Plant Physiol. (1984) 74, 795-799 0032-0889/84/74/0795/05/$01.00/0 De Novo Maltotriose Biosynthesis from the Reducing End by Spinacia oleracea L. Chloroplasts Received for publication July 26, 1983 and in revised form November 28, 1983 JAMES C. LINDEN*I AND N. SCHILLING Department of Biochemistry and Biophysics, Iowa State University, Ames, Iowa 50011 (J. C. L.); and Botanisches Institut der Universitaet Muenchen, 8000 Muenchen 19, Federal Republic ofGermany ABSTRACT The distribution of 14C in the various glucose residues of maltotriose was studied as a function of time of photosynthesis of isolated chloro- plasts of spinach (Spilnacia oeraca L.) using 4C02. The distribution of label showed that the reducing-end glucose residue was labeled first and the label subsequently distributed to the second and third glucose residues at approximately equal rates. A mechanism for the distribution of label and the synthesis of malto- triose from the reducing end is presented. The mechanism has postulated to be the same as that for the maltose synthase recently described by Schilling. Maltose biosynthesis from a-D-glucose-l-phosphate was char- acterized as involving two glucosyl intermediates by a double displace- ment mechanism with inversion of configuration. The mode of enzymic action by which maltosyl intermediates were transferred to glucosyl intermediates was consistent with the fractional distribution of radioac- tivity found in each glucose unit of maltotriose during short term photo- synthesis experiments. Maltodextrins have not been generally recognized as constit- uents of photosynthetic tissue. Indeed, the maltodextrins were not observed as such until green algae were labeled during photosynthesis in '4C02. Following photosynthesis of Chlorella and Scenedesmus in "4C02 and extraction of these tissues, Bas- sham and Calvin (2) prepared radioautograms of two-dimen- sional chromatograms of the extracts. The radioautograms showed a homologous series of compounds extending from the origin to sucrose. The homologous series was identified by French (6) as maltodextrins (i.e. a-l,4-linked D-glucose oligosaccha- rides). Kandler (12) also observed the maltodextrin series on two-dimensional chromatograms of various plant extracts follow- ing "'CO2 photosynthesis. Heber (9) reported tri-, tetra-, and pentasaccharides present in chloroplasts of Nitella, Valonia, and Chara. Maltotriose and maltotetraose represented a major per- centage of the excretion products of Oscillatoria redeker (11). Jensen also noted maltodextrins in extracts of isolated spinach chloroplasts after photosynthesis in "'CO2 (10). Maltose biosynthesis in vivo has been shown to involve a- glucose-I-P (18). A maltose synthase isolated from spinach was recently described by Schilling (19) in which maltose formation 'Supported by the Alexander von Humboldt Stiftung, Bad Godesberg, Federal Republic of Germany during the portion of the work conducted at the University of Munich. Current address: Department of Agricultural and Chemical Engineering, Colorado State University, Fort Collins, CO 80523. from aG-I-P2 was shown to involve two glucosyl-enzyme inter- mediates (,BG-E and ,BG-E') by a double displacement mecha- nism: 2 aG-l-P + E + E' .,BfG-E + ,BG-E' + 2 Pi ,BG-E + ,BG-E' = aG-f#G-E' + E aG-,BG-E' = aG-aG + E' (1) (2) (3) Robyt (17) has described a similar mechanism for polysaccharide biosynthesis involving transfer of activated monomer units to the reducing end of a growing polysaccharide chain covalently linked to the active site of the enzyme. The work reported in this paper describes the relationship of the above enzymic activity to a possible route of maltotriose biosynthesis. The labeling of maltotriose during photosynthesis in '4CO2 has been found to originate in the reducing end glucose moiety, and can be explained by extension of the glucosyl intermediate displacement mechanism described above. MATERIALS AND METHODS Chloroplast Preparation. Chloroplasts were isolated in pyro- phosphate buffer according to methods of Cockbum et al. (5) from fresh spinach (Spinacia oleracea L.) leaves purchased at the grocery. About 50 g of washed and chilled leaf laminae were homogenized in a Waring Blendor for 3 to 5 s in 200 ml of semifrozen buffer containing 0.333 M sorbitol, 0.005 M MgC12, 0.010 M Na4PjO7, and 0.004 M L-ascorbic acid. The latter was added after adjustment of the icy buffer to pH 6.6. The macerate was filtered through cheesecloth into 50 ml centrifuge tubes. The chloroplasts were pelleted at 0C from rest to 10OOg to rest in approximately 90 s. The supernatant was discarded and the pellet was suspended in 5 ml of ice-cold solution containing 0.333 M sorbitol, 0.001 M MgCl2, 0.001 M MnC12, 0.002 M ethylendinitrilotetracetic acid (EDTA), and 0.050 M Hepes. The pH of this buffer was 7.6 at 20C. The pellet was washed and centrifuged as above three times in this buffer at OC. Photosynthesis Procedures. Photosynthesis by the isolated chloroplasts was conducted in the pH 7.6 Hepes buffer following the procedure of Bassham et aL (4). Potassium bicarbonate and sorbitol were added to the suspension of chloroplasts so that 2 Abbreviations: aG- I-P, a-E-glucose- I -phosphate; aG*, alpha glucose containing '4C-label; E and E', independent active sites on maltose synthase enzyme(s); #G-E, beta glucosyl intermediate of maltose syn- thase; a-G-#G-E, beta maltosyl intermediate of enzyme; aG-a-G, alpha maltose; aG-aG-#G-E, beta maltotriosyl enzyme intermediate; aG-aG- aG*, maltotriose labeled in reducing end; G2, maltose; G3, maltotriose; G4, maltotetraose; Gs, maltopentaose; G7, maltoheptaose; G9, maltonon- aose; DP, degree of polymerization. 795 Downloaded from https://academic.oup.com/plphys/article/74/4/795/6079445 by guest on 12 February 2022
De Novo Maltotriose Biosynthesis from the Reducing Endby Spinacia oleracea L. Chloroplasts
Received for publication July 26, 1983 and in revised form November 28, 1983
JAMES C. LINDEN*I AND N. SCHILLINGDepartment ofBiochemistry and Biophysics, Iowa State University, Ames, Iowa 50011 (J. C. L.); andBotanisches Institut der Universitaet Muenchen, 8000 Muenchen 19, Federal Republic ofGermany
ABSTRACT
The distribution of 14C in the various glucose residues of maltotriosewas studied as a function of time of photosynthesis of isolated chloro-plasts of spinach (Spilnacia oeraca L.) using 4C02. The distribution oflabel showed that the reducing-end glucose residue was labeled first andthe label subsequently distributed to the second and third glucose residuesat approximately equal rates.A mechanism for the distribution of label and the synthesis of malto-
triose from the reducing end is presented. The mechanism has postulatedto be the same as that for the maltose synthase recently described bySchilling. Maltose biosynthesis from a-D-glucose-l-phosphate was char-acterized as involving two glucosyl intermediates by a double displace-ment mechanism with inversion of configuration. The mode of enzymicaction by which maltosyl intermediates were transferred to glucosylintermediates was consistent with the fractional distribution of radioac-tivity found in each glucose unit of maltotriose during short term photo-synthesis experiments.
Maltodextrins have not been generally recognized as constit-uents of photosynthetic tissue. Indeed, the maltodextrins werenot observed as such until green algae were labeled duringphotosynthesis in '4C02. Following photosynthesis of Chlorellaand Scenedesmus in "4C02 and extraction of these tissues, Bas-sham and Calvin (2) prepared radioautograms of two-dimen-sional chromatograms of the extracts. The radioautogramsshowed a homologous series of compounds extending from theorigin to sucrose. The homologous series was identified by French(6) as maltodextrins (i.e. a-l,4-linked D-glucose oligosaccha-rides). Kandler (12) also observed the maltodextrin series ontwo-dimensional chromatograms ofvarious plant extracts follow-ing "'CO2 photosynthesis. Heber (9) reported tri-, tetra-, andpentasaccharides present in chloroplasts of Nitella, Valonia, andChara. Maltotriose and maltotetraose represented a major per-centage of the excretion products of Oscillatoria redeker (11).Jensen also noted maltodextrins in extracts of isolated spinachchloroplasts after photosynthesis in "'CO2 (10).
Maltose biosynthesis in vivo has been shown to involve a-glucose-I-P (18). A maltose synthase isolated from spinach wasrecently described by Schilling (19) in which maltose formation
'Supported by the Alexander von Humboldt Stiftung, Bad Godesberg,Federal Republic ofGermany during the portion of the work conductedat the University ofMunich. Current address: Department ofAgriculturaland Chemical Engineering, Colorado State University, Fort Collins, CO80523.
from aG-I-P2 was shown to involve two glucosyl-enzyme inter-mediates (,BG-E and ,BG-E') by a double displacement mecha-nism:
2 aG-l-P + E + E' .,BfG-E + ,BG-E' + 2 Pi,BG-E + ,BG-E' = aG-f#G-E' + E
aG-,BG-E' = aG-aG + E'
(1)
(2)(3)
Robyt (17) has described a similar mechanism for polysaccharidebiosynthesis involving transfer of activated monomer units tothe reducing end of a growing polysaccharide chain covalentlylinked to the active site of the enzyme.The work reported in this paper describes the relationship of
the above enzymic activity to a possible route of maltotriosebiosynthesis. The labeling of maltotriose during photosynthesisin '4CO2 has been found to originate in the reducing end glucosemoiety, and can be explained by extension of the glucosylintermediate displacement mechanism described above.
MATERIALS AND METHODS
Chloroplast Preparation. Chloroplasts were isolated in pyro-phosphate buffer according to methods of Cockbum et al. (5)from fresh spinach (Spinacia oleracea L.) leaves purchased atthe grocery. About 50 g ofwashed and chilled leaf laminae werehomogenized in a Waring Blendor for 3 to 5 s in 200 ml ofsemifrozen buffer containing 0.333 M sorbitol, 0.005 M MgC12,0.010 M Na4PjO7, and 0.004 M L-ascorbic acid. The latter wasadded after adjustment of the icy buffer to pH 6.6.The macerate was filtered through cheesecloth into 50 ml
centrifuge tubes. The chloroplasts were pelleted at 0C from restto 10OOg to rest in approximately 90 s. The supernatant wasdiscarded and the pellet was suspended in 5 ml of ice-coldsolution containing 0.333 M sorbitol, 0.001 M MgCl2, 0.001 MMnC12, 0.002 M ethylendinitrilotetracetic acid (EDTA), and0.050 M Hepes. The pH of this buffer was 7.6 at 20C. The pelletwas washed and centrifuged as above three times in this bufferat OC.
Photosynthesis Procedures. Photosynthesis by the isolatedchloroplasts was conducted in the pH 7.6 Hepes buffer followingthe procedure of Bassham et aL (4). Potassium bicarbonate andsorbitol were added to the suspension of chloroplasts so that
2 Abbreviations: aG- I-P, a-E-glucose- I-phosphate; aG*, alpha glucosecontaining '4C-label; E and E', independent active sites on maltosesynthase enzyme(s); #G-E, beta glucosyl intermediate of maltose syn-thase; a-G-#G-E, beta maltosyl intermediate of enzyme; aG-a-G, alphamaltose; aG-aG-#G-E, beta maltotriosyl enzyme intermediate; aG-aG-aG*, maltotriose labeled in reducing end; G2, maltose; G3, maltotriose;G4, maltotetraose; Gs, maltopentaose; G7, maltoheptaose; G9, maltonon-aose; DP, degree of polymerization.
795
Dow
nloaded from https://academ
ic.oup.com/plphys/article/74/4/795/6079445 by guest on 12 February 2022
LINDEN AND SCHILLING
respective concentrations would remain 0.010 M in bicarbonateand 0.333 M in sorbitol after addition of radioactive bicarbonate.The 5 ml of chloroplast suspension were placed in a 3.3-cmdiameter, 24/25 standard taper, round bottom flask with sidearm (Thomas, Philadelphia) and were kept suspended by thegentle motion of a wrist-action shaker. The bottom of the flaskwas suspended in water thermostated at 20C. Illuminationthrough the water was a bank of six 61-cm daylight fluorescentbulbs. Photosynthesis was conducted for 15 to 20 min beforeinjection of 0.2 ml (13 gmol) of high specific activity [14C]NaHCO3 (40 oCi/flmol) into the suspension from a hypodermicsyringe by means of 20 cm of 1 mm polyethylene tubing whichpassed through the side arm of the flask. Samples of 0.4 ml wereremoved by the same means after intervals of time and injectedinto 1.6 ml of absolute methanol at 4C. After centrifugation atl0,OOOg for 10 min, the green supernatants and white pelletswere separated and stored at - 15C. In vivo photosynthesis ofspinach leaves in 14CO2 was conducted as described earlier ( 12).Paper Chromatography. For the analysis of the products of
photosynthesis in the alcoholic extracts in algae and chloroplasts,two-dimensional descending paper chromatography, as describedby Bassham and Kirk (3), was employed. To obtain the neutralmaltodextrins free from amino acids, organic phosphates andother ionic materials, the alcoholic extracts were deionized withAmberlite MB-3 mixed-bed ion-exchange resin which had beensaturated with glucose prior to use to reduce nonspecific bindingof maltodextrins to the resin. Following concentration, the sam-ples were chromatographed on water-washed Whatman No. 1 orWhatman 3 MM chromatography paper by multiple ascents insealed stainless steel tanks at 65°C as described by French et al.(7) in solvent containing 70% n-propyl alcohol:water (7:3, v/v).Radioautograms were prepared using Kodak No-Screen x-rayfilm. Chromatograms were developed for reducing compoundsby dipping in silver nitrate-acetone and alkali-methanol reagents(20). The distribution of radioactivity between the glucose unitsof maltotriose was estimated following a-amylolysis either onpaper by two-dimensional chromatography interspersed withp3-amylase as described by French et al. (8) or in test tubesfollowing elution of the radioactive maltotriose from the paperchromatograms. The f3-amylase digests were rechromatographedas described above for separation ofglucose and maltose hydrol-ysis products. The distribution of radioactivity in the maltoseproduct was determined following reduction with sodium boro-hydride and acid hydrolysis. Descending chromatography onWhatman 3 MM paper in solvent containing ethyl acetate:aceticacid: saturated boric acid solution (9:1:1, v/v/v) was used toseparate glucose and sorbitol (21). Areas from the chromato-grams which corresponded to radiographic spots were cut outand counted by scintillation spectrometry. Scintillation solutioncontaining 4.0 g PPO and 0.2 g POPOP per liter of toluene wasused for counting radioactivity on paper in a Packard Tri-Carbscintillation spectrometer.
RESULTSThe kinetics and distribution of labeling of maltotriose from
two photosynthesis experiments were studied. During both ex-periments, one conducted with continuous illumination and theother with a light-dark transition, the kinetics of maltotriose (G3)and maltotetraose (G4) labeling were nearly identical to eachother but quite distinct from the labeling kinetics of maltose (G2)and higher maltodextrins (OG5). This is illustrated in Figure 1for which data from the continuous illumination experiment ispresented. The counts of radioactive carbon appearing in malt-ose, maltotriose plus maltotetraose, maltopentaose, and malto-dextrins with DP > 8 are plotted as a function of time followinginjection of [r'4CNaHCO3 into the spinach chloroplast suspen-sion. The radioactivity appearing in maltohexaose, maltohep-
E
40
o
-20
I0
0 5 10 15 20 25 30 35 40 45TIME OF C - PHOTOSYNTHESIS (minutes)
FIG. 1. Kinetics of "'C-labeling photosynthetic products in spinachchloroplasts with continuous illumination. Quantities of radioactivityaccumulated with time in maltose (A), maltotriose plus maltotetraose(U), maltopentaose (0), and maltodextrins with DP > 8 which were notchromatographically resolved (C), from equal volume quantities of ex-tracts of spinach chloroplasts removed during a photosynthesis experi-ment in "CO2.
taose, and maltooctaose, which followed the same kinetics asthat in maltopentaose, were not plotted for clarity. Radioactivityappeared in maltose rapidly and continued to accumulate at thesame rate throughout the 45-min sampling period. Similar ki-netics were demonstrated for glucose, fructose, and sucrose (datanot presented), whereas the rate of radioactivity accumulation inthe maltodextrins other than maltose declined dramatically atabout 15 min. There was similarity in curves for the chromato-graphically unresolved maltodextrins (>G8) and those for themaltodextrins G5, G6, G7, and G8. The kinetics of G3 and G4were distinct from those of the maltodextrins 2G5. Label in G3and G4 continued to accumulate, whereas label in maltodextrinsoG5 remained at the same level or declined.The distribution of radioactivity in maltotriose from both
experiments was determined. After hydrolysis of G3 with,8-amylase, maltose and glucose were products; the reaction isshown in reaction 4.
aG-aG-aG' -- aG-#G + aG* (4)
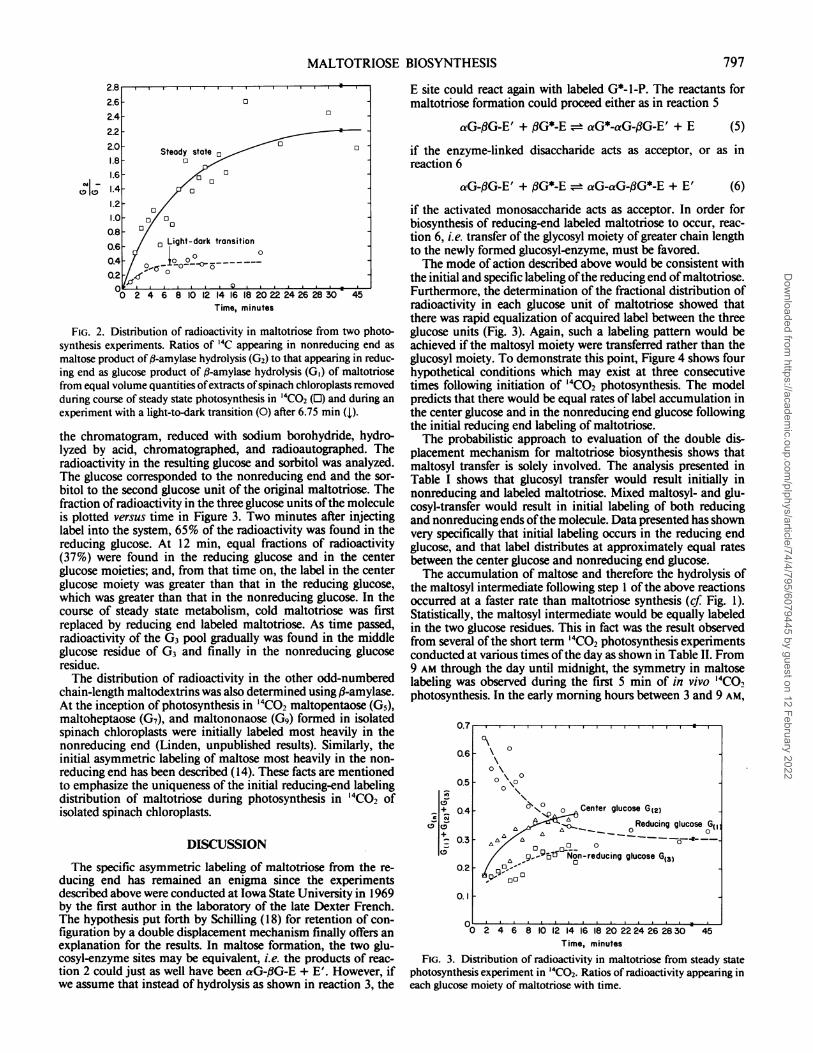
The ratio of radioactivities in maltose to glucose is plotted inFigure 2 as a function of time. Initially, the glucose was morehighly labeled than maltose. In fact, the curves obtained fromboth experiments extrapolate to zero in which case maltosewould have no label at zero time, and the reducing end of themolecule would have acquired label almost immediately afterinjection of radioactivity in the system. This would indicatesynthesis of maltotriose from the reducing end. It appeared fromthese observations that the labeling of maltotriose was veryspecific.The differences between the rate of labeling of maltotriose in
the two experiments may have been a result of the light-darktransition after 6.75 min of steady state photosynthesis in thelatter experiment. However, the initial slopes of the two curveswere unequal, which may indicate physiological differences inthe chloroplasts of the two preparations. The extent of labelingis apparently dependent upon the ability ofthe chloroplast to fixC02, and the fixation of CO2 is said to be dependent on thestructural integrity of the chloroplasts (1).The complete distribution of label in maltotriose from the
steady state experiment was determined. The maltose, which hadbeen hydrolyzed from maltotriose by ,3-amylase, was eluted from
796 Plant Physiol. Vol. 74, 1984
Dow
nloaded from https://academ
ic.oup.com/plphys/article/74/4/795/6079445 by guest on 12 February 2022
MALTOTRIOSE BIOSYNTHESIS
10 12 14 16 18 20 22 24 26 28 30 45Time, minutes
FIG. 2. Distribution of radioactivity in maltotriose from two photo-synthesis experiments. Ratios of '4C appearing in nonreducing end as
maltose product of ,B-amylase hydrolysis (G2) to that appearing in reduc-ing end as glucose product of ,B-amylase hydrolysis (GI) of maltotriosefrom equal volume quantities ofextracts ofspinach chloroplasts removedduring course of steady state photosynthesis in '4CO2 (0) and during an
experiment with a light-to-dark transition (0) after 6.75 min (1).
the chromatogram, reduced with sodium borohydride, hydro-lyzed by acid, chromatographed, and radioautographed. Theradioactivity in the resulting glucose and sorbitol was analyzed.The glucose corresponded to the nonreducing end and the sor-bitol to the second glucose unit of the original maltotriose. Thefraction ofradioactivity in the three glucose units ofthe moleculeis plotted versus time in Figure 3. Two minutes after injectinglabel into the system, 65% of the radioactivity was found in thereducing glucose. At 12 min, equal fractions of radioactivity(37%) were found in the reducing glucose and in the centerglucose moieties; and, from that time on, the label in the centerglucose moiety was greater than that in the reducing glucose,which was greater than that in the nonreducing glucose. In thecourse of steady state metabolism, cold maltotriose was firstreplaced by reducing end labeled maltotriose. As time passed,radioactivity of the G3 pool gradually was found in the middleglucose residue of G3 and finally in the nonreducing glucoseresidue.The distribution of radioactivity in the other odd-numbered
chain-length maltodextrins was also determined using ,8-amylase.At the inception of photosynthesis in '4C02 maltopentaose (G5),maltoheptaose (G7), and maltononaose (Gg) formed in isolatedspinach chloroplasts were initially labeled most heavily in thenonreducing end (Linden, unpublished results). Similarly, theinitial asymmetric labeling of maltose most heavily in the non-reducing end has been described (14). These facts are mentionedto emphasize the uniqueness of the initial reducing-end labelingdistribution of maltotriose during photosynthesis in "'CO2 ofisolated spinach chloroplasts.
DISCUSSION
The specific asymmetric labeling of maltotriose from the re-ducing end has remained an enigma since the experimentsdescribed above were conducted at Iowa State University in 1969by the first author in the laboratory of the late Dexter French.The hypothesis put forth by Schilling (18) for retention of con-figuration by a double displacement mechanism finally offers anexplanation for the results. In maltose formation, the two glu-cosyl-enzyme sites may be equivalent, i.e. the products of reac-tion 2 could just as well have been aG-#G-E + E'. However, ifwe assume that instead of hydrolysis as shown in reaction 3, the
E site could react again with labeled G*-1-P. The reactants formaltotriose formation could proceed either as in reaction 5
aG-,BG-E' + ,G*-E = aG*-aG-,G-E' + E (5)
if the enzyme-linked disaccharide acts as acceptor, or as inreaction 6
aG-,BG-E' + #G*-E = aG-aG-,BG*-E + E' (6)
if the activated monosaccharide acts as acceptor. In order forbiosynthesis of reducing-end labeled maltotriose to occur, reac-tion 6, i.e. transfer of the glycosyl moiety of greater chain lengthto the newly formed glucosyl-enzyme, must be favored.The mode of action described above would be consistent with
the initial and specific labeling ofthe reducing end of maltotriose.Furthermore, the determination of the fractional distribution ofradioactivity in each glucose unit of maltotriose showed thatthere was rapid equalization of acquired label between the threeglucose units (Fig. 3). Again, such a labeling pattern would beachieved if the maltosyl moiety were transferred rather than theglucosyl moiety. To demonstrate this point, Figure 4 shows fourhypothetical conditions which may exist at three consecutivetimes following initiation of 14C02 photosynthesis. The modelpredicts that there would be equal rates of label accumulation inthe center glucose and in the nonreducing end glucose followingthe initial reducing end labeling of maltotriose.The probabilistic approach to evaluation of the double dis-
placement mechanism for maltotriose biosynthesis shows thatmaltosyl transfer is solely involved. The analysis presented inTable I shows that glucosyl transfer would result initially innonreducing and labeled maltotriose. Mixed maltosyl- and glu-cosyl-transfer would result in initial labeling of both reducingand nonreducing ends ofthe molecule. Data presented has shownvery specifically that initial labeling occurs in the reducing endglucose, and that label distributes at approximately equal ratesbetween the center glucose and nonreducing end glucose.The accumulation of maltose and therefore the hydrolysis of
the maltosyl intermediate following step 1 of the above reactionsoccurred at a faster rate than maltotriose synthesis (cf Fig. 1).Statistically, the maltosyl intermediate would be equally labeledin the two glucose residues. This in fact was the result observedfrom several of the short term 14C02 photosynthesis experimentsconducted at various times ofthe day as shown in Table II. From9 AM through the day until midnight, the symmetry in maltoselabeling was observed during the first 5 min of in vivo 14CO0photosynthesis. In the early morning hours between 3 and 9 AM,
FIG. 3. Distribution of radioactivity in maltotriose from steady statephotosynthesis experiment in '4CO2. Ratios of radioactivity appearing ineach glucose moiety of maltotriose with time.
\ 0
0 \0
-%0° 0 Center glucose G1210 _ OReducing glucose G11
-000
oo Non-reducing glucose G(o
0°-'-Oo000
0. .
797
Dow
nloaded from https://academ
ic.oup.com/plphys/article/74/4/795/6079445 by guest on 12 February 2022
LINDEN AND SCHILLING
O-0- E E*
*0- El 0-E'0---
0-0- EI _-e'
O0- E'
E
0- .,EE E--O I ----
0--!0 E' EI
E,* + I
EI
E
0-0--0 +
E
FIG. 4. Representation of enzyme activesite(s) of the maltose synthase enzyme andreactions leading to biosynthesis of malto-triose in manner consistent with label distri-bution in maltotriose observed during pho-tosynthesis experiments in '4CO2- (0), Glu-cosyl moiety; (0), reducing end glucosyl moi-ety; (-), a- 1,4-glucosyl bond; (-), ,8-1 ,4-glu-cosyl linkage with the enzyme at two activesites E and E'; (*O), 14C label of glucosylmoiety.
t a 1000 sec
*OEI _.~
0-EI
O0E~~~~~I -E' 0 )E E
I
Table I. Probabilistic Analysis ofPotential Double DisplacementMechanismsfor Maltotriose Biosynthesis
Glucosyl moieties of maltotriose are represented as reducing endglucose, G((), center glucose, G(2), and nonreducing end glucose, G(3).Symbols 0 and 1, respectively, represent lack ofand potential for labelingof glucosyl moiety at given relative time with each possibility of malto-syl-, glucosyl-, or mixed maltosyl- and glucosyl-transfer in maltose syn-thase according to model presented in Figure 4.
the maltose formed during the first 5 min of such experimentswas asymmetrically labeled in the nonreducing end. The mobi-lization of photosynthate by the hydrolyzing enzymes in thechloroplast, which were described by Ponratz and Beck (16),leads to the accumulation of glucose and dextrins during thenighttime hours. The resulting pool then allows for the catalyzedreaction of the maltose glucosyltransferase ( 15) between labeledmaltose and glucose and yields asymmetrically labeled maltoseas shown in reaction 8.
aG*-IG* + iBG = aG*-3G + fBG* (8)
These results have demonstrated in vivo compatability ofmaltosesynthase and maltose glucosyltransferase activities. Maltotriosebiosynthesis which occurs by further transglucosylation reactionsof maltose synthase is also compatible with the demonstratedactivity of the purified maltose glucosyltransferase. The latterenzymic activity exhibited glucosyl transfer with maltose andmaltodextrins of DP > 5 but not with either maltotriose or
maltotetraose as donors (15). Schilling was unable to separatemaltose synthase and maltose glucosyltransferase activities byprotein purification procedures (19). The possibility that the twoactivities reside with the same protein raises interesting questionswith regard to substrate specificity and interrelationships of re-
actions.
E
+ l
E
Table II. Distribution ofRadioactivity in Maltose Formed in SpinachLeaves during the First 5 Minutes ofPhotosynthesis in '4CO2 at
Different Times ofDayThe quotients of '4C in nonreducing glucose units to '4C in reducing
end glucose units in maltose are given.
Time of Day '4CO2 Photosynthesis Quotient
min3AM 2 2.19
5 1.3210 1.22
9 AM 2 1.415 1.3410 1.09
12 noon 2 0.955 1.0110 0.98
4 PM 2 1.015 1.0910 0.95
7 PM 2 0.925 0.9510 1.09
12 midnight 2 0.955 1.0210 1.04
The most significant conclusion of the work is the in vivoevidence of oligosaccharide formation from the reducing end.The insertion mechanism given for maltotriose biosynthesis isdistinguished from general oligosaccharide polymerization reac-tions at the nonreducing end, or in the case of sucrosyl oligosac-charides at the end of the chain distal from sucrose in the caseof sucrosyl oligosaccharides (13). The results have also demon-strated that enzyme-bound oligosyl moieties may serve as thedonor molecule for further transglucosylation reactions. Forinstance, the transglucosylation involving a maltosyl-enzyme
(a)/
or
*o-E' ""( b)
798t a losec
O- EI
O.E'
0(-0'. E-_ I
E'
'b-I-P
,1-Pi
t 100 sec
IEI
G- I - P
P.--O
EI,
020 EI
Plant Physiol. Vol. 74, 1984
*G- I P
'11-.Pi
Dow
nloaded from https://academ
ic.oup.com/plphys/article/74/4/795/6079445 by guest on 12 February 2022
MALTOTRIOSE BIOSYNTHESIS
intermediate yielded maltotriose; the transfer of a maltotriosyl-enzyme complex, which would yield maltotetraose, has beendemonstrated (19). Maltotetraose is the smallest oligosaccharidewhich may serve as primer for polymerizations leading to amy-lose. On the other hand, the insertion mechanism described heremay well be the synthetic route of amylose and not just primerfor other polymerization reactions.
Finally, neither maltose synthase (19) nor maltose glucosyltransferase activities (15) were specifically isolated from extractsof spinach chloroplasts-whole leaf extracts were used in eachcase. The implication of maltose synthase in in vivo maltotriosebiosynthesis by isolated spinach chloroplasts by results presentedin this paper localizes the activity to the chloroplast.
Acknowledgments-The authors wish to acknowledge the helpful discussionswith Professor Dexter French and Dr. John F. Robyt of Iowa State University,Department of Biochemistry and Biophysics, where the photosynthesis and productdistribution studies were conducted in 1968 and also with Professor Otto Kandlerof the Botanisches Institut der Universitaet Muenchen where the hypotheses forthe enzymic mechanisms were developed.
LITERATURE CITED
1. AKAZAWA T 1965 Starch, inulin, and other reserve polysaccharides. In J.Bonner JE Varner, eds., Plant Biochemistry, Academic Press, Inc., NewYork, NY, pp. 258-293
2. BASSHAM JA, M CALVIN 1957 The path of carbon in photosynthesis. Prentice-Hall, Englewood Cliffs, New Jersey
3. BASSHAM JA, M KIRK 1964 Photosynthesis of amino acids. Biochim BiophysActa 90: 553-562
4. BASSHAM JA, M KIRK, RG JENSEN 1968 Photosynthesis by isolated chloro-plasts. I. Diffusion of labeled photosynthetic intermediates between isolatedchloroplasts and suspending media. Biochim Biophys Acta 153: 211-218
5. COCKBURN W, PA WALKER, CW BALDRY 1968 The isolation of spinach
chloroplasts in pyrophosphate media. Plant Physiol 43: 1415-14186. FRENCH D 1960 Determination of starch structure by enzymes. Chim Biol Soc
Bull 12: 1677-17007. FRENCH D, JL MANCUSI, M ABDULLAH, GL BRAMMER 1965 Separation of
starch oligosaccharides by high temperature paper chromatography. J Chro-matogr 19: 445-447
8. FRENCH D, AO PULLEY, M ABDULLAH, JC LINDEN 1966 Two-dimensionalpaperchromatography interspersed with reaction on the paper. J Chromatogr24: 271-276
9. HEBER U 1957 Zur Frage der Lokalization von loeslichen Zuckern in derPflanzenzelle. Ber Dtsch Bot Ges 70: 371-381
10. JENSEN RG, JA BASSHAM 1966 Photosynthesis by isolated chloroplasts. ProcNatl Acad Sci USA 56: 1095-1101
11. JUETTNER F, T MATUSCHER 1978 The release of low molecular weight com-pounds by the phytoplankton in a eutrophic lake. Water Res 12: 251-255
12. KANDLER 0 1964 Moeglichkeiten zur Verwendung von 14C fuer chemotax-onomische Untersuchung. Ber Dtsch Bot Ges 77: 62-73
13. KANDLER 0, H HOPF 1980 Oligosaccharides based on sucrose (sucrosyl oligo-saccharides) In PK Stumpf, EE Conn, eds., The Biochemistry of Plants, Vol3. Academic Press, New York, pp. 348-383
14. LINDEN JC, N SCHILLING, H BRACKENHOFER, 0 KANDLER 1975 Asymmetriclabeling of maltose during photosynthesis in "CO2. Z Pflanzenphysiol 76:176-181
15. LINDEN JC, W TANNER, 0 KANDLER 1974 Properties of glucosyltransferaseand glucan transferase from spinach. Plant Physiol 54: 752-757
16. PONGRATZ P, E BECK 1978 Diurnal oscillation ofamylolytic activity in spinachchloroplasts. Plant Physiol 62: 678-690
17. ROBYT JF 1979 Mechanisms involved in the biosynthesis of polysaccharides.Trends Biochem Sci Feb: 47-49
18. SCHILLING N, 0 KANDLER 1975 a-Glucose-l-phosphate, a precursor in thebiosynthesis of maltose in higher plants. Trans Biochem Soc 3: 985-987
19. SCHILLrNG N 1982 Characterization of maltose biosynthesis from a->-glucose-I-phosphate in Spinacia oleracea. L. Planta 154: 87-93
20. TREVELYAN WE, DP PRocrOR, JS HARRISON 1950 Detection of sugars onpaper chromatograms. Nature 166: 444445
21. WALKER GJ, WJ WHELAN 1957 The mechanism of carbohydrase action. 4.The mechanism of D-enzyme action. Biochem J 67: 548-551
799
Dow
nloaded from https://academ
ic.oup.com/plphys/article/74/4/795/6079445 by guest on 12 February 2022