Novel Interactions of CAPS (Ca2�-dependent ActivatorProtein for Secretion) with the Three Neuronal SNAREProteins Required for Vesicle Fusion*
Received for publication, May 14, 2010, and in revised form, August 16, 2010 Published, JBC Papers in Press, September 8, 2010, DOI 10.1074/jbc.M110.145169
Neil J. Daily, Kristin L. Boswell, Declan J. James, and Thomas F. J. Martin1
From the Department of Biochemistry, University of Wisconsin-Madison, Madison, Wisconsin 53706
CAPS (aka CADPS) is required for optimal vesicle exocytosisin neurons and endocrine cells where it functions to primethe exocytic machinery for Ca2�-triggered fusion. Fusionis mediated by trans complexes of the SNARE proteinsVAMP-2, syntaxin-1, and SNAP-25 that bridge vesicle andplasma membrane. CAPS promotes SNARE complex forma-tion on liposomes, but the SNARE binding properties ofCAPS are unknown. The current work revealed that CAPSexhibits high affinity binding to syntaxin-1 and SNAP-25 andmoderate affinity binding to VAMP-2. CAPS binding is spe-cific for a subset of exocytic SNARE protein isoforms andrequires membrane integration of the SNARE proteins.SNARE protein binding by CAPS is novel and mediated byinteractions with the SNAREmotifs in the three proteins. TheC-terminal site for CAPS binding on syntaxin-1 does notoverlap the Munc18-1 binding site and both proteins can co-reside on membrane-integrated syntaxin-1. As expected fora C-terminal binding site on syntaxin-1, CAPS stimulatesSNARE-dependent liposome fusion with N-terminal trun-cated syntaxin-1 but exhibits impaired activity with C-termi-nal syntaxin-1 mutants. Overall the results suggest thatSNARE complex formation promoted by CAPS may be medi-ated by direct interactions of CAPS with each of the threeSNARE proteins required for vesicle exocytosis.
Peptide and neurotransmitter release from endocrine cellsand neurons occurs by the regulated exocytic fusion of secre-tory vesicles with the plasma membrane (1). Membrane fusionis mediated by trans complexes of SNARE proteins that bridgethe vesicle and plasma membrane to promote close membraneapposition and bilayer mixing (2). The SNARE complexes forendocrine and neuronal vesicle fusion consist of parallel bun-dles of four �-helical SNAREmotifs with one helix contributedby the vesicle v-SNARE vesicle-associatedmembrane protein 2(VAMP-2)2 (aka synaptobrevin), one helix contributed by the
plasma membrane t-SNARE syntaxin-1, and two helices con-tributed by the plasma membrane t-SNARE SNAP-25 (3).Based on a central layer residue (arginine or glutamine) andhelix position in the SNARE bundle, these proteins are alsoclassified as R-, Qa-, and QbQc-SNAREs, respectively (4). Invitro studies suggested a “zipper” model for the assembly ofternary SNARE complexes involving the pairing of syntaxin-1(Qa) with SNAP-25 (QbQc) followed by the N- to C-terminalinsertion of the R-SNARE VAMP-2 (5, 6).Vesicle exocytosis is a sequential multi-step process that
involves the initial tethering of vesicles to the plasma mem-brane (7). Only a subset of membrane-attached vesicles rapidlyfuse in response to Ca2� elevations and these are considered tobe primed in steps that render the exocytic machinery fusion-competent (8). Priming involves the disassembly of cis SNAREcomplexes and the assembly of loose trans SNARE complexesin advance of fusion triggering (9–11). The in vivo pathway fortrans SNARE complex assembly is unknown but may involvethe zippering of VAMP-2 into syntaxin-1/SNAP-25 het-erodimers as suggested from in vitro studies (12).The assembly of SNARE complexes is catalyzed by essential
accessory factors. At intracellular sites of membrane fusion inthe secretory pathway, SNARE complex assembly is catalyzedby Sec-1/Munc18 proteins acting in concert with a heteroge-neous class of tethering factors that directly bind SNAREproteins(13, 14). For regulated vesicle exocytosis, Munc18-1 partici-pates in promoting SNARE complex assembly (15) with otheraccessory proteins. Genetic and biochemical studies indicatethat proteins in theMunc13/CAPS family function in the prim-ing of vesicles in endocrine and neural cells (8). CAPS-1 is amulti-domain 1289 residue protein that reconstitutes regulatedvesicle exocytosis in permeable neuroendocrine cells (16).Regions in the C-terminal half of CAPS exhibit homology toa corresponding C-terminal region of Munc13-1 that wasreported to bind syntaxin-1 (17, 18). CAPS contains a centralpleckstrin homology (PH) domain that interacts with phospha-tidylinositol 4,5-bisphosphate (PI(4,5)P2) (19), a lipid that isrequired for priming in exocytosis (20, 21). CAPS markedlyaccelerates SNARE-dependent liposome fusion in vitro whenPI(4,5)P2 is present in the syntaxin-1/SNAP-25-containingacceptor liposomes,which is dependent on the PHdomain (22).The stimulation of liposome fusion by CAPS is preceded by the
* This work was supported, in whole or in part, by National Institutes of HealthGrant DK040428 (to T. F. J. M.) and by an AHA fellowship (to D. J. J.).
1 To whom correspondence should be addressed: Dept. of Biochemistry, Uni-versity of Wisconsin, 433 Babcock Drive, Madison, WI 53706. Tel.: 608-263-2427; E-mail: [email protected].
2 The abbreviations used are: VAMP-2, vesicle-associated membrane pro-tein 2; CAPS(aka CADPS), Ca2�-dependent activator protein for secre-tion; SNAP-25, synaptosome-associated protein of 25 kDa; �-SNAP,�-soluble N-ethylmaleimide-sensitive factor attachment protein; SPR,surface plasmon resonance; MPB-PE, 1,2-dioleoyl-sn-glycero-3-phos-phoethanolamine-N-[4-(p-maleimidophenyl)butyramide]; NBD-PE, N-
(7-nitro-2–1,3-benzoxadiazol-4-yl)-1,2-dipalmitoyl phosphatidyleth-anolamine; Rh-PE, N-(lissamine rhodamine B sulfonyl)-1,2-dipalmitoylphosphatidylethanolamine; pf, protein-free; PC, phosphatidylcholine; PS,phosphatidylserine; PI 4,5-P2, phosphatidylinositol-4,5-bisphosphate.
increased assembly of trans SNARE complexes on the lipo-somes (23).A potential mechanism by which CAPS could promote
SNARE complex formation would be through direct SNAREprotein binding, however the SNARE binding properties ofCAPS are unknown. The current work reveals that CAPS bindsindependently to each of the three SNAREproteins required forvesicle exocytosis. CAPS binds to the SNARE motifs in each ofthe SNARE proteins and exhibits specificity for a subset of exo-cytic SNARE protein isoforms. The functional significance ofCAPS interactionswith the SNARE (plus linker) domain of syn-taxin-1 was shown in liposome fusion studies. We suggest thatCAPS promotes the priming of vesicle exocytosis by drivingtrans SNARE complex formation through direct interactionswith three SNARE motifs.
EXPERIMENTAL PROCEDURES
Materials—1,2-dioleoyl phosphatidylcholine (DOPC), 1,2-dioleoyl phosphatidylserine (DOPS), N-(7-nitro-2–1,3-benzoxadiazol-4-yl)-1,2-dipalmitoyl phosphatidylethanolamine(NBD-PE), N-(lissamine rhodamine B sulfonyl)-1,2-dipalmitoylphosphatidylethanolamine (Rh-PE), L-�-phosphatidylinositol-4,5-bisphosphate (PI 4,5-P2), and 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine-N-[4-(p-maleimidophenyl)butyramide](MPB-PE) were purchased fromAvanti Polar Lipids, Inc.Plasmids, Recombinant Protein Expression, and Purification—
Plasmid constructs pTW34 to coexpress rat syntaxin-1A plusN-terminally His-tagged mouse SNAP-25B and pTW2 toexpress C-terminal His-tagged mouse VAMP-2 were kindlyprovided by J.E. Rothman and T.Weber. Proteins were purifiedas described previously (2). Plasmid constructs pET28a toexpress C-terminally His-tagged syntaxin-1A-(183–288) andpET10 to express C-terminally His-tagged VAMP-2-(1–94)were generously provided by E. R. Chapman. Syntaxin-(183–288) was purified as described previously (24). pGEX-KG toexpress syntaxin-1A-(191–266) and constructs to express syn-taxins -3, -4, -5, and -6 were kindly provided by R. H. Scheller.pGEX-2T plasmids encoding GST-rat syntaxin-1A-(1–264),GST-rat syntaxin-1A-(1–177), GST-mouse SNAP-25B, GST-SNAP-25A-(1–100), GST-SNAP-25B-(101–206), GST-Munc-18-1, and pGEX4T1 plasmids encoding N-terminally GST-tagged rat syntaxin-1, -2, -3, -4, -6, and syntaxin H3–1A, -3, -4,-5, -6 were used to express proteins for purification by glutathi-one-agarose chromatography (25) with elution by thrombincleavage for 4 h at room temperature in reconstitution bufferwithout glycerol (25 mM HEPES-KOH pH 7.4, 100 mM KCl, 1mM DTT) followed by neutralization with 2 mM PMSF orAEBSF. pET10 plasmids encoding His-tagged VAMP-2-(1–94)and pET28a plasmids encoding His-tagged complexin wereused to express and purify proteins by Ni-NTA chromatogra-phy with elution into reconstitution buffer without glycerol.His-tagged �-SNAP in plasmid pQE9 (Qiagen) was expressedin Escherichia coli JM109 cells and purified by Ni-NTA chro-matography. Transmembrane-containing proteins were elutedinto buffer containing 1% octylglucoside (OG). Recombinantproteins that were cross-linked to MPB-PE were eluted intobuffers in which 1 mM DTT was substituted with 1 mM TCEP.All plasmids were transformed into E. coli BL-21(DE3) cells for
protein expression and purification. C-terminally Myc-His-tagged rat CAPS protein was produced in Sf9 insect cells or byHEK293 cell expression as described previously (19, 26).Preparation of Proteoliposomes—For liposome fusion assays,
proteoliposomes were formed by co-micellization in the pres-ence of either VAMP-2, co-expressed syntaxin-1 and SNAP-25B or syntaxin-(183–288) and SNAP-25B as previouslydescribed (2, 22). A lipid film containing 1.5 �mol of DOPC:DOPS in an 85:15 mole ratio was resuspended with 500 �lof SNARE proteins in elution buffer (25 mM HEPES-KOH pH7.4, 400 mM KCl, 500 mM imidazole-OAc, pH 7.4, 10% glycerol(w/v), 1.0% �-octylglucoside). For liposomes containing syn-taxin-1-(183–288)/SNAP-25B, proteins were produced sepa-rately but preincubated for 1 h on ice to allow heterodimerformation. Proteoliposomes made under these conditions had�40 copies/liposome of syntaxin/SNAP-25. For liposomes thatcontained 5mol%PI 4,5-P2, we usedDOPC:PI 4,5-P2mol ratiosof 95:5. For fluorescent donor liposomes, we incorporated�100 copies of VAMP-2 per liposome corresponding to synap-tic vesicle densities (27). The dried lipid film containing aDOPC:DOPS:Rh-PE:NBD-PE lipid mix in an 82:15:1.5:1.5 molratio was resuspended with 500 �l of 280 �g/ml VAMP-2 pro-tein diluted in elution buffer. All lipidmixtures contained 2�Ciof [3H]-1,2-dipalmitoyl phosphatidylcholine (NEN, Cam-bridge, UK) to enable determination of lipid recoveries andstandardization of fusion reactions. Proteoliposomes were dia-lyzed overnight at 4 °C in reconstitution buffer and stirringwithBio-beads� (Bio-Rad), mixed with an equal volume of 80%Nycodenz� (a.k.a. Accudenz�) (Accurate Chemical and Scien-tific Corp.), overlaidwith 30 and 0%Accudenz in reconstitutionbuffer without glycerol, and centrifuged at 45,000 rpm for 4 h at4 °C using an SW 50.1 rotor (Beckman Corp). Proteoliposomes(400 �l) at the 0–30% Accudenz interface were collected, ali-quoted, and flash frozen. The liposome SNARE content wasassessed by SDS-PAGE and visualized byCoomassie Blue stain-ing using BSA standards for comparison.Proteoliposomes were also formed by the extrusion method
(28). A lipid film containing 4.5 �mol of DOPC:DOPS in an85:15 mole ratio was resuspended in 300 �l of reconstitutionbuffer. The mixture was vortexed for 5 min, subjected to 5freeze-thaw cycles and extruded (MiniExtruder, Avanti PolarLipids) for 33 passes through a 0.1 �m Nuclepore� track-etched polycarbonate membrane filter (Whatman) to produce100 nm diameter proteoliposomes. The mixture was dividedinto 100-�l aliquots, mixed with 200 �l of SNARE proteins inelution buffer, incubated for 30 min at room temperature, anddiluted with 300 �l reconstitution buffer. The liposome mix-ture was dialyzed and separated by Accudenz gradient centri-fugation as described above. Proteoliposomes made by extru-sion had �160 copies/liposome of syntaxin/SNAP-25 or �360copies/liposome of VAMP-2.For cross-linking SNAP-25 to liposomes, extruded lipo-
somes were made as described above with the following modi-fications. Lipid mixtures contained a DOPC:DOPS:MPB-PE(Avanti Polar Lipids) lipid mix with a 80:15:5 mole ratio.MPB-PE is a lipid moiety which contains a maleimido groupthat reacts with the thiol group of cysteines to form a stablethioether bond as described (29). MPB-PE was cross-linked to
CAPS Interacts with Membrane-anchored SNARE Proteins
NOVEMBER 12, 2010 • VOLUME 285 • NUMBER 46 JOURNAL OF BIOLOGICAL CHEMISTRY 35321
the native cysteines of SNAP-25-(1–206) and -(1–100) and aS115Cmutationwas used for SNAP-25-(101–206). 1mMTCEPwas substituted for 1 mM DTT in buffers used before cross-linking steps. SNAP-25 proteins were incubated with the lipo-somemixture for 1 h at room temperature and quenchedwith 1mM �-mercaptoethanol for 10 min at room temperature. Thesyntaxin H3-(191–266)-Cys was cross-linked to liposomes in asimilar manner.Liposome Binding Assay—0.45 mM SNARE-containing lipo-
somes were resuspended in reconstitution buffer without glyc-erol for a total volume of 75 �l. CAPS protein was added atconcentrations indicated in figure legends. After mixing, thereactions were incubated at room temperature for 30 min andthen mixed with an equal volume of 80% (w/v) Accudenz andpipetted into the bottom of a Beckman Ultra-ClearTM centri-fuge tube (5� 41mm). Themixture was overlaid with 350�l of30% (w/v) Accudenz, and reconstitution buffer without glyc-erol (20 �l) was pipetted on top of the density gradient. Thegradients were centrifuged at 45,000 rpm using a SW50.1 rotor(Beckman) for 1–4 h at 4 °C (maximum liposome recovery wasachieved within 30 min of centrifugation). The top of the gra-dient 40–60�l was collected and 10,000 DPMof the recoveredliposomes were run on SDS-PAGE and analyzed by SYPRO�Ruby (Invitrogen) or Western blotting. Western blots wereprobed with CAPS antibody, syntaxin-1 antibodies (HPC-1,Sigma-Aldrich; R31 from R. Jahn), VAMP-2 antibody (from R.Jahn), SNAP-25 antibody (polyclonal, Life Span Biosciences,Seattle, WA; monoclonal SMI-81, Covance, Madison, WI).Gels stained with SYPRO� Ruby were analyzed on a Typhoon9410 (Amersham Biosciences). For experiments displaying allfractions, 100 �l of reconstitution buffer without glycerol wasoverlaid on top of the gradient and the gradients were dividedinto 8 or 10 equal samples of 60 or 75 �l after centrifugation.Lipid Mixing Fusion Assay—Lipid mixing fusion assays
between fluorescent donor liposomes and non-fluorescentacceptor liposomes was performed as described (2). Lipid mix-ing was reported by the loss of FRET between fluorescent lipids(NBD-PE and Rh-PE) in VAMP-2 donor liposomes that occursupon fusion with the non-fluorescent t-SNARE acceptor lipo-somes. The standard assay used 0.45 mM acceptor and 0.225mM donor liposomes in a total volume of 75 �l reconstitutionbuffer without glycerol. CAPS protein was added at a final con-centration of 1�M.Negative controls were prepared for all con-ditions by substituting t-SNARE acceptor liposomes with pro-tein-free (pf) liposomes to detect non-SNARE mediated lipid-mixing. Reactions were assembled on ice and flick-mixedbefore addition to 96 well FluoroNunc plates (Nunc). Fusionwas detected by measuring dequenching of NBD fluorescence(excitation, 465 nm; emission, 535 nm) every 90 s at 37 °C in aTecan Infinite F500 microplate spectrofluorometer (Tecan,Mannedorf, Switzerland). After 2 h, fusion reactions were sol-ubilized with 0.5% w/v dodecyl-D-maltoside and fluorescencereadings were recorded for an additional 10 min. Lipid mixingresults were quantified as previously described (22).Surface Plasmon Resonance—Surface plasmon resonance
was monitored with a BIACORE 2000 (Pharmacia BiosensorAB, Uppsala, Sweden) on aNTA sensor chip (Biacore AB, Upp-sala, Sweden). The sensor chip has a carboxymethylated dex-
tran matrix pre-immobilized with NTA (Qiagen). His-taggedCAPS was immobilized via Ni2�-NTA chelation, and proteinswere examined for binding by flowing over the chip at a con-centration of 20 �M.
RESULTS
CAPS-1 Binds Membrane-integrated Syntaxin-1—CAPS-1accelerates SNARE-dependent fusion between donor VAMP-2-containing and acceptor syntaxin-1/SNAP-25-containingliposomes (23). Fusion stimulated by CAPS is preceded by theformation of trans SNARE complexes that bridge the liposomes(23). These results suggested that CAPS may interact with oneor more of the SNARE proteins but our previous studies failedto detect interactions of CAPS with truncated soluble SNAREproteins. It is increasingly apparent that full-lengthmembrane-integrated SNARE proteins exhibit properties different fromtruncated soluble SNAREproteins (30, 31). Thus, in the currentwork, we employed full-length membrane-integrated SNAREproteins. The SNARE proteins were reconstituted into PC/PSliposomes by dialysis or by direct detergent dilution methods(see “Experimental Procedures”) at densities similar to thosefound in PC12 cells (23). There was stable incorporation of theSNARE proteins into liposomes by either method as indicatedby their resistance to extraction by 0.1 M sodium bicarbonate,pH 11 buffer.
FIGURE 1. CAPS binds syntaxin-1-containing liposomes with high affin-ity. A, schematic diagram of the liposome binding assay by buoyant densitygradient centrifugation. B, 0.5 �M CAPS was incubated with protein-free orsyntaxin-1-containing PC/PS liposomes (250 nM syntaxin) and bound CAPS(fractions 1–3) was separated from free CAPS (fractions 6 – 8) by gradient cen-trifugation. Fractions were analyzed by immunoblotting for CAPS. Represen-tative results for three experiments are shown. C, indicated concentrations ofCAPS were incubated with syntaxin-containing PC liposomes formed by co-micellization (27– 40 copies syntaxin/liposome) and bound CAPS was ana-lyzed by immunoblotting. A non-linear regression curve fit to the data(mean � S.E., n � 3) indicated Kd � 220 nM and Bmax � 0.57. CAPS binding toprotein-free liposomes was negligible.
CAPS Interacts with Membrane-anchored SNARE Proteins
35322 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 46 • NOVEMBER 12, 2010
CAPS binding to proteoliposomes was analyzed by separat-ing liposome-bound from free CAPS by buoyant density flota-tion (Fig. 1A). CAPS bound to full-length syntaxin-1-contain-ing liposomes whereas little binding to protein-free liposomeswas detected at these liposome concentrations (Fig. 1B).Because the dissociation of bound CAPS was minimal duringthe liposome flotation, wewere able to derive equilibriumbind-ing constants. Binding isotherms indicated that CAPS bound tosyntaxin-1 with a Kd � 220 nM and a Bmax � 0.6 mol of CAPSpermol syntaxin-1 (Fig. 1C). Thus, CAPS exhibited high affinityinteractions with membrane-integrated syntaxin-1.Syntaxin-1 is comprised of several domains (Fig. 2A) includ-
ing an N-terminal triple helical domain (Habc), a helicalSNARE domain (H3), and a C-terminal transmembranedomain (TMD). Munc13-1, which exhibits limited sequencehomology to CAPS, was reported to bind the N-terminaldomain of syntaxin-1 (17). By contrast, we found that CAPSinteracted with C-terminal domains in syntaxin-1. CAPSbound to a membrane-integrated syntaxin-(183–288) proteinthat contained the H3, membrane-proximal linker and TMDdomains but lacked N-terminal domains (Fig. 2B). We alsodeterminedwhether the TMDparticipated in CAPS binding bycross-linking an H3-linker-containing protein, syntaxin-(191–
266)-Cys lacking the TMD, to lipo-somes via a maleimide PE lipidanchor. CAPS was found to bindequally well to the cross-linkedH3-linker protein and to the mem-brane-integrated H3-linker protein(Fig. 2C). These results revealed thatthe H3 plus membrane-proximallinker domain comprised the bind-ing site on syntaxin-1 for CAPS.To independently verify CAPS
binding to the H3-linker domain,we conducted surface plasmon res-onance (SPR) studies utilizingimmobilized CAPS and soluble syn-taxin-1 protein fragments. Theresults showed that CAPS inter-acted with soluble C-terminalsyntaxin-(191–266) but not withsoluble N-terminal syntaxin-(1–177) (Fig. 2D). CAPS also failed tointeract with the soluble cytoplas-mic domain syntaxin-(1–264) likelybecause it is in a “closed” conforma-tion that occludes interactions withtheH3domain (32). The SPR resultsconfirmed that CAPS interactswith the H3-linker domain of syn-taxin-1. However, very high (20�M) syntaxin fragment concentra-tions were needed to detect bind-ing and we were unable to obtainbinding constants because of highbackgrounds. Nonetheless, it ap-peared that CAPS exhibits very
low affinity interactions with a soluble syntaxin H3-linkerdomain protein. This contrasts with the much higher affinityinteraction of CAPS with membrane-integrated H3-linkerand indicates that membrane is an important co-determi-nant for CAPS binding to syntaxin-1. PI(4,5)P2 is essentialfor the CAPS promotion of trans SNARE complex formationand SNARE-dependent liposome fusion (22, 23). However,CAPS binding to syntaxin-1 was not dependent on PI(4,5)P2inclusion in the liposomes.To determinewhether CAPS interactions with theH3-linker
domain of syntaxin-1 are sufficient to promote fusion, we gen-erated acceptor liposomes that contained a truncated syntaxin-(183–288) that lacks the N-terminal domain. This was recon-stituted with SNAP-25 in PI(4,5)P2-containing liposomes tocompare with liposomes containing full-length syntaxin plusSNAP-25 with PI(4,5)P2 in a liposome fusion assay (Fig. 2E).The results showed that CAPS was able to drive fusion ofVAMP-2-containing donor liposomes with syntaxin(183–288)/SNAP-25-containing acceptor liposomes (Fig. 2F). Thestudies of this section identify the H3-linker domain of syn-taxin-1 as the binding site for CAPS and indicate that CAPSactivity in SNARE-dependent liposome fusion does not requirethe N-terminal domain of syntaxin-1.
FIGURE 2. CAPS binds the C-terminal H3 plus linker domain of syntaxin-1. A, diagram showing syntaxin-1-(1–288) and syntaxin-1-(183–288) with domains. B, 1 �M CAPS was incubated with protein-free, syntaxin-(1–288)- or syntaxin-(183–288)-containing PC/PS liposomes. Bound was separated from free CAPS by gradientcentrifugation and detected by immunoblotting with CAPS antibody (representative of three experiments).C, 1 �M CAPS was incubated with protein-free (85:15 PC/PS or 80:15:5 PC/PS/MPB-PE) liposomes or withsyntaxin-(183–288) or syntaxin-(191–266-Cys)-containing liposomes. Bound fractions were analyzed by immu-noblotting for CAPS (representative of three experiments). D, CAPS was immobilized to a surface plasmonresonance Ni2�-NTA chip and syntaxin-1 fragments were tested for binding by flowing over the chip at 20 �M
(mean � S.E., n � 3). E, liposomes containing syntaxin-(183–288)/SNAP-25 or syntaxin-(1–288)/SNAP-25 com-posed of the indicated lipids were run on SDS-PAGE and stained with Coomassie G-250. F, liposome fusionassays were conducted with a common set of donor VAMP-2-containing liposomes and with acceptor PC/PI4,5-P2 liposomes consisting of: protein-free (x), syntaxin-(1–288)/SNAP-25 (�/f), or syntaxin-(183–288)/SNAP-25 (E/F). Incubations were at 37 °C in the presence (f, F) or absence (�, E) of 1 �M CAPS.
CAPS Interacts with Membrane-anchored SNARE Proteins
NOVEMBER 12, 2010 • VOLUME 285 • NUMBER 46 JOURNAL OF BIOLOGICAL CHEMISTRY 35323
Syntaxin Isoform Specificity of CAPS Binding—To assess thespecificity of CAPS interactions with syntaxins, we preparedfull-length syntaxins-1, -2, -3, -4, and -6 in PC/PS liposomes forbinding studies. CAPS bound to syntaxins-1, -2, and -4 but notto syntaxins-3 and -6 (Fig. 3A). Sequence comparisons (Fig. 3B)indicated that amino acids Tyr-243, Lys-260, and Lys-265 insyntaxin-1 were conserved in syntaxins-2 and -4 but not in syn-taxins-3 or -6. Mutation of these residues in syntaxin-1 to theircognates in syntaxin-3 produced a chimeric protein (syntaxin-1YKK/HQL) that exhibited impairedCAPS binding (Fig. 3C). Todetermine if CAPS binding to the H3-linker domain was alsoimpaired by these mutations, we utilized SPR to test CAPSbinding to mutant H3-linker domain proteins (Fig. 3D). CAPSbinding toH3-linker domain proteins was strongly inhibited bythe triple mutation of Tyr-243, Lys-260, and Lys-265 as well asby the double mutation at Lys-260 and Lys-265. Indeed, thesinglemutation at Lys-260 seemed to be responsible for the lossof binding (Fig. 3D). Inhibition of binding was also observed by
mutation of Lys-252 to Ala, as isfound in syntaxin-6, but not bymutation of more N-terminal resi-dues in H3 (Fig. 3D). The resultsindicate that C-terminal sites in theH3 domain extending into themembrane-proximal linker regioncomprise part of the binding site forCAPS on syntaxin-1.To determine whether reduced
syntaxin-1 binding affected CAPSfunction, we prepared acceptorliposomeswith full-length syntaxin-1(K260Q/K265L) plus SNAP-25 fortesting in liposome fusion. Lipo-some fusion stimulated by CAPSwas impaired in reactions contain-ing the syntaxin-1(K260Q/K265L)mutant (Fig. 3E). That these muta-tions strongly decreased CAPSbinding to syntaxin-1 (Figs. 3D) butdid not fully impair CAPS-stimu-lated liposome fusion (Fig. 3E) maybe attributed to CAPS interactionswith SNAP-25 in the acceptor lipo-somes (see below). Overall, theresults support the conclusion thatCAPS interacts with a subset of exo-cytic syntaxin isoforms by bindingto membrane-proximal regions ofthe H3-linker domain.CAPS Binds SNAP-25—Binding
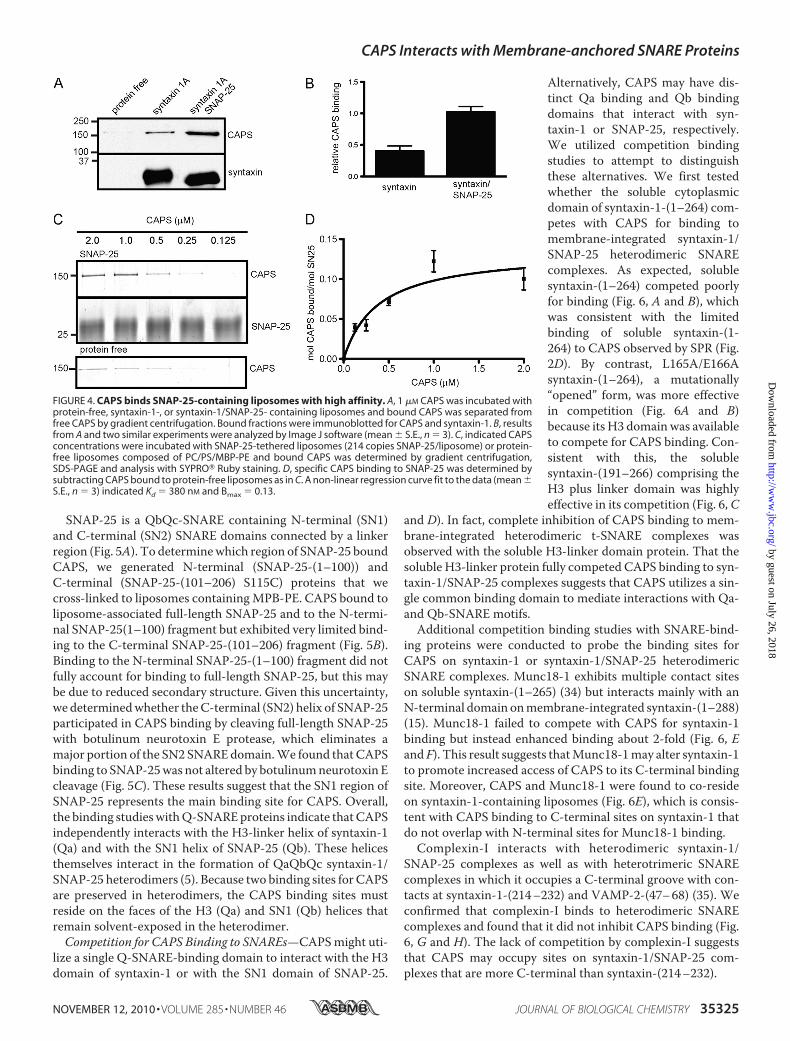
studies with syntaxin-1-containingversus syntaxin-1/SNAP-25-con-taining liposomes showed increasedCAPS binding to the latter (Fig. 4A).CAPS binding to membrane-inte-grated syntaxin-1/SNAP-25 was2-fold greater than to membrane-integrated syntaxin-1 (Fig. 4B). This
suggested that SNAP-25 might constitute a separate bindingsite for CAPS. To directly assess binding to SNAP-25 in theabsence of syntaxin-1, we generated SNAP-25-containing lipo-somes by cross-linking the native cysteine residues of SNAP-25to the maleimide lipid anchor MPB-PE. Liposome flotationstudies revealed that CAPS bound to SNAP-25-containingliposomes to a greater extent than to protein-free liposomes(Fig. 4C). Binding isotherms, which were constructed afterbackground subtraction of CAPS bound to protein-free lipo-somes, indicated an apparent Kd of �400 nM and a Bmax of 0.14mol of CAPS per mol of SNAP-25 (Fig. 4D). SubstoichiometricCAPS interactions with SNAP-25 may be due to the limitedsecondary structure of the helical domains of SNAP-25 in theabsence of syntaxin-1 (33). We note that SPR studies withimmobilized CAPS were unable to detect similar affinity inter-actions with soluble SNAP-25 (not shown). This indicates thatmembrane is an important co-determinant for CAPS bindingto SNAP-25 as was observed for syntaxin-1 binding.
FIGURE 3. CAPS binding is disrupted by mutations in membrane-proximal regions of syntaxin-1. A, 0.5 �M
CAPS was incubated with syntaxin-1, -2, -3, -4, and -6-containing PC/PS liposomes and separated by gradientcentrifugation. Bound CAPS was quantified by immunoblotting with CAPS antibody and normalized to theamounts of syntaxin (mean � S.E., n � 3). B, sequence alignment (ClustalW) of Rattus norvegicus syntaxinisoforms. Amino acid residues (Tyr-243, Lys-260, Lys-265) that are conserved in syntaxin-1, -2, and -4, but not insyntaxin-3 and -6 are indicated by *. Other amino acids (His-199, Met-229, Lys-252) selected for mutagenesisare indicated by †. C, 1 �M CAPS was incubated with protein-free, syntaxin-1- or syntaxin-1(YKK/HQL)-contain-ing PC/PS liposomes and separated by gradient centrifugation for immunoblotting with CAPS antibody.D, CAPS was immobilized on an SPR chip and the indicated syntaxin-(191–266) mutants were analyzed forbinding by flowing over the chip at 20 �M (n � 2– 6). E, liposome fusion assays were conducted with VAMP-2-containing donor liposomes and acceptor PC/PI 4,5-P2 liposomes containing SNAP-25 with syntaxin-1 (E/F) orsyntaxin(K260Q/K265L) (‚/Œ) or with protein-free liposomes (x). Incubations were conducted at 37 °C in thepresence (F, Œ) or absence (E, ‚) of 1 �M CAPS.
CAPS Interacts with Membrane-anchored SNARE Proteins
35324 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 46 • NOVEMBER 12, 2010
SNAP-25 is a QbQc-SNARE containing N-terminal (SN1)and C-terminal (SN2) SNARE domains connected by a linkerregion (Fig. 5A). To determinewhich region of SNAP-25 boundCAPS, we generated N-terminal (SNAP-25-(1–100)) andC-terminal (SNAP-25-(101–206) S115C) proteins that wecross-linked to liposomes containingMPB-PE. CAPS bound toliposome-associated full-length SNAP-25 and to the N-termi-nal SNAP-25(1–100) fragment but exhibited very limited bind-ing to the C-terminal SNAP-25-(101–206) fragment (Fig. 5B).Binding to the N-terminal SNAP-25-(1–100) fragment did notfully account for binding to full-length SNAP-25, but this maybe due to reduced secondary structure. Given this uncertainty,we determinedwhether theC-terminal (SN2) helix of SNAP-25participated in CAPS binding by cleaving full-length SNAP-25with botulinum neurotoxin E protease, which eliminates amajor portion of the SN2 SNAREdomain.We found that CAPSbinding to SNAP-25was not altered by botulinumneurotoxin Ecleavage (Fig. 5C). These results suggest that the SN1 region ofSNAP-25 represents the main binding site for CAPS. Overall,the binding studieswithQ-SNAREproteins indicate that CAPSindependently interacts with the H3-linker helix of syntaxin-1(Qa) and with the SN1 helix of SNAP-25 (Qb). These helicesthemselves interact in the formation of QaQbQc syntaxin-1/SNAP-25 heterodimers (5). Because two binding sites for CAPSare preserved in heterodimers, the CAPS binding sites mustreside on the faces of the H3 (Qa) and SN1 (Qb) helices thatremain solvent-exposed in the heterodimer.Competition for CAPS Binding to SNAREs—CAPSmight uti-
lize a single Q-SNARE-binding domain to interact with the H3domain of syntaxin-1 or with the SN1 domain of SNAP-25.
Alternatively, CAPS may have dis-tinct Qa binding and Qb bindingdomains that interact with syn-taxin-1 or SNAP-25, respectively.We utilized competition bindingstudies to attempt to distinguishthese alternatives. We first testedwhether the soluble cytoplasmicdomain of syntaxin-1-(1–264) com-petes with CAPS for binding tomembrane-integrated syntaxin-1/SNAP-25 heterodimeric SNAREcomplexes. As expected, solublesyntaxin-(1–264) competed poorlyfor binding (Fig. 6, A and B), whichwas consistent with the limitedbinding of soluble syntaxin-(1-264) to CAPS observed by SPR (Fig.2D). By contrast, L165A/E166Asyntaxin-(1–264), a mutationally“opened” form, was more effectivein competition (Fig. 6A and B)because its H3 domain was availableto compete for CAPS binding. Con-sistent with this, the solublesyntaxin-(191–266) comprising theH3 plus linker domain was highlyeffective in its competition (Fig. 6,C
and D). In fact, complete inhibition of CAPS binding to mem-brane-integrated heterodimeric t-SNARE complexes wasobserved with the soluble H3-linker domain protein. That thesoluble H3-linker protein fully competed CAPS binding to syn-taxin-1/SNAP-25 complexes suggests that CAPS utilizes a sin-gle common binding domain to mediate interactions with Qa-and Qb-SNARE motifs.Additional competition binding studies with SNARE-bind-
ing proteins were conducted to probe the binding sites forCAPS on syntaxin-1 or syntaxin-1/SNAP-25 heterodimericSNARE complexes. Munc18-1 exhibits multiple contact siteson soluble syntaxin-(1–265) (34) but interacts mainly with anN-terminal domain onmembrane-integrated syntaxin-(1–288)(15). Munc18-1 failed to compete with CAPS for syntaxin-1binding but instead enhanced binding about 2-fold (Fig. 6, Eand F). This result suggests thatMunc18-1may alter syntaxin-1to promote increased access of CAPS to its C-terminal bindingsite. Moreover, CAPS and Munc18-1 were found to co-resideon syntaxin-1-containing liposomes (Fig. 6E), which is consis-tent with CAPS binding to C-terminal sites on syntaxin-1 thatdo not overlap with N-terminal sites for Munc18-1 binding.Complexin-I interacts with heterodimeric syntaxin-1/
SNAP-25 complexes as well as with heterotrimeric SNAREcomplexes in which it occupies a C-terminal groove with con-tacts at syntaxin-1-(214–232) and VAMP-2-(47–68) (35). Weconfirmed that complexin-I binds to heterodimeric SNAREcomplexes and found that it did not inhibit CAPS binding (Fig.6, G and H). The lack of competition by complexin-I suggeststhat CAPS may occupy sites on syntaxin-1/SNAP-25 com-plexes that are more C-terminal than syntaxin-(214–232).
FIGURE 4. CAPS binds SNAP-25-containing liposomes with high affinity. A, 1 �M CAPS was incubated withprotein-free, syntaxin-1-, or syntaxin-1/SNAP-25- containing liposomes and bound CAPS was separated fromfree CAPS by gradient centrifugation. Bound fractions were immunoblotted for CAPS and syntaxin-1. B, resultsfrom A and two similar experiments were analyzed by Image J software (mean � S.E., n � 3). C, indicated CAPSconcentrations were incubated with SNAP-25-tethered liposomes (214 copies SNAP-25/liposome) or protein-free liposomes composed of PC/PS/MBP-PE and bound CAPS was determined by gradient centrifugation,SDS-PAGE and analysis with SYPRO� Ruby staining. D, specific CAPS binding to SNAP-25 was determined bysubtracting CAPS bound to protein-free liposomes as in C. A non-linear regression curve fit to the data (mean �S.E., n � 3) indicated Kd � 380 nM and Bmax � 0.13.
CAPS Interacts with Membrane-anchored SNARE Proteins
NOVEMBER 12, 2010 • VOLUME 285 • NUMBER 46 JOURNAL OF BIOLOGICAL CHEMISTRY 35325
�-SNAP interacts with heterotrimeric SNARE complexesand with the H3 domain of syntaxin-1 (36). Although the bind-ing sites for�-SNAP have not been precisely determined, it wasproposed that �-SNAP binds to the C-terminal half of the helixbundle of heterotrimeric SNARE complexes (37). �-SNAP wasfound to very effectively inhibit CAPS binding to syntaxin-1/SNAP-25 heterodimers (Fig. 6, G and H). This result supportsthe conclusion that CAPS binds C-terminal sites in t-SNAREheterodimers.
CAPS Also Binds VAMP-2—CAPS promotes the formationof trans SNARE complexes that bridge liposomes (23). Thismight result from CAPS interactions with Q-SNARE syntaxin-1/SNAP-25 heterodimers to promote favorable sites for theinsertion of VAMP-2 to form trans complexes. Alternatively,CAPS might also interact with VAMP-2 to facilitate VAMP-2binding to syntaxin-1/SNAP-25 heterodimers. We tested forpossible CAPS-VAMP-2 interactions by conducting bindingstudies with VAMP-2-containing liposomes using buoyantdensity gradient flotation (Fig. 7A). At liposome concentrationsand copy numbers similar to those used for syntaxin-1 orSNAP-25 binding, CAPS binding to VAMP-2 was limited.However, CAPS binding was evident as the concentration ofVAMP-2-containing liposomes was increased and it exceededthe binding to protein-free liposomes (Fig. 7A). Binding iso-therms constructed after background subtractions (Fig. 7, Band C) revealed that CAPS binding to membrane-integratedVAMP-2 was saturable and exhibited an apparent Kd �1.5 �M
(Fig. 7C). To determine the site on VAMP-2 that mediatedCAPS binding, we prepared liposomes in which VAMP-2 wascleaved by botulinum neurotoxin D. These liposomes, whichcontained a membrane-inserted C-terminal VAMP-2-(60–116) fragment, exhibited strongly reduced CAPS binding (Fig.7D). This result indicates that CAPS binding requires part ofthe 59 residue N-terminal segment of VAMP-2.
DISCUSSION
CAPS is essential for optimal Ca2�-triggered vesicle exocy-tosis where it functions in priming reactions that precede trig-gered membrane fusion (19, 38, 39). Priming involves theassembly of trans SNARE complexes but the assembly pathwayutilized and the accessory factors that catalyze it remain uncer-tain. CAPS promotes SNARE-dependent liposome fusion invitro (22, 23), which may represent priming reactions that pro-ceed into fusion in the absence of downstream regulators suchas synaptotagmin and complexin. CAPS stimulates the forma-tion of SNARE complexes that bridge the liposomes (23) andthe current work was undertaken to identify direct CAPS inter-actions with SNARE proteins. The major conclusion from thiswork is that CAPS binds to each of the SNARE proteins that arerequired for regulated vesicle exocytosis. The results suggestthe possibility that CAPS may promote trans SNARE complexassembly through direct binding interactions with each of thethree SNARE proteins. This possibility will need to be tested infuture work.General features of vesicle targeting to and fusion with
acceptor membranes have been characterized at many traffick-ing steps in the secretory pathway. The core machinery for ves-icle targeting and fusion consists of members from conservedfamilies of Rab, Sec-1/Munc18, and SNARE proteins (40). Atmany membrane trafficking stations, the initial interaction ofvesicles with target membranes is mediated by diverse stage-specific tethering factors or multi-subunit tethering complexes(14). These tethering complexes are frequently regulated byvesicle Rab proteins and they commonly interact with, orinclude, Sec-1/Munc18 family members. Some tethering fac-tors that initially link vesicle and target membranes have beenshown to directly interact with SNARE proteins and to catalyze
FIGURE 5. CAPS binds the SN1 domain of SNAP-25. A, diagram showingSNAP-25-(1–206), SN1 fragment (1–100) and SN2 fragment (101–206). B, 1�M CAPS was incubated with protein-free-liposomes or with SNAP-25-(1–206)-, SNAP-25-(1–100)- or SNAP-25-(101–206)-tethered liposomes inPC/PS/MPB-PE and separated by gradient centrifugation. Bound fractionswere immunoblotted for CAPS and SNAP-25. Representative results forthree experiments are shown. C, 1 �M CAPS was incubated with protein-free or with SNAP-25-containing liposomes in PC/PS/MPB-PE that wereuntreated or treated with botulinum neurotoxin E. Bound fractions iso-lated by gradient centrifugation were immunoblotted for CAPS and SNAP-25. Representative results for three experiments are shown.
CAPS Interacts with Membrane-anchored SNARE Proteins
35326 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 46 • NOVEMBER 12, 2010
trans SNARE complex formation. In ER-Golgi trafficking,Rab1-regulated p115 bridges membranes and promotes transSNARE complex formation through direct interactions withtheQ-SNARE syntaxin-5 and the R-SNAREGOS-28 (13). p115also interacts with other SNARE proteins involved in ER-Golgitrafficking via its SNAREmotif-like C-terminal coiled-coil (13).
For vesicle fusion in the regulatedsecretory pathway, stage-specifictethering complexes have not beenidentified. Mechanisms for theattachment of synaptic or dense-core vesicles to the plasma mem-brane are poorly understood (7).However, CAPS and Munc13 pro-teins, which are both essential forregulated vesicle exocytosis at astage following vesicle attachment,may be the functional homologuesof the stage-specific tethering fac-tors that prime membrane fusion.Consistent with this, C-terminaldomains shared between CAPS andMunc13 proteins were reported toexhibit weak homologies to sub-units of several tethering factors(41).The SNARE protein binding
properties of Munc13 proteins havenot been fully determined. It wasoriginally proposed that a C-termi-nal domain of Munc13-1 interactedwith the N-terminal domain of syn-taxin-1 (17), but this was not con-firmed in biochemical studies (42).Instead, in direct binding studies, aC-terminal domain of Munc13-1was found to interact with het-erodimeric and heterotrimericSNARE complexes but interactionswith individual SNARE proteinswas not evident (45, 48). By con-trast, the current work revealeddirect interactions between CAPSand each of the individual SNAREproteins required for regulated ves-icle exocytosis.A novel and important feature of
CAPS was revealed in finding thatCAPS interacts with the SNAREmotifs in the two Q-SNARE pro-teins. These comprised the H3SNARE plus linker domain of syn-taxin-1 and the SN1SNAREdomainof SNAP-25. High affinity CAPSbinding required the membrane-in-tegration ormembrane-tethering ofthese SNARE proteins, whereasonly very low affinity interactions
occurred with soluble versions of the Q-SNARE proteins. Inaddition to the structural differences in membrane-integratedSNAREs responsible for this difference (31), it is likely thatCAPS participates in forming a ternary CAPS-SNARE-mem-brane complex. The sites identified for CAPS binding in syn-taxin-1 are membrane-proximal. The linker domain in syn-
FIGURE 6. Competition for CAPS binding to syntaxin-1/SNAP-25 heterodimers. A, 0.3 �M CAPS was incu-bated with syntaxin-1/SNAP-25-containing liposomes in the presence of 0 –5 �M syntaxin-(1–264) or syntaxin-(1–264) L165A/E166A. Bound fractions were analyzed by SDS-PAGE and SYPRO� Ruby staining. B, results fromA were analyzed by Image J software for competition by syntaxin-(1–264) (f) and syntaxin-(1–264) L165A/E166A (�) (mean � S.E., n � 3). C, 0.3 �M CAPS was incubated with syntaxin/SNAP-25-containing liposomes asin A in the presence of 0 –20 �M syntaxin H3(191–266). D, results from C were analyzed by Image J software(mean � S.E., n � 3). E, Munc-18-1 and CAPS (1 �M) were incubated together with syntaxin-containing lipo-somes and bound fractions were analyzed on SDS-PAGE by immunoblotting for CAPS and Munc-18-1. F, resultsfrom E were analyzed by Image J software. G, 0.3 �M CAPS was incubated with syntaxin/SNAP-25-containingliposomes in the presence of 0 –10 �M �-SNAP or complexin-I. Bound fractions were analyzed by SDS-PAGE andSYPRO� Ruby staining. H, results from G were analyzed by Image J software for �-SNAP (‚) and complexin-I (f)(mean � S.E., n � 3).
CAPS Interacts with Membrane-anchored SNARE Proteins
NOVEMBER 12, 2010 • VOLUME 285 • NUMBER 46 JOURNAL OF BIOLOGICAL CHEMISTRY 35327
taxin-1 forms part of a continuous helix with the H3 domain(43), and residues required for CAPS binding are on the sameface of the H3 domain (Lys-252) and linker (Lys-260) close tothe membrane. Binding of the SN1 domain of SNAP-25 didnot prevent CAPS interactions with syntaxin-1 but rathercontributed an additional binding site for CAPS in het-erodimeric Q-SNARE complexes (Fig. 4). Liposome fusionpromoted by CAPS was previously shown to requireSNAP-25 (23), and the current work showed that the C-ter-minal H3-linker domain of syntaxin-1 is sufficient for CAPSstimulation of fusion (Fig. 2). C-terminal mutations in syn-taxin-1 impair CAPS function in fusion (Fig. 3), which isconsistent with binding studies that show direct H3-linkerbinding by CAPS and strong binding competition by�-SNAP. These results for CAPS are different from but com-plementary to results for Munc18-1 that showed stimulationof SNARE-dependent liposome fusion requiring the N ter-minus of syntaxin-1 (15).The finding that CAPS interacts with SNARE motifs in both
syntaxin-1 and SNAP-25 suggests several potential novelmechanisms for howCAPSmay participate in SNARE complexformation during priming. Because CAPS can oligomerize (16,44) andmay utilize a commonQa- andQb-binding site on eachmonomer (Fig. 6D), CAPS could function in several ways toregulate the Q-SNARE proteins syntaxin-1 and SNAP-25.CAPS might promote Q-SNARE heterodimer formationthrough Qa and Qb binding by each monomer in a CAPSoligomer. This is unlikely to be the key rate-limiting step
promoted by CAPS because CAPSstimulates SNARE-dependentfusion with proteoliposomes thatcontain pre-formed Q-SNARE het-erodimers (23). However, becausethe helices in Q-SNARE het-erodimers may improperly align inan antiparallel manner, oligomericCAPS could stabilize parallel-aligned heterodimers as has beensuggested for several accessory pro-teins including Munc13 (45). Alter-natively, by interacting with themembrane-proximal regions of hel-ices on syntaxin-1 and SNAP-25 inSNARE heterodimers, CAPS oli-gomers could function to cluster ororganizeQ-SNAREheterodimers atfusion sites for optimal insertion ofvesicle VAMP-2 (22). This possibil-ity could account for the observedability of CAPS to promote lipo-some fusion at extremely lowQ-SNARE densities (23). Theseproposed direct actions of CAPS onQ-SNAREs will need to be explicitlytested.In accord with recent views that
Munc18-1 regulates N-terminaldomains of syntaxin-1 (15, 46),
Munc18-1 failed to compete with CAPS for binding to syn-taxin-1 but it unexpectedly enhanced CAPS interactions withsyntaxin-1 (Fig. 6F). This finding suggests that Munc18-1 mayalter the conformation of syntaxin-1 to enable CAPS binding.Alternatively, CAPS and Munc18-1 might interact so thatMunc18-1 binding to syntaxin-1 would provide an additionalCAPS binding site. Nonetheless, that both Munc18-1 andCAPS were observed to co-reside on syntaxin-1-containingliposomes (Fig. 6E) is consistent with the assignments of N-ter-minal and C-terminal binding sites for Munc18-1 and CAPS,respectively. Because our liposome flotation studies of SNAREbinding did not achieve complete stoichiometry, we cannot ruleout possible binding of Munc18-1 and CAPS to different syn-taxin-1-containing liposomes; however, it was clear that thereis at least some overlap because Munc18-1 strongly affectedCAPS binding (Fig. 6F). Overall, the results indicate that bind-ing sites for Munc18-1 and CAPS on Q-SNARE heterodimersare complementary.CAPS also interacts with the membrane-integrated
R-SNARE VAMP-2 (Fig. 7). VAMP-2 is not completely struc-tured in isolation (31) and it is unclear whether the reducedaffinity and decreased stoichiometry for CAPS binding reflectsthe partly unfolded state for VAMP-2 on liposomes. CAPSinteractions with VAMP-2, unlike those for syntaxin-1 C-ter-minal domain binding, were with N-terminal domains ofVAMP-2 as indicated by the ability of botulinum neurotoxin Dto abolish binding. A “zipper”model for heterotrimeric SNAREcomplex assembly proposes the N- to C-terminal insertion of
FIGURE 7. CAPS binds the N-terminal domain of VAMP-2. A, 1 �M CAPS was incubated with 0.25, 0.5, and 1.0�M VAMP-2 on PC/PS liposomes formed by co-micellization (90 copies of VAMP-2/liposome) or with corre-sponding amounts of protein-free liposomes. Bound fractions isolated by gradient centrifugation were immu-noblotted for CAPS and VAMP-2. Representative results for three experiments are shown. B, indicated concen-trations of CAPS were incubated with VAMP-2-containing or protein-free liposomes and separated by gradientcentrifugation followed by SDS-PAGE and SYPRO� Ruby staining. C, CAPS bound to protein-free liposomes wassubtracted from CAPS bound to VAMP-2 liposomes and the data (mean � S.E., n � 3) were fit to a non-linearregression curve (Bmax � 0.04, Kd � 1.49 �M, R2 � 0.903). D, 1 �M CAPS was incubated with protein-free orVAMP-2-containing liposomes that were untreated or treated with botulinum neurotoxin D. Bound fractionsfrom gradients were immunoblotted for CAPS and VAMP-2. Representative results for three experiments areshown.
CAPS Interacts with Membrane-anchored SNARE Proteins
35328 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 46 • NOVEMBER 12, 2010
VAMP-2 into syntaxin-1/SNAP-25 heterodimers (5, 6). Thiswould imply an N-terminal nucleation sequence on VAMP-2that is initially integrated into the SNARE helix bundle.Through its N-terminal VAMP-2 interactions, CAPS bound toheterodimeric Q-SNARE complexes could be involved in cata-lyzing the zippering of VAMP-2 into recipient complexes.CAPS is the only protein essential for regulated vesicle exo-
cytosis that has been shown to exhibit direct independent inter-actions with each of the SNARE proteins required for vesiclefusion. However, recent studies have demonstrated thatMunc18-1, in addition to its interactions with Q-SNARE het-erodimers (Fig. 6 and Ref. 45), also interacts with VAMP-2 (47).The binding site for Munc18-1 on VAMP-2 is C-terminalwhereas that for CAPS is N-terminal. Thus, studies onMunc18-1 and CAPS now indicate that their binding sites onQ-SNARE heterodimers and on the R-SNARE VAMP-2 arecomplementary. This suggests a possible mechanism for prim-ing in which both factors bound to syntaxin-1/SNAP-25 het-erodimers play a catalytic role in the insertion of VAMP-2 toform trans SNARE complexes. CAPS andMunc18-1 might actcooperatively with CAPS assisting in the initial nucleation oftheN terminus ofVAMP-2 intoQ-SNARE recipient complexesfollowed by C-terminal “zippering” assisted by Munc18-1.In summary, we describe for the first time the unique inter-
actions of CAPS with each of the SNARE proteins required forregulated vesicle exocytosis. CAPS binding to Q-SNAREs ismediated by C-terminal, membrane-proximal interactionswith two of three SNAREmotifs in the heterodimeric complex.By contrast, CAPS binds N-terminal portions of the R-SNAREVAMP-2. These findings provide potential insight into mecha-nisms by which CAPS could promote trans SNARE complexformation for priming vesicle exocytosis.
Acknowledgments—SPR data were obtained at the University ofWis-consin Biophysics Instrumentation Facility established with grantsfrom the National Science Foundation (BIR-9512577) and NationalInstitutes of Health (S10 RR13790).
REFERENCES1. Malsam, J., Kreye, S., and Sollner, T. H. (2008) Cell Mol. Life Sci. 65,
2814–28322. Weber, T., Zemelman, B. V., McNew, J. A., Westermann, B., Gmachl, M.,
Parlati, F., Sollner, T. H., and Rothman, J. E. (1998) Cell 92, 759–7723. Sutton, R. B., Fasshauer,D., Jahn, R., andBrunger, A. T. (1998)Nature395,
347–3534. Fasshauer, D., Sutton, R. B., Brunger, A. T., and Jahn, R. (1998) Proc. Natl.
Acad. Sci. U.S.A. 95, 15781–157865. Fasshauer, D., and Margittai, M. (2004) J. Biol. Chem. 279, 7613–76216. Pobbati, A. V., Stein, A., and Fasshauer, D. (2006) Science 313, 673–6767. Verhage, M., and Sørensen, J. B. (2008) Traffic 9, 1414–14248. Wojcik, S. M., and Brose, N. (2007) Neuron 55, 11–249. Klenchin, V. A., and Martin, T. F. (2000) Biochimie 82, 399–40710. Rettig, J., and Neher, E. (2002) Science 298, 781–78511. Sørensen, J. B. (2005) Trends Neurosci. 28, 453–45512. Walter, A.M.,Wiederhold, K., Bruns, D., Fasshauer, D., and Sørensen, J. B.
(2010) J. Cell Biol. 188, 401–41313. Shorter, J., Beard, M. B., Seemann, J., Dirac-Svejstrup, A. B., and Warren,
G. (2002) J. Cell Biol. 157, 45–6214. Sztul, E., and Lupashin, V. (2009) FEBS Lett. 583, 3770–378315. Shen, J., Tareste, D. C., Paumet, F., Rothman, J. E., and Melia, T. J. (2007)
Cell 128, 183–19516. Walent, J. H., Porter, B. W., and Martin, T. F. (1992) Cell 70, 765–77517. Betz, A., Okamoto, M., Benseler, F., and Brose, N. (1997) J. Biol. Chem.
272, 2520–252618. Koch, H., Hofmann, K., and Brose, N. (2000) Biochem. J. 349, 247–25319. Grishanin, R. N., Kowalchyk, J. A., Klenchin, V. A., Ann, K., Earles, C. A.,
Chapman, E. R., Gerona, R. R., and Martin, T. F. (2004) Neuron 43,551–562
20. Hay, J. C., Fisette, P. L., Jenkins, G. H., Fukami, K., Takenawa, T., Ander-son, R. A., and Martin, T. F. (1995) Nature 374, 173–177
21. Milosevic, I., Sørensen, J. B., Lang, T., Krauss, M., Nagy, G., Haucke, V.,Jahn, R., and Neher, E. (2005) J. Neurosci. 25, 2557–2565
22. James, D. J., Khodthong, C., Kowalchyk, J. A., and Martin, T. F. (2008)J. Cell Biol. 182, 355–366
23. James, D. J., Kowalchyk, J., Daily, N., Petrie, M., and Martin, T. F. (2009)Proc. Natl. Acad. Sci. U.S.A. 106, 17308–17313
24. Schuette, C. G., Hatsuzawa, K., Margittai, M., Stein, A., Riedel, D., Kuster,P., Konig, M., Seidel, C., and Jahn, R. (2004) Proc. Natl. Acad. Sci. U.S.A.101, 2858–2863
25. Fasshauer, D., Eliason,W. K., Brunger, A. T., and Jahn, R. (1998) Biochem-istry 37, 10354–10362
26. Grishanin, R. N., Klenchin, V. A., Loyet, K. M., Kowalchyk, J. A., Ann, K.,and Martin, T. F. (2002) J. Biol. Chem. 277, 22025–22034
27. Takamori, S., Holt, M., Stenius, K., Lemke, E. A., Grønborg,M., Riedel, D.,Urlaub,H., Schenck, S., Brugger, B., Ringler, P.,Muller, S. A., Rammner, B.,Grater, F., Hub, J. S., De Groot, B. L., Mieskes, G., Moriyama, Y., Klingauf,J., Grubmuller, H., Heuser, J., Wieland, F., and Jahn, R. (2006) Cell 127,831–846
28. Mayer, L. D., Hope, M. J., and Cullis, P. R. (1986) Biochim. Biophys. Acta858, 161–168
29. McNew, J. A., Weber, T., Parlati, F., Johnston, R. J., Melia, T. J., Sollner,T. H., and Rothman, J. E. (2000) J. Cell Biol. 150, 105–117
30. Lang, T., Margittai, M., Holzler, H., and Jahn, R. (2002) J. Cell Biol. 158,751–760
31. Ellena, J. F., Liang, B., Wiktor, M., Stein, A., Cafiso, D. S., Jahn, R., andTamm, L. K. (2009) Proc. Natl. Acad. Sci. U.S.A. 106, 20306–20311
32. Misura, K. M., Scheller, R. H., and Weis, W. I. (2001) J. Biol. Chem. 276,13273–13282
33. Fasshauer, D., Otto, H., Eliason, W. K., Jahn, R., and Brunger, A. T. (1997)J. Biol. Chem. 272, 28036–28041
34. Misura, K.M., Scheller, R. H., andWeis,W. I. (2000)Nature 404, 355–36235. Chen, X., Tomchick, D. R., Kovrigin, E., Arac, D., Machius, M., Sudhof,
T. C., and Rizo, J. (2002) Neuron 33, 397–40936. Hayashi, T., Yamasaki, S., Nauenburg, S., Binz, T., andNiemann,H. (1995)
EMBO J. 14, 2317–232537. Marz, K. E., Lauer, J. M., and Hanson, P. I. (2003) J. Biol. Chem. 278,
27000–2700838. Jockusch, W. J., Speidel, D., Sigler, A., Sørensen, J. B., Varoqueaux, F.,
Rhee, J. S., and Brose, N. (2007) Cell 131, 796–80839. Liu, Y., Schirra, C., Stevens, D. R., Matti, U., Speidel, D., Hof, D., Bruns, D.,
Brose, N., and Rettig, J. (2008) J. Neurosci. 28, 5594–560140. Grosshans, B. L., Ortiz, D., and Novick, P. (2006) Proc. Natl. Acad. Sci.
U.S.A. 103, 11821–1182741. Pei, J., Ma, C., Rizo, J., and Grishin, N. V. (2009) J. Mol. Biol. 391, 509–51742. Basu, J., Shen, N., Dulubova, I., Lu, J., Guan, R., Guryev, O., Grishin, N. V.,
Rosenmund, C., and Rizo, J. (2005) Nat. Struct. Mol. Biol. 12, 1017–101843. Stein, A., Weber, G., Wahl, M. C., and Jahn, R. (2009) Nature 460,
525–52844. Nojiri,M., Loyet, K.M., Klenchin, V. A., Kabachinski, G., andMartin, T. F.
(2009) J. Biol. Chem. 284, 18707–1871445. Weninger, K., Bowen,M. E., Choi, U. B., Chu, S., and Brunger, A. T. (2008)
Structure 16, 308–32046. Burkhardt, P., Hattendorf, D. A., Weis, W. I., and Fasshauer, D. (2008)
EMBO J. 27, 923–93347. Xu, Y., Su, L., and Rizo, J. (2010) Biochemistry 49, 1568–157648. Guan, R., Dai, H., and Rizo, J. (2008) Biochemistry 47, 1474–1481
CAPS Interacts with Membrane-anchored SNARE Proteins
NOVEMBER 12, 2010 • VOLUME 285 • NUMBER 46 JOURNAL OF BIOLOGICAL CHEMISTRY 35329