See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/331985034 Nitrate-NRT1.1B-SPX4 cascade integrates nitrogen and phosphorus signalling networks in plants Article in Nature Plants · April 2019 DOI: 10.1038/s41477-019-0384-1 CITATIONS 125 READS 1,769 21 authors, including: Some of the authors of this publication are also working on these related projects: Towards Understanding of Molecular Basis of Abiotic Stresses in Plants View project Rice molecular genetics View project B. Hu Chinese Academy of Sciences 38 PUBLICATIONS 2,841 CITATIONS SEE PROFILE Zhimin Jiang South China Agricultural University 7 PUBLICATIONS 315 CITATIONS SEE PROFILE Wang Wei Chinese Academy of Sciences 35 PUBLICATIONS 1,375 CITATIONS SEE PROFILE Zhihua Zhang Institute of Genetics and Developmental Biology, CAS 17 PUBLICATIONS 794 CITATIONS SEE PROFILE All content following this page was uploaded by Chengcai Chu on 27 March 2019. The user has requested enhancement of the downloaded file.
Transcript
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/331985034
Nitrate-NRT1.1B-SPX4 cascade integrates nitrogen and phosphorus signalling
networks in plants
Article in Nature Plants · April 2019
DOI: 10.1038/s41477-019-0384-1
CITATIONS
125READS
1,769
21 authors, including:
Some of the authors of this publication are also working on these related projects:
Towards Understanding of Molecular Basis of Abiotic Stresses in Plants View project
Rice molecular genetics View project
B. Hu
Chinese Academy of Sciences
38 PUBLICATIONS 2,841 CITATIONS
SEE PROFILE
Zhimin Jiang
South China Agricultural University
7 PUBLICATIONS 315 CITATIONS
SEE PROFILE
Wang Wei
Chinese Academy of Sciences
35 PUBLICATIONS 1,375 CITATIONS
SEE PROFILE
Zhihua Zhang
Institute of Genetics and Developmental Biology, CAS
17 PUBLICATIONS 794 CITATIONS
SEE PROFILE
All content following this page was uploaded by Chengcai Chu on 27 March 2019.
The user has requested enhancement of the downloaded file.
1State Key Laboratory of Plant Genomics, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing, China. 2College of Life Sciences, University of Chinese Academy of Sciences, Beijing, China. 3Biogle Genome Editing Center, Changzhou, China. 4State Key Laboratory of Molecular Developmental Biology, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing, China. 5School of Agriculture, Henan University of Science and Technology, Luoyang, China. 6Botanical Institute, Cluster of Excellence on Plant Sciences, University of Cologne, Cologne, Germany. 7These authors contributed equally: Bin Hu, Zhimin Jiang, Wei Wang, Yahong Qiu. *e-mail: [email protected]; [email protected]
Nutrient balance is essential for maintaining optimal human health, and it can be achieved by consuming a balanced diet1–
3. For plants, maintaining a proper nutrient balance, which depends on the coordinated acquisition of various mineral nutri-ents, is also required. Nitrogen (N) and phosphorus (P) are the two most required mineral nutrients for plants and are the most widely used fertilizers in agricultural production. The proper ratio of sup-plied N and P is critical for maximizing the grain yield of crops, and has an even greater influence on plant performance than the abso-lute supply of N or P4–6. Moreover, it was reported that variations in the N:P supply ratio significantly affects their uptake. Notably, an increased N:P supply ratio greatly promotes the uptake of P in wetland graminoids7. Therefore, plants must have evolved strategies to coordinate the utilization of N and P under continuously varying nutritional environments. The molecular mechanisms underlying N–P interactions, however, remain largely elusive.
Nitrate and phosphate are not only the main available forms of N and P for plants but they also function as signalling molecules to regulate gene expression and activate nutritional responses. The signalling pathways of nitrate and phosphate have been extensively studied in the model plants Arabidopsis thaliana and rice. Nitrate is sensed by the dual-affinity nitrate transporter AtNRT1.1/CHL1/AtNPF6.3 (AT1G12110) through the phosphorylation of Thr101; this step is regulated by the concentration of nitrate and modulates the primary nitrate response in Arabidopsis8,9. In rice, OsNRT1.1B/OsNPF6.5 (NRT1.1B; LOC_Os10g40600), the functional homologue
of AtNRT1.1, has also been demonstrated to mediate nitrate signal transduction10. The NIN-like protein AtNLP7 (AT4G24020) is a central transcription factor that directly controls the expression of a subset of nitrate-responsive genes in Arabidopsis11. Interestingly, the nuclear localization of AtNLP7 is promoted by nitrate, transducing the nitrate signal to downstream responses. A recent study12 revealed that this process involves nitrate-triggered Ca2+ signalling mediated by Ca2+-sensor protein kinases. However, it remains unclear how nitrate sensing is transduced from the plasma membrane sensor to the downstream components, which greatly restricts our under-standing of the full mechanism of nitrate signalling.
In contrast to nitrate signalling, phosphate signalling is nor-mally activated by phosphate starvation, which increases the expression of phosphate starvation-induced (PSI) genes, leading to improved phosphate utilization. In this signalling pathway, the MYB-CC transcription factor PHR1 (in Arabidopsis, AT4G28610) and its related homologues (in other plants) act as central regula-tors to activate the expression of PSI genes13. It has been shown that OsPHR2 (PHR2; LOC_Os07g25710), the PHR1 homologue in rice, responds to variations in phosphate status by changing its subcel-lular localization14. PHR2 is sequestered in the cytosol under high phosphate conditions and moves to the nucleus when phosphate is limited. A SPX domain-containing protein, OsSPX4 (SPX4; LOC_Os03g61200), interacts with PHR2 in the cytosol and prevents its translocation into the nucleus. Phosphate starvation leads to SPX4 degradation via the 26S proteasome, resulting in the release
Nitrate–NRT1.1B–SPX4 cascade integrates nitrogen and phosphorus signalling networks in plantsBin Hu 1,7*, Zhimin Jiang1,7, Wei Wang1,7, Yahong Qiu1,2,7, Zhihua Zhang1,2, Yongqiang Liu1,2, Aifu Li1,2, Xiaokai Gao1, Linchuan Liu1, Yangwen Qian3, Xiahe Huang4, Feifei Yu1, Sai Kang1, Yiqin Wang1, Junpeng Xie1,2, Shouyun Cao1, Lianhe Zhang5, Yingchun Wang4, Qi Xie1, Stanislav Kopriva 6 and Chengcai Chu 1,2*
To ensure high crop yields in a sustainable manner, a comprehensive understanding of the control of nutrient acquisition is required. In particular, the signalling networks controlling the coordinated utilization of the two most highly demanded mineral nutrients, nitrogen and phosphorus, are of utmost importance. Here, we reveal a mechanism by which nitrate activates both phosphate and nitrate utilization in rice (Oryza sativa L.). We show that the nitrate sensor NRT1.1B interacts with a phosphate signalling repressor SPX4. Nitrate perception strengthens the NRT1.1B–SPX4 interaction and promotes the ubiquitination and degradation of SPX4 by recruiting NRT1.1B interacting protein 1 (NBIP1), an E3 ubiquitin ligase. This in turn allows the key tran-scription factor of phosphate signalling, PHR2, to translocate to the nucleus and initiate the transcription of phosphorus utili-zation genes. Interestingly, the central transcription factor of nitrate signalling, NLP3, is also under the control of SPX4. Thus, nitrate-triggered degradation of SPX4 activates both phosphate- and nitrate-responsive genes, implementing the coordinated utilization of nitrogen and phosphorus.
of PHR2 to activate the expression of PSI genes. Nonetheless, the components mediating SPX4 degradation remain elusive. Notably, SPX proteins directly bind inositol polyphosphates (InsPs), the concentration of which reflects phosphate availability in the cell15. Thus, InsPs modulate SPX4–PHR2 interactions and regulate phos-phate signalling15. Besides SPX4, other nuclear-localized SPX pro-teins, SPX1 and SPX2, can block PHR1 and PHR2 from binding to the promoters of the PSI genes through the formation of the SPX–PHR complex in a phosphate-dependent manner16,17. These reports collectively indicate that plants employ multiple strategies to sense phosphate availability.
Despite substantial progress in our understanding of the individ-ual pathways of nitrate and phosphate signalling, their direct cross-talk is still poorly understood. In Arabidopsis, NLA (AT1G02860) and NIGT1/HRS1 (AT1G13300) have been shown to be involved in this process18,19. Very recently, NIGT1 was further demonstrated to regulate the N starvation response as well as to incorporate P star-vation signalling to repress nitrate uptake20,21. However, the molec-ular mechanism of how nitrate and phosphate signals integrate is still far from being fully understood. In this study, we uncover the signalling pathway by which nitrate activates both phosphate and nitrate responses. Nitrate signals are perceived by the nitrate sen-sor NRT1.1B, leading to the ubiquitination and degradation of the repressor protein SPX4. Subsequently, central transcription factors of both nitrate and phosphate signalling are released, ensuring coor-dinated utilization of N and P to maintain nutrient balance in plants.
ResultsNitrate promotes coordinated utilization of N and P by activat-ing phosphate signalling. To investigate the interaction and coor-dinated utilization of N and P, we cultivated rice under varying nitrate and phosphate supply. Under low nitrate conditions, high phosphate supply did not promote plant growth and biomass gain (Fig. 1a,b), whereas under high nitrate conditions, it stimulated growth and a significant biomass increase of rice (Fig. 1a,b). These observations suggested that nitrate plays an essential role in activat-ing phosphate utilization. As expected, phosphate acquisition was significantly enhanced by high nitrate supply (Fig. 1c). To explore the molecular mechanism of this process, we examined the tran-script levels of phosphate transporter (PT) genes in rice plants cul-tured under different nitrate supply conditions. The PT genes OsPT2 (LOC_Os03g05640) and OsPT6 (LOC_Os08g45000) as well as the PSI genes OsIPS1 (AY568759) and OsIPS2 (BI796836) were upregu-lated by high nitrate supply irrespective of the phosphate concentra-tion (Supplementary Fig. 1). Interestingly, the expected induction of PSI genes by P limitation occurred only under high nitrate sup-ply and was repressed under low nitrate conditions (Supplementary Fig. 1). Furthermore, we performed short-term nitrate induction to test whether nitrate can act as a signal to directly trigger the expres-sion of phosphate-responsive genes. The expression of OsPT2, OsPT6, OsIPS1 and OsIPS2 was indeed induced by short-term nitrate treatment (Fig. 1d). Thus, nitrate can directly activate phos-phate starvation signalling, through which the increased N supply coordinately induces P uptake to achieve an optimal N–P nutrient balance and promote growth.
NRT1.1B is required for nitrate-triggered phosphate signalling. Our previous work has shown that NRT1.1B is the functional homo-logue of AtNRT1.1 in rice and acts as a transceptor to sense nitrate signals and to activate the nitrate response10. Thus, we further tested whether the nitrate-triggered phosphate signalling also depends on NRT1.1B. To this end, we examined the nitrate-induced expression of PSI genes in both wild-type and nrt1.1b mutant plants. Nitrate induction of PSI genes was nearly abolished in the nrt1.1b mutant (Fig. 2a), indicating an impairment of nitrate-triggered phosphate signalling caused by the loss-of-function of NRT1.1B. In contrast,
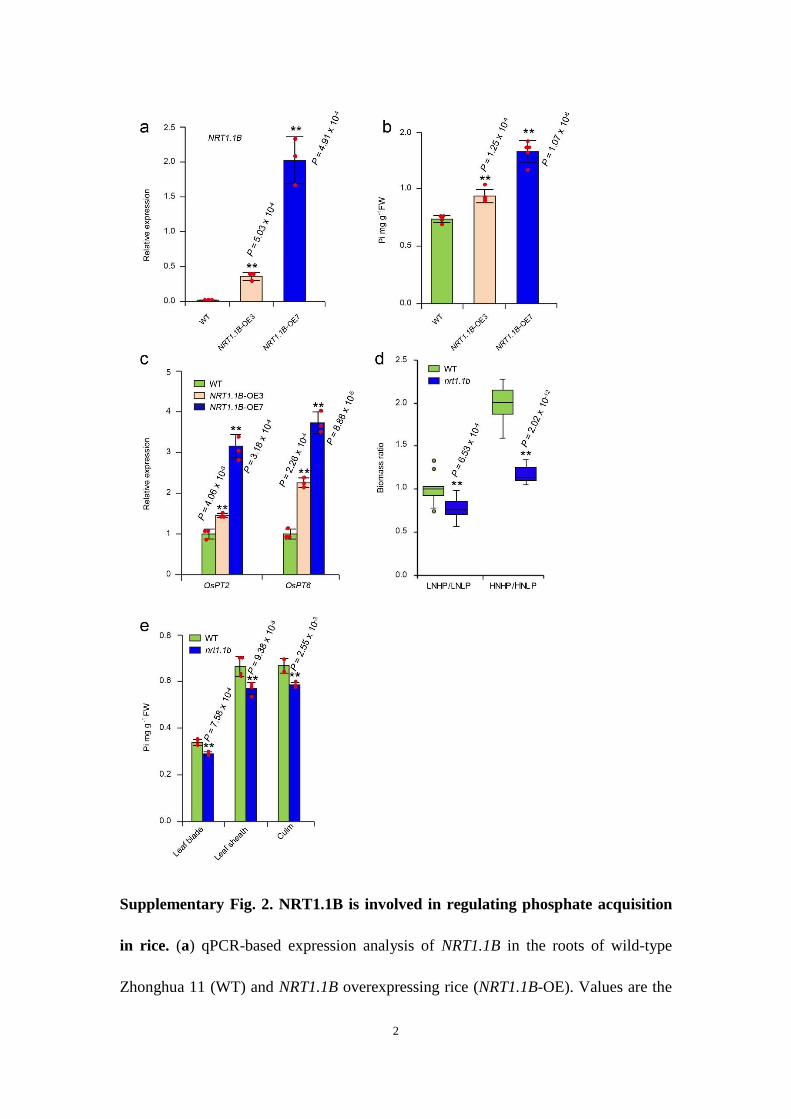
phosphate concentrations and expression of PSI genes were signifi-cantly increased in NRT1.1B-overexpressing plants (Supplementary Fig. 2a–c). Under long-term cultivation, the nrt1.1b mutant did not show significant biomass gain following high phosphate sup-ply under high nitrate conditions compared with wild-type plants (Fig. 2b,c; Supplementary Fig. 2d). Compared with the wild type, growth retardation of the nrt1.1b mutant was most significant when grown under the high-nitrate, high-phosphate condition (Fig. 2b,c), providing further evidence to support the critical role of NRT1.1B in coordinating N and P utilization. Additionally, phosphate con-centrations in leaf blade, leaf sheath and culm of nrt1.1b mutants grown in the field were consistently lower than that of wild-type plants (Supplementary Fig. 2e). Taken together, these results dem-onstrate that the nitrate-activated phosphate response is dependent on NRT1.1B.
Nitrate induces PSI gene expression through the SPX4–PHR2 module. SPX4 is a central component of the phosphate signalling pathway in rice, acting as a repressor to prevent the nuclear localiza-tion of PHR214. Considering the effect of nitrate on phosphate signal-ling, we investigated whether SPX4 is involved in this process. SPX4 transcript levels were not affected by nitrate induction (Fig. 3a), but the protein accumulation of SPX4 was decreased, as demonstrated by the fluorescence intensity of enhanced green fluorescent pro-tein (eGFP)-tagged SPX4 in rice protoplasts (Fig. 3b). The nitrate-induced degradation of SPX4 was further verified in transgenic rice expressing the SPX4–FLAG fusion protein. This degradation was repressed by MG132, a 26S proteasome inhibitor (Fig. 3c), indicat-ing that ubiquitination might be involved in nitrate-induced SPX4 degradation. Furthermore, nitrate promoted a shift in the localiza-tion of PHR2 from the cytosol to the nucleus (Fig. 3d). This cytoplas-mic–nuclear shuttling of PHR2 was inhibited by the co-expression of SPX4 (Fig. 3e). In addition, in both SPX4-overexpressing plants and phr2 mutant plants, nitrate-induced phosphate signalling was repressed compared with the wild-type plants (Supplementary Fig. 3a–d). A large number of nitrate-inducible PSI genes, including OsPT3 (LOC_Os10g30770), OsPT8 (LOC_Os10g30790), OsPT10 (LOC_Os06g21950), OsPT13 (LOC_Os04g10800), OsPAP10 (LOC_Os01g56880) and RNasePD2 (LOC_Os08g33710), were also repressed in the phr2 mutant (Supplementary Fig. 3e,f). This result further demonstrates the function of PHR2 in activating the nitrate-induced phosphate response. These results reveal that SPX4–PHR2, the central regulatory module of phosphate signalling, mediates nitrate-induced phosphate signalling.
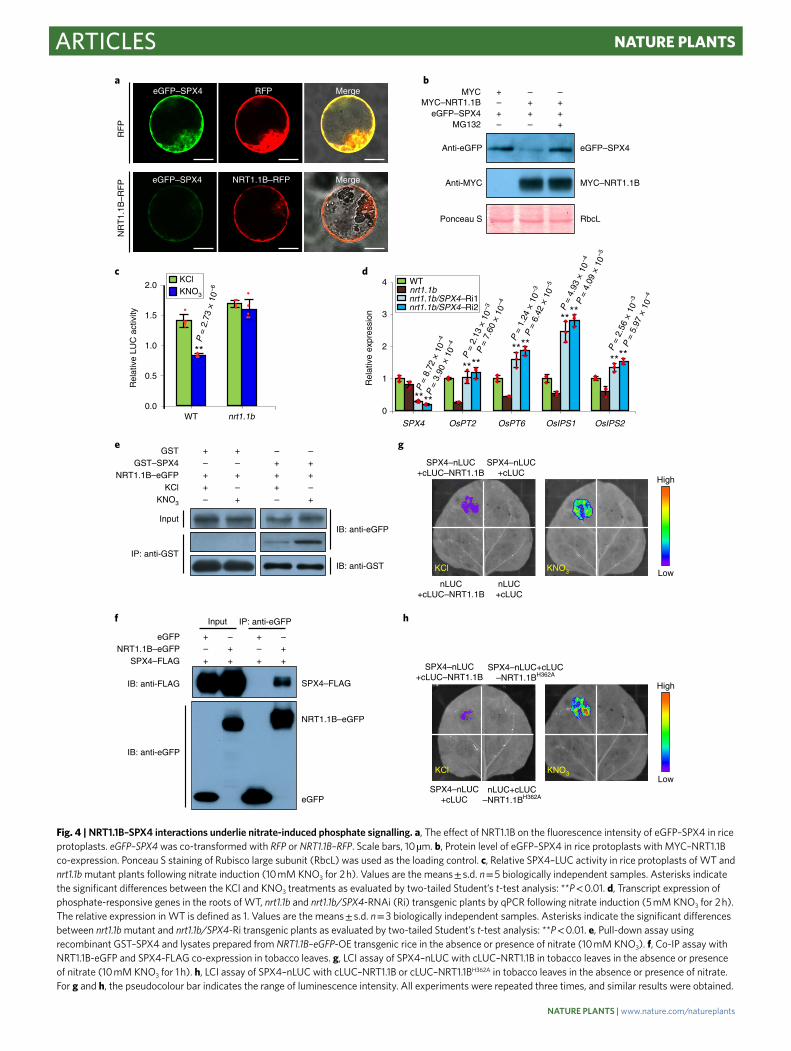
NRT1.1B mediates nitrate-triggered SPX4 degradation. We next investigated the relationship between NRT1.1B and SPX4 in nitrate-triggered phosphate signalling. Interestingly, we found that the co-expression of NRT1.1B decreased the fluorescence intensity of eGFP–SPX4 in rice protoplasts (Fig. 4a), indicat-ing that NRT1.1B might promote the degradation of the SPX4 protein. Correspondingly, in rice protoplasts, the protein level of eGFP–SPX4 was markedly decreased when co-expressed with NRT1.1B, and this effect was repressed by MG132 (Fig. 4b), fur-ther suggesting the involvement of ubiquitination in this process. Moreover, compared with the wild type, nitrate-induced degra-dation of SPX4–luciferase (LUC) was repressed in protoplasts of nrt1.1b mutant plants (Fig. 4c). These results indicated that NRT1.1B works upstream of SPX4 and mediates its degradation in response to nitrate. To provide genetic evidence for this relation-ship, we reduced SPX4 expression using RNA interference (RNAi) in nrt1.1b mutant plants (nrt1.1b/SPX4-RNAi) (Fig. 4d). Indeed, the expression of PSI genes was upregulated in nrt1.1b/SPX4-RNAi transgenic plants compared with nrt1.1b mutant plants (Fig. 4d). That is, SPX4 knock-down rescued the impairment of phosphate signalling caused by loss-of-function of NRT1.1B. These results
collectively demonstrate that NRT1.1B mediates the partial degra-dation of SPX4, through which nitrate can activate phosphate sig-nalling to coordinate N and P utilization.
Nitrate promotes NRT1.1B–SPX4 interactions. Given the func-tional relationship between NRT1.1B and SPX4, we speculated that these two components may form a complex. In rice protoplasts, a large proportion of eGFP–SPX4 was localized to the plasma membrane, especially under nitrate treatment (Supplementary Fig. 4a). Moreover, SPX4 largely co-localized in the plasma mem-brane with NRT1.1B (Supplementary Fig. 4b), indicating a possible interaction between NRT1.1B and SPX4. Thus, both in vitro and in vivo NRT1.1B–SPX4 interaction assays were performed. The in vitro pull-down assay showed co-purification of NRT1.1B–eGFP
with recombinant glutathione S-transferase (GST)–SPX4 fusion protein (Supplementary Fig. 4c). Importantly, nitrate enhanced the in vitro interaction (Fig. 4e; Supplementary Fig. 4d). Co-immunoprecipitation (co-IP) assays in tobacco (Nicotiana ben-thamiana) leaves also confirmed that NRT1.1B interacts with SPX4 in vivo (Fig. 4f). Additionally, luciferase complementation imaging (LCI) assays in tobacco leaves showed that co-expressing C-terminal luciferase fragment (cLUC)–NRT1.1B and SPX4–N-terminal lucif-erase fragment (nLUC) produced a weak fluorescence signal, which was highly enhanced in the presence of nitrate (Fig. 4g). Previous studies have revealed that His356 is a crucial nitrate-binding site in AtNRT1.1, and this site is conserved between AtNRT1.1 and NRT1.1B22,23. To further address whether the perception of nitrate is required for enhancing NRT1.1B–SPX4 interactions,
LN
LP HP
HN
LP HP
a b
0
1
2
3
4
LN HN
Upt
ake
activ
ity (
Pi μ
g g–1
roo
t FW
h–1
)
c
0.000
0.004
0.008
0.012
0.016
0 0.25 0.5 1 2 4
OsPT2
0.000
0.004
0.008
0.012
0.016
0 0.25 0.5 1 2 4
OsPT6
Rel
ativ
e ex
pres
sion
Rel
ativ
e ex
pres
sion
0.00
0.05
0.10
0.15
0.20
0 0.25 0.5 1 2 4
OsIPS1
0.00
0.10
0.20
0.30
0.40
0 0.25 0.5 1 2 4
OsIPS2
KNO3
KCl
KNO3
KCl
d
P = 2.23 × 10–4
LN HN
HPLP
** **
6
5
4
3
2
1
0
Bio
mas
s (g
)
P = 2.37 × 10–19
KNO3
KCl
KNO3
KCl
Rel
ativ
e ex
pres
sion
Rel
ativ
e ex
pres
sion
Time (h) Time (h)
Time (h) Time (h)
Fig. 1 | Nitrate activates phosphate utilization in rice. a, Growth performance of Zhonghua 11 wild-type plants (WT) under different N and P conditions for 2 months. High nitrate (HN), 5 mM KNO3; low nitrate (LN), 0.2 mM KNO3; high phosphate (HP), 0.18 mM KH2PO4; low phosphate (LP), 0.018 mM KH2PO4. Scale bars, 40 cm. Images are representative of 12 rice plants. b, Biomass analysis of WT plants under different N and P conditions. n = 12 biologically independent samples. Boxes represent the median values and the first and third quartiles; whiskers represent the minimum and maximum values. Asterisks indicate the significant differences between different P treatments under certain N conditions as evaluated by two-tailed Student’s t-test analysis: **P < 0.01. c, Phosphate (Pi) uptake activity assay of rice seedlings under LN and HN conditions. FW, fresh weight. Values are the means ± s.d. n = 4 biologically independent samples. Asterisks indicate the significant differences between LN and HN treatments as evaluated by two-tailed Student’s t-test analysis: **P < 0.01. d, Nitrate induction (5 mM KNO3) assay of phosphate-responsive genes in the roots of WT plants by quantitative PCR (qPCR). The growth media contained 0.18 mM KH2PO4 and KCl was used as the negative control. Values are the means ± s.d. n = 3 biologically independent samples.
we mutated the corresponding His362 in NRT1.1B (H362A), and found that this mutation prevented the interaction of these two proteins irrespective of nitrate (Fig. 4h). These results demonstrate that NRT1.1B–SPX4 physically interacts in vivo and that this inter-action is facilitated by the perception of nitrate by NRT1.1B. Thus, SPX4 also participates in the nitrate signalling pathway.
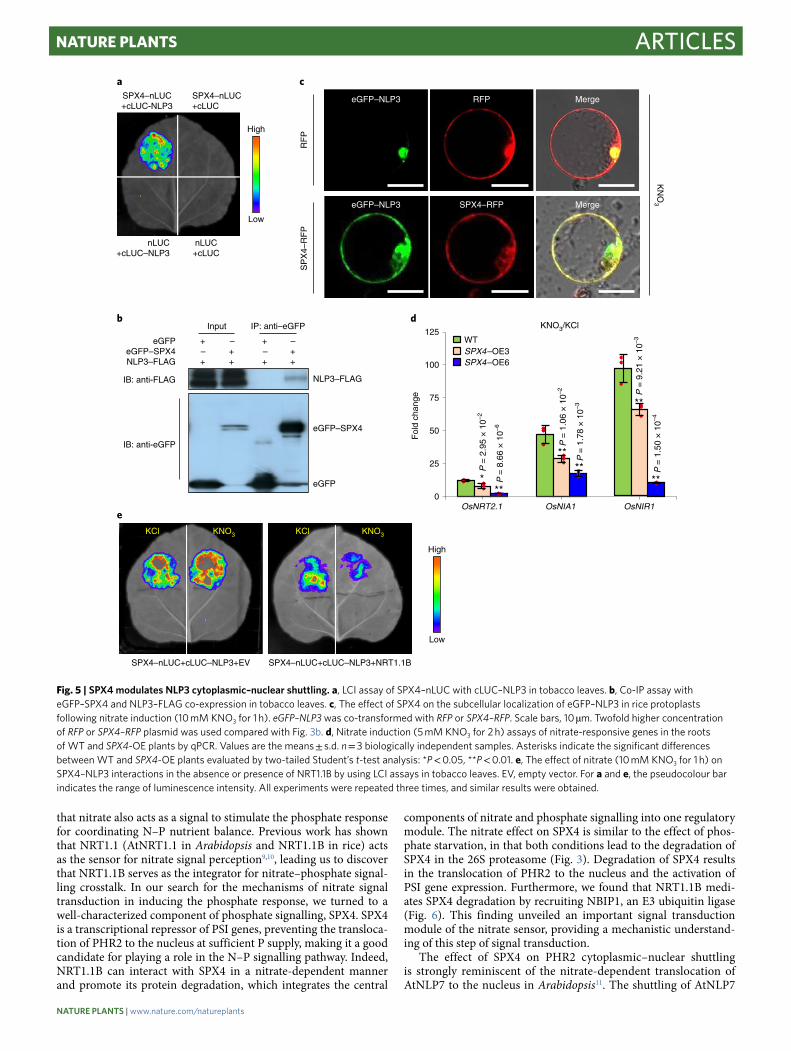
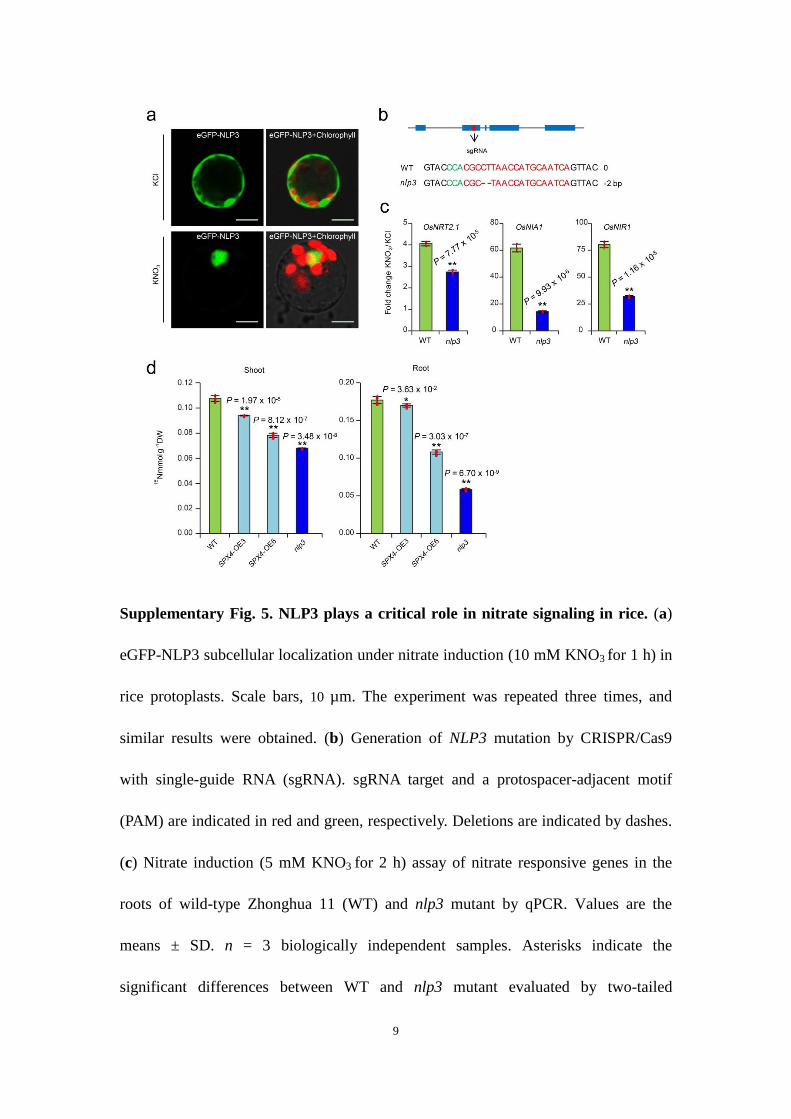
SPX4 is involved in the nitrate signalling pathway by modulating NLP3 cytoplasmic–nuclear shuttling. Our results revealed that SPX4 is directly downstream of NRT1.1B in nitrate signal trans-duction. Given that SPX4 plays an important role in modulating the cytoplasmic–nuclear shuttling of PHR2, we hypothesized that SPX4 might also regulate the cytoplasmic–nuclear shuttling of the NLP transcription factor and represent the missing link between the nitrate sensor and transcription factors in nitrate signal transduc-tion. In rice, OsNLP3 (NLP3; LOC_Os1g13540) is the closest homo-logue to AtNLP724. The nuclear localization of NLP3 was greatly facilitated by nitrate treatment (Supplementary Fig. 5a), and its loss-of-function mutant displayed a severe defect in the induction of nitrate-responsive genes (Supplementary Fig. 5b,c), indicating that NLP3 plays a critical role in nitrate signalling in rice. Interestingly, both LCI and co-IP assays in tobacco leaves showed that SPX4 interacts with NLP3 in vivo (Fig. 5a,b). In rice protoplasts, SPX4 co-expression repressed the nitrate-induced cytoplasmic–nuclear
shuttling of NLP3 (Fig. 5c). Correspondingly, the induction of typical nitrate-responsive genes, including OsNRT2.1 (LOC_Os02g02170), OsNIA1 (LOC_Os08g36480) and OsNIR1 (LOC_Os01g25484), was repressed in SPX4-overexpressing rice (Fig. 5d). In uptake experi-ments using 15N-nitrate labelling, both SPX4-overexpressing plants and nlp3 mutants displayed lower 15N accumulation than wild-type plants (Supplementary Fig. 5d). Moreover, nitrate repressed SPX4–NLP3 interactions in a NRT1.1B-dependent manner, as shown by the LCI assays (Fig. 5e), indicating that nitrate promotes the release of NLP3 from SPX4 in NRT1.1B-mediated nitrate signalling. All these results demonstrate that SPX4 connects nitrate signal percep-tion via NRT1.1B and downstream nitrate response activation via NLP3 in the nitrate signal transduction pathway.
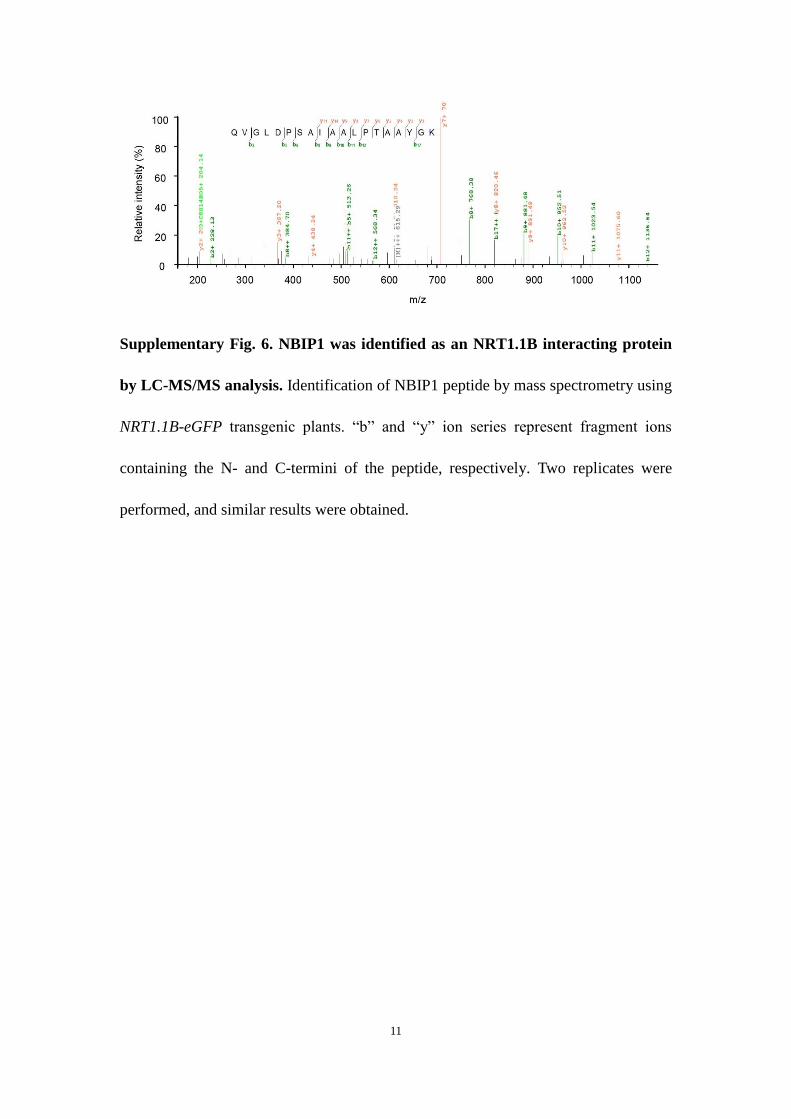
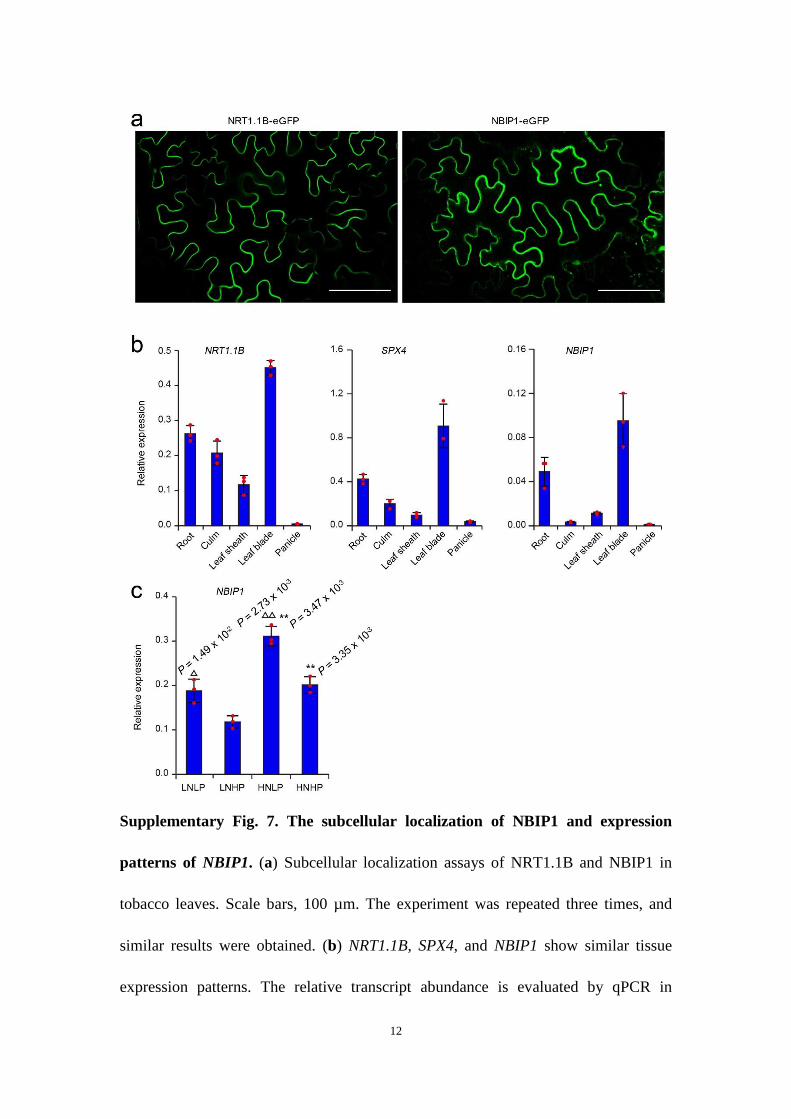
NRT1.1B recruits an E3 ubiquitin ligase to mediate SPX4 degra-dation. To explore the mechanism of NRT1.1B-mediated SPX4 deg-radation, we identified NRT1.1B-interacting proteins using co-IP coupled with mass spectrometry (IP–MS). Among the NRT1.1B-interacting proteins (NBIPs), a putative RING-type E3 ubiquitin ligase, NBIP1, attracted the most attention, as NRT1.1B-mediated SPX4 degradation depends on the 26S proteasome (Supplementary Fig. 6). Similar to NRT1.1B, NBIP1 protein predominantly local-ized to the plasma membrane, as demonstrated in rice protoplasts and in tobacco leaves (Fig. 6a; Supplementary Fig. 7a), and NBIP1
a
bLN, LP HN, LPLN, HP HN, HP
WT nrt1.1b WT nrt1.1b WT nrt1.1b WT nrt1.1b
0
2
4
6
8
OsIPS1 OsIPS2OsPT2 OsPT6
Fol
d ch
ange
** **
**
**
nrt1.1b
WT
c
4
6
5
3
2
1
0
Bio
mas
s (g
)
KNO3/KCl
LN, LP LN, HP HN, LP HN, HP
nrt1.1b
WT
***
****
P =
5.3
0 ×
10–4
P =
2.4
3 ×
10–4
P =
3.9
9 ×
10–5
P =
2.8
2 ×
10–4
P =
1.2
5 ×
10–2
P =
5.1
2 ×
10–1
0
P =
3.7
9 ×
10–4
P =
2.5
6 ×
10–2
0
Fig. 2 | NRT1.1B is involved in nitrate-induced phosphate signalling. a, Nitrate induction (5 mM KNO3 for 2 h) assay of phosphate-responsive genes in the roots of WT and nrt1.1b mutant plants by qPCR. Values are the means ± s.d. n = 3 biologically independent samples. b, Growth performance of WT and nrt1.1b mutant plants under different N and P conditions. Scale bars, 40 cm. Images are representative of 12 rice plants. c, Biomass analysis of WT and nrt1.1b mutant plants under different N and P conditions. The different N and P growth conditions are the same as those presented in Fig. 1. n = 12 biologically independent samples. Boxes represent the median values and the first and third quartiles; whiskers represent the minimum and maximum values. Asterisks indicate significant differences between WT and the nrt1.1b mutant as evaluated by two-tailed Student’s t-test analysis: *P < 0.05, **P < 0.01.
displayed similar tissue expression patterns as NRT1.1B and SPX4 (Supplementary Fig. 7b). The transcript expression of NBIP1 was upregulated by low phosphate and high nitrate treatments (Supplementary Fig. 7c). NBIP1-overexpressing plants showed upreg-ulation of both PSI and nitrate-inducible genes (Fig. 6b). Moreover, these plants displayed a significant increase in phosphate concentra-tion and a typical phosphate toxicity phenotype, which were also observed in the spx4 mutant (Supplementary Fig. 8). These results indicate that NBIP1 may be involved in regulating both phosphate and nitrate signalling. The co-IP assay in tobacco leaves confirmed that NRT1.1B interacts with NBIP1 in vivo (Supplementary Fig. 9a). Moreover, the co-IP assay of NRT1.1B, NBIP1 and SPX4 further revealed that these three components can form a complex in vivo (Fig. 6c). However, SPX4 and NBIP1 showed a weak interaction in the absence of NRT1.1B in the co-IP assay (Supplementary Fig. 9b), indicating that NRT1.1B plays a critical role in recruiting SPX4 and NBIP1 into a complex in vivo. The in vitro ubiquitination assay con-firmed that NBIP1 has E3 ubiquitin ligase activity (Fig. 6d), and can ubiquinate SPX4 (Fig. 6e). Moreover, NBIP1 significantly pro-moted the degradation of eGFP–SPX4 in protoplasts of wild-type
plants (Fig. 6f), whereas this process was repressed in protoplasts of the nrt1.1b mutant (Supplementary Fig. 10). Together, these results demonstrate that in the presence of nitrate, NRT1.1B recruits SPX4 to facilitate its ubiquitination and degradation by NBIP1. Thus, the NRT1.1B–NBIP1–SPX4 regulatory module explains the mecha-nism that can integrate the nitrate–phosphate signalling network, from nitrate signal perception (NRT1.1B), signal transduction (NRT1.1B–NBIP1–SPX4–PHR2/NLP3) and nitrate–phosphate responses (expression of nitrate- and phosphate-responsive genes).
DiscussionN is the most important mineral nutrient driving plant growth and crop yield. However, to reach its full potential in improving plant performance, other essential mineral nutrients, such as P, need to be sufficiently available. Because nutrient supply in soil is highly fluctuating, to achieve optimal and coordinated utilization of N and P, integration of N and P signalling into a combined network is required25. Nitrate has long been recognized as having a dual role as a nutrient and a signal to modulate the expression of more than a hundred genes, primarily for nitrate utilization26. We have revealed
0.00
0.02
0.04
0.06
0.08
0 0.25 0.5 1 2 4
SPX4
KNO3
KCl
Rel
ativ
e ex
pres
sion
a
c
KN
O3
KC
l
PHR2–eGFP PHR2–eGFP+Chlorophyll
PHR2–eGFP PHR2–eGFP+Chlorophyll
d
KNO3KClKNO3 +MG132
SPX4–FLAG
ACTIN
Anti-FLAG
Anti-ACTIN
KC
l
eGFP–SPX4 eGFP–SPX4+Chlorophyll
beGFP–SPX4 eGFP–SPX4+Chlorophyll
KN
O3
PHR2–eGFP+SPX4–RFPPHR2–eGFP
PHR2–eGFP PHR2–eGFP+RFP
KN
O3
RF
P
e
SP
X4-
RF
P
Time (h)
Fig. 3 | The SPX4PHR2 module is regulated by nitrate treatment. a, Nitrate induction (5 mM KNO3) assay of SPX4 transcript expression in the roots of WT plants by qPCR. Values are the means ± s.d. n = 3 biologically independent samples. b, eGFP–SPX4 fluorescence following nitrate induction in rice protoplasts. c, Protein level of SPX4–FLAG in roots of SPX4–FLAG-overexpressing (OE) transgenic rice after nitrate induction. d, PHR2–eGFP subcellular localization following nitrate induction in rice protoplasts. e, The effect of SPX4 on the subcellular localization of PHR2–eGFP in rice protoplasts following nitrate induction. PHR2–eGFP was co-transformed with red fluorescent protein (RFP) or SPX4–RFP. Scale bars, 10 µm. Twofold higher concentration of RFP or SPX4–RFP plasmid was used compared to b. Nitrate induction was 5 mM KNO3 for 2 h in c, 10 mM KNO3 for 1 h in b, d and e. KCl was used as the negative control. All experiments were repeated three times, and similar results were obtained.
Fig. 4 | NRT1.1B–SPX4 interactions underlie nitrate-induced phosphate signalling. a, The effect of NRT1.1B on the fluorescence intensity of eGFP–SPX4 in rice
protoplasts. eGFP–SPX4 was co-transformed with RFP or NRT1.1B–RFP. Scale bars, 10�μm. b, Protein level of eGFP–SPX4 in rice protoplasts with MYC–NRT1.1B
co-expression. Ponceau S staining of Rubisco large subunit (RbcL) was used as the loading control. c, Relative SPX4–LUC activity in rice protoplasts of WT and
nrt1.1b mutant plants following nitrate induction (10�mM KNO3 for 2�h). Values are the means�±�s.d. n�=�5 biologically independent samples. Asterisks indicate
the significant differences between the KCl and KNO3 treatments as evaluated by two-tailed Student’s t-test analysis: **P�<�0.01. d, Transcript expression of
phosphate-responsive genes in the roots of WT, nrt1.1b and nrt1.1b/SPX4-RNAi (Ri) transgenic plants by qPCR following nitrate induction (5�mM KNO3 for 2�h).
The relative expression in WT is defined as 1. Values are the means�±�s.d. n�=�3 biologically independent samples. Asterisks indicate the significant differences
between nrt1.1b mutant and nrt1.1b/SPX4-Ri transgenic plants as evaluated by two-tailed Student’s t-test analysis: **P�<�0.01. e, Pull-down assay using
recombinant GST–SPX4 and lysates prepared from NRT1.1B–eGFP-OE transgenic rice in the absence or presence of nitrate (10�mM KNO3). f, Co-IP assay with
NRT1.1B-eGFP and SPX4-FLAG co-expression in tobacco leaves. g, LCI assay of SPX4–nLUC with cLUC–NRT1.1B in tobacco leaves in the absence or presence
of nitrate (10�mM KNO3 for 1�h). h, LCI assay of SPX4–nLUC with cLUC–NRT1.1B or cLUC–NRT1.1BH362A in tobacco leaves in the absence or presence of nitrate.
For g and h, the pseudocolour bar indicates the range of luminescence intensity. All experiments were repeated three times, and similar results were obtained.
that nitrate also acts as a signal to stimulate the phosphate response for coordinating N–P nutrient balance. Previous work has shown that NRT1.1 (AtNRT1.1 in Arabidopsis and NRT1.1B in rice) acts as the sensor for nitrate signal perception9,10, leading us to discover that NRT1.1B serves as the integrator for nitrate–phosphate signal-ling crosstalk. In our search for the mechanisms of nitrate signal transduction in inducing the phosphate response, we turned to a well-characterized component of phosphate signalling, SPX4. SPX4 is a transcriptional repressor of PSI genes, preventing the transloca-tion of PHR2 to the nucleus at sufficient P supply, making it a good candidate for playing a role in the N–P signalling pathway. Indeed, NRT1.1B can interact with SPX4 in a nitrate-dependent manner and promote its protein degradation, which integrates the central
components of nitrate and phosphate signalling into one regulatory module. The nitrate effect on SPX4 is similar to the effect of phos-phate starvation, in that both conditions lead to the degradation of SPX4 in the 26S proteasome (Fig. 3). Degradation of SPX4 results in the translocation of PHR2 to the nucleus and the activation of PSI gene expression. Furthermore, we found that NRT1.1B medi-ates SPX4 degradation by recruiting NBIP1, an E3 ubiquitin ligase (Fig. 6). This finding unveiled an important signal transduction module of the nitrate sensor, providing a mechanistic understand-ing of this step of signal transduction.
The effect of SPX4 on PHR2 cytoplasmic–nuclear shuttling is strongly reminiscent of the nitrate-dependent translocation of AtNLP7 to the nucleus in Arabidopsis11. The shuttling of AtNLP7
Fig. 5 | SPX4 modulates NLP3 cytoplasmic–nuclear shuttling. a, LCI assay of SPX4–nLUC with cLUC–NLP3 in tobacco leaves. b, Co-IP assay with eGFP–SPX4 and NLP3–FLAG co-expression in tobacco leaves. c, The effect of SPX4 on the subcellular localization of eGFP–NLP3 in rice protoplasts following nitrate induction (10 mM KNO3 for 1 h). eGFP–NLP3 was co-transformed with RFP or SPX4–RFP. Scale bars, 10 µm. Twofold higher concentration of RFP or SPX4–RFP plasmid was used compared with Fig. 3b. d, Nitrate induction (5 mM KNO3 for 2 h) assays of nitrate-responsive genes in the roots of WT and SPX4-OE plants by qPCR. Values are the means ± s.d. n = 3 biologically independent samples. Asterisks indicate the significant differences between WT and SPX4-OE plants evaluated by two-tailed Student’s t-test analysis: *P < 0.05, **P < 0.01. e, The effect of nitrate (10 mM KNO3 for 1 h) on SPX4–NLP3 interactions in the absence or presence of NRT1.1B by using LCI assays in tobacco leaves. EV, empty vector. For a and e, the pseudocolour bar indicates the range of luminescence intensity. All experiments were repeated three times, and similar results were obtained.
is essential for its function in nitrate-controlled transcription, but the mechanism is unknown. Therefore, we hypothesized that SPX4 might also control the localization of NLP factors. Indeed, we revealed that the nuclear localization of NLP3, the NLP
member in rice that is most closely related to AtNLP7, is hindered by the co-expression of SPX4 (Fig. 5). Having linked NRT1.1B nitrate sensing and SPX4 degradation into a single regulatory module, this finding adds a critical piece of information to
Fig. 6 | NRT1.1B recruits NBIP1 to mediate SPX4 ubiquitination and degradation. a, Subcellular localization of NBIP1–eGFP in rice protoplasts. The transformed protoplasts were stained with FM4-64 for 10 min. Scale bars, 10 µm. b, Transcript expression of phosphate- and nitrate-responsive genes in the roots of WT and NBIP1-OE plants cultured in no-nitrate basal nutrient solution (see ‘Plant materials and growth conditions’ in Methods) by qPCR. The relative expression in WT plants is defined as 1. Values are the means ± s.d. n = 3 biologically independent samples. Asterisks indicate the significant differences between WT and NBIP1-OE plants as evaluated by two-tailed Student’s t-test analysis: *P < 0.05, **P < 0.01. c, Co-IP assay with eGFP–SPX4, NBIP1–FLAG and MYC–NRT1.1B co-expression in tobacco leaves. d, In vitro self-ubiquitination assay of transmembrane domain truncated NBIP1 (NBIP1T). Ubiquitin (Ub)-attached proteins were detected with an anti-Ub antibody (upper gel) and an anti-MBP antibody (lower gel). Ubn, ubiquitin conjugates. e, Detection of the ubiquitination of GST–SPX4 by NBIP1T in vitro. An anti-GST antibody was used for western blotting for detecting substrates. Asterisk indicates the ubiquitinated GST–SPX4 protein. f, Protein level of eGFP–SPX4 in rice protoplasts with NBIP1–FLAG co-expression. Ponceau S staining was used as the loading control. All experiments were repeated three times, and similar results were obtained.
complete the nitrate signalling pathway, from signal perception to downstream transcriptional responses. Additionally, the parallel modulation of PHR2 and NLP3 by SPX4 enables the coordinated activation of both nitrate and phosphate responses. Therefore, our work establishes a nitrate–phosphate interaction signalling path-way, whereby the nitrate signal is sensed by NRT1.1B and enhances the association of NRT1.1B, SPX4 and NBIP1, resulting in the ubiq-uitination and degradation of SPX4. This enables the subsequent release of PHR2 and NLP3 into the nucleus to activate the expres-sion of both phosphate- and nitrate-responsive genes (Fig. 7). Such a mechanism underlying N–P nutrient balance has enormous sig-nificance for plants to adapt to the changing nutritional environ-ment. It also provides an important paradigm to understand the signal transduction networks for other mineral nutrients, and will contribute to an in-depth understanding of the mechanism under-lying coordinated utilization of different nutrients in plants.
The interaction between NRT1.1B and SPX4 is critical for estab-lishing the nitrate signalling cascade. Therefore, it is essential that these two proteins are expressed in the same cells. NRT1.1B is mainly expressed in the epidermal and vascular tissues of the pri-mary roots10, in which SPX4 is also highly expressed (Supplementary Fig. 11). It has also been observed that NRT1.1B is a plasma mem-brane-integrated protein, but it has several loop regions exposed in the cytosol. It is possible that these loops constitute a binding domain for interacting with SPX4 close to the intracellular layer of the cell membrane. Thus, we speculate that nitrate triggers a struc-tural alteration of NRT1.1B protein and facilitates the exposure of the loop-containing domain, thereby strengthening the interaction between NRT1.1B and SPX4. Interestingly, our previous work had revealed that natural variation in NRT1.1B might contribute to the divergent ways that rice subspecies use nitrate10. Combined with the findings presented in this study, it will be intriguing to investigate whether the allelic variation of NRT1.1B also contributes to phos-phate utilization divergence in different rice varieties. Notably, the
amino-acid change in NRT1.1B between japonica and indica sub-species of O. sativa occurs in the central loop region, which may constitute the SPX4-interacting domain.
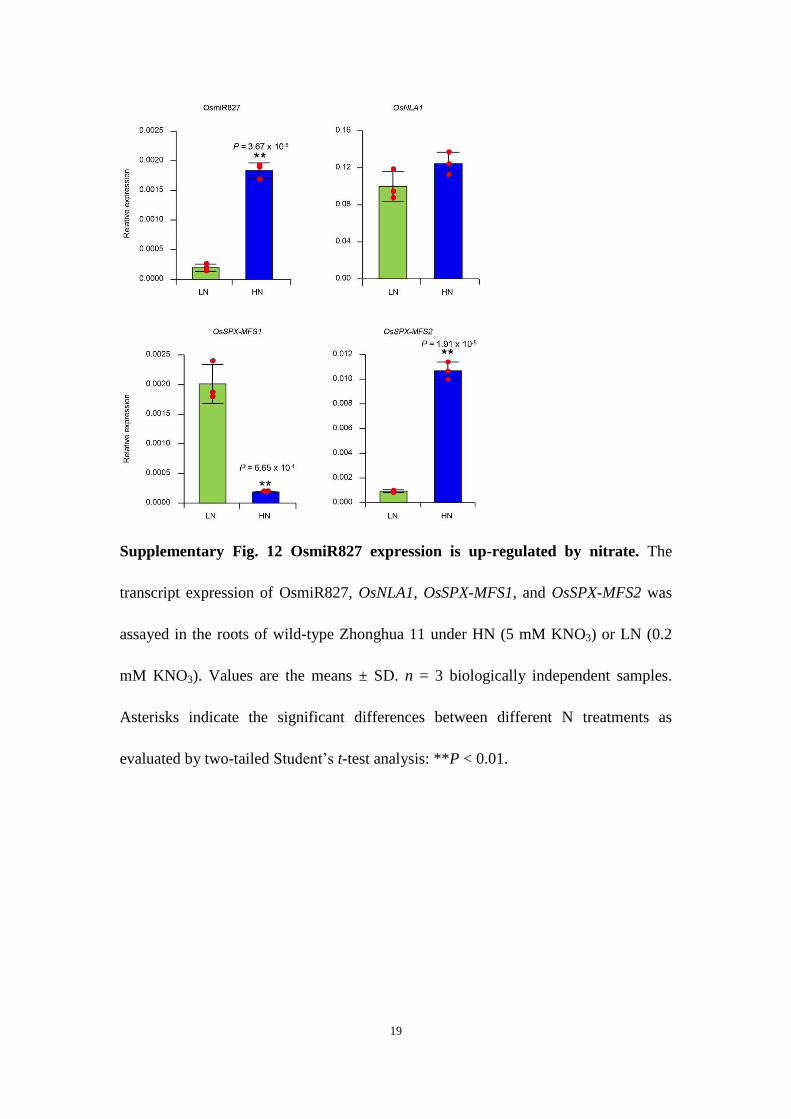
The microRNA827 (miR827, AT3G59884)–NLA (AT1G02860) module has been suggested to be involved in nitrate-dependent phosphate homeostasis in Arabidopsis18. In rice, although both OsmiR827 (MI0007051) and OsNLA1 (LOC_Os07g47590) are involved in phosphate homeostasis, OsmiR827 cannot target OsNLA1 to regulate its expression27,28. Interestingly, we found that OsmiR827 could be induced by nitrate, while the expression of OsNLA1 did not show significant alteration under different nitrate conditions (Supplementary Fig. 12). Interestingly, OsSPX–MFS1 (LOC_Os04g48390) and OsSPX–MFS2 (LOC_Os02g45520), the targets of OsmiR827, displayed an opposite nitrate-responsive pat-tern (Supplementary Fig. 12). That is, OsSPX–MFS1 was repressed by nitrate while OsSPX–MFS2 was induced by nitrate. As OsmiR827 is one of the target genes of PHR2, OsmiR827 may amplify the output of nitrate-induced phosphate signalling. Additionally, increasing evidence indicates that the plant hormone cytokinin plays important roles in both phosphate and nitrate signalling29–31. Biosynthesis of cytokinin can be induced by nitrate in the roots32. However, exogenous cytokinin application represses the expres-sion of PSI genes33,34. These reports suggest that cytokinin may be involved in modulating nitrate-triggered phosphate signalling. Under nitrate induction, cytokinin possibly prevents the overactiva-tion of PSI genes to avoid excessive phosphate uptake and detrimen-tal phosphate overaccumulation in cells.
In this study, we focused on dissecting the mechanism of how nitrate stimulates phosphate signalling under a certain phosphate condition. Interestingly, we also noted that nitrate-induced expres-sion of PSI genes was greatly promoted under low phosphate condi-tions (Supplementary Fig. 2). Previous work has revealed that SPX4 acts as an intracellular phosphate sensor via binding InsPs15. It is possible that the accumulation of InsPs caused by high phosphate
Nitrate
NRT1.1B
Ub
Ub
SPX4
SPX4
Phosphate response
Nitrate response
PHR2
SPX4PHR2
SPX4
SPX4
NLP3
NLP3
NLP3
NLP3
SPX4
NLP3
SPX4
PHR2
PHR2PHR2
NRT1.1B
Ub
NBIP1
UbUb
NBIP1
26Sproteasome
Fig. 7 | Proposed model of N–P coordinated responses. In the absence or at low levels of nitrate (left), SPX4 is stable and binds to PHR2 and NLP3, and represses the downstream phosphate and nitrate responses. At high levels of nitrate (right), perception of nitrate leads to NRT1.1B–NBIP1-mediated SPX4 ubiquitination and then degradation by the 26S proteasome. This process releases PHR2 and NLP3, which coordinately activate downstream phosphate and nitrate responses, respectively.
supply can be sensed by SPX4, resulting in an increased SPX4 affin-ity to PHR2, thereby blocking the activation of PSI genes. Moreover, in high phosphate conditions, the nuclear-localized SPX1 and SPX2 can bind PHR2 and prevent it from activating PSI genes. In con-trast, under low phosphate conditions, this repression is released, and the PHR2-triggered gene expression is higher16,17. Recent work has shown that NIGT1 is involved in activating the expression of PT genes under high-nitrate and low-phosphate conditions in Arabidopsis20, which also provides an important explanation for N–P interactive regulation under varying phosphate availability. Additionally, NIGT1 can function as a repressor to inhibit nitrate uptake under phosphate deficiency in Arabidopsis21, which might contribute to maintaining the N–P balance under nutrient limita-tion. Alternatively, the NRT1.1B–SPX4 module may modulate the optimal N–P acquisition under a sufficient nutrient condition.
Given the critical role of SPX4 in phosphate sensing and in link-ing the nitrate sensor and the downstream transcription factors that control the nitrate response, it is possible that the NRT1.1B–SPX4 module is also involved in phosphate-regulated nitrate response. Indeed, we found that phosphate availability can modulate the expression of nitrate-responsive genes (Supplementary Fig. 13a), and the NRT1.1B–SPX4 module is involved in this process. This was demonstrated by the greatly repressed phosphate-regulated nitrate response observed in both the nrt1.1b mutant and SPX4-overexpressing plants (Supplementary Fig. 13b). Under high nitrate conditions (together with high or low phosphate supply), the expression of nitrate-responsive genes was repressed by high phosphate supply (Supplementary Fig. 13a). This result is consis-tent with the accumulation of SPX4 protein under high phosphate conditions (Supplementary Fig. 13c). Conversely, the expression of nitrate-responsive genes was upregulated by high phosphate supply under low nitrate conditions (Supplementary Fig. 13a), even though the SPX4 protein still accumulated to a higher level (Supplementary Fig. 13c). These results indicate that plants employ a highly sophis-ticated mechanism to implement phosphate-regulated nitrate responses in a nitrate-dependent manner, which might not only be associated solely with the SPX4 protein levels but also determined by phosphate-modulated SPX4-interacting affinity. Phosphate or InsPs could enhance the interaction between SPX4 and PHR2 (Supplementary Fig. 14a), although they did not significantly influ-ence the interaction between SPX4 and NRT1.1B (Supplementary Fig. 14b). It may be that the dominating regulatory action of SPX4 would switch between its protein level and the interacting affinity under different N and P conditions, therefore conferring distinct patterns of phosphate-regulated nitrate responses. More research is needed to address this intriguing but complicated issue.
MethodsPlant materials and growth conditions. The wild-type rice used in this study was Zhonghua 11 (O. sativa L. japonica, ZH11). The nrt1.1b mutant was previously identified in our laboratory10, and phr2, spx4 and nlp3 mutants were generated by Biogle35 using the clustered regularly interspaced short palindromic repeat (CRISPR)–Cas9-mediated editing method in ZH11. For short-term (1–3 weeks) hydroponic culture (for nitrate-induction assays, gene expression analyses, Pi concentration measurement and rice protoplast preparation), rice seedlings were cultivated in a growth chamber at a photoperiod of 12 h (light)–12 h (dark) (~200 μmol m−2 s−1) at 28 °C/25 °C and ~70% humidity. The basal nutrient solution consists of the following macronutrients (in mM): (NH4)2SO4 (0.25), MgSO4·7H2O (0.54), CaCl2·2H2O (0.36), K2SO4 (0.1), KH2PO4 (0.18) and Na2SiO3·9H2O (1.6). The nutrient solution also contained the following micronutrients (in μM): MnCl2·4H2O (9.14), H3BO3 (46.2), (NH4)6Mo7O24·4H2O (0.08), ZnSO4·7H2O (0.76), CuSO4·5H2O (0.32) and Fe(II)-EDTA (40), with the pH adjusted to 5.8. The nutrient solution was renewed every day. KNO3 was added to the basal nutrient solution at indicated concentrations for different treatments. For long-term (2–3 months) hydroponic culture (for growth observation and biomass analyses), rice plants were grown in an artificial weather room with a photoperiod of 12 h (light)–12 h (dark) (~300 μmol m−2 s−1) at 28 °C/25 °C and ~40% humidity. During long-term hydroponic culture, the N- and P-free basal nutrient solutions were supplemented with KNO3 (0.2 mM and 5 mM) and KH2PO4 (0.018 mM and
0.18 mM). The K+ concentration was adjusted with KCl to keep consistency among different conditions.
Construction of vectors and generation of transgenic rice. To generate NRT1.1B–RFP, SPX4–RFP, eGFP–SPX4, PHR2–eGFP and eGFP–NLP3, the coding regions (CDS) of NRT1.1B (japonica type), SPX4, PHR2 and NLP3 were amplified and inserted into pSAT6-RFP-N1 (CD3–1108), pCAMBIA2300–35S-eGFP-N and pCAMBIA2300–35S-eGFP-C, respectively. For the NBIP1 subcellular localization assay, the CDS of NBIP1 was amplified and cloned into pCAMBIA2300–35S-eGFP-C to generate NBIP1–eGFP. For the in vivo degradation assays in rice protoplasts, the CDS of NRT1.1B was amplified and cloned into pCAMBIA1300–221–35S-MYC to generate MYC–NRT1.1B. To generate SPX4–FLAG, the CDS of SPX4 was amplified and cloned into pCAMBIA1300–221–35S-FLAG to generate pCAMBIA1300–35S–SPX4–FLAG. The full-length CDS of firefly luciferase (fLUC) was amplified from pGreenII0800–LUC36 and inserted into pCAMBIA1300–35S–SPX4–FLAG to replace FLAG with fLUC. The resulting vector was named pCAMBIA1300–35S–SPX4–LUC, and used for estimating SPX4 protein stability in the rice protoplast system.
To generate the constructs of SPX4–nLUC, cLUC–NRT1.1B, cLUC–NLP3 and cLUC–PHR2 for investigating protein interactions in tobacco (N. benthamiana), the CDS of SPX4, NRT1.1B, NLP3 and PHR2 were amplified and cloned into pCAMBIA1300–nLUC and pCAMBIA1300-cLUC, respectively37. The construct of cLUC–NRT1.1BH362A was generated by site-directed mutagenesis.
For co-IP assays in tobacco, the construct of NRT1.1B–eGFP was obtained from our previous work10. To generate NLP3–FLAG, the CDS of NLP3 was amplified and cloned into pCAMBIA1300–221–35S–FLAG to generate pCAMBIA1300–35S–NLP3–FLAG. For in vitro ubiquitination assays, the GST–SPX4 construct was generated by cloning the CDS of SPX4 into pGEX-4T-3, and MBP–NBIP1T was constructed by cloning a truncated NBIP1 (181–717 bp) into pMAL-c2X.
To generate the SPX4 RNAi vector for rice genetic transformation, a specific sequence with a fusion fragment of SPX4 CDS and the 3′ untranslated region was amplified. The resulting PCR product was inserted into pUCC-RNAi vector in both the sense and antisense orientation, and then the fragment containing an artificial inverted-repeat sequence of SPX4 was transferred into pCAMBIA2301–ACTIN1 for the SPX4 RNAi construct38. For overexpression, the full-length CDS of NRT1.1B and SPX4 was amplified and cloned into pCAMBIA2300–35S, while NBIP1 was amplified and clones into pCAMBIA1300–221–35S–FLAG. The resulting vectors were pCAMBIA2300–35S–NRT1.1B, pCAMBIA2300–35S–SPX4 (for nitrate induction assay), and pCAMBIA1300–35S-NBIP1–FLAG. These constructs, together with pCAMBIA1300–35S–SPX4-FLAG (for SPX4 protein stability assay) and pCAMBIA2300–35S–PHR2–eGFP (for gene expression assay), were introduced into Agrobacterium strain AGL1. The wild-type ZH11 was used as the recipients for Agrobacterium-mediated transformation as described previously to generate the transgenic rice39. All the primers used to generate the constructs are listed in Supplementary Table 1, and all constructs were confirmed by sequencing. Homozygous T3 plants were use for the following experiments.
Measurement of Pi concentration. The shoots of about 15 rice seedlings were collected together for the Pi concentration assay, and at least 5 such pools were analysed as independent biological replicates. The Pi concentration was measured using a previously described procedure40,41. To analyse the Pi concentration of rice plants grown in the field, four independent pools of leaf blades, leaf sheaths and culms from five independent rice plants each were collected.
Pi uptake activity assay. To investigate the Pi uptake activity under different nitrate concentrations, rice seedlings were first grown in the basal nutrient solution for 2 weeks. Before the Pi uptake assay, rice seedlings were pretreated with fresh basal nutrient solution for an additional 2 h and then transferred to basal nutrient solution containing 0.2 mM or 5 mM KNO3 for 3 h. Pi uptake was determined as the amount of Pi depleted from the solution, and the uptake activity was calculated as the amount of Pi uptake per unit fresh weight of roots per unit time.
Labelling with 15N-nitrate for determination of 15N accumulation. For the 15N-nitrate accumulation assay, rice seedlings were cultured in the basal nutrient solution for 10 days. Before the 15N-accumulation assay, rice seedlings were pretreated with basal nutrient solution containing 2.5 mM (NH4)2SO4 for 48 h under continuous light conditions, and then transferred to the same nutrient solution supplemented with 5 mM 15N-labelled KNO3 (98 atom % 15N; Sigma-Aldrich, no. 335134) for 24 h. At the end of the labelling process, the roots were rinsed with 0.1 mM CaSO4 for 1 min and separated from the shoots. Roots and shoots were collected and ground into powder, and then the 15N content was determined using an isotope ratio mass spectrometer equipped with an elemental analyser (Thermo Finnigan Delta Plus XP; Flash EA 1112). For each of the 4 biological replicates, 20 seedlings were collected and measured.
Short-term nitrate induction assay of phosphate and nitrate-responsive genes. For all the short-term nitrate induction assays in this study, the rice seedlings were treated as follows. The seedlings were first cultured in basal nutrient solution (0.25 mM (NH4)2SO4, 0.18 mM KH2PO4) as described above for 3 weeks. Before
nitrate induction, the seedlings were pretreated with basal nutrient solution containing 2.5 mM (NH4)2SO4 and 0.18 mM KH2PO4 for 48 h under continuous light conditions, and then transferred to the same nutrient solution supplemented with 5 mM KNO3 or KCl. Roots of rice seedlings were collected at the indicated timepoints for gene expression analyses.
Subcellular localization assay. Nitrate-free rice protoplasts were used in all subcellular localization assays. Rice seedlings were grown in the basal nutrient solution for 10 days, and rice protoplasts were isolated and transfected as described previously42,43. To analyse SPX4 degradation and PHR2 nuclear retention triggered by nitrate in rice protoplasts, nitrate-free rice protoplasts were transfected with 35S–eGFP–SPX4 or 35S–PHR2–eGFP plasmid DNA, and then incubated in W5 solution (154 mM NaCl, 125 mM CaCl2, 5 mM KCl, 0.18 mM KH2PO4 and 2 mM MES at pH 5.7) for 12 h. Transfected rice protoplasts were spun down and subsequently treated with W5 solution containing 10 mM KNO3 or KCl for 1 h, and then fluorescence signals were captured using a confocal laser-scanning microscope (TCS SP5, Leica). To investigate the effect of SPX4 on PHR2 and NLP3 localization under nitrate induction, nitrate-free rice protoplasts were co-transfected with 35S–PHR2–eGFP/35S-eGFP–NLP3 and RFP empty vector control or 35S–SPX4–RFP (at a ratio of 1:2) and incubated in W5 solution for 12 h. Rice protoplasts were spun down for 3 min at 200 × g, treated with W5 solution containing 10 mM KNO3 for 1 h and then used for observation as described above. For analysis of NRT1.1B-mediated SPX4 degradation in rice protoplasts, 35S–eGFP–SPX4 plasmid DNA was co-transfected into nitrate-free rice protoplasts with 35S–RFP empty vector control or 35S–NRT1.1B–RFP and incubated in W5 solution for 12 h. Transfected protoplasts were spun down, resuspended in W5 solution again, and observed as described above. To confirm the subcellular localization pattern, the indicated constructs were also transiently expressed in N. benthamiana leaves by Agrobacterium-mediated infiltration (strain GV3101) as described previously44. For FM4-64 staining, transfected rice protoplasts were incubated with FM4-64 (with a final concentration of 10 µM) at 4 °C for 10 min. Excitation laser wavelengths of 488 nm, 514 nm and 563 nm were used for imaging GFP, FM4-64, and RFP signals, respectively. Images presented in the figures are representative of at least five protoplasts or three tobacco leaves.
SPX4 protein stability assay in transgenic rice. To analyse SPX4 stability in response to nitrate stimulus, transgenic rice plants harbouring 35S–SPX4–FLAG were cultured in basal nutrient solution for 14 days. Subsequently, transgenic plants were pretreated with basal nutrient solution containing 2.5 mM (NH4)2SO4 for 48 h under continuous light conditions and then subjected to the same nutrient solution supplemented with 5 mM KNO3 or KCl for 2 h. Roots of transgenic rice were collected and frozen in liquid nitrogen. Total protein was obtained by disrupting roots in 1× SDS buffer (60 mM Tris-HCl (pH 8.5), 2% SDS (w/v), 2.5% glycerol (v/v), 0.13 mM EDTA, 1× complete protease inhibitor and 10 μM MG132). Immunoblot (IB) analysis was performed using anti-FLAG antibody (Sigma, F1804) for detecting the SPX4–FLAG protein level, and anti-Actin antibody (Easybio, no. BE0027) as a protein loading control.
SPX4 protein stability assay in rice protoplasts. To confirm that NRT1.1B promotes SPX4 degradation, the plasmid DNA of 35S–eGFP–SPX4 was co-transfected with 35S–MYC–NRT1.1B or 35S–MYC into nitrate-free rice protoplasts and incubated in W5 solution for 12 h. Transfected protoplasts were subsequently treated with 20 μM MG132 or dimethyl sulfoxide for 1 h and then collected in protein extraction buffer as described above. The lysates were centrifuged at 18,000 × g for 10 min at 4 °C. The supernatant was used for immunoblotting using anti-MYC and anti-GFP antibodies. To investigate whether NBIP1-mediated SPX4 degradation depends on NRT1.1B, nitrate-free rice protoplasts were isolated from wild-type ZH11 and nrt1.1b mutant plants, and co-transfected with the 35S–eGFP–SPX4 plasmid and either the 35S–NBIP1–FLAG or 35S–FLAG plasmid. After incubation in W5 solution for 12 h, rice protoplasts were collected and lysed in protein extraction buffer followed by SDS–PAGE and immunoblot analysis using the indicated antibodies.
Luciferase activity assay in rice protoplasts. For analysis of the SPX4 degradation rate in rice protoplasts, the plasmid DNA of 35S–SPX4–LUC and PTRL (containing a 35S–rLUC cassette for normalization purpose) were co-transfected into nitrate-free rice protoplasts isolated from wild-type ZH11 and nrt1.1b mutant plants. After incubation in W5 solution for 12 h, transfected protoplasts were treated with 10 mM KNO3 or KCl for 2 h. Activities of firefly luciferase (fLUC) and Renilla luciferase (rLUC) were assayed using a Dual-Luciferase Reporter Assay System kit (Promega). The data are presented as the ratio of fLUC/rLUC activity of five biological replicates.
Firefly LCI assays. To investigate the protein interaction between SPX4 and NRT1.1B or NRT1.1BH362A, the LCI assays were carried out as previously described37. Briefly, a Agrobacterium tumefaciens strain GV3101 carrying the indicated constructs was incubated in Luria–Bertani medium at 28 °C for 16 h, and the culture was pelleted and re-suspended in infiltration buffer (10 mM MES (pH 5.6), 10 mM MgCl2 and 0.2 mM acetosyringone) to a final concentration
of absorbance (A) at 600 of 1.0. The Agrobacterium cultures were kept at room temperature for at least 3 h without shaking. For co-infiltration, equal volumes of Agrobacterium suspensions carrying the indicated constructs were infiltrated into tobacco leaves. After infiltration, plants were cultured at 23 °C for 48 h at 16 h (light)–8 h (dark) photoperiod. The infiltrated leaves were first treated with 10 mM KNO3 or KCl for 1 h, and then sprayed with 1 mM luciferin (Sigma, L9504) supplemented with 0.01% Triton X-100 in darkness for 3 min before detection of luminescence. Images of luminescence were captured using a low-light-cooled CCD imaging apparatus (NightOWL II LB983). Images presented in the figures are representative of at least five tobacco leaves.
For LCI assays between SPX4 and NLP3, the method described above was carried out except without KNO3 or KCl treatment. To investigate the effect of nitrate and NRT1.1B on SPX4–NLP3 interactions, 35S–SPX4–nLUC and 35S–cLUC–NLP3 were co-infiltrated with equal volumes of 35S–MYC or 35S–MYC–NRT1.1B. The nitrate treatment and luciferase fluorescence image capture were performed as described above. For LCI assays between SPX4 and PHR2/NRT1.1B with phosphate or InsP6 treatments, 35S–SPX4–nLUC and 35S–cLUC–PHR2/NRT1.1B were co-infiltrated into tobacco leaves pretreated with 2 mM KNO3 for 2 days. The infiltrated leaves were treated with phosphate (0.18 mM KH2PO4 or 0.18 mM KCl for 1 h) or InsP6 (300 µM InsP6 or H2O for 1 h) and images of luciferase fluorescence was captured as described above.
RNA isolation and qPCR analysis. Total RNA was isolated using TRIzol reagent (Invitrogen) according to the manufacturer’s instructions. DNA removal and reverse transcription reactions were performed using ReverTra Ace qPCR RT Master Mix (Toyobo). qPCR assays were performed using SYBR Green Real-Time PCR Master Mix reagent (Toyobo) on a Chromo4 real-time PCR detection system according to the manufacturer’s instructions (Bio-Rad, CFX96). qPCR data were analysed using Opticon monitor software (Bio-Rad). Three biological replicates were performed for each gene. Rice ACTIN1 was used as the internal control for all analyses. Primers for qPCR are listed in Supplementary Table 1.
Co-IP assays. Total protein was extracted from infiltrated tobacco leaves with protein extraction buffer (50 mM Tris-HCl (pH 7.5), 100 mM NaCl, 1 mM EDTA, 10 mM NaF, 5 mM Na3VO4, 0.25% Triton X-100, 0.25% NP-40, 1 mM PMSF, 1× protease inhibitor cocktail and 10 μM ΜG132), and then incubated with 20 μl anti-GFP agarose beads (Chromotek, gta-20) for 2 h at 4 °C. The beads were washed five times with wash buffer (50 mM Tris-HCl (pH 7.5), 100 mM NaCl, 1 mM EDTA, 1 mM PMSF and 1× protease inhibitor cocktail), and the precipitated proteins were eluted with 2× SDS loading buffer at 95 °C for 3 min. The samples were subjected to immunoblot analysis using the indicated antibodies. For all immunoblot analyses for NRT1.1B–eGFP, the protein samples were not boiled.
Semi-in vivo pull-down assay. Total protein was extracted from the shoots of 35S–NRT1.1B–eGFP transgenic plants with protein extraction buffer as described above. After centrifugation at 15,000 × g for 10 min at 4 °C, the supernatants were incubated with glutathione agarose-bound GST or GST–SPX4 protein in the presence of 10 mM KNO3 or KCl for 4 h at 4 °C. The beads were washed five times with wash buffer and boiled with 2× SDS loading buffer at 95 °C for 3 min. The samples were detected by anti-GST and anti-GFP antibodies.
Preparation of recombinant proteins. To obtain recombinant proteins, the plasmids GST–SPX4 and MBP–NBIP1T (the full-length NBIP1 protein was expressed with very low efficiency in Escherichia coli because of the transmembrane domain in its amino terminus; therefore, the transmembrane domain-truncated NBIP1 fragment was cloned into pMAL-c2X to generate the MBP–NBIP1T construct) were transformed into E. coli strain BL21 (DE3). The bacterial cells were cultivated at 37 °C with shaking of 150–200 r.p.m. When the A600 reached 0.4–1.0, the bacterial cells were induced by 0.2 mM isopropylthiogalactoside at 37 °C for 3 h. Cells were collected and disrupted using a high pressure homogenizer (JNBIO) at 4 °C. The recombinant proteins were affinity-purified with amylose resin (NEB) or Glutathione–Sepharose (GE) according to manufacturer’s instructions.
In vitro ubiquitination assay. The in vitro ubiquitination assays were performed as described previously45,46. For the in vitro self-ubiquitination assay, 200 ng MBP–NBIP1T protein was mixed with 50 ng of Arabidopsis E1 (AtE1, At5g06460), 50 ng of human E2 (UbcH5b) and 2 μg of Arabidopsis ubiquitin (UBQ14, At4g02890) in 30 μl of ubiquitination reaction buffer (50 mM Tris-HCl (pH 7.4), 10 mM MgCl2, 5 mM ATP, 2 mM dithiothreitol) at 30 °C for 1.5 h. The reactions were stopped by adding 2× SDS loading buffer and boiled at 95 °C for 5 min. Proteins were separated by SDS–PAGE, followed by immunoblot analysis using anti-ubiquitin and anti-MBP antibodies.
To determine whether NBIP1 mediates SPX4 ubiquitination, 300 ng of purified GST–SPX4 was incubated with 50 ng of AtE1, 50 ng of human E2, 250 ng of MBP–NBIP1T and 2 μg ubiquitin in 30 μl of ubiquitination reaction buffer at 30 °C for 2 h. Reactions were stopped by adding 2× SDS loading buffer, and the protein samples were subjected to SDS–PAGE and immunoblot analysis with an anti-GST antibody.
MS analyses. Affinity purification and subsequent MS analysis were performed as described previously47,48. The 35S–eGFP and NRT1.1B–eGFP transgenic rice were cultured in basal nutrient solution supplemented with 0.25 mM (NH4)2SO4 and 1 mM KNO3 for 10 days. About 5 g of shoots from rice seedlings were harvested, ground to fine powder in liquid nitrogen and resuspended in protein extraction buffer as described above. Immunoprecipitations of GFP fusion proteins were performed as described for co-IP assay. The bound proteins were eluted from the beads with 2× SDS loading buffer by boiling at 95 °C for 3 min. Protein samples were separated by 10% SDS–PAGE and stained with Coomassie Brilliant Blue R-250. The entire lane of each sample was divided into six segments and subjected to in-gel digestion with trypsin. The peptides were analysed by liquid chromatography–tandem MS using a TripleTOF 5600 system (AB SCIEX) coupled online to an Eksigent nanoLC Ultra in information-dependent mode. The proteins were identified by searching the O. sativa database from IRGSP (http://rgp.dna.affrc.go.jp/IRGSP/) using ProteinPilot software v.4.2 with a 1% false discovery rate.
Reporting Summary. Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availabilityThe data that support the findings of this study are available from the corresponding authors upon request.
Received: 24 July 2018; Accepted: 8 February 2019; Published: xx xx xxxx
References 1. Young, V. R. Nutritional balance studies: indicators of human requirements or
of adaptive mechanisms? J. Nutr. 116, 700–703 (1986). 2. Hill, J. O. et al. Nutrient balance in humans: effects of diet composition.
Am. J. Clin. Nutr. 54, 10–17 (1991). 3. Carreiro, A. L. et al. The macronutrients, appetite, and energy intake.
Annu. Rev. Nutr. 36, 73–103 (2016). 4. Güsewell, S. N:P ratios in terrestrial plants: variation and functional
significance. New Phytol. 164, 243–266 (2004). 5. Khan, F. et al. Effect of different levels of nitrogen and phosphorus on the
phenology and yield of maize varieties. Am. J. Plant Sci. 5, 2582–2590 (2014). 6. Luo, X. et al. Nitrogen: phosphorous supply ratio and allometry in five alpine
plant species. Ecol. Evol. 6, 8881–8892 (2016). 7. Güsewell, S. Responses of wetland graminoids to the relative supply of
nitrogen and phosphorus. Plant Ecol. 176, 35–55 (2005). 8. Liu, K. H. & Tsay, Y. F. Switching between the two action modes of the
dual-affinity nitrate transporter CHL1 by phosphorylation. EMBO J. 22, 1005–1013 (2003).
9. Ho, C. H., Lin, S. H., Hu, H. C. & Tsay, Y. F. CHL1 functions as a nitrate sensor in plants. Cell 138, 1184–1194 (2009).
10. Hu, B. et al. Variation in NRT1.1B contributes to nitrate-use divergence between rice subspecies. Nat. Genet. 47, 834–838 (2015).
11. Marchive, C. et al. Nuclear retention of the transcription factor NLP7 orchestrates the early response to nitrate in plants. Nat. Commun. 4, 1713 (2013).
12. Liu, K. H. et al. Discovery of nitrate–CPK–NLP signalling in central nutrient–growth networks. Nature 545, 311–316 (2017).
13. Rubio, V. A conserved MYB transcription factor involved in phosphate starvation signaling both in vascular plants and in unicellular algae. Genes Dev. 15, 2122–2133 (2001).
14. Lv, Q. et al. SPX4 negatively regulates phosphate signaling and homeostasis through its interaction with PHR2 in rice. Plant Cell 26, 1586–1597 (2014).
15. Wild, R. et al. Control of eukaryotic phosphate homeostasis by inositol polyphosphate sensor domains. Science 352, 986–990 (2016).
16. Puga, M. I. et al. SPX1 is a phosphate-dependent inhibitor of phosphate starvation response 1 in arabidopsis. Proc. Natl Acad. Sci. USA 111, 14947–14952 (2014).
17. Wang, Z. et al. Rice SPX1 and SPX2 inhibit phosphate starvation responses through interacting with PHR2 in a phosphate-dependent manner. Proc. Natl Acad. Sci. USA 111, 14953–14958 (2014).
18. Kant, S., Peng, M. & Rothstein, S. J. Genetic regulation by NLA and microRNA827 for maintaining nitrate-dependent phosphate homeostasis in Arabidopsis. PLoS Genet. 7, e1002021 (2011).
19. Medici, A. et al. Atnigt1/hrs1 integrates nitrate and phosphate signals at the Arabidopsis root tip. Nat. Commun. 6, 6274 (2015).
20. Kiba, T. et al. Repression of nitrogen-starvation responses by members of the Arabidopsis GARP-type transcription factor NIGT1/HRS1 subfamily. Plant Cell 30, 925–945 (2018).
21. Maeda, Y. et al. A NIGT1-centred transcriptional cascade regulates nitrate signalling and incorporates phosphorus starvation signals in Arabidopsis. Nat. Commun. 9, 1376 (2018).
22. Sun, J. et al. Crystal structure of the plant dual-affinity nitrate transporter NRT1.1. Nature 507, 73–77 (2014).
23. Parker, J. L. & Newstead, S. Molecular basis of nitrate uptake by the plant nitrate transporter NRT1.1. Nature 507, 68–72 (2014).
24. Chardin, C., Girin, T., Roudier, F., Meyer, C. & Krapp, A. The plant RWP-RK transcription factors: key regulators of nitrogen responses and of gametophyte development. J. Exp. Bot. 65, 5577–5587 (2014).
25. Lopez-Arredondo, D. L., Leyva-Gonzalez, M. A., Alatorre-Cobos, F. & Herrera-Estrella, L. Biotechnology of nutrient uptake and assimilation in plants. Int. J. Dev. Biol. 57, 595–610 (2013).
26. Wang, R. et al. Genomic analysis of the nitrate response using a nitrate reductase-null mutant of Arabidopsis. Plant Physiol. 136, 2512–2522 (2004).
27. Lin, S. I. et al. Complex regulation of two target genes encoding SPX-MFS proteins by rice miR827 in response to phosphate starvation. Plant Cell Physiol. 51, 2119–2131 (2010).
28. Yue, W. et al. OsNLA1, a RING-type ubiquitin ligase, maintains phosphate homeostasis in Oryza sativa via degradation of phosphate transporters. Plant J. 90, 1040–1051 (2017).
29. Lopez-Bucio, J., Cruz-Ramirez, A. & Herrera-Estrella, L. The role of nutrient availability in regulating root architecture. Curr. Opin. Plant Biol. 6, 280–287 (2003).
30. Sánchez-Calderón, L., Chacón-López, A., Alatorre-Cobos, F., Leyva-González, M. A. & Herrera-Estrella, L. in Transporters and Pumps in Plant Signaling (eds Markus Geisler & Kees Venema) 191–224 (Springer, 2011).
31. Poitout, A. et al. Responses to systemic nitrogen signaling in Arabidopsis roots involve trans-Zeatin in shoots. Plant Cell 30, 1243–1257 (2018).
32. Takei, K. et al. AtIPT3 is a key determinant of nitrate-dependent cytokinin biosynthesis in Arabidopsis. Plant Cell Physiol. 45, 1053–1062 (2004).
33. Martin, A. C. et al. Influence of cytokinins on the expression of phosphate starvation responsive genes in Arabidopsis. Plant J. 24, 559–567 (2000).
34. Wang, X. M. et al. Cytokinin represses phosphate-starvation response through increasing of intracellular phosphate level. Plant Cell Environ. 29, 1924–1935 (2006).
35. Lu, Y. et al. Genome-wide targeted mutagenesis in rice using the CRISPR/Cas9 system. Mol. Plant 10, 1242–1245 (2017).
36. Hellens, R. P. et al. Transient expression vectors for functional genomics, quantification of promoter activity and RNA silencing in plants. Plant Methods 1, 13 (2005).
37. Chen, H. et al. Firefly luciferase complementation imaging assay for protein–protein interactions in plants. Plant Physiol. 146, 368–376 (2008).
38. Luo, A. et al. EUI1, encoding a putative cytochrome P450 monooxygenase, regulates internode elongation by modulating gibberellin responses in rice. Plant Cell Physiol. 47, 181–191 (2006).
39. Hiei, Y., Ohta, S., Komari, T. & Kumashiro, T. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence-analysis of the boundaries of the T-DNA. Plant J. 6, 271–282 (1994).
40. Wang, C. et al. Involvement of OsSPX1 in phosphate homeostasis in rice. Plant J. 57, 895–904 (2009).
41. Hu, B. et al. Leaf tip necrosis1 plays a pivotal role in the regulation of multiple phosphate starvation responses in rice. Plant Physiol. 156, 1101–1115 (2011).
42. Yoo, S. D., Cho, Y. H. & Sheen, J. Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nat. Protoc. 2, 1565–1672 (2007).
43. Zhang, Y. et al. A highly efficient rice green tissue protoplast system for transient gene expression and studying light/chloroplast-related processes. Plant Methods 7, 30 (2011).
44. Hecker, A. et al. Binary 2in1 vectors improve in planta (co)localization and dynamic protein interaction studies. Plant Physiol. 168, 776–787 (2015).
45. Zhao, Q., Liu, L. & Xie, Q. In vitro protein ubiquitination assay. Methods Mol. Biol. 876, 163–172 (2012).
46. Zhao, Q. et al. A plant‐specific in vitro ubiquitination analysis system. Plant J. 74, 524–533 (2013).
47. Wang, Z. et al. SCFSAP controls organ size by targeting PPD proteins for degradation in Arabidopsis thaliana. Nat. Commun. 7, 11192 (2016).
48. van Dongen, W., Van Heerde, L., Boeren, S. & De Vries, S. C. Identification of brassinosteroid signaling complexes by coimmunoprecipitation and mass spectrometry. Methods Mol. Biol. 1564, 145–154 (2017).
AcknowledgementspCAMBIA1300–nLUC/cLUC vectors were kindly provided by J.-M. Zhou, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences. The SPX4pro–gSPX4–GUS transgenic rice was kindly provided by C. Mao, College of Life Sciences, Zhejiang University. The authors thank T. O. Jobe (University of Cologne) for proofreading the manuscript. This work was supported by grants from the Chinese Academy of Sciences (XDA08010400), the National Natural Sciences Foundation of
China (31500975 and 31771348) and the China Postdoctoral Science Foundation (2017M610126). Research in the laboratory of S. Kopriva was supported by Deutsche Forschungsgemeinschaft (EXC 1028).
Author contributionsB.H., Z.J. and W.W. designed research, performed experiments, analysed the data and wrote the manuscript. Y. Qiu, Z.Z., Y.L., A.L., X.G., L.L., Y. Qian, X.H., F.Y., S. Kang, Yiq. W., J.X., S.C., L.Z., Ying. W., Q.X. and S. Kopriva conducted some of the experiments. C.C. designed research, wrote the manuscript and supervised the project.
Competing interestsThe authors declare no competing interests.
Additional informationSupplementary information is available for this paper at https://doi.org/10.1038/s41477-019-0384-1.
Reprints and permissions information is available at www.nature.com/reprints.
Correspondence and requests for materials should be addressed to B.H. or C.C.
Journal peer review information: Nature Plants thanks Dong Liu, Nicholaus von Wiren, Ying Liu, Ricardo Giehl and other anonymous reviewers for their contribution to the peer review of this work.
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Nitrate–NRT1.1B–SPX4 cascade integrates nitrogen and phosphorus signalling networks in plantsBin Hu 1,7*, Zhimin Jiang1,7, Wei Wang1,7, Yahong Qiu1,2,7, Zhihua Zhang1,2, Yongqiang Liu1,2, Aifu Li1,2, Xiaokai Gao1, Linchuan Liu1, Yangwen Qian3, Xiahe Huang4, Feifei Yu1, Sai Kang1, Yiqin Wang1, Junpeng Xie1,2, Shouyun Cao1, Lianhe Zhang5, Yingchun Wang4, Qi Xie1, Stanislav Kopriva 6 and Chengcai Chu 1,2*
1State Key Laboratory of Plant Genomics, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing, China. 2College of Life Sciences, University of Chinese Academy of Sciences, Beijing, China. 3Biogle Genome Editing Center, Changzhou, China. 4State Key Laboratory of Molecular Developmental Biology, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing, China. 5School of Agriculture, Henan University of Science and Technology, Luoyang, China. 6Botanical Institute, Cluster of Excellence on Plant Sciences, University of Cologne, Cologne, Germany. 7These authors contributed equally: Bin Hu, Zhimin Jiang, Wei Wang, Yahong Qiu. *e-mail: [email protected]; [email protected]
SUPPLEMENTARY INFORMATION
In the format provided by the authors and unedited.