THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1993 by The American Society for Biochemistry and Molecular Biology, Inc. Vol. 268, No. 14, Issue of May 15, pp. 10252-10259,1993 Printed in U. S. A. The C-terminal Domain of the Plasma Membrane Ca2’ Pump Contains Three High Affinity Ca2’ Binding Sites* (Received for publication, August 14, 1992, and in revised form, December 24, 1992) Francesco Hofmann,Peter James, Thomas VorherrS, and Ernest0 CarafoliB From the Institute of Biochemistry, Swiss Federal Institute of Technology (ETH), 8092 Zurich, Switzerland The C-terminal portion of the plasma membrane Ca2+- ATPase contains different regulatory domains. A re- combinant C-terminal fragment of the human plasma membrane CaZ’-ATPase l b isoform (E1079-P1180) was used to study the role of two acidic amino acid stretches located on either side of the calmodulin bind- ing domain, corresponding to synthetic peptides A18 (Vorherr, T., James, P., Krebs, J., Enyedi, A., Mc- Cormick, D. J., Penniston, J. T., and Carafoli, E. (1990) Biochemistry 29, 355-365) and B28 (James, P., Pruschy, M., Vorherr, T., Penniston, J. T., and Carafoli, E. (1989) Biochemistry 28, 4253-4258), re- spectively. The molecular mass of the recombinant C-terminal fragment, as determined by electrosprayionization mass spectrometry, was higher by 39 mass units than the calculated value (12,055 Da). This difference was the result of an EGTA-insensitive Ca2+ ion, which was located bychymotryptic proteolysis in a fragment cor- responding to the last 37 amino acidsof the expressed protein. Fluorescence experiments on the dansylated recombinantC-terminalfragmenttitratedwithin- creasing amountsof free Ca2+ revealed two additional Ca2+ binding sites with affinities corresponding to KD values of about 30 and 300 nM, respectively. Stains All spectra of different synthetic peptides, corresponding to subdomainsof the expressed protein, indicated that the site with the KD of 30 nM was probably located in the acidic sequence on the N-terminal side of the cal- modulin binding domain (peptide A18); the site with the 300 nM KD was apparently located on the C-ter- minal side of the calmodulin binding domain (peptide B28) or, alternatively, formed by the cooperation of distant residues of the domain. The C-terminal portion of the plasma membrane Ca2+ pump (for review, see Carafoli (1992)) contains various regulatory domains. This distinguishes the pump from all other pumps of the P-class (Pedersen and Carafoli, 1987a, 1987b), e.g. the sarco-(endo-)plasmicreticulum Ca‘+ pump (for review, see Inesi (1985)). The C terminally located calmodulin binding domain (James et al., 1988) plays a role in a number of regulatory mechanisms. It acts as an intramolecular inhibitor by binding close to the active site of the ATPase in the * The work was made possible by Swiss National Science Foun- dation Grant 31.30858.91. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. t Present address: Hoffmann-LaRoche AG, Basel, Switzerland. § To whom correspondence should be addressed: Institute of Bio- chemistry, Swiss Federal Institute of Technology (ETH), Universi- tatstrasse 16, 8092 Zurich, Switzerland. absence of calmodulin (Falchetto et al., 1991); it is involved in the oligomerization of thepump(Vorherr et al., 1991) which results in its activation; it participates in the response to activating phospholipids (Brodin et al., 1992); and it is the target site for the Ca2+-dependent protease calpain (James et al., 1989a). Finally, the C-terminal part of the PMCA’ also contains the target sites for kinase A (James et al., 1989b) and for kinase C (Wang et al., 1991). The C-terminal portion of the pump contains two highly acidic regions at the two sides of the CaM binding site (Shull and Greeb, 1988; Verma et al., 1988; Greeb and Shull, 1989; Strehler et al., 1990). The acidic character of these two sequences and their similarity to the first and fourth Ca‘+ binding sites of CaM (Verma et al., 1988) suggest their possible role in the binding of Ca2+. The aim of the work described here was to test this possibility. To this purpose, a recombinant C-terminal fragment of the Ca”-ATPase isoform l b (E1079-P1180; Verma et al., 1988) containing the CaM binding domain and the two adjacent acidic regions was expressed in Escherichia coli. The recom- binant fragment was indeed found to bind three Ca2+ ions with high affinity. Two were apparently constitutively bound to the domain since one resisted treatment with EGTA and the other was bound with an affinity corresponding toa KD of about 30 nM. The KD of the third Ca2+ binding site (about 300 nM) was in the range of the overall Ca2+ affinityof the pump, suggesting the possibility of reversible binding under native conditions and thus a role of the site in the regulation of the activityof the pump. EXPERIMENTAL PROCEDURES Materials-The vectors M13mp18/19, M13 primers, restriction enzymes, dNTPs, and CNBr-activated Sepharose 4B were from Phar- macia, Uppsala, Sweden. Taq polymerase and Taq buffer were ob- tained from Bohringer Mannheim, Rotkreuz, Switzerland. The se- quencing kit was from U. S. Biochemical Corp. The vector pJLA 502 was obtained from Medac, Hamburg, Germany. Mineral oil, Hepes, MOPS,dansyl chloride, phenylmethylsulfonyl fluoride, Triton X- 100, and L-a-phosphatidylcholine were purchased from Sigma. The second antibody (goat anti-rabbit peroxidase) was from Nordic Im- munological Laboratories, Tilburg, The Netherlands. 45Ca2+ was ob- tained from Amersham International, Amersham, United Kingdom. Spectrapor membrane tubing was purchased from Spectrum Medical Industries Inc., Los Angeles. Nitrocellulose was from Schleicher & Schuell,Dassel, Germany. Preparative and analytical HPLC were carried out using Applied Biosystems equipment (Foster City, CA) with Nucleosil reversed phase materials packed in Macherey & Nagel columns (Oensingen, Switzerland). Acetonitrile and 1-propanol for HPLC were obtained from Riedel de Haen, Seelze, Germany. Stains All and all other chemicals were of the best quality available from Fluka AG, Buchs, Switzerland. The abbreviations used are:PMCA, plasma membrane Caz+ ATP- ase; h, human; CaM, calmodulin; dNTP, deoxynucleotide triphos- phate; dansyl, 5-dimethylaminonaphthalene-1-sulfonyl; ESI, elec- trospray ionization; HPLC, high performance liquid chromatography; MOPS, 3-(N-morpholino)propanesulfonic acid Tricine, N- tris(hydroxymethy1)methylglycine. 10252
Transcript
THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1993 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 268, No. 14, Issue of May 15, pp. 10252-10259,1993 Printed in U. S. A.
The C-terminal Domain of the Plasma Membrane Ca2’ Pump Contains Three High Affinity Ca2’ Binding Sites*
(Received for publication, August 14, 1992, and in revised form, December 24, 1992)
Francesco Hofmann, Peter James, Thomas VorherrS, and Ernest0 CarafoliB From the Institute of Biochemistry, Swiss Federal Institute of Technology (ETH), 8092 Zurich, Switzerland
The C-terminal portion of the plasma membrane Ca2+- ATPase contains different regulatory domains. A re- combinant C-terminal fragment of the human plasma membrane CaZ’-ATPase l b isoform (E1079-P1180) was used to study the role of two acidic amino acid stretches located on either side of the calmodulin bind- ing domain, corresponding to synthetic peptides A18 (Vorherr, T., James, P., Krebs, J., Enyedi, A., Mc- Cormick, D. J., Penniston, J. T., and Carafoli, E. (1990) Biochemistry 29, 355-365) and B28 (James, P., Pruschy, M., Vorherr, T., Penniston, J. T., and Carafoli, E. (1989) Biochemistry 28, 4253-4258), re- spectively.
The molecular mass of the recombinant C-terminal fragment, as determined by electrospray ionization mass spectrometry, was higher by 39 mass units than the calculated value (12,055 Da). This difference was the result of an EGTA-insensitive Ca2+ ion, which was located by chymotryptic proteolysis in a fragment cor- responding to the last 37 amino acids of the expressed protein. Fluorescence experiments on the dansylated recombinant C-terminal fragment titrated with in- creasing amounts of free Ca2+ revealed two additional Ca2+ binding sites with affinities corresponding to KD values of about 30 and 300 nM, respectively. Stains All spectra of different synthetic peptides, corresponding to subdomains of the expressed protein, indicated that the site with the KD of 30 nM was probably located in the acidic sequence on the N-terminal side of the cal- modulin binding domain (peptide A18); the site with the 300 nM KD was apparently located on the C-ter- minal side of the calmodulin binding domain (peptide B28) or, alternatively, formed by the cooperation of distant residues of the domain.
The C-terminal portion of the plasma membrane Ca2+ pump (for review, see Carafoli (1992)) contains various regulatory domains. This distinguishes the pump from all other pumps of the P-class (Pedersen and Carafoli, 1987a, 1987b), e.g. the sarco-(endo-)plasmic reticulum Ca‘+ pump (for review, see Inesi (1985)). The C terminally located calmodulin binding domain (James et al., 1988) plays a role in a number of regulatory mechanisms. It acts as an intramolecular inhibitor by binding close to the active site of the ATPase in the
* The work was made possible by Swiss National Science Foun- dation Grant 31.30858.91. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
t Present address: Hoffmann-LaRoche AG, Basel, Switzerland. § To whom correspondence should be addressed: Institute of Bio-
chemistry, Swiss Federal Institute of Technology (ETH), Universi- tatstrasse 16, 8092 Zurich, Switzerland.
absence of calmodulin (Falchetto et al., 1991); it is involved in the oligomerization of the pump (Vorherr et al., 1991) which results in its activation; it participates in the response to activating phospholipids (Brodin et al., 1992); and it is the target site for the Ca2+-dependent protease calpain (James et al., 1989a). Finally, the C-terminal part of the PMCA’ also contains the target sites for kinase A (James et al., 1989b) and for kinase C (Wang et al., 1991). The C-terminal portion of the pump contains two highly acidic regions at the two sides of the CaM binding site (Shull and Greeb, 1988; Verma et al., 1988; Greeb and Shull, 1989; Strehler et al., 1990). The acidic character of these two sequences and their similarity to the first and fourth Ca‘+ binding sites of CaM (Verma et al., 1988) suggest their possible role in the binding of Ca2+. The aim of the work described here was to test this possibility. To this purpose, a recombinant C-terminal fragment of the Ca”-ATPase isoform l b (E1079-P1180; Verma et al., 1988) containing the CaM binding domain and the two adjacent acidic regions was expressed in Escherichia coli. The recom- binant fragment was indeed found to bind three Ca2+ ions with high affinity. Two were apparently constitutively bound to the domain since one resisted treatment with EGTA and the other was bound with an affinity corresponding to a KD of about 30 nM. The KD of the third Ca2+ binding site (about 300 nM) was in the range of the overall Ca2+ affinity of the pump, suggesting the possibility of reversible binding under native conditions and thus a role of the site in the regulation of the activity of the pump.
EXPERIMENTAL PROCEDURES
Materials-The vectors M13mp18/19, M13 primers, restriction enzymes, dNTPs, and CNBr-activated Sepharose 4B were from Phar- macia, Uppsala, Sweden. Taq polymerase and Taq buffer were ob- tained from Bohringer Mannheim, Rotkreuz, Switzerland. The se- quencing kit was from U. S. Biochemical Corp. The vector pJLA 502 was obtained from Medac, Hamburg, Germany. Mineral oil, Hepes, MOPS, dansyl chloride, phenylmethylsulfonyl fluoride, Triton X- 100, and L-a-phosphatidylcholine were purchased from Sigma. The second antibody (goat anti-rabbit peroxidase) was from Nordic Im- munological Laboratories, Tilburg, The Netherlands. 45Ca2+ was ob- tained from Amersham International, Amersham, United Kingdom. Spectrapor membrane tubing was purchased from Spectrum Medical Industries Inc., Los Angeles. Nitrocellulose was from Schleicher & Schuell, Dassel, Germany. Preparative and analytical HPLC were carried out using Applied Biosystems equipment (Foster City, CA) with Nucleosil reversed phase materials packed in Macherey & Nagel columns (Oensingen, Switzerland). Acetonitrile and 1-propanol for HPLC were obtained from Riedel de Haen, Seelze, Germany. Stains All and all other chemicals were of the best quality available from Fluka AG, Buchs, Switzerland.
The abbreviations used are: PMCA, plasma membrane Caz+ ATP- ase; h, human; CaM, calmodulin; dNTP, deoxynucleotide triphos- phate; dansyl, 5-dimethylaminonaphthalene-1-sulfonyl; ESI, elec- trospray ionization; HPLC, high performance liquid chromatography; MOPS, 3-(N-morpholino)propanesulfonic acid Tricine, N - tris(hydroxymethy1)methylglycine.
10252
Ca2+ Binding to the Ca2+ Pump 10253
Expression of Part of the C-terminal Domain of the hPMCAlb- The oligonucleotides for the amplification of the cDNA segment coding for the C-terminal domain of the pump were synthesized by the phosphite triester method (Letsinger et al., 1975; Beaucage and Caruthers, 1981) on a Pharmacia Gene Assembler using the procedure and the reagents supplied by Pharmacia LKB Biotechnology (Upps- ala, Sweden). The 5' primer was 25 nucleotides long and contained an NcoI restriction site and a methionine codon followed by the codons of the first 6 amino acids of the fragment to be expressed 5'- GGCCATGGAAGAAATACCTGAGGAG-3'. The 3' primer was 30 bases long, contained the complementary sequence of the codons for the last 6 amino acids of the fragment to be expressed followed by a stop codon and an NcoI restriction site: 3"GCATTGAGGTCAG-
The purification was carried out by reverse phase HPLC on a C- 18,5-pm, 300-A column applying the following gradient: 0-2 min 10% B, 2-42 min to 80% B, 42-47 min to 100% B. A contained 5% acetonitrile in 100 mM triethylammonium acetate, pH 7.0, and B contained 20% acetonitrile in 100 mM triethylammonium acetate, pH 7.0. The purified oligonucleotides were resuspended in water at a concentration of 0.4 nmollpl.
The amplification of the DNA coding for the fragment to he expressed was performed by polymerase chain reaction (Mullis and Faloona, 1987). 20 ng of a cDNA fragment isolated from a human teratoma NTera2D cell library (Verma et al., 1988) coding for the complete C terminus of the pump isoform hPMCAlb was used as a template. 20 pmol of each primer and 2.5 units of Taq polymerase were added to the template, and the reaction was carried out in Taq polymerase buffer in the presence of 200 p~ dNTPs. The mixture was covered with mineral oil, and 30 polymerase chain reaction cycles were performed (denaturation, 94 "C, 40 s; annealing, 55 "C, 40 s; elongation, 72 "C, 40 s). In the first cycle the denaturation time was elongated to 1.5 min. The molecular mass of the amplified material was checked on an agarose gel.
The amplified material was subcloned into the pJLA 502 vector (Schauder et al., 1987) at an NcoI restriction site, and the plasmid DNA was prepared as described in Maniatis et al. (1982). Nucleotide sequencing (Sanger et al., 1977) on single-stranded template DNA after subcloning an EcoRI-XhoI (blunt end) fragment into M13mp18/ 19 vector (Messing, 1983) digested with EcoRI and SmaI permitted the detection of colonies containing the insert in the correct orien- tation and to confirm the DNA sequence of the amplified material. Sequenase version 2 DNA polymerase, dGTP mixes, and universal MI3 primers were used as suggested by the supplier of the sequencing kit.
pJLA 502-derived expression plasmids were used to transform the protease-deficient E. coli strain Lon-hptr-teP (Goff et d . , 1984). Expression was carried out in LB medium containing 50 pg/ml ampicillin and 13 pg/ml tetracycline. Transformed E. coli cells were grown at 30 "C until an A,,,., of 1.0 was reached, and then the temperature was shifted to 42 "C to induce expression. After 16 h the cells were harvested by centrifugation.
Purification of the Expressed C-terminal Fragment-The harvested cells were resuspended in extraction buffer containing 20 mM Hepes, pH 7.5, 8 M urea, 130 mM NaCI, 0.1 mM CaCl,, 0.2 mM phenylmeth- ylsulfonyl fluoride, 0.1% Triton X-100 and sonicated in 50-ml por- tions for 15 min to extract the proteins. The sonicated material was centrifuged for 5 min at 3,000 rpm in a Sorvall SS34 rotor (Du Pont), and the supernatant was loaded on a Sepharose 4B-CaM column equilibrated with extraction buffer. The column was first washed with 3 bed volumes of 20 mM Hepes, pH 7.5,130 mM NaCI, 0.1 mM CaCl,, 0.4% Triton X-100; the second wash step was performed with 3 bed volumes of 20 mM Hepes, pH 7.5, 1 M NaCI, 0.1 mM CaC12 and was followed by a third overnight wash step with 5 bed volumes of 20 mM Hepes, pH 7.5, 130 mM NaCI, 0.1 mM CaC12. Elution was carried out with 20 mM Hepes, pH 7.5,130 mM NaCI, 2 mM NaEDTA, 4 M urea.
The expressed protein was purified further by reverse phase pre- parative HPLC on a C-8, 10 pm, 300 A, 250 X 22-mm column, by applying the following gradient 0-10 min 10% B, 10-40 min to 75% B, 40-50 min 75% B, 50-65 min to 80% B, 65-70 min to 100% B. A contained 0.1% trifluoroacetic acid, and B contained 50% 1-propanol and 0.05% trifluoroacetic acid. The purified material was lyophilized and stored at -20 "C.
SDS-Gel Electrophoresis and Immunostaining-The purified ma- terial was checkedon a 15% SDS-polyacrylamide gel (Laemmli, 1970). Part of the gel was stained with Coomassie Blue, and part was electroblotted onto nitrocellulose (Towbin et al., 1979). Immunostain- ing was carried out using a polyclonal antibody raised against the
GAGGTATCGGTACCTTC- 5'.
synthetic CaM binding peptide C20W as described previously (Vor- herr et a t , 1991).
Mass Spectroscopy and Sequencing-Electrospray ionization mass spectra were generated using a Finnigan MAT model 710 triple quadrupole mass spectrometer (Finnigan MAT, San Jose, CA) with a 20-kV conversion dynode and a 4,000 m/z mass range. The mass accuracy of the data acquired was routinely better than 0.01%.
Tryptic and Asp-N protease digestions of the protein were carried out using a 1:50 w/w pr0tease:suhstrate ratio for 18 h at 37 "C in 0.1 mM NaHC03, pH 8.0. A partial chymotryptic digestion was carried out using the same protein ratio but for only 5 min. The reaction was stopped by adding pure trifluoroacetic acid to yield a final pH of 2.0 followed by injection into an Applied Biosystems model 140A HPLC system. Peptides were separated on a Applied Biosystems aquapore C-8, 300-A reversed phase column using a gradient over 30 min, from 0.1% trifluoroacetic acid in water to 70% acetonitrile in water with 0.06% trifluoroacetic acid. A flow rate of 30 pl/min was used with a postcolumn split diverting 2 pl/min into the mass spectrometer. Spectra were accumulated with a scan rate of 400 atomic mass units/ s from 200 to 2,000 with a nominal resolution of 1,000.
Amino Acid Analysis-The concentration of peptide Met-A18-B28 in the solution used to run the induced coupled argon plasma calcium emission spectra was determined by amino acid analysis using an Applied Biosystems Derivatizer 420A with on-line phenylthiocarba- moyl detection with the Applied Biosystems Analyzer 130A.
Dansylation of the Expressed C-terminal Fragment of the Pump and Fluorescence Measurements-The lyophilized expressed protein was solubilized at a concentration of 0.5 mg/ml and dansylated in the presence of 100 p~ dansyl chloride as described previously (Vorherr et al., 1990). Absorption measurements at 320 nm using a molar extinction coefficient of 3,400 M" cm" (Chen, 1968) showed incor- poration of about 1.17 mol of dansyl chloride/mol of expressed peptide.
Fluorescence measurements of the dansylated expressed C-termi- nal fragment were performed with a SPEX fluorolog 1680 double- wavelength spectrophotometer connected to a DMlB coordinator (Metuchen, NY) (Vorherr et d. , 1990).
45Ca2+ Ouerlay-A Tricine gel was run according to the usual procedure (Schagger and von Jagow, 1987). The blotting onto nitro- cellulose and the 45Ca2+ overlay were carried out as described by Maruyama et al. (1984). The overlay was performed using 0.1 mCi of 45Ca2f, corresponding to a caz+ concentration of 88 pM.
Stains All Spectra-Stains All was dissolved at a concentration of 0.5 mM in ethylene glycol, and the spectra were recorded in 2 mM MOPS, pH 7.2,30% ethylene glycol as described by Caday and Steiner (1985). The peptides were incubated with 20 ~ L M Stains All in 4 ml of buffer at room temperature for 1 h with occasional stirring. The Stains All to peptide molar ratio was 1O:l in all experiments. After the incubation, the spectra were recorded from 700 to 400 nm using a Shimadzu UV-3000 spectrophotometer (Kyoto, Japan).
CaZf-ATPase Assay-The CaZ+-ATPase activity was measured by following the release of inorganic phosphate by the colorimetric method of Lanzetta et al. (1979). The calpain-digested Ca2+-ATPase was preincubated with the different peptides at different free CaZ+ concentrations in 20 mM Hepes, pH 7.2, 100 mM KC1, 1 mM MgCI,, 6.56 mM EGTA as indicated in the figure legends. The ATPase activity was measured at 660 nm after incubation at 37 "C for 20 or 40 min in the presence of 1 mM ATP.
CaZf-ATPase Preparation and Digestion with Calpain-CaM-free erythrocyte ghosts were prepared from outdated human erythrocyte concentrates obtained from the local Red Cross station, and the Ca2+ ATPase was isolated by CaM affinity chromatography (Niggli et al., 1979). L-a-Phosphatidylcholine was used as the stabilizing phospho- lipid in the EDTA eluate. The concentration of the purified Ca2+- ATPase was 200 pg/ml. The 124-kDa CaM-insensitive, activated Ca2+-ATPase fragment was obtained by calpain digestion as described by Falchetto et al. (1991) and used without further purification.
RESULTS
Construction of the Expression Vector for the C-terminal Fragment fMet-AlB-B28) of the hPMCAlb Isoform-The ex- pressed domain corresponded to the amino acid sequence contained between E-1079 and P-1180 of the hPMCAlb iso- form (Verma et al., 1988) preceded by a methionine residue. The 18 N-terminal amino acids of the domain, following the initiation Met, corresponded to synthetic peptide A18 (Vor-
10254 Cu2+ Binding to the Ca2+ Pump 1079 T C T T
t i i t t t t i i t i T T A A A A A A t t tf t tt ii ’i“
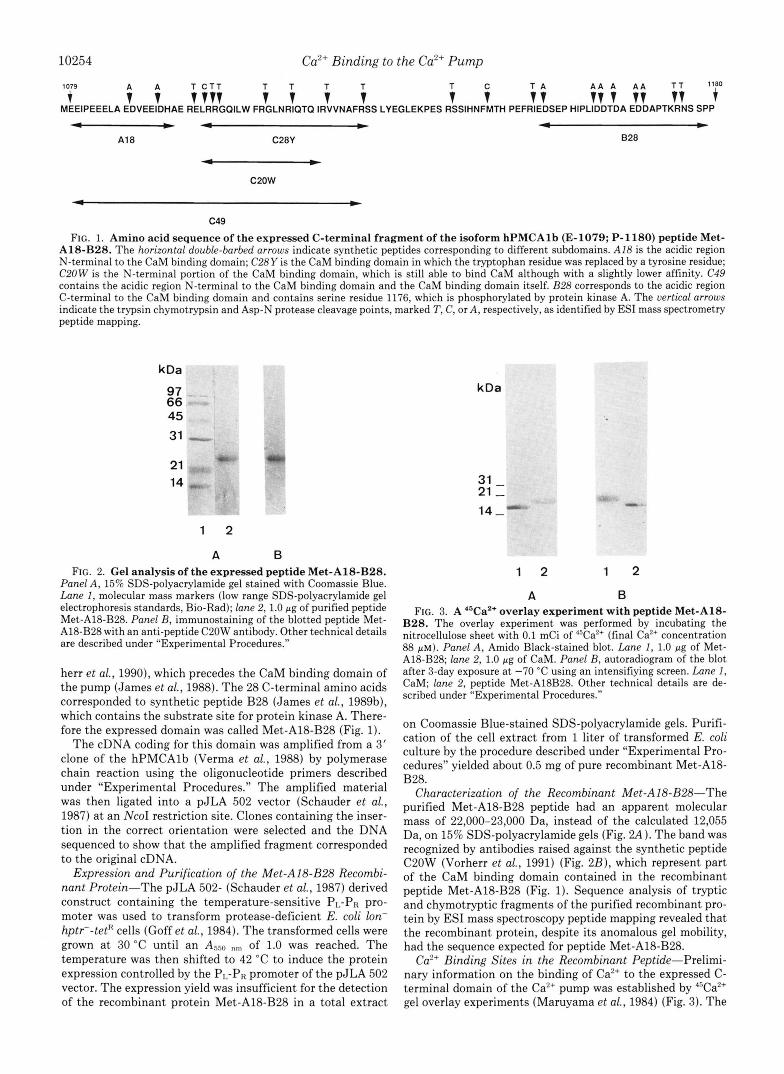
FIG. 1. Amino acid sequence of the expressed C-terminal fragment of the isoform hPMCAlb (E-1079; P-1180) peptide Met- A18-B28. The horizontal double-barbed arrows indicate synthetic peptides corresponding to different subdomains. A18 is the acidic region N-terminal to the CaM binding domain; C28Y is the CaM binding domain in which the tryptophan residue was replaced by a tyrosine residue; C20W is the N-terminal portion of the CaM binding domain, which is still able to bind CaM although with a slightly lower affinity. C49 contains the acidic region N-terminal to the CaM binding domain and the CaM binding domain itself. 828 corresponds to the acidic region C-terminal to the CaM binding domain and contains serine residue 1176, which is phosphorylated by protein kinase A. The uertical arrows indicate the trypsin chymotrypsin and Asp-N protease cleavage points, marked T, C, or A , respectively, as identified by ESI mass spectrometry peptide mapping.
kDa
97 66 45
31
21 14
wu t
1 2
A B FIG. 2. Gel analysis of the expressed peptide Met-A18-B28.
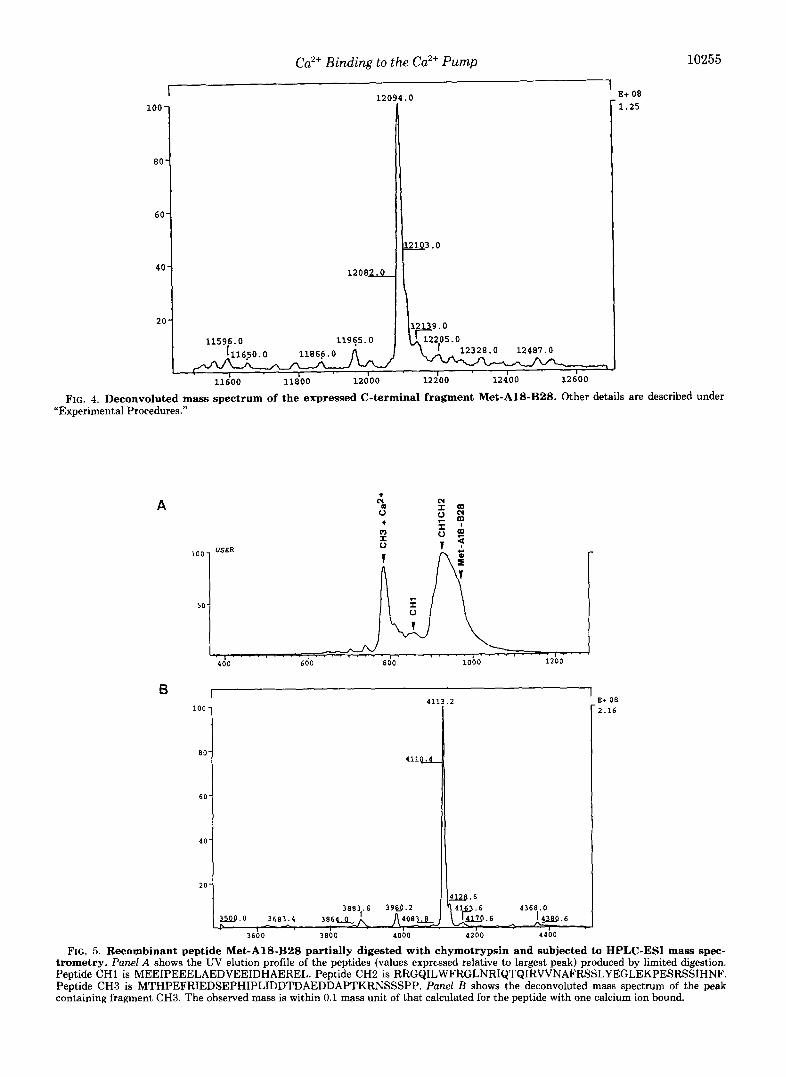
Panel A, 15% SDS-polyacrylamide gel stained with Coomassie Blue. Lane I, molecular mass markers (low range SDS-polyacrylamide gel electrophoresis standards, Bio-Rad); lane 2,l.O pg of purified peptide Met-A18-B28. Panel B, immunostaining of the blotted peptide Met- A18-B28 with an anti-peptide C20W antibody. Other technical details are described under “Experimental Procedures.”
herr et al., 1990), which precedes the CaM binding domain of the pump (James et al., 1988). The 28 C-terminal amino acids corresponded to synthetic peptide B28 (James et al., 1989b), which contains the substrate site for protein kinase A. There- fore the expressed domain was called Met-A18-B28 (Fig. 1).
The cDNA coding for this domain was amplified from a 3‘ clone of the hPMCAlb (Verma et al., 1988) by polymerase chain reaction using the oligonucleotide primers described under “Experimental Procedures.” The amplified material was then ligated into a pJLA 502 vector (Schauder et al., 1987) a t an NcoI restriction site. Clones containing the inser- tion in the correct orientation were selected and the DNA sequenced to show that the amplified fragment corresponded to the original cDNA.
Expression and Purification of the Met-A18-B28 Recombi- nant Protein-The pJLA 502- (Schauder et al., 1987) derived construct containing the temperature-sensitive PL-PR pro- moter was used to transform protease-deficient E. coli lon- hptr--tetK cells (Goff et al., 1984). The transformed cells were grown at 30 “C until an A,,,) ”,,, of 1.0 was reached. The temperature was then shifted to 42 “C to induce the protein expression controlled by the PI.-PR promoter of the pJLA 502 vector. The expression yield was insufficient for the detection of the recombinant protein Met-A18-B28 in a total extract
kDa
31 - 21 - 14-
1 2 1 2
A B FIG. 3. A 4aCa2+ overlay experiment with peptide Met-A18-
B28. The overlay experiment was performed by incubating the nitrocellulose sheet with 0.1 mCi of 45Ca’+ (final Ca2’ concentration 88 p ~ ) . Panel A , Amido Black-stained blot. Lane 1, 1.0 pg of Met- A18-B28; lane 2, 1.0 pg of CaM. Panel 8, autoradiogram of the blot after 3-day exposure at -70 “C using an intensifiying screen. Lane I, CaM; lane 2, peptide Met-A18B28. Other technical details are de- scribed under “Experimental Procedures.”
on Coomassie Blue-stained SDS-polyacrylamide gels. Purifi- cation of the cell extract from 1 liter of transformed E. coli culture by the procedure described under “Experimental Pro- cedures’’ yielded about 0.5 mg of pure recombinant Met-A18- B28.
Characterization of the Recombinant Met-AlB-B28--The purified Met-A18-B28 peptide had an apparent molecular mass of 22,000-23,000 Da, instead of the calculated 12,055 Da, on 15% SDS-polyacrylamide gels (Fig. Z4). The band was recognized by antibodies raised against the synthetic peptide C20W (Vorherr et al., 1991) (Fig. 2B), which represent part of the CaM binding domain contained in the recombinant peptide Met-AWB28 (Fig. 1). Sequence analysis of tryptic and chymotryptic fragments of the purified recombinant pro- tein by ESI mass spectroscopy peptide mapping revealed that the recombinant protein, despite its anomalous gel mobility, had the sequence expected for peptide Met-A18-B28.
ea2+ Binding Sites in the Recombinant Peptide-Prelimi- nary information on the binding of Ca2+ to the expressed C- terminal domain of the Ca“ pump was established by 45Ca2+ gel overlay experiments (Maruyama et al., 1984) (Fig. 3). The
Ca'+ Binding to the Ca2+ Pump 10255
I 1 E+ 08 I 12094.0 100 -
80-
60-
1.25
I
I
I I I
FIG. 4. Deconvoluted mass spectrum of the expressed C-terminal fragment Met-AlS-B28. Other details are described under "Experimental Procedures."
A
100
50
4d0 600 860 1000
6 I ' E+06 100
eo
6 0
40
20
411
4 l l U
2
W . 6
2.16
36b0 3 8 b O 40bO 42b0 44bO
FIG. 5. Recombinant peptide Met-A18-B2S partially digested with chymotrypsin and subjected to HPLC-ESI mass spec- trometry. Panel A shows the UV elution profile of the peptides (values expressed relative to largest peak) produced by limited digestion. Peptide CHI is MEEIPEEELAEDVEEIDHAEREL. Peptide CH2 is RRGQILWFRCLNRIQTQIRVVNAFRSSLYEGLEKPESRSSIHNF. Peptide CH3 is MTHPEFRIEDSEPHIPLIDDTDAEDDAPTKRNSSSPP. Panel B shows the deconvoluted mass spectrum of the peak containing fragment CH3. The observed mass is within 0.1 mass unit of that calculated for the peptide with one calcium ion bound.
10256 Ca2+ Binding to the Ca2+ Pump
1.7 1 I
1 '.IF" .o
Free Calcium / Magnesium nM
FIG. 6. Fluorescence titrations of dansyl-Met-A18-B28 with Ca2+. The dansylated peptide Met-A18-B28 (30 nM) in 20 mM Hepes, pH 7.2, 130 mM KC1, 0.5 mM EGTA was titrated with CaCI2 or MgCI,. The free Ca2+ concentration was calculated with the help of a computer program using a KL, of 0.21 PM for the Ca2+.EGTA complex (Martell and Smith, 1974). The figure shows a representative titration with CaC1, (filled circles) or MgClz (filled squares). The relative fluorescence intensities are plotted against the concentrations of free Ca2+ or free Mg".
autoradiogram in Fig. 3 shows that peptide Met-A18-B28 bound Ca2+; CaM was used as a positive control. The different gel mobility of the peptide Met-A18-B28 in this experiment, as compared with that shown in Fig. 2, was because of the use of a Tricine gel system.
The molecular mass of peptide Met-A18-B28 determined by mass spectroscopy was 39 mass units higher than the calculated mass of 12,055 Da (Fig. 4). Since the sequence of the expressed protein had been confirmed by mass spectros- copy peptide mapping, it was thought that this difference could be caused by a tightly bound Ca2+ ion. However, treat- ment of peptide Met-Al8-BZ8 with 0.5 mM EGTA did not eliminate the extra 39 mass units; only exhaustive trypsin and Asp-N protease digestions of the recombinant peptide brought the sum of the molecular masses of the proteolytic peptides exactly to the calculated 12,055 Da. That the extra 39 mass units determined by mass spectroscopy corresponded to a bound Ca2+ ion was then established directly by running an induced coupled argon plasma calcium emission spectrum (data not shown); a 2 PM peptide Met-A18-B28 solution (concentration determined by amino acid analysis) showed the presence of 3 PM Ca2+ over the background concentration of the water used to solubilize peptide Met-A18-B28 (0.75 PM), indicating that 1.5 mol of Caz+ were bound per mol of recombinant peptide. Mass spectroscopy peptide mapping of a partial chymotryptic digest of peptide Met-A18-B28 showed that the additional 39 mass units were associated with a chymotryptic fragment corresponding to the 37 C-terminal amino acids of the peptide (Figs. 1 and 5). The synthetic peptides B28, A18, and C49 did not bind Ca2+ and showed the expected molecular masses (data not shown).
When dansyl-Met-A18-B28 was titrated with increasing amounts of free Caz+ in a buffer containing 0.5 mM EGTA, a two-step fluorescence enhancement was observed (Fig. 6), indicating the presence of two additional Ca2+ binding sites. The two sites bound Ca2+ with KD values of about 30 and 300 nM, respectively. When the same experiment was carried out with increasing amounts of free Mg2+ instead of Ca2+ no
fluorescence changes were observed (Fig. 6). The expressed C-terminal domain contains two acidic re-
gions: one is located N terminally to the CaM binding domain and corresponds to the synthetic peptide A18 (Fig. I), the other one is located C terminally to the CaM binding domain and corresponds partially to the synthetic peptide B28 (Fig. 1).
The binding of Ca2+ to synthetic peptides A18, C49, and B28 (Fig. 1) was thus explored, taking advantage of the properties of the Stains All dye (Caday and Steiner, 1985), which interacts with acidic amino acids sequences in a Ca2+- dependent manner if the sequence represents a Caz+ binding site. The aim of the work was to locate the two putative Ca2+ binding sites identified in the fluorescence experiment. The Stains All spectra of both peptides A18 and B28 showed the typical J-band (610-650 nm) which corresponds to the binding of the dye within an acidic amino acid stretch. In analogy to those demonstrated for Ca2+-binding proteins (Caday and Steiner, 1985) the Stains All spectra of peptides A18 and B28 observed in the presence of EGTA showed a decrease of the absorption at the J-band after the addition of 1 mM CaCl,, indicating that both peptides bound Ca2+ (Fig. 7 A ) . The spectrum of peptide C49 (Fig. 11, which contains the acidic region A18 and the CaM binding domain, did not absorb in the J-band region, at variance with the results on the peptide A18. However, the J-band appeared in the Stains All spectrum of peptide C49 after the addition of 0.5 mM EGTA and decreased upon the addition of 1 mM CaCl, (Fig. 7A). A comparison of the Stains All spectra of peptides A18, C49, and B28 in presence and absence of EGTA (Fig. 7B) showed that the absorption at the J-band region was increased by EGTA for peptides A18 and C49, whereas almost no effect was seen with peptide B28. Thus, the peptides A18 and C49 were able to bind the traces of Ca2+ present in the medium in the absence of EGTA. The control peptide C20W, which represents part of the CaM binding domain and does not possess the features of a Ca2+ binding site, did not show absorption in the J-band region in the presence and in the absence of EGTA (data not shown).
Effects of the Recombinant Peptide Met-A18-B28 on the ATPase Activity-The C-terminal domain of the plasma membrane Ca2+ pump acts as an autoinhibitor of the activity of the pump (James et al., 1989a; Falchetto et al., 1991); a 124-kDa fragment obtained by calpain digestion (James et al., 1989a), which lacks the CaM binding domain and is fully active in the absence of CaM, is markedly inhibited by the synthetic versions of the CaM binding domain (peptides C28Y, C28W, C28A; Vorherr et al. (1990), Falchetto et al. (1991)). The recombinant peptide Met-A18-B28 also inhibited the ATPase activity of the 124-kDa calpain fragment (Fig. 8A), at 10 PM free Ca", as demonstrated previously with peptide C28Y. It inhibited by 50% at a concentration of 20 PM, whereas peptide C28Y had the same effect at a concen- tration of 10 PM. Since synthetic peptide C28Y does not contain Ca2+ binding sites, whereas peptide Met-A18-B28 apparently contains three, the effects of Ca2+ on the inhibition of the 124-kDa fragment of the Ca2+ pump were investigated. 10 PM peptide C28Y inhibited at higher Ca2+ concentrations than peptide Met-A18-B28 (Fig. 8B); the inhibitory effect of peptide Met-Al8-BZ8 became evident a t free Ca2+ concentra- tions higher than 150 nM, whereas that by peptide c28Y was only observed a t free Ca2+ concentrations higher than 300 nM. Increases in the concentration of C28Y did not affect the Ca2+ dependence of the inhibition (data not shown).
Ca2+ Binding to the Ca2+ Pump
A
10257
FIG. 7. Panel A , Stains All spectra of the peptides AH, B28, and C49. The spectra were recorded in the presence of 0.5 mM EGTA (solid line) and after ad- dition of 1.5 mM CaClz (dashed line). Panel B, Stains All spectra of peptides A18, B28, and C49 recorded in the pres- ence (solid line) and the absence (dashed line) of 0.5 mM EGTA.
B
600 700 600 700 600 700
0.8
0.6
0.4
0.2
0.0
600 7 0 0 6 0 0 700
WAVELENGHT [nm]
DISCUSSION
The C-terminal portion of the plasma membrane Ca2+ ATPase contains regulatory domains that have been charac- terized in some detail (James et al., 1988, 1989a, 198913; Vorherr et al., 1991; Falchetto et al., 1991). The region also contains two negatively charged sequences on either side of the CaM binding domain; the studies reported here show that they contain three Ca2+ binding sites.
The recombinant protein Met-A18-B28, corresponding to about 70% of the C-terminal domain of the hPMCAlb isoform (E1079-P1180) was used in the study. As expected, it cross- reacted with an antibody raised against the synthetic peptide C20W, corresponding to a subdomain of the expressed pro- tein; however, its mobility in a 15% SDS-polyacrylamide gel corresponded to a molecular mass of 22,000-23,000 Da instead of the calculated 12,055 Da. This anomalous gel mobility had already been observed for other proteins (Takano et al., 1988; Kleinschmidt et al., 1986), including the C-terminal domains of different plasma membrane Ca2+ ATPase isoforms (Kessler et al., 1992). Probably, the negatively charged stretches pres- ent in the region caused an electrostatic repulsion of SDS, resulting in decreased gel mobility; the expected amino acid sequence of the expressed product was confirmed by ESI mass spectrometry peptide mapping. The possibility that the 22- 23-kDa mass was caused by the formation of a dimer unre- solved by the gel system was ruled out by the absence of a dimeric species in both the ESI and matrix-assisted laser
desorption time-of-flight spectra (data not shown). The first indication that peptide Met-A18-B28 was able to
bind Ca2+ tightly came from the detection of 39 additional mass units when determining its mass by mass spectroscopy. The presence of a bound Caz+ was confirmed by the measure- ments of the CazC content of a 2.0 p M solution of the peptide by induced coupled argon plasma Ca2+ emission spectrum. However, the fact that 1.5 Ca2+ ions were detected per mole- cule of peptide Met-A18-B28 in the Ca2+ emission spectrum indicated at least one other Ca2+ binding site, which was not saturated under these conditions (see below). Since the first bound Ca2+ was insensitive to EGTA treatment, its binding site either had an extremely high affinity for Ca2+ ( i e . a subnanomolar KO) or was buried in the protein and therefore inaccessible to chelating agents. ESI mass spectroscopy pep- tide mapping analysis of a chymotryptic digest of the recom- binant peptide showed that its 37 C-terminal amino acids retained the ability to bind Ca2+ in such a tight (i .e. EGTA- insensitive) manner. However, the analysis also showed that synthetic peptide B28, which is contained in the aforemen- tioned 37-amino acid stretch, did not bind Ca2+ with equal affinity, since its mass corresponded exactly to that calculated from its sequence. Exhaustive trypsin and Asp-N protease digestions of peptide Met-A18-B28 also resulted in the loss of the bound calcium.
Titrations of dansyl-Met-A18-B28 in the presence of 0.5 mM EGTA with increasing amounts of Ca2+ showed two additional Ca2+ binding sites; the site with the higher affinity
10258 Ca2+ Binding to the Ca'+ Pump
Met-A18-828 concentration nM
0.0 ! I 0 100 200 300 400
Free calcium concentration nM FIG. 8. Effects of recombinant peptide Met-A18-B28 on the
activity of the 124-kDa fragment of the PM Ca2+-ATPase. The fragment was obtained by calpain digestion (James et al., 1989a). Panel A , 3 pg of calpain-digested Ca2+-ATPase was preincubated for 20 min at room temperature with different amounts of recombinant Met-A18-B28; the free Ca2+ concentration was 10 FM. The ATPase activity was measured after a 20-min incubation with 1 mM ATP at 37 "C and plotted as a percentage of its maximum against the con- centration of peptide Met-A18-B28 (representative curue for three experiments with very similar results). Panel B, effect of Ca2+ on the inhibition of the activity of the 124-kDa fragment by the recombinant peptide Met-A18-B28 and by peptide C28Y. The free Ca2+ concentra- tions were calculated with the help of a computer program using a K D of 0.21 FM for the Ca2+.EGTA complex (Martell and Smith, 1974). 3 Kg of calpain-digested Ca2+ ATPase was preincubated in the absence (circles) or presence of the recombinant peptide Met-AWB28 (20 p ~ ; triangles) and of peptide C28Y (10 p ~ ; squares) for 20 min at room temperature. The ATPase activity was measured after a 40-min incubation with 1 mM ATP at 37 "C and plotted against the free Ca2' concentration. The curues represent the average of four experiments; the error bars show the range between the maximal and minimal values obtained for each point in the four experiments.
had a KD of about 30 nM, i.e. this site was probably also constitutively present, considering that the cytoplasmic free Ca2+ concentration in the resting state is of the order of 100- 200 nM. The other Ca2+ binding site had a Ku of about 300 nM, i.e. it would be compatible with its reversible binding under in vivo conditions and thus with a role in regulation.
The Stains All dye experiments on different synthetic pep- tides corresponding to subdomains of peptide Met-A18-B28 identified the two acidic regions at the two sides of the CaM binding domain (A18 and B28) as Ca2+ binding sites. The finding that the binding of Stains All to peptide A18, and particularly to peptide C49, showed a clear sensitivity to
EGTA whereas the interaction of Stains All with peptide B28 was almost insensitive to it indicated that the Ca2+ binding site with a KD for Ca2+ of about 30 nM was N-terminal to the CaM binding domain, i.e. in the region corresponding to synthetic peptide A18. However, the complete sensitivity of the interaction of peptide C49 with Stains All to EGTA as compared with the only partial sensitivity of peptide A18 indicated that the latter did not contain all of the residues necessary for the binding and/or did not have the conforma- tion required to coordinate Ca2+ as tightly as peptide C49 or the recombinant Met-A18-B28. Peptide B28 bound Ca2+ with an affinity lower than that of peptide A18 since almost no changes in Stains All binding were brought about by the addition of EGTA. Peptide B28 thus either contained the Ca" binding site with lower affinity (KD about 300 nM) or formed part of the constitutive Ca2+ binding site that did not bind Ca2+ as tightly as the chymotryptic fragment that was 9 amino acids longer. If the second alternative were correct, the Caz+ binding site with a KD of about 300 nM could be formed with the contribution of (acidic) residues unrelated to the two acidic stretches in the primary structure.
In considering the results of experiments of this type, one has to be aware of the inherent limitations of the extrapola- tions of peptide work to the function of the intact protein. Bearing this in mind, a discussion of the possible functional implications of the results appears nevertheless permissible.
The tightly bound Caz+ at the A18 region and at the 37 C- terminal amino acids of peptide Met-A18-B28 probably has a structural role, since the affinity of the sites makes it almost impossible for them to be free of Ca2+ in vivo. For this reason, these sites have been defined in this work as constitutive binding sites. Different roles, e.g. structural stabilization of important domains in the neighborhood, or protection against proteolysis of exposed regions, have been postulated for con- stitutively bound Ca2+ ions in other proteins (Strynadka and James, 1991). This would be in line with the concept of structural differences in pump isoforms alternatively spliced at site C (Strehler, 1991), since the isoforms spliced in version "a" would lack the Ca'+ binding site on the C-terminal side of the CaM binding domain. The sequence of the A18 region is almost completely conserved in all isoforms, with the ex- ception of isoform 4, in which 2 amino acids (one a glutamic acid) are missing, and 2 more are replaced. One of the replace- ments is a glutamic acid for a lysine (Strehler, 1991). Although the precise location of the Ca2+ binding site defined here as constitutive in the A18 segment is not known, it appears possible that the replacements in isoform 4 would affect it, i.e. this site could be absent in this isoform.
By contrast, the Ca2+ binding site with the lower affinity is likely to play a functional role. A comparison of the Ca2+ effects on the inhibition of the ATPase activity of the calpain- truncated pump by a synthetic CaM binding domain (peptide C28Y) and by the recombinant peptide described here showed that the latter inhibited by about 50% at a concentration of 20 WM; at concentrations higher than 50 I .~M the inhibitory effect decreased, probably because of the formation of peptide aggregates in this concentration range. High molecular mass complexes (> IO5 Da) were in fact detected in ultracentrifu- gation experiments performed under these conditions. More- over, similar recombinant peptides corresponding to C-ter- minal domains of different hPMCA isoforms showed a strong tendency to form aggregates (Kessler et al., 1992). The recom- binant peptide inhibited optimally at free ca2+ concentrations higher than 150 nM, whereas the inhibition by peptide C28Y only became detectable at free Ca2+ concentrations higher than 300 nM. The finding would in principle support the
Ca2+ Binding to the Ca2+ Pump 10259
concept of a regulatory Ca2+ binding site in peptide Met-A18- B28, but alternative possibilities must also be considered. For example, Ca2+ could induce a conformational change of the 124-kDa calpain fragment, favoring its interaction with the CaM binding domain. It seems nevertheless attractive to suggest that the autoinhibitory function of the C-terminal portion of the pump (Falchetto et al., 1991) could be related to the presence of a Ca2+ binding site with a KD in the range of the overall Ca2+ affinity of the pump. This Ca2+ binding site could somehow control the access of Ca2+ to its catalytic site, which is presumably formed by some of the transmem- brane helices of the pump (Clarke et al., 1989). The absence of this Ca2+ binding site in the calpain-truncated pump or its displacement in the intact pump activated by CaM would permit free, unregulated access of Ca2+ to the catalytic site. Thus, the C-terminal domain of different pump isoforms lacking the putative regulatory Ca2+ binding site would differ both structurally (see above) and functionally. As for other possible Ca2+ binding sites in the pump molecule, e.g. the two EF-hand-type sequences (residues 22-33 and 310-321 of the hPMCAlb isoform) present in the N-terminal portion (Verma et al., 1988), they could also participate in the putative chan- neling pathway. One additional role of the Ca2+ binding sites in the C-terminal region of the pump could be the modulation of calpain sensitivity. The interaction of calpain with PEST sequences in the pump apparently requires a bridging Ca2+ ion (Wang et al., 1989); the acidic regions N- and C-terminal to the CaM binding domain indeed have high PEST sequences scores.
Having identified three Ca2+ binding sites in the C-terminal peptide of the pump, work is currently under way to essay their functional significance with site-directed mutagenesis work on the intact enzyme.
Acknowledgments-We are indebted to Werner Boll, Institute of Biochemistry, Swiss Federal Institute of Technology, Zurich, for the synthesis of the oligonucleotides and to Frank Arndt, Institute of Biotechnology, for help in setting up the large scale E. coli cultures.
REFERENCES Beaucage, S. L., and Caruthers, M. H. (1981) Tetrahedron Lett. 2 2 , 1859-1862 Brodin, P., Falchetto, R., Vorherr, T., and Carafoli, E. (1992) Eur, J. Biochem.
Caday, C. G., and Steiner, R. F. (1985) J . Biol. Chen . 260,5985-5990 Carafoli, E. (1992) J. Bid. Chem. 2 6 7 , 2115-2118
204,939-946
Chen, R. F. (1968) And. Biochem. 25,412-416 Clarke, D. M., Loo, T. W., Inesi, G., and MacLennan, D. H. (1989) Nature
Falchetto, R., Vorherr, T., Brunner, J., and Carafoli, E. (1991) J . Biol. Chem.
Goff, S. A., Lawrence, C. P., and Goldberg, A. L. (1984) Proc. Natl. Acad. Sci.
Greeb, J., and Shull, G. E. (1989).J. Bioi. Chem. 264, 18569-18576 Inesl, G. (1985) Annu. Reu. Physzol. 47,573-601 James, P., Maeda, M., Fischer, R., Verma, A. K., Krebs, J., Penniston, J. T.,
James, P., Vorherr, T., Krebs, J., Morelli, A,, Castello, G., McCormick, D. J., and Carafoli, E. (1988) J . Biol. Chem. 263,2905-2910
Penniston, J. T., De Flora, A,, and Carafoli, E. (1989a) J. Biol. Chem. 2 6 4 ,
239,476-478
266,2930-2936
U. S. A. 81,6647-6651
James, P., Pruschy, M., Vorherr, T., Penniston, J. T., and Carafoli, E. (1989b)
Kessler, F., Falchetto, R., Heim, R., Meili, R., Vorherr, T., Strehler, E.E., and
Kleinschmidt, J. A., Digwall, C., Maier, G., and Franke, W. W. (1986) EMBO
Laemmli, U. K. (1970) Nature 227,680-685 Lanzetta, P. A., Alvarez, L. J., Reinach, P. S., and Candia, 0. A. (1979) Anal.
Biochem. 100.95-97 Letsinger, R. L. Finnan, J. L. Heavner, G. A,, and Lunsford, W. B. (1975) J.
Am. Chen. Sic. 97, 3278-3179 Maniatis, T., Fritsch E. F., and Sambrook J. (1982) Molecular C h i n A
Martell, A., and Smith, R. (1974) Critical Stability Constants, Vol. 1, p, 269, Spring Harbor, NY
Maruyama, K., Mikawa, T., and Ebashi, S. (1984) J. Biochem. (Tokyo) 9 5 , Plenum Press, New York
511-519
8289-8296
Biochemrstry 28,4253-4258
Carafoli, E. (1992) Biochemistry 31, 11785-11792
J. 5,3547-3552
Messing, J. (1983) Methods Enzymol. 101 , 20-78 Mullis, K. B., and Faloona, F. A., (1987) Methods Enzymol. 155 , 335-350 Niggli, V., Penniston, J. T., and Carafoli, E. (1979) J . Biol. Chem. 254 , 9955-
~~ .~~
9958 Pedeisen, P. L., and Carafoli, E. (1987a) Trends Biochem. Sci. 12 146-150 Pedersen, P. L., and Carafoli, E. (1987b) Trends Biochem. Sci. 12: 186-189 Sanger, F., Nicklen, S., and Coulsen, A. R. (1977) Proc. Natl. Acad. Sci. U. S. A.
Schagger, H., and von Jagow, G. (1987) Anal. Biochem. 166,368-379 Schauder, B., Bliicker, H., Frank, R., and McCarthy, K. E. G. (1987) Gene
Shull, G. E.. and Greeb. J. (1988) J. Bbl. Chem. 263. 8646-8657
74,5463-5467
(Amst.) 5 2 , 279-283
Strehler, E:E. (1991) J: Membr. 'Bioi. 120 , 1-15- ' ~ ~ ~ ~ " . Strehler, E. E., James, P., Fischer, R. Heim R. Vorherr, T. Filoteo A. G.,
Strynadka, N. C. J., and James, M. N. G. (1991) Curr. Opin. Struct. Biol. 1 , Penniston, J. T., and Carafoli, E. (1i90) J. B i o i Chem. 265,'2835-&42
9n-w A Takano, E., Maki M., Mori, H., Hatanaka, M., Marti T. Titani K., Kannagi,
Towbln, M., Staehelin, T., and Gordon, J. (1979) Proc. Natl. Acad. Sei. U. S. A.
"I_ " _ R., poi, T., and'Murachi, T. (1988) Biochemistry 27, 1'96441932
76 A ? 5 L A ? 5 A Verma, A. K., Filoteo, A. G., Stanford, D. R., Wieben, E. D. Strehler, E. E.,
Fiscber, R., Heim, R., Vogel, G., Mathews, S., Strehler-Paie, M.-A,, James,
Chem. 263,14152-14159 P., Vorherr, T., Krebs, J., Penniston, J. T., and Carafoli, E. (1988) J. Biol.
Vorherr, T., James, P., Krebs J., Enyedi A. McCormick, D. J., Penniston, J.
Vorherr, T., Kessler, T., Hofmann, F.. and Carafoli, E. (1991) J . Bid. Chem. T., and Carafoli, E. (1990) hiochemist6 29,355-365
Wang, K. K. W., Villalobo, A., and Roufogalis, B. D. (1989) Biochem. J. 262 , 266,22-27
Wan K K W Wri ht, L. C., Machan, C. L., Allen, B. G., Conigrave, A. D., 693-706
anfRoufogal& B. 6. (1991) J. Biol. Chem. 266,9078-9085