Sci Parasitol 17(1-2):26-35, June 2016 ISSN 1582-1366 ORIGINAL RESEARCH ARTICLE 26 Nematodes and cestodes from the desert hedgehog, Paraechinus aethiopicus (Ehrenberg) in Central Saudi Arabia, revealed by SEM and microscopy, with notes on histopathology Omar M. Amin 1 , Richard A. Heckmann 2 1 – Institute of Parasitic Diseases, 11445 E. Via Linda, # 2-419, Scottsdale, AZ 85259, USA. 2 – Department of Biology, 401 WIDB, Brigham Young University, Provo, UT 84602, USA. Correspondence: Tel. 480-767-2522, Fax 480-767-5855, E-mail [email protected]Abstract. Specimens of two species of nematodes and one species of tapeworms were collected from the desert hedgehog, Paraechinus aethiopicus (Ehrenberg), in Central Saudi Arabia between summer of 2014 and spring of 2015. Females of Rictularia plagiostoma (Wedl, 1861) (Rictularidae: Nematoda) (new host record) and females of a species of Thubunaea Seurat, 1914 (Physalopteridae: Nematoda) (new record in Saudi Arabia), and of Mathevotaenia erinacei Meggitt, 1920 (Anoplocephalidae: Cestoda) (new geographical record) were collected from the intestinal tract of hedgehogs. The present report includes comparative descriptions of the Saudi material and is augmented by SEM scans. Coinfections with acanthocephalan parasites are briefly noted. Histopathological sections of the Thubunaea-host interface are also provided and a migratory tissue invading phase of larvae is demonstrated. The histopathology shows eggs hatching in the host and encysted larvae migrating into the muscle layers of the intestine causing limited damage to the host. The inflammatory response of the parasites especially the larval stages of the nematodes is characteristic of granular parasitic enteritis. Keywords: Rictularia plagiostoma; Thubunaea sp.; Mathevotaenia erinacei; Paraechinus aethiopicus; Saudi Arabia. Received 21.01.2016. Accepted 18.04.2016. Introduction Surveys of mammals and their parasitic fauna in Central Saudi Arabia (CSA) were conducted on regular monthly intervals in 2014 and 2015. Both host and parasite faunas of CSA and the Kingdom are poorly documented. The host, the desert hedgehog, Paraechinus aethiopicus (Ehrenberg, 1832) is one of the smallest of hedgehogs in the world. It is common in Saudi Arabia and other Middle Eastern and North African countries (Wikipedia, 2015). The present report documents new records and updated descriptions of nematode and tapeworm species from this host in Saudi Arabia, a land virgin for new discoveries.
Transcript
Sci Parasitol 17(1-2):26-35, June 2016 ISSN 1582-1366
ORIGINAL RESEARCH ARTICLE
26
Nematodes and cestodes from the desert hedgehog, Paraechinus
aethiopicus (Ehrenberg) in Central Saudi Arabia, revealed by SEM and
microscopy, with notes on histopathology
Omar M. Amin1, Richard A. Heckmann2
1 – Institute of Parasitic Diseases, 11445 E. Via Linda, # 2-419, Scottsdale, AZ 85259, USA.
2 – Department of Biology, 401 WIDB, Brigham Young University, Provo, UT 84602, USA.
Abstract. Specimens of two species of nematodes and one species of tapeworms were collected from the
desert hedgehog, Paraechinus aethiopicus (Ehrenberg), in Central Saudi Arabia between summer of 2014 and
spring of 2015. Females of Rictularia plagiostoma (Wedl, 1861) (Rictularidae: Nematoda) (new host record)
and females of a species of Thubunaea Seurat, 1914 (Physalopteridae: Nematoda) (new record in Saudi
Arabia), and of Mathevotaenia erinacei Meggitt, 1920 (Anoplocephalidae: Cestoda) (new geographical record)
were collected from the intestinal tract of hedgehogs. The present report includes comparative descriptions of
the Saudi material and is augmented by SEM scans. Coinfections with acanthocephalan parasites are briefly
noted. Histopathological sections of the Thubunaea-host interface are also provided and a migratory tissue
invading phase of larvae is demonstrated. The histopathology shows eggs hatching in the host and encysted
larvae migrating into the muscle layers of the intestine causing limited damage to the host. The inflammatory
response of the parasites especially the larval stages of the nematodes is characteristic of granular parasitic
enteritis.
Keywords: Rictularia plagiostoma; Thubunaea sp.; Mathevotaenia erinacei; Paraechinus aethiopicus; Saudi
Arabia.
Received 21.01.2016. Accepted 18.04.2016.
Introduction Surveys of mammals and their parasitic fauna in Central Saudi Arabia (CSA) were conducted on regular monthly intervals in 2014 and 2015. Both host and parasite faunas of CSA and the Kingdom are poorly documented. The host, the desert hedgehog, Paraechinus aethiopicus
(Ehrenberg, 1832) is one of the smallest of hedgehogs in the world. It is common in Saudi Arabia and other Middle Eastern and North African countries (Wikipedia, 2015). The present report documents new records and updated descriptions of nematode and tapeworm species from this host in Saudi Arabia, a land virgin for new discoveries.
Sci Parasitol 17(1-2):26-35, June 2016 ISSN 1582-1366
ORIGINAL RESEARCH ARTICLE
27
Material and methods Desert Hedgehogs, P. aethiopicus, were collected by live traps monthly between August, 2014 and March, 2015 mostly in Central Saudi Arabia at Unaizah (26°5′2.4″N 43°59′38.4″E), Al Qassim Province. They were overdosed with anesthetic agent before dissection. Parasites were collected from the incised digestive tract under a dissecting scope. Acanthocephalans, tapeworms, and nematodes were placed in 70% ethanol to process for microscopy and SEM, and in neutral buffered formalin for histopathological sectioning. The nematodes were found in the stomach but the acanthocephalans and tapeworms were located in the intestine. The total number of hedgehogs examined was 89 animals (50 males and 39 females) weighing 354.7 ± 92.3 gm. Of those animals, parasites were collected from 30 individuals; a prevalence of 33.71% in the population studied. Infected hedgehogs (18 males and 12 females) weighed between 166 and 495 gm (303.8 ± 80.3). For microscopical examination, specimens were placed in water overnight or until fully extended then fixed in 70% ethanol. Worms were punctured with a fine needle and subsequently stained in Mayer’s acid carmine, destained in 4% hydrochloric acid in 70% ethanol, dehydrated in ascending concentrations of ethanol (24 hr each), and cleared in 100% xylene then in 50% Canada balsam and 50% xylene (24 hr each). Whole worms were then mounted in Canada balsam. Optical microscopic images were captured using an Olympus BH2 compound light microscope (Olympus Optical Co., Tokyo, Japan) hooked to an AmScope camera MU900 (United Scope, Irvine, California). Measurements are in micrometers, unless otherwise noted; the range is followed by the mean values between parentheses. Width measurements represent maximum width. Trunk length does not include proboscis, neck, or bursa. For SEM studies, specimens previously fixed in 70% ethanol were placed in critical-point drying baskets and dehydrated using ethanol series of 95% and 3 N 100% for at least 10 min
per soak followed by critical point drying (Lee, 1992). Samples were mounted on SEM sample mounts, gold coated and observed with a scanning electron microscope (XL30 ESEMFEG; FEI, Hillsboro, Oregon). Digital images of the structures were obtained using digital imaging software attached to a computer. For histological sections, infected host tissue was fixed in 10% buffered formalin and after dehydration and blocking, the specimens were processed using standard methods (Bancroft and Gamble, 2001; Kiernan, 2002). The paraffin blocked tissue was sectioned at 4-6 microns, placed on glass slides and stained with hematoxylin and eosin (HE). Additional sections were stained with Mallory’s trichrome to emphasize pathological responses to the parasite (Galigher and Kozloff, 1971). The prepared glass slides were viewed with an LSM laser (Carl Zeiss, Thornwood, New York) equipped compound light microscope with representative pictures taken at varying magnifications with a digital camera. HE is a standard stain for tissue whereas Mallory’s trichrome helps differentiate granular tissue typical of parasite invasion. Voucher specimens were deposited in the University of Nebraska’s State Museum’s Harold W. Manter Laboratory (HWML) collection in Lincoln, Nebraska, USA. Results Two species of nematodes and one species of cestode were collected from the stomach and the intestine of the desert hedgehog in Central Saudi Arabia between August 2014 and March, 2015. A new species of an acanthocephalan of the genus Moniliformis Travassos, 1915 was also collected from hedgehog intestines but reported elsewhere (Amin et al., 2016). The lighter weight of infected hedgehogs (303.8 ± 80.3 gm) compared to that of examined animals (354.7 ± 92.3 gm) corresponded with the possible impact of infection on host overall health and weight. Rictularia plagiostoma (Wedl, 1861) (Willemoes-Suhm, 1873) of Parona, 1898 (Rictularidae: Nematoda)
Sci Parasitol 17(1-2):26-35, June 2016 ISSN 1582-1366
ORIGINAL RESEARCH ARTICLE
28
(syn. Rictularia fallax Jägerskiöld, 1909-Yorke & Maplestone, 1926; Pterygodermatitis plagiostoma (Wedl, 1861); Ophiostoma spinosum Will.-Suhm, 1869) Twelve gravid females were collected from the stomach of 5 hedgehogs. No males were found. Most reports of this and related species of nematodes refer only to females. The description of our worms is comparable to that of Hall (1916) who also reported no males. Description Oral opening dorsal and transverse with single esophageal tooth (figure 1, arrow). Buccal capsule well developed. Body surface covered with thin circular nearly overlapping plates (figure 4). Two rows of prominent cuticular combs along the ventral side anteriorly (figure 2) followed by posterior spines (figure 3). Vulva anterior. Eggs usually contain developing embryo. Females 9.12-15.00 (11.79) mm long X 0.20-0.37 (0.26) mm wide, Esophagus 2.53-3.62 (2.98) mm long. Nerve ring 200-272 (242) from anterior end. Anus 208 from posterior end (figure 4). Vulva 3.00-3.25 (3.13) mm from anterior. Eggs 40-50 (46) long X 27-35 (30) wide. Cuticular combs in 48-55 (51) circles with the highest combs being only 87 deep, and 112 long (figure 2). Same height of combs near vulva. Specimens. Voucher specimens in the HWML coll. No. 101912 (2 slides). Remarks. Measurements of our specimens from P. aethiopicus in Saudi Arabia are comparable to those in the original account of Hall (1916) from the Mentawai squirrel, Sciurus (=Callosciurus) melanogaster Thomas, in Mentawai, Sumatra. His specimens were 20.00 mm long X 0.75 mm wide, esophagus was 2.96 mm long, nerve ring was 240 from anterior end, anus was 200 from posterior end, vulva was 3.44 mm from anterior end, eggs were almost identical, 44 X 27, cuticular combs were in 50 circles with highest combs being 110 deep and 110 long.
To the best of our knowledge, Rictularia plagiostoma is reported from the desert hedgehog, P. aethiopicus, for the first time. It has been reported in other species of hedgehogs including the long-eared hedgehog, Hemiechinus auritus Gmelin from Egypt (Ammar, 2015; Ashour, 1980; Mikhall, 1967; Yamaguti, 1961; Fahmy et al., 1971). This species of nematode has many subspecies found elsewhere in the Steppe zone from East Ukraine to Mongolia, Gobi Desert (Kazakhstan) in the north and from Libya to West Pakistan in the south (Syria, Lebanon, Palestine, Israel, Jordan, Iran, Iraq, Afghanistan, also Cyprus and Turkey), Kirghizistan, Turkmenistan, Tadjikistan, Southern China, Xinjiang, and probably Transcaucasia (Armenia, Republic of Georgia, Azerbaijan) and SW European Russia (Hutterer, 2005). Other species of Rictularia Froelich, 1802 such as Rictularia shaldybini Skrjabin, Sobolev, Ivaschkin, 1967 in Mongolia have been reported from H. auritus. The presence of hedgehogs in the reported habitat in Saudi Arabia and their role in maintaining zoonotic infections of nematode and cestode parasites are documented here for the first time. Mammalian hosts are infected with larval Rictularia nematodes upon the ingestion of infected arthropod intermediate hosts. For instance, the white-footed mouse Peromyscus leucopus noveboracensis (Fischer) in Ohio become infected with Rictularia coloradensis Hall, 1916 by feeding on camel crickets Ceuthophilus g. gracilipes (Haldeman, 1850) and wood cockroachs Parcoblatta mrginica (Brunner von Wattenwyl, 1865) naturally infected with the larvae (Oswald, 1956). Various arthropods constituted major components of the diet of P. aethiopicus in Algeria (Khaldi et al., 2013). Thubunaea Seurat, 1914 (Thubunaeinae: Physalopteridae: Nematoda) Thubunaea sp. Forty female specimens of an unidentified species of the nematode genus Thubunaea Seurat, 1914 were collected from the stomach of 12 desert hedgehogs. About half of these specimens were used for SEM. No males were found.
Sci Parasitol 17(1-2):26-35, June 2016 ISSN 1582-1366
ORIGINAL RESEARCH ARTICLE
29
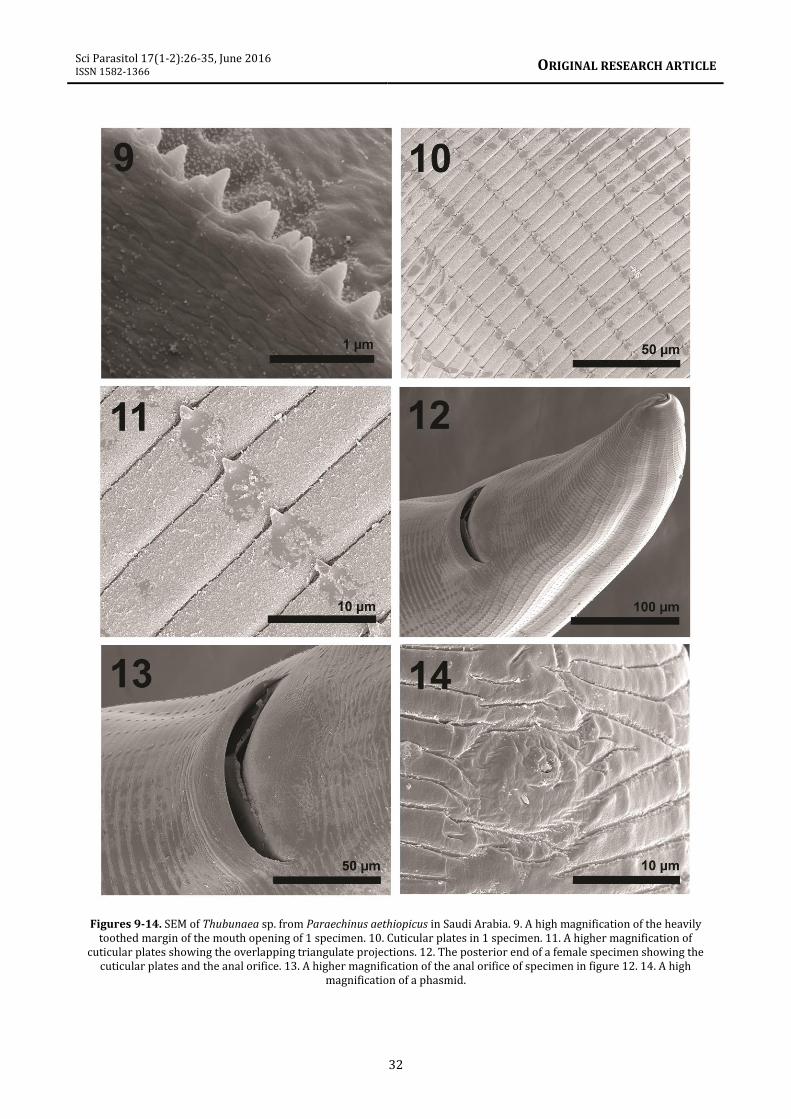
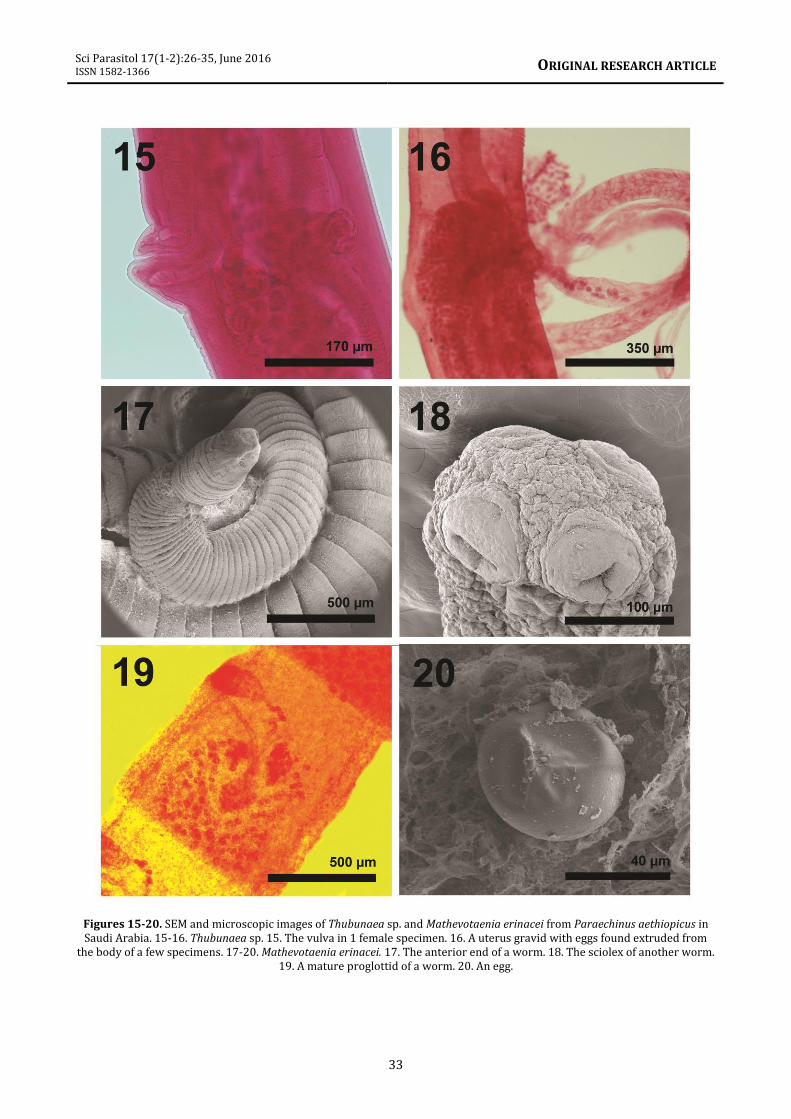
Description Medium size worms with cuticle covered with transverse striations of thin circular nearly overlapping plates, except on the oral surface, the vulva, and the anal orifice (figures 5, 6, 10-16). Cuticular plates broader posteriorly, with triangulate dentacles at regular intervals in longitudinal rows (figures 10, 11). Oral end broad, dome-shaped with symmetrical oral orifice (figures 5, 6). Deeply set obovate mouth (figures 6, 8) with 2 simple lateral lips armed with externo-lateral, interno-lateral, and submedian teeth, and heavily toothed outer margin (figures 8, 9). Oral orifice with 4 prominent submedian amphids (figures 5, 6, arrows). Gravid females 21.70-31.25 (25.90) mm long X 0.22-0.57 (0.38) mm wide. Pharynx 312-342 (322) long. Entire esophagus 6.25-7.55 (6. 68) mm long, muscular and glandular portions not readily separated. Nerve ring 230-310 (260) from anterior end. Excretory duct directed anteriorly, opening 340-400 (375) from anterior end, just behind nerve ring. Reproductive system with 2 ovaries, looping uteri (figure 16) and a vulva slightly more posterior to middle of trunk. Vulva with elevated lips (figure 15), post-equatorial, 13.00-16.25 (14.46) mm from anterior end. Anus, a lateral slit, opening 200-260 (240) from blunt conical posterior end; no caudal spike (figures 12, 13). Paired lateral phasmids just anterior to anal opening (figure 14). Ovoid thick-shelled eggs, 37-50 (44) long X 25-30 (27) wide, with larvae. Specimens. Voucher specimens in the HWML coll. No. 101913 (4 slides). Remarks. These nematodes were placed in Thubunaea Seurat, 1914 based primarily on resemblances in the morphology of anterior end, symmetrical oral orifice, and cuticular striations, among other features. They showed some similarities to T. pudica Seurat, 1914 but varied in such features as the length of the vestibule and esophagus which corresponded with worm length and the position of the vulva. Thubunaea pudica was previously reported from reptiles in Morocco (Chabaud and Golvan, 1957) and Egypt (Moravec et al., 1987; Myers et al., 1962). The apical view of the Moroccan specimens reported by Chabaud and Golvan
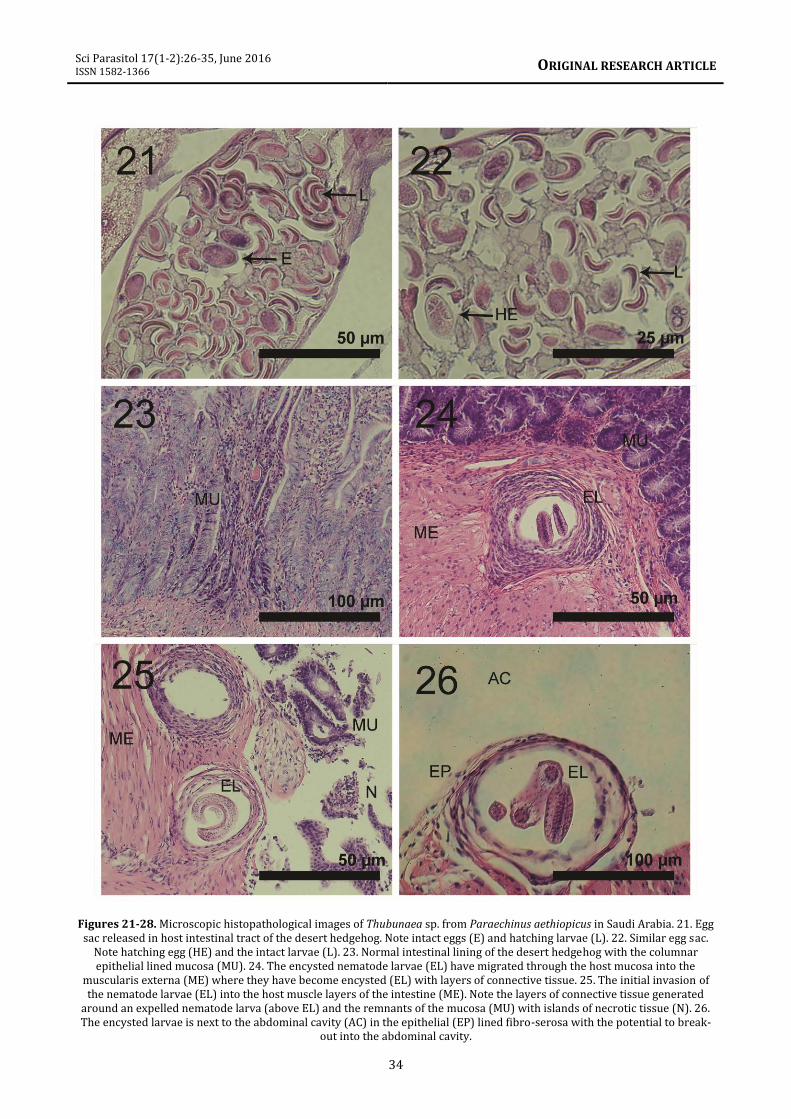
(1957; figure 6) is identical to that in our material. The Egyptian specimens reported by Moravec et al. (1987) were considerably shorter than ours. Insects such as cockroaches appear to be the reservoirs of larval forms of species of Thubunaea (see Singh et al., 2011). Feeding of hedgehogs on insects infected with larval forms or eggs of Thubunacaea sp. will initiate an intestinal infection which appears to be invasive to gut and muscle tissues. Histopathology Histopathological findings are presented in figures 21 to 26 which appear to be a type of necrotic enteritis. The main pathology is caused by the larval stages. Figures 21 and 22 demonstrate eggs and hatching larvae from the female egg sac of the nematode. Figure 22 shows the development from egg to larva (HE). The whole mount with an extended uterus is depicted in figure 16. The hatching eggs (figure 22) develop into 1st stage larvae that invade the host mucosa (figure 23) and migrate into the muscle layers (figure 24). In the host muscularis externa, the larvae continue to develop displacing the surrounding smooth muscle fibers. The host responds by enclosing the larvae with connective tissue isolating the nematode from the host tissue (figures 24, 25). Numerous layers of collagenous fibers are generated around the larvae (figure 24) which isolated the parasite from host tissue. There is a limited blood loss and necrotic tissue in the invaded host muscularis externa. The nematode larvae continue to move to the outer layers of the host intestine (figure 25) up to the edge of the outer fibro-serosa. From here the larvae can escape into the host abdominal cavity (figure 26). From the initial nematode larvae (figure 22) to the outer fibro-serosa, the worm has gone through several cell divisions depicted by the numerous nuclei in the latter stages (figure 26). There is limited damage to the host intestine compared to the other helminths interfacing in the same host intestine. Figure 25 displays necrotic epithelial tissue due to the initial invasion. The action of this nematode on host tissue is similar to other round worms such as Trichinella spiralis (Owen, 1835) in pigs and bears which also
Sci Parasitol 17(1-2):26-35, June 2016 ISSN 1582-1366
ORIGINAL RESEARCH ARTICLE
30
develop a fibrous capsule around the larvae (Faust et al., 1975). Boggs et al. (1990) noted the presence of physalopterid nematode larvae in the pectoral musculature of bobwhite quail, Colinus virginianus (Linn.). These larvae were unencapsulated and were associated with necrosis and minor granulomatous response. This situation is similar to what we observed except that the larvae of Thubunaea become encysted. The histopathology is typical of most parasitic invasions with prominent granular cells within the host necrotic intestinal tissue. Collagenous fibers surround the encysted larval stages of the nematode. Mathevotaenia erinacei (Meggitt, 1820) Spassky, 1951 (Anoplocephalidae: Cestoda) (syn: Mathevotaenia amphisbeteta (Meggit, 1924); M. erinacei rodentium Joyeux, 1937) About 10 worms were recovered from 4 hedgehog individuals. Description Worms of moderate size (figure 17) with somewhat rectangular scolex (260 long by 300 wide), lacking rostellum, with 4 relatively large, oval, muscular suckers (156 in diameter) not sunken at base of scolex and without outgrowths (figure 18). Excretory stems in 2 pairs. Testes numerous in posterior part of proglottid, near rounded (62 long by 52 wide). Cirrus sac elongated (416 long by 177 wide); atrium irregularly alternating on right or left anterior margins of proglottids. Female genital ducts pass ventral to excretory stem and with no sphincter or sucker-like muscular organ (figure 19). Ovary bilobed and median (330 by 127); vitellaria behind ovary. Eggs round, about 40 in diameter (figure 20). Specimens. Voucher specimens in the HWML Coll. No. 101914 (2 slides). Remarks. Mathevotaenia erinacei has been previously reported from P. aethiopicus in other Middle Eastern countries such as Algeria (Khaldi et al., 2012).
It was also found in other species of hedgehogs such as H. auritus in Iraq (Al-Zihiry, 2009). Our record from Saudi Arabia is new. Our specimens were similar but somewhat variable from those reported from other hosts elsewhere. For instance, specimens from the hedgehog H. auritus in Iraq had relatively larger scolex (530-610 long by 440-510 wide) and suckers 190-270 in diameter, but smaller internal structures including testes (37-39 by 24-31), cirrus sac (170-200 by 40-70), and ovary (100-140 by 190-220) (Al-Zihiry, 2009). Mathevotaenia erinacei has an indirect life cycle utilizing hedgehogs and related mammals as definitive hosts and insects as intermediate hosts. This has been demonstrated in infections of the North African hedgehog Atelerex algirus (Lereboullet, 1842) in Algeria (Khaldi et al., 2012) but interestingly not in P. aethiopicus (our host in Saudi Arabia) in the same locations. Acknowledgments We are grateful to Dr. Charles Bursey, Pennsylvania State University at Sharon for his initial ideas regarding the generic diagnosis of the nematode parasites. This work was supported by funding from the King Saud University, Vice Deanship of Scientific Research (Research group project No. RGP-VPP-020), by the Department of Microbiology and Molecular Biology, Brigham Young University (BYU), Provo, Utah, and by an Institutional Grant from the Parasitology Center, Inc. (PCI), Scottsdale, Arizona. Thanks to the Histology Laboratory, Utah Valley Regional Medical Center, Mike Downey Director, for the histopathology slide preparation. We are also grateful to Michael Standing and Stephen Hunsaker, BYU Electron Optics Center and Bean Museum, respectively, for their professional help with electron optics equipment and plate preparation and editing. Dr. Osama Mohammed and Dr. Abdulaziz Alagaili, KSU Mammals Research, Department of Zoology, College of Science, King Saud University, Riyadh, Saudi Arabia were instrumental in making resource material available for this study.
Sci Parasitol 17(1-2):26-35, June 2016 ISSN 1582-1366
ORIGINAL RESEARCH ARTICLE
31
Figures 1-8. SEM and microscopic images of Rictularia plagiostoma and Thubunaea sp. from Paraechinus aethiopicus in Saudi Arabia. 1-4. Rictularia plagiostoma. 1. The anterior end of a female specimen showing the single esophageal tooth (arrow). 2.
The anterior portion of a female specimen showing the cuticular combs on the inner margin of the specimen. 3. Posterior cuticular spines on the trunk of another specimen. 4. The posterior end of a specimen showing that anal orifice and cuticular plates. 5-8. Thubunaea sp. 5. The anterior end showing 2 amphids (arrows). Note the absence of cuticular plates at the apical end. 6. A second perspective of the anterior end of another worm showing 1 amphid (arrow) and the mouth (apical end). 7. A
high magnification of an amphid. 8. A high magnification of a deeply set mouth opening.
Sci Parasitol 17(1-2):26-35, June 2016 ISSN 1582-1366
ORIGINAL RESEARCH ARTICLE
32
Figures 9-14. SEM of Thubunaea sp. from Paraechinus aethiopicus in Saudi Arabia. 9. A high magnification of the heavily toothed margin of the mouth opening of 1 specimen. 10. Cuticular plates in 1 specimen. 11. A higher magnification of
cuticular plates showing the overlapping triangulate projections. 12. The posterior end of a female specimen showing the cuticular plates and the anal orifice. 13. A higher magnification of the anal orifice of specimen in figure 12. 14. A high
magnification of a phasmid.
Sci Parasitol 17(1-2):26-35, June 2016 ISSN 1582-1366
ORIGINAL RESEARCH ARTICLE
33
Figures 15-20. SEM and microscopic images of Thubunaea sp. and Mathevotaenia erinacei from Paraechinus aethiopicus in Saudi Arabia. 15-16. Thubunaea sp. 15. The vulva in 1 female specimen. 16. A uterus gravid with eggs found extruded from
the body of a few specimens. 17-20. Mathevotaenia erinacei. 17. The anterior end of a worm. 18. The sciolex of another worm. 19. A mature proglottid of a worm. 20. An egg.
Sci Parasitol 17(1-2):26-35, June 2016 ISSN 1582-1366
ORIGINAL RESEARCH ARTICLE
34
Figures 21-28. Microscopic histopathological images of Thubunaea sp. from Paraechinus aethiopicus in Saudi Arabia. 21. Egg sac released in host intestinal tract of the desert hedgehog. Note intact eggs (E) and hatching larvae (L). 22. Similar egg sac.
Note hatching egg (HE) and the intact larvae (L). 23. Normal intestinal lining of the desert hedgehog with the columnar epithelial lined mucosa (MU). 24. The encysted nematode larvae (EL) have migrated through the host mucosa into the
muscularis externa (ME) where they have become encysted (EL) with layers of connective tissue. 25. The initial invasion of the nematode larvae (EL) into the host muscle layers of the intestine (ME). Note the layers of connective tissue generated
around an expelled nematode larva (above EL) and the remnants of the mucosa (MU) with islands of necrotic tissue (N). 26. The encysted larvae is next to the abdominal cavity (AC) in the epithelial (EP) lined fibro-serosa with the potential to break-
out into the abdominal cavity.
Sci Parasitol 17(1-2):26-35, June 2016 ISSN 1582-1366
ORIGINAL RESEARCH ARTICLE
35
References Al-Zihiry K.J.K. 2009. First record of cestode
Mathevotaenia erinacei Meggitt, 1920 from hedgehog Hemiechinus auritus in Basrah Province, Iraq. J. Thi-Qar Sci. 1:20-26.
Amin O.M., Heckmann R.A., Mohammed O., Evans R.P. 2016. Morphological and molecular descriptions of Moniliformis saudi n. sp. (Acanthocephala: Moniliformidae) from the desert hedgehog, Paraechinus aethiopicus (Ehrenberg) in Saudi Arabia, with a key to species and notes on histopathology. Folia (in press).
Ammar K.N.A. 2015. Spirurid infection Rictularia affinis Jaegerskiold, 1909 (Spirurida: Rictularidae) from Egyptian fox Vulpus niloticus E. Geoffroy Saint Hilaire, 1803 (Mammalia: Canida). Danish J. Med. Biol. Sci. 15-28.
Ashour A.A. 1980. Ultrastructural and other studies on the intestinal nematodes of small mammals from Egypt. Thesis (Zool.), Faculty Sc., Ain Shams Univ., Cairo, Egypt.
Bancroft J.D., Gamble M. 2001. Theory and Practice of Histological Techniques. 5th Ed. Edingburough, UK, Churchill Livingston, 800 pp.
Boggs J.F., Peoples A.D., Lachmiller R.L., Elangbam C.S., Qualis C.W. Jr. 1990. Occurrence and pathology of Physalopterid larvae infections in Bobwhite Quail from Western Oklahoma. Proc. Oklahoma Acad. Sci. 70:29-31.
Chabaud A.G., Golvan Y.J. 1957. Miscellanea helminthological Maroccana XXIV, nématodes parasites de lézards de la Forèt de Nefifik. Arch. l'Institut Pasteur Maroc 7:447-469.
Fahmy M.A., Mikhall J.W., McConnell E. 1971. A survey of helminth parasites collected from Egyptian small mammals. J. Egypt. Soc. Parasitol. 1:47-58.
Faust E.C., Beaver P.C., Jung R.C. 1975. Animal agents and vectors of human diseases. Les and Fabigin Philadelphia, 479 pp.
Galigher A.E., Kozloff E.N. 1971. Essentials of Practical Microtechnique. 2nd Ed. Philadelphia, Pennsylvania, Lee and Febiger, 531 pp.
Hall M.C. 1916. Nematode parasites of mammals of the orders Rodentia, Lagomorpha, and Hyracoidae. Proc. U.S. Nat. Mus. 50:1-258.
Hutterer R. 2005. (Wilson D.E., Reeder D.M., eds.) Mammal species of the world: a taxonomic and geographic reference (3rd ed.). Baltimore, Maryland, Johns Hopkins Univ. Press, 215 pp.
Kiernan J.A. 2002. Histological and histochemical methods: theory and practice. Edinburgh, UK: Churchill Livingston, 520 pp.
Khaldi M., Torres J., Samso B., Miquel J., Biche M., Benyettou M., Borech G., Benelkadi H.A., Ribas A. 2012. Endoparasites (helminths and Coccidians) in the hedgehogs, Atelerix algirus and Paraechinus aethiopicus from Algeria. African Zool. 47:48-54.
Khaldi M., Barech G., Espadaler X. 2013. Importance of the myrmecofauna in the diet of hedgehogs (Mammalia: Erinaceidae) from Algeria. 5th Central Eur. Workshop Myrmecology, Innsbruck, Austria.
Lee R.E. 1992. Scanning Electron Microscopy and X-Ray Microanalysis. Englewood Cliffs, New Jersey, Prentice Hall, pp. 1-458.
Mikhall J.W. 1967. Studies on helminths of small mammals. M. Sc. Thesis, Faculty Sci., Cairo Univ., Egypt.
Moravec F., Baruš V., Ryšavý B. 1987. Some parasitic nematodes, excluding Heterakidae and Pharyngodonidae, from amphibians and reptiles in Egypt. Folia Parasitol. 34:255-267.
Myers B.J., Kuntz R.E., Wells W.H. 1962. Helminth parasites of reptiles, birds and mammals in Egypt. VII. Check list of nematodes collected from 1948-1955. Canad. J. Zool. 40:531-538.
Oswald V.H. 1956. Studies on the life history of Rictularia coloradensis Hall, 1916 (Nematoda: Thelaziidae), a parasite of Peromyscus leucopus noveboracensis (Fischer). Dissert. Abst. 16(8):1535 (Record no. 19560801983).
Singh H.S., Malti, Chaudhary A. 2011. Larval form of the genus Thubunaea Seurat, 1914 from the body cavity of an insect, Supella sp., at Meerut (U.P.), India. J. Appl. Nat. Sci. 3:54-57.
























![An overview of cercariae from the Egyptian inland water snailsoaji.net/articles/2017/2154-1511162691.pdf · microfilaria among some cestodes and nematodes, respectively[1,2]. Generally,](https://static.documents.pub/doc/80x56/5b52b0c47f8b9af4408de203/an-overview-of-cercariae-from-the-egyptian-inland-water-microfilaria-among-some.jpg)