however>
the damage to synovialis by nonrheumatoid lymphocytes does not
appear to be tissue specific. :
A number of minor facets concerning the experim ental p ro
cedure employed in this study- rem ain to be considered.
.. Control and treatm ent diffusion chambers were often placed in
the sam e rat%, thus reducing the effect of biological variation of nu tri
tional status among m em bers of the inbred ra t strain .
-Since the cham bers were picked at random for implantation,
interaction between the two chambers would have caused a random
necrotic resu lt in Tables 8 and 9. That interaction was not detected
(Table 10) is reasonable since Weaver et al. (33) noted no differences
in challenge resu lts due to the immunization status of the homologous' " ' ■ > : : ' -
chamber hosts. - v.. - -
The staining procedure with hematoxylin and eosin always p ro
duced over staining of undamaged nuclei, thus eliminating the critic ism
that hypochromic nuclei were not necrotic but phly under stained. P o ss i
bilities of relative staining variations due to tim e and composition of - ;
stain were eliminated within one experim ental run by staihing all slides
and a l l filte rs simultaneously. Variations between experim ental runs
were cut down by rigidly fixing all staining tim es and using fresh solu
tions .
Page 57
; - : . 5 0
.Other sources of e rro r in this study mainly, involved the paraffih
sections. They were heterogenous, since necrosis existed in various
form s. Also, some of the fragm ents were sectioned closer to the
i center than others, etc. Reporting resu lts in term s of a plus system
prevented prejudicing of resu lts .
Quantitation of the degree of necrosis Of the chamber contents
is extrem ely vital to further work. Detection and quantitation Of some
property produced by necrosis, ra ther than quantitation, of the loss of
some healthy function, would allow experimental, variation to; produce
resu lts beyond the capability of visual estim ation.
Page 58
5 1
Table 11. Challenge Necrosis - - Lymphocytes and T issue Compared.
. 1 . . ' . ' ' ' t _ '
. ’ ' t ' .Experiment ,
: . L* + 't ,’ . Corrected Average ? • N ecrosis1._ . -t L + 't ;
• : : " V;': h ; : ■ ' L* r
TT . t .:. ■■■ :y^;
'R IB .,: ; : d. -'d" . .'"2,0 - .2.3 ' -y -I d'2.0 0.0
R I9 ' I;, 1.8 2,2 : :■ o.o 0.3
RI- IO' " / . 1.0 1,3 ' 1.0 1.0
■■ Hi i i ' - ' '■ 3.0 2.0 ; l,o 0,7 •
•Average 1 2.0 ' -2,0/ \ 1.0 0.5 ,
Note: Rheumatoid: £«* - lymphocytes, t* - synovialis; nonrheurnatoid: L lym phocytes,/t - synovialis; corrected necrosis ® challenge necrpsis - eohtrol necrosis. '% . - ' ' '
Page 59
CHAPTER V
'StMMARY
Cix synovial tis su es , - two rheumatoid and four nonrheumatoid,
were challenged within diffusion cham bers by inclusion of lymphocytes
obtained from subiects with and without rheumatoid a rth ritis
In. chambers containing rheumatoid lymphocytes in contact with
either type of tissue, necrosis averaged three tim es that in controls i,
.1 The necrosis was observed to occur without extensive ce ll-to -eell con
tact- Following are the four possible m echanisms, here arranged in
descending order . F irs t, am immunologic mechanism: peripheral blood
lymphocytes were possibly capable of producing or releasing a diffusible
, high-affinity:: antibody. Second, an artefact: a difference in the two types
of lymphocytes existed in the requirem ent f o r .some essential nutrient
present only in low concentrations in the diffusion cham ber fluid. This
would resu lt in different degrees of starvation of the immobile, cen tra l
ized tissue fragment by the mobile lymphocytes at the periphery of the
chamber. Third, an artefact: a difference in stability of lymphocytes
existed when in the presence of tissue metabolic w astes, resulting in
: dichotomous; degrees of poisoning of the chamber' flu id , by the• te rm in a l'; •
5 2
Page 60
release of toxins. Fourth? an immunologic mechanism: initial necro
sis was contact produced. The term inal release of diffusible toxins
caused the observed necrosis and lys'ed the adjoining. lymphocytes.
More experim ental work on an exact quantitative basis is needed to
arrive at a definite conclusion concerning the cause and meaning of the
observed necrosis attendant upon challenge with rheumatoid lympho-
Page 61
LITERATURE CITED
ChicagoL :Holbrook, W, P=, and Hill, D. F„: Manual of Rheumatic D iseases„
' Chicago, Y ear' Book Publisherh', Ihc.-, ■ 1952, p. 15.
2„ ’ Collins, D. H o , : : The Pathology of A rticular and Spinal DiseasesJ . London, - Edward Arnold and C o,, 1949, pp. 169-205.- ■
3» ■Gardner,-: E. ': The structure and function of jo in ts. In Hollander,'..J . Lo, Ed., : A rthritis and Allied Conditions« Philadelphia,
. ' .. . Lea and Febiger, 1960, pp. 44-48. .
4. Maxiniow,, A. A.,- and Bloom, W. : Joints and synovial m embranes.In A Textbook of Histology. Philadelphia, WV B. Saunders
-Co., 1957, pp. 154-157.
5. de Forest, G. K., -Mucci, M„. B„,. and Boisvert, P . L. : Two year.
6. Allison,. N., and Ghormley, R. H. : Diagnosis in Joint Disease; aClinical and Pathological Study of A rth ritis . New York,
. Wm. Wood and Co., 1931, p. 196.
7. Roberts, K. B., and Gowans, J . L ..: The localization of antigens
9. Burnet, P. M. : The Integrity of the Body. Cambridge, Harvard ' . - University P ress,. 1962,; p.. 54. . , -
comparative study of se ria l hemagglutination tests done on groups of rheumatoid a rth ritis patiehts. A rth ritis and Rheumatism 1: 387-409, 1958.
and the sites of antibody formation. In Florey, H. W., Ed General Pathology. Philadelphia, W. B. Saunders Co., 1958, pp. 729-736. ■: ; ■
. i
8. Gitlin, p . , Janeway, C. A., Apt, L ., and Craig, J . M. :: Agammaglobulinemia. In Lawrence, ELS., Ed. : Cellular and Humoral Aspects of the Hypersensitive States. New York, H oeber-H arper, 1959, pp. 375-441.
54
Page 62
55
10. Robinson, Wo D. : The etiology of rheumatoid a rth ritis . InHollander, J , L,, Ed, r A rthritis and Allied Conditions, Philadelphia, Lea and Febiger, 1980, p, 171, v
11. Medawar, P, B, : Theories of immunological tolerance. InWolstenholme, G, E, W,, and O’Connor, M,, Eds, for . Ciba Foundation Symposium: Cellular Aspects of lmmun-
. ityv Boston, L ittle, Brown and Co, j 1959, pp. 134-156,
12. Hess, E, V,, Ashworth, C, T ,,. and Ziff, M,;:- T ransfer of. an autoimmune nephrosis in the r a t by means of lymph node cells, J; Exp, Med,-115; 421-480, 1982, •; y'v'
13. Waksman, B. H , : A comparative histbpathological study of delayedhypersensitive reactions. In Wolstenholme, G, E. W., and O’Connor,. M., Eds. for Ciba Foundation Symposium: Cellular Aspects of Immunityo Boston, L ittle, Brown and Co.,
' 1959, pp.-288-295. ; :
14. " - v I .:! A comparative histopathological study of delayed.. .hypersensitive reactions. In Wolstenholme, G. Eo. W., and
O’Connor,. M,, Eds. for Ciba FoundationHymposium: Cellular Aspects of Immunity. Boston, L ittle, Brown and Co.,
. ' ' 1959, p. 2B3. : ■ , -'V.k. . - ;
15. Rosenberg, E. F. : The pathology of rheumatoid a r th r itis . InHollander, J. L., Ed, : A rthritis and Allied Conditions,
r, Philadelphia, Lea and Febiger, 1960, p: 183. ' , ,
16. Waksman, B. H. : : A comparative histopathological study of delayedhypersensitive reactions. In Wolstenholme, G. E. W., and
. O’Conndr, M., Eds. for Ciba Foundation Symposium: Cellular Aspects of Immunity, 1959, pp. 315-319, •
17. GOrer, P. A. : Some reactions of H-2 antibodies in vitro and in■■■ ' V vivo. Ann. N.Y. Acad. Sc, # : 707-721, 1958. ' /V' : ;
18. Amos, D. B ,.: The agglutination of mouse leukocytes by iso- ,/ / ; i mmune se ra . B rit.-J. Exp. Path. 34: 455-470, 1953.
19. Gorer , P . A. : Interaction between sessile and hum oral antibodies;- ' : - -..k , in homograft reactions. In 'Wolstenholme, G.- E, W., and :
O’Connor, M., Eds. for Ciba Foundation Symposium: Cel- k-kk-'-'k-'k;'Vk lular Aspects of Immunity. Boston, L ittle, Brown and .Co.,
■ - Vk kA.ks :-v-?1959V ASl-SSS. ; ^ ; v,:.k ,-::...;k
Page 63
5 6
20= Robinson, ,W.. D. : The etiology of rheumatoid a rth ritis . InHollander, J . L., Ed. : A rthritis and Allied GonditionS. x
, ' ’ Philadelphia, Lea and Febiger, i960, p. 173. ' v x': .::"v;
, 21. ■ Kunkel, H. G’., Franklin, E. C., and M uller-Eberhard, H. J. : : :Studies on the isolation and characterization Of the ’'rheu^ matoid facto r.n J . Glin. Invest. 38: 424-434, 1959.
22. Lospalluto, J ., and Ziff, M., : Chromatographic studies of the - , :rheumatoid factor. J . Exp. Med. 110: 169-186, 1959. ; :
23. M ellors, R. C., Nowoslawski, A., and Korngold, L. : Rheumatoidarth ritis and the cellu lar drigin. of rheumatoid factors. ;
' : Amer. J. Path. 39: 533-546, 1961. . . -x, , \ '
24. Vaughan, J . H. : The study of the rheumatoid factor. In ;Hollander, J. L., Ed..: A rthritis and Allied Conditions.
. ■' ' ■■■.. Philadelphia,' Lea and Febiger, 1960, pp. 92-95. ■ ■ V/Vx ;:x;
25. 'Good, R, A., Rot stein, J .> and M azzitello, W. F. : The sim ul- '''d-'dxxtaneous occurrence of rheumatoid a rth ritis and agamma- x : "
; , : globulinemia, J. Lab. and Clin. Med.'49: 343-357, 1957. ixx'..'',y
26. Vaughan, ■ J . H, , and H arris, J , : Transfusion of rheumatoid -' 1 : plaSma and cells. A rthritis and Rheumatism 2: 51-53,
;; 27.x,:Gdod,xR. A;?;:;Varco,; S. J . T ransplantation studies in patients with agammaglobulinemia. ,
' : Ann, N.Y. Acad. Sc. 64: 882-928, 1957. "x- '' ; :
28. Medawar, P. B, : Reactions t o .homologous tissue antigens in ■: - / ax , relation-to;hypersensitivity. In Lawrence, H., 8., E d , : :x;x:'
Cellular and Humoral. Aspects of the Hypersensitive : 'r '.' ' '/'■ ^States.vNew1 York,. H peber-H arper, 1959, p. 516. % ' .
V 2dv^&Stehisonv- : fP W iW trah sfe r of transplantation immunity.Proc. Roy. Soc. London, se r . B., 142: 72-87, 1954.
30. Billingham,: R .;E .,-B rent, .L,, -and Medawar,, P . B. : Quantitativestudies on tissue transplantation immunity. EL- The ■ ■ 'origin, strength and duration Of actively and adoptively acquired immunity. P roc. Roy. Soc.-London, se r. B., '■ .
- 143: 58-80,- 1954.
Page 64
57
' 31. Algire, G. H., Weaver, J. M., and Prehn, R. TV: Growth of ..cells in vivo in diffusion cham bers. I. Survival of hom.0- . grafts in immunized mice. J . Nat. Cancer Inst. 15: 493-507
/ 1954. ‘ ~ : v- v
.32.", Prehn, R. T., Weaver,- J . M., and A lgire, G. H. : The diffusion chamber technique applied to a study of the nature of homograft resistance. J . Nat. Cancer Inst. 15: 509-517,
. 1954, . ; : ' . " ' -
33. Weaver, J . M., Algire, G. H., and Prehn, R. T. : The growth ofcells in vivo in diffusion cham bers. II. The role of cells in the destruction of homografts in mice, J . Nat. Cancer Inst. 15: 1737-1767, 1955.
34. Dillingham, R. E ., and Sparrow, E .. M. : Studies on the nature of- immunity to homologous grafted skin, with special r e fe r
ence to the use of pure epiderm al grafts. J . Exp. Biol.. 31: 16-39, ,1954. ' ' ' V: ' ' • ' V , ,
35. Trowe 11, O, A/': The culture of m ature organs in a synthetic ' ' ' ' medium. Exp. Cell. Res. 16: 118-147, 1959. -
36. Scothorne, R. J ., and Nagy, I. : In vitro studies of the interactionof lymph node and homologous tissue^ Ann. N. Y. Acad,Sc, 87: 149-155, 1960. / ; V "
37. Medawar, P. B. : Tests by tissue culture methods in the natureof immunity to grafted skin; Quart. J . Microscop. Sc. 89:
: 239-252, 1948. " ; .'- i - ' V:
38. Govaerts, A. : Cellular antibodies in kidney homotrahsplantation. ;J . Immunol. 85: 516-522, 1960. :
39. Rosenau, W,, and Moon, H. D. : Lysis of homologous celis by ; ;sensitized lymphocytes in tissue culture. J . Nat, Cancer Inst. 27: 471-483,' 1961. : : ' / - - ' \
40.-:- Algire. 6 . H-. : In vivo cell culture. Univ. Michigan Med. Bull. ; - vv 22: 517-523, .1956. ' ■ , . ; / ' ' ' . , ;
41. Algire, G. E.,% W eaver,- 3,. M., and P rehn, K. T / : Studies on tissue,homotransplantatioh in mice using diffusion chamber,
■Vx methods. - Ann. N.Y.. Acad. Sc. 64: T 0 0 9 -l# 3 , T956x ' \
Page 65
58
42o Algire, G. Ho:: Diffusion chamber techniques for studies of ce llular immunity. Ann. N.Y.: Acad.: Sc. 69: ..603-667, 1957'.
43. A lgire, G. H. : Summary of studies of transplantation of homologous tissues. Federation P roc. 16: 601-602, 1957.
44.: Algire, G. H. B orders, M. L., and Evans, V. J . : Studies ofheterografts in diffusion chambers in m ice. J . Nat.Cancer Inst. 20: 1187-1201, 1958. :
- ■ 1 , ; ' ' ■ ‘ : n • : '. 45, Rezzesi, F. D. : Eine Methode zur Zuchtung der Gewebe in vivo.
■ Archiv. fur Exper. Zellforschung 13: 258-281, 1932.■■ ■ . : " ? ? - ... • ■. ' ! ; t ? ' ' , - ,
46. Bisceglieh V. : Uber die an ti^op lastische Immunitat. H .. MitteilurigUber die Wachstumsfahigkeit der heterologen Geshwulste in erwachsenen T ieren nach Eihpflanzung in Kollodium- sackchen. Z tschr. fur K rebsforsch. 40: 141-158, 1933.
47. Goldhaber, P^.: Osteogenic induction across M illipore filte rs in: \ vivo. Science 133: 2065-2067, 1961. !, '• • , •,
48. v P etrak is , N. L.^ . Davis and Lucia, S., P. : The in vivo differ? ,entiation of human leukocytes into histiocytes, fibroblasts,
' - and fat cells in subcutaneous diffusion cham bers. Blood: ..-'.-.IT: 109-118,.: 1961 iv ; ,• /. f. ^'d.- ::y
49. P etrak is, N. L. : . In vivo cultivation of leukocytes in diffusion. chambers: Requirement of ascorbic acid for differentia
tion of mononuclear leukocytes to fibroblasts. Blood 18:v y ^ ' -: y V. ■■■■ . ‘ y.""''
50. Odone, D. T ., F o rste r, I . E ., and Epstein, W. Y._: Fibroblastformation froni peripheral white cells of patients with various connective tissue diseases. A rth ritis and Rheum atism 4: 428y 1961.
51. Epstein, W. V. and Ross, M. : Sequence of morphologic changesin rheumatoid lymph node cells grown in M illipore Cham- bers. A rthritis and Rheumatism 4: 109-110, 1961.
52. RowseU, E. V.,: The effects of dusts on peritoneal cells withindiffusion cham bers. J. Path. Bact. 80: 337-344, 1960,
53. Brooks, J . R., Sturgis, S. H., and Hill, G. J . : An evaluation ofendocrine tissue homotransplantation in the M illipore
Page 66
59
chamber: with a note on tissue adaption to the host, Aniio , . : Acad. Sc. 87: 482-500, 1980. . - .
54. Najarian, J . S., and Feldman^ J . D. :. Passive tran sfe r of tran s-plantation immunity: 1 : T ritiated lymphoid:cells. H. Lymphoid'cells in M illipore chambers. J . Exp.-Med.115: ■ 1083-1098, 1962. ' ■ .
55. Holttb, M. : Morphological changes in lymphocytes cultivated indiffusion chambers during the prim ary response to a p ro tein antigen. In Holub, M .,'and Jaroskova, L., Eds. : Mechanisms of Antibody Formation, Prague Symposium. Prague, Publishing House of the Czechoslovak Academy of Sciences, 1960, pp. 30-33. • - ■
56. Shelton, E ., and Rice, M. : Growth of norm al peritoneal cells in. diffusion chambers: a study in cell modulation. Amer. J .
' , : / % / Anat.::105: 281-341, 1959, ; - ,
57. Shelton, E. : Differences in the potentialities of norm al and m alig-v nant lymphocytes grown in diffusion cham bers. Pathologies;
; Biologie ^ 542-544,; 19i61.: v ■; > , ; ,
58. Shelton, E., and Rice M. : Studies on mouse lymphomas, n .Behavior of three lymphomas in diffusion chambers in relation to their invasive capacity in the host. J . Nat.
: : Cancer Inst. 21: 137-161, 1958, ' . . : ' •
59. E arle , W. R._ : Production of malignancy in v itro . IV. The mousefibroblast cultures and changes seen- in the living c e lls .
• J . Nat. Cancer Inst. 4: 165-212, ■1943.
60. Jago, M. r A simple method for the separation of living lymphocytes from norm al human blood. B rit. J , Haemat. 2:. 439,;
61. Amos,; D. B . : Possible relationships between the cytotoxic effectsof isoantibody and host ceil function. Ann. N.Y. Acad. Sc.
' ; : : ; : 87: 273-292, I960: V ■ . ;v \ ; , . r
62. Amos, D. B., and Wakefield, J. D„ : Growth of mouse ascitestumor cells in diffusion cham bers. I. v Studies of growth ra te of cells and the ra te of entry of isoantibody. J . Nat. Cancer Inst. 21: 657-670, 1958.
Page 67
6 0
§;3.y-: Amos, D. B,, and Wakefield, J. D. : GroMh of mouse ascitestum or cells in diffusion cham bers» n . Lysis and. growth inhibition by diffusible isoantibody. J . Nat. Cancer Inst.
; ' :v- : 22: 1077-1092, 1959.' . '
64. Gabourel, J . B. : Cell culture in vtyo. II. Behavior of L -fibro-blasts in diffusion chambers in resis tan t hosts. Cancer
. Research 21: 506-5.09,; 1961. .
65. Algire, G. H..: Growth inhibition of homografts of a plasm a cell : j ■ : neoplasm in cell-im penetrable diffusion chambers in
hyperimmunized m ice. '■ •’j . ; Nat. Cancer Inst. 23: 435 -440, -; - : : : ,66.:. Karush, F., and Risen, H. N. : A'theory, of delayed hypersensi-
tivity. The main features of. this phenornenon are expli- / v'; - " ° ’V cable in te rm s of high-affinity humoral antibody. Science/: V '■ ' ,136 : 1032-1039, 1962.., ' . ' ' ■
67.: Lawrence, H. S. : The transfer of hypersensitiyity of the delayedtype in man. In Lawrence, ELS.., Ed. : Cellular and Humoral Aspects of the Hypersensitive S tates. New York,
: H oeber-Harper, 1959,' pp.' 279-318. ' ' ■ ' ; '
68v Fishman, M. : Antibody formation in vitro. J . Exp. Med. 114:• 837-856, 1961., ; ' : ■
69. Raffel, S. : Mechanisms of acquired immunity. Serologic m anifestations of antibody activity. Lysis and complement.In Immunity. New York, Appleton-Century Crofts, 1961, pp. 162-172. . ' - ; ■ : . ■
70. M iller, JT. M .,: Vaughan,:;;J. .H:., "aid :Favour,. C. B. :• The role ofi,.. ' complement in.the- lysis of leukocytes by tuberculoproteirio
' P roc. Soc. Exp. Biol, and Med.. 71: ■ 592-597,, 1949. ,.
71.. Schar, V. B., and M eier, R . : Die Bedeutung des Komplementes • ■ fur den Charakter der Antigen-Antikorper -Reaktion an
der isolierten T ierzelle (Mikrofilmanalyze). Schweiz. Z tschr. f. Allg. Pathol. un Bakteriol. 14: 618-622, 1951.
Page 68
6 1
72. Pulvertaft, R;. J. V., Doniach, D., Roitt, L M., and Hudson, R. V.: Cytotoxic effects of Hashimoto serum on human thyroid cells in tissue culture. Prelim inary Communication The Lancet, pp. 214-216, 1959 (August 29).
7'3. Gorer, P . A., and O’Gorman, P.;: The cytotoxic activity of iso- antibodies in mice. Transpl. Bull. 3_: 142-143, 1956.
Page 69
Figure 1. Rheumatoid synovialis cultured alone. (Note the large number of undamaged nuclei.)
Legend:Figures 1, 2, and 3: Hematoxylin and eosin stained paraffin sections; 300X.
Page 70
63
Figure 2.
Figure 3.
. *
; * ..V " ' * ,
,£B* *e™ . w * •*
* y . i .
' M
e •
# I> t '
i f
fti M* •
Rheumatoid synovialis cultured with nonrheumatoid lymphocytes. (Note the large number of undamaged nuclei.)
Rheumatoid synovialis cultured with rheumatoid lymphocytes. (Note the absence of undamaged nuclei.)
Page 71
64
Figure 4. Nonrheumatoid synovialis cultured alone. (Note numerous undamaged nuclei.)
Legend:Figures 4, 5, and 6: Hematoxylin and eosin stainedparaffin sections; 300X.
Page 72
Figure 5. Nonrheumatoid synovialis cultured with non-rheumatoid lymphocytes. (Note the undamaged nuclei.)
Figure 6. Nonrheumatoid synovialis cultured with rheumatoidlym phocytes. (Note the scarcity of undamagednuclei.)
Page 73
66
Figure 7. Rheumatoid lymphocytes cultured alone. (Note the numerous undamaged nuclei.)
Legend:Figures 7, 8, and 9: Hematoxylin and eosin stainedM illipore filter surfaces; 300X.
Page 74
Figure 8. Rheumatoid lymphocytes cultured near rheumatoidsynovialis. (Note reduction in number of undamaged nuclei.)
Figure 9. Rheumatoid lymphocytes cultured near nonrheuma-toid synovialis. (Note scarcity of undamaged nuclei.)
Page 75
68
Figure 10. Nonrheumatoid lymphocytes cultured alone. (Note large numbers of undamaged nuclei.)
Legend:Figures 10, 11, and 12: Hematoxylin and eosin stainedM illipore filter surfaces; 300X.
Page 76
69
w e- • ' _ w S H B B M H I
V \* . • > : '
* A «
I <* -„•*•*. %• » 1 A *
7 i *✓ • A,
<♦
>E*; -
. i n ; ? ♦‘ •srt ••
Figure 11. Nonrheumatoid lymphocytes cultured near rheum atoid synovialis. (Note large numbers of undamaged nuclei.)
Figure 12. Nonrheumatoid lymphocytes cultured near nonrheumatoid synovialis. (Note the large numbers of undamaged nuclei.)