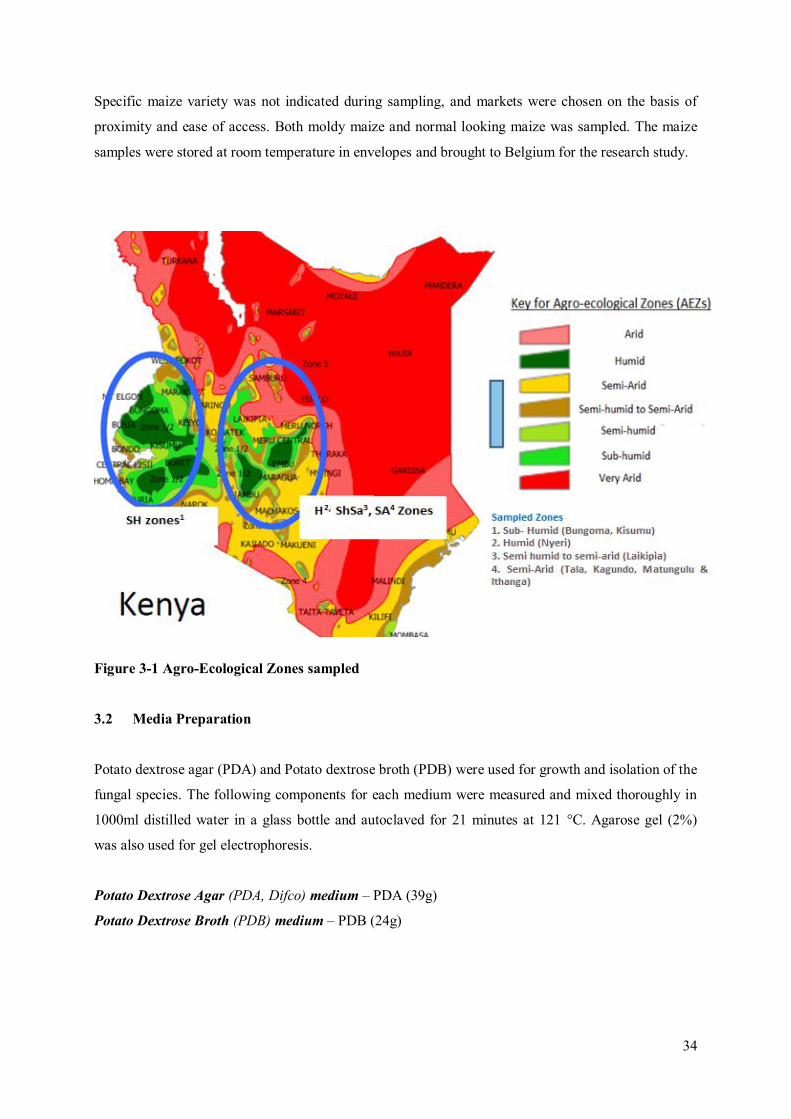
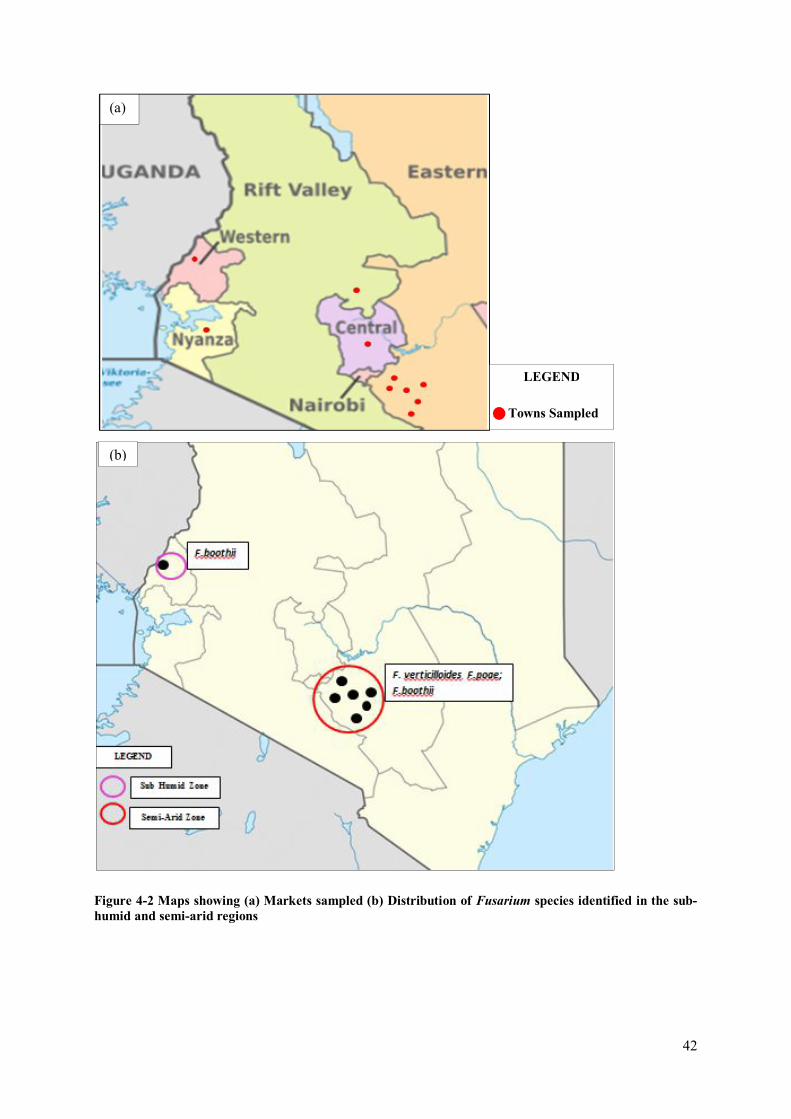
Faculty of Bioscience Engineering Academic year 2014 – 2015 Occurrence of Mycotoxigenic Fungi in Maize from Food Commodity Markets in Kenya Evalyne Nyakio Kibe Promoters: Prof. dr. ir. Monica Hofte Master’s dissertation submitted in fulfillment of the requirements for the degree of Master of Science in Nutrition and Rural Development, Main subject: Public Health Nutrition
Transcript
Faculty of Bioscience Engineering
Academic year 2014 – 2015
Occurrence of Mycotoxigenic Fungi in Maize from Food
Commodity Markets in Kenya
Evalyne Nyakio Kibe
Promoters: Prof. dr. ir. Monica Hofte
Master’s dissertation submitted in fulfillment of the requirements for the degree of Master of Science in Nutrition and Rural Development,
Main subject: Public Health Nutrition
Copyright
“All rights reserved. The author and the promoters permit the use of this Master’s Dissertation for consulting purposes and copying of parts for personal use. However, any other use falls under the limitations of copyright regulations, particularly the stringent obligation to explicitly mention the source when citing parts out of this Master’s dissertation.”
Chronic dietary exposure to Fusarium toxins can cause a variety of toxic effects in both humans and
animals. Among the identified FUMs, fumonisin B1 (FB1) is the most prevalent with proven
immunotoxic, hepatotoxic, neurotoxic and nephrotoxic effects (IARC, 1993; 2002). ZEA is an
estrogenic toxin may lead to different changes in the reproductive system while DON is known to
have an immunotoxic effect and may affect changes in brain neurochemicals. When T-2 and HT-2
toxins are ingested, they can influence the incidence of several effects: nausea, abdominal pain,
dizziness, dermal necrosis, inhibition of protein synthesis (CAST, 2003).
18
2.4.2 Aflatoxins
Aflatoxins are organic chemical compound derivatives formed by a polyketide pathway. The fungi
responsible for these toxins are Aspergillus flavus and Aspergillus parasiticus. Aspergillus bombycis,
Aspergillus ochraceoroseus, Aspergillus nomius, and Aspergillus pseudotamari are also aflatoxin
producing species but are encountered less frequently (Richard, 2009; Peterson et al., 2001).
Aflatoxins occur mostly in tropical regions with high humidity and temperature, and they accumulate
post-harvest when food commodities are stored under conditions that promote fungal growth. The hot
and humid tropical climate in SSA provides ideal conditions for growth of toxigenic Aspergillus spp.
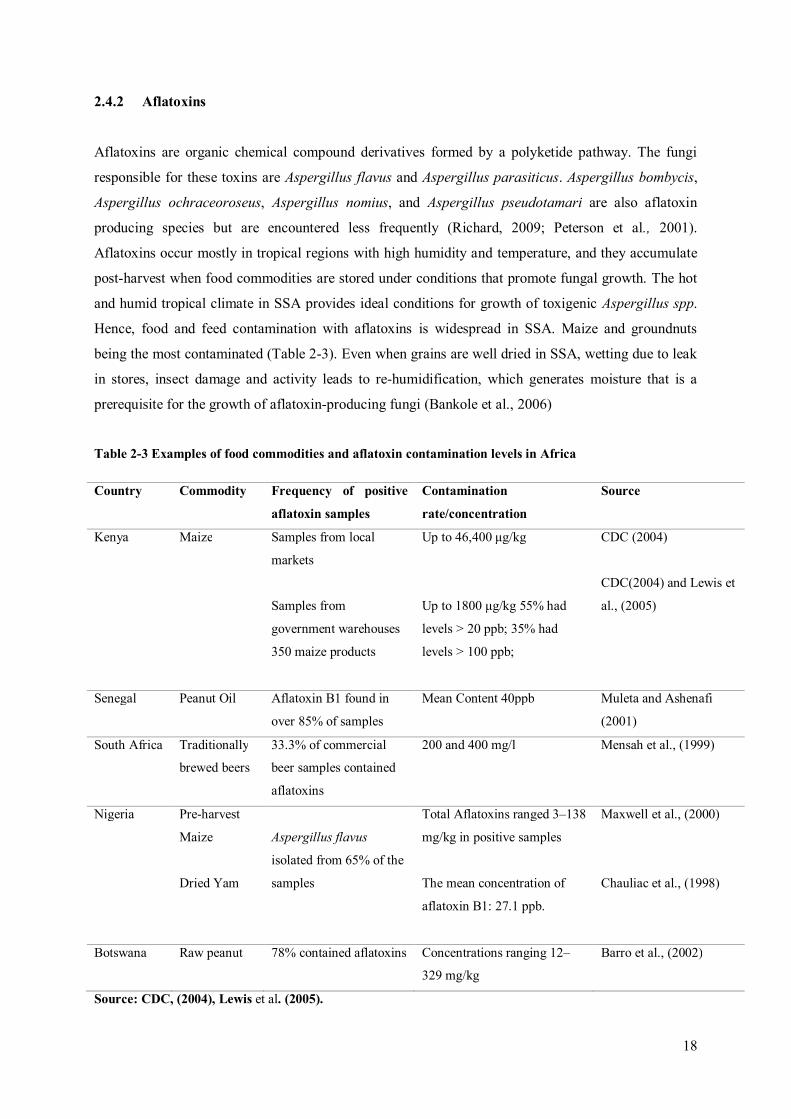
Hence, food and feed contamination with aflatoxins is widespread in SSA. Maize and groundnuts
being the most contaminated (Table 2-3). Even when grains are well dried in SSA, wetting due to leak
in stores, insect damage and activity leads to re-humidification, which generates moisture that is a
prerequisite for the growth of aflatoxin-producing fungi (Bankole et al., 2006)
Table 2-3 Examples of food commodities and aflatoxin contamination levels in Africa
Country Commodity Frequency of positive
aflatoxin samples
Contamination
rate/concentration
Source
Kenya Maize Samples from local
markets
Samples from
government warehouses
350 maize products
Up to 46,400 μg/kg
Up to 1800 μg/kg 55% had
levels > 20 ppb; 35% had
levels > 100 ppb;
CDC (2004)
CDC(2004) and Lewis et
al., (2005)
Senegal Peanut Oil Aflatoxin B1 found in
over 85% of samples
Mean Content 40ppb Muleta and Ashenafi
(2001)
South Africa Traditionally
brewed beers
33.3% of commercial
beer samples contained
aflatoxins
200 and 400 mg/l Mensah et al., (1999)
Nigeria Pre-harvest
Maize
Dried Yam
Aspergillus flavus
isolated from 65% of the
samples
Total Aflatoxins ranged 3–138
mg/kg in positive samples
The mean concentration of
aflatoxin B1: 27.1 ppb.
Maxwell et al., (2000)
Chauliac et al., (1998)
Botswana Raw peanut 78% contained aflatoxins Concentrations ranging 12–
329 mg/kg
Barro et al., (2002)
Source: CDC, (2004), Lewis et al. (2005).
19
Aflatoxins have been implicated in acute aflatoxicosis, carcinogenicity, growth retardation, neonatal
jaundice and immunological suppression in humans (Figure 2-7). The toxigenic ability of aflatoxins
differs qualitatively and quantitatively depending on strain within each aflatoxigenic species (Okoth et
al., 2012). The four major aflatoxins are namely B1, B2, G1 and G2 with the classification based on
their fluorescence under UV light and relative chromatographic mobility during thin-layer
chromatography (Bhat et al., 2010). Aflatoxin B1 is reported as the most toxic in the aflatoxins group,
Research indicates that it is the most potent chemical liver carcinogen known to be naturally occurring.
Specific P450 enzymes in the liver metabolize aflatoxin into a reactive oxygen species (aflatoxin-8,9-
epoxide), which can then bind to proteins and cause acute toxicity (aflatoxicosis) or to DNA and
induce liver cancer (Wild and Gong 2010; Wu and Khlangwiset 2010; Hamid et al., 2013).
Figure 2-7 Aflatoxin and disease pathways in humans (Wu, 2010)
20
2.4.3 Ochratoxins
Ochratoxins are nephrotoxins produced by a diverse species of molds and were first described in 1965.
Ochratoxins ingestion through dietary exposure represents a serious health issue and is associated with
several human and animal diseases including porcine nephropathy, Human Endemic Nephropathies
and urinary tract tumors in humans. OTA exposure causes a disease known ochratoxicosis, whose
primary target is the kidney. As evidenced by epidemiological studies, OTA may be involved in the
pathogenesis of different forms of human nephropathies, including kidney cancer (Marquardt &
Frohlich 1992; Ringot et al. 2006; Pfohl-Leszkowicz & Manderville 2007).
Tumour incidence data from long-term animal studies also provide reasons for concern about the
effect of OTA exposure on the human population. Due to these reasons, OTA was classified as a
possible carcinogen (Group 2B) to humans by The International Agency for Research on Cancer
(IARC 1993). They are immune-suppressive nature, teratogenic and have fertility inhibition,
mutagenic and carcinogenic effects. Ochratoxins are common in cereals, and other starch rich foods
and also can be found in coffee, spices and dried fruits (Zinedine et al., 2007). This toxic compound It
is produced by fungi namely Aspergillus and Penicillium genera which occurs in wide range of
products (Ruadrew et al., 2013), and they grow in wide range of conditions i.e. substrate, pH,
temperature, and moisture. Some crops contaminated by Ochratoxin A are medical herbs, coffee,
cocoa, oats, wheat, and nuts. Also, fresh produce like tomatoes and animal products such as cheese,
and meat from animals feed on contaminated grains (Haighton et al., 2012).
21
2.5 Mycotoxin Problem in Africa
Worldwide, crops are affected by fungal growth, and this not only does it have serious economic
consequences, but also enormous health effects for both human and animals due to the contamination
of food and feed with mycotoxins. Mycotoxin formation may begin in pre-harvest infected in the field
and be continued or commence postharvest due to improper postharvest practices (Logrieco et al.,
2003; Wagacha and Muthomi, 2008). Environmental conditions leading to fungal proliferation include
high temperature and humidity, monsoons, unseasonal rains during harvest and floods. Also, poor
harvesting and storage practices and improper transportation, marketing, and processing also
contribute to fungal growth. These climatic conditions, as well as the improper postharvest practices,
are characteristic in most parts of Africa. Hence, exposure to mycotoxins is high as the diets in these
countries consist mainly of crops (maize) susceptible to toxigenic fungi and consequently their
produced mycotoxins (Lewis et al., 2005).
Losses from rejected shipments and lower prices for inferior quality can be devastating for developing
countries export markets. Direct costs to farmers include reduced income as a result of losses in yield,
low prices for poor quality products, increased livestock mortality and reduced livestock productivity,
fertility and immunity (Wagacha and Muthomi, 2008). An important additional effect could be the
cost of reduced labor force due to illness and costs from hospitalization or other health care services as
a consequence of acute exposure to the toxins. Hence, strategies that will lower mycotoxins levels in
foods will not only reduce costs in health care but also provide better income and international export
opportunities for low and middle-income countries (Bryden, 2007).
Management of mycotoxins contamination usually will include good agricultural practices during
production, harvest, and storage. These constitute of practices like crop rotation, pest control,
irrigation, proper drying and removal of damaged kernels. Mycotoxins are relatively very stable;
although certain processing practices have been in practice to reduce the level of contamination. Long-
term strategies like breeding for resistance to toxigenic fungi are also very promising (Bryden, 2007;
Wagacha and Muthomi, 2008).
22
The knowledge that mycotoxins have serious effects on humans, animals and countries’ economies
has not only led to strategies for reducing mycotoxins contamination, but also to the establishment of
regulations on mycotoxins levels in food and feed. Worldwide, approximately 100 countries had
developed specific limits for mycotoxins in food and feed by the end of 2003, which represent
approximately 87% of world inhabitants (FAOSTAT, 2007). In Africa, the majority of the countries
have no specific mycotoxins regulations, although the problem is undeniable. However, mycotoxins
issues in Africa can only be effectively addressed when regarded in the overall context of local food
safety, health, and agricultural issues. The establishment of mycotoxins regulations will have limited
effects in terms of health protection for subsistent farmers reliant on their crops. In addition, adequate
resources to afford improved varieties, fertilizers and insecticides and information on good agricultural
practices and resistance of plant cultivars to fungal infection for small farmers need to be addressed
through appropriate strategies and policies (Golob, 2007; Shepard, 2004).
2.5.1 Fusarium in Africa
Mycotoxins from Fusarium species in the past have usually been associated with temperate cereals.
This is because these fungi require slightly lower temperatures for growth and mycotoxin production
than the aflatoxigenic Aspergillus species. Though, extensive data now exists to indicate that
contamination of cereal grains with a number of Fusarium mycotoxins is on a global scale (Muller and
Schwadorf, 1993; Chulze et al., 1996; Viquez et al., 1996). Despite the prevalence of fumonisins in
maize and the importance of maize as a food staple, there is inadequate information available on the
natural occurrence of fumonisins in maize consumed by rural populations in sub-Saharan Africa, with
the exception of South Africa. Surveys of maize from rural smallholder farms in the Transkei region
of South Africa were conducted in 1985 and 1989. High incidences and levels of fumonisin B1 were
found in both good-quality and mouldy maize (Rheeder et al., 1992). A study in Benin indicated a
high prevalence of F. verticillioides strains, which are high fumonisin producers (Fandohan et al.,
2005). Doko et al. (1995) , in their study comparing fumonisin contamination in different African
countries, already noted Benin as a high occurrence area since they found high total fumonisin levels
(3 mg/kg) in maize samples. The highest FB1 levels produced by isolates of F. verticillioides reported
so far are 17,900 mg/kg from South Africa (Alberts et al., 1990).
23
Adejumo et al., (2007) reported that F. verticillioides were the most commonly fungi in Nigerian
maize, other Fusarium species isolated included F. sporotrichioides, F. graminearum , F.
pallidoroseum , F. compactum , F. equiseti, F. acuminatum , F. subglutinans and F. oxysporum .
Gamanya et al., (2001) conducted a survey to determine the levels of fumonisins in maize in
Zimbabwe.The study carried out in Zimbabwe’s different ecological zones (wet Region I, moderately
wet Region II and dry Region III) observed that the incidence of fungal contamination can be clearly
linked to high rainfall and high relative humidities. A general comparison between cereals and peanuts
shows that the distribution of F. moniliform in maize was significantly higher than in other crops.
The study showed that the incidence of F. moniliforme and other Fusarium species and levels of FB1
decreased from regions with high rainfall and annual moderate temperatures to low rainfall regions.
This corresponds with studies previously carried out in tropical regions such as Transkei, South Africa,
where F. moniliforme and FB incidences were correlated and the relationship of its distribution to
climatic conditions established (Sydenham et al., 1990). This implies that for effective control of
Fusarium infection in crops and mycotoxin production may indeed require focusing on particular
agricultural regions.
Wagacha et al., (2010) reported the occurrence of 19 different Fusarium species in wheat in Kenya
with F. boothii, F. poae, F scirpi, F. chlamydosporum, F. graminearum, and F. anthrosporioides
accounting for 80% of contamination. Major Fusarium-related mycotoxins such as deoxynivalenol
(DON), nivalenol (NIV), zearalenone (ZEA), T2-toxin and HT2-toxin have been reported in wheat
kernels sampled from fields in different wheat-growing regions of Kenya (Muthomi et al. 2002, 2007a,
2008). O’Donnell et al., (2008) identified a new species in Ethiopia (F. aethiopicum) which produce
15ADON.
Multi-locus sequence typing (MLST) suggests that this species with the closely related Fusarium
acaciae-mearnsii may be endemic to Africa and Australia. Boutigny et al., (2011) conducted a more
recent extensive study in South Africa where only F. boothii was found in maize (15ADON). In wheat
and barley, 85% of the isolates were F. graminearum 15ADON type. However, it is important to note
that a majority of the research studies mainly based on random surveys of farmers’ stores and retail
markets, mostly basing data measurements on a relatively small number of samples.
24
The number of investigations on FFSC and FGSC diversity in Africa is still limited. Fusarium and
their mycotoxins in maize have been conducted in numerous parts of the world, namely in the USA,
South America, Europe and South Africa. There is still limited research on the occurrence of Fusarium
and its toxins in maize in Africa with the exception of South Africa (Gamanya and Sibanda, 2001;
Ngoko et al., 2001; Kpodo et al., 2000; Doko et al., 1995; Kedera et al., 1999). There is a great need
for additional investigations on the continent, at least where maize production and consumption are
predominant.
2.6 Food Safety and Health Hazard Implications Associated with Mycotoxins
The occurrence of mycotoxins in food and feed includes potential risks for the health of both humans
and animals. Mycotoxins are potential carcinogenic, mutagenic and teratogenic compounds (Wagacha
and Muthomi, 2008). Chronic exposure to mycotoxins has also been associated with impaired growth
in children (Gong et al., 2002; Gong et al., 2004; Kimanya et al., 2010), neural tube defects in unborn
children (Marasas et al., 2004) and immunosuppression (Turner et al., 2009), which results in
vulnerability to other infectious diseases.
Aflatoxins have been implicated in acute aflatoxicosis, carcinogenicity, growth retardation, neonatal
jaundice and immunological suppression in SSA. Acute aflatoxicosis usually associated with
extremely high doses of aflatoxin is characterized by hemorrhage, acute liver damage, edema, and
death in humans. Several reported cases of acute aflatoxicosis in Africa and Asia are mainly associated
with consumption of contaminated home-grown maize as indicated in Table 2-4. According to Miller
(2008), 40% of the productivity lost to diseases in developing countries is due to diseases aggravated
by aflatoxins. Unfortunately, many of the people in the region are not even aware of the effect of
consuming moldy products. Due to the low literacy levels and other socio-economic factors, even if
steps were taken to make food products safe, the consumers might be unwilling to pay extra costs, and
may still prefer to buy the cheap commodities.
25
Table 2-4 Aflatoxicosis in Maize consuming countries
Population Fatalities Samples Estimated Intakea References
397 Patients in
Western India, >180
villages (1974)
106 dead
27% Fatality
Maize from affected
households contained
aflatoxin(type
unspecified) levels
between 6250-15,600
ppb
6.25-15.6 ppm
aflatoxins and 350g
maize/day equates to
2.19-5.46 mg
aflatoxins/kg/day
Krishnamachari, K..
A. et al. (1975)
20 cases in Machakos
district, Kenya (1981)
12 dead
60% Fatality;
Maize from homes
with fatalities had
AFB1levels 3200 –
12,000 ppb.
3.2-12 ppm AFB1 and
350g maize/day
equates to 1.12-4.2 mg
AFB1/kg/day
Ngindu, A. et al.
(1982)
317 case in Eastern
Kenya (Makueni,
Kitui, Machakos, and
Thika) and a case-
control study of 40
cases with acute
jaundice and 80
village controls
125 dead
39% Fatality
Case-control
Study- 29 cases
alive at time of
blood sampling,
an additional 7
dead by August
2004
GM of Total AF in
stored maize; 354.53
ppb in case and 44.14
ppb in control
households
Intakes 5-20 ppm were
associated with
fatality and 350g
maize/day equates to
1.75-7mg
Aflatoxins/kg/day
PROMEC Unit.
(2001)
GM, Geometrical Mean aAssumed body weight of 60kg
b Blood samples collected after an average of 33 days of onset of symptoms c Household maize collected after an average of 33 days of onset of symptoms (9-112 days)
Epidemiological studies of human populations exposed to diets naturally contaminated with aflatoxins
revealed an association between the high incidence of liver cancer in Africa and dietary intake of
aflatoxins. Hepatitis B and C infections coupled with aflatoxin exposure; which are common in sub-
Saharan Africa has shown to raises the risk of liver cancer by more than ten-fold as compared to either
chronic exposure alone (Turner et al., 2005). In addition, preliminary evidence suggests that there may
be an interaction between chronic mycotoxin exposure and malnutrition, immunosuppression,
impaired growth, and diseases such as malaria and HIV/AIDS (Gong et al., 2003, 2004). A recent
study in Ghana indicated that higher levels of aflatoxin B1-albumin adducts in plasma were associated
with lower percentages of certain leukocyte immunophenotypes (Jiang et al., 2005) while another
research study in Gambian children found an association between serum aflatoxin albumin levels and
reduced salivary secretory IgA levels (Turner et al., 2005).
26
Studies on mycotoxins-linked diseases in animals show equine leuko- encephalomalacia in horses and
pulmonary oedema in pigs (Kellerman et al., 1990), liver and kidney cancer in mice and rats and
immunosuppression (Oswald et al., 2005). In 1993, the International Agency for Research on Cancer
(IARC) had also categorized mycotoxins into different groups with respect to their carcinogenicity as
shown in Table 2-5 for Fusarium toxins (IARC, 2002).
Table 2-5 Carcinogenicity Risk evaluated by IARC for Fusarium Mycotoxins
Toxins Degree of evidence of
carcinogenicity
Overall evaluation
In human In animals
Toxins derived from
F. graminearum,
F. culmorum,
F. crookwellense
I
Group 3
Zearalenone ND L
Nivalenol I
Fusarenone X I
Deoxynivalenol I
Toxins derived from
F. sporotrichioides
ND
Group 3
T-2 toxin L
Toxins derived from
F. moniliforme
I
S
Group 2B
Fumonisin B1 L
Fumonisin B2 I
Fusarin C L
*Source IARC, (1993, 2002)
I, insufficient evidence; L, limited evidence; ND, no adequate data; S, sufficient evidence.
Group 2B=possibly carcinogenic to humans; Group 3= not classifiable as to its carcinogenicity to humans.
27
2.7 Mitigation Strategies for Mycotoxins
Stored and processed food commodities such as maize, sorghum, millet, and barley carry a broad
range of microorganisms. The different species of microorganisms will depend on field climatic
conditions and harvesting procedures. Extrinsic and intrinsic factors influence mycotoxins production.
Extrinsic factors include factors such as temperature, water availability and gas compositions (Magan
et al., 2003). Since mycotoxins are known to have detrimental effects on post-harvest food losses as
well as to the health of consumers, a number of mitigation strategies have been developed to prevent
growth of fungi as well as to decontaminate and detoxify food, which contaminated by mycotoxin
(Kabak et al., 2006).
To mitigate and reduce impact of mycotoxins in food and feed chain needs comprehensive research in
order to understand crop biology, agronomy, fungal ecology, harvesting methods, storage conditions
and detoxification methods of mycotoxin (Bryden, 2009), importantly the hazard analysis critical
control point systems (HACCP) is the key element in reduction of mycotoxins (Aldred et al., 2004).
Grain with high moisture should not be held in wagon or trucks for more than six hours instead should
be dried to moisture content level of 12-13% to stop production of aflatoxin (Sumner & Lee, 2009).
Mould, fungi, and aflatoxins are usually at higher levels in the fine material, thus by removing fine
material will reduce aflatoxin levels by 50%.
Different preparation methods of cereal before milling show significance reduction of contamination
of mycotoxins in flour, such methods are like hand sorting of maize and washing play vital part in
reducing mycotoxins during preparation of complementary food (Van der Westhuizen et al., 2011).
Sorting has proven that it can remove a large percentage of aflatoxin contaminated grain, but reduction
may also be through food processing procedures such as washing, wet and dry milling, grain clearing,
dehulling, roasting, baking, frying and extrusion cooking. Other possible methods used to mitigate
mycotoxin production in cereals includes avoiding prolonged harvesting, and long drying period on
the field have been linked with higher aflatoxin levels in corn in Benin (Hell et al., 2003).
Other potential methods includes application of antifungal such as synthetic antioxidants (Farnochi et
al., 2005), educating population on risk of mycotoxin contaminated diet, Sanitation, smoking (Bankole
& Adebanjo, 2003), use of essential oils (Nguefack et al., 2004), natural phenolic compounds (Bakan
et al.,2003) or the use of modified atmospheres (Ellis et al., 1993; Ellis et al.,1994). Changing degrees
of efficiency have in mitigation of mycotoxins have been achieved, which have not necessarily
resulted in commercial success. Besides, a majority of these studies have been carried out on artificial
media, and their effects would still need to be validated on corn (Samapundo et al., 2006).
28
2.7.1 Reducing Mycotoxin Exposure
Fundamental to developing prevention strategies, it is crucial to have an understanding of the
interaction between the fungus and the host plant. For instance, Aspergillus spp. will infect the maize
crop in the field but aflatoxins continue to accumulate post-harvest under poor storage conditions,
which favor fungal growth and toxin production. Therefore, post-harvest interventions may contribute
significantly to controlling aflatoxin. The majority of the toxin is present at the time of harvest. Thus,
control of FB requires more attention to pre-harvest practices and the subsequent effects of processing
and preparation of foodstuffs (Humpf. et al., 2004).
Recommendations to shift the traditional diet away from commodities prone to contamination can
reduce chronic exposure. For instance, in China economic developments resulted in reduced maize
consumption (IIASA, 2009) formerly the primary source of aflatoxin exposure in regions such as
Qidong County. Populations in some of the poorest countries facing the highest risk of mycotoxin
exposure due to consumption of contaminated staple foods are trapped by poverty and the lack of
alternatives, making it virtually impossible to replace the contaminated food with that of good quality
(Shephard, 2008). The lack of established regulatory mechanisms in these countries makes the
situation worse, the reliance on staple foods highly contaminated with mycotoxins exposes them to
high doses beyond established toxicological safe limits (Table 2-6).
Table 2-6 Toxicological Safe Limits for Mycotoxins
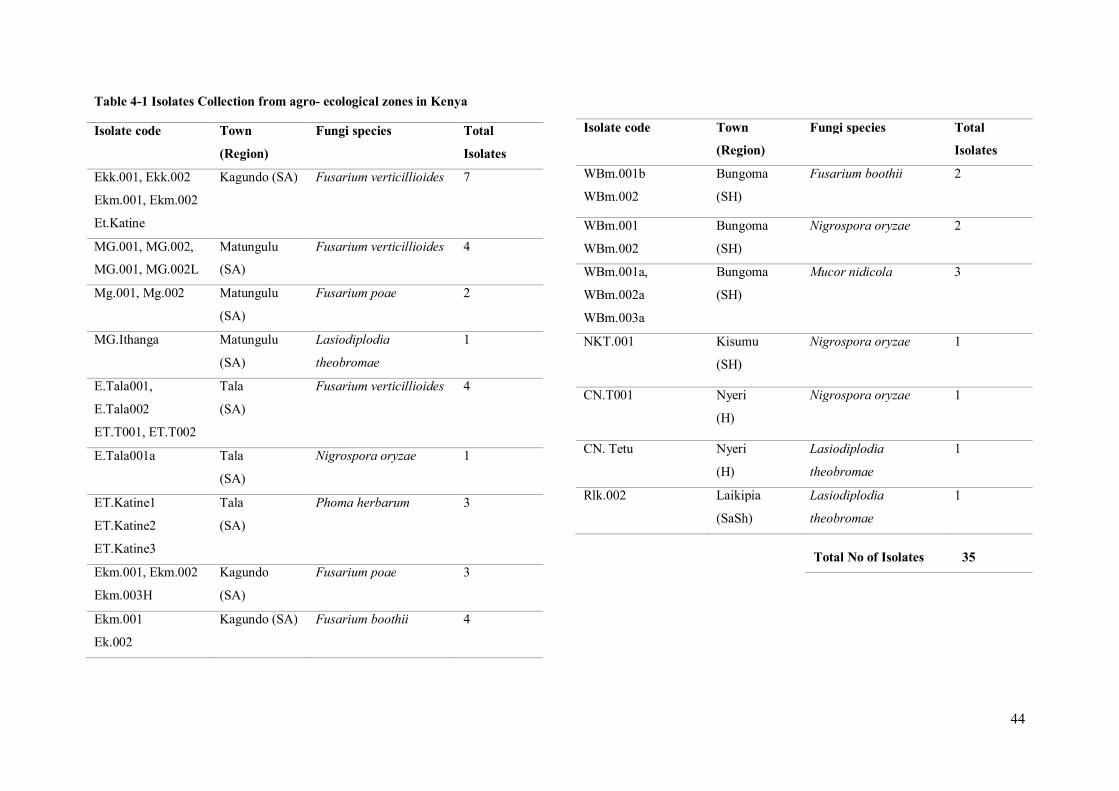
Table 4-1 Isolates Collection from agro- ecological zones in Kenya
Isolate code Town
(Region)
Fungi species Total
Isolates
Ekk.001, Ekk.002
Ekm.001, Ekm.002
Et.Katine
Kagundo (SA) Fusarium verticillioides 7
MG.001, MG.002,
MG.001, MG.002L
Matungulu
(SA)
Fusarium verticillioides 4
Mg.001, Mg.002 Matungulu
(SA)
Fusarium poae 2
MG.Ithanga Matungulu
(SA)
Lasiodiplodia
theobromae
1
E.Tala001,
E.Tala002
ET.T001, ET.T002
Tala
(SA)
Fusarium verticillioides 4
E.Tala001a Tala
(SA)
Nigrospora oryzae 1
ET.Katine1
ET.Katine2
ET.Katine3
Tala
(SA)
Phoma herbarum 3
Ekm.001, Ekm.002
Ekm.003H
Kagundo
(SA)
Fusarium poae 3
Ekm.001
Ek.002
Kagundo (SA) Fusarium boothii 4
Isolate code Town
(Region)
Fungi species Total
Isolates
WBm.001b
WBm.002
Bungoma
(SH)
Fusarium boothii 2
WBm.001
WBm.002
Bungoma
(SH)
Nigrospora oryzae 2
WBm.001a,
WBm.002a
WBm.003a
Bungoma
(SH)
Mucor nidicola 3
NKT.001 Kisumu
(SH)
Nigrospora oryzae 1
CN.T001 Nyeri
(H)
Nigrospora oryzae 1
CN. Tetu Nyeri
(H)
Lasiodiplodia
theobromae
1
Rlk.002 Laikipia
(SaSh)
Lasiodiplodia
theobromae
1
Total No of Isolates 35
45
4.3 Molecular Analysis
4.3.1 PCR Amplification
To confirm the morphometric identifications of the isolates collected (Table 4-3), PCR amplification
was done. The resultant query length of amplified products of ITS and EF ranged from approximately
520 – 678 (Table 4-1).Sequences of these amplified products were compared with those deposited in
the NCBI GenBank and Fusarium-ID database. Sequences from isolates of these species showed an
identity ranging from 99% to 100%; Table 4-1 shows the tabulated results. The distribution of
Fusarium species was observed to be mostly in the Semi-arid and sub-humid regions.
The results also identified Fusarium verticillioides as the major contaminant of the infected maize
kernels. Fusarium verticillioides was distributed across the various towns in the semiarid regions,
indicating that it may be an endemic contaminant in maize grown in these regions. Fusarium poae and
Fusarium boothii were also isolated from the maize kernels. The F. boothii was isolated from samples
collected both in the semi-arid and sub-humid region in Bungoma while the F. poae was isolated from
maize kernels collected in the semi-arid regions.
46
Table 4-2 Pathogenic Fungi Species Identified
AEZ Region Town Code Pathogen Identity (ITS) Identity
(TEF 1- alpha)
Humid (H) Central Nyeri CN.Tetu
CN.Tetu1
Nigrospora oryzae
Lasiodiplodia theobromae
99%
100%
-
-
Sub Humid (SH) Western Bungoma WB.Makhonge
WB.Makhonge1
Mucor nidicola
Fusarium boothii
99%
99%
-
100%
Kisumu Nigrospora oryzae 99% -
Semi-Arid (SA) Eastern Tala ET.Tala1
ET.Tala
Fusarium verticillioides
Giberrella moniliformis
99%
99%
99%
99%
Kagundo EK.Mbilini
EK.Mbilini1
EK.Kitwii
Fusarium poae
Fusarium boothii
Fusarium verticillioides
100%
99%
99%
100%
99%
100%
Matungulu ET. Matungulu Phoma herbarum
Fusarium poae
100%
100%
-
100%
Ithanga MG.Ithanga
MG.Ithanga1
Lasiodiplodia theobromae
Fusarium verticillioides
100%
100%
-
99%
SemiArid-SemiHumid (SaSh) Central Laikipia RL.Kieni Nigrospora oryzae 100% -
47
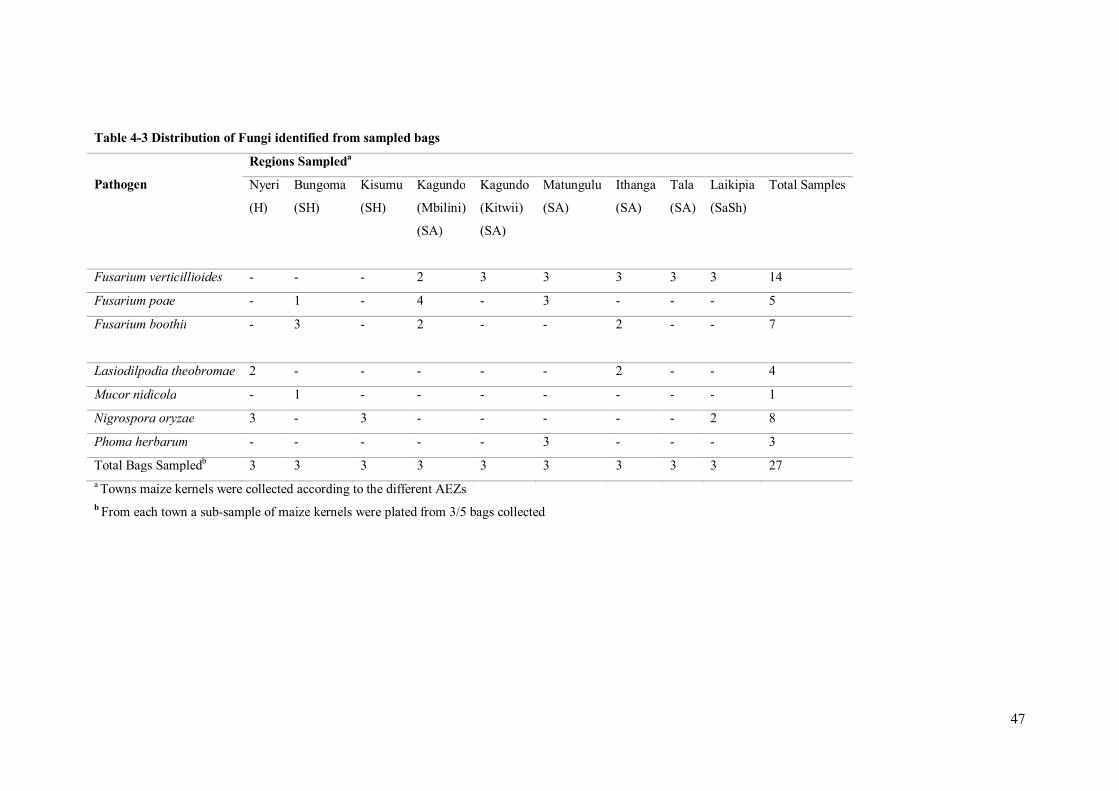
Table 4-3 Distribution of Fungi identified from sampled bags
Pathogen
Regions Sampleda
Nyeri
(H)
Bungoma
(SH)
Kisumu
(SH)
Kagundo
(Mbilini)
(SA)
Kagundo
(Kitwii)
(SA)
Matungulu
(SA)
Ithanga
(SA)
Tala
(SA)
Laikipia
(SaSh)
Total Samples
Fusarium verticillioides - - - 2 3 3 3 3 3 14
Fusarium poae - 1 - 4 - 3 - - - 5
Fusarium boothii - 3 - 2 - - 2 - - 7
Lasiodilpodia theobromae 2 - - - - - 2 - - 4
Mucor nidicola - 1 - - - - - - - 1
Nigrospora oryzae 3 - 3 - - - - - 2 8
Phoma herbarum - - - - - 3 - - - 3
Total Bags Sampledb 3 3 3 3 3 3 3 3 3 27 a Towns maize kernels were collected according to the different AEZs b From each town a sub-sample of maize kernels were plated from 3/5 bags collected
48
4.3.2 Screening for the ability to produce Mycotoxins
The primer pairs tri13F/R used were designed by comparing the published sequences for this gene
from known NIV and DON-producers (Accession# AF336365, AF366366, and AY057841–
AY057844). Tri13 and Tri17 genes are responsible for oxygenation and acetylation of the C-4 residue
of the trichothecene backbone respectively. They are used to identify the putative chemotype of each
of the isolates. DNA from isolates were subjected to PCRs with the primer pairs, and each of these
DNAs generated amplicons with fragments of sizes 415bp and 234bp (Waalwijk et al., 2003).
The isolates Fusarium poae and Fusarium boothii generated a 415bp fragment (NIV-producers) and a
fragment of 234bp (DON-producers) respectively.
Figure 4-3 Chemotypes for Fusarium boothii (rep1) Ekm.001, (rep2) WBm.001b and Fusarium poae (rep1)
Mg.001, (rep2) Ekm.003H
The Fusarium isolates were identified as B-Type trichothecenes producers’; Fusarium boothii
classified as 15ADON chemotype; the Fusarium poae isolates were both from the semi-arid regions,
and only one isolate was successfully identified as a NIV chemotype. This suggests that maize in the
semi-arid and humid regions is most likely to be contaminated with B-Type trichothecenes particularly
NIV + acetylated derivatives and DON+15ADON. Fusarium boothii and Fusarium poae are members
of the FGSC species which are known to produce B-Type trichothecenes such as deoxynivalenol
(DON, known as vomitoxin), nivalenol (NIV) and their acetylated derivatives (Ward et al., 2002).
49
5 CHAPTER FIVE: DISCUSSION
The maize samples from the semi-arid and sub-humid zones were highly contaminated with members
of Fusarium graminearum species complex (FGSC) and the Fusarium fujikuroi species complex
(FFSC). The study successfully identified Fusarium poae, Fusarium verticillioides, Fusarium boothii
as the major mycotoxin producing fungi from the collected maize samples. Fusarium verticillioides
(FFSC strain) was predominant (33%), followed by FGSC strains: Fusarium boothii (17%) and
Fusarium poae (12%). The results correspond with findings of Muthomi and Mutitu (2003); Fusarium
spp. was isolated at high frequencies in cereal grain (wheat). In addition, a field survey conducted for
Fusarium spp. in the major maize-growing areas of Kenya in 1993 found F. moniliforme to be
predominant (82% of isolates from maize), followed by F. graminearum (9% of isolates) and F.
subglutinans which was 7% of isolates (Kedera, 1994).
Wagacha et al., (2010) reported the occurrence of 19 different Fusarium species in wheat in Kenya
with F. boothii, F. poae, F scirpi, F. chlamydosporum, F. graminearum, and F. anthrosporioides
accounting for 80% of contamination. MacDonald and Chapman (1997) also reported a high incidence
of F. graminearum (9% of the kernels tested) and of F. moniliforme (14% of the kernels tested), in a
survey of maize grain purchased from market stalls and roadside traders in central and western Kenya.
Studies hence indicate that Fusarium spp. is a predominant pathogen in Kenyan maize.In addition,
they suggest that the FGSC and FFSC strains are endemic in Kenya. Futhermore, O’Donell et al.,
(2008) identified a new species in Ethiopia (F. aethiopicum) which produce 15ADON.
Pathogenic and mycotoxin-producing Fusarium species isolated from the maize kernels: F.
verticillioides, F.boothii, and F. poae suggests that infection in the sampled regions is due to a
complex of Fusarium species. According to Marasas (1991), F. graminearum, F. poae, and F.
verticillioides are considered the most toxic Fusarium species. F. graminearum complex which
includes Fusarium boothii is the most significant producer of DON and ZEA. The multiple
contaminations of maize with different mycotoxigenic fungi indicate a potential risk of contamination
of the grain with various mycotoxins like trichothecenes, T-2 toxin, zearalenone, fumonisins,
moniliformin.
50
The major mycotoxins that contaminate small-grain cereals are the typeA trichothecenes T-2 and HT-
2, primarily produced by F. poae and the typeB trichothecenes DON (or vomitoxin) and NIV
produced mainly by F. graminearum. Comparative analysis of the gene clusters associated with the
biosynthesis, in DON-producers and NIV-producer, identify the genes tri13 and tri7 as vital in the
synthesis of either NIV or DON. Gene sequences from both genes were used to develop primers that
were utilized to screen Fusarium boothii and Fusarium poae chemotypes, characterizing them as
DON- and NIV-producers respectively. (Waalwijk et al., 2003)
Harvested maize grains in the tropical zones are infected and invaded by mycelium and spores of
diverse group of pathogenic fungal species including mainly Fusarium, Aspergillus and Penicillium
that can come into contact, grow and compete for food when the environmental conditions are
favorable. Many research studies have highlighted the interaction of Fusarium species with other
fungi. Findings from Velluti et al. (2000) showed that populations of F. verticillioides and F.
proliferatum, the most important fumonisin producers, are markedly reduced by the presence of F.
graminearum , and that fumonisin B1(FB1) production by them can be significantly inhibited as well
in the presence of F. graminearum.
On the other hand, Marin et al. (1998) found that F. verticillioides and F. proliferatum are generally
very competitive and dominant against Aspergillus flavus and Penicillium spp. especially at a water
activity of more than 0.96. This inhibition can lead to significantly reduced aflatoxin contamination in
infected grains (Zummo and Scott, 1992). This may be indicated by the higher incidence of Fusarium
verticillioides in the maize samples and lack of Aspergillus flavus and Penicillium spp in the isolates.
Though this too could be explained by the size of the samples collected.
Fusarium verticillioides is an endophyte of maize that has long-term associations with the host plant.
Therefore, the symptomless infection can exist throughout the plant in leaves, stems, roots, grains, and
its presence as in many cases ignored since it does not cause visible damage to the plant. Hence, this
indicates that some strains of F. verticillioides may produce disease in maize while others do not. The
strain infects corn at all stages of development, either via infected seeds, the silk channel or wounds,
causing grain rot during both the pre- and postharvest periods. F. verticillioides strains are high
fumonisin, producers. This appeals for more attention and suggests that farmers should adopt adequate
postharvest management procedures to assure satisfactory quality of the stored maize.
51
The Fusarium trichothecenes have been divided into type A trichothecenes, characterized by a
functional group other than a ketone at C-8, and type B- trichothecenes with only the carbonyl at C-8.
The type A trichothecenes include T2 and HT2, mainly produced by strains of F. sporotrichioides, F.
acuminatum, and F. poae (Logrieco et al., 2002). Fusarium graminearum species complex comprised
of at least nine distinct, cryptic species including Fusarium acaciae-mearnsii, F. asiaticum, F.
austroameri-canum, F. boothii, F. meridonale, F. mesoamericanum. Members of this complex are
known to produce mycotoxins including the trichothecenes deoxynivalenol (DON) along with its
acetylated derivatives and nivalenol (Goswami et al., 2005).
The co-occurrence of the different Fusarium mycotoxins may result in additive and synergistic effects.
Multiple mycotoxins may lead to synergistic toxicity which is greater than the total of the toxicities of
each mycotoxin (Speijers, 2004). The number and type of mycotoxins in a sample depends on
Fusarium strains present and their toxigenicity as well as environmental factors. The legislative limits
for Fusarium mycotoxins range from 500μg/kg-2000μg/kg for deoxynivalenol, 30-1000μg/kg for
zearalenone and 100μg/kg for T-2 toxin. This implies that the Kenyan maize products could be
contaminated with low but significant levels of the Fusarium mycotoxins.
.
52
6 CHAPTER SIX: CONCLUSIONS AND RECOMMENDATIONS
This research study suggests that Fusarium graminearum species complex (FGSC) and the Fusarium
fujikuroi species complex (FFSC) are the principal contaminants of maize grain in Kenya. More than
50% of the samples tested were infected with Fusarium species which usually produce a broad range
of toxins specifically trichothecenes (Type A and Type B), zearalenone and fumonisins. This
underscores the need for more vigilance and implementation of preventive measures that reduce the
risk of toxin accumulation in the field and contaminated maize. There is still insufficient information
available on the occurrence of Fusarium spp. and its toxins in Kenya; focus has been mainly on
aflatoxin control.
Few African countries with the exception of South Africa, have conducted research studies on FGSC
and FFSC diversity in cereal grains. Most of these studies mainly are based on random surveys of
farmers’ stores and informal markets with data measurements based on small numbers of samples
(Shephard et al., 1996). This too is a limitation in this research study. However to be saluted are efforts
investigating fumonisins contamination in maize and maize-based foods in some African countries:
Benin, Cameroon, Ghana, Kenya, Zambia and Zimbabwe, Tanzania (Shephard et al., 1996; Kimanya
et al.2010; Doko et al., 1995; Hell et al., 1995; Kedera et al., 1999; Ngoko et al., 2001). Though, there
is still a great need for more investigations on the continent, mainly in the maize production and
consumption zones.
Planting improved maize cultivars, combined with good crop management and post-harvest handling
practices should be explored to deter the proliferation of fungal species and reduce the risk of
mycotoxins contamination. Novel control strategies should be further investigated and applied in the
field rather than in artificial media. The use of an endophytic bacterium (Bacillus mojavensis) as a
biological control agent on maize seed (Bacon and Hinton, 2000) and the use of non-mycotoxin
producing strains of F. verticillioides aiming to minimise fumonisin levels in maize (Plattner et al.,
2000) has been reported. Additional investigations are however needed to render some of those
technologies more applicable and feasible for use by farmers in the field.
53
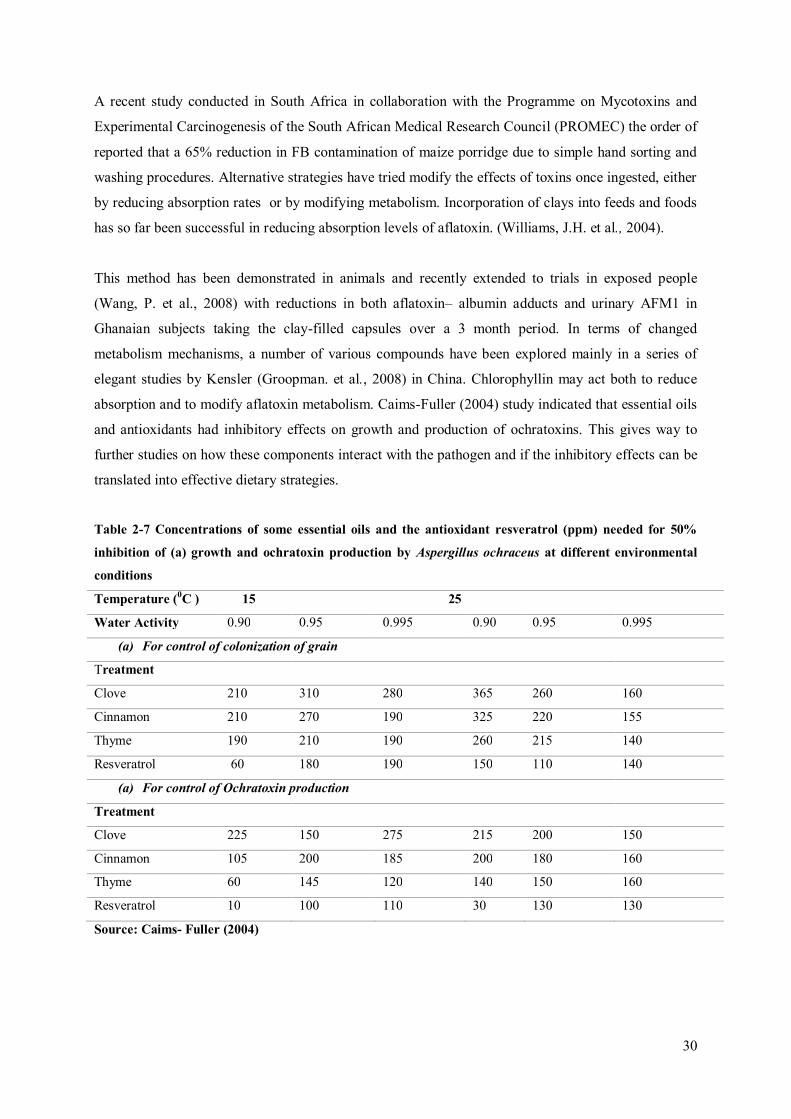
Recently essential oils (Table 2-7) have been proposed for application post-harvest to prevent both
fungal growth and mycotoxin production after positive evaluation of their effects on artificial media.
These effects, however, may not be translated to the same extent on the actual food product owing to
interactions that may occur in the more complex food matrix compared to those in artificial media. But
could they be an option as a dietary strategy?
PCR-based detection and identification provides qualitative information about the presence or absence
of a certain fungus and provides a cheaper option. This can be used for food and feed quality control
as the technology has the power to provide insight into the mycotoxigenic potential of samples
analysed. This information can then be used to decide whether a lot should go further down the
process of production or should be retained for further analysis of mycotoxins. The emerging topic of
conjugated mycotoxins is also of great interest especially with the current technologies in plant
breeding resistant varieties. Host plant for fungi is constantly changing as plants are being bred for
fungi/toxin resistance and different nutrient profile; hence continually new masked toxins are being
discovered. It is essential to investigate infection pathways of pathogenic fungi, as this is the basis of
good management and postharvest handling practices.
Biocontrol, resistance breeding still remain as important strategies to minimize chronic exposure to
mycotoxins in the developing world. The aflasafe biocontrol method can reduce aflatoxin
contamination in corn and groundnuts by 80–90%, in some cases even as much as 99% (IITA, 2011).
Further investigation on the biocontrol potential of atoxigenic Fusaria strains is recommended.
A significant challenge is the lack of consistent standards in African countries and elsewhere in the
world, it is important that governments support the development of harmonized standards in the region.
Dietary diversification by the Kenyan population, reducing reliance on the maize crop has been
suggested. This is not a realistic mitigation measure in the communities whose maize cultivation and
consumption is heavily embedded in their cultural norms.
54
LIST OF REFERENCES
Abass, A. B., Ndunguru, G., Mamiro, P., Alenkhe, B., Mlingi, N., & Bekunda, M. (2014). Post-harvest
food losses in a maize-based farming system of semi-arid savannah area of Tanzania. Journal of
Stored Products Research, 57, 49-57
Adejumo, T. O., Hettwer, U., & Karlovsky, P. (2007). Occurrence of Fusarium species and
trichothecenes in Nigerian maize. International journal of food microbiology, 116(3), 350-357.
Aldred, D., Magan, N., & Olsen, M. (2004). The use of HACCP in the control of mycotoxins: the case
of cereals. Mycotoxins in food: detection and control. Boca Raton, Fla.: CRC Press. p, 139-73.
Aoki, T., Ward, T. J., Kistler, H. C., & O’Donnell, K. (2012). Systematics, phylogeny and
trichothecene mycotoxin potential of Fusarium head blight cereal pathogens. Mycotoxins, 62(2), 91-
102.
Atehnkeng, J., Ojiambo, P. S., Ikotun, T., Sikora, R. A., Cotty, P. J., & Bandyopadhyay, R. (2008).
Evaluation of atoxigenic isolates of Aspergillus flavus as potential biocontrol agents for aflatoxin in
maize. Food Additives and Contaminants, 25(10), 1264-1271.
Bakan, B., Bily, A. C., Melcion, D., Cahagnier, B., Regnault-Roger, C., Philogène, B. J., & Richard-
Molard, D. (2003). Possible role of plant phenolics in the production of trichothecenes by Fusarium
graminearum strains on different fractions of maize kernels. Journal of agricultural and food
chemistry, 51(9), 2826-2831.
Bankole, S., Schollenberger, M., & Drochner, W. (2006). Mycotoxins in food systems in Sub-Saharan
Africa: A review. Mycotoxin Research, 22(3), 163-169.
Bankole, S. A., & Adebanjo, A. (2004). Mycotoxins in food in West Africa: current situation and
possibilities of controlling it. African Journal of Biotechnology, 2(9), 254-263.
Barnett H. L., Barry B., Hunter F. (1998). Illustrated Genera of Imperfect Fungi (4th Ed.). American
Pathology Society. USA
Bhat R.V, Shetty P.H, Rao P.A, Rao VS. (2010). A foodborne disease outbreak due to the
consumption of moldy sorghum and maize containing fumonisin mycotoxins. Journal of Clinical
Toxicology. 1997; 35: 249–55.
55
Bryden, L.W. (2009). Mycotoxins and mycotoxicosis, significance, occurrence and mitigation in the
food chain. General, Applied and Systems Toxicology, 3:6
Cairns-Fuller, V. (2004). Dynamics and control of ochratoxigenic strains of Penicillium verrucosum
and Aspergillus ochraceus in the stored grain ecosystem (Doctoral Dissertation, Cranfield University).
CDC (Centers for Disease Control and Prevention) 2004. Outbreak of aflatoxin poisoning—eastern
and central provinces, Kenya, January–July 2004.
CIMMYT, International Maize and Wheat Improvement Center, (2009). Effective Grain Storage for
Better Livelihoods of African Farmers Project. http://www.cimmyt.org/fr/programs-and-units/global-
maize-program/projects/effective grain-storage-project (accessed 12th June 2015)
Creppy, E.E. (2002). Update of the survey, regulation and toxic effects of mycotoxins in Europe.
Toxicology Letters, 127, 1–3, 19–28
Daniel, J. H., Lewis, L. W., Redwood, Y. A., Kieszak, S., Breiman, R. F., Flanders, W. D. &
McGeehin, M. A. (2011). Comprehensive Assessment of Maize Aflatoxin Levels in Eastern Kenya.
Environmental health perspectives, 119(12), 1795.
De Boevre, M., Jacxsens, L., Lachat, C., Eeckhout, M., Di Mavungu, J. D., Audenaert, K. & De
Saeger, S. (2013). Human exposure to mycotoxins and their masked forms through cereal-based foods
in Belgium. Toxicology letters, 218(3), 281-292.
Delgado, J.E., & Wolt, J.D. (2010). Fumonisin B1 and implications in nursery swine productivity: A
quantitative exposure assessment. Journal of Animal Science, 88(11) 3767-3777.