Characterization of A Novel Groundbreaking Radioresistance Pathway in the Endometrial Cancer Stem Cell Niche: The Interplay of WNT, Cyclooxygenase-2 (COX-2), and Hypoxia Inducible Factor-1α (HIF-1α) NELLY LIN & KERCHENG CHEN
Transcript
Characterization of A Novel Groundbreaking Radioresistance Pathway in the Endometrial Cancer Stem Cell Niche: The Interplay of WNT, Cyclooxygenase-2 (COX-2), and Hypoxia Inducible Factor-1α (HIF-1α)
NELLY LIN & KERCHENG CHEN
Introduction
Issues with Current Medical Treatments for Cancer: Radiation TherapyCancer is a disease recognized for its resilience to treatment. Such resilience is displayed through cancer’s ability to evade the effects of radiation, especially in the form of patients that suffer from local reoccurrences or metastasis after treatment.
In an effort to understand radiation therapy’s failures, we have isolated several problems responsible for cancer’s radioresistance: 1. The intrinsically radioresistant nature of cancer stem
cells (CSCs)
2. A hypoxic microenvironment encompassing the CSC, which acts as a barrier against radiotherapy
3. Arrest of the CSC in the G phase of the cell cycle, inhibiting radiation’s effects on replicating DNA
4. A return to normal conditions of the cancer after radiation treatment
Cancer Stem Cells (CSCs) Cancer Stem Cells (CSCs): are rare undifferentiated cancer cells that contribute to the initiation and maintenance of malignant tissue.
Four Properties of CSCs: that allow for tumor formation, maintenance, regeneration, and survival.
1. Self-Renewal
2. Extensive Proliferation
3. Differentiation
4. Tumorigenicity
A Paradigm Shift in Stem Cell Biology: Researchers have shifted their focus from targeting cancer cells to targeting CSCs, as a result of the CSC Hypothesis.
Cancer Stem Cell Hypothesis- The mutation of stem cells, rather than the mutation of normal cells causes growth of malignant tumors.
This small subpopulation of cells is able to withstand the influence of radiation (intrinsically radioresistant).
The Cancer Stem Cell Niche:Hypoxia Diminishes Radiation Therapy’s Potency
CSCs radioresitant characteristics are exacerbated by a hypoxia microenvironment that surrounds the CSCs, which creates a barrier that protects the CSC from radiation.
Hypoxia: the absence of oxygen, greatly hinders radiotherapy’s efficacy. Oxygen is a necessary component in forming DNA damaging free radicals . Without free radicals, radiation therapy cannot eliminate the source of cancer, the CSCs.
CSC Microenvironment/Niche: an architectural construct of differentiated cells surrounding a pluripotent CSC.
• The CSC niche specializes in maintaining the tumorigenetic properties of the CSC by inducing a variety of signals.
• Studies suggest that these regulatory signals (signaling pathways) are major factors in the protection of CSCs.
Hypoxia Inducible Factor-1α (HIF-1α)
The hypoxic microenvironment is maintained through the activation of the Hypoxia Inducible Factor-1 (HIF-1), a transcription factor that controls adaptation and survival of cells to hypoxia.
Let us understand
How Radiation Therapy Works:
1. Radiation is able to break DNA molecules directly, through
molecular changes in DNA’s double-strand structure
2. Or indirectly, through the ionization of water molecules which
results in the formation of free radicals that subsequently
causes DNA breakage.
Breakages in DNA results in the death of the cancer cell.
The Interplay of WNT, Cyclooxygenase-2 (COX-2), and Hypoxia Inducible Factor-1α (HIF-1α) Signaling
PathwaysCyclooxygenase-2 (COX-2)
It has been made known that DNA is the most vulnerable to radiation while in the S phase (replication) of the cell cycle.
As such, we propose that since CSCs are radioresistant, CSCs are in the G phase of the cell cycle, a period when DNA is not replicating.
Cyclooxygenase-2 (COX-2): Recent studies have identified cyclooxygenase-2 (COX-2), to be the signal responsible for inducing cell cycle arrest (G phase).
COX-2 is also know to aid in the radiation-induced bystander effect, which allows non-irradiated cells in the proximity of irradiated cells display effects of a cell affected by radiation.
* Encourages every cell to arrest in G phase*
WNT
Clearly though, these cancerous cells do not remain arrested in the cell cycle. Unfortunately, they continue to replicate and proliferate. A missing link to this pathway is apparent.
Canonical WNT Signaling Pathway: we theorize that the Wnt signaling pathway plays a role incell cycle reentry.
COX-2 & HIF-1α
HIF-1’s α subunit was found to bind directly to an HRE located on the COX-2 promoter, suggesting that COX-2 up-regulation is dependent on HIF-1 induction.
Hypothesis:A Novel Radioresistance PathwayWe propose that upon radiation exposure CSCs up-regulate COX-2 production, which
arrests cells in the G phase to inhibit DNA replication and radiation damage.
COX-2 triggers the activation of the MAPK pathway, which leads to HIF-1 activation.
HIF-1 metabolizes oxygen and free radicals, diminishing radiation therapy’s effect.
Once all free radicals are metabolized, HIF-1 down-regulates, thus down-regulating COX-2.
The low levels of COX-2 releases the cell from arrest.
Wnt expression will allow cells cycle reentry.
Materials & Methods
Cell Culture Techniques1. Sample Acquisition: Fresh tumor samples from endometrial cancer patients
undergoing surgical procedures (radical hysterectomy) at SUNY Downstate Medical Center at Brooklyn, NY were isolated.
2. Dissociation of Tumor into Single Cell Suspensions: The tumor was minced mechanically into small 2 mm cubes with crossed scalpels. Forcing tumor chunks through stainless steel wire mesh produced a brei of tumor cells.
3. Cell Lines Used: Cultures of AG1522 normal human skin fibroblasts were also used in this study.
Hybrid Spheroid Assay4. Hybrid Spheroid Formation and Maintenance • The Hybrid Spheroid Assay (HSA), a functional assay for identifying CSCs based on stem cell
growth dynamics, was used as a culture assay. • The AG1522 normal human skin fibroblasts and the endometrial tumor cells were mixed at
a concentration of 80% fibroblast and 20% tumor cells.• Cells were plated onto 96 well Ultra-Low attachment (ULA) plates with 150 cells/well. ULA
plates selectively inhibits cellular attachment using a layer of covalently bound hydrogel.
5. Identification of HS containing EnCSCs : Spheroid growth measurements were taken to document changes in HS volume and size using the 20x HMC obj.of a Nikon microscope.
Profiling the Expression of WNT, COX-2, and HIF-1α
6. Immunofluorescence Analysis Pre-irradiation: HSs were incubated with the following antibodies: Oct-4-AF647
Post-irradiation: Separate multiwell plates of spheroids were treated to 2 Gy
doses of 250 kVp X-rays from a Philips RT 250 therapy machine. The immunofluorescence staining protocol was repeated.
Image Acquisition: Images were generated using respective excitation
wavelengths and confocal microscopy. HSs were imaged using an Olympus FluoviewFV1000 Laser Scanning Biological Microscope housed on the Olympus IX81 Inverted Microscope.
This graph shows the growth curves for HSs containing irradiated EnCSCs. Based on
the number of HSs that grew, compared to non-irradiated HSs, a survival fraction for
individual patients can be determined.
Model of Ideal EnCSC Hybrid Spheroid Growth Curves
Wnt and Fzd Expression colocalized to epithelial EnCSCs and mesenchymal EnCSCs, respectively.
A) A merge image of all the stains in the HS. B) DAPI stain to identify all the cells in the HS. C) DAPI and Nanog to identify all the epithelial EnCSCs within the niche. D)
DAPI and Oct4 to show the mesenchymal EnCSCs within the niche (colozalization of stains creates pinkish tinge). E) panK localized to same regions as Nanog to show that
Nanog+ EnCSCs are in fact of epithelial origin. F) Distribution of Fzd receptor within the niche overlaps primarily with distribution of mesenchymal EnCSCs. Some expression
noted to overlap with epithelial EnCSCs. G) Wnt production localized to Nanog+ regions. H) Wnt colocalizes with Nanog. Oct4 primarily colocalizes with Fzd.
WNT Expression in the EnCSC Niche
Wnt and Fzd Expression in the EnCSC Niche.
A) A merge image of all of the stains in the HS.
B) DAPI stain to identify all the cells in the HS.
C) Nanog with DAPI to identify epithelial EnCSCs in the EnCSC niche.
D)Oct4 stain with Nanog stain to distinguish between mesenchymal and epithelial
EnCSCs.
E1) Oct4 with Nanog and panK to further distinguish between epithelial and
mesenchymal EnCSCs.
E2) Nanog with Fzd to show distribution of Fzd receptor in the niche cells.
F) Wnt and Fzd to show regions of Wnt expression and Fzd expression within the niche.
G)Colocalization of Nanog and Wnt expression.
H) Colocalization of Oct4 and Fzd expression
WNT Expression in the EnCSC Niche
Characterization of Wnt Expression within the EnCSC Niche. The
epithelial EnCSC are responsible for Wnt production and secretion within the
niche. The primary targets for Wnt are the mesenchymal EnCSCs which bind
Wnt via the Fzd receptor through paracrine signaling. The presence of Fzd
receptor on epithelial EnCSCs suggests that they themselves are a target of
Wnt signaling as well through autocrine signaling.
Paracrine and Autocrine Signaling in the EnCSC Niche of
WNT Expression
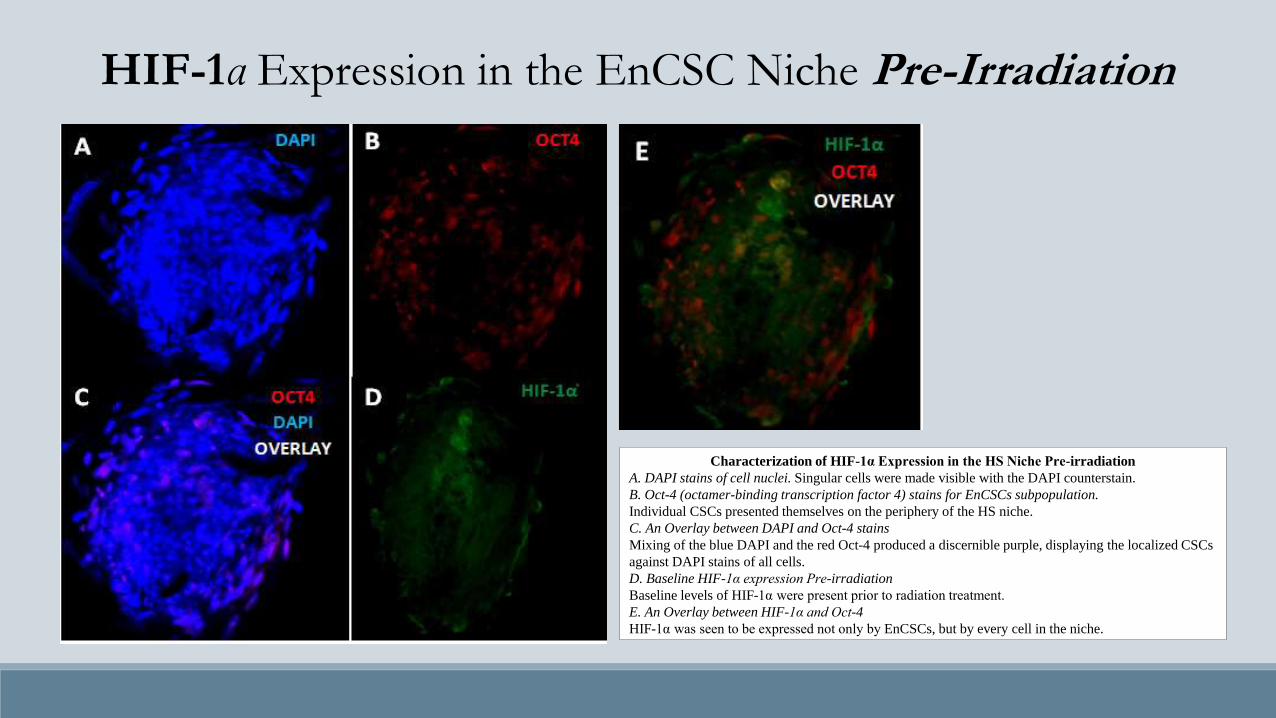
Characterization of HIF-1α Expression in the HS Niche Pre-irradiation
A. DAPI stains of cell nuclei. Singular cells were made visible with the DAPI counterstain.
B. Oct-4 (octamer-binding transcription factor 4) stains for EnCSCs subpopulation.
Individual CSCs presented themselves on the periphery of the HS niche.
C. An Overlay between DAPI and Oct-4 stains
Mixing of the blue DAPI and the red Oct-4 produced a discernible purple, displaying the localized CSCs
against DAPI stains of all cells.
D. Baseline HIF-1α expression Pre-irradiation
Baseline levels of HIF-1α were present prior to radiation treatment.
E. An Overlay between HIF-1α and Oct-4
HIF-1α was seen to be expressed not only by EnCSCs, but by every cell in the niche.
HIF-1α Expression in the EnCSC Niche Pre-Irradiation
Characterization of COX-2 Expression in the HS Niche Pre-irradiation
A. Novel image of the EnCSC Niche. Merged image showing DAPI, Oct-4, panK, HIF-1α and COX-2 stains. B. An
Overlay between DAPI and Oct-4 stains. Oct-4 expression lies on an outer ring of the niche, suggesting the location of
EnCSCs to be on the outer edge of the HS. C. An Overlay between DAPI and panK stains. PanKeratin was centralized
in the niche, staining for differentiated epithelial cells.
COX-2 Expression in the EnCSC NichePre-Irradiation
Characterization of COX-2 Expression in the HS Niche Post-irradiation
A. DAPI stains of cell nuclei. Singular cells are made visible with the DAPI
counterstain.
B. An Overlay between panK and Oct-4 stains: on one hand, panK (panKeratin) stains
for EnCSCs’ differentiated epithelial progeny, on the other, Oct-4 (octamer-binding
transcription factor 4) stains for EnCSCs subpopulation. EnCSCs was seen to encircle
its differentiated progeny, residing the periphery of the HS niche. Epithelial
endometrial cells lay localized in the center of the HS niche.
C. Expression of COX-2 Post-Irradiation.
COX-2 was up-regulated several hours after initial exposure to 2Gy of radiation
D. An Overlay between panK, Oct-4, and COX-2 stains. COX-2 was recognized to be
expressed in both CSCs and progeny cancer cells.
COX-2 Expression in the EnCSC NichePost-Irradiation
COX-2 Expression in the EnCSC Niche Linked to Epithelial EnCSCs Post-irradiation
A. An overlay image showing various expression profiles within the HS.
B. Stains of DAPI & NANOG
DAPI (blue) identifies the cells within this HS. NANOG (cyan) identifies the presence of
epithelial EnCSCs within the niche.
C. Mesenchymal vs. Epithelial EnCSCs
The image shows the location of Oct-4+ mesenchymal EnCSCs and NANOG+ epithelial
EnCSCs within the niche.
D. Vimentin staining
Vimentin staining to show that peripheral Oct-4+ CSCs are of mesenchymal origin.
E. panK staining
panK+ epithelial cells confirms that NANOG+ cells are of epithelial origin.
F. COX-2 expression post-irradiation in correlation with NANOG+ expression.
Characterization of HIF-1α and COX-2 expression Post-irradiation
A. Merged image of all the expression profiles in the HS.
B. Stain of Oct-4+ mesenchymal EnCSCs & NANOG+ epithelial
EnCSCs.
C. COX-2 expression post-irradiation in the lower hemisphere of the
HS.
D. HIF-1 expression in the lower hemisphere of the HS, as well as near
the middle, with minor expression at the top right of the HS.
E. Interplay between COX-2 expression and NANOG expression
F. Interplay between HIF-1 expression and NANOG expression.
HIF-1α & COX-2 Expression in the EnCSC Niche Post-Irradiation
Discussion
The Core Circuitry Behind Radioresistance
• Basal levels of HIF-1α present within EnCSCs serve as a natural protection mechanism against damaging effects of radiation exposure by limiting the production of free radicals in the intracellular environment, thereby reducing the potentially damaging consequences of radiation-induced free radical production which can amplify DNA double strand breaks.
• Upon exposure to radiation, the CSCs up-regulate COX-2.
• COX-2 arrests the CSC in the cell cycle, thereby making it non-proliferative and increasing its radioresistance. By arresting the cell cycle, COX-2 ensures that the CSC’s DNA is no longer susceptible to radiation induced damage.
• Extracellular secretion of COX-2 in CSC cross-talk within the niche ensures that all other EnCSCs within the niche are also arrested during the cell cycle.
• COX-2 up-regulation and the accumulation of its downstream effector, prostaglandin E2 (PGE2) leads to an up-regulation of HIF-1α which promotes various outcomes.
• HIF-1α acts as a free radical scavenger and eliminates any free radicals that may exist in order to minimize the effects of radiation exposure to the CSC's DNA.
• Up-regulation of PGE2 and HIF-1α ultimately lead to MAPK/ERK signaling up-regulation which leads to increased survival and proliferation of the EnCSCs.
• Once HIF-1α metabolizes all free radicals, its expression is down regulated followed by down regulation of COX-2 which releases the cell from arrest. Release from arrest allows for Wnt expression to be reactivated thus causing cell cycle reentry and minimizing possible DNA damage.
Limitations & Future Research• Studies will be done to accurately measure levels of expression by measuring mRNA
expression of Wnt, COX-2, and HIF-1α to gain a better understanding of the expression profiles, something we were not able to do in this study.
• Potential studies will also be focused on designing methods to inhibit Wnt, COX-2, and HIF-1α in order to make the EnCSCs radiosensitive, and to potentially take these new treatments to clinical trials.
• The study was limited by not being able to test out the hypothesis using COX-2 inhibitors in mice.
Significance• Discovery of this pathway enables future designs to eliminating
radioresistance through the pathway’s manipulation, thus altering the efficacy of radiation therapy to potentially save countless lives of patients with resistant metastasis and secondary reoccurrences.
• Each year thousands of patients fail treatments because of the inherent radioresistance of their CSCs. If this pathway can be targeted and the resistant CSCs can be made sensitive, we have a chance at saving or prolonging numerous lives.
References1. Diehn, M., Cho, R. W., Lobo, N. A., Kalisky, T., Dorie, M. J., Kulp, A. N., ... & Clarke, M. F. (2009).
Association of reactive oxygen species levels and radioresistance in cancer stem cells. nature, 458(7239), 780-783.
2. Wicha, M. S., Liu, S., & Dontu, G. (2006). Cancer stem cells: an old idea—a paradigm shift. Cancer research, 66(4), 1883-1890.
3. Overgaard, J., & Horsman, M. R. (1996, January). Modification of hypoxia-induced radioresistance in tumors by the use of oxygen and sensitizers. InSeminars in radiation oncology (Vol. 6, No. 1, pp. 10-21). WB Saunders.
4. Zhou, H., Ivanov, V. N., Gillespie, J., Geard, C. R., Amundson, S. A., Brenner, D. J., ... & Hei, T. K. (2005). Mechanism of radiation-induced bystander effect: role of the cyclooxygenase-2 signaling pathway. Proceedings of the National Academy of Sciences of the United States of America, 102(41), 14641-14646.
5. Mothersill, C., & Seymour, C. (2001). Radiation-induced bystander effects: past history and future directions. Radiation research, 155(6), 759-767.
7. Greenhough, A., Smartt, H. J., Moore, A. E., Roberts, H. R., Williams, A. C., Paraskeva, C., & Kaidi, A. (2009). The COX-2/PGE2 pathway: key roles in the hallmarks of cancer and adaptation to the tumour
8. Djordjevic, B., & Lange, C. S. (2006). Cell-cell interactions in spheroids maintained in suspension. Acta Oncologica, 45(4), 412-420.
9. Hugo, H., Ackland, M. L., Blick, T., Lawrence, M. G., Clements, J. A., Williams, E. D., & Thompson, E. W. (2007). Epithelial—mesenchymal and mesenchymal—epithelial transitions in carcinoma progression. Journal of cellular physiology, 213(2), 374-383.
10. Nagy, M. A. (2010). HIF-1 is the Commander of Gateways to Cancer. Journal of Cancer Science & Therapy.
11. Sakanaka, C., Sun, T. Q., & Williams, L. T. (1999). New steps in the Wnt/beta-catenin signal transduction pathway. Recent progress in hormone research, 55, 225-236.
12. Leung, J. Y., Kolligs, F. T., Wu, R., Zhai, Y., Kuick, R., Hanash, S., ... & Fearon, E. R. (2002). Activation of AXIN2 Expression by β-Catenin-T Cell Factor A FEEDBACK REPRESSOR PATHWAY REGULATING Wnt SIGNALING. Journal of Biological Chemistry, 277(24), 21657-21665.
13. Araki, Y., Okamura, S., Hussain, S. P., Nagashima, M., He, P., Shiseki, M., ... & Harris, C. C. (2003). Regulation of cyclooxygenase-2 expression by the Wnt and ras pathways. Cancer research, 63(3), 728-734.
14. Yang, W., Yan, H. X., Chen, L., Liu, Q., He, Y. Q., Yu, L. X., ... & Wang, H. Y. (2008). Wnt/β-catenin signaling contributes to activation of normal and tumorigenic liver progenitor cells. Cancer research, 68(11), 4287-4295.
15. Komiya, Y., & Habas, R. (2008). Wnt signal transduction pathways. Organogenesis, 4(2), 68.
16. Ward, J. F. (1988). DNA damage produced by ionizing radiation in mammalian cells: identities, mechanisms of formation, and reparability. Progress in nucleic acid research and molecular biology, 35, 95-125.
17. Rosen, E. M., Fan, S., Goldberg, I. D., & Rockwell, S. (2000). Biological basis of radiation sensitivity. Part 2: Cellular and molecular determinants of radiosensitivity. Oncology (Williston Park, NY), 14(5), 741-57. Hugo, H., Ackland, M. L., Blick, T., Lawrence, M. G., Clements, J. A., Williams, E. D., & Thompson, E. W. (2007). Epithelial—mesenchymal and mesenchymal—epithelial transitions in carcinoma progression. Journal of cellular physiology, 213(2), 374-383.