Olfactory Ensheathing Glia: an investigation of factors affecting responsiveness of these cells in vitro and in vivo Thalles R.B. De Mello This thesis is presented for the degree of Doctor of Philosophy at The University of Western Australia School of Anatomy & Human Biology and School of Animal Biology 2006
Transcript
Olfactory Ensheathing Glia: an investigation of factors affecting responsiveness of these cells in
vitro and in vivo
Thalles R.B. De Mello
This thesis is presented for the degree of
Doctor of Philosophy
at The University of Western Australia
School of Anatomy & Human Biology and
School of Animal Biology
2006
ii
Abstract
Olfactory ensheathing glia (OEG) have been demonstrated to improve
functional and anatomical outcomes after injury to the nervous system and are
currently being trialled clinically. This thesis presents the investigation of two
important issues in OEG biology. The first study (Chapter 2) investigates
effects of different members of the neuregulin (NRG) family of molecules on
the proliferation of OEG, as a means of quickly obtaining large numbers of
cells for clinical or experimental use. We report that NRG-1β, but not NRG-
2α or NRG-3, has a significant proliferative effect. Furthermore, we report for
the first time that use of different mitogens (forskolin and pituitary extract)
commonly used to expand these cells in vitro, can have a significant effect on
the responsiveness of OEG to added NRG in subsequent mitogenic assays.
OEG grown initially with forskolin and pituitary extract exhibited increased
basal proliferation rates in comparison to OEG originally expanded without
these factors, and this increased rate of proliferation was sustained for at least
6 days following their withdrawal from the culture medium. We also report
for the first time the expression pattern of ErbB2, ErbB3 and ErbB4 receptors
on p75-selected OEG, and investigate their contribution to the NRG mitogenic
effect by the use of inhibitory ErbB antibodies.
Our second study (Chapter 3) seeks to clarify the role of OEG in promoting
myelination of central nervous system neurons. In this study we have
iii
investigated the myelinating ability of OEG derived from embryonic (EEG),
postnatal (PEG) and adult tissue (AEG) both in vitro and in vivo. OEG
selected by p75-immunopanning were co-cultured with dissociated cultures of
TrkA-dependant embryonic dorsal root ganglion (DRG) neurons. EEG, but
not AEG or PEG, successfully myelinated DRG neurons in the presence of
serum and/or ascorbate. AEG also failed to myelinate GDNF-dependant
embryonic DRG cultures, and growth factor-independent adult DRG cultures.
Transplantation of OEG into lysolecithin demyelinated spinal cord
demonstrated distinct ultrastructural differences between transplants of OEG
derived from animals of different ages. Furthermore, we demonstrate that
clearance of degraded myelin from the lesion site appears to be more effective
when animals are transplanted with EEG rather than AEG or Schwann cell
preparations. These results suggest that myelinating potential of OEG in vitro
and behaviour of these cells following transplantation in vivo are
developmentally regulated.
Together the two studies presented here constitute important evidence that
variations in extraction and expansion protocols can have a drastic effect on
behaviour of OEG both in vitro and in vivo, and arguably that these
differences may constitute a large source of variation between results
observed by different laboratories utilising OEG.
iv
Thesis Structure
This document is composed of four chapters.
Chapter 1 – Introduction
Chapter 1 constitutes an overview of literature covering the olfactory system,
an introduction to the use of olfactory ensheathing glia as cellular transplant
therapy to repair models of lesioned central nervous system, an overview of
myelin and the key myelin proteins investigated in this study, and an overview
of the role of neuregulins in the biology of peripheral nervous system
development.
Chapters 2 and 3
Chapters 2 and 3 describe in detail two separate studies investigating different
aspects of the biology of olfactory ensheathing glia. These chapters constitute
papers being currently prepared for submission to prominent scientific
journals (Glia and Journal of Neuroscience). As a result, they are self-
contained units detailing the studies described herein and follow a standard
paper format of a brief introduction, the materials and methods utilised, a
detailed description of the results, and a brief discussion highlighting issues of
primary importance.
v
Chapter 4 – Extended Discussion
Chapter 4 comprises an extended examination of the implications of the
results described in chapters 2 and 3 in the context of the available literature
of ensheathing cell biology. Wherever possible, the author has sought to
minimise duplication of points already covered in chapters 2 and 3. Rather,
this section attempts to expand upon those issues, and seeks to bring a sense
of context to the findings presented in this document. It is an attempt to
bridge the gap between reader and author, and details many of the thoughts,
ideas and questions arising as a result of this work, including suggestions by
the author regarding future studies that seek to investigate the biology of
p<0.001). No significant difference in the percentage of unmyelinated axons
was seen between medium alone, Schwann cell or AEG groups (p>0.05).
Despite the differences in appearance and myelination state between the
various groups, no significant difference was detected between total numbers
of axons present in each group (p>0.05).
Closer inspection of the lesion site in the AEG and EEG transplant groups
revealed that in all cases where peripheral myelin is seen, the myelinating
glial cell is present in a 1:1 relationship with the axon (Figure 13). These cells
Chapter 3 – Age-dependent myelination by OEG
93
possess a basal lamina (Figure 13d) and resemble Schwann cells.
Unfortunately, none of our transplanted Schwann cells or AEG demonstrated
any visible bluo-gal reaction product. Transplanted EEG demonstrated faint
electron dense precipitate (Figure 13c) characteristic of LacZ reaction
(Franklin and Barnett, 1991; Weis et al., 1991; Sekerkova et al., 1997; Boyd et
al., 2004a) but this was deemed too pale for accurate counts.
Ultrastructurally, unmyelinating AEG and EEG were recognisable by their
large amount of cytoplasm and numerous processes surrounding nearby axons
(Figure 13 b-c). Interestingly, the EEG transplant group always exhibited
EEG enveloping large clusters of myelinating cells that resembled Schwann
cells in appearance, but the EEG never contacted axons directly (Figure 13c).
Our observations support the data reported by Boyd et al. (2004a), who
described that unpurified EEG transplants form channels around central cores
of axons myelinated in a 1:1 relationship by glia resembling Schwann cells.
Discussion
We have shown that OEG derived from embryonic, but not adult or postnatal
animals, are capable of myelinating TrkA-dependent DRG neurons in vitro.
Furthermore, we have shown that the inability of AEG to myelinate DRG
axons in vitro is not related to axonal calibre. We have also demonstrated that
both AEG and EEG behave differently when transplanted into demyelinated
Chapter 3 – Age-dependent myelination by OEG
94
spinal cord in vivo. These results indicate a developmental regulation of OEG
behaviour and responses to factors both in vitro and in vivo.
Myelination by OEG in vitro
We have shown that adult and postnatally derived OEG cannot be induced to
myelinate TrkA-dependant DRG neurons under culture conditions containing
serum, ascorbate and progesterone. This finding agrees with that previously
reported by Plant et al. (2002), who failed to induce myelination of AEG
under conditions containing serum and ascorbate. Here we extend the
findings and report that whereas both AEG and PEG do not myelinate in the
presence of serum, ascorbate and progesterone in vitro, that embryonically
derived OEG do form myelin. Previously, others have shown that unpurified
populations of EEG can myelinate TrkA-dependent cultures under medium
conditions containing serum (Devon and Doucette, 1995). Here we report for
the first time that EEG preparations are also able to myelinate DRG neurites
under conditions containing ascorbate alone. Although we cannot
categorically rule out the possibility that contaminating Schwann cells in the
DRG preparation are responsible, we were unable to observe myelination in
the medium alone, AEG and PEG groups. We also cannot rule out the
possibility that Schwann cell precursors are responsible for the myelination
observed in the ascorbate-treated EEG group, though myelination in the
presence of serum alone (without ascorbate) would indicate that Schwann cell
precursors are not completely responsible for our observations. The data
Chapter 3 – Age-dependent myelination by OEG
95
suggests that if EEG are responsible for myelinating DRG neurites, then the
ability may be associated with the age of the animal and may be indicative of
a developmentally regulated variation in the extracted cells.
Our subsequent studies further support this hypothesis and demonstrate that
AEG appear incapable of myelination in vitro. AEG demonstrated no
preference for either axons or the collagen substrate in GDNF-dependent
DRG cultures, or in growth factor-independent adult DRG cultures. AEG also
demonstrated no visible staining for MAG, CNP or MBP in either GDNF or
growth factor-independent neuronal co-culture systems, either in the presence
of ascorbate or serum. Interestingly, AEG cultures constitutively express P0
protein both before and after co-culture with neurons, thus indicating that at
least in part they still possess intrinsic features that indicate they may be able
to myelinate neurons. It remains to be seen if AEG will in future be able to be
induced to myelinate if cultured under different medium conditions, or if the
basal expression of P0 by AEG is a developmental remnant from a stage
where EEG had the capacity to myelinate.
Myelination by OEG in vivo
We have performed for the first time a comparative in vivo study of the
myelination ability of OEG derived from animals of two different ages.
Lysolecithin-induced demyelinated areas of the dorsal funiculus bear a
distinctly varied ultrastructural appearance in all treatment groups examined at
Chapter 3 – Age-dependent myelination by OEG
96
both light and electron microscopic levels. Medium only controls exhibit
large number of axons surrounded by degraded myelin profiles, and very few
axons that are unmyelinated or enveloped by mature myelin sheaths. Our
Schwann cell controls demonstrate a marked increase in levels of intact
mature myelin, a finding that is supported by previous studies of Schwann
cells in demyelinating lesions of the CNS (Honmou et al.., 1996). The AEG
transplant group displays a similar proportion of myelinated profiles to the
Schwann cell transplant group, though a distinct difference is observed in the
ultrastructural appearance of the lesion site itself. Many previous in vivo
studies have argued that OEG (both purified and unpurified preparations) are
able to myelinate demyelinated neurons, and have consistently indicated that
OEG myelin is indistinguishable from Schwann cell myelin in terms of
conduction velocity, immunoreactivity and morphology (Franklin et al., 1996;
Barnett et al., 2000; Imaizumi et al., 2000a, b; Kato et al., 2000).
The peripheral myelin observed in our AEG transplant group is
ultrastructurally indistinguishable from Schwann cell myelin (Friede and
Samorajski, 1968), and several researchers have proposed that much of the
observed myelination may be due to increased recruitment of endogenous
Schwann cells into the lesion site (Boyd et al., 2004a, b; Ramer et al., 2004a;
Richter et al., 2005). Furthermore, it is well established that lysolecithin
induced lesions cause short term permeability to the blood brain barrier,
allowing for invasion of immune response cells into the lesion site (Ousman
Chapter 3 – Age-dependent myelination by OEG
97
and David, 2000). Although the timeframe in our experiments was designed
to avoid this phase, it is possible that transplantation was performed at a time
when such effects had not completely subsided, thus facilitating movement of
Schwann cells from the periphery into the site of AEG transplanted cells.
However, given the failure of our Lv-LacZ label, we cannot accurately
confirm that purified AEG are not responsible for myelination in our
lysolecithin induced lesions. Nevertheless, GFP-labelled AEG have been
shown to be responsible for the majority of peripheral-like myelin following
transplantation into spinal cord lesions (Sasaki et al., 2004) although
contamination by Schwann cells during OEG isolation remains to be
excluded.
The finding that spinal cords transplanted with purified AEG exhibit a random
distribution of myelinated and unmyelinated axons throughout the lesion site
is of interest. By contrast, two other groups have previously demonstrated
that animals transplanted with AEG form clusters of myelinating cells that are
sometimes surrounded by unmyelinating AEG (Li et al., 1997, 1998, 2003b;
Sasaki et al., 2004), though another group has failed to report these
observations (Takami et al., 2002). However, the studies used spinal cord
transection rather than lysolecithin demyelination as described here. It is
possible that the cluster arrangement is only seen in lesions where active
neuronal degeneration is taking place, rather than simple remyelination as in
our system. Most importantly however, is the fact that such clusters of
Chapter 3 – Age-dependent myelination by OEG
98
myelinated axons have been consistently observed in experimental models
utilising cells derived from younger animals. For example, clusters or
channels of myelinating axons are often seen in transplants of PEG into
chronically demyelinated spinal cord (Franklin et al., 1996; Smith et al., 2001;
Lakatos et al., 2003b), and transected spinal cord (Imaizumi et al., 2000b).
Indeed, the work reported here on EEG confirms the work of Boyd et al.,
(2004a) showing that transplanted EEG do not contact axons, but rather
surround clusters of myelinating cells resembling Schwann cells.
The age-dependent differences in behaviour of OEG transplants are further
emphasised by our observations on the myelinated status of axons within the
lesion site. Our results demonstrate that axons in the EEG and AEG
transplant groups possess a similar proportion of intact myelin to the Schwann
cell group. However, the proportion of axons surrounded by loose
uncompacted myelin is significantly less in EEG groups, and the proportion of
axons that are completely unmyelinated is correspondingly higher. The
observations indicate that OEG transplants behave in a developmentally-
dependant manner that can not only dramatically modify the ultrastructural
appearance of the lesion site, but also the extent of repair to the damaged
myelin. These findings are also suggestive of a developmental shift in the
properties of OEG populations, and suggest that age-related differences may
yet be found in the bulb in vivo despite recent findings (Magavi et al., 2005).
Chapter 3 – Age-dependent myelination by OEG
99
Acknowledgements
This work was supported by the UWA Small Grant, The Neurotrauma
Research Program of Western Australia and the Ramaciotti Foundation. Dr.
Giles Plant is an NHMRC RD Wright Research Fellow (ID# 303265) and A.
Prof. Sarah Dunlop is an NH&MRC Senior Research Fellow (ID# 254670).
Special thanks to Dr. Patrick Wood (The Miami Project to Cure Paralysis,
University of Miami School of Medicine, Miami, Florida), and to Dr. Bruce
Trapp (Department of Neurosciences, Lerner Research Institute, Cleveland,
Ohio) for providing us with several of the antibodies utilized in this study.
The RT97 monoclonal antibody, developed by Dr. John Wood, was obtained
from the Developmental Studies Hybridoma Bank developed under the
auspices of the NICHD and maintained by the Sandoz Institute for Medical
Research, London, UK. We would also like to thank A. Prof. Janet Keast
(University of New South Wales, Sydney, Australia) for providing us with
instruction in the culturing of adult DRG neurons. We also thank Michael
Archer, School of Animal Biology, The University of Western Australia for
assistance with the ultrastructural work.
100
Figure 1. Bluo Gal staining of adult OEG visualised under bright field microscopy. The
arrowhead indicates a cluster of densely labeled cells. The small arrow indicates an
unlabelled OEG. This image has been contrast enhanced for clear visualization of the
cells. Scale bar = 50 µm.
Figure 2. Confirmation of myelination by Schwann cells and unpurified EEG in a TrkA
selected DRG neuron co-culture system. Unpurified EEG (A, B, C) co-cultured with
dissociated neurons express MBP in the presence of 15% (v/v) FBS (A). Schwann cells
(D, E, F) also express MBP when co-cultured in the presence of 50 µg/ml ascorbate (D).
Red staining indicates presence of S100 protein in unpurified EEG co-cultures (B) and
Schwann cell co-cultures (E). The polyclonal S100 antibody used binds to both neuronal
and glial forms of S100, and provides a picture of all the cells within this system.
Hoechst 33343 staining is indicated for unpurified EEG (C) and Schwann cell (F) co-
101
cultures. A close association of glial cells with neuronal axons can be seen in both of
these cultures (arrows). Scale bar = 200 µm.
Figure 3. Immunofluorescence of glial cell/neuron co-cultures grown in the presence of
15% (v/v) FBS. Green denotes MBP staining, red denotes S100 staining and blue
denotes Hoechst 33343 staining of all cell nuclei. Shown are Schwann cells (A, E, I),
EEG (B,F,J), PEG (C,G,K) and AEG (D,H,L) co-cultures. Note that MBP staining is
only present in co-cultures containing purified EEG (B), but is absent from cultures
containing Schwann cells, PEG or AEG (A, C, D respectively). Also note that Hoechst
33343 staining appears to be tightly localised to neuronal axons in cultures containing
Schwann cells (I) and EEG (J) (arrows), but not in cultures containing PEG (K) or AEG
(L). Scale bar = 200 µm.
102
Figure 4. Co-cultures of TrkA-dependent embryonic DRG neurons with glial cells in the
presence of myelinating factors. Shown are phase images of co-cultures containing
Schwann cells in the presence of 50 µg/ml ascorbate (A), and p75-selected EEG (B),
unpurified EEG (C), p75 selected PEG (D) and p75 selected AEG (E) all cultured in the
presence of 15% (v/v) FBS. Shown in (F) is a purified AEG co-culture stained with
RT97 anti-neurofilament antibody (red) and a polyclonal S100 antibody (green) with
Hoechst 33343 (blue). Tubular structures characteristic of myelin sheaths are visible in
A, B and C. PEG and AEG (D and E respectively) (arrows) appear to show no
preference for neurons within the co-culture. This is more clearly evidenced by the
immunofluorescent image of an AEG co-culture shown in (F). Note that axons in (F)
appear orange-yellow due to the presence of both RT97 and S100 in the filaments. Scale
bar = 200 µm.
103
Figure 5. Quantitation of MBP levels detected on co-cultured TrkA-dependent
embryonic DRG neurons. (A) Nine defined fields were analysed in each DRG culture.
The illustrated culture was stained with Sudan Black and is provided courtesy of Dr.
Giles Plant. (B) Three lines were drawn within each defined field, and the number of
MBP positive axons crossing these lines were counted. The counted myelin segments per
field were averaged to obtain a measure of the amount of myelin in each culture. (C)
104
Indicated are levels of myelin present in cultures without glial cells, and in neuronal
cultures containing either Schwann cells, EEG, PEG or AEG. The different treatment
groups indicated include groups of either medium alone (-A), medium containing 50
µg/ml ascorbate (+A) or medium containing 15% (v/v) FBS (+S). Detectable levels of
myelin were only observed in co-cultures containing Schwann cells in the presence of
ascorbate, and in co-cultures containing EEG in the presence of either serum or
ascorbate. (D) Indicated are levels of myelin present in co-cultures containing 50 µg/ml
ascorbate, 15% (v/v) FBS, and 20 mM progesterone. The addition of progesterone does
not stimulate either PEG or AEG to myelinate, and does not significantly increase EEG
myelination (p>0.1). In all groups n=4-6. Error bars indicate ± s.e.m.
Figure 6. Co-culture of glia with GDNF-selected embryonic DRG neurons in the
presence of serum. Shown are Schwann cell co-cultures (A), and AEG co-cultures (B, C,
D). Images C and D illustrate the same field at different magnifications. Green denotes
MBP staining (not detectable), red denotes S100 staining and blue denotes Hoechst
33343 staining of all cell nuclei (colour composite appears as pink in the image). All
four images were taken from the edges of the culture where axons were present at lower
densities. AEG appear to associate into clusters, though no preference of these clusters
for either axons or collagen substrate was observed. No MBP immunoreactivity was
observed under these culture conditions. Scale bar = 200 µm.
105
Figure 7. Co-culture of AEG with GDNF-selected embryonic DRG neurons in the
presence of serum. AEG do not preferentially associate with neurons at the densely
populated centre of the neuronal cultures (A, C). Image B demonstrates the same field
depicted in A, and was immunostained with antibodies against β-III tubulin (red), MBP
(green, not detectable) and treated with Hoechst 33343 (blue) which stains all cell nuclei.
Field D illustrates the same field depicted in C, and was immunostained with antibodies
aganst β-III tubulin (red), MAG (green, not detectable) and treated with Hoechst 33343
(blue). Neither MBP nor MAG immunoreactivity is visible, and no preferrential
association of AEG with either neurons or substrate is observed. Scale bar = 200 µm.
106
Figure 8. AEG cultured in the presence of 1 ng/ml GDNF. Field A was immunostained
with antibodies against p75 (green) and S100 (red). Nearly 100% of all cells were
positive for these two markers. Field B was immunostained with antibodies against p75
(green) and GFAP (red), and once again nearly 100% of cells analysed in our purified
AEG cultures expressed both of these proteins. Field C was immunostained with
antibodies against P0 (red) and p75 (green). Nearly 100% of AEG express basal levels of
P0. Scale bar = 100 µm.
Figure 9. Co-culture of AEG with growth factor-independent adult DRG neurons in the
presence of serum. This field was immunostained with antibodies against P0 (red) and
RT97 neurofilament (green), and treated with Hoechst 33343 (blue) which stains all cell
nuclei. AEG express basal levels of P0, but demonstrate no preferential association with
either neurons or substrate. Scale bar = 200 µm.
107
Figure 10. Toluidine Blue staining of demyelinated dorsal funiculus at 19 days
(continued next page).
108
Figure 10. Toluidine Blue staining of demyelinated dorsal funiculus at 19 days.
Indicated are the medium control (A, B), Schwann cell transplant group (C, D), AEG
transplant group (E, F) and EEG group (G, H). Note that axons are evenly distributed
throughout the lesion site in the control (A, B) and Schwann cell groups (C, D), with
more myelinated profiles evident in the Schwann cell group. This same distribution is
not evident in the AEG group (E, F), with axons appearing less frequently and irregularly
spaced. The presence of large quantities of cytoplasm and extracellular matrix is evident
(large arrows). The EEG group (G, H) appears to demonstrate clusters of neurons and
glia that are not seen in any of the other groups. Inflammatory cells are present in
abundance (small arrows). Scale bars = 200 µm.
109
Figure 11. Electron Micrographs of demyelinated dorsal funiculus. Shown are
representative images of the medium control group (A), Schwann cell transplant group
(B), AEG transplant group (C) and EEG group (D). Axons are densely distributed
throughout the lesion in both medium control (A) and Schwann cell groups (B), with
degraded myelin profiles abundantly present in the medium control group. Large
quantities of cytoplasm and extracellular matrix separate individual axons throughout the
lesion site in the AEG transplant group (C). Axons in the EEG group are distinctly
fasciculated (arrows), and very small amounts of degraded myelin profiles are present
(D). Scale bar = 10 µm.
110
Figure 12. Quantification of myelination state. Shown are percentage counts of axons
from within the lesion site of the four different transplant groups. Axons possessing
intact myelin, degraded myelin or an unmyelinated state are represented. All transplanted
glial cell types significantly decrease the amount of degraded myelin present, and
increase levels of intact myelin. EEG in particular demonstrate a large decrease in
degraded myelin profiles and a corresponding increase in both unmyelinated and intact
myelin profiles. In all cases n=3 counted images at x1600 magnification. * indicates p <
0.05, ** indicates p < 0.01, *** indicates p < 0.001. All indicated statistical comparisons
were made against the corresponding medium only control groups. The table indicates
raw counts.
111
Figure 13. Electron micrographs of demyelinated dorsal funiculus. Depicted are AEG
transplant groups (A, B) and EEG transplant groups (C, D). Myelinated profiles are
abundant in the AEG group (A), with peripheral type myelin seen in a 1:1 relationship
with axons (black arrows). (B) AEG (white arrows) are present and discerned by their
ability to extend processes (small black arrows) around several axons (*). (C) EEG
(large arrow) are never seen to directly myelinate axons, but rather to engulf cells that
maintain a 1:1 relationship with axons (white arrowheads). (D) The myelinating cells
maintain a 1:1 relationship with axons (*) and possess a basal lamina (small arrows), a
characteristic feature of Schwann cells. Scale bars for A and C = 8 µm. Scale bars for B
and D = 2 µm.
CHAPTER 4
Extended Discussion
Chapter 4 – Extended Discussion
113
Part I
Summary
We have presented here two studies analysing different aspects of OEG
biology. In the first study (Chapter 2) we reported the effects of added
neuregulin (NRG) isoforms on the proliferation of OEG in vitro. We
demonstrated that culture conditions during the purification and expansion
phases, prior to performing a mitogenic assay, are crucial determinants of the
responsiveness of adult olfactory ensheathing glia (AEG) to added NRG.
Added mitogens to the culture medium such as forskolin and pituitary extract
can mask the responsiveness of AEG to NRG, and increase the base
proliferation rate of AEG for at least six days subsequent to their removal.
We have also reported the expression and activation patterns of ErbB
receptor subpopulations in purified p75-selected AEG, investigated their
functional role to AEG proliferation by use of ErbB antibody inhibitors, and
have related our data to previously published results in the literature. This
comparison illustrates that perceived differences in the published expression
of ErbB receptors may be attributable to variations in age of preparation and
purification techniques employed in other laboratories.
Influence of purification techniques on ErbB receptor expression
One indication that purification techniques may have an important role in
observations of OEG biology stems from our reported ErbB expression
profile utilising p75-purified AEG (Chapter 2). Here we reported that
Chapter 4 – Extended Discussion
114
ErbB2, ErbB3 and ErbB4 are expressed by AEG. These results are contrary
to previously published observations utilising postnatal olfactory ensheathing
glia (PEG; P2-P7) purified by selection for the O4 antigen and expanded
under similar culture conditions (Pollock et al., 1999; Thompson et al., 2000)
(summarised in Table 1). The discrepancy between the expression profiles of
AEG and PEG is not altogether unexpected in light of recently published
studies.
Table 1. Studies reporting mitogenic effect of NRG-1β on OEG and/or ErbB receptor
expression on OEG. Listed are the tissue source of cells utilised, the purification
techniques, culture conditions under which cells were expanded, and a summary of the
observations. Although several studies have reported increased proliferation of OEG in
the presence of NRG-1β, only a few of those studies performed concentration dose
response curves (DRC) for OEG proliferation.
Study Cells used Purification Culture Observations Chuah et al., 2000 OB-PEG AraC BPE weak proliferative DRC Pollock et al., 1999 OB-PEG O4 FACS proliferative DRC express ErbB2 + ErbB4 not ErbB3 Alexander OB-PEG O4 FACS proliferation et al., 2002 Thompson OB-PEG O4 FACS ErbB4 mRNA et al., 2000 no ErbB3 mRNA Moreno-Flores OB-P21 unpurified BPE express ErbB2 strongly, et al., 2003 forskolin ErbB3 + ErbB4 weakly Yan et al., 2001a OB-AEG p75 IP proliferative DRC Yan et al., 2001b OB-AEG p75 IP proliferation Key: OB-xEG = OEG derived from the olfactory bulb. OB-P21 = OEG derived from the olfactory bulb of young P21 animals. IP = immunopanning. FACS = fluorescence activated cell sorting. BPE = bovine pituitary extract. AraC = cytosine arabinoside. DRC = NRG-1β dose response curve.
Chapter 4 – Extended Discussion
115
Previous studies have already shown that different populations of OEG are
likely to be present within the olfactory nerve layer of the olfactory bulb (Au
et al., 2002). According to Kumar et al., (2005), only 21.3% of cells
extracted from the adult olfactory bulb are O4-positive, and of these only up
to 5% co-express p75. They also report that less than 10% of O4-positive
cells express GFAP in culture whereas 70-80% of p75 positive cells express
this marker, indicating that O4-positive and p75-positive cells may constitute
distinct populations from within the olfactory bulb. Wewetzer et al., (2005)
has also suggested that selection of PEG using O4 antigen does not occur by
means of a marker that is specific to PEG, but rather by selection of PEG that
have attached fragments of olfactory receptor neurons on their cell surface.
Although the authors also report that O4-selected PEG internalise these
axonal fragments and begin to express p75 after several days in culture, they
did not carry out comparative stains with other OEG markers such as GFAP
or S100 (Wewetzer et al., 2005). As such, the possibility remains that
selection of OEG for the O4 antigen may select a mixed population of OEG
and other cell types with O4 positive fragments of their cell surface, such as
astrocytes, fibroblasts, non-myelinating Schwann cells or oligodendrocyte
precursors.
Although the study of Moreno-Flores et al., (2003) appears to agree with our
reported expression of ErbB receptors (Table 1), one must be careful in
interpreting their study as confirmation of AEG receptor expression. In their
Chapter 4 – Extended Discussion
116
study, Moreno-Flores et al., (2003) utilised an unpurified population of
young P21 OEG. These unpurified cells were expanded in the presence of
pituitary extract and forskolin prior to analysis. Although it is possible that
an unpurified population would contain a higher proportion of OEG that are
p75-positive prior to extraction from the olfactory bulb, this population also
includes a large number of olfactory fibroblasts or meningeal cells (Barber
and Lindsay, 1982).
The question as to proliferation of contaminants is particularly important
even in purified preparations of OEG. Several different approaches have
been devised for purification of OEG. These include removal of rapidly
proliferating cells by treatment with anti-mitotics (Vincent et al., 2003),
treatment with neurotrophins (Bianco et al., 2004), positive immunoselection
for cells that express the p75 low affinity neurotrophin receptor (Ramon-
Cueto and Nieto-Sampedro, 1994; Gudino-Cabrera and Nieto-Sampedro,
1996; Ramon-Cueto et al., 1998) or the O4 antigen (Barnett et al., 1993),
negative selection of contaminating cells by means such as sorting for the
Thy-1.1 fibroblast marker (Chuah and Au, 1993; Gudino-Cabrera and Nieto-
Sampedro, 1996), differential attachment of astrocytes, macrophages and
microglia (Nash et al., 2001; Wang et al., 2005), and removal of
contaminants by a short period of trypsinisation (Ramon-Cueto and Nieto-
Sampedro, 1992). Despite the different purification techniques employed, all
of these studies still report the presence of contaminating cells in their
Chapter 4 – Extended Discussion
117
preparations: 8% for treatment with cytosine arabinoside (Vincent et al.,
2003), 20% for unpurified populations treated with neurotrophins (Bianco et
al., 2004), 3% for p75 magnetic activated cell sorting (MACS) (Gudino-
Cabrera and Nieto-Sampedro, 1996), 5-15% for p75 immunopanning
(Ramon-Cueto and Nieto-Sampedro, 1994), 3-30% for O4 selection (Barnett
et al., 1993; Franceschini and Barnett, 1996; Riddell et al., 2004), 7-20% for
differential attachment (Nash et al., 2001; Lipson et al., 2003), and 5-28%
with methods utilising removal of fibroblasts as the primary means of
purification (Chuah and Au, 1993; Gudino-Cabrera and Nieto-Sampedro,
1996). It is currently unknown if these contaminating cells express ErbB
receptors, or if these cells are able to proliferate at a rate comparable to that
of OEG once mitogens such as pituitary extract and forskolin are included in
the culture media.
Influence of tissue age on ErbB receptor expression
The age of animals from which OEG preparations are derived is another
possible variable that may account for contradictory reports on ErbB
expression. Studies utilising PEG report the expression of ErbB2 and ErbB4,
but not ErbB3 mRNA and protein (Pollock et al., 1999; Thompson et al.,
2000). Our results indicate that ErbB2 and ErbB3 mRNA and protein are
strongly expressed, and that ErbB4 protein but not mRNA is detectable in
AEG cultures (Table 2). We cannot disregard the notion that variations in
the age of preparation may play a role in this observed difference between
Chapter 4 – Extended Discussion
118
ours and other published results. This hypothesis is supported in part by the
study of Moreno-Flores et al., (2003), who utilized young P21 animals. The
authors report that ErbB2, ErbB3 and ErbB4 proteins are expressed, in
agreement with our findings and differing from studies utilising cells derived
from younger animals (Pollock et al., 1999; Thompson et al., 2000).
Other circumstantial evidence supports the hypothesis that cultured OEG
may retain developmentally regulated expression patterns (Table 2). For
example, PEG have been reported to express TrkB, but not TrkA or TrkC
receptor protein and mRNA (Woodhall et al., 2001; Vincent et al., 2003).
Meanwhile, AEG have been reported to express both protein and mRNA for
TrkA, TrkB and TrkC receptors (Bianco et al., 2004). Nevertheless, given
that these cells have been purified with different techniques, these observed
correlations do not in any way constitute proof that age of preparation may
have an effect on the expression profile of OEG in vitro. Detailed
comparative studies utilising standardised culture and purification protocols
will be nescessary to ascertain this hypothesis. However, further evidence
for potential developmentally regulated differences between OEG
populations is discussed in Part II of this chapter.
Chapter 4 – Extended Discussion
119
Table 2. Reported expression patterns across three different ages of preparations. Included are only those factors for which enough data is available to allow a comparison to be made between ages of preparation. Indicated in bold are negative results reported for the studies listed. Key: prot = reported protein expression, sec = reported secretory protein, mRNA = reported expression of mRNA. Neurotrophin PEG P21 AEG NGF prot2, sec1, mRNA1 prot4, mRNA3 BDNF prot2, sec1, mRNA1 prot4, sec6, mRNA3,5,
Study Cells used Purification Culture Cond. 1Woodhall et al., 2001 OB-PEG AraC BPE 2Vincent et al., 2003 OB-PEG AraC BPE 3Lipson et al., 2003 OB-AEG DI 4Liu et al., 2005 LP-AEG DI 5Byrnes et al., 2005 OB-AEG DI 6Ruitenberg et al., 2003 OB-AEG p75 IP BPE, forskolin 7Bianco et al., 2004 LP-AEG unpurified 8Pollock et al., 1999 OB-PEG O4 FACS 9Thompson et al., 2000 OB-PEG O4 FACS 10de Mello et al., (Chapter 2) OB-AEG p75 IP with and without
BPE and forskolin
11Moreno-Flores OB-P21 unpurified BPE, forskolin et al., (2003) Key: OB-xEG = OEG derived from the olfactory bulb. LP-PEG = PEG derived from the olfactory lamina propria. AraC = cytosine arabinoside. IP = immunopanning, DI = differential attachment to remove macrophages and microglia. FACS = fluorescence activated cell sorting. CML = complement mediated lysis. BPE = bovine pituitary extract.
Chapter 4 – Extended Discussion
120
Observed Mitogenic Effect of NRG on AEG
Several studies have previously reported that NRG-1β is a potent mitogen for
OEG (Pollock et al., 1999; Chuah et al., 2000; Yan et al., 2001a, b;
Alexander et al., 2002). However, only three performed dose response
curves to NRG-1β (Table 1), and the only study that is directly comparable to
our experiments in terms of age of preparation, purification methodology,
and culture conditions is that of Yan et al., (2001a). Not surprisingly, we
obtain comparable results to theirs for proliferation of AEG, with similar
dose response curves and similar levels of proliferation prior to addition of
NRG to the culture medium. Our observations indicate that peak
proliferation in the presence of NRG-1β was approximately 19%, 19% in the
presence of 2 µM forskolin, 38% in the presence of bovine pituitary extract,
45% in the presence of DF10S+mit medium (contains serum, forskolin and
pituitary extract), 45% in the presence of both NRG-1β and pituitary extract,
42% in the presence of both NRG-1β and forskolin, and 42% in the presence
of foskolin and pituitary extract. From these results we see that there is no
significant difference on AEG proliferation between any of the groups where
growth factors were tested in combination (Chapter 2). Furthermore, there is
no added benefit of including bovine pituitary extract together with NRG-1β
in the culture medium. This is likely due to the presence of NRG-1β in
pituitary extract mixtures (Raff et al., 1978; Brockes et al., 1980).
Interestingly, NRG-1β and forskolin act together to produce an additive
effect on proliferative rates, though their combined rate of proliferation is
Chapter 4 – Extended Discussion
121
once again not significantly different to that observed with pituitary extract
alone. The significance of these findings lies in the applicability of our
results to the rapid enhancement of AEG proliferation in vitro.
We suggest here that the utilisation of bovine pituitary extract in culture
media may be superfluous for the short-term proliferation of AEG. The
proliferative effect produced by a combination of NRG-1β and forskolin is
comparable to that of DF10S+mit medium. Furthermore, it has been
previously demonstrated that addition of a forskolin/NRG-1β combination to
cultured PEG is able to remove the proliferation arrest induced by astrocyte
conditioned medium, and that this proliferative effect is maintained over long
periods in culture (Alexander et al., 2002). Removal of pituitary extract from
AEG expansion protocols would eliminate another potential source of
variability found in experimental procedures, as the exact contents of
pituitary extracts vary depending on individual stock numbers and
manufacturers. Furthermore, pituitary extract contains other AEG
proliferative factors including FGF and PDGF, whose effect on other aspects
of AEG biology are still unknown (Gospodarowics et al., 1983; Ueno et al.,
1986; Halper et al., 1992; Chuah and Teague, 1999; Pollock et al., 1999; Au
and Roskams, 2003; Yan et al., 2001a, 2003).
We do however hesitate to recommend that the combination of NRG-1β and
forskolin be used to rapidly proliferate OEG preparations in vitro. Firstly,
Chapter 4 – Extended Discussion
122
we do not yet know whether p75-selected preparations from younger animals
(EEG or PEG) express the various ErbB receptor subtypes, or if NRG-1β is
capable of exerting a mitogenic effect upon those cells. Secondly, we do not
at this stage fully understand the implications of utilising these factors to
proliferate OEG in vitro, and the subsequent impact on proliferation and axon
interactions after transplantation in vivo. For example, application of cAMP
analogues to astrocytes in vitro have been found to induce measurable effects
post-transplantation (Chu et al., 1999). The converse is also true, with
injections of cAMP analogues in vivo demonstrating distinct effects several
days later on the extracted cells in vitro (Neumann et al., 2002). Vincent et
al., (2003) have also demonstrated that alterations of intracellular cAMP
levels can provoke drastic morphologic shifts in cultured PEG, and
hypothesised that OEG may be incredibly plastic cells capable of adjusting to
new environments by adopting a variety of different phenotypes. Given that
forskolin is a cAMP-inducing agent (Seamon et al., 1981; Fradkin et al.,
1982), we would strongly recommend that further studies be performed into
the effects of forskolin on cultured OEG in vitro and after transplantation in
vivo.
Chapter 4 – Extended Discussion
123
Part II
Summary
Our second study (Chapter 3) investigated the ability of OEG derived from
three different ages of animals to myelinate DRG neurons in vitro, and to
remyelinate the demyelinated dorsal funiculus of rats in vivo. We
demonstrated that age of preparation is a possible determinant of myelination
potential of OEG, and that this potential is unaffected by either axonal calibre
or the addition of factors known to enhance Schwann cell myelination in
vitro. That is, in our hands, EEG are able to produce myelin in vitro but PEG
and AEG are not. Furthermore, our results show that preparations derived
from embryonic animals interact very differently with the demyelinated
spinal cord environment, and that clearance of degraded myelin appears to be
more effective when animals are transplanted with EEG rather than AEG or
Schwann cell preparations.
Interaction of OEG with axons
The failure of our Lv-LacZ label to provide a strong consistent signal was
unfortunate in that we were consequently not able to answer the question as
to whether OEG can directly myelinate axons in vivo. Though faint label
could be visualised within non-myelinating cells of the EEG transplant
groups, no label was seen in any of the myelinating cells. Furthermore, no
label was visualised in electron micrographs of Schwann cell and AEG-
Chapter 4 – Extended Discussion
124
transplanted groups. This brings to light the question of whether the
lentivirus label is being downregulated in cells closely associated with axons,
and whether AEG do indeed have the capacity to myelinate as suggested by
several researchers (Sasaki et al., 2004; Li et al., 1997, 1998, 2003b).
All the data to date indicate that AEG appear to lack the ability to myelinate
axons in vitro under all culture conditions tested so far (Plant et al., 2002; de
Mello et al., Chapter 3). Here we provide, to our knowledge for the first
time, that the ability of OEG to myelinate DRG axons in vitro may be
dependant primarily on the age of the animal from which the glial cells were
extracted. Furthermore, we have demonstrated that there is an age-dependent
variation in the behaviour of OEG following transplantation into a
demyelinating lesion of the adult rat spinal cord, and that OEG derived from
embryonic animals appear to be significantly more effective at inducing
clearance of degraded myelin from chemically demyelinated dorsal
funiculus. Still unadressed by our study however, is the underlying
mechanism as to why EEG appear to be significantly better at preserving the
integrity of the lesion site following a demyelinating lesion, and which
mechanisms are responsible for the increased clearance of degraded myelin
profiles observed within our EEG group. Our data suggest that the answer to
this question appears to lie in the ability of the different transplant groups to
induce phagocytosis of degraded myelin within the lesion site.
Chapter 4 – Extended Discussion
125
Mechanisms of action by OEG in vivo
Though our study did not investigate the effect on recruitment of phagocytic
cells into the demyelinated area, it nevertheless remains likely that the
observed increase in clearance of degraded myelin must be, at least in part,
due to the effect of EEG on recruited cells. In a normal lysolecithin
demyelination model, the inflammatory response is quickly activated leading
to invasion of T cells, neutrophils and monocytes within the first 6-12 hours
(Kume et al., 1992; Ousman and David, 2000). Though many of the
invading cells remain present only transiently, activated macrophages will
continue to clear degraded myelin from the target area up to 4 weeks post-
lesion (Gilson and Blakemore, 1993; Pavelko et al., 1998). It is possible that
EEG are chemotactic for these inflammatory cells, may induce other CNS
cells to increase production of such chemotactic factors, or may prolong the
initial period of monocyte recruitment and activation.
No studies to date have focused on the ability of OEG to produce factors that
may be chemotactic for macrophages, a mechanism that appears to be of
critical importance given the suggestion by some researchers that a robust
macrophage response may be associated with efficient remyelination and
clearance of the lesion site (Graca and Blakemore, 1986; Perry et al., 1987;
David et al., 1990; George and Griffin, 1994; Morell et al., 1998; Rapalino et
al., 1998). A number of molecules such as tumour necrosis factor (TNF)-α,
interleukin (IL)-1, and IL-6 have been shown to increase recruitment and
Chapter 4 – Extended Discussion
126
activation of monocytes and microglia in the spinal cord (Giulian et al., 1989;
Schnell et al., 1999b; Klusman and Schwab, 1997; Smith et al., 1998). Other
candidate molecules that have been implicated in chemoattraction of
monocytes or in mediation of myelin phagocytosis include IP-10 (Luster and
Ravetch, 1987), MAC-2 (Reichert et al., 1994), monocyte chemoattractant
protein (MCP)-1α (Toews et al., 1998), IL-8, IL-10, growth-related oncogene
(GRO)-α, macrophage inflammatory protein (MIP)-1α, and granulocyte
macrophage-colony stimulating factor (GM-CSF) (Bartholdi and Schwab,
1997; Von Zahn et al, 1997; McTigue et al., 1998; Smith et al., 1998;
Ousman and David, 2001; Ma et al, 2002). Further studies will be necessary
to examine the ability of OEG either to directly express these or other unique
molecules, or to induce their expression at the site of OEG transplantation.
Furthermore, extracellular molecules such as VCAM-1, ICAM-1,
fibronectin, laminin and collagen type I have been associated with increased
adhesion of monocytes and/or increased functional activity of macrophages
and microglia (Newman and Tucci, 1990; Chamak and Mallat, 1991; Carlos
et al., 1991; Ley, 1996). Though several studies have thus far indicated that
AEG are capable of depositing large quantities of laminin and fibronectin in
vitro (Ramon-Cueto and Nieto-Sampedro, 1992; Sonigra et al., 1999; Au and
Roskams, 2003) and in vivo (Ramer et al., 2004a, b), the expression profiles
of other extracellular molecules after transplantation of OEG into lesioned
spinal cord remains unknown. To date, no comparison has been made
Chapter 4 – Extended Discussion
127
between EEG and older OEG preparations regarding the production of these
molecules, nor have any studies broadened their scope to include the other
molecules mentioned above.
Another possibility that has yet to be addressed is the effect of OEG
preparations on the integrity of the blood-brain barrier and its subsequent
effect on infiltration of endogenous cells into the lesion site (Andersson et al.,
1992; Riva-Depaty et al., 1994), or the simple fact that increased myelin
clearance may be attributable at least in part to direct phagocytosis by
recruited Schwann cells (Holtzman and Novikoff, 1965; Stoll et al., 1989;
Reichert et al., 1994; Fernandez-Valle et al., 1995; Liu et al., 1995b; Hirata et
al., 1999). Direct phagocytosis by OEG is another factor that must be
considered, given their ability to internalise degraded axonal material in vitro
(Wewetzer et al., 2005) and in injury models of the olfactory bulb in vivo
(Chuah et al., 1995; Susuki et al., 1996; Li et al., 2005). Though none of
these possibilities have been addressed in our study, they remain open for
investigation in future studies of OEG transplantation in the spinal cord.
Finally, the effect of OEG transplantation on the expression of molecules
directly associated with myelin destruction and phagocytosis during nerve
degeneration remains completely unknown. Apolipoprotein E (ApoE) for
example, has been implicated with recycling of myelin lipids for axonal
regeneration (reviewed by Vance et al., 2000). ApoE is produced by
Chapter 4 – Extended Discussion
128
macrophages and fibroblasts in response to nerve injury (Snipes et al., 1986;
Boyles et al., 1989; Saada et al., 1995), at which time Schwann cells
proximate to damaged neurites upregulate expression of ApoE low density
lipoprotein receptors (Rothe and Muller, 1991; Vance et al., 2000).
Unfortunately, no information is currently available on the expression or
induction of ApoE by OEG, though such studies are urgently needed. For
example, ApoE production by olfactory fibroblasts would go a long way
towards explaining why Li et al., (1997, 1998, 2003b) and other groups
(Appendix A) insist that unpurified (ie. fibroblast containing) populations of
OEG have a greater potential to promote regrowth and repair of lesioned
spinal cord than purified preparations. Conversely, a detailed analysis of low
density lipoprotein receptor expression by OEG would provide valuable
clues as to the underlying mechanisms by which these cells promote
improved clearance of myelin.
In any case, it stands to reason that the ability of OEG to clear degraded
myelin profiles quickly and efficiently may be one of the central aspects
behind their ability to promote axonal regeneration in the lesioned spinal
cord. It is well known that in the PNS regeneration can only proceed after
injured tissue components have been cleared via Wallerian degeneration
(Hirata and Kawabuchi, 2002). Furthermore, several studies have
demonstrated that macrophages can alter the nonpermissive adult CNS to a
state that permits axonal growth after injury (David et al., 1990; Lazarov-
Chapter 4 – Extended Discussion
129
Spiegler et al., 1996, 1998; Prewitt et al., 1997; Rapalino et al., 1998), a
situation reflected by the lack of secondary damage during macrophage
recruitment in lysolecithin demyelination models (Hall, 1972; Gregson,
1989; Jeffery and Blakemore, 1995). Future studies exploring this possibility
are encouraged, with special attention paid to the differences of the CNS
macrophage response upon transplantation of OEG derived from various
stages of development. However, any such future studies must pay special
attention to the type of injury used, as macrophage responses in the CNS
have been reported to vary depending on the type and location of injury
(Hirata et al., 1999; Schnell et al., 1999a).
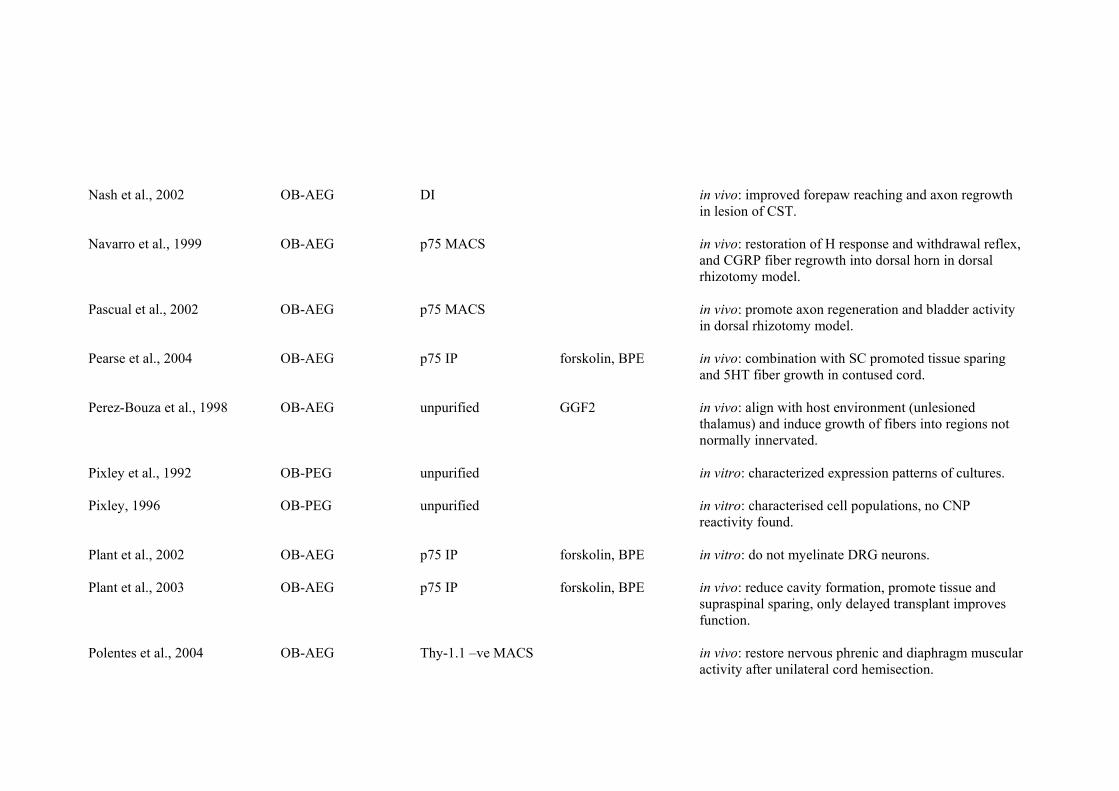
Influence of preparation age on promotion of axon growth
Unfortunately, no clear picture has emerged from the published literature
relating the age from which OEG were extracted and the extent of
functional/axonal regeneration observed. One reason for this lies with the
paucity of published studies that have investigated the use of
purified/unpurified EEG transplants into the lesioned CNS (Smale et al.,
1996; Boyd et al., 2004a). Furthermore, no clear interpretation of the
literature can be made between studies utilising PEG and AEG. This is due
to the large number of other variables present between the various studies
undertaken to date, including type and extent of lesion, location of lesion,
method of purification and expansion of the transplanted cells, time after
Chapter 4 – Extended Discussion
130
injury at which the transplant was carried out, and parameters utilised to
determine functional and anatomical recovery.
For example, several studies have utilised dorsal funiculus transection or
crush injury to analyse the effects of OEG transplantation (Li et al., 1997,
1998, 2003a; Imaizumi et al., 2000b; Nash et al., 2002; Shen et al., 2002;
Keyvan-Fouladi et al., 2003; Andrews and Stelzner, 2004; Polentes et al.,
2004; Sasaki et al., 2004, 2006; Ramer et al., 2004a; Richter et al., 2005;
Ruitenberg et al., 2005a). Of these, none have utilised EEG transplantation
and only three have utilised PEG (Imaizumi et al., 2000b; Ramer et al.,
2004a; Richter et al., 2005), the remainder using AEG (Li et al., 1997, 1998,
2003a; Nash et al., 2002; Shen et al., 2002; Keyvan-Fouladi et al., 2003;
Andrews and Stelzner, 2004; Polentes et al., 2004; Sasaki et al., 2004, 2006;
Ruitenberg et al., 2005a). Two of the studies utilising PEG transplantation
used cells derived from the olfactory bulb of rats (Imaizumi et al., 2000b;
Richter et al., 2005), whereas the third study utilised mouse PEG derived
from the lamina propria (Ramer et al., 2004a). Finally, the three studies on
PEG utilised different purification and/or expansion techniques. Imaizumi et
al., (2000b) performed a purification method similar to that used by Chuah
and Au (1993), where PEG are treated with cytosine arabinoside to remove
some contaminating cells, immunoadsorbed with antiserum to Thy-1.1 to
remove fibroblasts, and expanded rapidly by the addition of bovine pituitary
extract to the culture medium. Ramer et al., (2004a) utilised PEG, purified
Chapter 4 – Extended Discussion
131
by Thy-1.1 complement mediated lysis and expanded without added
mitogens in the culture medium. Richter et al., (2005) utilised PEG purified
by p75 immunopanning and expansion of cells in medium without mitogens
added. The type and location of lesion performed in either study was also
different, with Imaizumi et al., (2000b) performing transverse cuts along the
dorsal aspect of the lumbar spinal cord, whereas Ramer et al., (2004a) and
Richter et al., (2005) performed a dorsolateral funiculus crush of the cervical
spinal cord. Finally, the means by which anatomical and functional recovery
are assessed also varied greatly between each different study. The large
number of variables is also reflected in studies utilising transplantation of
AEG into the damaged dorsal funiculus, making interpretation of the
literature superficial at best.
The only model that can be compared and contrasted effectively is the
transplantation of OEG of different ages into the lesioned dorsal roots of the
spinal cord. Several studies have investigated the ability of OEG to restore
functional and anatomical connections in this lesion system (Ramon-Cueto
and Nieto-Sampedro, 1994; Navarro et al., 1999; Taylor et al., 2001; Pascual
et al., 2002; Gomez et al., 2003; Li et al., 2004; Ramer et al., 2004b; Riddell
et al., 2004). Suspensions of AEG were utilised in some of these studies
(Ramon-Cueto and Nieto-Sampedro, 1994; Navarro et al., 1999; Taylor et
al., 2001; Pascual et al., 2002; Gomez et al., 2003; Li et al., 2004) whereas
PEG were used in others (Ramer et al., 2004b; Riddell et al., 2004). The
Chapter 4 – Extended Discussion
132
results of these studies are summarised in Table 3. Briefly, restoration of
function and regrowth of ascending sensory axons past the dorsal root entry
zone and into the spinal cord was only observed in groups transplanted with
p75-selected AEG (Ramon-Cueto and Nieto-Sampedro, 1994; Navarro et al.,
1999; Taylor et al., 2001; Pascual et al., 2002) and unpurified AEG (Li et al.,
2004), but not AEG that were purified by removal of fastly adhering cells
and negative selection of fibroblasts (Gomez et al., 2003). Groups that did
not utilize AEG but used PEG instead (Ramer et al., 2004b; Riddell et al.,
2004) did not observe any significant regrowth of axons into the spinal cord
or restoration of function.
Gomez et al., (2003) suggested some of the differences observed between
these studies could be accounted for by variations in the extent of rhizomy in
nearby roots that could have allowed spared axons to sprout into the lesioned
area. This explanation is feasible, explaining why regrowth of fibers is
observed in studies utilising rhizotomy of 1-3 roots (Ramon-Cueto and
Nieto-Sampedro, 1994; Navarro et al, 1999; Pascual et al., 2002; Li et al.,
2004), but fails to account for the observed regrowth in studies where 7
dorsal roots were transected (Taylor et al., 2001), or the lack of regrowth in
other studies where only one dorsal root was severed (Riddell et al., 2004).
Another possibility not addressed by the authors is that the age animal from
which the cells are derived, or the purification techniques utilised, may be an
Chapter 4 – Extended Discussion
133
important factor accounting for the differences observed between these
studies. It is noteworthy that functional and anatomical regeneration is
observed only in studies utilising p75-selected AEG, or unpurified AEG
(Table 3). Groups utilising PEG (Ramer et al., 2004b; Riddell et al., 2004)
have failed to observe significant functional or anatomical recovery, as has
one study utilising AEG purified by negative immunoselection of fibroblasts
(Gomez et al., 2003). Two of the studies reporting negative results utlised
methods to eliminate contaminating fibroblasts from their cultures (Gomez et
al., 2003; Ramer et al., 2004b) whereas the third study utlised selection for
the O4 antigen as the primary means of purification (Riddell et al., 2004).
Selection for the O4 antigen may select for a subpopulation of OEG that do
not possess the full regenerative potential of p75-selected OEG (Kumar et al.,
2005). Our laboratory has also recently demonstrated that cultured
preparations of olfactory bulb cells possess different subpopulations of
fibroblasts, and that not all of these populations may express Thy-1.1 (Sophie
Callender, unpublished observations). This is of particular importance as any
contaminating cells in the preparation may proliferate once in contact with
damaged spinal cord tissue (Woodhouse et al., 2005) and become an
unknown factor during the repair stage in vivo. These observations highlight
the importance of future studies in defining and standardising the basic
methodology surrounding the extraction and purification of cells for
transplantation, an aspect that has often been neglected in OEG studies to
date.
Chapter 4 – Extended Discussion
134
Table 3. Studies utilising transplantation of OEG into transected spinal cord dorsal roots.
Study Cells used Purification Observations Ramon-Cueto and OB-AEG p75 IP Regeneration and ingrowth of Nieto-Sampedro, 1994 ascending sensory fibers into
contralateral dorsal horn Navarro et al., 1999 OB-AEG p75 MACS Restoration of spinal reflex
arcs. Regenerating ascending axons crossed DREZ
Taylor et al., 2001 OB-AEG p75 IP restoration of biceps reflex
activity and sensory input Pascual et al., 2002 OB-AEG p75 MACS restoration of sensory stimuli
implying regrowth of sensory axons through DREZ
Li et al., 2004 OB-AEG unpurified regrowth through DREZ, into
grey matter of dorsal horn and ascending dorsal columns
Gomez et al., 2003 OB-AEG DI, Thy1.1 No significant increase in
–ve MACS sensory afferent regrowth Riddell et al., 2004 OB-PEG O4 FACS No increase in ascending
fiber ingrowth. No detectable post-synaptic activity
Ramer et al., 2004b LP-PEG Thy1.1 CML No increase in sensory
afferent regrowth. Key: OB-xEG = OEG derived from the olfactory bulb. LP-PEG = PEG derived from the olfactory lamina propria. IP = immunopanning, MACS = magnetic activate cell sorting. FACS = fluorescence activated cell sorting. CML = complement mediated lysis. DREZ = dorsal root entry zone.
Despite these interesting correlations in published studies in vivo, there is no
evidence to suggest that age of preparation is a crucial factor influencing the
growth promoting (as opposed to myelinating) abilities of OEG in vitro.
Promotion of axonal growth by OEG has been reported in embryonic (Denis-
Donini and Estenoz, 1988; Kafitz and Greer; 1998, 1999), postnatal (Chuah
and Au, 1994; Le Roux and Reh, 1994; Tisay and Key, 1999; Van Den Pol
Chapter 4 – Extended Discussion
135
and Santarelli, 2003), young P21 (Moreno-Flores et al., 2003) and adult
preparations (Ramon-Cueto et al., 1993; Sonigra et al., 1999; Gudino-
Cabrera and Nieto-Sampedro, 2000; Gomez et al., 2003; Lipson et al., 2003;
Agrawal et al., 2004; Kumar et al., 2005; Leaver et al., 2006) (Table 4).
Furthermore, a comparative study by Goodman et al., (1993) has found that
selected immortalised cell lines of both PEG and AEG are able to promote
growth of embryonic chick retinal ganglion cells similarly, and concluded
that OEG appear to retain their ability to promote growth throughout
development. It would however be interesting to undertake a comparative
study between primary cultures of AEG, PEG and EEG.
Three studies to date have reported that diffusible factors released by OEG
are able to promote neurite growth in vitro (Le Roux and Reh, 1994; Kafitz
and Greer, 1998, 1999; Chung et al., 2004). One group reported a strong
growth promoting effect by unpurified EEG conditioned medium (Kafitz and
Greer, 1998, 1999), whereas only a weak growth promoting effect was
observed by groups utilising unpurified PEG (Le Roux and Reh, 1994;
Chung et al., 2004). One further study has reported that PEG are capable of
releasing axonal growth-promoting neurotrophic factors in vivo (Chuah et al.,
2004). Meanwhile, studies investigating AEG populations in vitro (Sonigra
et al., 1999; Lipson et al., 2003; Leaver et al., 2006) or PEG cultures
eliminated from fibroblasts (Chuah and Au, 1994) have failed to detect an
effect of OEG diffusible factors on neurite growth. To date, no study has
Chapter 4 – Extended Discussion
136
contrasted the effect of preparation age on the extent of neurite growth
promotion, nor has a detailed study been performed contrasting age of
preparation on release of diffusible factors by OEG.
Table 4. Studies investigating the promotion of neuron growth by primary OEG cultures.
OEG are cultured in DMEM supplemented with 10% FCS in all studies presented.
Additional factors present in the culture medium are listed where appropriate.
Study Cells used Purification Culture Growth Agrawal et al., 2004 OB-AEG unpurified Yes Kumar et al., 2005 OB-AEG p75 IP forskolin Yes ONR-AEG and GGF2 Gomez et al., 2003 OB-AEG DI, Thy1.1 Yes
–ve MACS Gudino-Cabrera and OB-AEG unpurified Yes Nieto-Sampedro, 2000 Ramon-Cueto et al., 1993 OB-AEG unpurified Yes Leaver et al., 2006 OB-AEG p75 IP forskolin Yes. Contact BPE mediated only. Lipson et al., 2003 OB-AEG DI Yes. Contact mediated only Sonigra et al., 1999 OB-AEG unpurified Yes. Contact mediated only Moreno-Flores et al., 2003 OB-P21 unpurified forskolin Yes BPE Chuah et al., 2004 (in vivo) OB-PEG AraC BPE Yes. Diffusible
factors. Chung et al., 2004 OB-PEG and AraC BPE Yes. Diffusible OM-PEG factors Chuah and Au, 1994 OB-PEG AraC and BPE Yes. Contact
Thy1.1 IA mediated only Le Roux and Reh, 1994 OB-PEG unpurified Yes. Diffusible factors
Chapter 4 – Extended Discussion
137
Van Den Pol and OB-PEG unpurified Yes Santarelli, 2003 Denis-Donini and EEG unpurified Yes Estenoz, 1988 Kafitz and Greer, 1998, 1999 EEG unpurified progesterone Yes. Diffusible corticosterone factors Key: AraC = cytosine arabinoside. OB-xEG = OEG derived from the olfactory bulb. ONR-AEG = AEG derived from the olfactory nerve rootlet on the intracranial side of cribiform plate. OM-PEG = PEG derived from the olfactory mucosa. GGF2 = glial growth factor 2. BPE = bovine pituitary extract. DI = differential attachment to remove macrophages and microglia. IA = immunoabsorption with antibody. IP = immunopanning. MACS = magnetic cell sorting.
Importance of Neuroglial Arrangement
Finally, another interesting aspect that remains open for investigation in this
field is the effect of neuroglial arrangement on the regeneration of axons
within the spinal cord. Several studies have indicated that OEG may require
specific alignment with respect to elongating fibres in order to promote
regrowth. Williams et al., (2004) demonstrated this quite clearly in a ZnSO4
irrigation lesion within the nasal cavity. The results of that study
demonstrated for the first time that OEG maintain their cytoarchitecture and
retain open channels through which regenerating axons from the olfactory
epithelium subsequently regrow. Their findings were recently supported by
Li et al., (2005), who found that OEG channels in which axons were
previously located were maintained even after severance of directed
projections from the cribriform plate to the olfactory bulb. This
groundbreaking in vivo work by these two groups in turn confirm previous
observations that orientation of OEG with respect to the growing axons may
Chapter 4 – Extended Discussion
138
be a critical factor in neuronal guidance (Sonigra et al., 1999; Van Den Pol
and Santarelli, 2003). Though significant progress has been made to date
utilising suspensions of isolated cells transplanted into lesions of the CNS,
this recent work brings to light another possible means of improving
functional outcomes, provided a means can be found to reproduce these
specific neuroglial arrangements and open channels prior to transplantation
of tissue bridges across the damaged area.
Furthermore, questions arise as to how such organised OEG structures
respond when the age of the preparation is varied. Is the reason why we
observe clear channels of EEG in our lysolecithin demyelinated animals due
to use of cells that are still more responsive to axonal cues and thus able to
rearrange themselves easily into growth promoting clusters/channels around
the axons of the spinal cord? Can AEG be induced to form such
arrangements in cultured matrices prior to transplantation into the cord, and if
so do these arrangements encourage directed regrowth of fibres as the work
of Williams et al., (2004) and Li et al., (2005) suggests they would? Taking
a step backwards, what would we have observed in our demyelinated model
had we also transected the cord? Would the transplanted EEG in our model
maintain their neuroglial arrangements following spinal transection and
subsequent degeneration of axons similarly to OEG located in the olfactory
system? Further studies into transplantation of olfactory nerve layer or
lamina propria into the lesioned spinal cord would provide a logical
Chapter 4 – Extended Discussion
139
framework for further studies, utilising olfactory tissues that have been
previously cleared of axons but that still retain an AEG glial arrangement that
may be conducive to growth. Another simpler approach has already been
initiated by De Mello and colleagues utilising time-lapse video microscopy to
answer the basic question of whether OEG from different age preparations
align themselves with growing axons in vitro, or whether growing axons
merely follow OEG paths already set prior to axonal contact.
Future Directions
Future studies analysing the regenerative potential of OEG derived from
animals of different ages will have to be performed with careful
consideration of all of the above. Several well designed studies have already
compared the benefits of acute vs delayed transplantation of AEG into the
spinal cord (Plant et al., 2003; Lopez-Vales et al., 2006), the benefits of
injecting the OEG directly into the lesion site or at points located more
distally to the injury (Andrews and Stelzner, 2004; Ramer et al., 2004b;
Richter et al., 2005), and the benefits of utilising PEG derived from the
olfactory bulb compared to PEG derived from the lamina propria of the
olfactory mucosa (Richter et al., 2005). Whereas all of these studies have
been controlled sufficiently to allow comparative analyses of the treatments
in question, there are still many factors that will need to be systematically
addressed by future reserch:
Chapter 4 – Extended Discussion
140
1) Does the age animal from which the OEG preparation is derived have a
significant effect on tissue sparing, cavity formation, axonal regeneration and
sprouting of spinal cord axons in a partial transection model, a contusion
model, a dorsal rhizotomy model and/or in a complete transection model?
2) Do culture conditions prior to transplantation affect the observed
functional/anatomical effects of the transplanted cells?
3) Does the location of the olfactory pathway (olfactory mucosa vs olfactory
bulb) from which the cells are derived have an effect on regeneration or
sparing in the aforementioned models? How are these responses affected by
the age of preparation?
4) Does the neuroglial arrangement of the transplanted cells affect the observed
functional/anatomical recovery?
5) Does acute vs delayed transplantation affect regenerative responses? How
does the site of injection affect the observed responses?
Concluding Remarks
We have demonstrated here in two studies that OEG populations are cells
possessing remarkable plasticity. The manner in which these cells respond
both in vitro and in vivo appears to be primarily dependant upon the age of
the animal from which they were extracted. Further investigation of the
literature seems to indicate that these age-dependant variations in behaviour
are not restricted to myelination potential and integration within the
Chapter 4 – Extended Discussion
141
demyelinated spinal cord, but that they are potentially a large source of
variation between results by different laboratories in other models of CNS
injury. We have also demonstrated in Chapter 2 that the conditions in which
the cells are grown following extraction can influence their responsiveness to
growth factors in vitro, and that these effects are persistent for a minimum of
six days after withdrawal of the impingent culture conditions. Every
laboratory working with OEG utilises their own individual means of
expanding these cells post-extraction. In this manner, the same cell type
extracted with similar protocols in two different laboratories may in fact also
behave significantly differently in subsequent experiments.
This wide variation in different aspects of in vitro and in vivo experiments
employing OEG should be discussed in appropriate forums if we are to
reduce variability of results in this field and increase cross applicability of
collected results. Another problem that remains unaddressed by our two
studies presented here is the issue of OEG purification techniques employed
in different laboratories. Though more studies have recently begun to
address these issues (Kumar et al., 2005), much work remains to be done to
contrast the effects of different purification techniques on the regenerative
potential of OEG. In the past 15 years giant strides forward have been made
into the study of OEG biology, but much remains to be done before clinical
transplantation of these cells can be performed with support of the existing
body of knowledge fully behind it.
APPENDIX A
Appendix A
143
Included in this appendix is a table summarising the various methods utilised
to culture OEG. Understanding the impact of variations such as source of the
cells, age of animal from which they are extracted, purification methods, and
mitogenic factors added to the tissue culture medium during the expansion
phases in vitro are vital to understanding the impact of each of these studies in
the field of OEG biology. Included in this table are all studies located by the
author that have utilised OEG derived from rodent tissue. Not included here
are studies utilising immortalised cell lines, cells derived from sources other
than the rodent, and studies that have utilised biopsies of olfactory tissue.
This table provides a quick reference guide for comparison of results obtained
from different laboratories utilising transplanted or cultured OEG.
Appendix A
144
Abbreviation Key Cells used If multiple cell types are listed, the authors have utilised multiple preparations as part of their study.
OE-xEG = OEG derived from the olfactory neuroepithelium ONR-xEG = OEG derived from olfactory nerve rootlets OM-xEG = OEG derived from the olfactory mucosa LP-xEG = OEG derived from the lamina propria of the olfactory
mucosa Purification If multiple purification methods are listed then the authors have compared use of the various treatments. Where used, the term 'and' indicates that those purification methods were used sequentially in the preparation.
AraC = treatment with cytosine arabinoside CML = complement mediated lysis DI = purification by differential attachment rates of
macrophages and fibroblasts FACS = Fluorescence activated cell sorting IA = immunoadsorption IP = immunopanning MACS = magnetic cell sorting NT-3 = purification was performed by the addition of
neurotrophin-3 to the culture medium BDNF = purification was performed by the addition of BDNF
to the culture medium. ST = removal of contaminants by a short period of
trypsinisation Culture conditions Culture medium in all cases is comprised of DMEM or DMEM/F12 (50:50 v/v) supplemented with 5%-10% FCS. Listed here are additional growth factors added to the culture medium to which the author would like to draw special attention. Factors such as L-Glutamine, glucose, gentamycin, etc, are not included here.
Observations A brief summary of the major findings of the study, with emphasis on cultured and transplanted results. 5HT = serotonergic axons bFGF = basic fibroblast growth factor
TH = tyrosine hydroxilase X-EB = X irradiation and Ethidium Bromide
Study Cells used Purification Culture Observations Agrawal et al., 2004 OB-AEG unpurified in vivo: improvement in SLA increase TH +ve fiber
density in 6-OHDA lesion of nigrostriatal pathway. In vitro: improved growth of TH +ve neurons.
Alexander et al., 2002 OB-PEG O4 FACS in vitro: investigated long-term proliferation effects of
different combinations of mitogens to 22 days. Andrews and OB-AEG DI in vivo: promotion of growth in bilateral crush of cord is Stelzner, 2004 additive with promotion by sciatic nerve conditioning
lesion. Au and Roskams, 2002 OM-PEG Thy-1.1 CML in vitro: describe culturing technique for mouse PEG. Au and Roskams, 2003 LP-PEG Thy-1.1 CML in vitro: characterised expression profile differs to OB-
xEG. bFGF is mitogenic but not in long-term. Barakat et al., 2005 OB-AEG p75 IP forskolin, BPE in vivo: poor survival, promotion of growth, and
restoration of function in contused cord. Barber and Lindsay, 1982 OB-AEG, OB-PEG unpurified in vitro: characterise expression and morphology of OM-AEG, OM-PEG cultures from bulb and mucosa, from adult and neonate. Barnett et al., 1993 OB-PEG O4 FACS in vitro: describe culturing technique and cell
characteristics.
Bianco et al., 2004 LP-AEG unpurified in vitro: NT-3 promotes proliferation of GFAP +ve NT-3 olfactory cells. Expression profile analysis performed. BDNF
Boyd et al., 2004a OB-EEG unpurified in vivo: clip compression injury, do not myelinate spinal cord. Boyd et al., 2006 OB-EEG unpurified proteomic: EEG but not SC express calponin. In vivo: do not associate with axons in compressed cord. Byrnes et al., 2005 OB-AEG DI in vitro: low power laser irradiation changes expression
profile over 21 day period. Cao et al., 2004 OB-AEG p75 IP forskolin, BPE in vivo: promoted sprouting, growth past lesion and
function improvement in complete transection. Retroviral increase GDNF expression promotes growth.
Choi and Raisman, 2005 OB-AEG unpurified and ST forskolin, BPE in vivo: improved eye closure, but not motoneuron
regrowth in severed facial nerve. Chuah and Au, 1993 OB-PEG AraC and BPE in vitro: describe culturing technique and cell Thy-1.1 IA characteristics. Chuah and Au, 1994 OB-PEG AraC and BPE in vitro: promote growth of ORN
Thy1.1 IA (contact mediated only). Chuah and Teague, 1999 OB-PEG AraC and DI BPE in vitro: report bFGF proliferation DRC. Chuah et al., 2000 OB-PEG AraC BPE in vitro: GGF2 is mitogenic and promotes ECM
deposition. Chuah et al., 2004 OB-PEG AraC BPE in vivo: encapsulated cells into dorsal transection.
Promote regrowth when PEG both in and out of capsules.
Chung et al., 2004 OB-PEG and AraC BPE in vitro: promote growth of CN (mediated by contact OM-PEG and diffusible factors) Collazos-Castro et al., 2005 OB-AEG DI and in vivo: no regrowth of CST axons and no improvement -ve Thy-1.1 MACS of function in contused cord. Deni-Donini and OB-EEG unpurified in vitro: promote growth of dopaminergic neurons from Estenoz, 1988 substantia nigra. Devon and Doucette, 1992 OB-EEG unpurified in vitro: myelinate DRG neurons. Devon and Doucette, 1995 OB-EEG unpurified in vitro: myelinate DRG neurons without ascorbate. Doucette, 1993b OB-EEG unpurified in vitro: describe phenotypic features and effect of ACM
or cAMP analogues. Doucette and Devon, 1994 OB-PEG unpurified in vitro: attempt to promote expression of myelinating
phenotype (neuron-free cultures). MBP not expressed under any condition.
Doucette and Devon, 1995 OB-EEG unpurified in vitro: attempt to promote expression of myelinating
phenotype (neuron-free cultures). MBP not expressed under any condition.
Dunning et al., 2004 OB-PEG AraC and ST forskolin, in vivo: tracked cells via MRI. Report remyelination of and p75 IP heregulin X-EB cord. Fairless et al., 2005 OB-PEG O4 FACS FGF2, ACM, in vitro: move easily over astrocyte cultures, SC do not. forskolin, BPE N-cadherin important for SC but not PEG movement.
Fouad et al., 2005 OB-AEG p75 IP forskolin, BPE in vivo: combined therapy with SC and chondroitinase in complete cord transection, increased functional outcomes and growth of 5HT fibers.
Franceschini and OB-PEG O4 FACS ACM in vivo: p75 and E-N-CAM define two populations of Barnett, 1996 OEG in bulb. In vitro: Majority of cells p75-ve but gain
p75 over time in culture. Garcia-Alias et al., 2004 OB-AEG p75 MACS in vivo: improve tissue preservation, restoration of
behavioral skills and physiological outcome in photochemical lesion of cord.
Gomez et al., 2003 OB-AEG DI and Thy1.1 in vivo: no regrowth in dorsal rhizotomy model.
–ve MACS in vitro: promote growth of DRG neurons. Gudino-Cabrera and OB-AEG p75 MACS in vivo: tranplanted cells migrate long distances in Nieto-Samepedro, 1996 Thy-1.1 –ve MACS unlesioned hippocampus. In vitro: viability of cells
maintained after freezing. Gudino-Cabrera and OB-AEG unpurified in vitro: report tanycytes and pituicytes express similar Nieto-Sampedro, 2000 markers to AEG. All three glial types promote growth
of DRG neurons. Guntinas-Lichius et al., 2001 OB-PEG AraC forskolin in vivo: increased number of motoneurons but no
improvement of whisking behaviour in facial nerve axotomy model.
Hayat et al., 2003a OB-AEG unpurified forskolin, in vitro: investigated intracellular Ca2+ handling in cells
GGF2 resting or in contact with RGC. Blocking Ca2+ influx decreases axon growth.
Hayat et al., 2003b OB-AEG unpurified forskolin, in vitro: G proteins regulate calcium signalling GGF2 and thus RGC neurite growth.
Imaizumi et al., 1998 OB-PEG unpurified in vivo: remyelinate X-EB cord, improve conduction
velocity. Imaizumi et al., 2000b OB-PEG AraC and BPE in vivo: regrow and remyelinate transected dorsal Thy-1.1 IA column, improve conduction velocity. Jani and Raisman, 2004 OB-AEG unpurified in vitro: investigated proliferative rates of cells to 21 OM-AEG days. p75+ve cells from OM maintained high
proliferation rates. Johansson et al., 2005 OB-AEG DI in vivo: combined with mesocenphalic embryonic tissue
in 6-OHDA lesion of nigrostriatal pathway. Improved function, survival and regrowth of dopaminergic fibers.
Kafitz and Greer, 1998 OB-EEG unpurified progesterone, in vitro: promote growth of ORN (mediated by contact corticosterone and diffusible factors) Kafitz and Greer, 1999 EEG unpurified progesterone, in vitro: promote growth of ORN (mediated by contact corticosterone and diffusible factors) Keyvan-Fouladi et al., 2003 OB-AEG unpurified in vivo: improved forepaw reaching, angiogenesis and
CST axon regrowth in electrolytic lesion of dorsal CST. Kumar et al., 2005 OB-AEG p75 IP forskolin, in vitro: p75 selected AEG better at promoting RGC
ONR-AEG p75 –ve IP GGF2 growth, no difference in regrowth by p75+ve OB-AEG vs ONR-AEG.
Lakatos et al., 2000 OB-PEG O4 FACS in vitro: PEG but not SC intermix well with astrocytes. SC but not PEG induce CSPG expression in astrocytes.
Lakatos et al., 2003a OB-PEG O4 FACS forskolin, in vitro: lesser astrocytic response than SC in unlesioned heregulin spinal cord. Lakatos et al., 2003b OB-PEG • unpurified forskolin, in vivo: remyelinate X-EB cord, myelination improves if • AraC and ST heregulin meningeal cells included.
and p75 IP Leaver et al., 2006 OB-AEG p75 IP forskolin, BPE in vitro: promote RGC growth (contact mediated only) Lee et al., 2004 OB-AEG unpurified in vivo: tracked cells via MRI, no function improvement
after complete transection of cord. Le Roux and Reh, 1994 OB-PEG unpurified in vitro: promote growth of CN (mediated by contact and diffusible factors) Li et al., 1997 OB-AEG unpurified in vivo: improve forepaw reaching, myelinate and
regrow axons after focal CST lesion. Li et al., 1998 OB-AEG unpurified in vivo: myelinate and regrow axons after focal CST
lesion, highly angiogenic. Li et al., 2003a OB-AEG unpurified in vivo: improve supraspinal control, breathing and
climbing in dorsal hemisection of cord. Li et al., 2003b OB-AEG unpurified in vivo: axons regrow through transected optic nerve. Li et al., 2004 OB-AEG unpurified in vivo: axons regrow into dorsal horn and columns in
dorsal rhizotomy model.
Lipson et al., 2003 OB-AEG DI in vitro: promote growth of embryonic sympathetic
neurons and Remak's ganglia (contact mediated only). Neurotrophic factor expression analysed in vitro and in vivo.
Liu et al., 1995a OB-PEG AraC and BPE in vivo: migrate towards bulb when injected into Thy-1.1 IA olfactory epithelium. In vitro: migrate towards olfactory
bulb due to soluble factors released by the bulb. Liu et al., 2005 OM-AEG DI in vitro: characterisation of primary cultures
(neurotrophic factors and mitosis) Lopez-Vales et al., 2004 OB-AEG p75 MACS in vivo: improve functional outcomes and MEP, and
increase angiogenesis in photochemically injured cord. Lopez-Vales et al., 2006 OB-PEG p75 MACS in vivo: acute vs delayed transplants in completely
transected cord. Both improved MEP and regrowth of 5HT and NOR fibers, acute transplants better than delayed at improving locomotor function.
Lu et al., 2001 LP-AEG unpurified in vivo: promote growth 5HT fibers, improve
electrophysiological outcome and functional recovery in complete transection.
Moreno-Flores et al., 2003 OB-P21 unpurified BPE, forskolin in vitro: primary cultures express ErbB2, ErbB3 and
ErbB4 protein, and expression increases when medium contains BPE. Promote growth of RGC neurons.
Nash et al., 2001 OB-AEG DI in vitro: describe culturing technique.
Nash et al., 2002 OB-AEG DI in vivo: improved forepaw reaching and axon regrowth in lesion of CST.
Navarro et al., 1999 OB-AEG p75 MACS in vivo: restoration of H response and withdrawal reflex,
and CGRP fiber regrowth into dorsal horn in dorsal rhizotomy model.
Pascual et al., 2002 OB-AEG p75 MACS in vivo: promote axon regeneration and bladder activity
in dorsal rhizotomy model. Pearse et al., 2004 OB-AEG p75 IP forskolin, BPE in vivo: combination with SC promoted tissue sparing
and 5HT fiber growth in contused cord. Perez-Bouza et al., 1998 OB-AEG unpurified GGF2 in vivo: align with host environment (unlesioned thalamus) and induce growth of fibers into regions not normally innervated. Pixley et al., 1992 OB-PEG unpurified in vitro: characterized expression patterns of cultures. Pixley, 1996 OB-PEG unpurified in vitro: characterised cell populations, no CNP
reactivity found. Plant et al., 2002 OB-AEG p75 IP forskolin, BPE in vitro: do not myelinate DRG neurons. Plant et al., 2003 OB-AEG p75 IP forskolin, BPE in vivo: reduce cavity formation, promote tissue and
supraspinal sparing, only delayed transplant improves function.
Polentes et al., 2004 OB-AEG Thy-1.1 –ve MACS in vivo: restore nervous phrenic and diaphragm muscular
activity after unilateral cord hemisection.
Pollock et al., 1999 OB-PEG O4 FACS in vitro: neuregulin is strong mitogen and survival factor. Express ErbB2 and ErbB4 but not ErbB3.
Ramer et al., 2004a LP-PEG Thy-1.1 CML in vivo: decrease cavity and scar formation, increased
growth of 5HT and TH +ve axons, increased angiogenesis and increased recruitment of SC in dorsolateral crush of cord.
Ramer et al., 2004b LP-PEG Thy1.1 CML . in vivo: no increased sensory afferent ingrowth in
dorsal rhizotomy model. Ramon-Cueto and OB-AEG unpurified and ST in vitro: characterisation of primary cultures. Nieto-Sampedro, 1992 Ramon-Cueto and OB-AEG p75 IP in vivo: ingrowth of ascending sensory fibers into Nieto-Sampedro, 1994 contralateral dorsal horn in dorsal rhizotomy model. Ramon-Cueto et al., 1993 OB-AEG unpurified in vitro: characterisation of primary cultures, promote
growth of ORN.
Ramon-Cueto et al., 1998 OB-AEG p75 IP forskolin, BPE in vivo: combined therapy with SC in completely transected cord, CGRP and 5HT axons regrow.
Ramon-Cueto et al., 2000 OB-AEG p75 IP forskolin, BPE in vivo: improve function and proprioceptive response,
and induce regrowth of NOR and 5HT axons in completely transected cord.
Resnick et al., 2003 OB-AEG p75 IP forskolin, BPE in vivo: no restoration of function in contusion injury.
Richter et al., 2005 LP-PEG Thy-1.1 CML in vivo: increase angiogenesis, decrease cavity formation OB-PEG p75 IP (LP-PEG>OB-PEG), increase sprouting (LP>OB) and
autonomy (LP only) in dorsolateral funiculus crush. Riddell et al., 2004 OB-PEG O4 FACS FGF2, heregulin, in vivo: no increase in ascending fiber ingrowth, and no forskolin, ACM detectable post-synaptic activity in dorsal rhizotomy
model. Ruitenberg et al., 2002 OB-AEG p75 IP forskolin, BPE in vivo: expression induced by viral vectors stable after
implantation into spinal cord dorsolateral RST lesion. Ruitenberg et al., 2003 OB-AEG p75 IP forskolin, BPE in vitro: promote growth DRG axons (contact only) in vivo: adenoviral production of BDNF and NT-3
increases RST sprouting and function in unilateral RST transection.
Ruitenberg et al., 2005a OB-AEG p75 IP forskolin, BPE in vivo: promote tissue sparing but no regrowth of axons
past lesion in dorsal hemisection. Virally modified NT-3 producing AEG promote long distance axon regrowth.
Sasaki et al., 2004 OB-AEG DI in vivo: improve hindlimb locomotion, remyelinate
transected dorsal funiculus. Sasaki et al., 2006 OB-AEG DI in vivo: decreased neuronal loss, improved locomotor
activity and increased BDNF levels in transected dorsal funiculus.
Santos-Silva and OB-AEG unpurified in vitro: express CNP but not MBP. Cavalcante, 2001
Shen et al., 2002 OB-AEG unpurified in vivo: improve function, MEP, and growth of MBP +ve fibers in completely transected cord.
Smale et al., 1996 OB-EEG unpurified in vivo: cholinergic axons grow into grafts in fimbria-
fornix lesion. Smith et al., 2001 OB-PEG O4 FACS in vivo: remyelinate X-EB cord. Sonigra et al., 1999 OB-AEG unpurified in vitro: promote RGC growth (contact mediated only) Takami et al., 2002 OB-AEG p75 IP forskolin, BPE in vivo: no significant regrowth of axons or
improvement of function in contused cord. Promote tissue sparing.
Taylor et al., 2001 OB-AEG p75 IP in vivo: restoration of biceps reflex activity and sensory
input in dorsal rhizotomy model. Thompson et al., 2000 OB-PEG O4 FACS in vitro: express ErbB4 but not ErbB3 mRNA. Express
mRNA for NRG-1 but do not release it.
Tisay and Key, 1999 OE-PEG unpurified in vitro: promote ORN growth which is potentiated by laminin.
Van Den Pol and OB-PEG unpurified in vitro: promote growth of granule cell neurons in Santarelli, 2003 parallel to long axis of PEG. SC avoid astrocytes
Verdu et al., 2001 OB-AEG p75 MACS in vivo: reduced gliosis and cystic cavitation in
photochemical lesion of cord.
Verdu et al., 2003 OB-AEG p75 MACS in vivo: prevent loss of parenchyma, no improvement of function, increased nociceptive w/drawal, and increased MEP and SSEP in photochemical lesion of cord.
Vincent et al., 2003 OB-PEG AraC BPE in vitro: morphology changes can be controlled by
cAMP and endothelin-1. Neurotrophin profile unchanged by culture conditions.
Vincent et al., 2005 OB-PEG AraC BPE in vitro: PEG are more closely related to SC than to
astrocytes according to microarray analysis.
Wang et al., 2005 OB-AEG DI in vitro: describe culturing technique and cell characteristics.
Wewetzer et al., 2001 OB-PEG AraC forskolin in vitro: express CNTF mRNA and its receptor.
Forskolin increases CNTF mRNA and decreases receptor mRNA. CNTF not mitogenic.
Wewetzer et al., 2005 OB-PEG O4 MACS in vitro: O4 reactivity due to axonal fragments on p75 MACS surface of cell. PEG phagocytose these fragments in
vivo and in vitro. p75 expression increases as O4 decreases.
Woodhall et al., 2001 OB-PEG AraC BPE in vitro: express GDNF, BDNF and NGF proteins,
secrete BDNF and NGF, express TrkB mRNA. Woodhall et al., 2003 OB-PEG AraC BPE in vivo: report mRNA changes following
implantation into dorsal transection of cord
Woodhouse et al., 2005 OB-PEG AraC BPE in vitro: cultured with stabbed or contused cord. Contaminant proliferation high in presence of cord tissue. High apoptosis of PEG with acutely injured cord.
Xia et al., 2005 OB-PEG AraC and DI BPE in vitro: galanin inhibits proliferation, report mRNA
expression of galanin receptors. Yan et al., 2001a OB-AEG p75 IP in vitro: proliferation DRC to heregulin, FGF2, PDGF-
BB and IGF-1. All are mitogenic in presence of serum, only heregulin and FGF2 in absence of serum.
Yan et al., 2001b OB-AEG p75 IP in vitro: hepatocyte growth factor promotes dose-
Yan et al., 2003 OB-PEG p75 IP in vitro: proliferation DRC to LPA, tested in
combination with other factors. LPA promotes migration/proliferation.
References
159
REFERENCES Adlkofer K, Lai C (1999) Role of neuregulins in glial development. Glia 29:104-111. Aguayo AJ, Charron L, Bray GM (1976) Potential of Schwann cells from unmyelinated nerves to produce myelin: a quantitative ultrastructural and autoradiographic study. J Neurocytol 5:565-573. Agrawal HC, Sprinkle TJ, Agrawal D (1990) 2’-3’ cyclic nucleotide-3’-phosphodiesterase in peripheral nerve myelin is phosphorylated by a phorbol ester-sensitive protein kinase. Biochem Biophys Res Comm 170:817-823. Agrawal AK, Shukla S, Chaturvedi RK, Seth K, Srivastava N, Ahmad A, Seth PK (2004) Olfactory ensheathing cell transplantation restores functional deficits in rat model of Parkinson's disease: a cotransplantation approach with fetal ventral mesencephalic cells. Neurobiol Dis 16:516-526. Alexander CL, Fitzgerald UF, Barnett SC (2002) Identification of growth factors that promote long-term proliferation of olfactory ensheathing cells and modulate their antigenic phenotype. Glia 37:349-364. Altman J, Das GD (1965) Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J Comp Neurol 124:319-335. Anderson DJ (1993) Cell and molecular biology of neural crest cell lineage diversification. Curr Opin Neurobiol 3:8-13. Andersson PB, Perry VH, Gordon S (1992) The acute inflammatory response to lipopolysaccharide in CNS parenchyma differs from that in other body tissues. Neurosci 48:169-186. Anderton BH, Breinburg D, Downes MJ, Green PJ, Tomlinson BE, Ulrich J, Wood JN, Kahn J (1982) Monoclonal antibodies show that neurofibrillary tangles and neurofilaments share antigenic determinants. Nature 298:84-86. Andrews MR, Stelzner DJ (2004) Modification of the regenerative response of dorsal column axons by olfactory ensheathing cells or peripheral axotomy in adult rat. Exp Neurol 190:311-327. Au E, Roskams AJ (2002) Culturing olfactory ensheathing glia from the mouse olfactory epithelium. Methods Mol Biol 198:49-54. Au E, Roskams AJ (2003) Olfactory ensheathing cells of the lamina propria in vivo and in vitro. Glia 41:224-236.
References
160
Au WW, Treloar HB, Greer CA (2002) Sublaminar organisation of the mouse olfactory bulb nerve layer. J Comp Neurol 446:68-80. Bamber NI, Li H, Lu X, Oudega M, Aebischer P, Xu XM (2001) Neurotrophins BDNF and NT-3 promote axonal re-entry into the distal host spinal cord through Schwann cell-seeded min-channels. Eur J Neurosci 13:257-268. Barber PC (1981) Regeneration of vomeronasal nerves into the main olfactory bulb in the mouse. Brain Res 216:239-251. Barber PC (1982) Neurogenesis and regeneration in the primary olfactory pathway of mammals. Bibl Anat 23:12-25. Barber PC, Lindsay RM (1982) Schwann cells of the olfactory nerves contain glial fibrillary acidic protein and resemble astrocytes. Neurosci 7:3077-3090. Barber PC, Raisman G (1978) Replacement of receptor neurones after section of the vomeronasal nerves in the adult mouse. Brain Res 147:297-313. Barnett SC, Roskams AJ (2002) Olfactory ensheathing cells. Isolation and culture from the rat olfactory bulb. Methods Mol Biol 198:41-48. Barnett SC, Alexander CL, Iwashita Y, Gilson JM, Crowther J, Clark L, Dunn LT, Papanastassiou V, Kennedy PGE, Franklin RJM (2000) Identification of a human olfactory ensheathing cell that can effect transplant-mediated remyelination of demyelinated CNS axons. Brain 123:1581-1588. Barnett SC, Hutchins A-M, Noble M (1993) Purification of olfactory nerve ensheathing cells from the olfactory bulb. Dev Biol 155:337-350. Barakat DJ, Gaglani SM, Neravetla SR, Sanchez AR, Andrade CM, Pressman Y, Puzis R, Garg MS, Bunge MB, Pearse DD (2005) Survival, integration, and axon growth support of glia transplanted into the chronically contused spinal cord. Cell Transplant 14:225-240. Bartholdi D, Schwab ME (1997) Expression of pro-inflammatory cytokine and chemokine mRNA upon experimental spinal cord injury in mouse: an in situ hybridization study. Eur J Neurosci 9:1422-1438. Baulieu E-E, Schumacher M (2000) Progesterone as a neuroactive neurosteroid, with special reference to the effect of progesterone on myelination. Steroids 65:605-612.
References
161
Beattie M, Bresnahan JC, Komon J, Tovar CA, Van Meter M, Anderson DK, Faden AI, Hsu CY, Noble LJ, Salzman S, Young W (1997) Endogenous repair after spinal cord contusion injuries in the rat. Exp Neurol 148:453-463. Ben-Baruch N, Yarden Y (1994) Neu differentiation factors: a family of alternatively spliced neuronal and mesenchymal factors. Proc Soc Exp Biol Med 208:221-227. Benfey M, Aguayo AJ (1982) Extensive elongation of axons from rat brain into peripheral nerve grafts 296:150-152. Bianco JI, Perry C, Harkin DG, Mackay-Sim A, Feron F (2004) Neurotrophin 3 promotes purification and proliferation of olfactory ensheathing cells from human nose. Glia 45:111-123. Bixby JL, Lilien J, Reichardt LF (1988) Identification of the major proteins that promote neuronal process outgrowth on Schwann cells in vitro. J Cell Biol 107:353-361. Blakemore WF (1977) Remyelination in the spinal cord of the cat following intraspinal injections of lysolecithin. J Neurol Sci 33:31-43. Blanes T (1898) Sobre algunos puntos dudosos de la estructura del bulbo olfatorio. Rev Trim Microg 3:99-127. Blesch A, Tuszynski MH (2003) Cellular GDNF delivery promotes growth of motor and dorsal column sensory axons after partial and complete spinal cord transections and induces remyelination. J Comp Neurol 467:403-417. Boruch AV, Conners JJ, Pipitone M, Deadwyler G, Storer PD, Devries GH, Jones KJ (2001) Neurotrophic and migratory properties of an olfactory ensheathing cell line. Glia 33:225-229. Bovolenta P, Wandosell F, Nieto-Sampedro M (1992) CNS glial scar tissue: a source of molecules which inhibit central neurite outgrowth. Prog Brain Res 94:367-379. Boyd JG, Lee J, Skihar V, Doucette R, Kawaja MD (2004a) LacZ-expressing olfactory ensheathing cells do not associate with myelinated axons after implantation into the compressed spinal cord. PNAS 101:2162-2166. Boyd JG, Doucette R, Kawaja MD (2004b) Defining the role of olfactory ensheathing cells in facilitating axon remyelination following damage to the spinal cord. FASEB Journal 19:694-703.
References
162
Boyd JG, Jahed A, McDonald TG, Krol KM, Van Evk JE, Doucette R, Kawaja MD (2006) Proteomic evaluation reveals that olfactory ensheathing cells but not Schwann cells express calponin. Glia 53:434-440. Boyles JK, Zoellner CD, Anderson LJ, Kosik LM, Pitas RE, Wesgraber KH, Hull DY, Mahley RW, Gebicke-Haerter PJ, Ignatius MJ, Shooter EM (1989) A role for apolipoprotein E, apolipoprotein A-I, and low density lipoprotein receptors in cholesterol transport during regeneration and remyelination of the rat sciatic nerve. J Clin Invest 83:1015-31. Bradbury EJ, Moon LD, Popat RJ, King VR, Bennet GS, Patel PN, Fawcett JW, McMahon SB (2002) Chondroitinase ABC promotes functional recovery after spinal cord injury. Nature 416:636-640. Braun PE, Sandillon F, Edwards A, Matthieu JM, Privat A (1988) Immunocytochemical localization by electron microscopy of 2'3'-cyclic nucleotide 3'-phosphodiesterase in developing oligodendrocytes of normal and mutant brain. J Neurosci 8:3057-3066. Bregman BS, Coumans JV, Dai HN, Kuhn PL, Lynskey J, mcAtee M, Sandhu F (2002) Transplants and neurotrophic factors increase regeneration and recovery of function after spinal cord injury. Prog Brain Res 137:257-273. Bregman BS, Kunkel-Bagden E, Schnell L, Dai HN, Gao D, Schwab ME (1995) Recovery from spinal cord injury mediated by antibodies to neurite growth inhibitors. Nature 378:498-501. Bregman BS, McAtee M, Dai HN, Kuhn PL (1997) Neurotrophic factors increase axonal growth after spinal cord injury and transplantation in the adult rat. Exp Neurol 148:475-494. Britsch S, Li L, Kirchhoff S, Theuring F, Brinkmann V, Birchmeier C, Riethmacher D (1998) The ErbB2 and ErbB3 receptors and their ligand, neuregulin-1, are essential for development of the sympathetic nervous system. Genes Dev 12:1825-1836. Brockes JP, Lemke GE, Balzer DR Jr (1980a) Purification and preliminary characterization of a glial growth factor from the bovine pituitary. J Biol Chem 255:8374-8377. Brockes JP, Raff MC, Nishiguchi DJ, Winter J (1980b) Studies on cultured rat Schwann cells. III. Assays for peripheral myelin proteins. J Neurocytol 9:67-77.
References
163
Brosamle C, Huber AB, Fiedler M, Skerra A, Schwab ME (2000) Regeneration of lesioned corticospinal tract fibers in the adult rat induced by a recombinant, humanized IN-1 antibody fragment. J Neurosci 20:8061-8068. Brook GA, Plate D, Franzen R, Martin D, Moonen G, Schoenen J, Schmitt AB, Noth J, Nacimiento W (1998) Spontaneous longitudinally orientated axonal regeneration is associated with the Schwann cell framework within the lesion site following spinal cord compression injury of the rat. J Neurosci Res 53:51-65. Bunge MB, Williams AK, Wood PM (1982) Neuron-Schwann cell interaction in basal lamina formation. Dev Biol 92:449-460. Burden SJ (1998) The formation of neuromuscular synapses. Genes Dev 12:133-148. Burden S, Yarden Y (1997) Neuregulins and their receptors: a versatile signaling molecule in organogenesis and oncogenesis. Neuron 18:847-855. Busfield SJ, Michnick DA, Chickering TW, Revett TL, Ma J, Woolf EA, Comrack CA, Dussault BJ, Woolf J, Goodearl ADJ, Gearing DP (1997) Characterization of a neuregulin-related gene, Don-1, that is highly expressed in restricted regions of the cerebellum and hippocampus. Mol Cell Biol 17:4007-4014. Byrnes KR, Wu X, Waynant RW, Ilev IK, Anders JJ (2005) Low power laser irradiation alters gene expression of olfactory ensheathing cells in vitro. Lasers Surg Med 37:161-171. Cai D, Shen Y, De Bellard M, Tang S, Filbin MT (1999) Prior exposure to neurotrophins blocks inhibition of axonal regenration by MAG and myelin via a CAMP-dependent mechanism. Neuron 22:89-101. Calof AL, Chikaraishi DM (1989) Analysis of neurogenesis in a mammalian neuroepithelium: proliferation and differentiation of an olfactory neuron precursor in vitro. Neuron 3:115-127. Cao L, Liu L, Chen Z-Y, Wang L-M, Ye J-L, Qiu H-Y, Lu C-L, He C (2004) Olfactory ensheathing cells genetically modified to secrete GDNF to promote spinal cord repair. Brain 127:535-549. Carlos T, Kovach N, Schwartz B, Rosa M, Newman B, Wayne E, Benjamin C, Osborn L, Lobb R, Harlan J (1991) Human monocyte bind to two cytokine-induced adhesion ligands on cultured human endothelial cells: Endothelial-leukocyte adhesion molecule-1 and vascular cell adhesion molecule-1. Blood 77:2266-2271.
References
164
Carpenter G (2003) Nuclear localization and possible functions of receptor tyrosine kinases. Curr Opin Cell Biol 15:143-148. Carr VM, Farbman AI (1992) Ablation of the olfactory bulb up-regulates the rate of neurogenesis and induces precocious cell death in olfactory epithelium. Exp Neurol 115:55-59. Carraway KL III, Burden SJ (1995) Neuregulins and their receptors. Curr Opin Neurobiol 5:606-612. Carraway KL III, Cantley LC (1994) A neu acquaintance for erbB3 and erbB4: a role for receptor heterodimerization in growth signalling. Cell 78:5-8. Carraway KL III, Sliwkowski MX, Akita R, Platko JV, Guy PM, Nuijens A, Diamonti AJ, Vandlen RL, Cantley LC, Cerione RA (1994) The erbB3 gene product is a receptor for heregulin. J Biol Chem 269:14303-14306. Carraway KL III, Weber JL, Unger MJ, Ledesma J, Yu N, Gassmann M, Lai C (1997) Neuregulin-2, a new ligand of ErbB3/ErbB4-receptor tyrosine kinases. Nature 387:512-516. Carroll SL, Miller ML, Frohnert PW, Kim SS, Corbett JA (1997) Expression of neuregulins and their putative receptors, ErbB2 and ErbB3, is induced during Wallerian degeneration. J Neurosci 17:1642-1659. Chamak B, Mallat M (1991) Fibronectin and laminin regulate the in vitro differentiation of microglial cells. Neurosci 45:513-527. Chang H, Riese DJ 2nd, Gilbert W, Stern DF, McMahan UJ (1997) Ligands for ErbB-family receoptors encoded by a neuregulin-like gene. Nature 387:509-512. Chau CH, Shum DK, Li H, Pei J, Lui YY, Wirthlin L, Chan YS, Xu XM (2004) Chondroitinase ABC enhances axonal regrowth through Schwann cell-seeded guidance channels after spinal cord injury. FASEB J 19:194-196. Chen A, Xu XM, Kleitman N, Bunge MB (1996) Methylprednisolone administration improves axonal regeneration into Schwann cell grafts in transected adult rat thoracic spinal cord. Exp Neurol 138:261-276. Cheng H, Cao Y, Olson L (1996) Spinal cord repair in adult paraplegic rats: partial restoration of hind limb function. Science 273:510-513.
References
165
Choi D, Raisman G (2005) Disorganization of the facial nucleus after nerve lesioning and regeneration in the rat: effects of transplanting candidate reparative cells to the site of injury. Neurosurg 56:1093-1100. Chu J, Hatton J, Su H (1999) Effects of epidermal growth factor and dibutyryl cyclic adenosine monophosphate on the migration pattern of astrocytes grafted into adult rat brain. Neurosurg 45:859-866. Chuah MI, Au C (1991) Olfactory Schwann cells are derived from precursor cells in the olfactory epithelium. J Neurosci Res 29:172-180. Chuah MI, Au C (1993) Cultures of ensheathing cells from neonatal rat olfactory bulbs. Brain Res 601:213-220. Chuah MI, Au C (1994) Olfactory cell cultures on ensheathing cell monolayers. Chem Senses 19:25-34. Chuah MI, Teague R (1999) Basic fibroblast growth factor in the primary olfactory pathway: mitogenic effect on ensheathing cells. Neurosci 88:1043-1050. Chuah MI, Tennent R, Jacobs I (1995) Response of olfactory Schwann cells to intranasal zinc sulfate irrigation. J Neurosci Res 42:470-478. Chuah MI, Cossins J-M, Woodhall E, Tennent R, Nash G, West AK (2000) Glial growth factor 2 induces proliferation and structural changes in ensheathing cells. Brain Res 857:265-274. Chuah MI, Choi-Lundberg D, Weston S, Vincent AJ, Chung RS, Vickers JC, West AK (2004) Olfactory ensheathing cells promote collateral axonal branching in the injured adult rat spinal cord. Exp Neurol 185:15-25. Chung RS, Woodhouse A, Fung S, Dickson TC, West AK, Vickers JC, Chuah MI (2004) Olfactory ensheathing cells promote neurite sprouting of injured axons in vitro by direct cellular contact and secretion of soluble factors. CMLS 61:1238-1245. Collazos-Castro JE, Muneton-Gomez VC, Nieto-Sampedro M (2005) Olfactory glia transplantation into cervical spinal cord contusion injuries. J Neurosurg Spine 3:308-317. Corfas G, Rosen KM, Aratake H, Krauss R, Fischbach GD (1995) Differential expression of ARIA isoforms in the rat brain. Neuron 14:103-115. Costanzo RM, Graziadei PP (1983) A quantitative analysis of changes in the olfactory epithelium following bulbectomy in hamster. 215:370-381.
References
166
Coumans JV, Lin TT, Dai HN, MacArthur L, McAtee M, Nash C, Bregman BS (2001) Axonal regeneration and functional recovery after complete spinal cord transection in rats by delayed treatment with transplants and neurotrophins. J Neurosci 21:9334-9344. Crovello CS, Lai C, Cantley LC, Carraway KL III (1998) Differential signaling by the epidermal growth factor-like growth factors neuregulin-1 and neuregulin-2. J Biol Chem 273:26954-26961. Cummings BJ, Uchida N, Tamaki SJ, Salazar DL, Hooshmand M, Summers R, Gage FH, Anderson AJ (2005) Human neural stem cells differentiate and promote locomotor recovery in spinal cord-injured mice. Proc Natl Acad Sci USA 102:14069-14074. David S, Aguayo AJ (1981) Axonal elongation into peripheral nervous system "bridges" after central nervous system injury in adult rats. Science 214:931-933. David S, Bouchard C, Tsatas O, Giftochristos N (1990) Macrophages can modify the nonpermissive nature of the adult mammalian central nervous system. Neuron 5:463-469. De Mello TR, Dunlop S, Plant GW (2003) Myelination Potential of Olfactory Ensheathing Glia. Society for Neuroscience Meeting, New Orleans, USA. De Mello TR, Busfield S, Dunlop S, Plant GW (2005) Proliferative effects of neuregulins on olfactory ensheathing glia. Australian Neuroscience Society, Perth, Western Australia. Denis-Donini S, Estenoz M (1988) Interneurons versus efferent neurons: heterogeneity in their neurite outgrowth response to glia from several brain regions. Dev Biol 130:237-249. Dergham P, Ellezam B, Essagian C, Avedissian H, Lubell WD, McKerracher L (2002) Rhosignalling pathway targeted to promote spinal cord repair. J Neurosci 22:6570-6577. Devon R, Doucette R (1992) Olfactory ensheathing cells myelinate dorsal root ganglion neurites. Brain Res 589:175-179. Devon R, Doucette R (1995) Olfactory ensheathing cells do not require L-ascorbic acid in vitro to assemble a basal lamina or to myelinate dorsal root ganglion neurites. Brain Res 688:223-229.
References
167
Dong Z, Brennan A, Liu N, Yarden Y, Lefkowitz G, Mirsky R, Jessen KR (1995) neu differentiation factor is a neuron-glia signal and regulates survival, proliferation and maturation of rat Schwann cell precursors. Neuron 15:585-596. Dong Z, Dean C, Walters JE, Mirsky R, Jessen KR (1997) Response of Schwann cells to mitogens in vitro is determined by pre-exposure to serum, time in vitro and developmental stage. Glia 20:219-230. Doucette JR (1984) The glial cells in the nerve fiber layer of the rat olfactory bulb. Anat Rec 210:385-391. Doucette R (1989) Development of the nerve fiber layer in the olfactory bulb of mouse embryos. J Comp Neurol 285:514-527. Doucette R (1990) Glial influences on axonal growth in the primary olfactory system. Glia 3:433-449. Doucette R (1991) PNS-CNS transitional zone of the first cranial nerve. J Comp Neurol 312:451-466. Doucette R (1993a) Glial cells in the nerve fiber layer of the main olfactory bulb of embryonic and adult animals. Microsc Res Tech 24:113-130. Doucette R (1993b) Glial progenitor cells of the nerve fiber layer of the olfactory bulb: effect of astrocyte growth media. J Neurosci Res 35:274-287. Doucette R (1995) Olfactory ensheathing cells: potential for glial cell transplantation into areas of CNS injury. Histol Histopathol 10:503-507. Doucette R, Devon R (1994) Media that support the growth and differentiation of oligodendrocytes do not induce olfactory ensheathing cells to express a myelinating phenotype. Glia 10:296-310. Doucette R, Devon R (1995) Elevated intracellular levels of cAMP induce olfactory ensheathing cells to express Gal-C and GFAP but not MBP. Glia 13:130-140. Dunning MD, Lakatos A, Loizou L, Kettunen M, ffrench-Constant C, Brindle KM, Franklin RJM (2004) Superparamagnetic iron oxide-labeled Schwann cells and olfactory ensheathing cells can be traced in vivo by magnetic resonance imaging and retain functional properties after transplantation into the CNS. J Neurosci 24:9799-9810.
References
168
Eldridge CF, Bunge MB, Bunge RP, Wood PM (1987) Differentiation of axon-related Schwann cells in vitro. I. Ascorbic acid regulates basal lamina assembly and myelin formation. J Cell Biol 105:1023-1034. Erickson SL, O’Shea KS, Ghaboosin N, Loverro L, Frantz G, Bauer M, Lu LH, Moore MW (1997) ErbB3 is required for normal cerebellar and cardiac development: A comparison with erbB2 and heregulin deficient mice. Development 124:4999-5011. Falls DL, Harris DA, Johnson FA, Morgan MM, Corfas G, Fischbach GD (1990) Mr 42,000 ARIA: A protein that may regulate the accumulation of acetylcholine receptors at developing chick neuromuscular junctions. CSH Symp Quant Biol 55:397-406. Falls DL, Rosen KM, Corfas G, Lane WS, Fischbach GD (1993) ARIA, a protein that stimulates acetylcholine receptor synthesis, is a member of the neu ligand family. Cell 72:801-815. Farbman AI, Squinto LM (1985) Early development of olfactory receptor cell axons. Brain Res 351:205-213. Fairless R, Frame MC, Barnett SC (2005) n-Cadherin differentially determines Schwann cell and olfactory ensheathing cell adhesion and migration responses upon contact with astrocytes. Mol & Cell Neurosci 28:253-263. Fernandez-Valle C, Bunge RP, Bunge MB (1995) Schwann cells degrade myelin and proliferate in the absence of macrophages: evidence from in vitro studies of Wallerian degeneration. J Neurocytol 24:667-679. Feron F, Perry C, Cochrane J, Licina P, Nowitzke A, Urquhart S, Geraghty T, Mackay-Sim A (2005) Autologous olfactory ensheathing cell transplantation in human spinal cord injury. Brain 128:2951-2960. Field PM, Li Y, Raisman G (2003) Ensheathment of the olfactory nerves in the adult rat. J Neurocytol 32:317-324. Figlewicz DA, Quarles RH, Johnson D, Barbarash GR, Sternberger NH (1981) Biochemical demonstration of the myelin-associated glycoprotein in the peripheral nervous system. J Neurochem 37:749-758. Filbin MT, Walsh FS, Trapp BD, Pizzey JA, Tennekoon GI (1990) Role of myelin P0 protein as a homophilic adhesion molecule. Nature 344:871-872. Filbin MT (2003) Myelin-associated inhibitors of axonal regeneration in the adult mammalian CNS. Nat Rev Neurosci 4:703-713.
References
169
Fishman PS, Kelley JP (1984) The fate of severed corticospinal axons. Neurology 34:1161-1167. Fouad K, Schnell L, Bunge, MB, Schwab ME, Liebscher T, Pearse DD (2005) Combining Schwann cell bridges and olfactory-ensheathing glia grafts with chondroitinase promotes locomotor recovery after complete transection of the spinal cord. J Neurosci 25:1169-1178. Fradkin JE, Cook GH, Kilhoffer MC, Wolff J (1982) Forskolin stimulation of thyroid adenylate cyclase and cyclic 3’,5’-adenosine monophosphate accumulation. Endocrinology 111:849-856. Franceschini IA, Barnett, SC (1996) Low-Affinity NGF-receptor and E-N-CAM expression define two types of olfactory nerve ensheathing cells that share a common lineage. Dev Biol 173:327-343. Franklin RJM, Barnett SC (1991) The electron microscopic appearance of the β-galactosidase reaction product. Acta Neuropathol 81:686-687. Franklin RJ, Gilson JM, Franceschini IA, Barnett SC (1996) Schwann cell-like myelination following transplantation of an olfactory bulb-ensheathing cell line into areas of demyelination in the adult CNS. Glia 17:217-224. Fregien NL, White LA, Bunge MA, Wood PA (2004) Forskolin increases neuregulin receptors in human Schwann cells without increasing receptor mRNA. Glia 49:24-35. Friede RL, Samorjski T (1968) Myelin formation in the sciatic nerve of the rat: a quantitative electron microscopic, histochemical and radioautographic study. J Neuropathol Exp Neurol 27:546-570. Fruttiger M, Montag D, Schachner M, Martini R (1995) Crucial role for the myelin-associated glycoprotein in the maintenance of axon-myelin integrity. Eur J Neurosci 7:511-515. Gage FH (2000) Mammalian neural stem cells. Science 287:1433-1438. Garbay B, Heape AM, Sargueil F, Cassagne C (2000) Myelin synthesis in the peripheral nervous system. Prog Neurobiol 61:267-304. Garcia-Alias G, Lopez-Vales R, Fores J, Navarro X, Verdu E (2004) Acute transplantation of olfactory ensheathing cells or Schwnann cells promotes recovery after spinal cord injury in the rat. J Neurosci Res 75:632-641.
References
170
Garratt AN, Britsch S, Birchmeier C (2000) Neuregulin, a factor with many functions in the life of a Schwann cell. BioEssays 22:987-996. Gavazzi I, Kumar RDC, McMahon SB, Cohen J (1999) Growth responses of different subpopulations of adult sensory neurons to neurotrphic factors in vitro. Eur J Neurosci 11:3405-3414. George R, Griffin JW (1994) Delayed macrophage responses and myelin clearance during Wallerian degeneration in the central nervous system: the dorsal radiculotomy model. Exp Neurol 129:225-236. Ghoumari AM, Ibanez C, El-Etr M, Leclerc P, Eychenne B, O'Malley BW, Baulieu EE, Schumacher M (2003) Progesterone and its metabolites increase myelin basic protein expression in organotypic slice cultures of rat cerebellum. J Neurochem 86:848-859. Giese KP, Martini R, Lemke G, Soriano P, Schachner M (1992) Mouse P0 gene disruption leads to hypomyelination, abnormal expression of recognition molecules, and degeneration of myelin and axons. Cell 71:565-576. Gilson J, Blakemore WF (1993) Failure of remyelination in areas of demyelination produced in the spinal cord of old rats. Neurpathol Appl Neurobiol 19:173-181. Giulian D, Chen J, Ingeman JE, George JK, Noponen M (1989) The role of mononuclear phagocytes in wound healing after traumatic injury to adult mammalian brain. J Neurosci 8:4416-4429. Goldsmith HS, de la Torre JC (1992) Axonal regeneration after spinal cord transection and reconstruction. Brain Res 589:217-224. Gomez VM, Averill S, King V, Yang Q, Perez ED, Chacon SC, Ward R, Nieto-Sampedro M, Priestley J, Taylor J (2003) Transplantation of olfactory ensheathing cells fails to promote significant axonal regeneration from dorsal roots into the rat cervical cord. J Neurocytol 32:53-70. Goodman MN, Silver J, Jacobberger JW (1993) Establishment and neurite outgrowth properties of neonatal and adult rat olfactory bulb glial cell lines. Brain Res 619:199-213. Goodearl AD, Davis JB, Mistry K, Minghetti L, Otsu M, Waterfield MD, Stroobant P (1993) Purification of multiple forms of glial growth factor. J Biol. Chem 268:18095-18102. Gospodarowicz D, Cheng J, Lirette M (1983) Bovine brain and pituitary fibroblast growth factors: comparison of their abilities to support the
References
171
proliferation of human and bovine vascular endothelial cells. J Cell Biol 97:1677-1685. Gould E, Reeves AJ, Graziano MSA, Gross CG (1999) Neurogenesis in the neocortex of adult primates. Science 286:548-552. Graca DL, Blakemore WF (1986) Delayed remyelination in rat spinal cord following ethidium bromide injection. Neuropathol Appl Neurobiol 12:593-605. GrandPre T, Li S, Strittmatter SM (2002) Nogo-66 receptor antagonist peptide promotes axonal regeneration. Nature 417:547-551. Graziadei PPC (1973) Vertebrates olfactory mucosa. In: The ultrastructure of sensory organs (Friedman I ed), pp269-305. North-Holland Publishing Co: London. Graziadei PPC, Monti Graziadei GA (1978) Continuous nerve cell renewal in the olfactory system. In: Handbook of sensory physiology, Vol. 9. (Jacobson M, ed.), pp55-82. Springer Verlag: Berlin. Graziadei PP, Monti Graziadei GA (1979) Neurogenesis and neuron regeneration in the olfactory system of mammals. I. Morphological aspects of differentiation and structural organization of the olfactory sensory neurons. J Neurocytol 8:1-8. Graziadei PPC, Monti Graziadei GA (1980) Neurogenesis and neuron regeneration in the olfacotory system of mammals. III. Deafferentation and reinnervation of the olfactory bulb following section of the fila olfactoria in rat. J Neurocytol 9:145-162. Graziadei PPC, Levine RR, Monti Graziadei GA (1978) Regeneration of olfactory axons and synapse formation in the forebrain after bulbectomy in neonatal mice. Proc Natl Acad Sci USA 75:5230-5234. Greenfield S, Brostoff S, Eylar EH, Morell P (1973) Protein composition of myelin of the peripheral nervous system. J Neurochem 20:1207-1216. Gregson NA (1989) Lysolipids and membrane damage: lysolecithin and its interaction with myelin. Biochem Soc Trans 17:280-283. Grinspan JB, Marchionni MA, Reeves M, Coulaloglou M, Scherer SS (1996) Axonal interactions regulate Schwann cell apoptosis in developing peripheral nerve: neuregulin receptors and the role of neuregulins. J Neurosci 16:6107-6118.
References
172
Gudino-Cabrera G, Nieto-Sampedro M (1996) Ensheathing cells: large scale purification from adult olfactory bulb, freeze-preservation and migration of transplanted cells in adult brain. Rest Neurol Neurosci 10:25-34. Gudino-Cabrera G, Nieto-Sampedro M (2000) Schwann-like macroglia in adult rat brain. Glia 30:49-63. Guntinas-Lichius O, Angelov DN, Tomov TL, Dramiga J, Neiss WF, Wewetzer K (2001) Transplantation of olfactory ensheathing cells stimulates the collateral sprouting from axotomized adult rat facial motoneurons. Exp Neurol 172:70-80. Guntinas-Lichius O, Wewetzer K, Tomov TL, Azzolin N, Kazemi, S, Streppel M, Neiss WF, Angelov DN (2002) Transplantation of olfactory mucosa minimizes axonal branching and promotes the recovery of vibrissae motor performance after facial nerve repair in rats. J Neurosci 15:7121-7131. Guth L, Zhang Z, Roberts E (1994) Key role for pregnenolone in combination therapy that promotes recovery after spinal cord injury. Proc Natl Acad Sci USA 91:12308-12312. Guy PM, Platko JV, Cantley LC, Cerione RA, Carraway KL III (1994) Insect cell-expressed p180erbB3 possesses an impaired tyrosine kinase activity. Proc Natl Acad Sci USA 91:8132-8136. Hahn AF, Whitaker JN, Kachar B, Webster HdeF (1987) P2, P1 and P0 myelin protein expression in developing rat sixth nerve: a quantitative immunocytochemical study. J Comp Neurol 260:501-512. Hall SM (1972) The Effect of injections of lysophosphatidylcholine into white matter of the adult mouse spinal cord. J Cell Sci 10:535-546. Halper J, Parnell PG, Carter BJ, Ren P, Scheithauer BW (1992) Presence of growth factors in human pituitary. Lab Invest 66:639-645. Harauz G, Ishiyama N, Hill CMD, Bates IR, Libich DS, Fares C (2004) Myelin basic protein – diverse conformational states of an intrinsically unstructured protein and its roles in myelin assembly and multiple sclerosis. Micron 35:503-542. Harvey AR (1994) Expression of low affinity NGF (p75) receptors in rat superior colliculu: studies in vivo, in vitro, and in fetal tectal grafts. Exp Neurol 130:237-249.
References
173
Hayat S, Wigley CB, Robbins J (2003a) Intracellular calcium handling in rat olfactory ensheathing cells and its role in axonal regeneration. Neurosci 259-270. Hayat S, Thomas A, Afshar F, Sonigra R, Wigley CB (2003b) Manipulation of olfactory ensheathing cell signalling mechanisms: effects on their support for neurite regrowth from adult CNS neurons in culture. Glia 44:232-241. Herrera LP, Casas CE, Bates ML, Guest JD (2005) Ultrastructural study of the primary olfactory pathway in Macaca fascicularis. J Comp Neurol 488:427-441. Hirata K, Kawabuchi M (2002) Myelin phagocytosis by macrophages and nonmacrophages during Wallerian degeneration. Microsc Res Tech 57:541-547. Hirata K, Mitoma H, Ueno N, He J, Kawabuchi M (1999) Differential response of macrophage subpopulations to myelin degradation in the injured rat sciatic nerve. J Neurocytol 28:685-695. Ho WH, Armanini MP, Nuijens A, Phillips HS, Osheroff PL (1995) Sensory and motor neuron-derived factor. A novel heregulin variant highly expressed in sensory and motot neurons. J Biol Chem 270:14523-14532. Hoke A, Ho T, Crawford TO, LeBel C, Hilt D, Griffin JW (2003) Glial cell line-derived neurotrophic factor alters axon Schwann cell units and promotes myelination in unmyelinated nerve fibers. J Neurosci 23:561-567. Holmes WE, Sliwkowski MX, Akita RW, Henzel WJ, Lee J, Park JW, Yansura D, Abadi N, Raab H, Lewis GD, Shepard HM, Kuang W-J, Wood WI, Goeddel DV, Vandlen RL (1992) Identification of heregulin, a specific activation of p185erbB2. Science 256:1205-1210. Holtzman E, Novikoff AB (1965) Lysosomes in the rat sciatic nerve following crush. J Cell Biol 27:651-669. Honmou O, Felts PA, Waxman SG, Kocsis JD (1996) Restoration of normal conduction properties in demyelinated spinal cord axons in the adult rat by transplantation of exogenous Schwann cells. J Neurosci 16:3199-3208. Huang H, Chen L, Hongmei W, Bo X, Bingehen L, Rui W, Jian Z, Feng Z, Zheng C, Ying L, Yinglun S, Wei H, Shuyi P, Junzhao S (2003) Influence of patients age on functional recovery after transplantation of olfactory ensheathing cells into injured spinal cord injury. Chin Med J 116:1488-1491.
References
174
Hughes JT, Brownell B (1963) Aberrant nerve fibres within the spinal cord. J neurol Neurosurg Psychiat 26:528-534. Imaizumi T, Lankford KL, Wazman SG, Greer CA, Kocsis JD (1998) Transplanted olfactory ensheathing cells remyelinate and enhance axonal conduction in the demyelinated dorsal columns of rat spinal cord. J Neurosci 18:6176-6185. Imaizumi T, Lankford KL, Burton WV, Fodor WL, Kocsis JD (2000a) Xenotransplantation of transgenic pig olfactory ensheathing cells promotes axonal regenration in rat spinal cord. Nature Biotech 18:949-953. Imaizumi T, Lankford KL, Kocsis JD (2000b) Transplantation of olfactory ensheathing cells or Schwann cells restores rapid and secure conduction across the transected spinal cord. Brain Res 854:70-78. Jani HR, Raisman G (2004) Ensheathing cell cultures from the olfactory bulb and mucosa. Glia 47:130-137. Jeffery ND, Blakemore WF (1995) Remyelination of mouse spinal cord axon demyelinated by local injection of lysolecithin. J Neurocytol 24:775-781. Jessen KR, Mirsky R (1992) Schwann cells: early lineage, regulation of proliferation and control of myelin formation. Curr Opin Neurobiol 2:575-581. Johansson S, Lee I-H, Olson L, Spenger C (2005) Olfactory ensheathing glial co-grafts improve functional recovery in rats with 6-OHDA lesions. Brain 128:2961-2976. Johnson PW, Abramow-Newerly W, Seilheimer B, Sadoul R, Tropak MB, Arquint M, Dunn RJ, Schachner M, Roder JC (1989) Recombinant myelin-associated glycoprotein confers neural adhesion and neurite outgrowth function. Neuron 3:377-385. Jones JT, Akita RW, Sliwkowski MX (1999) Binding specificities and affinities of egf domains for ErbB receptors. FEBS Letters 447:227-231. Jones RB, Gordus A, Krall JA, MacBeath G (2006) A quantitative protein interaction network for the ErbB receptors using protein microarrays. Nature 439:168-174. Jung-Testas I, Schumacher M, Robel P, Baulieu EE (1994) Actions of steroid hormones on glial cells of the central and peripheral nervous system. J Steroid Biochem 48:145-154.
References
175
Kafitz KW, Greer CA (1998) The influence of ensheathing cells on olfactory receptor cell neurite outgrowth in vitro. Ann NY Acad Sci 855:266-269. Kafitz KW, Greer CA (1999) Olfactory ensheathing cells promote neurite extension from embryonic olfactory receptor cells in vitro. Glia 25:99-110. Kato T, Honmou O, Uede T, Hashi K, Kocsis JD (2000) Transplantation of human olfactory ensheathing cells elicits remyelination of demyelinated rat spinal cord. Glia 30:209-218. Katsuki M, Sato M, Kimura M, Yokoyama M, Kobayashi K, Nomura T (1988) Conversion of normal behavior of shiverer by myelin basic protein antisense cDNA in transgenic mice. Science 241:593-595. Keyvan-Fouladi N, Raisman G, Li Y (2003) Functional repair of the corticospinal tract by delayed transplantation of olfactory ensheathing cells in adult rats. J Neurosci 23:9428-9434. Kim HA, Rosenbaum T, Marchionni MA, Ratner N, DeClue JE (1995) Schwann cells from neurofibromin deficient mice exhibit activation of p21ras, inhibition of cell proliferation and morphological changes. Oncogene 11:325-335. Kim HA, DeClue JE, Ratner N (1997) cAMP-dependent protein kinase A is required for Schwann cell growth: interactions between the cAMP and neuregulin/tyrosine kinase pathways. J Neurosci Res 49:236-247. Kita Y, Mayer J, Zamborelli T, Hara S, Rohde M, Watson E, Koski R, Ratzkin B, Nicolson M (1995) Boactive synthetic peptide of NDF/Heregulin. Biochem Biophys Res Comm 210:441-451. Kleitman N, Wood PM, Bunge RP (1998) Tissue culture methods for the study of myelination. In: Culturing nerve cells (Banker G, Goslin K, ed), pp545-594. Cambridge: London MIT Press. Klusman I, Schwab ME (1997) Effects of pro-inflammatory cytokins in experimental spinal cord injury. Brain Res 762:173-184. Kobayashi NR, Fan DP, Giehl KM, Bedard AM, Wiegand SJ, Tetzlaff W (1997) BDNF and NT-4/5 prevent atrophy of rat rubrospinal neurons after cervical axotomy, stimulate GAP-43 and Talpha1-tubulin mRNA expression, and promote axonal regeneration. J Neurosci 17:9583-9595. Koenig HL, Schumacher M, Ferzaz B, Do Thi AN, Ressouches A, Guennoun R, Jung-Testas I, Robel P, Akwa Y, Baulieu E-E (1995) Progesterone synthesis and myelin formation by Schwann cells. Science 268:1500-1503.
References
176
Kraus MH, Issing W, Miki T, Popescu NC, Aaronson SA (1989) Isolation and characterization of ERBB3, a third member of the ERBB/ epidermal growth factor receptor family: Evidence for overexpression in a subset of human mammary tumors. Proc Natl Acad Sci USA 86:9193-9197. Kromer LF, Cornbrooks CJ (1985) Transplants of Schwann cell cultures promote axonal regeneration in the adult mammalian brain. Proc Natl Acad Sci USA 82:6330-6334. Kuhlengel KR, Bunge MB, Bunge RP, Burton H (1990) Implantation of cultured sensory neurons and Schwann cells into lesioned neonatal rat spinal cord. II. Implant characteristics and examination of corticospinal tract growth. J Comp Neurol 293:74-91. Kumar R, Hayat S, Felts P, Bunting S, Wigley C (2005) Functional differences and interactions between phenotypic subpopulations of olfactory ensheathing cells in promoting CNS axonal regeneration. Glia 50:12-20. Kume N, Cybulsky MI, Gimbrone MA Jr (1992) Lysophosphatidylcholine, a component of atherogenic lipoproteins, induces mononuclear leukocyte adhesion molecules in cultured human and rabbit arterial endothelial cells. J Clin Invest 90:1138-1144. Lakatos A, Franklin RJ, Barnett SC (2000) Olfactory ensheathing cells and Schwann cells differ in their in vitro interactions with astrocytes. Glia 32:214-25. Lakatos A, Barnett SC, Franklin JM (2003a) Olfactory ensheathing cells induce less host astrocyte response and chondroitin sulphate proteoglycan expression than Schwann cells following transplantation into adult CNS white matter. Exp Neurol 184:237-246. Lakatos A, Smith PM, Barnett SC, Franklin RJM (2003b) Meningeal cells enhance limited CNS remyelination by transplanted olfactory ensheathing cells. Brain 126:598-609. Lazarov-Spiegler O, Solomon AS, Zeev-Brann AB, Hirschberg DL, Lavie V, Schwartz M (1996) Transplantation of activated macrophages overcomes central nervous system regrowth failure. FASEB J 10:1296-12302. Lazarov-Spiegler O, Solomon AS, Schwartz M (1998) Peripheral nerve-stimulated macrophages simulate a peripheral nerve-like regenerative response in rat transected optic nerve. Glia 24:329-337.
References
177
Leaver SG, Harvey AR, Plant GW (2006) Adult olfactory ensheathing glia promote the long-distance growth of adult retinal ganglion cell neurites in vitro. Glia In press. Lee I-H, Bulte JWM, Schweinhardt P, Douglas T, Trifunovski A, Hofstetter C, Olson L, Spenger C (2004) In vivo magnetic resonance tracking of olfactory ensheathing glia grafted into the rat spinal cord. Exp Neurol 187:509-516. Lee MJ, Brennan A, Blanchard A, Zoidl G, Dong Z, Tabernero A, Zoidl C, Dent MAR, Jessen KR, Mirsky R (1997) P0 is constitutively expressed in the rat neural crest and embryonic nerves and is negatively and positively regulated by axons to generate non-myelin-forming and myelin-forming Schwann cells respectively. Mol Cell Neurosci 8:336-350. Lemke G (1988) Unwrapping the genes of myelin. Neuron 1:535-543. Lemke G (1996) Neuregulins in Development. Mol Cell Neurosci 7:247-262. Lemke G, Axel R (1985) Isolation and sequence of a cDNA encoding the major structural protein of peripheral myelin. Cell 40:501-508. Lemke GE, Brockes JP (1984) Identification and purification of glial growth factor. J Neurosci 4:75-83. Le Roux PD, Reh TA (1994) Regional differences in glial-derived factors that promote dendritic outgrowth from mouse cortical neurons in vitro. J Neurosci 14:4639-4655. Levi ADO, Bunge RP, Lofgren JA, Meima L, Hefti F, Nikolics K, Sliwkowski MX (1995) The influence of heregulins on human Schwann cell proliferation. J Neurosci 15:1329-1340. Ley K (1996) Molecular mechanisms of leukocyte recruitment in the inflammatory process. Cardiovasc Res 32:733-742. Li Y, Raisman G (1994) Schwann cells induce sprouting in motor and sensory axons in the adult rat spinal cord. J Neurosci 14:4050-4063. Li S, Strittmatter SM (2003) Delayed systemic Nogo-66 receptor antagonist promotes recovery from spinal cord injury. J Neurosci 23:4219-4227. Li Y, Field PM, Raisman G (1997) Repair of adult rat corticospinal tract by transplants of olfactory ensheathing cells. Science 277:2000-2002.
References
178
Li Y, Field PM, Raisman G (1998) Regeneration of adult rat corticospinal axons induced by transplanted olfactory ensheathing cells. J Neurosci 18:10514-10524. Li Y, Decerchi P, Raisman G (2003a) Transplantation of olfactory ensheathing cells into spinal cord lesions restores breathing and climbing. J Neurosci 23:727-731. Li Y, Sauve Y, Li D, Lund RD, Raisman G (2003b) Transplanted olfactory ensheathing cells promote regeneration of cut adult rat optic nerve axons. J Neurosci 27:7783-7788. Li Y, Carlstedt T, Berthold, C-H, Raisman G (2004) Interaction of transplanted olfactory-ensheathing cells and host astrocytic processes provides a bridge for axons to regenerate axons across the dorsal root entry zone. Exp Neurol 188:300-308. Li Y, Field PM, Raisman G (2005) Olfactory ensheathing cells and olfactory nerve fibroblasts maintain continuous open channels for regrowth of olfactory nerve fibres. Glia 52:245-251. Liesi P (1985a) Do neurons in the vertebrate CNS migrate on laminin? EMBO J 4:1163-1170. Liesi P (1985b) Laminin-immunoreactive glia distinguish regenerative adult CNS systems from non-regenerative ones. EMBO J 4:2505-2511. Lin WC, Sanchez HB, Deerlinck T, Morris JK, Ellisman M, Lee KF (2000) Aberrant development of motor axons and neuromuscular synapses in erbB2-deficient mice. Proc Natl Acad Sci USA 97:1299-1304. Lindsay RM (1988) Nerve growth factors (NGF, BDNF) enhance axonal regeneration but are not required for survival of adult sensory neurons. J Neurosci 8:2394-2405. Lipson AC, Widenfalk J, Lindqvist E, Ebendal T, Olson L (2003) Neurotrophic properties of olfactory ensheathing glia. Exp Neurol 180:167-171. Liu KL, Chuah MI, Lee KKH (1995a) Soluble factors from the olfactory bulb attract olfactory Schwann cells. J Neurosci 15:990-1000. Liu HM, Yan LH, Yang YJ (1995b) Schwann cell properties: 3. C-fos expression, bFGF production, phagocytosis and proliferation during Wallerian degeneration. J Neuropathol Exp Neurol 54:487-496.
References
179
Liu JB, Tang TS, Gong AH, Sheng WH, Yang JC (2005) The mitosis and immunocytochemistry of olfactory ensheathing cells from nasal olfactory mucosa. Clin J Traumatol 8:306-310. Longart M, Liu Y, Karavanova I, Buonanno A (2004) Neuregulin-2 is developmentally regulated and targeted to dendrites of central neurons. J Comp Neurol 472:156-172. Lopez-Vales R, Garcia-Alias G, Fores J, Navarro X, Verdu E (2004) Increased expression of cyco-oxygensase 2 and vascular endothelial growth factor in lesioned spinal cord by translanted olfactory ensheathing glia. J Neurotrauma 21:1031-1043. Lopez-Vales R, Fores J, Verdu E, Navarro X (2006) Acute and delayed transplantation of olfactory ensheathing cells promote partial recovery after complete transection of the spinal cord. Neurobiol Dis 21:57-68. Lu J, Feron F, Ho SM, Mackay-Sim A, Waite PM (2001) Transplantation of nasal olfactory tissue promotes partial recovery in paraplegic adult rats. Brain Res 889:344-357. Lu J, Feron F, Mackay-Sim A, Waite PME (2002) Olfactory ensheathing cells promote locomotor recovery after delayed transplantation into transected spinal cord. Brain 125:14-21. Lu P, Yang H, Jones LL, Filbin MT, Tuszynski MH (2004) Combinatorial therapy with neurotrophins and cAMP promotes axonal regeneration beyond sites of spinal cord injury. J Neurosci 24:6402-6409. Luster AD, Ravetch JV (1987) Genomic characterization of a gamma-interferon-inducible gene (IP-10) and identification of an interferon-inducible hypersensitive site. Mol Cell Biol 7:3723-3731. Ma M, Wei T, Boring L, Charo IF, Ransohoff RM, Jakeman LB (2002) Monocyte recruitment and myelin removal are delayed following spinal cord injury in mice with CCR2 chemokine receptor deletion. J Neurosci Res 68:691-702. Mackay-Sim A (2005) Olfactory ensheathing cells and spinal cord repair. J Med 54:8-14. Mackay-Sim A, Kittel P (1991) Cell dynamics in the adult mouse olfactory epithelium: a quantitative autoradiographic study. J Neurosci 11:979-984.
References
180
Madison R, da Silva CF, Dikkes P, Chiu TH, Sidman RL (1985) Increased rate of peripheral nerve regeneration using bioresorbable nerve guides and a laminin-containing gel. Exp Neurol 88:767-772. Magavi SSP, Mitchell BD, Szentirmai O, Carter BS, Macklis JD (2005) Adult-born and preexisting olfactory granule neurons undergo distinct experience-dependent modification of their olfactory responses in vivo. J Neurosci 25:10729-10739. Mahanthappa NK, Anton ES, Matthew WD (1996) Glial growth factor 2, a soluble neuregulin, directly increases Schwann cell motility and indirectly promotes neurite outgrowth. J Neurosci 16:4673-4683. Marchionni MA, Goodearl AD, Chen MS, Bermingham-McDonogh O, Kirk C, Hendricks M, Danehy F, Misumi D, Sudhalter J, Kobayashi K, Wroblewski D, Lynch C, Baldassare M, Hiles I, Davis JB, Hsuan JJ, Totty NF, Otsu M, BcBurney RN, Waterfield MD, Stoobant P, Gwynne D (1993) Glial growth factors are alternatively spliced erbB2 ligands expressed in the nervous system. Nature 362:312-318. Marin-Padilla M, Amieva MR (1989) Early neurogenesis of the mouse olfactory nerve: Golgi and electron microscopic studies. J Comp Neurol 288:339-352. Martin D, Robe P, Franzen R, Delree P, Schoenen J, Stevenaert A, Moonen G (1996) Effects of Schwann cell transplantation in a contusion model of rat spinal cord injury. J Neurosci Res 45:588-597. Martini R, Bollensen E, Schachner M (1988) Immunocytological localization of the major peripheral nervous system glycoprotein P0 and the L2/HNK-1 and L3 carbohydrate structures in developing and adult mouse sciatic nerve. Dev Biol 129:330-338. Martini R, Mohajeri MH, Kasper S, Giese KP, Schachner M (1995) Mice doubly deficient in the genes for P0 and myelin basic protein show that both proteins contribute to the formation of the major dense line in peripheral nerve myelin. J Neurosci 15:4488-4495. McDonald JW, Liu XZ, Qu Y, Liu S, Mickey SK, Turetsky D, Gottlieb DI, Choi DW (1999) Transplanted embryonic stem cells survive, differentiate and promote recovery in injured rat spinal cord. Nat Med 5:1410-1412. McKerracher L, David S, Jackson DL, Kottis V, Dunn RJ, Braun PE (1994) Identification of myelin-associated glycoprotein as a major myelin-derived inhibitor of neurite growth. Neuron 13:805-811.
References
181
McTigue DM, Tani M, Krivacic K, Chernosky A, Kelner GS, Macicjewski D, Maki R, Ransohoff RM, Stokes BT (1998) Selective chemokine mRNA accumulation in the rat spinal cord after contusion injury. J Neurosci Res 53:368-376. Melcangi RC, Ballabio M, Cavarretta I, Gonzalex LC, Leonelli E, Veiga S, Martini L, Magnaghi V (2003) Effects of neuroactive steroids on myelin of peripheral nervous system. J Steroid Biochm & Mol Biol 85:323-327. Menei P, Montero-Menei C, Whittemore SR, Bunge RP, Bunge MB (1998) Schwann cells genetically modified to secrete human BDNF promote enhanced axonal regrowth across transected adult rat spinal cord. Eur J Neurosci 10:607-621. Meyer D, Birchmeier C (1995) Multiple essential functions of neuregulin in development. Nature 378:386-390. Michailov G, Sereda MW, Brinkmann BG, Fischer TM, Haug B, Birchmeier C, Role L, Lai C, Schwab MH, Nave K-A (2004) Axonal neuregulin-1 regulates myelin sheath thickness. Science 304:700-3. Miragall F, Kadmon G, Husmann M, Schachner M (1988) Expression of cell adhesion molecules in the olfactory system of the adult mouse: presence of the embryonic form of N-CAM. Dev Biol 129:516-531. Mirsky R, Jessen KR (1999) The neurobiology of Schwann cells. Brain Path 9:293-311. Molineaux SM, Engh H, De Ferra F, Hudson L, Lazzarini RA (1986) Recombination within the myelin basic protein gene created the dysmyelinating shiverer mouse mutation. PNAS 83:7542-7546. Morell P, Barrett CV, Mason JL, Toews AD, Hostettler, JD, Knapp GW, Matsushima GK (1998) Gene expression in brain during cuprizone-induced demyelination and remyelination. Mol Cell Neurosci 12:220-227. Moreno-Flores MT, Lim F, Martin-Bermejo J, Diaz-Nido J, Avila J, Wandosell F (2003) Immortalized olfactory ensheathing glia promote axonal regeneration of rat retinal ganglion neurons. J Neurochem 85:861-871. Morris JK, Lin W, Hauser C, Marchuk Y, Getman D, Lee KF (1999) Rescue of the cardiac defect in erbB2 mutant mice reveals essential roles of erbB2 in peripheral nervous system development. Neuron 23:273-283.
References
182
Morrissey TK, Kleitman N, Bunge RP (1991) Isolation and functional characterisation of Schwann cells derived from adult peripheral nerve. J Neurosci 11:2433-2442. Morrissey TK, Levi AD, Nuijens A, Sliwkowski MX, Bunge RP (1995) Axon-induced mitogenesis of human Schwann cells involves heregulin and p185erbB2. Proc Natl Acad Sci USA 92:1431-1435. Mu X, Azbill RD, Springer JE (2000) Riluzole and methylprednisolone combined treatment improves functional recovery in traumatic spinal cord injury. J Neurotrauma 17:773-780. Mukhopadhyay G, Doherty P, Wash FS, Crocker PR, Filbin MT (1994) A novel role for myelin-associated clycoprotein as an inhibitor of axonal regeneration. Neuron 13:757-767. Murrell W, Bushell GR, Livesey J, McGrath J, MacDonald KP, Bates PR, Mackay-Sim A (1996) Neurogenesis in adult human. Neuroreport 76:1189-1194. Nash HH, Borke RC, Anders JJ (2001) New method of purification for establishing primary cultures of ensheathing cells from the adult olfactory bulb. Glia 34:81-87. Nash HH, Borke RC, Anders JJ (2002) Ensheathing cells and methylprednisolone promote axonal regeneration and functional recovery in the lesioned adult rat spinal cord. J Neurosci 22:7111-7120. Namiki J, Kojima A, Tator CH (2000) Effect of brain-derived neurotrophic factor, nerve growth factor, and neurotrophin-3 on functional recovery and regeneration after spinal cord injury in adult rats. J Neurotrauma 17:1219-1231. Navarro X, Valero A, Gudino G, Fores J, Rodriguez FJ, Verdu E, Pascual R, Cuadras J, Nieto-Sampedro M (1999) Ensheathing glia transplants promote dorsal root regeneration and spinal reflex restitution after multiple lumbar rhizotomy. Annals Neurol 45:207-215. Neumann S, Bradke F, Tessier-Lavigne M, Basbaum AI (2002) Regeneration of sensory axons within the injured spinal cord induced by intraganglionic cAMP elevation. 34:885-893. Newman SL, Tucci, MA (1990) Regulation of human monocyte/macrophage function by extracellular matrix. Adherence of monocytes to collagen matrices enhances phagocytosis of opsonised bacteria by activation of complement receptors and enhancement of Fc receptor function. J Clin Invest 86:703-714.
References
183
Ni C-Y, Murphy MP, Golde TE, Carpenter G (2001) γ-Secretase cleavage and nuclear localization of ErbB-4 receptor tyrosine kinase. Science 294:2179-2181. Nikulina E, Tidwell JL, Dai HN, Bregman BS, Filbin MT (2004) The phosphodiesterase inhibitor rolipram delivered after a spinal cord lesion promotes axonal regeneration and functional recovery. Proc Natl Acad Sci USA 101:8786-8790. Norgren RB Jr, Ratner N, Brackenbury R (1992) Development of olfactory nerve glia defined by a monoclonal antibody specific for Schwann cells. Dev Dynamics 194:231-238. Novikova LN, Novikov LN, Kellerth JO (2002) Differential effects of neurotrphins on neuronal survival and axonal regeneration after spinal cord injury in adult rats. J Comp Neurol 452:255-263. Novikova LN, Novikov LN, Kellerth JO (2003) Biopolymers and biodegradable smart implants for tissue regeneration after spinal cord injury. Curr Opin Neurol 16:711-715. Ousman SS, David S (2000) Lysophosphatidylcholine induces rapid recruitment and activation of macrophages in the adult mouse spinal cord. Glia 30:92-104. Ousman SS, David S (2001) MIP-1α, MCP-1, GM-CSF, and TNF-α control the immune cell response that mediates rapid phagocytosis of myelin from the adult mouse spinal cord. J Neurosci 21:4649-4656. Pascual JI, Gudino-Cabrera G, Insausti R, Nieto-Sampedro M (2002) Spinal implants of olfactory ensheathing cells promote axon regeneration and bladder activity after bilateral lumbosacral dorsal rhizotomy in the adult rat. J Urol 167:1522-1526. Pavelko KD, van Engelen BGM, Rodriguez M (1998) Acceleration in the rate of CNS remyelination in lysolecithin-induced demyelination. J Neurosci 18:2498-2505. Pearse DD, Pereira FC, Marcillo AE, Bates ML, Berrocal YA, Filbin MT, Bunge MA (2002) cAMP and Schwann cells promote axonal growth and functional recovery after spinal cord injury. Nat Medicine 10:610-616. Pearse DD, Marcillo AE, Oudega M, Lynch MP, Wood PM, Bunge MB (2004) Transplantation of Schwann cells and olfactory ensehathing glia after
References
184
spinal cord injury: does pretreatment with methylprednisolone and interleukin-10 enhance recovery? J Neurotrauma 21:1223-1239. Peirano RI, Goerich DE, Riethmacher D, Wegner M (2000) Protein zero gene expression is regulated by the glial transcription factor Sox10. Mol Cell Biol 20:3198-3209. Peles E, Bacus SS, Koski RA, Lu HS, Wen D, Ogden SG, Levy RB, Yarden Y (1992) Isolation of the neu/HER-2 stimulatory ligand: A 44 kd glycoprotein that induces differentiation of mammary tumor cells. Cell 69:205-216. Perez-Bouza A, Wigley CB, Nacimiento W, Noth J, Brook GA (1998) Spontaneous orientation of transplanted olfactory gloia influences axonal regeneration. Neuroreport 9:2971-2975. Perry VH, Brown MC, Gordon S (1987) The macrophage response to central and peripheral nerve injury. A possible role for macrophages in regeneration. J Exp med 164:1218-1223. Pinkas-Kramarski R, Soussan L, Waterman H, Levkowitz G, Alroy I, Klapper L, Lavi S, Seger R, Ratzkiin BJ, Sela M, Yarden Y (1996) Diversification of Neu differentiation factor and epidermal growth factor signalling by combinatorial receptor interactions. EMBO J 15:2452-2467. Pixley SK (1992) The olfactory nerve contains two populations of glia, identified both in vivo and in vitro. Glia 5:269-284. Pixley SK (1996) Characterization of olfactory receptor neurons and other cell types in dissociated rat olfactory cell cultures. Int J Devl Neurosci 14:823-839. Pizzo DP, Paban V, Coufal NG, Gage FH, Thal LJ (2004) Long-term production of choline acetyltransferase in the CNS after transplantation of fibroblasts modified with a regulatable factor. Brain Res Mol Brain Res 126:1-13. Plant GW, Currier PF, Cuervo EP, Bates ML, Pressman Y, Bunge MB, Wood PM (2002) Purified adult ensheathing glia fail to myelinate axons under culture conditions that enable Schwann cells to form myelin. J Neurosci 22:6083-6091. Plant GW, Christensen CL, Oudega M, Bunge MB (2003) Delayed transplantation of olfactory ensheathing glia promotes sparing/regeneration of supraspinal axons in the contused adult rat spinal cord. J Neurotrauma 20:1-16.
References
185
Plowman GD, Whitney GS, Neubauer MG, Green JM, McDonald VL, Todaro GJ, Shoyab M (1990) Molecular cloning and expression of an additional epidermal growth factor receptor-related gene. Proc Natl Acad Sci USA 87:4905-4909. Plowman GD, Culouscou JM, Whitney GS, Green JM, Carlton GW, Foy L, Neubauer MG, Shoyab M (1993) Ligand-specific activation of HER4/p180erbB4, a fourth member of the epidermal growth factor receptor family. Proc Natl Acad Sci USA 90:1746-1750. Polentes J, Stamegna JC, Nieto-Sampedro M, Gauthier P (2004) Phrenic rehabilitation and diaphragm recovery after cervical injury and transplantation of olfactory ensheathing cells. Neurobiol Dis 16:638-653. Pollock GS, Franceschini IA, Graham G, Marchionni MA, Barnett SC (1999) Neuregulin is a mitogen and survival factor for olfactory bulb ensheathing cells and an isoform is produced by astrocytes. Eur. J. Neurosci 11:761-768. Prewitt CM, Niesman IR, Kane CJ, Houle JD (1997) Activated macrophage/microglial cells can promote the regeneration of sensory axons into the injured spinal cord. Exp Neurol 148:433-443. Privat A, Jacque C, Bourre JM, Dupouey P, Baumann N (1979) Absence of the major dense line in myelin of the mutant mouse “shiverer”. Neurosci Lett 12:107-112. Purves-Tyson TD, Keast JR (2004) Rapid actions of estradiol on cyclic amp response-element binding protein phosphorylation in dorsal root ganglion neurons. Neurosci 129:629-637. Quarles RH (2005) Comparison of CNS and PNS myelin proteins in the pathology of myelin disorders. J Neurol Sci 228:187-189. Quarles RH, Everly JL, Brady RO (1972) Demonstration of a glycoprotein which is associated with a purified myelin fraction from rat brain. Biochem Biophys Res Commun 47:491-497. Qiu J, Cai D, Dai H, McAtee M, Hoffman PN, Bregman BS, Filbin MT (2002) Spinal axon regeneration induced by elevation of cyclic AMP. Neuron 34:895-903. Rabchevsky AG, Streit WJ (1997) Grafting of cultured microglial cells into the lesioned spinal cord of adult rats enhances neurite outgrowth. J Neurosci Res 47:34-48.
References
186
Rabinovich SS, Seledtsov VI, Poveschenko OV, Senuykov VV, Taraban VY, Yarochno VI, Kolosov NG, Savchenko SA, Kozlov VA (2003) Transplantation treatment of spinal cord injury patients. Biomed Pharmacother 57:428-433. Radtke C, Akiyama A, Brokaw J, Lankford KL, Wewetzer K, Fodor WL, Kocsis JD (2003) Remyelination of the nonhuman primate spinal cord by tranplantation of H-transferase transgenic adult pig olfactory ensheathing cells. FASEB J 18:335-7 Raff MC, Abney E, Brockes JP, Hornby-Smith A (1978) Schwann cell growth factors. Cell 15:813-822. Raisman G (1985) Specialized neuroglial arrangement may explain the capacity of vomeronasal axons to reinnervate central neurons. Neuroscience 14:237-254. Ramer LM, Au E, Richter MW, Liu J, Tetzlaff W, Roskams AJ (2004a) Peripheral olfactory ensheathing cells reduce scar and cavity formation and promote regeneration after spinal cord injury. J Comp Neurol 473:1-15. Ramer LM, Richter MW, Roskams AJ, Tetzlaff W, Ramer MS (2004b) Peripherally-derived olfactory ensheathing cells do not promote primary afferent regeneration following dorsal root injury. Glia 47:189-206. Ramon-Cueto A, Nieto-Sampedro M (1992) Glial cells from adult rat olfactory bulb: immunocytochemical properties of pure cultures of ensheathing cells. Neurosci 47:213-220. Ramon-Cueto A, Nieto-Sampedro M (1994) Regeneration into the spinal cord of transected dorsal root axons is promoted by ensheathing glia transplants. Exp Neurol 127:232-244. Ramon-Cueto A, Valvede F (1996) Olfactory bulb ensheathing glia: a unique cell type with axonal growth-promoting properties. Glia 14:163-173. Ramon-Cueto A, Perez J, Nieto-Sampedro M (1993) In vitro enfolding of olfactory neurites by p75 NGF receptor positive ensheathing cells from adult rat olfactory bulb. Eur J Neurosci 5:1172-1180. Ramon-Cueto A, Plant GW, Avila J, Bunge MB (1998) Long-distance axonal regeneration in the transected adult rat spinal cord is promoted by olfactory ensheathing glia transplants. J Neurosci 18:3803-3815.
References
187
Ramon-Cueto A, Cordero MI, Santos-Benito FF, Avila J (2000) Functional recovery of paraplegic rats and motor axon regeneration in their spinal cords by olfactory ensheathing glia. Neuron 25:425-435. Rapalino O, Lazarov-Spiegler O, Agranov E, Velan GJ, Yoles E, Fraidakis M, Solomon A, Gepstein R, Katz A, Belkin M, Hadani M, Schwartz M (1998) Implantation of homologous macrophages results in partial recovery of paraplegic rats. Nature Med 4:814-821. Readhead C, Popko B, Takahashi N, Shine HD, Saavedra RA, Sidman RL, Hood L (1987) Expression of a myelin basic protein gene in transgenic shiverer mice: correction of the dysmyelinating phenotype. Cell 48:703-712. Reichert F, Saada A, Rotshenker S (1994) Peripheral nerve injury induces Schwann cells to express two macrophage phenotypes: Phagocytosis and the galactose-specific lectin MAC-2. J Neurosci 14:3231-3245. Reier PJ, Perlow MJ, Guth L (1983) Development of embryonic spinal cord transplants in the rat. Brain Res 312:201-219. Resnick DK, Cechvala CF, Yan Y, Witwer BP, Sun D, Zhang S (2003) Adult olfactory ensheathing cell transplantation for acute spinal cord injury. J Neurotrauma 20:279-285. Reynolds ES (1963) The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J Cell Biol 17:208-212. Reynolds R, Wilkin GP (1988) Development of macroglial cells in rat cerebellum. II. An in situ immunohistochemical study of oligodendroglial lineage from precursor to mature myelinating cell. Development 102:409-425. Richardson PM, McGuinness UM, Aguayo AJ (1980) Axons from CNS neurons regenerate into PNS grafts. Nature 284:264-265. Richardson PM, McGuinness UM, Aguayo AJ (1982) Peripheral nerve autografts to the rat spinal cord: studies with axonal tracing methods. Brain Res 237:147-162. Richardson PM, Issa VM, Aguayo AJ (1984) Regeneration of long spinal axons in the rat. J Neurocytol 13:165-182. Richter MW, Fletcher PA, Liu J, Tetzlaff W, Roskams AJ (2005) Lamina propria and olfactory bulb ensheathing cells exhibit differential integration and migration and promote differential axon sprouting in the lesioned spinal cord. J Neurosci 25:10700-10711.
References
188
Riddell JS, Enriquez-Denton M, Toft A, Fairless R, Barnett SC (2004) Olfactory ensheathing cell grafts have minimal influence on regeneration at the dorsal root entry zone following rhizotomy. Glia 150-167. Riethmacher D, Sonnenberg-Riethmacher E, Brinkmann V, Yamaai T, Lewin GR, Birchmeier C (1997) Severe neuropathies in mice with targeted mutations in the ErbB3 receptor. Nature 725-730. Rimer M, Prieto AL, Weber JL, Colasante C, Ponomareva O, Fromm L, Schwab MH, Lai C, Burden SJ (2004) Neuregulin-2 is synthesized by motor neurons and terminal Schwann cells and activates acetylcholine receptor transcription in muscle cells expressing ErbB4. Mol Cell Neurosci 26:271-281. Riva-Depaty I, Fardeau C, Mariani J, Bouchaud C, Delhaye-Bouchaud, N (1994) Contribution of peripheral macrophages and microglia to the cellular reaction after mechanical or neurotoxin-induced lesions of the rat brain. Exp Neurol 128:77-87. Rosenbluth J (1980) Central myelin in the mouse mutant shiverer. J Comp Neurol 194:639-648. Rothe T, Muller HW (1991) Uptake of endoneurial lipoprotein into Schwann cells and sensory neurons is mediated by low density lipoprotein receptors and stimulated after axonal injury. J Neurochem 57:2016-2025. Ruitenberg MJ, Plant GW, Christensen CL, Blits B, Niclou SP, Harvey AR, Boer GJ, Verhaagen J (2002) Viral vector-mediated gene expression in olfactory ensheathing glia implants in the lesioned rat spinal cord. Gene Ther 9:135-146. Ruitenberg MJ, Plant GW, Hamers FPT, Wortel J, Blits B, Dijkhuizen PA, Gispen WH, Boer GJ, Verhaagen J (2003) Ex vivo adenoviral vector-mediated neurotrophin gene transfer to olfactory ensheathing glia: effects on rubrospinal tract regeneration, lesion size, and functional recovery after implantation in the injured rat spinal cord. J Neurosci 23:7045-7058. Ruitenberg MJ, Levison DB, Lee SV, Verhaagen J, Harvey AR, Plant GW (2005a) NT-3 expression from engineered olfactory ensheathing glia promotes spinal sparing and regeneration. Brain 128:839-853. Ruitenberg MJ, Vukovic, Sarich J, Busfield SJ, and Plant GW (2005b) Olfactory ensheathing cells: characteristics, genetic engineering and therapeutic potential. J Neurotrauma (In press).
References
189
Rutkowski JL, Kirk CJ, Lerner MA, Tennekoon GI (1995) Purification and expansion of human Schwann cells in vitro. Nature Med 1:80-83. Saada A, Dunaevsky-Hutt A, Aamar A, Reichert F, Rotschenker S (1995) Fibroblasts that reside in mouse and frog injured peripheral nerves produce apolipoproteins. J Neurochem 64:1996-2003. Sandrock Jr AW, Dryer SE, Rosen KM, Gozani SN, KMramer R, Theill LE, Fischbach GD (1997) Maintenance of acetylcholine receptor number by neuregulins at the neuromuscular junction in vivo. Science 276:599-603. Santos-Benito FF, Ramon-Cueto A (2003) Olfactory ensheathing glia transplantation: a therapy to promote repair in the mammalian central nervous system. Anat Rec 271B:77-85. Santos-Silva A, Cavalcante LA (2001) Expression of the non-compact myelin protein 2',3'-cyclic nucleotide 3'-phosphodiesterase (CNPase) in olfactory bulb ensheathing glia from explant cultures. Neurosci Res 40:189-193. Sasaki M, Lankford KL, Zemedkun M, Kocsis JD (2004) Identified olfactory ensheathing cells transplanted into the transected dorsal funiculus bridge the lesion and form myelin. J Neurosci 24:485-8493. Sasaki M, Hains BC, Lankford KL, Waxman, SG, Kocsis JD (2006) Protection of corticospinal tract neurons after dorsal spinal cord transection and engraftment of olfactory ensheathing cells. Glia 53:352-359. Schnell L, Schwab ME (1990) Axonal regeneration in the rat spinal cord produced by an antibody against myelin-associated neurite growth inhibitors. Nature 343:269-272. Schnell L, Schneider R, Kolbeck R, Barde YA, Schwab ME (1994) Neurotrophin-3 enhances sprouting of corticospinal tract during development and after adult spinal cord lesion. Nature 367:170-173. Schnell L, Fearn S, Klassen H, Schwab ME, Perry VH (1999a) Acute inflammatory responses to mechanical lesions in the CNS: differences between brain and spinal cord. Eur J Neurosci 11:3648-3658. Schnell L, Fearn S, Schwab ME, Perry VH, Anthony DC (1999b) Cytokine-induced acute inflammation in the brain and spinal cord. J Neuropathol Exp Neurol 58:245-254. Seamon KB, Padgett W, Daly JW (1981) Forskolin: unique diterpene activator of adenylate cyclase in membranes and in intact cells. Proc Natl Acad Sci USA 78:3363-3367.
References
190
Sekerkova G, Katarova Z, Joo F, Wolff JR, Prodan S, Szabo G (1997) Visualisation of β-galactosidase by enzyme and immunohistochemistry in the olfactory bulb of transgenic mice carrying the LacZ transgene. J Histochem & Cytochem 45:1147-1155. Shah NM, Anderson DJ (1997) Integration of multiple instructive cues by neural crest stem cells reveals cell-intrinsic biases in relative growth factor responsiveness. Proc Natl Acad Sci USA 94:11369-11374. Shah NM, Marchionni MA, Isaacs I, Stroobant P, Anderson DJ (1994) Glial growth factor restricts mammalian neural crest stem cells to a glial fate. Cell 77:349-360. Shen H, Tang Y, Wu Y, Chen Y, Cheng Z (2002) Influences of olfactory ensheathing cells transplantation on axonal regeneration in spinal cord of adult rats. Chin J Traumatol 5:136-141. Shepherd GM, Greer CA (1998) Olfactory bulb. In: The synaptic organisation of the brain (Shepherd GM ed), pp159-203. Oxford University Press: New York Sims TJ, Gilmore SA (1983) Interactions between intraspinal Schwann cells and the cellular constituents normally occurring in the spinal cord: an ultrastructural study in the irradiated rat. Brain Res 276:17-30. Sliwkowski MX, Schaefer G, Akita RW, Lofgren JA, Fitzpatrick VD, Nuijens A, Fendly BM, Cerione RA, Vandlen RL, Carraway KL III (1994) Coexpression of erbB2 and erbB3 proteins reconstitutes a high affinity receptor for heregulin. J Biol Chem 269:14661-14665. Smale KA, Doucette R, Kawaja MD (1996) Implantation of olfactory ensheathing cells in the adult rat brain following fimbria-fornix transection. Exp Neurol 137:225-233. Smith ME, van der Maesen K, Somera FP (1998) Macrophage and microglia responses to cytokines in vitro: phagocytic activity, proteolytic enzyme release, and free radical production. J Neurosci Res 54:68-78. Smith PM, Sim FJ, Barnett SC, Franklin RJ (2001) SCIP/Oct-6, Krox-20, and desert hedgehog mRNA expression during CNS remyelination by tranplanted olfactory ensheathing cells. Glia 36:342-353. Smith PM, Lakatos A, Barnett SC, Jeffery ND, Franklin RJM (2002) Cryopreserved cells isolated from the adult canine olfactory bulb are capable
References
191
of extensive remyelination following transplantation into the adult rat CNS. Exp Neurol 176:402-406. Snipes GJ, McGuire CB, Norden JJ, Freeman JA (1986) Nerve injury stimulates the secretion of apolipoprotein E by nonneuronal cells. Proc Natl Acad Sci USA 83:1130-1134. Sonigra RJ, Brighton PC, Jacoby J, Hall S, Wigley CB (1999) Adult rat olfactory nerve ensheathing cells are effective promoters of adult central nervous system neurite outgrowth in coculture. Glia 25:256-269. Sprinkle TJ (1989) 2',3'-cyclic nucleotide 3'-phosphodiesterase, an oligodendrocyte-Schwann cell and myelin-associated enzyme of the nervous system. Crit Rev Neurobiol 4:235-301. Stoll G, Griffin JW, Li CY, Trapp BD (1989) Wallerian degeneration in the peripheral nervous system: participation of both Schwann cells and macrophages in myelin degradation. J Neuroctyol 18:671-683. Sun TS, Ren JX, Shi JG (2005) Repair of acute spinal cord injury promoted by transplantation of olfactory ensheathing glia. Zhongguo Yi Xue ke Xue Yuan Xue Bao 27:143-147. Susuki Y, Takeda M, Farbman AI (1996) Supporting cells as phagocytes in the olfactory epithelium after bulbectomy. J Comp Neurol 376:509-517. Syroid DE, Maycox PR, Burrola PG, Liu N, Wen D, Lee K-F, Lemke G, Kilpatrick TJ (1996) Cell death in the Schwann cell lineage and its regulation by neuregulin. Proc Natl Acad Sci USA 93:9229-9234. Takami T, Oudega M, Bates ML, Wood PM, Kleitman N, Bunge MB (2002) Schwann cell but not olfactory ensheathing glia transplants improve hindlimb locomotor performance in the moderately contused adult rat thoracic spinal cord. J Neurosci 22:6670-6681. Tang S, Woodhall RW, Shen YJ, deBellard ME, Saffell JL, Doherty P, Walsh FS, Filbin MT (1997) Soluble myelin-associated glycoprotein (MAG) found in vivo inhibits axonal regeneration. Mol Cell Neurosci 9:333-346. Taylor JS, Muneton-Gomez VC, Eguia-Recuero R, Nieto-Sampedro M (2001) Transplants of olfactory ensheathing cells promote functional repair of multiple dorsal rhizotomy. Prog Brain Res 132:641-654. Tennent R, Chuah MI (1996) Ultrastructural study of ensheathing cells in early development of olfactory axons. Dev Brain Res 95:135-139.
References
192
Thompson RJ, Roberts B, Alexander CL, Williams SK, Barnett SC (2000) Comparison of neuregulin-1 expression in olfactory ensheathing cells, Schwann cells and astrocytes. J Neurosci Res 61:171-85. Tisay KT, Key B (1999) The extracellular matrix modulates olfactory neurite outgrowth on ensheathing cells. J Neurosci 19:9890-9899. Toews AD, Barrett C, Morell P (1998) Monocyte chemoattractant protein 1 is responsible for macrophage recruitment following injury to sciatic nerve. J Neurosci Res 53:260-267. Trachtenberg JT, Thompson WJ (1996) Schwann cell apoptosis at developing neuromuscular junctions is regulated by glial growth factor. Nature 379:174-177. Tzahar E, Levkowitz G, Karunagaran D, Yi L, Peles E, Lavi S, Chang D, Liu N, Yayon A, Wen D, Yarden Y (1994) ErbB-3 and erbB-4 function as the respective low and high affinity receptor of all neu differentiation factor/heregulin isoforms. J Biol Chem 269:25226-25233. Tzakos AG, Troganis A, Theodorou V, Tselios T, Svarnas C, Matsoukas J, Apostopoulos V, Georthanassis IP (2005) Structure and function of the myelin proteins: current status and perspectives in relation to multiple sclerosis. Curr Med Chem 12:1569-1587. Ueno N, Baird A, Esch F, Shimasaki S, Ling N, Guillemin R (1986) Prufication and partial characterization of a mitogenic factor from bovine liver: structureal homology with basic fibroblast growth factor. Regul Pept 16:135-145. Valverde F, Lopez-Mascaraque L (1991) Neuroglial arrangements in the olfactory glomeruli of the hedgehog. J Comp Neurol 307:658-674. Van Den Pol AN, Santarelli JG (2003) Olfactory ensheathing cells: time lapse imaging of cellular interactions, axonal support, rapid morphologic shifts, and mitosis. J Comp Neurol 458:175-194. van der Geer P, Hunter T, Lindberg RA (1994) Receptor protein-tyrosine kinases and their signal transduction pathways. Annu Rev Cell Biol 10:251-337. Vance JE, Campenot RB, Vance DE (2000) The synthesis and transport of lipids for axonal growth and nerve regeneration. Biochim Biophys Acta 1486:84-96.
References
193
Vartanian T, Goodearl A, Viehover A, Fischbach G (1997) Axonal neuregulin signals cells of the oligodendrocyte lineage through activation of HER4 and Schwann cells through HER2 and HER3. J Cell Biol 137:211-220. Verdu E, Garcia-Alias G, Fores J, Gudino-Cabrera G, Muneton VC, Nieto-Sampedro M, Navarro X (2001) Effects of ensheathing cells transplanted into photochemically damaged spinal cord. Neuroreport 12:2303-2309. Verdu E, Garcia-Alias G, Fores J, Lopez-Vales R, Navarro X (2003) Olfactory ensheathing cells transplanted in lesioned spinal cord prevent loss of spinal cord parenchyma and promote functional recovery. Glia 42:275-286. Vincent AJ, West AK, Chuah MI (2003) Morphological plasticity of olfactory ensheathng cells is regulated by cAMP and endothelin-1. Glia 41:393-403. Vincent AJ, Taylor JM, Choi-Lundberg DL, West AK, Chuah MI (2005) Genetic expression profile of olfactory ensheathing cells is distinct from that of Schwann cells and astrocytes. Glia 51:132-147. Vogel US, Thompson RJ (1988) Molecular structure, localization and possible functions of the myelin-associated enzyme 2’,3’-cyclic nucleotide 3’-phosphodiesterase. J Neurochem 50:1667-1677. von Zahn J, Moller T, Kettenmann H, Nolte C (1997) Microglial phagocytosis is modulated by pro- and anti-inflammatory cytokines. Neuroimmunol 8:3851-3856. Wang CT, H SH, Yang H, Zhang P, Fei LL, You SW, Ju G (2005) Culture, purification and biological characters of olfactory ensheathing cells from adult transgenic mice. Shi Yan Sheng Wu Xue Bao 38:340-346. Weidner N, Blesch A, Grill RJ, Tuszynski MH (1999) Nerve growth factor-hypersecreting Schwann cell grafts augment and guide spinal cord axonal growth and remyelinate central nervous system axons in a phenotypically appropriate manner that correlates with expression of L1. J Comp Neurol 413:495-506. Weis J, Fine SM, David C, Savarirayan S, Sanes JR (1991) Integration site-dependent expression of a transgene reveals specialized features of cells associated with neuromuscular junction. J Cell Biol 113:1385-1397. Weltzien HU (1979) Cytolytic and membrane-perturbing properties of lysophosphatidylcholine. Biochim Biophys Acta 559:259-287. Wen D, Peles E, Cupples R, Suggs SV, Bacus SS, Luo Y, Trail G, Hu S, Silbiger SM, Levy RB, Koski RA, Lu HS, Yarden Y (1992) Neu
References
194
differentiation factor: a transmembrane glycoprotein containing an EGF domain and an immunoglobulin homology unit. Cell 69:559-572. Wen D, Suggs SV, Karunagaran D, Liu N, Cupples RL, Luo Y, Janssen AM, Ben-Baruch N, Trollinger DB, Jacobsen VL, Meng S-Y, Lu HS, Hu S, Chang D, Yang W, Yanigahara D, Koski RA, Yarden Y (1994) Structural and functional aspects of the multiplicity of Neu differentiation factors. Mol Cell Biol 14:1909-1919. Wewetzer K, Grothe C, Claus P (2001) In vitro expression and regulation of ciliary neurotrophic factor and its α receptor subunit in neonatal rat olfactory ensheathing cells. Neurosci Lett 306:165-168. Wewetzer K, Kern N, Ebel C, Radtke C, Brandes G (2005) Phagocytosis of O4+ axonal fragments in vitro by p75- neonatal rat olfactory ensheathing cells. Glia 49:577-587. Wiggins RC, Benjamins JA, Morell P (1975) Appearance of myelin proteins in rat sciatic nerve during development. 89:99-106. Williams SK, Franklin RJM, Barnett SC (2004) Response of olfactory ensheathing cells to the degeneration and regeneration of the peripheral olfactory system and the involvement of neuregulins. J Comp Neurol 470:50-62. Wilson KCP, Raisman G (1980) Age-related changes in the neurosensory epithelium of the mouse vomeronasal organ: extended period of post-natal growth in size and evidence for rapid cell turnover in the adult. Brain Res 185:103-113. Woerly S, Doan VD, Sosa N, de Vellis J, Espinosa-Jeffrey A (2004) Prevention of gliotic scar formation by NeuroGel allows partial endogenous repair of transected cat spinal cord. J Neurosci Res 75:262-272. Woldeyesus MT, Britsch S, Riethmacher D, Xu L, Sonnenberg-Riethmacher E, Abou-Rebyeh F, Harvey R, Caroni P, Birchmeier C (1999) Genes Dev 2538-2548. Wolpowitz D, Mason TBA, Dietrich P, Mendelsohn MA, Scherer SS (2000) Cysteine-rich domain isoforms of the neuregulin-1 gene are required for maintenance of peripheral synapses. Neuron 25:79-91. Wood JN, Anderton BH (1981) Monoclonal antibodies to mammalian neurofilaments. Bioscience Reports 1:263-268.
References
195
Woodhall E, West AK, Chuah MI (2001) Cultured olfactory ensheathing cells express nerve growth factor, brain-derived neurotrophic factor, glia cell line-derived neurotrophic factor and their receptors. Mol Brain Res 88:203-213. Woodhall E, West AK, Vickers JC, Chuah MI (2003) Olfactory ensheathing cell phenotype following implantation in the lesioned spinal cord. CMLS 60:2241-2253. Woodhouse A, Vincent AJ, Kozel MA, Chung RS, Waite PM, Vickers JC, West AK, Chuah MI (2005) Spinal cord tissue affects ensheathing cell proliferation and apoptosis. Neuroreport 16:737-740. Woodruff RH, Franklin JM (1999) The expression of myelin protein mRNAs during remyelination of lysolecithin-induced demyelination. Neuropathol Appl Neurobiol 25:226-235. Wrathall JR, Kapoor V, Kao CC (1984) Observation of cultured peripheral non-neuronal cells implanted into the transected spinal cord. Acta Neuropathol Berl 64:203-212. Xia C-Y, Yuan C-X, Yuan C-G (2005) Galanin inhibits the proliferation of glial olfactory ensheathing cells. Neuropeptides 39:453-459. Xu XM, Guenard V, Kleitman N, Aebischer P, Bunge MB (1995) A combination of BDNF and NT-3 promotes supraspinal axonal regeneration into Schwann cell grafts in adult rat thoracic spinal cord. Exp Neurol 134:261-272. Xu XM, Chen A, Guenard V, Kleitman N, Bunge MB (1997) Bridging Schwann cell transplants promote axonal regeneration from both the rostral and caudal stumps of transected adult rat spinal cord. J Neurocytol 26:1-16. Yan H, Bunge MB, Wood PM, Plant GW (2001a) Mitogenic response of adult rat olfactory ensheathing glia to four growth factors. Glia 33:334-342. Yan H, Nie X, Kocsis JD (2001b) Hepatocyte growth factor is a mitogen for olfactory ensheathing cells. J Neurosci Res 66:698-704. Yan H, Lu D, Rivkees SA (2003) Lysophosphatidic acid regulates the proliferation and migration of olfactory ensheathing cells in vitro. Glia 44:26-36. Yang X, Kuo Y, Devay P, Yu C, Role L (1998) A cysteine-rich isoform of neuregulin controls the level of expression of neuronal nicotinic receptor channels during synaptogenesis. Neuron 20:255-270.
References
196
Ye JH, Houle JD (1997) Treatment of the chronically injured spinal cord with neurotrophic factors can promote axonal regeneration from supraspinal neurons. Exp Neurol 143:70-81. Yin X, Peterson J, Gravel M, Braun PE, Trapp BD (1997) CNP overexpression induces aberrant oligodendrocyte membranes and inhibits MBP accumulation and myelin compaction. J Neurosci Res 50:238-247. Yoshino JE, Dinneen MP, Sprinkly TJ, DeVries GH (1985) Localization of 2’,3’-cyclic nucleotide 3’-phosphodiesterase on cultured Schwann cells. Brain Res 325:199-203. Zhang D, Sliwkowski MX, Mark M, Frantz G, Akita R, Sun Y, Hillan K, Crowley C, Brush J, Godowski PJ (1997) Neuregulin-3 (NRG3): a novel neural tissue-enriched protein that binds and activates ErbB4. Proc Natl Acad Sci USA 94:9562-9567. Zhang Y, Campbell G, Anderson PN, Martini R, Schachner M, Lieberman AR (1995) Molecular basis of interactions between regenerating adult rat thalamic axons and Schwann cells in peripheral nerve grafts I. Neural cell adhesion molecules. J Comp Neurol 361:193-209.