Proc. Nati. Acad. Sci. USA Vol. 91, pp. 12317-12321, December 1994 Biochemistry cDNA cloning of the Sm proteins D2 and D3 from human small nuclear ribonucleoproteins: Evidence for a direct D1-D2 interaction (autoimmunity/anti-Sm antibodies/splicing/protein-protein interactions) THOMAS LEHMEIER*, VERONICA RAKER*, HERBERT HERMANN, AND REINHARD LUHRMANNt Institut ffr Molekularbiologie und Tumorforschung, Philipps-Universitat Marburg, Emil-Mannkopff-Strasse 2, D-35037 Marburg, Germany Communicated by Joan A. Steitz, July 11, 1994 ABSTRACT The major small nuclear ribonucleoproteins (snRNPs) U1, U2, U4/U6, and U5 share a set of common proteins denoted B/B', D1, D2, D3, E, F, and G which play an important part in the biogenesis of the snRNPs. In addition, there is a link between the common proteins and autoimmunity; the three D proteins, together with B/B', are the major autoantigens for the so-called anti-Sm antibodies often produced by patients suffering from systemic lupus erythematosus. Here we describe the characterization of the human proteins D2 and D3 by cDNA cloning and immuno- logical methods. D2 and D3 are encoded by distinct genes and are 118 and 126 amino acids in length, respectively. Both proteins prepared by in vitro translation exhibit Sm epitopes and can be precipitated by anti-Sm autoantibodies. They react differently with various patient sera, in a manner consistent with the reaction pattern on immunoblots of the D proteins isolated from HeLa cells. D1 and D2 synthesized in vitro form specific complexes, a result that is significant for the assembly pathway of the various core proteins into an snRNP's core ribonucleoprotein structure. The D3 protein is homologous to the human D1 protein, showing an overall amino acid sequence identity of 29%, including two regions with over 60% identity. D2 has less than 15% sequence identity with D1 and D3. A data bank search revealed a striking similarity (with more than 40% sequence identity) between human D3 and a Saccharomyces cerevisiae gene, previously published as the 5' flanking gene of yeast pep3 [Preston, R. A., Manolson, M., Becherer, K., Weidenham- mer, E., Kirkpatrick, D., Wright, R. & Jones, E. (1991) Mol. Cell. Biol. 11, 5801-5812], suggesting that this gene encodes the yeast homologue of the human D3 protein. The small nuclear ribonucleoproteins (snRNPs) U1, U2, U4/U6, and U5 are essential for pre-mRNA splicing (1, 2). Their proteins can be classified into two categories: the common proteins, called Sm proteins (see below), which are shared by the four snRNPs, and the snRNP-specific proteins. U1 snRNPs contain three specific proteins de- noted 70k, A, and C. The 17S U2 snRNP particle and the U4/U6/U5 tri-snRNP particle contain more than 10 specific proteins each. These specific proteins are thought to me- diate snRNP particle-specific functions in pre-mRNA splic- ing (3, 4). The complexity of the core ribonucleoprotein (RNP) struc- ture of the snRNPs has only recently been appreciated. It was previously assumed that the core consisted of the proteins B/B', D, E, F, and G bound to the appropriate RNA. However, we have presented evidence that the D protein actually consists of three distinct polypeptides, named ac- cording to their apparent molecular masses: D1 (16 kDa), D2 (16.5 kDa), and D3 (18 kDa) (5). The presence of three D proteins appears to be evolutionarily conserved, as all three could be identified in snRNPs from man, mouse, and Dro- sophila (3, 5). More recently, a 69-kDa protein, which also binds to the core structure of the spliceosomal snRNPs and may therefore be considered to be an additional core protein, has been identified. However, the 69-kDa protein appears to interact transiently with snRNPs (6). Of the many core proteins, only the cDNAs for the E (7), D1 (8-10), and B/B' (11-18) proteins from metazoa have been cloned (for review see ref. 19), while in yeast, only the cDNA for the homologue of human D1 has been published (20). The snRNP core proteins are interesting for various rea- sons. First of all, they are essential for the biogenesis of the snRNP particles. They associate in the cytoplasm with the Sm binding site of newly transcribed U1, U2, U4, and U5 small nuclear RNAs (snRNAs) thereby forming the common core RNP domain (21, 22). Proteins E, F, and G and one or more of the D proteins appear to assemble independently of snRNA into a 6S heterooligomeric protein complex. This complex probably binds to the snRNA Sm site (refs. 23 and 24; for review see ref. 22). The nature of the protein-protein interactions in the core RNP structure of the snRNP is not yet understood. One or more of the core proteins create(s) a nuclear localization signal on the core RNP domain that is essential for the nuclear targeting of snRNP particles (25, 26). In addition, the core proteins probably provide a binding site for a methyltransferase that hypermethylates the m7G cap of snRNA to produce the m3G cap of the native snRNP particle (27). Finally, the core proteins are interesting from a clinical and immunological point of view. Patients suffering from sys- temic lupus erythematosus (SLE) often produce anti-Sm autoantibodies that react with the core proteins of spliceo- somal snRNPs, predominantly with the B and D proteins (28, 29). The core proteins are therefore also called Sm proteins. Of particular diagnostic value for SLE are autoantibodies against the D proteins (30). We have recently shown by immunoblotting that the anti-Sm sera from SLE patients contain populations of anti-Sm autoantibodies that react differentially with the D proteins; some antibodies recognize all of the D proteins, while others only recognize a subset (5). These results indicated that the D proteins are distinct polypeptides; however, the antigenic cross-reactivity sug- gested that the proteins may also share some structural features. As a prerequisite for the characterization of Sm epitopes and also for the further elucidation of the function of the D proteins in the biogenesis of the snRNPs, we have Abbreviations: snRNA, small nuclear RNA; snRNP, small nuclear ribonucleoprotein; RNP, ribonucleoprotein; mAb, monoclonal anti- body; SLE, systemic lupus erythematosus; RIPA, radioimmunopre- cipitation assay; RACE, rapid amplification of cDNA ends; TEMED, N,N,N',N'-tetramethylethylenediamine. *T.L. and V.R. contributed equally to this work. tTo whom reprint requests should be addressed. 12317 The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Transcript
Proc. Nati. Acad. Sci. USAVol. 91, pp. 12317-12321, December 1994Biochemistry
cDNA cloning of the Sm proteins D2 and D3 from human smallnuclear ribonucleoproteins: Evidence for a direct D1-D2 interaction
THOMAS LEHMEIER*, VERONICA RAKER*, HERBERT HERMANN, AND REINHARD LUHRMANNtInstitut ffr Molekularbiologie und Tumorforschung, Philipps-Universitat Marburg, Emil-Mannkopff-Strasse 2, D-35037 Marburg, Germany
Communicated by Joan A. Steitz, July 11, 1994
ABSTRACT The major small nuclear ribonucleoproteins(snRNPs) U1, U2, U4/U6, and U5 share a set of commonproteins denoted B/B', D1, D2, D3, E, F, and G which playan important part in the biogenesis of the snRNPs. Inaddition, there is a link between the common proteins andautoimmunity; the three D proteins, together with B/B', arethe major autoantigens for the so-called anti-Sm antibodiesoften produced by patients suffering from systemic lupuserythematosus. Here we describe the characterization of thehuman proteins D2 and D3 by cDNA cloning and immuno-logical methods. D2 and D3 are encoded by distinct genes andare 118 and 126 amino acids in length, respectively. Bothproteins prepared by in vitro translation exhibit Sm epitopesand can be precipitated by anti-Sm autoantibodies. Theyreact differently with various patient sera, in a mannerconsistent with the reaction pattern on immunoblots of the Dproteins isolated from HeLa cells. D1 and D2 synthesized invitro form specific complexes, a result that is significant forthe assembly pathway of the various core proteins into ansnRNP's core ribonucleoprotein structure. The D3 protein ishomologous to the human D1 protein, showing an overallamino acid sequence identity of 29%, including two regionswith over 60% identity. D2 has less than 15% sequenceidentity with D1 and D3. A data bank search revealed astriking similarity (with more than 40% sequence identity)between human D3 and a Saccharomyces cerevisiae gene,previously published as the 5' flanking gene of yeast pep3[Preston, R. A., Manolson, M., Becherer, K., Weidenham-mer, E., Kirkpatrick, D., Wright, R. & Jones, E. (1991) Mol.Cell. Biol. 11, 5801-5812], suggesting that this gene encodesthe yeast homologue of the human D3 protein.
The small nuclear ribonucleoproteins (snRNPs) U1, U2,U4/U6, and U5 are essential for pre-mRNA splicing (1, 2).Their proteins can be classified into two categories: thecommon proteins, called Sm proteins (see below), whichare shared by the four snRNPs, and the snRNP-specificproteins. U1 snRNPs contain three specific proteins de-noted 70k, A, and C. The 17S U2 snRNP particle and theU4/U6/U5 tri-snRNP particle contain more than 10 specificproteins each. These specific proteins are thought to me-diate snRNP particle-specific functions in pre-mRNA splic-ing (3, 4).The complexity of the core ribonucleoprotein (RNP) struc-
ture ofthe snRNPs has only recently been appreciated. It waspreviously assumed that the core consisted of the proteinsB/B', D, E, F, and G bound to the appropriate RNA.However, we have presented evidence that the D proteinactually consists of three distinct polypeptides, named ac-cording to their apparent molecular masses: D1 (16 kDa), D2(16.5 kDa), and D3 (18 kDa) (5). The presence of three D
proteins appears to be evolutionarily conserved, as all threecould be identified in snRNPs from man, mouse, and Dro-sophila (3, 5). More recently, a 69-kDa protein, which alsobinds to the core structure of the spliceosomal snRNPs andmay therefore be considered to be an additional core protein,has been identified. However, the 69-kDa protein appears tointeract transiently with snRNPs (6). Of the many coreproteins, only the cDNAs for the E (7), D1 (8-10), and B/B'(11-18) proteins from metazoa have been cloned (for reviewsee ref. 19), while in yeast, only the cDNA for the homologueof human D1 has been published (20).The snRNP core proteins are interesting for various rea-
sons. First of all, they are essential for the biogenesis of thesnRNP particles. They associate in the cytoplasm with theSm binding site of newly transcribed U1, U2, U4, and U5small nuclear RNAs (snRNAs) thereby forming the commoncore RNP domain (21, 22). Proteins E, F, and G and one ormore of the D proteins appear to assemble independently ofsnRNA into a 6S heterooligomeric protein complex. Thiscomplex probably binds to the snRNA Sm site (refs. 23 and24; for review see ref. 22). The nature of the protein-proteininteractions in the core RNP structure ofthe snRNP is not yetunderstood. One or more of the core proteins create(s) anuclear localization signal on the core RNP domain that isessential for the nuclear targeting ofsnRNP particles (25, 26).In addition, the core proteins probably provide a binding sitefor a methyltransferase that hypermethylates the m7G cap ofsnRNA to produce the m3G cap of the native snRNP particle(27).
Finally, the core proteins are interesting from a clinical andimmunological point of view. Patients suffering from sys-temic lupus erythematosus (SLE) often produce anti-Smautoantibodies that react with the core proteins of spliceo-somal snRNPs, predominantly with the B and D proteins (28,29). The core proteins are therefore also called Sm proteins.Of particular diagnostic value for SLE are autoantibodiesagainst the D proteins (30). We have recently shown byimmunoblotting that the anti-Sm sera from SLE patientscontain populations of anti-Sm autoantibodies that reactdifferentially with the D proteins; some antibodies recognizeall of the D proteins, while others only recognize a subset (5).These results indicated that the D proteins are distinctpolypeptides; however, the antigenic cross-reactivity sug-gested that the proteins may also share some structuralfeatures. As a prerequisite for the characterization of Smepitopes and also for the further elucidation of the function ofthe D proteins in the biogenesis of the snRNPs, we have
Abbreviations: snRNA, small nuclear RNA; snRNP, small nuclearribonucleoprotein; RNP, ribonucleoprotein; mAb, monoclonal anti-body; SLE, systemic lupus erythematosus; RIPA, radioimmunopre-cipitation assay; RACE, rapid amplification of cDNA ends;TEMED, N,N,N',N'-tetramethylethylenediamine.*T.L. and V.R. contributed equally to this work.tTo whom reprint requests should be addressed.
12317
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
12318 Biochemistry: Lehmeier et al.
isolated and characterized the cDNAs for the human D2 andD3 proteins.A
MATERIALS AND METHODSProtein Purification and Sequencing. The core snRNP pro-
teins D2 and D3 were isolated by purifying HeLa snRNPs andthen fractionating their proteins preparatively on high-TEMED (N,N,N',N'-tetramethylethylenediamine), SDS/12.5% polyacrylamide gels (5). The individual proteins wereelectroeluted from the high-TEMED gels as described pre-viously (5) and cleaved with cyanogen bromide. Separation ofthe peptides by HPLC was followed by peptide sequencingperformed with a gas phase sequencer. The following peptidesequences were obtained for D2: MXTEVPKSGKGK-KKSKPVNKDRYIXK and MFLXGDSVIVVLRN-PLIAGK; and for D3: MSIGV, MSNITVTY, MLKNAPM-LKXMK, MKNKNQGSGXGKAAILKAQVAAXGXGX-GXG and MGXGNIFQKR, where X represents an aminoacid that could not be unequivocally determined.
Oligonucleotide Design. D2-specific oligonucleotides to beused as PCR primers were designed as follows: D2-PCR1,5'-TTGAATTCTAA(G/A)CCNGTNAA(T/C)AA(G/A)-GA-3' (from the peptide sequence KPVNKD; the underlinedsequence is an EcoRI restriction site) and D2-PCR2: 5'-TTCTCGAGTGTCGACTCTTGCCGGCGATGAGCG-GGTT-3' (antisense from the initial DNA sequence informa-tion, see below and Fig. 1, nucleotides 364-384; the under-lined sequences are the restriction sites for Xho I and Sal I,respectively). The oligonucleotides designed from the D3peptides to be used for screening a cDNA library were
Isolation of the cDNA Clones. A AgtlO cDNA libraryconstructed from poly(A)+ mRNA isolated from HeLa S3cell line was screened for the D2 clone by using the rapidamplification of cDNA ends (RACE) PCR method (31). The3' RACE was carried out with the D2-PCR1 primer and anoligo(dT) primer containing a Sal I restriction site at the 5'end. Amplified DNA was fractionated on a 1% low meltingtemperature agarose gel, digested with EcoRI and Sal I, andinserted into pBluescript (Stratagene), and its nucleotidesequence was determined by using the Sequenase DNAsequencing system (United States Biochemical). The DNAsequence information from the appropriately amplified frag-ment (cDNA2a, nucleotides 292-496 in Fig. 1) was used todesign the oligonucleotide D2-PCR2. A 5' RACE usingD2-PCR2 and a vector-specific antisense primer that hybrid-ized to the vector DNA immediately 5' to the insert DNAresulted in the identification of fragment cDNA2b (nucleo-tides 1-394 in Fig. 1). The two cDNA fragments, cDNA2aand cDNA2b, were cleaved at an internal Nae I site presentin both cDNAs (GCCGGC, nucleotides 385-390 in Fig. 1) andthen ligated together. The full-length clone was digested withEcoRI and Sal I and inserted into pBluescript (pBLSD2).A AgtlO cDNA library constructed from human placenta
cDNA was screened with D3-oligol by standard hybridiza-tion screening (32). Plaques identified as potential positivesafter the initial screening with the D3-oligol probe wererescreened with D3-oligo2. Hybridization-positive plaquesfrom this second round of screening underwent a final
tThe sequences reported in this paper have been deposited in theGenBank data base (accession nos. U15008 and U15009).
screening with D3-oligo3. All positive plaques contained thesame insert, which was then inserted into pBluescript(pBLSD3).In Vitro Translation and Radioimmunoprecipitation Assays
(RIPAs). The cDNA for D1 (D45-2, generously provided byS. Hoch, see ref. 8) was inserted into pBluescript (pBLSD1).The plasmids containing the cDNAs for D1, D2, or D3(pBLSD1, pBLSD2, or pBLSD3) were linearized with Sal Iand transcribed with T7 RNA polymerase. One microgram ofin vitro-synthesized mRNA was translated by using a rabbitreticulocyte system according to the manufacturer's instruc-tions (Promega) in the presence of [35S]methionine (Amer-sham). For immunoprecipitation assays, proteins translatedin vitro were incubated first with the appropriate antibody for30 min at room temperature and then were added to 10 Al ofprotein A-Sepharose (Pharmacia), which had been preincu-bated for 2 h with 1% bovine serum albumin in phosphate-buffered saline (PBS, 150 mM NaCl/20 mM sodium phos-phate, pH 8.0). Incubation was continued in a total volume of400 ul of PBS for 2 h at 4°C with constant rotation. TheSepharose-bound antibodies were pelleted and washed fivetimes with 1 ml of IPP (10 mM Tris-HCl, pH 8.0/0.1%Nonidet P-40/150 mM NaCl, unless a different salt concen-tration is stated in the figure legend), with a transfer of theSepharose to a new tube after the second wash. Immuno-precipitates were then fractionated on a high-TEMED, SDS/12.5% polyacrylamide gel, which was then treated withAmplify (Amersham) and analyzed by fluorography.
RESULTS AND DISCUSSIONIsolation of the cDNA Clones for D2 and D3. The core
snRNP proteins D2 and D3 were isolated by purifying HeLasnRNPs and then fractionating their proteins preparativelyon high-TEMED, SDS/polyacrylamide gels (5). Amino acidsequences of various peptides from each protein were ob-tained by direct microsequencing. The amino acid sequenceinformation allowed us to design degenerate oligonucleotidesto be used for the subsequent library screening for cDNAs.The D2 clone was isolated from a HeLa AgtlO cDNA libraryusing the RACE PCR procedure (31). The cDNA clone for D3was identified by screening a human placenta AgtlO cDNAlibrary with 32P-labeled oligonucleotides.The identities ofthe D2 and D3 cDNAs were verified by the
following criteria. First, all of the peptide sequences obtainedfrom each of the native proteins were found within thepredicted amino acid sequences of the cDNA clones (Figs. 1and 2). Second, the D2 and D3 proteins prepared by in vitrotranslation comigrated with the native D2 and D3 proteins
31
91
CGGGAGTGAACGGAGAGCGTAGTGACCATC
ATGAGCCTCCTCAACAAGCCCAAGAGTGAGATGACCCCAGAGGAGCTGCAGAAGCGAGAGM S L L N K P K S E M T P E E L 0 K R E 20
GAGGAGGAATTTAACACCGGTCCACTCTCTGTGCTCACACAGTCAGTCAAGAACAATACCE E E F N T G P L S V L T 0 S V K N N T 40
151 CAAGTGCTCATCAACTGCCGCAACAATAAGAAACTCCTGGGCCGCGTGAAGGCCTTCGAT0 V L I N C R N N K K L L G R V K A F D 60
211 AGGCACTGCAACATGGTGCTGGAGAACGTGAAGGAGATGTGGACTGAGGTACCCAAGAGTR H C N M V L E N V K E M W I V P K.. 80
271 GGCAAGGGCAAGAAGAAGTCCAAGCCAGTCAACAAAGACCGCTACATCTCCAAGATGTTCa K a K K K S K P V N K n R Y S K M F 100
331 CTGCGCGGGGACTCAGTCATCGTGGTCCTGCGGAACCCGCTCATCGCCGGCAAGTAGGGGL R G D S V I V V R N P A 0 K END 118
FIG. 1. Nucleotide and predicted amino acid sequences of D2.The amino acid sequences obtained by microsequencing the nativeprotein are underlined (peptide sequences are also listed in Materialsand Methods). The potential poly(A)-addition signal is shown in boldtype. Nucleotides are numbered on the left and amino acids arenumbered on the right.
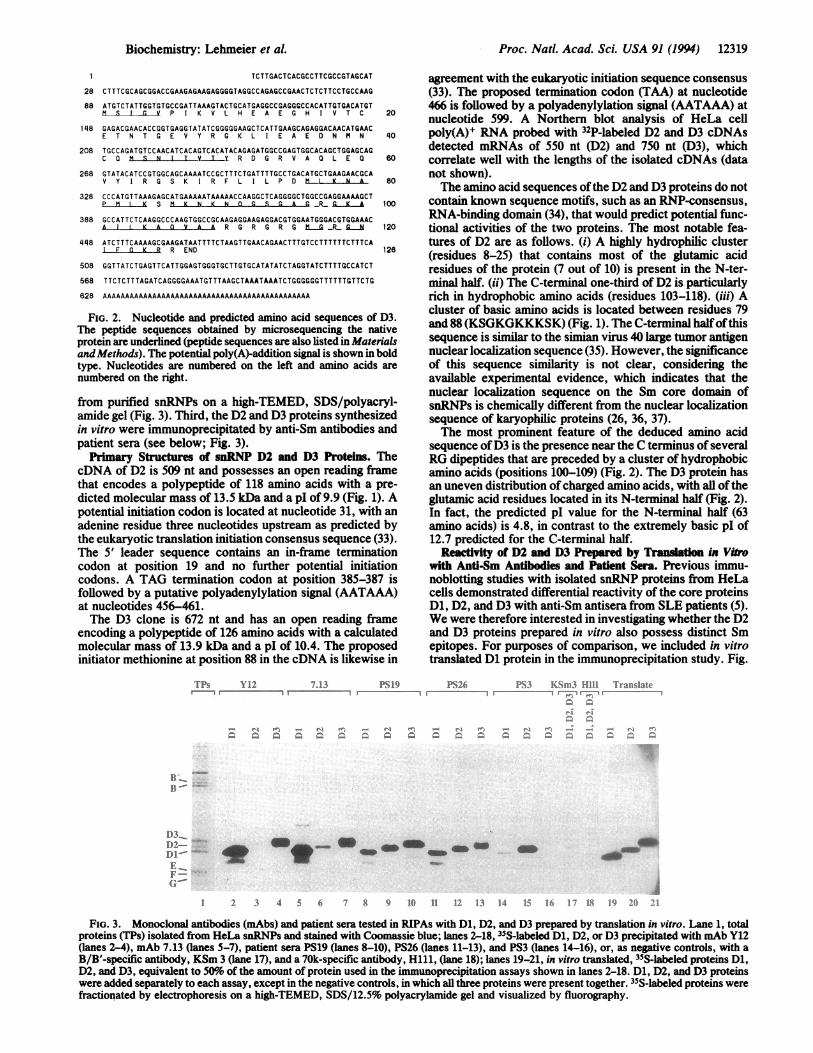
FIG. 2. Nucleotide and predicted amino acid sequences of D3.The peptide sequences obtained by microsequencing the nativeprotein are underlined (peptide sequences are also listed in Materialsand Methods). The potential poly(A)-addition signal is shown in boldtype. Nucleotides are numbered on the left and amino acids arenumbered on the right.
from purified snRNPs on a high-TEMED, SDS/polyacryl-amide gel (Fig. 3). Third, the D2 and D3 proteins synthesizedin vitro were immunoprecipitated by anti-Sm antibodies andpatient sera (see below; Fig. 3).Primary Structures of snRNP D2 and D3 Proteins. The
cDNA of D2 is 509 nt and possesses an open reading framethat encodes a polypeptide of 118 amino acids with a pre-dicted molecular mass of 13.5 kDa and a pI of 9.9 (Fig. 1). Apotential initiation codon is located at nucleotide 31, with anadenine residue three nucleotides upstream as predicted bythe eukaryotic translation initiation consensus sequence (33).The 5' leader sequence contains an in-frame terminationcodon at position 19 and no further potential initiationcodons. A TAG termination codon at position 385-387 isfollowed by a putative polyadenylylation signal (AATAAA)at nucleotides 456-461.The D3 clone is 672 nt and has an open reading frame
encoding a polypeptide of 126 amino acids with a calculatedmolecular mass of 13.9 kDa and a pI of 10.4. The proposedinitiator methionine at position 88 in the cDNA is likewise in
agreement with the eukaryotic initiation sequence consensus(33). The proposed termination codon (TAA) at nucleotide466 is followed by a polyadenylylation signal (AATAAA) atnucleotide 599. A Northern blot analysis of HeLa cellpoly(A)+ RNA probed with 32P-labeled D2 and D3 cDNAsdetected mRNAs of 550 nt (D2) and 750 nt (D3), whichcorrelate well with the lengths of the isolated cDNAs (datanot shown).The amino acid sequences ofthe D2 and D3 proteins do not
contain known sequence motifs, such as an RNP-consensus,RNA-binding domain (34), that would predict potential func-tional activities of the two proteins. The most notable fea-tures of D2 are as follows. (i) A highly hydrophilic cluster(residues 8-25) that contains most of the glutamic acidresidues of the protein (7 out of 10) is present in the N-ter-minal half. (ii) The C-terminal one-third of D2 is particularlyrich in hydrophobic amino acids (residues 103-118). (iii) Acluster of basic amino acids is located between residues 79and 88 (KSGKGKKKSK) (Fig. 1). The C-terminal halfofthissequence is similar to the simian virus 40 lare tumor antigennuclearlocalization sequence (35). However, the significanceof this sequence similarity is not clear, considering theavailable experimental evidence, which indicates that thenuclear localization sequence on the Sm core domain ofsnRNPs is chemically different from the nuclear localizationsequence of karyophilic proteins (26, 36, 37).The most prominent feature of the deduced amino acid
sequence ofD3 is the presence near the C terminus of severalRG dipeptides that are preceded by a cluster of hydrophobicamino acids (positions 100-109) (Fig. 2). The D3 protein hasan uneven distribution ofcharged amino acids, with all oftheglutamic acid residues located in its N-terminal half (Fig. 2).In fact, the predicted pI value for the N-terminal half (63amino acids) is 4.8, in contrast to the extremely basic pI of12.7 predicted for the C-terminal half.
Reactivity of D2 and D3 Prepared by Translation in Vitrowith Anti-Sm Antibodies and Patient Sera. Previous immu-noblotting studies with isolated snRNP proteins from HeLacells demonstrated differential reactivity ofthe core proteinsD1, D2, and D3 with anti-Sm antisera from SLE patients (5).We were therefore interested in investigating whether the D2and D3 proteins prepared in vitro also possess distinct Smepitopes. For purposes of comparison, we included in vitrotranslated D1 protein in the immunoprecipitation study. Fig.
FIG. 3. Monoclonal antibodies (mAbs) and patient sera tested in RIPAs with D1, D2, and D3 prepared by translation in vitro. Lane 1, totalproteins (TPs) isolated from HeLa snRNPs and stained with Coomassie blue; lanes 2-18, 35S-labeled D1, D2, or D3 precipitated with mAb Y12(lanes 2-4), mAb 7.13 (lanes 5-7), patient sera PS19 (lanes 8-10), PS26 (lanes 11-13), and PS3 (lanes 14-16), or, as negative controls, with aB/B'-specific antibody, KSm 3 (lane 17), and a 70k-specific antibody, Hill, (lane 18); lanes 19-21, in vitro translated, 35S-labeled proteins D1,D2, and D3, equivalent to 50%o of the amount of protein used in the immunoprecipitation assays shown in lanes 2-18. D1, D2, and D3 proteinswere added separately to each assay, except in the negative controls, in which all three proteins were present together. 35S-labeled proteins werefractionated by electrophoresis on a high-TEMED, SDS/12.5% polyacrylamide gel and visualized by fluorography.
Biochemistry: Lehmeier et al.
An_
12320 Biochemistry: Lehmeier et al.
3 shows typical examples of immunoprecipitation patternsobserved with anti-Sm patient sera preselected for reactivitywith one or more ofthe three snRNPD proteins. Several seraprecipitated all three D proteins-for example, sera PS19 andPS26 (lanes 8-13, Fig. 3). A few sera, such as serum PS3 (Fig.3, lanes 14-16), precipitated predominantly the D2 protein.Interestingly, the standard monoclonal anti-Sm antibody Y12precipitated D1 and D3 but not, or only to a limited degree,D2 (Fig. 3, lanes 2-4). Anti-Sm mAb 7.13, like mAb Y12,predominantly precipitates D1 and D3 with only a weakreactivity with D2 (Fig. 3, lanes 5-7). Consistent with im-munoblotting studies with D proteins isolated from HeLasnRNPs (5), these data show clearly that the in vitro trans-lation products D1, D2, and D3 possess distinct Sm epitopes,which either are unique to a single protein (as in the case ofD2) or are shared by subsets of the three D proteins (like theY12 epitope).
Specific Interaction Between Proteins D1 and D2 Preparedby Translation in Vitro. During immunoprecipitation studiesof the in vitro translated D proteins with patient sera, wenoticed a propensity of D1 and D2 proteins to interact witheach other. We have investigated the specificity of thisprotein complex formation in more detail by coimmunopre-cipitation analysis, taking advantage of the observation thatmAb Y12 precipitates D1 and D3 but not D2 (Fig. 3). WhenD1 and D2 were mixed prior to addition of mAb Y12, theantibody precipitated not only D1, as expected, but also D2(Fig. 4, lanes 4-6). In contrast, when D2 and D3 were testedtogether, D2 was not coprecipitated with D3 by mAb Y12above background (Fig. 4, compare lane 7 with lane 8). Thenegative control, mAb H111 specific for the U1 70k protein,showed no precipitation of D1, D2, or D3. Thus, these dataindicate that a specific interaction occurs between D2 andD1, which is not due to a general binding of D2 with otherbasic proteins. This finding is reinforced by the observationthat the association between D2 and Dl is observed even atsalt concentrations up to at least 350mM (Fig. 4, lane 5). Theaffinity of mAb Y12 for D1 was diminished at a salt concen-tration of 500 mM, so that the stability of the D1-D2 inter-action at 500mM NaCl cannot be determined (Fig. 4, lane 6).Also, increasing the amount of D2 in the assay while main-
'Tranislate
salt conIc. 1)2 cone.
LI2 x<< lX- IX X0 I Xlx XXXX1)1lX ''XXX XiXX _x _ XL.
taining a constant amount of D1 does not lead to a linearincrease of coimmunoprecipitated D2 (Fig. 4, lanes 11-14),which suggests that D1-D2 complex formation is stoichio-metric.The interaction between D1 and D2 is of interest when one
considers that an RNA-independent 6S complex consisting ofproteins E, F, G, and one or more of the D proteins has beenpreviously described (see Introduction). The protein-proteininteraction data presented here would suggest that the RNA-free, protein complex containing E, F, and G also containsboth D1 and D2 or D3 alone.
Proteins D1 and D3 Are Homologous. Since several anti-Smantibodies cross-react with two or more of the three Dproteins (see above and ref. 5), we were interested in inves-tigating possible homology among the three D proteins. Infact, an alignment of the amino acid sequence of the humanD1 and D3 proteins revealed an overall sequence identity of29% (Fig. 5A) with two stretches of clear homology (about60% sequence identity) as detailed below. At the C terminus,both proteins share a region rich in arginine and glycine(containing 11 of 17 identical residues). While the RG dipep-tide motif is repeated nine times in D1 (8), only four repeatsare found in D3 (Fig. SA). We note that similarRG repeats arealso present in the EBNA-1 and EBNA-2 proteins from theEpstein-Barr virus (39, 40) as well as in the yeast transcrip-tion factor SNF-2 (41).The second extensive region ofhomology between D1 and
D3 is located at the beginning of the C-terminal halves ofbothproteins. Interestingly, 13 of the 21 amino acids (62%) in theregions spanned by amino acids 54-74 ofD1 and 57-77 ofD3are identical (Fig. SA). Clearly, the significant homologybetween D1 and D3 suggests that the two genes may have hada common ancestor that duplicated and diverged duringevolution.
In contrast to the sequence similarities observed betweenthe D1 and D3 proteins, the D2 protein exhibits little overallidentity with either D1 (11%) or D3 (13%) (not shown).
A hDl1: hD3A10 .20 .30 .40 460 to0hD 1IKLVRFLNKLSHETVTIELKNGTOVHGTITGVDVSMNTHLKAVKNTLKNREPVOLETLSIRGN:: L VT E C :G : : MN : ::T :: OLE : IRG
FIG. 4. Interaction between D1 and D2, prepared by translationin vitro, as shown by RIPAs with mAb Y12. Lanes 1-3, in vitrotranslated, 35S-labeled proteins D1, D2, and D3, equivalent to 30%ofthe amount used in the immunoprecipitation assays shown in lanes4-11; lanes 4-6, RIPAs with D1 and D2 and increasing salt concen-tration in the wash buffer (150 mM NaCl, lane 4; 300 mM NaCl, lane5; and 500 mM NaCl, lane 6); lane 7, RIPA with D2 and D3; lane 8,RIPA with D2; lane 9, RIPA with D1; lane 10, RIPA with D1, D2, andD3 and the mAb H111, which is directed against the Ul-specific 70kprotein; lanes 11-13, RIPA with D1 and increasing amounts ofD2 andmAb Y12. The amount of D2 was increased either 2-fold (lane 12) or3-fold (lane 13) compared to the amount used in the assay shown inlane 11; lane 14, RIPA with three times the amount of D2 as in lane11 and no D1 or D3. 35S-labeled precipitated proteins were fraction-ated by electrophoresis on a high-TEMED, SDS/12.5% polyacryl-amide gel and visualized by fluorography.
C hD3:y3 .10 420 430 .40 S50 f60yD3 TMNGIPVKLLNEAOGHIVSLELTTGATYRGKLVESEDSKNVOLRDVIATEPOCAVTH1MDO
G: P: K: L EA: CHIV: E TG YRGKL: E ED tHN 0: :: T : C V :: OIM lSIGVPIKVLHEAEGHIVTCETNTGEVYRGKLIEAEDNMNCOMISNITVTYRDCRVADLED
FIG. 5. Homology between core proteins. Identical residues areshown in the middle row; residues belonging to one of the followinggroups are considered to be similar: (L V I M), (F Y W), (S T), (AG), (K R H), (ED Q N); as indicated by a colon. (A) Human D1 (hD1;Sm-D1, ref. 8) and human D3 (hD3); (B) human D2 (hD2) and humanB/B' (hB; Sm-B/B', ref. 11); (C) human D3 (hD3) and the putativeyeast D3 (yD3; the 5' flanking sequence to thepep3 gene, ref. 38); (D)human D3 (hD3) and yeast D1 (yDl; Smdpl, ref. 20).
Proc. Natl. Acad Sci. USA 91 (1994)
Y'12 11111 Y I -)
Proc. Natl. Acad. Sci. USA 91 (1994) 12321
However, the C-terminal portion of D2 shows significantamino acid identity with the N-terminal region of the humancore proteins B/B' as shown in Fig. 5B. A span of 29 residues(29-57 in B/B' and 54-84 in D2) has a 41% identity (Fig. 5B).The various degrees of identity found among subsets ofthe
human D1, D2, D3, and B/B' proteins are ofpotential interestfor mapping Sm epitopes on these proteins. For example, thecross-reactivity of mAb Y12 with the D1 and D3 proteinscorrelates with the homology found between these twoproteins, making the highly homologous regions shared be-tween D1 and D3 (see above) prime candidates for identifi-cation as the mAb Y12 epitope(s). Mutation analysis of thehuman D1 polypeptide has recently provided evidence thatthe RG-rich C terminus of D1 makes up a large part of themAb Y12 epitope (38). Epitope-mapping studies of D3,however, suggest that this RG-rich region in D3 plays only aminor role in mAb Y12 recognition, while a region located inthe middle of D3 makes up the major part of the epitope (H.Brahms, V.R., and R.L., unpublished data).The Human D3 Protein Possesses Striking Homolgy with a
Yeast Protein. A data-bank search with the human D3 se-quence revealed a protein sequence from Saccharomycescerevisiae, published as the flanking gene 5' to the pep3 gene(42) that displayed conspicuous similarity with D3. This genehas an open reading frame with the capacity to encode a 101amino acid protein with a predicted molecular mass of 11.2kDa and a pI of 10.0. Fig. 5C shows that the two proteinspossess significant similarity (>40% sequence identity) overalmost their entire length, except at their C-termini, wherethe RG-rich domain is missing from the yeast sequence.Significantly, several sequence blocks, 3-5 amino acids inlength, are 100% conserved. The high level of conservationbetween this protein and the human D3 polypeptide suggeststhat this could be a yeast D3 homologue. Indeed, recentexperiments provide evidence that this protein behaves as afunctional common snRNP protein in yeast (J. Roy, B.Zheng, B. Rymond, and J. Woolford, personal communica-tion). Recently, a homologue of the human D1 protein,denoted Smdpl, has been identified in the yeast S. cerevisiae(20). Smdpl has an amino acid sequence identity of35% withthe human D1 protein. The human D1 gene is able tofunctionally replace the yeast Smdpl gene (43). We note thatthe degree of identity between human D3 and the proteinencoded by the pep3 flanking gene is even greater than thatbetween human D1 and the yeast homologue of D1, Smdpl.The human D3 protein also displays significant sequence
similarity with the yeast Smdpl protein (21% identity), witha particularly high degree of sequence identity between theregions 56-77 in D3 and 81-101 in Smdpl (Fig. SD). Given thehigh degree of evolutionary conservation between yeast andman now observed for two snRNP core proteins, it istempting to suggest that the entire snRNP core structure maybe conserved between these species.We thank J. Steitz, D. Williams, S. Hoch, W. van Venrooi, and
H. Peter for their generous gifts of mAbs and patient sera, and wethank S. Hoch additionally for the D45-2 cDNA. We are grateful toK. Foulaki for advice on sequencing peptides with a gas-phasesequencer, U. Grundmann for the gift of a cDNA library, as well asW. Hackl and J. Lauber for their help throughout this work. Wethank I. Ochsner, D. Meyer, and M. Wicke for expert technicalassistance and C. Will, H. Brahms, and C. Marshallsay for theircritical reading of this manuscript. We also thank J. Woolford and B.Rymond for communicating results prior to publication. This workwas supported by grants from the Bundesministeruim fur Forschungund Technologie, the Deutsche Forschungsgemeinschaft, and theFonds der Chemischen Industrie.1. Steitz, J. A., Black, D. L., Gerke, V., Parker, K. A., Krfmer, A.,
Frendewey, D. & Keller, W. (1988) in Structure and Function ofMajor and Minor Small Nuclear Ribonucleoprotein Particles, ed.Birnstiel, M. L. (Springer, Berlin), pp. 115-154.
2. Moore, M. J., Query, C. C. & Sharp, P. A. (1993) in TheRNA World(Cold Spring Harbor Lab. Press, Plainview, NY), pp. 303-357.
3. Ldhrmann, R., Kastner, B. & Bach, M. (1990) Biochim. Biophys.Acta 1087, 265-292.
4. Behrens, S.-E., Tyc, K., Kastner, B., Reichelt, J. & LUhrmann, R.(1993) Mol. Cell. Biol. 13, 307-319.
5. Lehmeier, T., Foulaki, K. & Ltlhrmann, R. (1990) Nucleic AcidsRes. 18, 6475-6484.
6. Hackl, W., Fischer, U. & Luhrmann, R. (1993) J. Cell Biol. 124,261-272.
7. Stanford, D. R., Kehl, M., Perry, C. A., Holicky, E. L., Harvey,S. E., Rolhetter, N. M., Rehder, K., Jr., Ldhrmann, R. & Wieben,E. D. (1988) Nucleic Acids Res. 16, 10593-10605.
8. Rokeach, L. A., Haselby, J. A. & Hoch, S. A. (1988) Proc. Natd.Acad. Sci. USA 85, 4832-4836.
9. Renz, M., Heim, C., Braunling, O., Czichos, A., Wieland, C. &Seelig, H. P. (1989) Clin. Chem. 35, 1861-1863.
10. Cohen, P. L., Mitsuda, T. & Eisenberg, R. (1991) Mol. Biol. Rep.15, 158.
11. Rokeach, L. A., Jannatipour, M., Haselby, J. A. & Hoch, S. 0.(1989) J. Biol. Chem. 264, 5024-5030.
12. McAllister, G., Poby-Shemkovitz, A., Amara, S. G. & Lerner,M. R. (1989) EMBO J. 8, 1177-1181.
13. Schmauss, C., McAllister, G., Ohosone, Y., Hardin, J. A. &Lerner, M. R. (1989) Nucleic Acids Res. 17, 1733-1743.
14. Li, S., Klein, E. S., Russo, A. F., Simmons, D. M. & Rosenfeld,M. G. (1989) Proc. Natd. Acad. Sci. USA 86, 9778-9782.
15. Sharpe, N. G., Williams, D. G., Howarth, D. N., Coles, B. &Latchman, D. S. (1989) FEBS Lett. 250, 585-590.
16. van Dam, A., Winkel, I., Z4lstra-Baalbergen, J., Smeenk, R. &Cuypers, H. T. (1989) EMBO J. 8, 3853-3860.
17. Elkon, K. B., Hines, J. J., Chu, J.-L. & Parnassa, A. (1990) J.Immunol. 145, 636-643.
18. Ohosone, Y., Mimori, T., Griffith, A., Akizuki, M., Homma, M.,Craft, J. & Hardin, J. A. (1989) Proc. Natl. Acad. Sci. USA 86,4249-4253.
19. Rokeach, L. A. & Hoch, S. 0. (1992) Mol. Biol. Rep. 16, 165-174.20. Rymond, B. C. (1993) Proc. Natl. Acad. Sci. USA 90, 848-852.21. Mattaj, I. W. (1988) in Structure and Function ofMajor and Minor
Small Nuclear Ribonucleoprotein Particles, ed. Birastiel, M. L.(Springer, Berlin), pp. 100-114.
22. Zieve, G. W. & Sauterer, R. A. (1990) CRC Crit. Rev. Biochem.Mol. Biol. 25, 1-46.
23. Fisher, D. E., Conner, G. E., Reeves, W. H., Wisniewolski, R. &Blobel, G. (1985) Cell 42, 751-758.
24. Sauterer, R. A., Goyal, A. & Zieve, G. W. (1990) J. Biol. Chem.265, 1048-1058.
25. Mattaj, I. W. & De Robertis, E. M. (1985) Cell 40, 111-118.26. Fischer, U., Sumpter, V., Sekine, M., Satoh, T. & Lihrmann, R.
(1993) EMBO J. 12, 573-583.27. Plessel, G., Fischer, U. & LUhrmann, R. (1994) Mol. Cell. Biol. 14,
4160-4172.28. Lerner, E. A., Lerner, M. R., Janeway, J. A., Jr., & Steitz, J. A.
(1981) Proc. Natl. Acad. Sci. USA 78, 2737-2741.29. Habets, W. J., Berden, J. H. M., Hoch, S. 0. & van Venrooj,
W. J. (1985) Eur. J. Immunol. 15, 992-997.30. Tan, E. M. (1989) Adv. Immunol. 44, 93-152.31. Frohman, M. A., Dush, M. K. & Martin, G. R. (1988) Proc. Natl.
Acad. Sci. USA 85, 8998-9002.32. Davis, L. G., Dibner, M. D. & Battey, J. F. (1986) in Basic Meth-
ods in Molecular Biology (Elsevier, New York), pp. 130-205.33. Kozak, M. (1986) Cell 44, 283-292.34. Swanson, M. S. & Dreyfuss, G. (1988) Mol. Cell. Biol. 8, 2237-2241.35. Kalderon, D., Roberts, B. L., Richardson, W. D. & Smith, A. E.
(1984) Cell 39, 499-509.36. Michaud, N. & Goldfarb, D. (1992) J. Cell Biol. 116, 851-861.37. Fischer, U., Darzynkiewicz, E., Tahara, S. M., Dathan, N. A.,
Lfhrmann, R. & Mattaj, I. W. (1991) J. Cell Biol. 113, 705-714.38. Hirakata, M., Craft, J. & Hardin, J. A. (1993) J. Immunol. 150,
3592-3601.39. Baer, R., Bankier, A. T., Biggin, M. D., Deininger, P. L., Farrell,
P. J., Gibson, T. V., Hatfull, G., Hudson, G. S., Satchwell, S. C.,Sequin, C., Tuffnell, P. S. & Barrell, B. G. (1984) Nature (London)310, 207-211.
40. Cohen, J. I., Wang, F. & Kieff, E. (1991) J. Virol. 65, 2545-2554.41. Laurent, B. C., Treitel, M. A. & Carlson, M. (1991) Proc. Natl.
Acad. Sci. USA 88, 2687-2691.42. Preston, R. A., Manoloson, M. F., Becherer, K., Weidenhammer,
E., Kirkpatrick, D., Wright, R. & Jones, E. W. (1991) Mol. Cell.Biol. 11, 5801-5812.
43. Rymond, B. C., Rokeach, L. A. & Hoch, S. 0. (1993) NucleicAcids Res. 21, 3501-3505.