Research ArticleOlive (Olea europaea) Leaf Extract Induces Apoptosis andMonocyteMacrophage Differentiation in Human ChronicMyelogenous Leukemia K562 Cells Insight into the UnderlyingMechanism
Imen Samet1 Junkyu Han23 Lobna Jlaiel4 Sami Sayadi4 and Hiroko Isoda23
1 Graduate School of Life and Environmental Sciences University of Tsukuba Tennodai 1-1-1 Tsukuba Ibaraki 305-8572 Japan2 Faculty of Life and Environmental Sciences University of Tsukuba Tennodai 1-1-1 Tsukuba Ibaraki 305-8572 Japan3 Alliance of Research on North Africa University of Tsukuba Tennodai 1-1-1 Tsukuba Ibaraki 305-8572 Japan4 Laboratory of Environmental Bioprocesses Biotechnology Center of Sfax Sfax 3018 Tunisia
Correspondence should be addressed to Hiroko Isoda isodahirokogautsukubaacjp
Received 6 December 2013 Accepted 16 February 2014 Published 6 April 2014
Academic Editor Elisa Cabiscol
Copyright copy 2014 Imen Samet et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Differentiation therapy is an attractive approach aiming at reversing malignancy and reactivating endogenous differentiationprograms in cancer cells Olive leaf extract known for its antioxidant activity has been demonstrated to induce apoptosis inseveral cancer cells However its differentiation inducing properties and the mechanisms involved are still poorly understoodIn this study we investigated the effect of Chemlali Olive Leaf Extract (COLE) for its potential differentiation inducing effect onmultipotent leukemia K562 cells Results showed that COLE inhibits K562 cells proliferation and arrests the cell cycle at G0G1 andthen at G2M phase over treatment time Further analysis revealed that COLE induces apoptosis and differentiation of K562 cellstoward themonocyte lineage Microarray analysis was conducted to investigate the underlyingmechanism of COLE differentiationinducing effect The differentially expressed genes such as IFI16 EGR1 NFYA FOXP1 CXCL2 CXCL3 and CXCL8 confirmed thecommitment of K562 cells to the monocytemacrophage lineage Thus our results provide evidence that in addition to apoptosisinduction of differentiation is one of the possible therapeutic effects of olive leaf in cancer cells
1 Introduction
Several advances against cancer have been recently achievedthanks to different therapeutic modalities with radiation andchemotherapy being the most used so far Although thesetherapies have been proven successful against some tumorsthey are still highly toxic and nonspecific since their primarymode of action is DNA damage which results in severeadverse effects for normal cells [1] Differentiation inducingtherapy is therefore anticipated as a novel medical treatmentthat could reduce such adverse effects This new conceptwhich consists in forcingmalignant cells to undergo terminaldifferentiation instead of killing them through cytotoxicityhas so far gained a great interest especially for treatingleukemia Many compounds have been reported to induce
differentiation of leukemia cells and some of them are alreadyapproved for clinical use [2] Natural products have greatlycontributed to cancer therapy and a rising interest is beingattributed to the identification of new compounds from theplant resources with relevant effects against cancer develop-ment [3 4] Some of these compounds are now being used inclinical practice such as All-Trans Retinoic Acid Recent basicresearch studies and observational epidemiologic studiesstrongly support that the disease-preventing effects of naturalproducts are in part attributed to antioxidants even thoughtheir efficiency in vivo needs more investigations [5]
Olive leaves contain many potentially bioactive com-pounds that may have antioxidant antimicrobial antihyper-tensive antiviral anti-inflammatory hypoglycemic neuro-protective and anticancer properties [6ndash14] Olive leaf has
Hindawi Publishing CorporationOxidative Medicine and Cellular LongevityVolume 2014 Article ID 927619 16 pageshttpdxdoiorg1011552014927619
2 Oxidative Medicine and Cellular Longevity
gained the rising interest of the scientific and industrialcommunity due to its proved beneficial health propertiesand thus has emerged as commercially valuable nutraceu-ticals [15] The primary constituents which are believed tocontribute to the health benefits of olive leaves are Oleu-ropein Hydroxytyrosol as well as several other flavonoidssuch as Verbascoside Apigenin-7-glucoside and Luteolin-7-glucoside [14 16] Oleuropein the major constituent ofolive leaves has been shown to be a potent antioxidantIts radical scavenging activity has been well documented[6 17] Oleuropein has been shown to inhibit the oxida-tion of low density lipoproteins in vitro and in vivo [18]Jemai et al have demonstrated that polyphenols recoveredfrom olive leaf extracts Oleuropein Hydroxytyrosol andOleuropein aglycone exhibited a pronounced hypolipidemiceffect reduced the lipid peroxidation process and enhancedthe antioxidant defense system in experimental atherogenicmodel [19] Benavente-Garcıa et al [17] studied the antiox-idant activity of phenolic compounds from olive leaves andconcluded that olive phenolsmay exhibit synergistic behaviorin their radical scavenging capacity when mixed in thesame proportions as occur in the olive leaf extract Tworecent studies have focused on the bioavailability of oliveleaf phenolic compounds in human subjects and have cometo the conclusion that Oleuropein is rapidly absorbed andmetabolized to be mainly excreted as glucuronidated andsulfated Hydroxytyrosol suggesting that olive leaf extractcould exert benefits against oxidative stress-related processesin vivo [15 20]
In the prior studies olive leaf extract has been shown toexhibit an antitumor activity and to induce apoptosis path-ways in cancer cells little attention has been paid to its effecton the process of cancer cell differentiation Particularly oliveleaf has been reported to exhibit an antileukemia effect byinducing apoptosis in the acutemyeloid leukemiaHL-60 cells[13 21]
In this study we investigate the effect of olive leaf of themost abundant Tunisian variety Chemlali on the humanchronic myeloid leukemia K562 cells Compared to theleukemia cell lines used in previous studies [13 21] K562 cellscan be regarded as stem-like cells thanks to their pluripotency[22] and are known for their strong resistance to chemicalinducers [23]
We speculate that olive leaf extract may have anticarcino-genic property in K562 cells not only by inducing apoptosisbut also by inducing the commitment of leukemia cells to thematuration process in order to progressively give an apparentnormal cell life
2 Materials and Methods
21 Preparation of Olive Leaf Extract Collected olive leavesof Chemlali variety from the region of Sfax Tunisia wereair-dried and ground with a mixer Extraction was carriedby ethanol 70 (110 wv) in darkness for 2 weeks at roomtemperature The mixture was then centrifuged and filteredusing 045120583m filter (Millipore Japan) and stored at minus80∘Cuntil it was used
22 HPLC Analysis Chromatographic analyses were ach-ieved on an Agilent series 1260 HPLC-DAD instrument(Agilent Waldbronn Germany) The instrument includesa quaternary pump an online degasser an autosamplerand a thermostatically controlled column compartmentChromatographic separation was carried out on a ZOR-BAX Eclipse XDB-C18 column serial number USNH027266(46mm ID times 250mm times 35 120583m particle size) The elutionconditions were as follows mobile phase A (01 acetic acidin water) and mobile phase B (100 acetonitrile) flow rate of05mLmin sample injection volume of 10 120583L and operatingtemperature 40∘C The running gradient was as follows 0ndash22min 10ndash50 B 22ndash32min 50ndash100 B 32ndash40min100 B 40ndash44min 100ndash10 B Reequilibration durationlasted 6minThe DAD detector scanned from 190 to 400 nmand the samples were detected at 254 280 and 330 nm
23 Cell Line and Culture Conditions Human chronic leu-kemia cell line K562 was obtained from the Riken Cell Bank(Tsukuba Ibaraki Japan) Cells were cultured in RPMI 1640medium (Gibco) supplemented with 10 heat-inactivatedfetal bovine serum and maintained at 37∘C in a humidifiedincubator with 5 CO
2 The cells were pass-cultured every 3
days and used for experiments after reaching the exponentialgrowth phase
24 Cell Proliferation Assay Cell proliferation wasinvestigated by MTT (3-(45-dimethylthiazol-2-yl)-25-diphenyltetrazolium bromide) assay K562 cells were seededin 96-well plates at 20 times 104 cellsmL After incubation for24 h olive leave extract diluted in medium was added at finalconcentrations of 50 75 100 125 and 150120583gmL Controlcells were treated by ethanol at a final concentration of 03MTT was added after treatment for 24 48 and 72 h and theresulting formazan was completely dissolved by 100120583L of10 sodium dodecyl sulfate (SDS) for 24 h The absorbancewas determined at 570 nm in a multidetection microplatereader (Powerscan HT Dainippon Pharmaceutical NJUSA) Absorbance caused by the ability of the sample toreduce MTT or by its color was corrected using platesprepared in the same conditions in the absence of cells
25 Cell Viability Assay and Cell Morphological Changes Theviability of COLE treated cells was measured using flowcytometry according to the manufacturer instructions K562cells were seeded in 6-well plates at 20 times 104 cellsmL andtreated the following day by 50 100 and 150 120583gmL of COLEdiluted in medium and 03 ethanol in the case of thecontrol After incubation for the indicated time treated cellswere harvested suspended in Guava ViaCount reagent andallowed to be stained for at least 5min in darkness The cellnumber and viability were measured by Guava PCA flowcytometry (Guava Technologies CA USA) Morphologicalchanges were detected by observation under a phase contrastmicroscope (Leica Microsystem)
26 Cell Cycle Analysis The distribution of the cell cyclephases of treated and control cells was analysed by flow
Oxidative Medicine and Cellular Longevity 3
cytometry Briefly 20 times 104 cellsmL of K562 cells wereseeded in 6-well plates and treated by 100 and 150 120583gmLof COLE diluted in medium Control cells were treatedwith ethanol 03 After the desired time of incubationcells were harvested washed twice with PBS and fixed with70 ethanol at 4∘C for more than 12 h The fixed cells werethen centrifuged at 500timesg for 5min and washed with PBStwice Cell cycle reagent (Guava Technologies) was addedand the cells were kept in darkness for 30min at roomtemperature The population of cells in each cell cycle phasewas determined by a Guava PCA flow cytometry accordingto their DNA content
27 Annexin V Assay The induction of apoptosis in treatedcells was determined by measuring the externalization ofphosphatidylserine (PS) to the cell surface by flow cytometryK562 cells were seeded in 6 well plates and treated with 50100 and 150 ugmL of COLE diluted inmedium Control cellswere treated with ethanol 03 When reaching the desiredtreatment time cells were harvested and stained with GuavaNexin ReagentThen cells were incubated for 20min at roomtemperature in the dark and then acquired on theGuava PCAsystem
28 Cell Differentiation Assay Cell differentiation wasassessed by flow cytometry by measuring the expression ofCD11b and CD14 on the surface of K562 cells Cells wereseeded at 20 times 104 cellsmL in 6-well plates and incubatedfor 24 h The COLE diluted in medium was added at finalconcentrations of 50 100 and 150120583gmL and ethanol at03 in the case of control cells After incubation cellswere harvested washed twice with cold PBS and adjustedto the same number 10 times 105 The cells were then labeledwith phycoerythrin conjugated anti-CD14 anti-CD11banti-CD41 and anti-glycophorin A for 30min according tomanufacturerrsquos instructions The stained cells were washedtwice with cold PBS and resuspended in 500120583L PBS formeasurement
29 Total RNA Isolation K562 cells at a concentration of20 times 104 cellsmL were incubated for 24 h and then treatedwith 100 and 150 120583gmL of COLE for 72 h The control cellswere treated with ethanol at final concentration of 03The cells were then collected and washed with PBS DNA-free total RNA was isolated from the cells using Isogenreagent (Nippon Gene Co Tokyo Japan) following themanufacturerrsquos instructions
210 DNA Microarray Analysis Microarray hybridizationprobes were generated from isolated RNA samples Double-stranded cDNA was synthesized from 100 ng of total RNAusing the GeneAtlas 31015840 IVT Express Kit (Affymetrix Inc)Biotin-labeled aRNA was synthesized by in vitro transcrip-tion and purified 10 120583g of purified aRNA was then frag-mented using the GeneAtlas 31015840 IVT Express Kit and washybridized to the Affymetrix HG-U219 (Affymetrix) for 16 hat 45∘CThe chips were washed and stained in the GeneAtlasFluidics Station 400 (Affymetrix) and then imaged in the
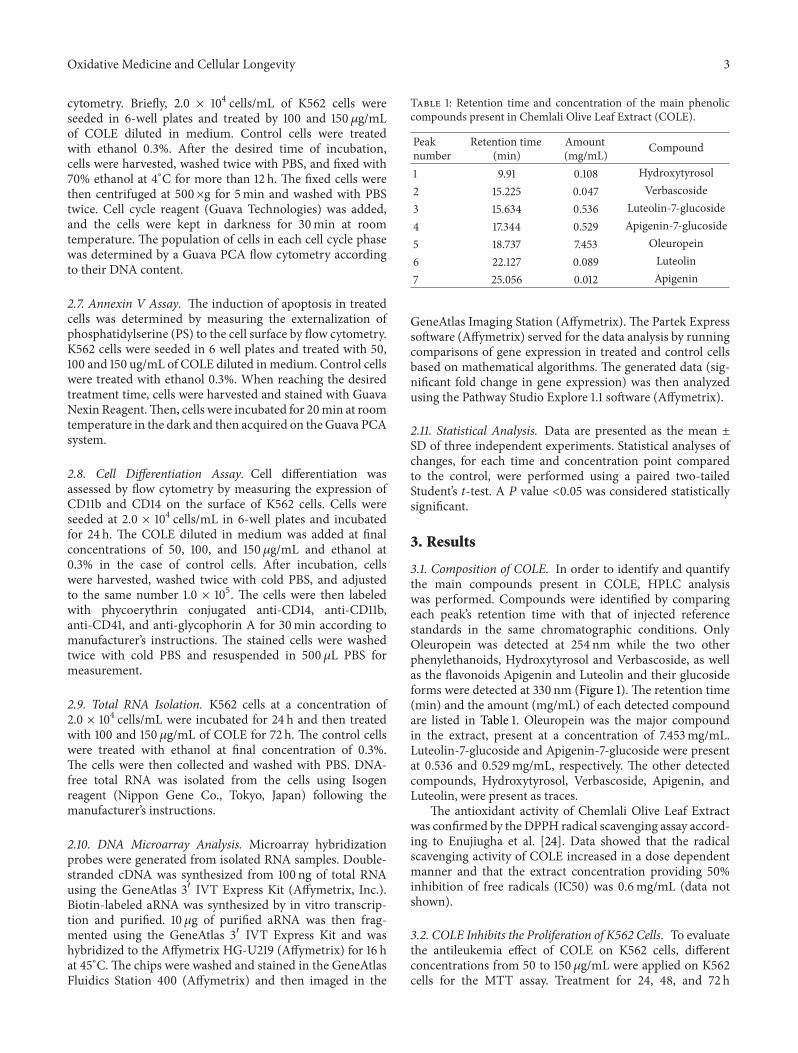
Table 1 Retention time and concentration of the main phenoliccompounds present in Chemlali Olive Leaf Extract (COLE)
GeneAtlas Imaging Station (Affymetrix) The Partek Expresssoftware (Affymetrix) served for the data analysis by runningcomparisons of gene expression in treated and control cellsbased on mathematical algorithms The generated data (sig-nificant fold change in gene expression) was then analyzedusing the Pathway Studio Explore 11 software (Affymetrix)
211 Statistical Analysis Data are presented as the mean plusmnSD of three independent experiments Statistical analyses ofchanges for each time and concentration point comparedto the control were performed using a paired two-tailedStudentrsquos t-test A 119875 value lt005 was considered statisticallysignificant
3 Results
31 Composition of COLE In order to identify and quantifythe main compounds present in COLE HPLC analysiswas performed Compounds were identified by comparingeach peakrsquos retention time with that of injected referencestandards in the same chromatographic conditions OnlyOleuropein was detected at 254 nm while the two otherphenylethanoids Hydroxytyrosol and Verbascoside as wellas the flavonoids Apigenin and Luteolin and their glucosideforms were detected at 330 nm (Figure 1) The retention time(min) and the amount (mgmL) of each detected compoundare listed in Table 1 Oleuropein was the major compoundin the extract present at a concentration of 7453mgmLLuteolin-7-glucoside and Apigenin-7-glucoside were presentat 0536 and 0529mgmL respectively The other detectedcompounds Hydroxytyrosol Verbascoside Apigenin andLuteolin were present as traces
The antioxidant activity of Chemlali Olive Leaf Extractwas confirmed by the DPPH radical scavenging assay accord-ing to Enujiugha et al [24] Data showed that the radicalscavenging activity of COLE increased in a dose dependentmanner and that the extract concentration providing 50inhibition of free radicals (IC50) was 06mgmL (data notshown)
32 COLE Inhibits the Proliferation of K562 Cells To evaluatethe antileukemia effect of COLE on K562 cells differentconcentrations from 50 to 150120583gmL were applied on K562cells for the MTT assay Treatment for 24 48 and 72 h
4 Oxidative Medicine and Cellular Longevity
5 254nm
0
200
400
600
800(m
AU)
5092 5330
6793
8008
9939
12511
14086
15191
16120
17086
18050
18737
19022
20132
21064
22125
23065
25779
26629
28065
29037
32310
34407
35982
36878
38286
41288
46112
0 10 20 30 40 50
(min)
(a)
1
2
3 4
6
7
330nm
5341
10429
11708
12683
13625
14768
15634
16254
16657
17344
17816
18026
18280 19230
19959
21050
22127
23051
46608
0
10
20
30
40
50
60
0 5 10 15 20 25 30 35 40 45
(mAU
)
(min)
(b)
Figure 1 HPLC chromatogram of Chemlali Olive Leaf Extract (COLE) at (a) 254 nm and (b) 330 nm Extraction was conducted with 70ethanol Peaks 1 Hydroxytyrosol 2 Verbascoside 3 Luteolin-7-glucoside 4 Apigenin-7-glucoside 5 Oleuropein 6 Luteolin 7 Apigenin
caused a significant decrease of the proliferation of K562 cellsin a dose dependent manner After 72 h of treatment with150 120583gmL of COLE the cell proliferation was inhibited to17 compared with the control cells (Figure 2(a))
Observation of the morphological changes of K562 cellsafter treatment with the different concentrations revealed anincrease in cell size especially in cells treated with 150120583gmLof COLE compared with control cells which kept the samemorphology and cell size (Figure 2(b))
33 COLE Inhibits the Growth Rate of K562 Cells To under-stand if the antiproliferative effect was led by cell death orgrowth inhibition we determined the cell number and cellviability after treatment with COLE by flow cytometry Thenumbers of K562 cells treated with 100 120583gmL and 150 120583gmLwere significantly less than the number of control cellsK562 cells treated with COLE exhibited a slow cell growthcompared to the control (Figure 3(a))
Results (Figure 3(b)) showed that treatment with150 120583gmL of COLE caused a slight decrease of the viabilityof K562 cells during the first 3 days with keeping more than
80 of viable cells Then the decrease became drastic andonly 20 of live cells remain at the 6th day of treatment bythe same concentration The viability of cells treated with100 120583gmL of COLE started to decrease significantly fromthe 4th day while treatment with 50120583gmL did not affect thecell viability during the whole period Taken together theseresults suggested that the antiproliferative effect of COLEat the concentrations of 100 120583gmL and 150 120583gmL mightbe conducted in part by inhibiting the growth rate and to alesser extent by causing cell death
34 COLE Modulates Cell Cycle Progression in K562 CellsSinceCOLE reduced the growth ofK562 cells we investigatedits effect on the cell cycle progression During the 1st and2nd day K562 cells treated with 100 and 150 120583gmL werearrested at G0G1 During the following days results showeda significant increase of G2M population in cell treated with100 and 150 120583gmL of COLE In the case of cells treatedwith 150120583gmL of COLE this increase was associated with asignificant decrease of G0G1 cell population throughout the3rd and 4th days of treatment (Table 2)
Oxidative Medicine and Cellular Longevity 5
0
20
40
60
80
100
120
24 48 72
Cel
l pro
lifer
atio
n (
of co
ntro
l)
Control50120583gmL75120583gmL
100120583gmL125120583gmL150120583gmL
lowastlowastlowast lowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
(h)
(a)
Control 50120583gmL
100120583gmL 150120583gmL
(b)
Figure 2 (a) Effect ofChemlaliOlive Leaf Extract (COLE) on the proliferation ofK562 cellsThe cells were treatedwith various concentrations(50 75 100 125 and 150120583gmL) of COLE for 24 48 and 72 h Cell proliferation was measured byMTT assay Control represents cells treatedwith 03 ethanol in medium Results are represented as the mean plusmn SD of three independent experiments lowast lowastlowast significantly differentfrom the control (119875 lt 005 and 119875 lt 001 resp) (b) Morphological observations of K562 cells after treatment with various concentrationsof Chemlali Olive Leaf Extract (COLE) for 72 h Cells were observed under a phase contrast microscope at 100x magnification Scale barsrepresent 100 120583m
0
2
4
6
8
10
12
14
16
18
20
Day 0 Day 1 Day 2 Day 3 Day 4 Day 6
Num
ber o
f via
ble c
ells
(times106)
Control 50120583gmL100 120583gmL 150120583gmL
(a)
0102030405060708090100
Day 0 Day 1 Day 2 Day 3 Day 4 Day 6
Viab
ility
()
Control50120583gmL
100 120583gmL150120583gmL
lowastlowastlowastlowast
lowastlowast
lowastlowast
lowastlowast
lowast lowast
(b)
Figure 3 Effect of Chemlali Olive Leaf Extract (COLE) on the cell number and viability of K562 cells (a) Number of viable cells aftertreatment with COLE after incubation up to 6 days (b) Percentage of viability in K562 cells treated with COLE K562 cells were treated at afinal concentration of 50 100 and 150120583gmL of COLE and incubated for different periods Cell number and cell viability were measured byflow cytometry Control represents cells treated with 03 ethanol in medium Results are represented as the mean plusmn SD of three independentexperiments lowast lowastlowast significantly different from the control (119875 lt 005 and 119875 lt 001 resp)
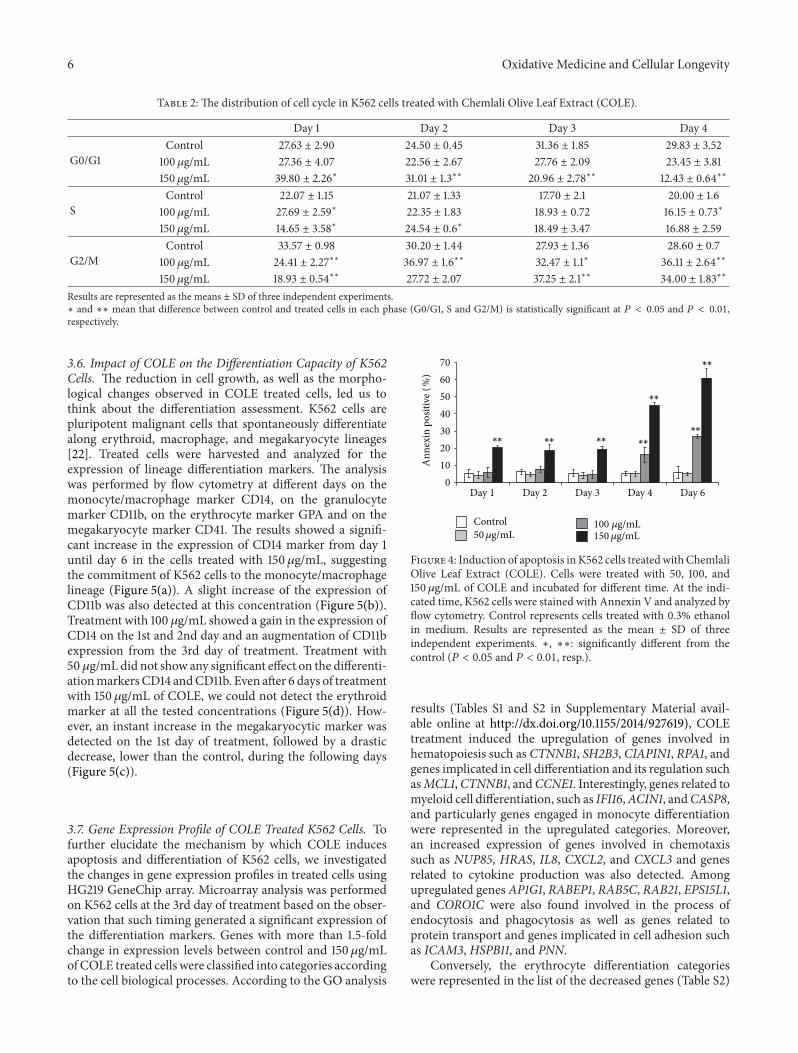
35 COLE Induces Apoptosis in K562 Cells The decreasein cell viability (Figure 3(b)) caused by COLE treatmentprompted us to investigate if the extract induces apoptosis inK562 cells Apoptosis is an important and active regulatorypathway of cell growth and proliferation resulting in somecharacteristic physiological changes Among these external-ization of phosphatidylserine (PS) is easily detected by flowcytometry after binding to the labeled Annexin VThe resultsindicated an increase in Annexin V positive cells from the1st day of incubation with 150 120583gmL of COLE in comparisonwith control cells (Figure 4) The percentage of Annexin V
cells remained stable during the first 3 days of treatmentwith 20 of total treated cells and interestingly increasedin the 4th and 6th days with 45 and 60 respectivelyTreatment with 100 120583gmL caused the apparition of apoptoticcells from the 4th day of incubation with a proportionof 162 This population increased in the 6th day up to267No significant difference in apoptotic cells amountwasdetected when cells were treated with 50 120583gmL until 6 daysof incubation
6 Oxidative Medicine and Cellular Longevity
Table 2 The distribution of cell cycle in K562 cells treated with Chemlali Olive Leaf Extract (COLE)
Results are represented as the means plusmn SD of three independent experimentslowast and lowastlowast mean that difference between control and treated cells in each phase (G0G1 S and G2M) is statistically significant at 119875 lt 005 and 119875 lt 001respectively
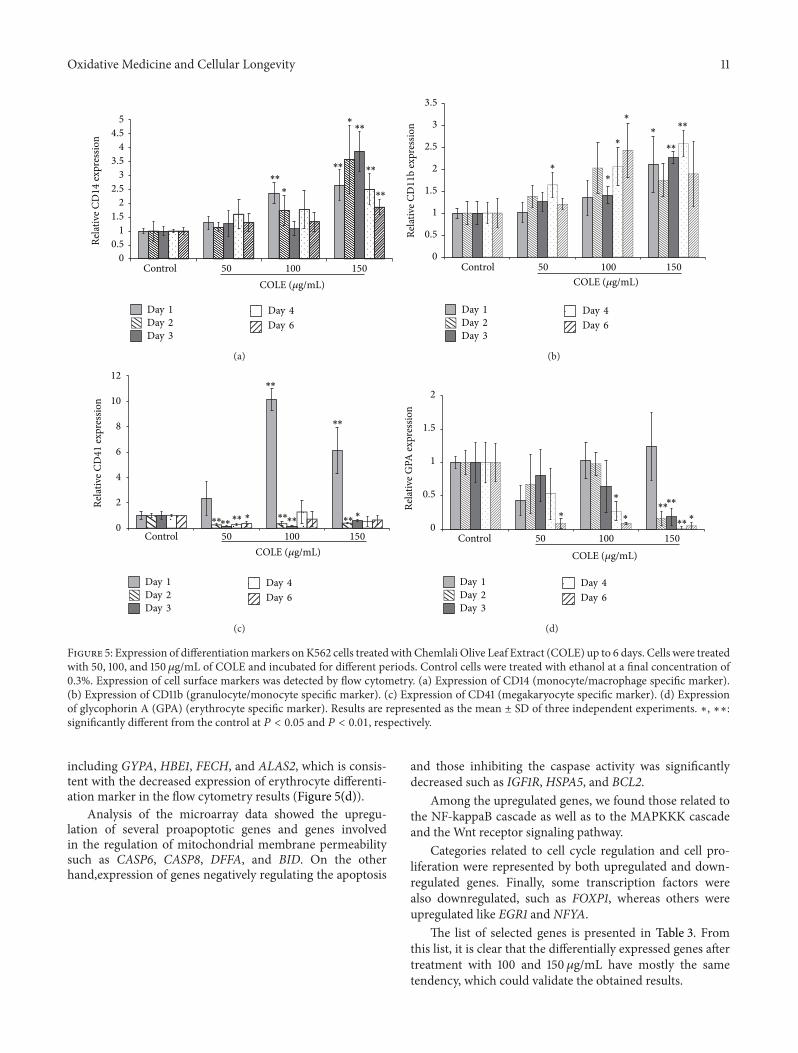
36 Impact of COLE on the Differentiation Capacity of K562Cells The reduction in cell growth as well as the morpho-logical changes observed in COLE treated cells led us tothink about the differentiation assessment K562 cells arepluripotent malignant cells that spontaneously differentiatealong erythroid macrophage and megakaryocyte lineages[22] Treated cells were harvested and analyzed for theexpression of lineage differentiation markers The analysiswas performed by flow cytometry at different days on themonocytemacrophage marker CD14 on the granulocytemarker CD11b on the erythrocyte marker GPA and on themegakaryocyte marker CD41 The results showed a signifi-cant increase in the expression of CD14 marker from day 1until day 6 in the cells treated with 150120583gmL suggestingthe commitment of K562 cells to the monocytemacrophagelineage (Figure 5(a)) A slight increase of the expression ofCD11b was also detected at this concentration (Figure 5(b))Treatment with 100 120583gmL showed a gain in the expression ofCD14 on the 1st and 2nd day and an augmentation of CD11bexpression from the 3rd day of treatment Treatment with50120583gmL did not show any significant effect on the differenti-ationmarkersCD14 andCD11b Even after 6 days of treatmentwith 150 120583gmL of COLE we could not detect the erythroidmarker at all the tested concentrations (Figure 5(d)) How-ever an instant increase in the megakaryocytic marker wasdetected on the 1st day of treatment followed by a drasticdecrease lower than the control during the following days(Figure 5(c))
37 Gene Expression Profile of COLE Treated K562 Cells Tofurther elucidate the mechanism by which COLE inducesapoptosis and differentiation of K562 cells we investigatedthe changes in gene expression profiles in treated cells usingHG219 GeneChip array Microarray analysis was performedon K562 cells at the 3rd day of treatment based on the obser-vation that such timing generated a significant expression ofthe differentiation markers Genes with more than 15-foldchange in expression levels between control and 150 120583gmLofCOLE treated cells were classified into categories accordingto the cell biological processes According to the GO analysis
0
10
20
30
40
50
60
70
Day 1 Day 2 Day 3 Day 4 Day 6
Ann
exin
pos
itive
(
)
Control50120583gmL
100 120583gmL150120583gmL
lowastlowast lowastlowast lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
Figure 4 Induction of apoptosis in K562 cells treatedwith ChemlaliOlive Leaf Extract (COLE) Cells were treated with 50 100 and150 120583gmL of COLE and incubated for different time At the indi-cated time K562 cells were stained with Annexin V and analyzed byflow cytometry Control represents cells treated with 03 ethanolin medium Results are represented as the mean plusmn SD of threeindependent experiments lowast lowastlowast significantly different from thecontrol (119875 lt 005 and 119875 lt 001 resp)
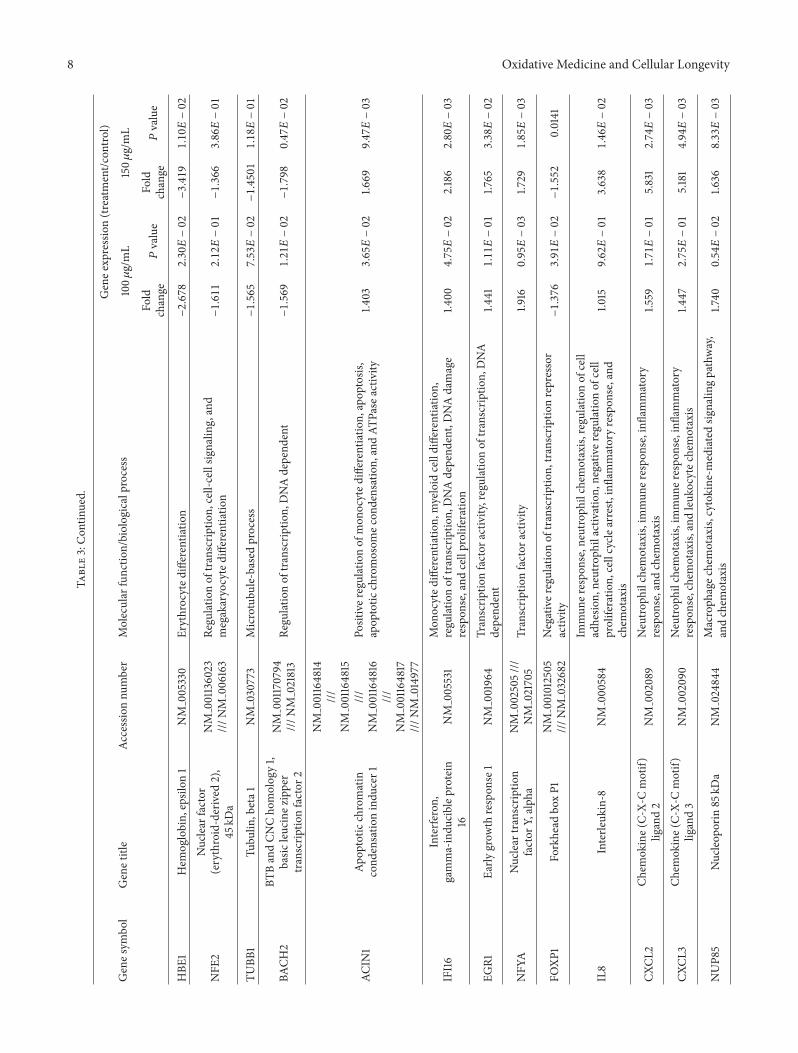
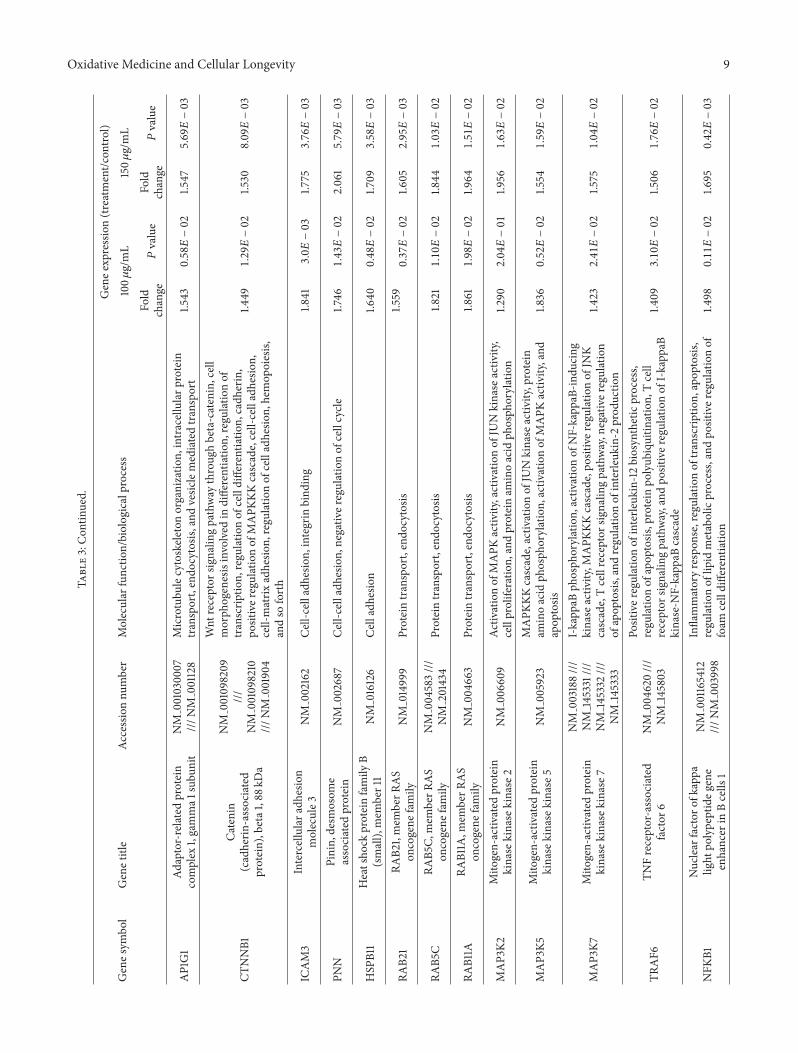
results (Tables S1 and S2 in Supplementary Material avail-able online at httpdxdoiorg1011552014927619) COLEtreatment induced the upregulation of genes involved inhematopoiesis such as CTNNB1 SH2B3 CIAPIN1 RPA1 andgenes implicated in cell differentiation and its regulation suchasMCL1CTNNB1 andCCNE1 Interestingly genes related tomyeloid cell differentiation such as IFI16ACIN1 andCASP8and particularly genes engaged in monocyte differentiationwere represented in the upregulated categories Moreoveran increased expression of genes involved in chemotaxissuch as NUP85 HRAS IL8 CXCL2 and CXCL3 and genesrelated to cytokine production was also detected Amongupregulated genes AP1G1 RABEP1 RAB5C RAB21 EPS15L1and CORO1C were also found involved in the process ofendocytosis and phagocytosis as well as genes related toprotein transport and genes implicated in cell adhesion suchas ICAM3 HSPB11 and PNN
Conversely the erythrocyte differentiation categorieswere represented in the list of the decreased genes (Table S2)
Figure 5 Expression of differentiationmarkers onK562 cells treatedwith Chemlali Olive Leaf Extract (COLE) up to 6 days Cells were treatedwith 50 100 and 150 120583gmL of COLE and incubated for different periods Control cells were treated with ethanol at a final concentration of03 Expression of cell surface markers was detected by flow cytometry (a) Expression of CD14 (monocytemacrophage specific marker)(b) Expression of CD11b (granulocytemonocyte specific marker) (c) Expression of CD41 (megakaryocyte specific marker) (d) Expressionof glycophorin A (GPA) (erythrocyte specific marker) Results are represented as the mean plusmn SD of three independent experiments lowast lowastlowastsignificantly different from the control at 119875 lt 005 and 119875 lt 001 respectively
including GYPA HBE1 FECH and ALAS2 which is consis-tent with the decreased expression of erythrocyte differenti-ation marker in the flow cytometry results (Figure 5(d))
Analysis of the microarray data showed the upregu-lation of several proapoptotic genes and genes involvedin the regulation of mitochondrial membrane permeabilitysuch as CASP6 CASP8 DFFA and BID On the otherhandexpression of genes negatively regulating the apoptosis
and those inhibiting the caspase activity was significantlydecreased such as IGF1R HSPA5 and BCL2
Among the upregulated genes we found those related tothe NF-kappaB cascade as well as to the MAPKKK cascadeand the Wnt receptor signaling pathway
Categories related to cell cycle regulation and cell pro-liferation were represented by both upregulated and down-regulated genes Finally some transcription factors werealso downregulated such as FOXP1 whereas others wereupregulated like EGR1 and NFYA
The list of selected genes is presented in Table 3 Fromthis list it is clear that the differentially expressed genes aftertreatment with 100 and 150120583gmL have mostly the sametendency which could validate the obtained results
12 Oxidative Medicine and Cellular Longevity
4 Discussion
Previous reports had indicated that olive leaf extract inducesapoptosis in the human leukemic cell lines HL-60 and Jurkatcells [13 21] In this study we give evidence that COLEexhibits its antileukemia effect by both inducing apoptosisand promoting differentiation of the multipotent humanleukemia K562 cells
Induction of differentiation as well as apoptosis has beenfrequently reported to be associatedwith a loss of proliferativecapacity of the cells It has been reported that olive leaf extractinhibits the proliferation of some cancer cell lines such asB16 melanoma cells HL-60 leukemia cells the breast cancerMCF-7 cells and the glioblastoma cells [12 25ndash27] Ourresults indicate that COLE inhibits the proliferation of K562cells in a dose dependent manner (Figure 2(a)) Howeverdespite the drastic decrease in cell proliferation the viabilityof cells treated with the highest concentration of COLE(150 120583gmL) remains relatively high (80) for up to 3 daysof treatment (Figure 3(b))
Since it is well established that the arrest of the cellcycle progression allows the cells to follow other processessuch as apoptosis and differentiation we then explored theeffect of COLE on the cell cycle distribution (Table 2) Oliveleaf extract has been shown to cause cell cycle arrest atG0G1 phase in both the breast cancer MCF-7 cells and themelanoma B16 cells [12 25] However the treatment timesapplied in both of these studies were limited to 48 h and 18 hrespectively In this study COLE treated cells were arrested atG0G1 on the 1st and 2ndday of treatment which is consistentwith the reported studies Prolonged incubation with COLEshowed an arrest of cell cycle at G2M phase (3rd and 4thday of incubation) The microarray analysis performed forthe 3rd day of treatment revealed a significant increase ofCHEK2 gene expression in COLE treated cells The encodedprotein regulates the cell cycle checkpoint arrest throughthe inhibition of the activity of CDC25A CDC25B andCDC25C [28] CDC25 proteins trigger the entry into mitosisat different points of the cell cycle by activating theCdk-cyclincomplexes CDC25A acts early in the cell cycle regulatingthe G1S transition whereas CDC25B and CDC25C act atG2M [29] Interestingly our results showed a decrease in theexpression of CDC25C gene in COLE treated cells associatedwith an increase in the expression of CDC25A which mayexplain the cell cycle arrest at G2M phase observed at the3rd day of treatment with COLE
Treatment with COLE increased the expression of CD14on the cell surface of treated cells indicating the differenti-ation of K562 cells toward the mono-macrophage lineage(Figure 5(a)) The increase in the percentage of cells positivefor CD11b (Figure 5(b)) confirms this hypothesis In factCD11b is expressed on both monocytes and granulocytesThus together with the increase of CD14 expression theexpression of CD11b suggests that treatment with COLEpromotes the commitment of K562 cells into the mono-cytemacrophage lineage at the expense of the granulocyticdifferentiationThe instant increase of the expression of CD41
on the 1st day of treatment and its absence during the fol-lowing days (Figure 5(c)) could be consistent with monocytedifferentiation instead of megakaryocyte differentiation sinceit has been reported that 120572IIb promoter which initiates thetranscription of CD41 is transcriptionally active in pluripo-tent myeloid progenitors in early stages of erythropoiesisand all along the megakaryocytic differentiation as well asto a lesser extent in the early stages of myelomonocyticdifferentiation and the late stages of erythropoiesis [30] Thisinstant increase could be also explained by the phenomenonof lineage conversion where some hematopoietic progenitorscan be converted into lineages other than their own bythe ectopic expression of some transcription factors [31]In this regard it was previously reported that commonlymphoid progenitors megakaryocyte erythroid progenitorsearly B cells and early T cells could be converted to thegranulocytemonocyte lineage by the induction of CEBP120572activity [32] Microarray results support the absence ofmegakaryocyte differentiation in the 3rd day of treatment Infact data showed a decrease in the expression of genes relatedtomegakaryopoiesis such as the essential regulator of plateletrelease NFE2 and its main candidate target gene TUBB1(encoding the megakaryocyte and platelet specific isoform of120573-tubulin (1205731)) [33] as well as the BACH2 transcription factorreported to be induced during megakaryocyte differentiation[34]
Interestingly results of microarray analysis (Table 3)showed the upregulation of several genes already reportedas key players in monocytemacrophage differentiation inthe COLE treated cells Among these genes the expressionof IFI16 was markedly increased Constitutively expressedin lymphoid cells IFI16 was shown to be associated withthe differentiation of human myeloid cells in response tointerferon gamma [35] Moreover IFI16 has been selectivelyexpressed during the differentiation of myeloid progenitorcells along monocytic lineage while it is downregulatedduring the commitment toward the granulocytic or erythroidlineage suggesting its potential involvement in the differenti-ation andmaturation of the monocytic lineage [36 37] Geneencoding for EGR-1 a positive modulator of macrophagedifferentiation was also significantly upregulated EGR-1 isknown to dictate development of myeloid progenitors alongthe macrophage lineage at the expense of development alongother lineages [38 39] Dauffy et al [40] reported thatIFI16-enforced expression in myeloid progenitors inducedthe expression of EGR-1 and conducted the macrophagedifferentiation in the absence of the macrophage colonystimulating factor (M-CSF) This suggests that EGR-1 couldbe regulated by the expression of IFI16 Treatment with COLEincreased as well the expression of NFYA encoding the Asubunit of the nuclear transcription factor Y whose synthesiswas previously reported during the terminal differentiationof monocytes to macrophages [38 41] The downregula-tion of FOXP1 in COLE treated K562 cells is consistentwith monocytemacrophage differentiation since it has beendemonstrated that the expression of FOXP1 was markedlydecreased in monocyte-induced differentiation of HL-60cells as well as in human peripheral blood monocytes and
Oxidative Medicine and Cellular Longevity 13
that the overexpression of FOXP1 prevented the morphologicmacrophage-like differentiation [42 43]
Monocytes and macrophages have several functionalcharacteristics including cell adhesion migration chemo-taxis and phagocytic activity Interestingly an enhancedexpression of the chemokines genes CXCL8 (IL8) CXCL2andCXCL3was found in COLE treated cells IL-8 productionhas been observed in vitro in a wide variety of cells includingmonocytes T lymphocytes neutrophils vascular endothe-lial cells dermal fibroblasts keratinocytes hepatocytes andhuman gastric cancer cells while the production of CXCL2andCXCL3was only described inmonocytes fibroblasts andendothelial cells [44 45] Another gene related to chemotaxiswas upregulated NUP85 which is an essential component ofthe nuclear pore complex and was reported to be involved inCCR2-mediated chemotaxis of monocytes [46]
COLE treatment also enhanced the expression of somegenes related to phagocytosis AP1G1 andRab proteins relatedgenes AP1G1 is a subunit of clathrin-associated adaptorprotein complex 1 whose role has been recently highlightedfor efficient phagocytosis at an early stage of phagosomeformation since it participates in the extension of the phago-cytic cup [47 48] The small GTPases Rab are key regulatorsof intracellular membrane trafficking Recent studies havedemonstrated that several RAB proteins play an importantrole in phagocytosis RAB proteins such as RAB11 Rab5 andRab 21 participate in the phagosome formation and matura-tion and are necessary for the phagocytic activity [48ndash51]
Consistently with the increase in Annexin V positivecells (Figure 4) treatment with COLE upregulated someproapoptotic genes such as CASP6 CASP8 DFFA and BIDand downregulated the apoptosis suppressor BCL2 as wellas the caspase inhibitors genes (Table 3) Olive leaf waspreviously shown to induce morphological changes thatare characteristic of apoptosis in leukemia cells but themechanisms underlying this effect were not well investi-gated [13 21] The increase in the proportion of apoptoticcells observed from the 4th day of treatment with COLEcould be explained by the programmed death of the fullydifferentiated cells (Figure 4) In fact this increase wasaccompanied by an important decrease in cell viability aswell as a diminution in CD14 expression (Figures 3(b) and5(a)) Elsewhere it is well established that in hematopoietictissues apoptosis is coupled to terminal differentiation ofmyeloid progenitors even though themechanism responsiblefor the activation of apoptosis during myeloid maturationis still poorly understood [52 53] It is also noteworthy tomention that several reports have highlighted the role ofcaspases in the terminal differentiation of a variety of celltypes [54] One example is the differentiation of human bloodmonocytes into macrophages a process that is blocked bysynthetic caspase inhibitors [55] It has been also reportedthat caspase 8 deletion in bone-marrow cells resulted in arrestof hemopoietic progenitor functioning and that its deletionin cells of the myelomonocytic lineage led to the arrest ofdifferentiation into macrophages and consequently to celldeath [56]
Among the differentially expressed genes in COLEtreated cells (Table 3) genes encoding the mitogen-activated
protein kinase kinase kinases (MAPKKKs) MAP3K2MAP3K5 and MAP3K7 were highly expressed whileMAP2K5 and MAPK14p38-120572 were downregulated TheMAPKKKs act at the upstream of the MAP kinase cascadewhich regulates important cellular processes such as geneexpression cell proliferation differentiation cell survivaland death The MAP3K5 activates the c-Jun N-terminalkinase (JNK) and p38 mitogen-activated protein kinases[57] while the MAP3K2 is reported to preferentially activateJNK [58 59] However the significant decrease in theMAPK14p38-120572 may suggest that the effect of COLE onK562 cells is likely to be mediated by the JNK MAPKsrather than the p38 MAPKs JNK plays an important rolein apoptosis pathways weather by activating proapoptoticor antiapoptotic genes Particularly it was shown thatprolonged but not transient JNK activation promotes thecell death [60] Our results showed that JKAMP gene whichencodes for a membrane-anchored regulator of the durationof JNK1 activity was upregulated (154 plusmn 0051) The elevatedexpression of JKAMP as reported by Kadoya et al [61]results in sustained JNK activity These findings may indicatethe potential role of JNK pathway in the COLE-inducedapoptosis in K562 cells
Thanks to their large regulatory domains some MAP-KKKs can interact with upstream regulators have functionssuch as ubiquitylation and be activated by relief of autoin-hibition and oligomerization [62] In this context MAP3K2has been reported to play an important role in NF-kappaBsignaling pathway by activating the I-kappaB kinases whichin turn phosphorylate the inhibitory factors of the nuclearfactor kappaB (NF-kappaB) resulting in their rapid ubiq-uitination and the liberation of NF-kappaB complex whichtranslocate from the cytoplasm to the nucleus [63] MAP3K7in association with other proteins could also be required forthe activation of NF-kappaB According to the GO analysis(Table S1) COLE treatment upregulated genes implicatedin NF-kappaB-related categories such as positive regulationof NF-kappaB transcription factor activity I-kappaB kinase-NF-kappaB cascade and its positive regulation and I-kappaBphosphorylation Genes encoding the signal transducers inthe NF-kappaB pathway TRAF5 and TRAF6 as well asthe activator of NF-kappaB signaling SNIP1 were upregu-lated Interestingly an increased expression of NFKB1 geneencoding for the precursor of the mature NF-kappaB p50was detected in COLE treated cells compared to untreatedK562 cells NFKB is well recognized as a central activator ofthe antiapoptotic cascades in response to external stimuli orintrinsic immune reactions and its prosurvival activity hasbeen implicated in a variety of biological processes [60 64]In the B cell lineage the activity of NF-120581B is required forthe completion of various developmental stages includingdifferentiation and the response of these cells to antigens [65]NF-kappaB also plays an important role in the survival anddevelopment of T cells [66] and has been recently reported tomediate the differentiation of mesenchymal stem cells [67]
Several studies have demonstrated a crosstalk betweenthe NF-kappaB and JNK pathways seen as an inhibitoryeffect of NF-kappaB on apoptosis through the suppressionof JNK activity and that this suppressive effect may occur
14 Oxidative Medicine and Cellular Longevity
through different mechanisms [60] This suggests that thebalance between JNK and NF-kappaB activities is crucial todetermine the cell fate survival or deathWehypothesize herethat COLE-induced apoptosis or differentiation is likely to bedetermined by the crosstalk between these two pathways inK562 cells Further investigations regarding protein expres-sion and activity are needed to confirm these findings
5 Conclusions
Our study demonstrates for the first time that olive leafextract exhibits an antileukemia effect on the human chronicmyeloid leukemia cells Olive leaf extract was shown toinhibit the proliferation of K562 cells by inducing cell cyclearrest apoptosis and differentiation toward the monocytelineage The induced expression of molecules involved indifferentiation toward the monocytemacrophage lineage aswell asmolecules related to apoptosis and cell cycle regulationconfirmed these findings and provides insights into themechanism by which olive leaf exhibits its antileukemiaeffect
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported by the JICA-JST Science andTechnology Research Partnership for Sustainable Develop-ment (SATREPS) Project ldquoValorization of Bio-Resources inSemi-Arid and Arid Land for Regional Developmentrdquo
References
[1] M Leszczyniecka T Roberts P Dent S Grant and P B FisherldquoDifferentiation therapy of human cancer basic science andclinical applicationsrdquo Pharmacology and Therapeutics vol 90no 2-3 pp 105ndash156 2001
[2] A I Spira and M A Carducci ldquoDifferentiation therapyrdquoCurrent Opinion in Pharmacology vol 3 no 4 pp 338ndash3432003
[3] S Nobili D Lippi E Witort et al ldquoNatural compounds forcancer treatment and preventionrdquo Pharmacological Researchvol 59 no 6 pp 365ndash378 2009
[4] S Ramos ldquoEffects of dietary flavonoids on apoptotic pathwaysrelated to cancer chemopreventionrdquo Journal of NutritionalBiochemistry vol 18 no 7 pp 427ndash442 2007
[5] V Hajhashemi G Vaseghi M Pourfarzam and A AbdollahildquoAre antioxidants helpful for disease preventionrdquo Research inPharmaceutical Sciences vol 5 no 1 pp 1ndash8 2010
[6] M Bouaziz and S Sayadi ldquoIsolation and evaluation of antiox-idants from leaves of a Tunisian cultivar olive treerdquo EuropeanJournal of Lipid Science andTechnology vol 107 no 7-8 pp 497ndash504 2005
[7] A P Pereira I C F R Ferreira F Marcelino et al ldquoPhenoliccompounds and antimicrobial activity of olive (Olea europaeaL Cv Cobrancosa) leavesrdquo Molecules vol 12 no 5 pp 1153ndash1162 2007
[8] L I Somova F O Shode P Ramnanan and A Nadar ldquoAnti-hypertensive antiatherosclerotic and antioxidant activity oftriterpenoids isolated from Olea europaea subspecies africanaleavesrdquo Journal of Ethnopharmacology vol 84 no 2-3 pp 299ndash305 2003
[9] V Micol N Caturla L Perez-Fons V Mas L Perez andA Estepa ldquoThe olive leaf extract exhibits antiviral activityagainst viral haemorrhagic septicaemia rhabdovirus (VHSV)rdquoAntiviral Research vol 66 no 2-3 pp 129ndash136 2005
[10] J Wainstein T Ganz M Boaz et al ldquoOlive leaf extract as ahypoglycemic agent in both human diabetic subjects and inratsrdquo Journal of Medicinal Food vol 15 no 7 pp 605ndash610 2012
[11] L Seddik TM Bah A AouesM Slimani andM BenderdourldquoElucidation of mechanisms underlying the protective effects ofolive leaf extract against lead-induced neurotoxicity in Wistarratsrdquo Journal of Toxicological Sciences vol 36 no 6 pp 797ndash809 2011
[12] Z Bouallagui J Han H Isoda and S Sayadi ldquoHydroxytyrosolrich extract from olive leaves modulates cell cycle progressionin MCF-7 human breast cancer cellsrdquo Food and ChemicalToxicology vol 49 no 1 pp 179ndash184 2011
[13] R Fares S Bazzi S E Baydoun and R M Abdel-Massih ldquoTheantioxidant and anti-proliferative activity of the Lebanese Oleaeuropaea extractrdquo Plant Foods for Human Nutrition vol 66 no1 pp 58ndash63 2011
[14] V Goulas V Exarchou A N Troganis et al ldquoPhytochemicalsin olive-leaf extracts and their antiproliferative activity againstcancer and endothelial cellsrdquo Molecular Nutrition and FoodResearch vol 53 no 5 pp 600ndash608 2009
[15] M de Bock E B Thorstensen J G Derraik H V HendersonP L Hofman and W S Cutfield ldquoHuman absorption andmetabolism of oleuropein and hydroxytyrosol ingested as olive(Olea europaea L) leaf extractrdquo Molecular Nutrition and FoodResearch vol 57 no 11 pp 2079ndash2085 2013
[16] R Briante M Patumi S Terenziani E Bismuto F Febbraioand R Nucci ldquoOlea europaea L leaf extract and derivativesantioxidant propertiesrdquo Journal of Agricultural and Food Chem-istry vol 50 no 17 pp 4934ndash4940 2002
[17] O Benavente-Garcıa J Castillo J Lorente A Ortuno and J ADel Rio ldquoAntioxidant activity of phenolics extracted from Oleaeuropaea L leavesrdquo Food Chemistry vol 68 no 4 pp 457ndash4622000
[18] S H Omar ldquoOleuropein in olive and its pharmacologicaleffectsrdquo Scientia Pharmaceutica vol 78 no 2 pp 133ndash154 2010
[19] H Jemai M Bouaziz I Fki A El Feki and S SayadildquoHypolipidimic and antioxidant activities of oleuropein and itshydrolysis derivative-rich extracts from Chemlali olive leavesrdquoChemico-Biological Interactions vol 176 no 2-3 pp 88ndash982008
[20] R Garcıa-Villalba M Larrosa S Possemiers F A Tomas-Barberan and J C Espın ldquoBioavailability of phenolics from anoleuropein-rich olive (Olea europaea) leaf extract and its acuteeffect on plasma antioxidant status comparison between pre-and postmenopausal womenrdquo European Journal of Nutrition2013
[21] J Anter Z Fernandez-Bedmar M Villatoro-Pulido et al ldquoApilot study on the DNA-protective cytotoxic and apoptosis-inducing properties of olive-leaf extractsrdquo Mutation Researchvol 723 no 2 pp 165ndash170 2011
[22] J A Sutherland A R Turner and P Mannoni ldquoDifferen-tiation of K562 leukemia cells along erythroid macrophage
Oxidative Medicine and Cellular Longevity 15
and megakaryocyte lineagesrdquo Journal of Biological ResponseModifiers vol 5 no 3 pp 250ndash262 1986
[23] K Schwenke H-P Peterson K-H Wangenheim and L EFeinendegen ldquoInduction of differentiation in erythroleukemicK562 cells by 120574-irradiationrdquo Leukemia Research vol 19 no 12pp 955ndash961 1995
[24] V N Enujiugha J Y Talabi S A Malomo and A I OlagunjuldquoDPPH radical scavenging capacity of phenolic extracts fromAfrican Yam Bean (Sphenostylis stenocarpa)rdquo Food and Nutri-tion Sciences vol 3 pp 7ndash13 2012
[25] S A Mijatovic G S Timotijevic D M Miljkovic et alldquoMultiple antimelanoma potential of dry olive leaf extractrdquoInternational Journal of Cancer vol 128 no 8 pp 1955ndash19652011
[26] L Abaza T P N Talorete P Yamada Y KuritaM Zarrouk andH Isoda ldquoInduction of growth inhibition and differentiation ofhuman leukemia HL-60 cells by a Tunisian Gerboui olive leafextractrdquo Bioscience Biotechnology and Biochemistry vol 71 no5 pp 1306ndash1312 2007
[27] B Tunca G Tezcan G Cecener et al ldquoOlea europea leafextract alters microRNA expression in human glioblastomacellsrdquo Journal of Cancer Research and Clinical Oncology vol 138no 11 pp 1831ndash1844 2012
[28] S Matsuoka M Huang and S J Elledge ldquoLinkage of ATM tocell cycle regulation by the Chk2 protein kinaserdquo Science vol282 no 5395 pp 1893ndash1897 1998
[29] C Frazer and P G Young ldquoPhosphorylation mediated regu-lation of Cdc25 activity localization and stabilityrdquo in ProteinPhosphorylation in Human Health C Huang Ed pp 395ndash436InTech 2012
[30] N Debili C Robin V Schiavon et al ldquoDifferent expression ofCD41 on human lymphoid andmyeloid progenitors from adultsand neonatesrdquo Blood vol 97 no 7 pp 2023ndash2030 2001
[31] H Nakajima ldquoRole of transcription factors in differentiationand reprogramming of hematopoietic cellsrdquo Keio Journal ofMedicine vol 60 no 2 pp 47ndash55 2011
[32] Y Fukuchi F Shibata M Ito et al ldquoComprehensive analysis ofmyeloid lineage conversion using mice expressing an inducibleform of CEBP120572rdquoThe EMBO Journal vol 25 no 14 pp 3398ndash3410 2006
[33] R A Shivdasani ldquoMolecular and transcriptional regulation ofmegakaryocyte differentiationrdquo Stem Cells vol 19 no 5 pp397ndash407 2001
[34] K Terui Y Takahashi J Kitazawa T TokiM Yokoyama and EIto ldquoExpression of transcription factors during megakaryocyticdifferentiation of CD34+ cells from human cord blood inducedby thrombopoietinrdquo Tohoku Journal of Experimental Medicinevol 192 no 4 pp 259ndash273 2000
[35] J A Trapani M Dawson V A Apostolidis and K A BrowneldquoGenomic organization of IFI16 an interferon-inducible genewhose expression is associated with human myeloid cell differ-entiation correlation of predicted protein domains with exonorganizationrdquo Immunogenetics vol 40 no 6 pp 415ndash424 1994
[36] M J Dawson N J Elwood R W Johnstone and J A TrapanildquoThe IFN-inducible nucleoprotein IFI 16 is expressed in cellsof the monocyte lineage but is rapidly and markedly down-regulated in other myeloid precursor populationsrdquo Journal ofLeukocyte Biology vol 64 no 4 pp 546ndash554 1998
[37] M Gariglio M De Andrea M Lembo et al ldquoThe murinehomolog of the HIN 200 family Ifi 204 is constitutivelyexpressed in myeloid cells and selectively induced in the
monocytemacrophage lineagerdquo Journal of Leukocyte Biologyvol 64 no 5 pp 608ndash614 1998
[38] A F Valledor F E Borras M Cullell-Young and A CeladaldquoTranscription factors that regulate monocytemacrophage dif-ferentiationrdquo Journal of Leukocyte Biology vol 63 no 4 pp405ndash417 1998
[39] K Krishnaraju B Hoffman and D A Liebermann ldquoEarlygrowth response gene 1 stimulates development of hematopoi-etic progenitor cells along the macrophage lineage at theexpense of the granulocyte and erythroid lineagesrdquo Blood vol97 no 5 pp 1298ndash1305 2001
[40] J Dauffy G Mouchiroud and R P Bourette ldquoThe interferon-inducible gene Ifi204 is transcriptionally activated in responseto M-CSF and its expression favors macrophage differentiationin myeloid progenitor cellsrdquo Journal of Leukocyte Biology vol79 no 1 pp 173ndash183 2006
[41] G Marziali E Perrotti R Ilari U Testa E M Coccia andA Battistini ldquoTranscriptional regulation of the ferritin heavy-chain gene the activity of the CCAAT binding factor NF-Y ismodulated in heme-treated Friend leukemia cells and duringmonocyte-to-macrophage differentiationrdquo Molecular and Cel-lular Biology vol 17 no 3 pp 1387ndash1395 1997
[42] C Shi X Zhang Z Chen et al ldquoIntegrin engagement regulatesmonocyte differentiation through the forkhead transcriptionfactor Foxp1rdquo Journal of Clinical Investigation vol 114 no 3 pp408ndash418 2004
[43] C Shi M Sakuma T Mooroka et al ldquoDown-regulation of theforkhead transcription factor Foxp1 is required for monocytedifferentiation andmacrophage functionrdquoBlood vol 112 no 12pp 4699ndash4711 2008
[44] D F Smith E Galkina K Ley and Y Huo ldquoGRO familychemokines are specialized for monocyte arrest from flowrdquoThe American Journal of Physiology Heart and CirculatoryPhysiology vol 289 no 5 pp H1976ndashH1984 2005
[45] J J Oppenheim O M Z Howard and E Goetzl ChemotacticFactors Neuropeptides and Other Ligands for Seven Transmem-brane Receptors 2000
[46] Y Terashima N Onai M Murai et al ldquoPivotal function forcytoplasmic protein FROUNT in CCR2-mediated monocytechemotaxisrdquo Nature Immunology vol 6 no 8 pp 827ndash8352005
[47] Y Lefkir M Malbouyres D Gotthardt et al ldquoInvolvement ofthe AP-1 adaptor complex in early steps of phagocytosis andmacropinocytosisrdquo Molecular Biology of the Cell vol 15 no 2pp 861ndash869 2004
[48] F Niedergang and P Chavrier ldquoSignaling and membranedynamics during phagocytosis many roads lead to the phago-somerdquo Current Opinion in Cell Biology vol 16 no 4 pp 422ndash428 2004
[49] D Cox D J Lee B M Dale J Calafat and S Green-berg ldquoA Rab11-containing rapidly recycling compartment inmacrophages that promotes phagocytosisrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 97 no 2 pp 680ndash685 2000
[50] O V Vieira C Bucci R E Harrison et al ldquoModulation of Rab5and Rab7 recruitment to phagosomes by phosphatidylinositol3-kinaserdquoMolecular andCellular Biology vol 23 no 7 pp 2501ndash2514 2003
[51] T Khurana J A Brzostowski and A R Kimmel ldquoARab21LIM-onlyCH-LIM complex regulates phagocytosis viaboth activating and inhibitory mechanismsrdquo The EMBO Jour-nal vol 24 no 13 pp 2254ndash2264 2005
16 Oxidative Medicine and Cellular Longevity
[52] S J Martin J G Bradley and T G Cotter ldquoHL-60 cellsinduced to differentiate towards neutrophils subsequently dievia apoptosisrdquo Clinical and Experimental Immunology vol 79no 3 pp 448ndash453 1990
[53] A Benito D Grillot G Nunez and J L Fernandez-Luna ldquoReg-ulation and function of Bcl-2 during differentiation-inducedcell death in HL-60 promyelocytic cellsrdquoThe American Journalof Pathology vol 146 no 2 pp 481ndash490 1995
[54] M Lamkanfi N Festjens W Declercq T V Berghe andP Vandenabeele ldquoCaspases in cell survival proliferation anddifferentiationrdquo Cell Death and Differentiation vol 14 no 1 pp44ndash55 2007
[55] O Sordet C Rebe S Plenchette et al ldquoSpecific involvement ofcaspases in the differentiation of monocytes into macrophagesrdquoBlood vol 100 no 13 pp 4446ndash4453 2002
[56] T-B Kang T Ben-Moshe E E Varfolomeev et al ldquoCaspase-8 serves both apoptotic and nonapoptotic rolesrdquo Journal ofImmunology vol 173 no 5 pp 2976ndash2984 2004
[57] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[58] J Cheng J Yang Y Xia M Karin and B Su ldquoSynergisticinteraction of MEK kinase 2 c-Jun N-terminal kinase (JNK)kinase 2 and JNK1 results in efficient and specific JNK1activationrdquo Molecular and Cellular Biology vol 20 no 7 pp2334ndash2342 2000
[59] J L Blank P Gerwins EM Elliott S Sather andG L JohnsonldquoMolecular cloning of mitogen-activated proteinERK kinasekinases (MEKK) 2 and 3 regulation of sequential phosphoryla-tion pathways involving mitogen-activated protein kinase andc-Jun kinaserdquo Journal of Biological Chemistry vol 271 no 10pp 5361ndash5368 1996
[60] S Papa F Zazzeroni C G Pham C Bubici and G FranzosoldquoLinking JNK signaling to NF-120581B a key to survivalrdquo Journal ofCell Science vol 117 no 22 pp 5197ndash5208 2004
[61] T Kadoya A Khurana M Tcherpakov et al ldquoJAMP a JUB N-terminal kinase 1 (JNK1)-associated membrane protein regu-lates duration of JNK activityrdquo Molecular and Cellular Biologyvol 25 no 19 pp 8619ndash8630 2005
[62] M Qi and E A Elion ldquoMAP kinase pathwaysrdquo Journal of CellScience vol 118 no 16 pp 3569ndash3572 2005
[63] Q Zhao and F S Lee ldquoMitogen-activated protein kinaseERKkinase kinases 2 and 3 activate nuclear factor-120581B through I120581Bkinase-120572 and I120581B kinase-120573rdquoThe Journal of Biological Chemistryvol 274 no 13 pp 8355ndash8358 1999
[64] H Namba V Saenko and S Yamashita ldquoNuclear factor-120581Bin thyroid carcinogenesis and progression a novel therapeutictarget for advanced thyroid cancerrdquo Arquivos Brasileiros deEndocrinologia e Metabologia vol 51 no 5 pp 843ndash851 2007
[65] B Feng S Cheng W S Pear and H-C Liou ldquoNF-kBinhibitor blocks B cell development at two checkpointsrdquoMedi-cal Immunology vol 3 article 1 2004
[66] S Gerondakis and U Siebenlist ldquoRoles of the NF-kappaBpathway in lymphocyte development and functionrdquoCold SpringHarbor Perspectives in Biology vol 2 no 5 Article ID a0001822010
[67] K P Bhat V Balasubramaniyan B Vaillant et al ldquoMesenchy-mal differentiation mediated by NF-kB promotes radiationresistance in glioblastomardquo Cancer Cell vol 24 no 3 pp 331ndash346 2013
gained the rising interest of the scientific and industrialcommunity due to its proved beneficial health propertiesand thus has emerged as commercially valuable nutraceu-ticals [15] The primary constituents which are believed tocontribute to the health benefits of olive leaves are Oleu-ropein Hydroxytyrosol as well as several other flavonoidssuch as Verbascoside Apigenin-7-glucoside and Luteolin-7-glucoside [14 16] Oleuropein the major constituent ofolive leaves has been shown to be a potent antioxidantIts radical scavenging activity has been well documented[6 17] Oleuropein has been shown to inhibit the oxida-tion of low density lipoproteins in vitro and in vivo [18]Jemai et al have demonstrated that polyphenols recoveredfrom olive leaf extracts Oleuropein Hydroxytyrosol andOleuropein aglycone exhibited a pronounced hypolipidemiceffect reduced the lipid peroxidation process and enhancedthe antioxidant defense system in experimental atherogenicmodel [19] Benavente-Garcıa et al [17] studied the antiox-idant activity of phenolic compounds from olive leaves andconcluded that olive phenolsmay exhibit synergistic behaviorin their radical scavenging capacity when mixed in thesame proportions as occur in the olive leaf extract Tworecent studies have focused on the bioavailability of oliveleaf phenolic compounds in human subjects and have cometo the conclusion that Oleuropein is rapidly absorbed andmetabolized to be mainly excreted as glucuronidated andsulfated Hydroxytyrosol suggesting that olive leaf extractcould exert benefits against oxidative stress-related processesin vivo [15 20]
In the prior studies olive leaf extract has been shown toexhibit an antitumor activity and to induce apoptosis path-ways in cancer cells little attention has been paid to its effecton the process of cancer cell differentiation Particularly oliveleaf has been reported to exhibit an antileukemia effect byinducing apoptosis in the acutemyeloid leukemiaHL-60 cells[13 21]
In this study we investigate the effect of olive leaf of themost abundant Tunisian variety Chemlali on the humanchronic myeloid leukemia K562 cells Compared to theleukemia cell lines used in previous studies [13 21] K562 cellscan be regarded as stem-like cells thanks to their pluripotency[22] and are known for their strong resistance to chemicalinducers [23]
We speculate that olive leaf extract may have anticarcino-genic property in K562 cells not only by inducing apoptosisbut also by inducing the commitment of leukemia cells to thematuration process in order to progressively give an apparentnormal cell life
2 Materials and Methods
21 Preparation of Olive Leaf Extract Collected olive leavesof Chemlali variety from the region of Sfax Tunisia wereair-dried and ground with a mixer Extraction was carriedby ethanol 70 (110 wv) in darkness for 2 weeks at roomtemperature The mixture was then centrifuged and filteredusing 045120583m filter (Millipore Japan) and stored at minus80∘Cuntil it was used
22 HPLC Analysis Chromatographic analyses were ach-ieved on an Agilent series 1260 HPLC-DAD instrument(Agilent Waldbronn Germany) The instrument includesa quaternary pump an online degasser an autosamplerand a thermostatically controlled column compartmentChromatographic separation was carried out on a ZOR-BAX Eclipse XDB-C18 column serial number USNH027266(46mm ID times 250mm times 35 120583m particle size) The elutionconditions were as follows mobile phase A (01 acetic acidin water) and mobile phase B (100 acetonitrile) flow rate of05mLmin sample injection volume of 10 120583L and operatingtemperature 40∘C The running gradient was as follows 0ndash22min 10ndash50 B 22ndash32min 50ndash100 B 32ndash40min100 B 40ndash44min 100ndash10 B Reequilibration durationlasted 6minThe DAD detector scanned from 190 to 400 nmand the samples were detected at 254 280 and 330 nm
23 Cell Line and Culture Conditions Human chronic leu-kemia cell line K562 was obtained from the Riken Cell Bank(Tsukuba Ibaraki Japan) Cells were cultured in RPMI 1640medium (Gibco) supplemented with 10 heat-inactivatedfetal bovine serum and maintained at 37∘C in a humidifiedincubator with 5 CO
2 The cells were pass-cultured every 3
days and used for experiments after reaching the exponentialgrowth phase
24 Cell Proliferation Assay Cell proliferation wasinvestigated by MTT (3-(45-dimethylthiazol-2-yl)-25-diphenyltetrazolium bromide) assay K562 cells were seededin 96-well plates at 20 times 104 cellsmL After incubation for24 h olive leave extract diluted in medium was added at finalconcentrations of 50 75 100 125 and 150120583gmL Controlcells were treated by ethanol at a final concentration of 03MTT was added after treatment for 24 48 and 72 h and theresulting formazan was completely dissolved by 100120583L of10 sodium dodecyl sulfate (SDS) for 24 h The absorbancewas determined at 570 nm in a multidetection microplatereader (Powerscan HT Dainippon Pharmaceutical NJUSA) Absorbance caused by the ability of the sample toreduce MTT or by its color was corrected using platesprepared in the same conditions in the absence of cells
25 Cell Viability Assay and Cell Morphological Changes Theviability of COLE treated cells was measured using flowcytometry according to the manufacturer instructions K562cells were seeded in 6-well plates at 20 times 104 cellsmL andtreated the following day by 50 100 and 150 120583gmL of COLEdiluted in medium and 03 ethanol in the case of thecontrol After incubation for the indicated time treated cellswere harvested suspended in Guava ViaCount reagent andallowed to be stained for at least 5min in darkness The cellnumber and viability were measured by Guava PCA flowcytometry (Guava Technologies CA USA) Morphologicalchanges were detected by observation under a phase contrastmicroscope (Leica Microsystem)
26 Cell Cycle Analysis The distribution of the cell cyclephases of treated and control cells was analysed by flow
Oxidative Medicine and Cellular Longevity 3
cytometry Briefly 20 times 104 cellsmL of K562 cells wereseeded in 6-well plates and treated by 100 and 150 120583gmLof COLE diluted in medium Control cells were treatedwith ethanol 03 After the desired time of incubationcells were harvested washed twice with PBS and fixed with70 ethanol at 4∘C for more than 12 h The fixed cells werethen centrifuged at 500timesg for 5min and washed with PBStwice Cell cycle reagent (Guava Technologies) was addedand the cells were kept in darkness for 30min at roomtemperature The population of cells in each cell cycle phasewas determined by a Guava PCA flow cytometry accordingto their DNA content
27 Annexin V Assay The induction of apoptosis in treatedcells was determined by measuring the externalization ofphosphatidylserine (PS) to the cell surface by flow cytometryK562 cells were seeded in 6 well plates and treated with 50100 and 150 ugmL of COLE diluted inmedium Control cellswere treated with ethanol 03 When reaching the desiredtreatment time cells were harvested and stained with GuavaNexin ReagentThen cells were incubated for 20min at roomtemperature in the dark and then acquired on theGuava PCAsystem
28 Cell Differentiation Assay Cell differentiation wasassessed by flow cytometry by measuring the expression ofCD11b and CD14 on the surface of K562 cells Cells wereseeded at 20 times 104 cellsmL in 6-well plates and incubatedfor 24 h The COLE diluted in medium was added at finalconcentrations of 50 100 and 150120583gmL and ethanol at03 in the case of control cells After incubation cellswere harvested washed twice with cold PBS and adjustedto the same number 10 times 105 The cells were then labeledwith phycoerythrin conjugated anti-CD14 anti-CD11banti-CD41 and anti-glycophorin A for 30min according tomanufacturerrsquos instructions The stained cells were washedtwice with cold PBS and resuspended in 500120583L PBS formeasurement
29 Total RNA Isolation K562 cells at a concentration of20 times 104 cellsmL were incubated for 24 h and then treatedwith 100 and 150 120583gmL of COLE for 72 h The control cellswere treated with ethanol at final concentration of 03The cells were then collected and washed with PBS DNA-free total RNA was isolated from the cells using Isogenreagent (Nippon Gene Co Tokyo Japan) following themanufacturerrsquos instructions
210 DNA Microarray Analysis Microarray hybridizationprobes were generated from isolated RNA samples Double-stranded cDNA was synthesized from 100 ng of total RNAusing the GeneAtlas 31015840 IVT Express Kit (Affymetrix Inc)Biotin-labeled aRNA was synthesized by in vitro transcrip-tion and purified 10 120583g of purified aRNA was then frag-mented using the GeneAtlas 31015840 IVT Express Kit and washybridized to the Affymetrix HG-U219 (Affymetrix) for 16 hat 45∘CThe chips were washed and stained in the GeneAtlasFluidics Station 400 (Affymetrix) and then imaged in the
Table 1 Retention time and concentration of the main phenoliccompounds present in Chemlali Olive Leaf Extract (COLE)
GeneAtlas Imaging Station (Affymetrix) The Partek Expresssoftware (Affymetrix) served for the data analysis by runningcomparisons of gene expression in treated and control cellsbased on mathematical algorithms The generated data (sig-nificant fold change in gene expression) was then analyzedusing the Pathway Studio Explore 11 software (Affymetrix)
211 Statistical Analysis Data are presented as the mean plusmnSD of three independent experiments Statistical analyses ofchanges for each time and concentration point comparedto the control were performed using a paired two-tailedStudentrsquos t-test A 119875 value lt005 was considered statisticallysignificant
3 Results
31 Composition of COLE In order to identify and quantifythe main compounds present in COLE HPLC analysiswas performed Compounds were identified by comparingeach peakrsquos retention time with that of injected referencestandards in the same chromatographic conditions OnlyOleuropein was detected at 254 nm while the two otherphenylethanoids Hydroxytyrosol and Verbascoside as wellas the flavonoids Apigenin and Luteolin and their glucosideforms were detected at 330 nm (Figure 1) The retention time(min) and the amount (mgmL) of each detected compoundare listed in Table 1 Oleuropein was the major compoundin the extract present at a concentration of 7453mgmLLuteolin-7-glucoside and Apigenin-7-glucoside were presentat 0536 and 0529mgmL respectively The other detectedcompounds Hydroxytyrosol Verbascoside Apigenin andLuteolin were present as traces
The antioxidant activity of Chemlali Olive Leaf Extractwas confirmed by the DPPH radical scavenging assay accord-ing to Enujiugha et al [24] Data showed that the radicalscavenging activity of COLE increased in a dose dependentmanner and that the extract concentration providing 50inhibition of free radicals (IC50) was 06mgmL (data notshown)
32 COLE Inhibits the Proliferation of K562 Cells To evaluatethe antileukemia effect of COLE on K562 cells differentconcentrations from 50 to 150120583gmL were applied on K562cells for the MTT assay Treatment for 24 48 and 72 h
4 Oxidative Medicine and Cellular Longevity
5 254nm
0
200
400
600
800(m
AU)
5092 5330
6793
8008
9939
12511
14086
15191
16120
17086
18050
18737
19022
20132
21064
22125
23065
25779
26629
28065
29037
32310
34407
35982
36878
38286
41288
46112
0 10 20 30 40 50
(min)
(a)
1
2
3 4
6
7
330nm
5341
10429
11708
12683
13625
14768
15634
16254
16657
17344
17816
18026
18280 19230
19959
21050
22127
23051
46608
0
10
20
30
40
50
60
0 5 10 15 20 25 30 35 40 45
(mAU
)
(min)
(b)
Figure 1 HPLC chromatogram of Chemlali Olive Leaf Extract (COLE) at (a) 254 nm and (b) 330 nm Extraction was conducted with 70ethanol Peaks 1 Hydroxytyrosol 2 Verbascoside 3 Luteolin-7-glucoside 4 Apigenin-7-glucoside 5 Oleuropein 6 Luteolin 7 Apigenin
caused a significant decrease of the proliferation of K562 cellsin a dose dependent manner After 72 h of treatment with150 120583gmL of COLE the cell proliferation was inhibited to17 compared with the control cells (Figure 2(a))
Observation of the morphological changes of K562 cellsafter treatment with the different concentrations revealed anincrease in cell size especially in cells treated with 150120583gmLof COLE compared with control cells which kept the samemorphology and cell size (Figure 2(b))
33 COLE Inhibits the Growth Rate of K562 Cells To under-stand if the antiproliferative effect was led by cell death orgrowth inhibition we determined the cell number and cellviability after treatment with COLE by flow cytometry Thenumbers of K562 cells treated with 100 120583gmL and 150 120583gmLwere significantly less than the number of control cellsK562 cells treated with COLE exhibited a slow cell growthcompared to the control (Figure 3(a))
Results (Figure 3(b)) showed that treatment with150 120583gmL of COLE caused a slight decrease of the viabilityof K562 cells during the first 3 days with keeping more than
80 of viable cells Then the decrease became drastic andonly 20 of live cells remain at the 6th day of treatment bythe same concentration The viability of cells treated with100 120583gmL of COLE started to decrease significantly fromthe 4th day while treatment with 50120583gmL did not affect thecell viability during the whole period Taken together theseresults suggested that the antiproliferative effect of COLEat the concentrations of 100 120583gmL and 150 120583gmL mightbe conducted in part by inhibiting the growth rate and to alesser extent by causing cell death
34 COLE Modulates Cell Cycle Progression in K562 CellsSinceCOLE reduced the growth ofK562 cells we investigatedits effect on the cell cycle progression During the 1st and2nd day K562 cells treated with 100 and 150 120583gmL werearrested at G0G1 During the following days results showeda significant increase of G2M population in cell treated with100 and 150 120583gmL of COLE In the case of cells treatedwith 150120583gmL of COLE this increase was associated with asignificant decrease of G0G1 cell population throughout the3rd and 4th days of treatment (Table 2)
Oxidative Medicine and Cellular Longevity 5
0
20
40
60
80
100
120
24 48 72
Cel
l pro
lifer
atio
n (
of co
ntro
l)
Control50120583gmL75120583gmL
100120583gmL125120583gmL150120583gmL
lowastlowastlowast lowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
(h)
(a)
Control 50120583gmL
100120583gmL 150120583gmL
(b)
Figure 2 (a) Effect ofChemlaliOlive Leaf Extract (COLE) on the proliferation ofK562 cellsThe cells were treatedwith various concentrations(50 75 100 125 and 150120583gmL) of COLE for 24 48 and 72 h Cell proliferation was measured byMTT assay Control represents cells treatedwith 03 ethanol in medium Results are represented as the mean plusmn SD of three independent experiments lowast lowastlowast significantly differentfrom the control (119875 lt 005 and 119875 lt 001 resp) (b) Morphological observations of K562 cells after treatment with various concentrationsof Chemlali Olive Leaf Extract (COLE) for 72 h Cells were observed under a phase contrast microscope at 100x magnification Scale barsrepresent 100 120583m
0
2
4
6
8
10
12
14
16
18
20
Day 0 Day 1 Day 2 Day 3 Day 4 Day 6
Num
ber o
f via
ble c
ells
(times106)
Control 50120583gmL100 120583gmL 150120583gmL
(a)
0102030405060708090100
Day 0 Day 1 Day 2 Day 3 Day 4 Day 6
Viab
ility
()
Control50120583gmL
100 120583gmL150120583gmL
lowastlowastlowastlowast
lowastlowast
lowastlowast
lowastlowast
lowast lowast
(b)
Figure 3 Effect of Chemlali Olive Leaf Extract (COLE) on the cell number and viability of K562 cells (a) Number of viable cells aftertreatment with COLE after incubation up to 6 days (b) Percentage of viability in K562 cells treated with COLE K562 cells were treated at afinal concentration of 50 100 and 150120583gmL of COLE and incubated for different periods Cell number and cell viability were measured byflow cytometry Control represents cells treated with 03 ethanol in medium Results are represented as the mean plusmn SD of three independentexperiments lowast lowastlowast significantly different from the control (119875 lt 005 and 119875 lt 001 resp)
35 COLE Induces Apoptosis in K562 Cells The decreasein cell viability (Figure 3(b)) caused by COLE treatmentprompted us to investigate if the extract induces apoptosis inK562 cells Apoptosis is an important and active regulatorypathway of cell growth and proliferation resulting in somecharacteristic physiological changes Among these external-ization of phosphatidylserine (PS) is easily detected by flowcytometry after binding to the labeled Annexin VThe resultsindicated an increase in Annexin V positive cells from the1st day of incubation with 150 120583gmL of COLE in comparisonwith control cells (Figure 4) The percentage of Annexin V
cells remained stable during the first 3 days of treatmentwith 20 of total treated cells and interestingly increasedin the 4th and 6th days with 45 and 60 respectivelyTreatment with 100 120583gmL caused the apparition of apoptoticcells from the 4th day of incubation with a proportionof 162 This population increased in the 6th day up to267No significant difference in apoptotic cells amountwasdetected when cells were treated with 50 120583gmL until 6 daysof incubation
6 Oxidative Medicine and Cellular Longevity
Table 2 The distribution of cell cycle in K562 cells treated with Chemlali Olive Leaf Extract (COLE)
Results are represented as the means plusmn SD of three independent experimentslowast and lowastlowast mean that difference between control and treated cells in each phase (G0G1 S and G2M) is statistically significant at 119875 lt 005 and 119875 lt 001respectively
36 Impact of COLE on the Differentiation Capacity of K562Cells The reduction in cell growth as well as the morpho-logical changes observed in COLE treated cells led us tothink about the differentiation assessment K562 cells arepluripotent malignant cells that spontaneously differentiatealong erythroid macrophage and megakaryocyte lineages[22] Treated cells were harvested and analyzed for theexpression of lineage differentiation markers The analysiswas performed by flow cytometry at different days on themonocytemacrophage marker CD14 on the granulocytemarker CD11b on the erythrocyte marker GPA and on themegakaryocyte marker CD41 The results showed a signifi-cant increase in the expression of CD14 marker from day 1until day 6 in the cells treated with 150120583gmL suggestingthe commitment of K562 cells to the monocytemacrophagelineage (Figure 5(a)) A slight increase of the expression ofCD11b was also detected at this concentration (Figure 5(b))Treatment with 100 120583gmL showed a gain in the expression ofCD14 on the 1st and 2nd day and an augmentation of CD11bexpression from the 3rd day of treatment Treatment with50120583gmL did not show any significant effect on the differenti-ationmarkersCD14 andCD11b Even after 6 days of treatmentwith 150 120583gmL of COLE we could not detect the erythroidmarker at all the tested concentrations (Figure 5(d)) How-ever an instant increase in the megakaryocytic marker wasdetected on the 1st day of treatment followed by a drasticdecrease lower than the control during the following days(Figure 5(c))
37 Gene Expression Profile of COLE Treated K562 Cells Tofurther elucidate the mechanism by which COLE inducesapoptosis and differentiation of K562 cells we investigatedthe changes in gene expression profiles in treated cells usingHG219 GeneChip array Microarray analysis was performedon K562 cells at the 3rd day of treatment based on the obser-vation that such timing generated a significant expression ofthe differentiation markers Genes with more than 15-foldchange in expression levels between control and 150 120583gmLofCOLE treated cells were classified into categories accordingto the cell biological processes According to the GO analysis
0
10
20
30
40
50
60
70
Day 1 Day 2 Day 3 Day 4 Day 6
Ann
exin
pos
itive
(
)
Control50120583gmL
100 120583gmL150120583gmL
lowastlowast lowastlowast lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
Figure 4 Induction of apoptosis in K562 cells treatedwith ChemlaliOlive Leaf Extract (COLE) Cells were treated with 50 100 and150 120583gmL of COLE and incubated for different time At the indi-cated time K562 cells were stained with Annexin V and analyzed byflow cytometry Control represents cells treated with 03 ethanolin medium Results are represented as the mean plusmn SD of threeindependent experiments lowast lowastlowast significantly different from thecontrol (119875 lt 005 and 119875 lt 001 resp)
results (Tables S1 and S2 in Supplementary Material avail-able online at httpdxdoiorg1011552014927619) COLEtreatment induced the upregulation of genes involved inhematopoiesis such as CTNNB1 SH2B3 CIAPIN1 RPA1 andgenes implicated in cell differentiation and its regulation suchasMCL1CTNNB1 andCCNE1 Interestingly genes related tomyeloid cell differentiation such as IFI16ACIN1 andCASP8and particularly genes engaged in monocyte differentiationwere represented in the upregulated categories Moreoveran increased expression of genes involved in chemotaxissuch as NUP85 HRAS IL8 CXCL2 and CXCL3 and genesrelated to cytokine production was also detected Amongupregulated genes AP1G1 RABEP1 RAB5C RAB21 EPS15L1and CORO1C were also found involved in the process ofendocytosis and phagocytosis as well as genes related toprotein transport and genes implicated in cell adhesion suchas ICAM3 HSPB11 and PNN
Conversely the erythrocyte differentiation categorieswere represented in the list of the decreased genes (Table S2)
Figure 5 Expression of differentiationmarkers onK562 cells treatedwith Chemlali Olive Leaf Extract (COLE) up to 6 days Cells were treatedwith 50 100 and 150 120583gmL of COLE and incubated for different periods Control cells were treated with ethanol at a final concentration of03 Expression of cell surface markers was detected by flow cytometry (a) Expression of CD14 (monocytemacrophage specific marker)(b) Expression of CD11b (granulocytemonocyte specific marker) (c) Expression of CD41 (megakaryocyte specific marker) (d) Expressionof glycophorin A (GPA) (erythrocyte specific marker) Results are represented as the mean plusmn SD of three independent experiments lowast lowastlowastsignificantly different from the control at 119875 lt 005 and 119875 lt 001 respectively
including GYPA HBE1 FECH and ALAS2 which is consis-tent with the decreased expression of erythrocyte differenti-ation marker in the flow cytometry results (Figure 5(d))
Analysis of the microarray data showed the upregu-lation of several proapoptotic genes and genes involvedin the regulation of mitochondrial membrane permeabilitysuch as CASP6 CASP8 DFFA and BID On the otherhandexpression of genes negatively regulating the apoptosis
and those inhibiting the caspase activity was significantlydecreased such as IGF1R HSPA5 and BCL2
Among the upregulated genes we found those related tothe NF-kappaB cascade as well as to the MAPKKK cascadeand the Wnt receptor signaling pathway
Categories related to cell cycle regulation and cell pro-liferation were represented by both upregulated and down-regulated genes Finally some transcription factors werealso downregulated such as FOXP1 whereas others wereupregulated like EGR1 and NFYA
The list of selected genes is presented in Table 3 Fromthis list it is clear that the differentially expressed genes aftertreatment with 100 and 150120583gmL have mostly the sametendency which could validate the obtained results
12 Oxidative Medicine and Cellular Longevity
4 Discussion
Previous reports had indicated that olive leaf extract inducesapoptosis in the human leukemic cell lines HL-60 and Jurkatcells [13 21] In this study we give evidence that COLEexhibits its antileukemia effect by both inducing apoptosisand promoting differentiation of the multipotent humanleukemia K562 cells
Induction of differentiation as well as apoptosis has beenfrequently reported to be associatedwith a loss of proliferativecapacity of the cells It has been reported that olive leaf extractinhibits the proliferation of some cancer cell lines such asB16 melanoma cells HL-60 leukemia cells the breast cancerMCF-7 cells and the glioblastoma cells [12 25ndash27] Ourresults indicate that COLE inhibits the proliferation of K562cells in a dose dependent manner (Figure 2(a)) Howeverdespite the drastic decrease in cell proliferation the viabilityof cells treated with the highest concentration of COLE(150 120583gmL) remains relatively high (80) for up to 3 daysof treatment (Figure 3(b))
Since it is well established that the arrest of the cellcycle progression allows the cells to follow other processessuch as apoptosis and differentiation we then explored theeffect of COLE on the cell cycle distribution (Table 2) Oliveleaf extract has been shown to cause cell cycle arrest atG0G1 phase in both the breast cancer MCF-7 cells and themelanoma B16 cells [12 25] However the treatment timesapplied in both of these studies were limited to 48 h and 18 hrespectively In this study COLE treated cells were arrested atG0G1 on the 1st and 2ndday of treatment which is consistentwith the reported studies Prolonged incubation with COLEshowed an arrest of cell cycle at G2M phase (3rd and 4thday of incubation) The microarray analysis performed forthe 3rd day of treatment revealed a significant increase ofCHEK2 gene expression in COLE treated cells The encodedprotein regulates the cell cycle checkpoint arrest throughthe inhibition of the activity of CDC25A CDC25B andCDC25C [28] CDC25 proteins trigger the entry into mitosisat different points of the cell cycle by activating theCdk-cyclincomplexes CDC25A acts early in the cell cycle regulatingthe G1S transition whereas CDC25B and CDC25C act atG2M [29] Interestingly our results showed a decrease in theexpression of CDC25C gene in COLE treated cells associatedwith an increase in the expression of CDC25A which mayexplain the cell cycle arrest at G2M phase observed at the3rd day of treatment with COLE
Treatment with COLE increased the expression of CD14on the cell surface of treated cells indicating the differenti-ation of K562 cells toward the mono-macrophage lineage(Figure 5(a)) The increase in the percentage of cells positivefor CD11b (Figure 5(b)) confirms this hypothesis In factCD11b is expressed on both monocytes and granulocytesThus together with the increase of CD14 expression theexpression of CD11b suggests that treatment with COLEpromotes the commitment of K562 cells into the mono-cytemacrophage lineage at the expense of the granulocyticdifferentiationThe instant increase of the expression of CD41
on the 1st day of treatment and its absence during the fol-lowing days (Figure 5(c)) could be consistent with monocytedifferentiation instead of megakaryocyte differentiation sinceit has been reported that 120572IIb promoter which initiates thetranscription of CD41 is transcriptionally active in pluripo-tent myeloid progenitors in early stages of erythropoiesisand all along the megakaryocytic differentiation as well asto a lesser extent in the early stages of myelomonocyticdifferentiation and the late stages of erythropoiesis [30] Thisinstant increase could be also explained by the phenomenonof lineage conversion where some hematopoietic progenitorscan be converted into lineages other than their own bythe ectopic expression of some transcription factors [31]In this regard it was previously reported that commonlymphoid progenitors megakaryocyte erythroid progenitorsearly B cells and early T cells could be converted to thegranulocytemonocyte lineage by the induction of CEBP120572activity [32] Microarray results support the absence ofmegakaryocyte differentiation in the 3rd day of treatment Infact data showed a decrease in the expression of genes relatedtomegakaryopoiesis such as the essential regulator of plateletrelease NFE2 and its main candidate target gene TUBB1(encoding the megakaryocyte and platelet specific isoform of120573-tubulin (1205731)) [33] as well as the BACH2 transcription factorreported to be induced during megakaryocyte differentiation[34]
Interestingly results of microarray analysis (Table 3)showed the upregulation of several genes already reportedas key players in monocytemacrophage differentiation inthe COLE treated cells Among these genes the expressionof IFI16 was markedly increased Constitutively expressedin lymphoid cells IFI16 was shown to be associated withthe differentiation of human myeloid cells in response tointerferon gamma [35] Moreover IFI16 has been selectivelyexpressed during the differentiation of myeloid progenitorcells along monocytic lineage while it is downregulatedduring the commitment toward the granulocytic or erythroidlineage suggesting its potential involvement in the differenti-ation andmaturation of the monocytic lineage [36 37] Geneencoding for EGR-1 a positive modulator of macrophagedifferentiation was also significantly upregulated EGR-1 isknown to dictate development of myeloid progenitors alongthe macrophage lineage at the expense of development alongother lineages [38 39] Dauffy et al [40] reported thatIFI16-enforced expression in myeloid progenitors inducedthe expression of EGR-1 and conducted the macrophagedifferentiation in the absence of the macrophage colonystimulating factor (M-CSF) This suggests that EGR-1 couldbe regulated by the expression of IFI16 Treatment with COLEincreased as well the expression of NFYA encoding the Asubunit of the nuclear transcription factor Y whose synthesiswas previously reported during the terminal differentiationof monocytes to macrophages [38 41] The downregula-tion of FOXP1 in COLE treated K562 cells is consistentwith monocytemacrophage differentiation since it has beendemonstrated that the expression of FOXP1 was markedlydecreased in monocyte-induced differentiation of HL-60cells as well as in human peripheral blood monocytes and
Oxidative Medicine and Cellular Longevity 13
that the overexpression of FOXP1 prevented the morphologicmacrophage-like differentiation [42 43]
Monocytes and macrophages have several functionalcharacteristics including cell adhesion migration chemo-taxis and phagocytic activity Interestingly an enhancedexpression of the chemokines genes CXCL8 (IL8) CXCL2andCXCL3was found in COLE treated cells IL-8 productionhas been observed in vitro in a wide variety of cells includingmonocytes T lymphocytes neutrophils vascular endothe-lial cells dermal fibroblasts keratinocytes hepatocytes andhuman gastric cancer cells while the production of CXCL2andCXCL3was only described inmonocytes fibroblasts andendothelial cells [44 45] Another gene related to chemotaxiswas upregulated NUP85 which is an essential component ofthe nuclear pore complex and was reported to be involved inCCR2-mediated chemotaxis of monocytes [46]
COLE treatment also enhanced the expression of somegenes related to phagocytosis AP1G1 andRab proteins relatedgenes AP1G1 is a subunit of clathrin-associated adaptorprotein complex 1 whose role has been recently highlightedfor efficient phagocytosis at an early stage of phagosomeformation since it participates in the extension of the phago-cytic cup [47 48] The small GTPases Rab are key regulatorsof intracellular membrane trafficking Recent studies havedemonstrated that several RAB proteins play an importantrole in phagocytosis RAB proteins such as RAB11 Rab5 andRab 21 participate in the phagosome formation and matura-tion and are necessary for the phagocytic activity [48ndash51]
Consistently with the increase in Annexin V positivecells (Figure 4) treatment with COLE upregulated someproapoptotic genes such as CASP6 CASP8 DFFA and BIDand downregulated the apoptosis suppressor BCL2 as wellas the caspase inhibitors genes (Table 3) Olive leaf waspreviously shown to induce morphological changes thatare characteristic of apoptosis in leukemia cells but themechanisms underlying this effect were not well investi-gated [13 21] The increase in the proportion of apoptoticcells observed from the 4th day of treatment with COLEcould be explained by the programmed death of the fullydifferentiated cells (Figure 4) In fact this increase wasaccompanied by an important decrease in cell viability aswell as a diminution in CD14 expression (Figures 3(b) and5(a)) Elsewhere it is well established that in hematopoietictissues apoptosis is coupled to terminal differentiation ofmyeloid progenitors even though themechanism responsiblefor the activation of apoptosis during myeloid maturationis still poorly understood [52 53] It is also noteworthy tomention that several reports have highlighted the role ofcaspases in the terminal differentiation of a variety of celltypes [54] One example is the differentiation of human bloodmonocytes into macrophages a process that is blocked bysynthetic caspase inhibitors [55] It has been also reportedthat caspase 8 deletion in bone-marrow cells resulted in arrestof hemopoietic progenitor functioning and that its deletionin cells of the myelomonocytic lineage led to the arrest ofdifferentiation into macrophages and consequently to celldeath [56]
Among the differentially expressed genes in COLEtreated cells (Table 3) genes encoding the mitogen-activated
protein kinase kinase kinases (MAPKKKs) MAP3K2MAP3K5 and MAP3K7 were highly expressed whileMAP2K5 and MAPK14p38-120572 were downregulated TheMAPKKKs act at the upstream of the MAP kinase cascadewhich regulates important cellular processes such as geneexpression cell proliferation differentiation cell survivaland death The MAP3K5 activates the c-Jun N-terminalkinase (JNK) and p38 mitogen-activated protein kinases[57] while the MAP3K2 is reported to preferentially activateJNK [58 59] However the significant decrease in theMAPK14p38-120572 may suggest that the effect of COLE onK562 cells is likely to be mediated by the JNK MAPKsrather than the p38 MAPKs JNK plays an important rolein apoptosis pathways weather by activating proapoptoticor antiapoptotic genes Particularly it was shown thatprolonged but not transient JNK activation promotes thecell death [60] Our results showed that JKAMP gene whichencodes for a membrane-anchored regulator of the durationof JNK1 activity was upregulated (154 plusmn 0051) The elevatedexpression of JKAMP as reported by Kadoya et al [61]results in sustained JNK activity These findings may indicatethe potential role of JNK pathway in the COLE-inducedapoptosis in K562 cells
Thanks to their large regulatory domains some MAP-KKKs can interact with upstream regulators have functionssuch as ubiquitylation and be activated by relief of autoin-hibition and oligomerization [62] In this context MAP3K2has been reported to play an important role in NF-kappaBsignaling pathway by activating the I-kappaB kinases whichin turn phosphorylate the inhibitory factors of the nuclearfactor kappaB (NF-kappaB) resulting in their rapid ubiq-uitination and the liberation of NF-kappaB complex whichtranslocate from the cytoplasm to the nucleus [63] MAP3K7in association with other proteins could also be required forthe activation of NF-kappaB According to the GO analysis(Table S1) COLE treatment upregulated genes implicatedin NF-kappaB-related categories such as positive regulationof NF-kappaB transcription factor activity I-kappaB kinase-NF-kappaB cascade and its positive regulation and I-kappaBphosphorylation Genes encoding the signal transducers inthe NF-kappaB pathway TRAF5 and TRAF6 as well asthe activator of NF-kappaB signaling SNIP1 were upregu-lated Interestingly an increased expression of NFKB1 geneencoding for the precursor of the mature NF-kappaB p50was detected in COLE treated cells compared to untreatedK562 cells NFKB is well recognized as a central activator ofthe antiapoptotic cascades in response to external stimuli orintrinsic immune reactions and its prosurvival activity hasbeen implicated in a variety of biological processes [60 64]In the B cell lineage the activity of NF-120581B is required forthe completion of various developmental stages includingdifferentiation and the response of these cells to antigens [65]NF-kappaB also plays an important role in the survival anddevelopment of T cells [66] and has been recently reported tomediate the differentiation of mesenchymal stem cells [67]
Several studies have demonstrated a crosstalk betweenthe NF-kappaB and JNK pathways seen as an inhibitoryeffect of NF-kappaB on apoptosis through the suppressionof JNK activity and that this suppressive effect may occur
14 Oxidative Medicine and Cellular Longevity
through different mechanisms [60] This suggests that thebalance between JNK and NF-kappaB activities is crucial todetermine the cell fate survival or deathWehypothesize herethat COLE-induced apoptosis or differentiation is likely to bedetermined by the crosstalk between these two pathways inK562 cells Further investigations regarding protein expres-sion and activity are needed to confirm these findings
5 Conclusions
Our study demonstrates for the first time that olive leafextract exhibits an antileukemia effect on the human chronicmyeloid leukemia cells Olive leaf extract was shown toinhibit the proliferation of K562 cells by inducing cell cyclearrest apoptosis and differentiation toward the monocytelineage The induced expression of molecules involved indifferentiation toward the monocytemacrophage lineage aswell asmolecules related to apoptosis and cell cycle regulationconfirmed these findings and provides insights into themechanism by which olive leaf exhibits its antileukemiaeffect
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported by the JICA-JST Science andTechnology Research Partnership for Sustainable Develop-ment (SATREPS) Project ldquoValorization of Bio-Resources inSemi-Arid and Arid Land for Regional Developmentrdquo
References
[1] M Leszczyniecka T Roberts P Dent S Grant and P B FisherldquoDifferentiation therapy of human cancer basic science andclinical applicationsrdquo Pharmacology and Therapeutics vol 90no 2-3 pp 105ndash156 2001
[2] A I Spira and M A Carducci ldquoDifferentiation therapyrdquoCurrent Opinion in Pharmacology vol 3 no 4 pp 338ndash3432003
[3] S Nobili D Lippi E Witort et al ldquoNatural compounds forcancer treatment and preventionrdquo Pharmacological Researchvol 59 no 6 pp 365ndash378 2009
[4] S Ramos ldquoEffects of dietary flavonoids on apoptotic pathwaysrelated to cancer chemopreventionrdquo Journal of NutritionalBiochemistry vol 18 no 7 pp 427ndash442 2007
[5] V Hajhashemi G Vaseghi M Pourfarzam and A AbdollahildquoAre antioxidants helpful for disease preventionrdquo Research inPharmaceutical Sciences vol 5 no 1 pp 1ndash8 2010
[6] M Bouaziz and S Sayadi ldquoIsolation and evaluation of antiox-idants from leaves of a Tunisian cultivar olive treerdquo EuropeanJournal of Lipid Science andTechnology vol 107 no 7-8 pp 497ndash504 2005
[7] A P Pereira I C F R Ferreira F Marcelino et al ldquoPhenoliccompounds and antimicrobial activity of olive (Olea europaeaL Cv Cobrancosa) leavesrdquo Molecules vol 12 no 5 pp 1153ndash1162 2007
[8] L I Somova F O Shode P Ramnanan and A Nadar ldquoAnti-hypertensive antiatherosclerotic and antioxidant activity oftriterpenoids isolated from Olea europaea subspecies africanaleavesrdquo Journal of Ethnopharmacology vol 84 no 2-3 pp 299ndash305 2003
[9] V Micol N Caturla L Perez-Fons V Mas L Perez andA Estepa ldquoThe olive leaf extract exhibits antiviral activityagainst viral haemorrhagic septicaemia rhabdovirus (VHSV)rdquoAntiviral Research vol 66 no 2-3 pp 129ndash136 2005
[10] J Wainstein T Ganz M Boaz et al ldquoOlive leaf extract as ahypoglycemic agent in both human diabetic subjects and inratsrdquo Journal of Medicinal Food vol 15 no 7 pp 605ndash610 2012
[11] L Seddik TM Bah A AouesM Slimani andM BenderdourldquoElucidation of mechanisms underlying the protective effects ofolive leaf extract against lead-induced neurotoxicity in Wistarratsrdquo Journal of Toxicological Sciences vol 36 no 6 pp 797ndash809 2011
[12] Z Bouallagui J Han H Isoda and S Sayadi ldquoHydroxytyrosolrich extract from olive leaves modulates cell cycle progressionin MCF-7 human breast cancer cellsrdquo Food and ChemicalToxicology vol 49 no 1 pp 179ndash184 2011
[13] R Fares S Bazzi S E Baydoun and R M Abdel-Massih ldquoTheantioxidant and anti-proliferative activity of the Lebanese Oleaeuropaea extractrdquo Plant Foods for Human Nutrition vol 66 no1 pp 58ndash63 2011
[14] V Goulas V Exarchou A N Troganis et al ldquoPhytochemicalsin olive-leaf extracts and their antiproliferative activity againstcancer and endothelial cellsrdquo Molecular Nutrition and FoodResearch vol 53 no 5 pp 600ndash608 2009
[15] M de Bock E B Thorstensen J G Derraik H V HendersonP L Hofman and W S Cutfield ldquoHuman absorption andmetabolism of oleuropein and hydroxytyrosol ingested as olive(Olea europaea L) leaf extractrdquo Molecular Nutrition and FoodResearch vol 57 no 11 pp 2079ndash2085 2013
[16] R Briante M Patumi S Terenziani E Bismuto F Febbraioand R Nucci ldquoOlea europaea L leaf extract and derivativesantioxidant propertiesrdquo Journal of Agricultural and Food Chem-istry vol 50 no 17 pp 4934ndash4940 2002
[17] O Benavente-Garcıa J Castillo J Lorente A Ortuno and J ADel Rio ldquoAntioxidant activity of phenolics extracted from Oleaeuropaea L leavesrdquo Food Chemistry vol 68 no 4 pp 457ndash4622000
[18] S H Omar ldquoOleuropein in olive and its pharmacologicaleffectsrdquo Scientia Pharmaceutica vol 78 no 2 pp 133ndash154 2010
[19] H Jemai M Bouaziz I Fki A El Feki and S SayadildquoHypolipidimic and antioxidant activities of oleuropein and itshydrolysis derivative-rich extracts from Chemlali olive leavesrdquoChemico-Biological Interactions vol 176 no 2-3 pp 88ndash982008
[20] R Garcıa-Villalba M Larrosa S Possemiers F A Tomas-Barberan and J C Espın ldquoBioavailability of phenolics from anoleuropein-rich olive (Olea europaea) leaf extract and its acuteeffect on plasma antioxidant status comparison between pre-and postmenopausal womenrdquo European Journal of Nutrition2013
[21] J Anter Z Fernandez-Bedmar M Villatoro-Pulido et al ldquoApilot study on the DNA-protective cytotoxic and apoptosis-inducing properties of olive-leaf extractsrdquo Mutation Researchvol 723 no 2 pp 165ndash170 2011
[22] J A Sutherland A R Turner and P Mannoni ldquoDifferen-tiation of K562 leukemia cells along erythroid macrophage
Oxidative Medicine and Cellular Longevity 15
and megakaryocyte lineagesrdquo Journal of Biological ResponseModifiers vol 5 no 3 pp 250ndash262 1986
[23] K Schwenke H-P Peterson K-H Wangenheim and L EFeinendegen ldquoInduction of differentiation in erythroleukemicK562 cells by 120574-irradiationrdquo Leukemia Research vol 19 no 12pp 955ndash961 1995
[24] V N Enujiugha J Y Talabi S A Malomo and A I OlagunjuldquoDPPH radical scavenging capacity of phenolic extracts fromAfrican Yam Bean (Sphenostylis stenocarpa)rdquo Food and Nutri-tion Sciences vol 3 pp 7ndash13 2012
[25] S A Mijatovic G S Timotijevic D M Miljkovic et alldquoMultiple antimelanoma potential of dry olive leaf extractrdquoInternational Journal of Cancer vol 128 no 8 pp 1955ndash19652011
[26] L Abaza T P N Talorete P Yamada Y KuritaM Zarrouk andH Isoda ldquoInduction of growth inhibition and differentiation ofhuman leukemia HL-60 cells by a Tunisian Gerboui olive leafextractrdquo Bioscience Biotechnology and Biochemistry vol 71 no5 pp 1306ndash1312 2007
[27] B Tunca G Tezcan G Cecener et al ldquoOlea europea leafextract alters microRNA expression in human glioblastomacellsrdquo Journal of Cancer Research and Clinical Oncology vol 138no 11 pp 1831ndash1844 2012
[28] S Matsuoka M Huang and S J Elledge ldquoLinkage of ATM tocell cycle regulation by the Chk2 protein kinaserdquo Science vol282 no 5395 pp 1893ndash1897 1998
[29] C Frazer and P G Young ldquoPhosphorylation mediated regu-lation of Cdc25 activity localization and stabilityrdquo in ProteinPhosphorylation in Human Health C Huang Ed pp 395ndash436InTech 2012
[30] N Debili C Robin V Schiavon et al ldquoDifferent expression ofCD41 on human lymphoid andmyeloid progenitors from adultsand neonatesrdquo Blood vol 97 no 7 pp 2023ndash2030 2001
[31] H Nakajima ldquoRole of transcription factors in differentiationand reprogramming of hematopoietic cellsrdquo Keio Journal ofMedicine vol 60 no 2 pp 47ndash55 2011
[32] Y Fukuchi F Shibata M Ito et al ldquoComprehensive analysis ofmyeloid lineage conversion using mice expressing an inducibleform of CEBP120572rdquoThe EMBO Journal vol 25 no 14 pp 3398ndash3410 2006
[33] R A Shivdasani ldquoMolecular and transcriptional regulation ofmegakaryocyte differentiationrdquo Stem Cells vol 19 no 5 pp397ndash407 2001
[34] K Terui Y Takahashi J Kitazawa T TokiM Yokoyama and EIto ldquoExpression of transcription factors during megakaryocyticdifferentiation of CD34+ cells from human cord blood inducedby thrombopoietinrdquo Tohoku Journal of Experimental Medicinevol 192 no 4 pp 259ndash273 2000
[35] J A Trapani M Dawson V A Apostolidis and K A BrowneldquoGenomic organization of IFI16 an interferon-inducible genewhose expression is associated with human myeloid cell differ-entiation correlation of predicted protein domains with exonorganizationrdquo Immunogenetics vol 40 no 6 pp 415ndash424 1994
[36] M J Dawson N J Elwood R W Johnstone and J A TrapanildquoThe IFN-inducible nucleoprotein IFI 16 is expressed in cellsof the monocyte lineage but is rapidly and markedly down-regulated in other myeloid precursor populationsrdquo Journal ofLeukocyte Biology vol 64 no 4 pp 546ndash554 1998
[37] M Gariglio M De Andrea M Lembo et al ldquoThe murinehomolog of the HIN 200 family Ifi 204 is constitutivelyexpressed in myeloid cells and selectively induced in the
monocytemacrophage lineagerdquo Journal of Leukocyte Biologyvol 64 no 5 pp 608ndash614 1998
[38] A F Valledor F E Borras M Cullell-Young and A CeladaldquoTranscription factors that regulate monocytemacrophage dif-ferentiationrdquo Journal of Leukocyte Biology vol 63 no 4 pp405ndash417 1998
[39] K Krishnaraju B Hoffman and D A Liebermann ldquoEarlygrowth response gene 1 stimulates development of hematopoi-etic progenitor cells along the macrophage lineage at theexpense of the granulocyte and erythroid lineagesrdquo Blood vol97 no 5 pp 1298ndash1305 2001
[40] J Dauffy G Mouchiroud and R P Bourette ldquoThe interferon-inducible gene Ifi204 is transcriptionally activated in responseto M-CSF and its expression favors macrophage differentiationin myeloid progenitor cellsrdquo Journal of Leukocyte Biology vol79 no 1 pp 173ndash183 2006
[41] G Marziali E Perrotti R Ilari U Testa E M Coccia andA Battistini ldquoTranscriptional regulation of the ferritin heavy-chain gene the activity of the CCAAT binding factor NF-Y ismodulated in heme-treated Friend leukemia cells and duringmonocyte-to-macrophage differentiationrdquo Molecular and Cel-lular Biology vol 17 no 3 pp 1387ndash1395 1997
[42] C Shi X Zhang Z Chen et al ldquoIntegrin engagement regulatesmonocyte differentiation through the forkhead transcriptionfactor Foxp1rdquo Journal of Clinical Investigation vol 114 no 3 pp408ndash418 2004
[43] C Shi M Sakuma T Mooroka et al ldquoDown-regulation of theforkhead transcription factor Foxp1 is required for monocytedifferentiation andmacrophage functionrdquoBlood vol 112 no 12pp 4699ndash4711 2008
[44] D F Smith E Galkina K Ley and Y Huo ldquoGRO familychemokines are specialized for monocyte arrest from flowrdquoThe American Journal of Physiology Heart and CirculatoryPhysiology vol 289 no 5 pp H1976ndashH1984 2005
[45] J J Oppenheim O M Z Howard and E Goetzl ChemotacticFactors Neuropeptides and Other Ligands for Seven Transmem-brane Receptors 2000
[46] Y Terashima N Onai M Murai et al ldquoPivotal function forcytoplasmic protein FROUNT in CCR2-mediated monocytechemotaxisrdquo Nature Immunology vol 6 no 8 pp 827ndash8352005
[47] Y Lefkir M Malbouyres D Gotthardt et al ldquoInvolvement ofthe AP-1 adaptor complex in early steps of phagocytosis andmacropinocytosisrdquo Molecular Biology of the Cell vol 15 no 2pp 861ndash869 2004
[48] F Niedergang and P Chavrier ldquoSignaling and membranedynamics during phagocytosis many roads lead to the phago-somerdquo Current Opinion in Cell Biology vol 16 no 4 pp 422ndash428 2004
[49] D Cox D J Lee B M Dale J Calafat and S Green-berg ldquoA Rab11-containing rapidly recycling compartment inmacrophages that promotes phagocytosisrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 97 no 2 pp 680ndash685 2000
[50] O V Vieira C Bucci R E Harrison et al ldquoModulation of Rab5and Rab7 recruitment to phagosomes by phosphatidylinositol3-kinaserdquoMolecular andCellular Biology vol 23 no 7 pp 2501ndash2514 2003
[51] T Khurana J A Brzostowski and A R Kimmel ldquoARab21LIM-onlyCH-LIM complex regulates phagocytosis viaboth activating and inhibitory mechanismsrdquo The EMBO Jour-nal vol 24 no 13 pp 2254ndash2264 2005
16 Oxidative Medicine and Cellular Longevity
[52] S J Martin J G Bradley and T G Cotter ldquoHL-60 cellsinduced to differentiate towards neutrophils subsequently dievia apoptosisrdquo Clinical and Experimental Immunology vol 79no 3 pp 448ndash453 1990
[53] A Benito D Grillot G Nunez and J L Fernandez-Luna ldquoReg-ulation and function of Bcl-2 during differentiation-inducedcell death in HL-60 promyelocytic cellsrdquoThe American Journalof Pathology vol 146 no 2 pp 481ndash490 1995
[54] M Lamkanfi N Festjens W Declercq T V Berghe andP Vandenabeele ldquoCaspases in cell survival proliferation anddifferentiationrdquo Cell Death and Differentiation vol 14 no 1 pp44ndash55 2007
[55] O Sordet C Rebe S Plenchette et al ldquoSpecific involvement ofcaspases in the differentiation of monocytes into macrophagesrdquoBlood vol 100 no 13 pp 4446ndash4453 2002
[56] T-B Kang T Ben-Moshe E E Varfolomeev et al ldquoCaspase-8 serves both apoptotic and nonapoptotic rolesrdquo Journal ofImmunology vol 173 no 5 pp 2976ndash2984 2004
[57] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[58] J Cheng J Yang Y Xia M Karin and B Su ldquoSynergisticinteraction of MEK kinase 2 c-Jun N-terminal kinase (JNK)kinase 2 and JNK1 results in efficient and specific JNK1activationrdquo Molecular and Cellular Biology vol 20 no 7 pp2334ndash2342 2000
[59] J L Blank P Gerwins EM Elliott S Sather andG L JohnsonldquoMolecular cloning of mitogen-activated proteinERK kinasekinases (MEKK) 2 and 3 regulation of sequential phosphoryla-tion pathways involving mitogen-activated protein kinase andc-Jun kinaserdquo Journal of Biological Chemistry vol 271 no 10pp 5361ndash5368 1996
[60] S Papa F Zazzeroni C G Pham C Bubici and G FranzosoldquoLinking JNK signaling to NF-120581B a key to survivalrdquo Journal ofCell Science vol 117 no 22 pp 5197ndash5208 2004
[61] T Kadoya A Khurana M Tcherpakov et al ldquoJAMP a JUB N-terminal kinase 1 (JNK1)-associated membrane protein regu-lates duration of JNK activityrdquo Molecular and Cellular Biologyvol 25 no 19 pp 8619ndash8630 2005
[62] M Qi and E A Elion ldquoMAP kinase pathwaysrdquo Journal of CellScience vol 118 no 16 pp 3569ndash3572 2005
[63] Q Zhao and F S Lee ldquoMitogen-activated protein kinaseERKkinase kinases 2 and 3 activate nuclear factor-120581B through I120581Bkinase-120572 and I120581B kinase-120573rdquoThe Journal of Biological Chemistryvol 274 no 13 pp 8355ndash8358 1999
[64] H Namba V Saenko and S Yamashita ldquoNuclear factor-120581Bin thyroid carcinogenesis and progression a novel therapeutictarget for advanced thyroid cancerrdquo Arquivos Brasileiros deEndocrinologia e Metabologia vol 51 no 5 pp 843ndash851 2007
[65] B Feng S Cheng W S Pear and H-C Liou ldquoNF-kBinhibitor blocks B cell development at two checkpointsrdquoMedi-cal Immunology vol 3 article 1 2004
[66] S Gerondakis and U Siebenlist ldquoRoles of the NF-kappaBpathway in lymphocyte development and functionrdquoCold SpringHarbor Perspectives in Biology vol 2 no 5 Article ID a0001822010
[67] K P Bhat V Balasubramaniyan B Vaillant et al ldquoMesenchy-mal differentiation mediated by NF-kB promotes radiationresistance in glioblastomardquo Cancer Cell vol 24 no 3 pp 331ndash346 2013
cytometry Briefly 20 times 104 cellsmL of K562 cells wereseeded in 6-well plates and treated by 100 and 150 120583gmLof COLE diluted in medium Control cells were treatedwith ethanol 03 After the desired time of incubationcells were harvested washed twice with PBS and fixed with70 ethanol at 4∘C for more than 12 h The fixed cells werethen centrifuged at 500timesg for 5min and washed with PBStwice Cell cycle reagent (Guava Technologies) was addedand the cells were kept in darkness for 30min at roomtemperature The population of cells in each cell cycle phasewas determined by a Guava PCA flow cytometry accordingto their DNA content
27 Annexin V Assay The induction of apoptosis in treatedcells was determined by measuring the externalization ofphosphatidylserine (PS) to the cell surface by flow cytometryK562 cells were seeded in 6 well plates and treated with 50100 and 150 ugmL of COLE diluted inmedium Control cellswere treated with ethanol 03 When reaching the desiredtreatment time cells were harvested and stained with GuavaNexin ReagentThen cells were incubated for 20min at roomtemperature in the dark and then acquired on theGuava PCAsystem
28 Cell Differentiation Assay Cell differentiation wasassessed by flow cytometry by measuring the expression ofCD11b and CD14 on the surface of K562 cells Cells wereseeded at 20 times 104 cellsmL in 6-well plates and incubatedfor 24 h The COLE diluted in medium was added at finalconcentrations of 50 100 and 150120583gmL and ethanol at03 in the case of control cells After incubation cellswere harvested washed twice with cold PBS and adjustedto the same number 10 times 105 The cells were then labeledwith phycoerythrin conjugated anti-CD14 anti-CD11banti-CD41 and anti-glycophorin A for 30min according tomanufacturerrsquos instructions The stained cells were washedtwice with cold PBS and resuspended in 500120583L PBS formeasurement
29 Total RNA Isolation K562 cells at a concentration of20 times 104 cellsmL were incubated for 24 h and then treatedwith 100 and 150 120583gmL of COLE for 72 h The control cellswere treated with ethanol at final concentration of 03The cells were then collected and washed with PBS DNA-free total RNA was isolated from the cells using Isogenreagent (Nippon Gene Co Tokyo Japan) following themanufacturerrsquos instructions
210 DNA Microarray Analysis Microarray hybridizationprobes were generated from isolated RNA samples Double-stranded cDNA was synthesized from 100 ng of total RNAusing the GeneAtlas 31015840 IVT Express Kit (Affymetrix Inc)Biotin-labeled aRNA was synthesized by in vitro transcrip-tion and purified 10 120583g of purified aRNA was then frag-mented using the GeneAtlas 31015840 IVT Express Kit and washybridized to the Affymetrix HG-U219 (Affymetrix) for 16 hat 45∘CThe chips were washed and stained in the GeneAtlasFluidics Station 400 (Affymetrix) and then imaged in the
Table 1 Retention time and concentration of the main phenoliccompounds present in Chemlali Olive Leaf Extract (COLE)
GeneAtlas Imaging Station (Affymetrix) The Partek Expresssoftware (Affymetrix) served for the data analysis by runningcomparisons of gene expression in treated and control cellsbased on mathematical algorithms The generated data (sig-nificant fold change in gene expression) was then analyzedusing the Pathway Studio Explore 11 software (Affymetrix)
211 Statistical Analysis Data are presented as the mean plusmnSD of three independent experiments Statistical analyses ofchanges for each time and concentration point comparedto the control were performed using a paired two-tailedStudentrsquos t-test A 119875 value lt005 was considered statisticallysignificant
3 Results
31 Composition of COLE In order to identify and quantifythe main compounds present in COLE HPLC analysiswas performed Compounds were identified by comparingeach peakrsquos retention time with that of injected referencestandards in the same chromatographic conditions OnlyOleuropein was detected at 254 nm while the two otherphenylethanoids Hydroxytyrosol and Verbascoside as wellas the flavonoids Apigenin and Luteolin and their glucosideforms were detected at 330 nm (Figure 1) The retention time(min) and the amount (mgmL) of each detected compoundare listed in Table 1 Oleuropein was the major compoundin the extract present at a concentration of 7453mgmLLuteolin-7-glucoside and Apigenin-7-glucoside were presentat 0536 and 0529mgmL respectively The other detectedcompounds Hydroxytyrosol Verbascoside Apigenin andLuteolin were present as traces
The antioxidant activity of Chemlali Olive Leaf Extractwas confirmed by the DPPH radical scavenging assay accord-ing to Enujiugha et al [24] Data showed that the radicalscavenging activity of COLE increased in a dose dependentmanner and that the extract concentration providing 50inhibition of free radicals (IC50) was 06mgmL (data notshown)
32 COLE Inhibits the Proliferation of K562 Cells To evaluatethe antileukemia effect of COLE on K562 cells differentconcentrations from 50 to 150120583gmL were applied on K562cells for the MTT assay Treatment for 24 48 and 72 h
4 Oxidative Medicine and Cellular Longevity
5 254nm
0
200
400
600
800(m
AU)
5092 5330
6793
8008
9939
12511
14086
15191
16120
17086
18050
18737
19022
20132
21064
22125
23065
25779
26629
28065
29037
32310
34407
35982
36878
38286
41288
46112
0 10 20 30 40 50
(min)
(a)
1
2
3 4
6
7
330nm
5341
10429
11708
12683
13625
14768
15634
16254
16657
17344
17816
18026
18280 19230
19959
21050
22127
23051
46608
0
10
20
30
40
50
60
0 5 10 15 20 25 30 35 40 45
(mAU
)
(min)
(b)
Figure 1 HPLC chromatogram of Chemlali Olive Leaf Extract (COLE) at (a) 254 nm and (b) 330 nm Extraction was conducted with 70ethanol Peaks 1 Hydroxytyrosol 2 Verbascoside 3 Luteolin-7-glucoside 4 Apigenin-7-glucoside 5 Oleuropein 6 Luteolin 7 Apigenin
caused a significant decrease of the proliferation of K562 cellsin a dose dependent manner After 72 h of treatment with150 120583gmL of COLE the cell proliferation was inhibited to17 compared with the control cells (Figure 2(a))
Observation of the morphological changes of K562 cellsafter treatment with the different concentrations revealed anincrease in cell size especially in cells treated with 150120583gmLof COLE compared with control cells which kept the samemorphology and cell size (Figure 2(b))
33 COLE Inhibits the Growth Rate of K562 Cells To under-stand if the antiproliferative effect was led by cell death orgrowth inhibition we determined the cell number and cellviability after treatment with COLE by flow cytometry Thenumbers of K562 cells treated with 100 120583gmL and 150 120583gmLwere significantly less than the number of control cellsK562 cells treated with COLE exhibited a slow cell growthcompared to the control (Figure 3(a))
Results (Figure 3(b)) showed that treatment with150 120583gmL of COLE caused a slight decrease of the viabilityof K562 cells during the first 3 days with keeping more than
80 of viable cells Then the decrease became drastic andonly 20 of live cells remain at the 6th day of treatment bythe same concentration The viability of cells treated with100 120583gmL of COLE started to decrease significantly fromthe 4th day while treatment with 50120583gmL did not affect thecell viability during the whole period Taken together theseresults suggested that the antiproliferative effect of COLEat the concentrations of 100 120583gmL and 150 120583gmL mightbe conducted in part by inhibiting the growth rate and to alesser extent by causing cell death
34 COLE Modulates Cell Cycle Progression in K562 CellsSinceCOLE reduced the growth ofK562 cells we investigatedits effect on the cell cycle progression During the 1st and2nd day K562 cells treated with 100 and 150 120583gmL werearrested at G0G1 During the following days results showeda significant increase of G2M population in cell treated with100 and 150 120583gmL of COLE In the case of cells treatedwith 150120583gmL of COLE this increase was associated with asignificant decrease of G0G1 cell population throughout the3rd and 4th days of treatment (Table 2)
Oxidative Medicine and Cellular Longevity 5
0
20
40
60
80
100
120
24 48 72
Cel
l pro
lifer
atio
n (
of co
ntro
l)
Control50120583gmL75120583gmL
100120583gmL125120583gmL150120583gmL
lowastlowastlowast lowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
(h)
(a)
Control 50120583gmL
100120583gmL 150120583gmL
(b)
Figure 2 (a) Effect ofChemlaliOlive Leaf Extract (COLE) on the proliferation ofK562 cellsThe cells were treatedwith various concentrations(50 75 100 125 and 150120583gmL) of COLE for 24 48 and 72 h Cell proliferation was measured byMTT assay Control represents cells treatedwith 03 ethanol in medium Results are represented as the mean plusmn SD of three independent experiments lowast lowastlowast significantly differentfrom the control (119875 lt 005 and 119875 lt 001 resp) (b) Morphological observations of K562 cells after treatment with various concentrationsof Chemlali Olive Leaf Extract (COLE) for 72 h Cells were observed under a phase contrast microscope at 100x magnification Scale barsrepresent 100 120583m
0
2
4
6
8
10
12
14
16
18
20
Day 0 Day 1 Day 2 Day 3 Day 4 Day 6
Num
ber o
f via
ble c
ells
(times106)
Control 50120583gmL100 120583gmL 150120583gmL
(a)
0102030405060708090100
Day 0 Day 1 Day 2 Day 3 Day 4 Day 6
Viab
ility
()
Control50120583gmL
100 120583gmL150120583gmL
lowastlowastlowastlowast
lowastlowast
lowastlowast
lowastlowast
lowast lowast
(b)
Figure 3 Effect of Chemlali Olive Leaf Extract (COLE) on the cell number and viability of K562 cells (a) Number of viable cells aftertreatment with COLE after incubation up to 6 days (b) Percentage of viability in K562 cells treated with COLE K562 cells were treated at afinal concentration of 50 100 and 150120583gmL of COLE and incubated for different periods Cell number and cell viability were measured byflow cytometry Control represents cells treated with 03 ethanol in medium Results are represented as the mean plusmn SD of three independentexperiments lowast lowastlowast significantly different from the control (119875 lt 005 and 119875 lt 001 resp)
35 COLE Induces Apoptosis in K562 Cells The decreasein cell viability (Figure 3(b)) caused by COLE treatmentprompted us to investigate if the extract induces apoptosis inK562 cells Apoptosis is an important and active regulatorypathway of cell growth and proliferation resulting in somecharacteristic physiological changes Among these external-ization of phosphatidylserine (PS) is easily detected by flowcytometry after binding to the labeled Annexin VThe resultsindicated an increase in Annexin V positive cells from the1st day of incubation with 150 120583gmL of COLE in comparisonwith control cells (Figure 4) The percentage of Annexin V
cells remained stable during the first 3 days of treatmentwith 20 of total treated cells and interestingly increasedin the 4th and 6th days with 45 and 60 respectivelyTreatment with 100 120583gmL caused the apparition of apoptoticcells from the 4th day of incubation with a proportionof 162 This population increased in the 6th day up to267No significant difference in apoptotic cells amountwasdetected when cells were treated with 50 120583gmL until 6 daysof incubation
6 Oxidative Medicine and Cellular Longevity
Table 2 The distribution of cell cycle in K562 cells treated with Chemlali Olive Leaf Extract (COLE)
Results are represented as the means plusmn SD of three independent experimentslowast and lowastlowast mean that difference between control and treated cells in each phase (G0G1 S and G2M) is statistically significant at 119875 lt 005 and 119875 lt 001respectively
36 Impact of COLE on the Differentiation Capacity of K562Cells The reduction in cell growth as well as the morpho-logical changes observed in COLE treated cells led us tothink about the differentiation assessment K562 cells arepluripotent malignant cells that spontaneously differentiatealong erythroid macrophage and megakaryocyte lineages[22] Treated cells were harvested and analyzed for theexpression of lineage differentiation markers The analysiswas performed by flow cytometry at different days on themonocytemacrophage marker CD14 on the granulocytemarker CD11b on the erythrocyte marker GPA and on themegakaryocyte marker CD41 The results showed a signifi-cant increase in the expression of CD14 marker from day 1until day 6 in the cells treated with 150120583gmL suggestingthe commitment of K562 cells to the monocytemacrophagelineage (Figure 5(a)) A slight increase of the expression ofCD11b was also detected at this concentration (Figure 5(b))Treatment with 100 120583gmL showed a gain in the expression ofCD14 on the 1st and 2nd day and an augmentation of CD11bexpression from the 3rd day of treatment Treatment with50120583gmL did not show any significant effect on the differenti-ationmarkersCD14 andCD11b Even after 6 days of treatmentwith 150 120583gmL of COLE we could not detect the erythroidmarker at all the tested concentrations (Figure 5(d)) How-ever an instant increase in the megakaryocytic marker wasdetected on the 1st day of treatment followed by a drasticdecrease lower than the control during the following days(Figure 5(c))
37 Gene Expression Profile of COLE Treated K562 Cells Tofurther elucidate the mechanism by which COLE inducesapoptosis and differentiation of K562 cells we investigatedthe changes in gene expression profiles in treated cells usingHG219 GeneChip array Microarray analysis was performedon K562 cells at the 3rd day of treatment based on the obser-vation that such timing generated a significant expression ofthe differentiation markers Genes with more than 15-foldchange in expression levels between control and 150 120583gmLofCOLE treated cells were classified into categories accordingto the cell biological processes According to the GO analysis
0
10
20
30
40
50
60
70
Day 1 Day 2 Day 3 Day 4 Day 6
Ann
exin
pos
itive
(
)
Control50120583gmL
100 120583gmL150120583gmL
lowastlowast lowastlowast lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
Figure 4 Induction of apoptosis in K562 cells treatedwith ChemlaliOlive Leaf Extract (COLE) Cells were treated with 50 100 and150 120583gmL of COLE and incubated for different time At the indi-cated time K562 cells were stained with Annexin V and analyzed byflow cytometry Control represents cells treated with 03 ethanolin medium Results are represented as the mean plusmn SD of threeindependent experiments lowast lowastlowast significantly different from thecontrol (119875 lt 005 and 119875 lt 001 resp)
results (Tables S1 and S2 in Supplementary Material avail-able online at httpdxdoiorg1011552014927619) COLEtreatment induced the upregulation of genes involved inhematopoiesis such as CTNNB1 SH2B3 CIAPIN1 RPA1 andgenes implicated in cell differentiation and its regulation suchasMCL1CTNNB1 andCCNE1 Interestingly genes related tomyeloid cell differentiation such as IFI16ACIN1 andCASP8and particularly genes engaged in monocyte differentiationwere represented in the upregulated categories Moreoveran increased expression of genes involved in chemotaxissuch as NUP85 HRAS IL8 CXCL2 and CXCL3 and genesrelated to cytokine production was also detected Amongupregulated genes AP1G1 RABEP1 RAB5C RAB21 EPS15L1and CORO1C were also found involved in the process ofendocytosis and phagocytosis as well as genes related toprotein transport and genes implicated in cell adhesion suchas ICAM3 HSPB11 and PNN
Conversely the erythrocyte differentiation categorieswere represented in the list of the decreased genes (Table S2)
Figure 5 Expression of differentiationmarkers onK562 cells treatedwith Chemlali Olive Leaf Extract (COLE) up to 6 days Cells were treatedwith 50 100 and 150 120583gmL of COLE and incubated for different periods Control cells were treated with ethanol at a final concentration of03 Expression of cell surface markers was detected by flow cytometry (a) Expression of CD14 (monocytemacrophage specific marker)(b) Expression of CD11b (granulocytemonocyte specific marker) (c) Expression of CD41 (megakaryocyte specific marker) (d) Expressionof glycophorin A (GPA) (erythrocyte specific marker) Results are represented as the mean plusmn SD of three independent experiments lowast lowastlowastsignificantly different from the control at 119875 lt 005 and 119875 lt 001 respectively
including GYPA HBE1 FECH and ALAS2 which is consis-tent with the decreased expression of erythrocyte differenti-ation marker in the flow cytometry results (Figure 5(d))
Analysis of the microarray data showed the upregu-lation of several proapoptotic genes and genes involvedin the regulation of mitochondrial membrane permeabilitysuch as CASP6 CASP8 DFFA and BID On the otherhandexpression of genes negatively regulating the apoptosis
and those inhibiting the caspase activity was significantlydecreased such as IGF1R HSPA5 and BCL2
Among the upregulated genes we found those related tothe NF-kappaB cascade as well as to the MAPKKK cascadeand the Wnt receptor signaling pathway
Categories related to cell cycle regulation and cell pro-liferation were represented by both upregulated and down-regulated genes Finally some transcription factors werealso downregulated such as FOXP1 whereas others wereupregulated like EGR1 and NFYA
The list of selected genes is presented in Table 3 Fromthis list it is clear that the differentially expressed genes aftertreatment with 100 and 150120583gmL have mostly the sametendency which could validate the obtained results
12 Oxidative Medicine and Cellular Longevity
4 Discussion
Previous reports had indicated that olive leaf extract inducesapoptosis in the human leukemic cell lines HL-60 and Jurkatcells [13 21] In this study we give evidence that COLEexhibits its antileukemia effect by both inducing apoptosisand promoting differentiation of the multipotent humanleukemia K562 cells
Induction of differentiation as well as apoptosis has beenfrequently reported to be associatedwith a loss of proliferativecapacity of the cells It has been reported that olive leaf extractinhibits the proliferation of some cancer cell lines such asB16 melanoma cells HL-60 leukemia cells the breast cancerMCF-7 cells and the glioblastoma cells [12 25ndash27] Ourresults indicate that COLE inhibits the proliferation of K562cells in a dose dependent manner (Figure 2(a)) Howeverdespite the drastic decrease in cell proliferation the viabilityof cells treated with the highest concentration of COLE(150 120583gmL) remains relatively high (80) for up to 3 daysof treatment (Figure 3(b))
Since it is well established that the arrest of the cellcycle progression allows the cells to follow other processessuch as apoptosis and differentiation we then explored theeffect of COLE on the cell cycle distribution (Table 2) Oliveleaf extract has been shown to cause cell cycle arrest atG0G1 phase in both the breast cancer MCF-7 cells and themelanoma B16 cells [12 25] However the treatment timesapplied in both of these studies were limited to 48 h and 18 hrespectively In this study COLE treated cells were arrested atG0G1 on the 1st and 2ndday of treatment which is consistentwith the reported studies Prolonged incubation with COLEshowed an arrest of cell cycle at G2M phase (3rd and 4thday of incubation) The microarray analysis performed forthe 3rd day of treatment revealed a significant increase ofCHEK2 gene expression in COLE treated cells The encodedprotein regulates the cell cycle checkpoint arrest throughthe inhibition of the activity of CDC25A CDC25B andCDC25C [28] CDC25 proteins trigger the entry into mitosisat different points of the cell cycle by activating theCdk-cyclincomplexes CDC25A acts early in the cell cycle regulatingthe G1S transition whereas CDC25B and CDC25C act atG2M [29] Interestingly our results showed a decrease in theexpression of CDC25C gene in COLE treated cells associatedwith an increase in the expression of CDC25A which mayexplain the cell cycle arrest at G2M phase observed at the3rd day of treatment with COLE
Treatment with COLE increased the expression of CD14on the cell surface of treated cells indicating the differenti-ation of K562 cells toward the mono-macrophage lineage(Figure 5(a)) The increase in the percentage of cells positivefor CD11b (Figure 5(b)) confirms this hypothesis In factCD11b is expressed on both monocytes and granulocytesThus together with the increase of CD14 expression theexpression of CD11b suggests that treatment with COLEpromotes the commitment of K562 cells into the mono-cytemacrophage lineage at the expense of the granulocyticdifferentiationThe instant increase of the expression of CD41
on the 1st day of treatment and its absence during the fol-lowing days (Figure 5(c)) could be consistent with monocytedifferentiation instead of megakaryocyte differentiation sinceit has been reported that 120572IIb promoter which initiates thetranscription of CD41 is transcriptionally active in pluripo-tent myeloid progenitors in early stages of erythropoiesisand all along the megakaryocytic differentiation as well asto a lesser extent in the early stages of myelomonocyticdifferentiation and the late stages of erythropoiesis [30] Thisinstant increase could be also explained by the phenomenonof lineage conversion where some hematopoietic progenitorscan be converted into lineages other than their own bythe ectopic expression of some transcription factors [31]In this regard it was previously reported that commonlymphoid progenitors megakaryocyte erythroid progenitorsearly B cells and early T cells could be converted to thegranulocytemonocyte lineage by the induction of CEBP120572activity [32] Microarray results support the absence ofmegakaryocyte differentiation in the 3rd day of treatment Infact data showed a decrease in the expression of genes relatedtomegakaryopoiesis such as the essential regulator of plateletrelease NFE2 and its main candidate target gene TUBB1(encoding the megakaryocyte and platelet specific isoform of120573-tubulin (1205731)) [33] as well as the BACH2 transcription factorreported to be induced during megakaryocyte differentiation[34]
Interestingly results of microarray analysis (Table 3)showed the upregulation of several genes already reportedas key players in monocytemacrophage differentiation inthe COLE treated cells Among these genes the expressionof IFI16 was markedly increased Constitutively expressedin lymphoid cells IFI16 was shown to be associated withthe differentiation of human myeloid cells in response tointerferon gamma [35] Moreover IFI16 has been selectivelyexpressed during the differentiation of myeloid progenitorcells along monocytic lineage while it is downregulatedduring the commitment toward the granulocytic or erythroidlineage suggesting its potential involvement in the differenti-ation andmaturation of the monocytic lineage [36 37] Geneencoding for EGR-1 a positive modulator of macrophagedifferentiation was also significantly upregulated EGR-1 isknown to dictate development of myeloid progenitors alongthe macrophage lineage at the expense of development alongother lineages [38 39] Dauffy et al [40] reported thatIFI16-enforced expression in myeloid progenitors inducedthe expression of EGR-1 and conducted the macrophagedifferentiation in the absence of the macrophage colonystimulating factor (M-CSF) This suggests that EGR-1 couldbe regulated by the expression of IFI16 Treatment with COLEincreased as well the expression of NFYA encoding the Asubunit of the nuclear transcription factor Y whose synthesiswas previously reported during the terminal differentiationof monocytes to macrophages [38 41] The downregula-tion of FOXP1 in COLE treated K562 cells is consistentwith monocytemacrophage differentiation since it has beendemonstrated that the expression of FOXP1 was markedlydecreased in monocyte-induced differentiation of HL-60cells as well as in human peripheral blood monocytes and
Oxidative Medicine and Cellular Longevity 13
that the overexpression of FOXP1 prevented the morphologicmacrophage-like differentiation [42 43]
Monocytes and macrophages have several functionalcharacteristics including cell adhesion migration chemo-taxis and phagocytic activity Interestingly an enhancedexpression of the chemokines genes CXCL8 (IL8) CXCL2andCXCL3was found in COLE treated cells IL-8 productionhas been observed in vitro in a wide variety of cells includingmonocytes T lymphocytes neutrophils vascular endothe-lial cells dermal fibroblasts keratinocytes hepatocytes andhuman gastric cancer cells while the production of CXCL2andCXCL3was only described inmonocytes fibroblasts andendothelial cells [44 45] Another gene related to chemotaxiswas upregulated NUP85 which is an essential component ofthe nuclear pore complex and was reported to be involved inCCR2-mediated chemotaxis of monocytes [46]
COLE treatment also enhanced the expression of somegenes related to phagocytosis AP1G1 andRab proteins relatedgenes AP1G1 is a subunit of clathrin-associated adaptorprotein complex 1 whose role has been recently highlightedfor efficient phagocytosis at an early stage of phagosomeformation since it participates in the extension of the phago-cytic cup [47 48] The small GTPases Rab are key regulatorsof intracellular membrane trafficking Recent studies havedemonstrated that several RAB proteins play an importantrole in phagocytosis RAB proteins such as RAB11 Rab5 andRab 21 participate in the phagosome formation and matura-tion and are necessary for the phagocytic activity [48ndash51]
Consistently with the increase in Annexin V positivecells (Figure 4) treatment with COLE upregulated someproapoptotic genes such as CASP6 CASP8 DFFA and BIDand downregulated the apoptosis suppressor BCL2 as wellas the caspase inhibitors genes (Table 3) Olive leaf waspreviously shown to induce morphological changes thatare characteristic of apoptosis in leukemia cells but themechanisms underlying this effect were not well investi-gated [13 21] The increase in the proportion of apoptoticcells observed from the 4th day of treatment with COLEcould be explained by the programmed death of the fullydifferentiated cells (Figure 4) In fact this increase wasaccompanied by an important decrease in cell viability aswell as a diminution in CD14 expression (Figures 3(b) and5(a)) Elsewhere it is well established that in hematopoietictissues apoptosis is coupled to terminal differentiation ofmyeloid progenitors even though themechanism responsiblefor the activation of apoptosis during myeloid maturationis still poorly understood [52 53] It is also noteworthy tomention that several reports have highlighted the role ofcaspases in the terminal differentiation of a variety of celltypes [54] One example is the differentiation of human bloodmonocytes into macrophages a process that is blocked bysynthetic caspase inhibitors [55] It has been also reportedthat caspase 8 deletion in bone-marrow cells resulted in arrestof hemopoietic progenitor functioning and that its deletionin cells of the myelomonocytic lineage led to the arrest ofdifferentiation into macrophages and consequently to celldeath [56]
Among the differentially expressed genes in COLEtreated cells (Table 3) genes encoding the mitogen-activated
protein kinase kinase kinases (MAPKKKs) MAP3K2MAP3K5 and MAP3K7 were highly expressed whileMAP2K5 and MAPK14p38-120572 were downregulated TheMAPKKKs act at the upstream of the MAP kinase cascadewhich regulates important cellular processes such as geneexpression cell proliferation differentiation cell survivaland death The MAP3K5 activates the c-Jun N-terminalkinase (JNK) and p38 mitogen-activated protein kinases[57] while the MAP3K2 is reported to preferentially activateJNK [58 59] However the significant decrease in theMAPK14p38-120572 may suggest that the effect of COLE onK562 cells is likely to be mediated by the JNK MAPKsrather than the p38 MAPKs JNK plays an important rolein apoptosis pathways weather by activating proapoptoticor antiapoptotic genes Particularly it was shown thatprolonged but not transient JNK activation promotes thecell death [60] Our results showed that JKAMP gene whichencodes for a membrane-anchored regulator of the durationof JNK1 activity was upregulated (154 plusmn 0051) The elevatedexpression of JKAMP as reported by Kadoya et al [61]results in sustained JNK activity These findings may indicatethe potential role of JNK pathway in the COLE-inducedapoptosis in K562 cells
Thanks to their large regulatory domains some MAP-KKKs can interact with upstream regulators have functionssuch as ubiquitylation and be activated by relief of autoin-hibition and oligomerization [62] In this context MAP3K2has been reported to play an important role in NF-kappaBsignaling pathway by activating the I-kappaB kinases whichin turn phosphorylate the inhibitory factors of the nuclearfactor kappaB (NF-kappaB) resulting in their rapid ubiq-uitination and the liberation of NF-kappaB complex whichtranslocate from the cytoplasm to the nucleus [63] MAP3K7in association with other proteins could also be required forthe activation of NF-kappaB According to the GO analysis(Table S1) COLE treatment upregulated genes implicatedin NF-kappaB-related categories such as positive regulationof NF-kappaB transcription factor activity I-kappaB kinase-NF-kappaB cascade and its positive regulation and I-kappaBphosphorylation Genes encoding the signal transducers inthe NF-kappaB pathway TRAF5 and TRAF6 as well asthe activator of NF-kappaB signaling SNIP1 were upregu-lated Interestingly an increased expression of NFKB1 geneencoding for the precursor of the mature NF-kappaB p50was detected in COLE treated cells compared to untreatedK562 cells NFKB is well recognized as a central activator ofthe antiapoptotic cascades in response to external stimuli orintrinsic immune reactions and its prosurvival activity hasbeen implicated in a variety of biological processes [60 64]In the B cell lineage the activity of NF-120581B is required forthe completion of various developmental stages includingdifferentiation and the response of these cells to antigens [65]NF-kappaB also plays an important role in the survival anddevelopment of T cells [66] and has been recently reported tomediate the differentiation of mesenchymal stem cells [67]
Several studies have demonstrated a crosstalk betweenthe NF-kappaB and JNK pathways seen as an inhibitoryeffect of NF-kappaB on apoptosis through the suppressionof JNK activity and that this suppressive effect may occur
14 Oxidative Medicine and Cellular Longevity
through different mechanisms [60] This suggests that thebalance between JNK and NF-kappaB activities is crucial todetermine the cell fate survival or deathWehypothesize herethat COLE-induced apoptosis or differentiation is likely to bedetermined by the crosstalk between these two pathways inK562 cells Further investigations regarding protein expres-sion and activity are needed to confirm these findings
5 Conclusions
Our study demonstrates for the first time that olive leafextract exhibits an antileukemia effect on the human chronicmyeloid leukemia cells Olive leaf extract was shown toinhibit the proliferation of K562 cells by inducing cell cyclearrest apoptosis and differentiation toward the monocytelineage The induced expression of molecules involved indifferentiation toward the monocytemacrophage lineage aswell asmolecules related to apoptosis and cell cycle regulationconfirmed these findings and provides insights into themechanism by which olive leaf exhibits its antileukemiaeffect
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported by the JICA-JST Science andTechnology Research Partnership for Sustainable Develop-ment (SATREPS) Project ldquoValorization of Bio-Resources inSemi-Arid and Arid Land for Regional Developmentrdquo
References
[1] M Leszczyniecka T Roberts P Dent S Grant and P B FisherldquoDifferentiation therapy of human cancer basic science andclinical applicationsrdquo Pharmacology and Therapeutics vol 90no 2-3 pp 105ndash156 2001
[2] A I Spira and M A Carducci ldquoDifferentiation therapyrdquoCurrent Opinion in Pharmacology vol 3 no 4 pp 338ndash3432003
[3] S Nobili D Lippi E Witort et al ldquoNatural compounds forcancer treatment and preventionrdquo Pharmacological Researchvol 59 no 6 pp 365ndash378 2009
[4] S Ramos ldquoEffects of dietary flavonoids on apoptotic pathwaysrelated to cancer chemopreventionrdquo Journal of NutritionalBiochemistry vol 18 no 7 pp 427ndash442 2007
[5] V Hajhashemi G Vaseghi M Pourfarzam and A AbdollahildquoAre antioxidants helpful for disease preventionrdquo Research inPharmaceutical Sciences vol 5 no 1 pp 1ndash8 2010
[6] M Bouaziz and S Sayadi ldquoIsolation and evaluation of antiox-idants from leaves of a Tunisian cultivar olive treerdquo EuropeanJournal of Lipid Science andTechnology vol 107 no 7-8 pp 497ndash504 2005
[7] A P Pereira I C F R Ferreira F Marcelino et al ldquoPhenoliccompounds and antimicrobial activity of olive (Olea europaeaL Cv Cobrancosa) leavesrdquo Molecules vol 12 no 5 pp 1153ndash1162 2007
[8] L I Somova F O Shode P Ramnanan and A Nadar ldquoAnti-hypertensive antiatherosclerotic and antioxidant activity oftriterpenoids isolated from Olea europaea subspecies africanaleavesrdquo Journal of Ethnopharmacology vol 84 no 2-3 pp 299ndash305 2003
[9] V Micol N Caturla L Perez-Fons V Mas L Perez andA Estepa ldquoThe olive leaf extract exhibits antiviral activityagainst viral haemorrhagic septicaemia rhabdovirus (VHSV)rdquoAntiviral Research vol 66 no 2-3 pp 129ndash136 2005
[10] J Wainstein T Ganz M Boaz et al ldquoOlive leaf extract as ahypoglycemic agent in both human diabetic subjects and inratsrdquo Journal of Medicinal Food vol 15 no 7 pp 605ndash610 2012
[11] L Seddik TM Bah A AouesM Slimani andM BenderdourldquoElucidation of mechanisms underlying the protective effects ofolive leaf extract against lead-induced neurotoxicity in Wistarratsrdquo Journal of Toxicological Sciences vol 36 no 6 pp 797ndash809 2011
[12] Z Bouallagui J Han H Isoda and S Sayadi ldquoHydroxytyrosolrich extract from olive leaves modulates cell cycle progressionin MCF-7 human breast cancer cellsrdquo Food and ChemicalToxicology vol 49 no 1 pp 179ndash184 2011
[13] R Fares S Bazzi S E Baydoun and R M Abdel-Massih ldquoTheantioxidant and anti-proliferative activity of the Lebanese Oleaeuropaea extractrdquo Plant Foods for Human Nutrition vol 66 no1 pp 58ndash63 2011
[14] V Goulas V Exarchou A N Troganis et al ldquoPhytochemicalsin olive-leaf extracts and their antiproliferative activity againstcancer and endothelial cellsrdquo Molecular Nutrition and FoodResearch vol 53 no 5 pp 600ndash608 2009
[15] M de Bock E B Thorstensen J G Derraik H V HendersonP L Hofman and W S Cutfield ldquoHuman absorption andmetabolism of oleuropein and hydroxytyrosol ingested as olive(Olea europaea L) leaf extractrdquo Molecular Nutrition and FoodResearch vol 57 no 11 pp 2079ndash2085 2013
[16] R Briante M Patumi S Terenziani E Bismuto F Febbraioand R Nucci ldquoOlea europaea L leaf extract and derivativesantioxidant propertiesrdquo Journal of Agricultural and Food Chem-istry vol 50 no 17 pp 4934ndash4940 2002
[17] O Benavente-Garcıa J Castillo J Lorente A Ortuno and J ADel Rio ldquoAntioxidant activity of phenolics extracted from Oleaeuropaea L leavesrdquo Food Chemistry vol 68 no 4 pp 457ndash4622000
[18] S H Omar ldquoOleuropein in olive and its pharmacologicaleffectsrdquo Scientia Pharmaceutica vol 78 no 2 pp 133ndash154 2010
[19] H Jemai M Bouaziz I Fki A El Feki and S SayadildquoHypolipidimic and antioxidant activities of oleuropein and itshydrolysis derivative-rich extracts from Chemlali olive leavesrdquoChemico-Biological Interactions vol 176 no 2-3 pp 88ndash982008
[20] R Garcıa-Villalba M Larrosa S Possemiers F A Tomas-Barberan and J C Espın ldquoBioavailability of phenolics from anoleuropein-rich olive (Olea europaea) leaf extract and its acuteeffect on plasma antioxidant status comparison between pre-and postmenopausal womenrdquo European Journal of Nutrition2013
[21] J Anter Z Fernandez-Bedmar M Villatoro-Pulido et al ldquoApilot study on the DNA-protective cytotoxic and apoptosis-inducing properties of olive-leaf extractsrdquo Mutation Researchvol 723 no 2 pp 165ndash170 2011
[22] J A Sutherland A R Turner and P Mannoni ldquoDifferen-tiation of K562 leukemia cells along erythroid macrophage
Oxidative Medicine and Cellular Longevity 15
and megakaryocyte lineagesrdquo Journal of Biological ResponseModifiers vol 5 no 3 pp 250ndash262 1986
[23] K Schwenke H-P Peterson K-H Wangenheim and L EFeinendegen ldquoInduction of differentiation in erythroleukemicK562 cells by 120574-irradiationrdquo Leukemia Research vol 19 no 12pp 955ndash961 1995
[24] V N Enujiugha J Y Talabi S A Malomo and A I OlagunjuldquoDPPH radical scavenging capacity of phenolic extracts fromAfrican Yam Bean (Sphenostylis stenocarpa)rdquo Food and Nutri-tion Sciences vol 3 pp 7ndash13 2012
[25] S A Mijatovic G S Timotijevic D M Miljkovic et alldquoMultiple antimelanoma potential of dry olive leaf extractrdquoInternational Journal of Cancer vol 128 no 8 pp 1955ndash19652011
[26] L Abaza T P N Talorete P Yamada Y KuritaM Zarrouk andH Isoda ldquoInduction of growth inhibition and differentiation ofhuman leukemia HL-60 cells by a Tunisian Gerboui olive leafextractrdquo Bioscience Biotechnology and Biochemistry vol 71 no5 pp 1306ndash1312 2007
[27] B Tunca G Tezcan G Cecener et al ldquoOlea europea leafextract alters microRNA expression in human glioblastomacellsrdquo Journal of Cancer Research and Clinical Oncology vol 138no 11 pp 1831ndash1844 2012
[28] S Matsuoka M Huang and S J Elledge ldquoLinkage of ATM tocell cycle regulation by the Chk2 protein kinaserdquo Science vol282 no 5395 pp 1893ndash1897 1998
[29] C Frazer and P G Young ldquoPhosphorylation mediated regu-lation of Cdc25 activity localization and stabilityrdquo in ProteinPhosphorylation in Human Health C Huang Ed pp 395ndash436InTech 2012
[30] N Debili C Robin V Schiavon et al ldquoDifferent expression ofCD41 on human lymphoid andmyeloid progenitors from adultsand neonatesrdquo Blood vol 97 no 7 pp 2023ndash2030 2001
[31] H Nakajima ldquoRole of transcription factors in differentiationand reprogramming of hematopoietic cellsrdquo Keio Journal ofMedicine vol 60 no 2 pp 47ndash55 2011
[32] Y Fukuchi F Shibata M Ito et al ldquoComprehensive analysis ofmyeloid lineage conversion using mice expressing an inducibleform of CEBP120572rdquoThe EMBO Journal vol 25 no 14 pp 3398ndash3410 2006
[33] R A Shivdasani ldquoMolecular and transcriptional regulation ofmegakaryocyte differentiationrdquo Stem Cells vol 19 no 5 pp397ndash407 2001
[34] K Terui Y Takahashi J Kitazawa T TokiM Yokoyama and EIto ldquoExpression of transcription factors during megakaryocyticdifferentiation of CD34+ cells from human cord blood inducedby thrombopoietinrdquo Tohoku Journal of Experimental Medicinevol 192 no 4 pp 259ndash273 2000
[35] J A Trapani M Dawson V A Apostolidis and K A BrowneldquoGenomic organization of IFI16 an interferon-inducible genewhose expression is associated with human myeloid cell differ-entiation correlation of predicted protein domains with exonorganizationrdquo Immunogenetics vol 40 no 6 pp 415ndash424 1994
[36] M J Dawson N J Elwood R W Johnstone and J A TrapanildquoThe IFN-inducible nucleoprotein IFI 16 is expressed in cellsof the monocyte lineage but is rapidly and markedly down-regulated in other myeloid precursor populationsrdquo Journal ofLeukocyte Biology vol 64 no 4 pp 546ndash554 1998
[37] M Gariglio M De Andrea M Lembo et al ldquoThe murinehomolog of the HIN 200 family Ifi 204 is constitutivelyexpressed in myeloid cells and selectively induced in the
monocytemacrophage lineagerdquo Journal of Leukocyte Biologyvol 64 no 5 pp 608ndash614 1998
[38] A F Valledor F E Borras M Cullell-Young and A CeladaldquoTranscription factors that regulate monocytemacrophage dif-ferentiationrdquo Journal of Leukocyte Biology vol 63 no 4 pp405ndash417 1998
[39] K Krishnaraju B Hoffman and D A Liebermann ldquoEarlygrowth response gene 1 stimulates development of hematopoi-etic progenitor cells along the macrophage lineage at theexpense of the granulocyte and erythroid lineagesrdquo Blood vol97 no 5 pp 1298ndash1305 2001
[40] J Dauffy G Mouchiroud and R P Bourette ldquoThe interferon-inducible gene Ifi204 is transcriptionally activated in responseto M-CSF and its expression favors macrophage differentiationin myeloid progenitor cellsrdquo Journal of Leukocyte Biology vol79 no 1 pp 173ndash183 2006
[41] G Marziali E Perrotti R Ilari U Testa E M Coccia andA Battistini ldquoTranscriptional regulation of the ferritin heavy-chain gene the activity of the CCAAT binding factor NF-Y ismodulated in heme-treated Friend leukemia cells and duringmonocyte-to-macrophage differentiationrdquo Molecular and Cel-lular Biology vol 17 no 3 pp 1387ndash1395 1997
[42] C Shi X Zhang Z Chen et al ldquoIntegrin engagement regulatesmonocyte differentiation through the forkhead transcriptionfactor Foxp1rdquo Journal of Clinical Investigation vol 114 no 3 pp408ndash418 2004
[43] C Shi M Sakuma T Mooroka et al ldquoDown-regulation of theforkhead transcription factor Foxp1 is required for monocytedifferentiation andmacrophage functionrdquoBlood vol 112 no 12pp 4699ndash4711 2008
[44] D F Smith E Galkina K Ley and Y Huo ldquoGRO familychemokines are specialized for monocyte arrest from flowrdquoThe American Journal of Physiology Heart and CirculatoryPhysiology vol 289 no 5 pp H1976ndashH1984 2005
[45] J J Oppenheim O M Z Howard and E Goetzl ChemotacticFactors Neuropeptides and Other Ligands for Seven Transmem-brane Receptors 2000
[46] Y Terashima N Onai M Murai et al ldquoPivotal function forcytoplasmic protein FROUNT in CCR2-mediated monocytechemotaxisrdquo Nature Immunology vol 6 no 8 pp 827ndash8352005
[47] Y Lefkir M Malbouyres D Gotthardt et al ldquoInvolvement ofthe AP-1 adaptor complex in early steps of phagocytosis andmacropinocytosisrdquo Molecular Biology of the Cell vol 15 no 2pp 861ndash869 2004
[48] F Niedergang and P Chavrier ldquoSignaling and membranedynamics during phagocytosis many roads lead to the phago-somerdquo Current Opinion in Cell Biology vol 16 no 4 pp 422ndash428 2004
[49] D Cox D J Lee B M Dale J Calafat and S Green-berg ldquoA Rab11-containing rapidly recycling compartment inmacrophages that promotes phagocytosisrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 97 no 2 pp 680ndash685 2000
[50] O V Vieira C Bucci R E Harrison et al ldquoModulation of Rab5and Rab7 recruitment to phagosomes by phosphatidylinositol3-kinaserdquoMolecular andCellular Biology vol 23 no 7 pp 2501ndash2514 2003
[51] T Khurana J A Brzostowski and A R Kimmel ldquoARab21LIM-onlyCH-LIM complex regulates phagocytosis viaboth activating and inhibitory mechanismsrdquo The EMBO Jour-nal vol 24 no 13 pp 2254ndash2264 2005
16 Oxidative Medicine and Cellular Longevity
[52] S J Martin J G Bradley and T G Cotter ldquoHL-60 cellsinduced to differentiate towards neutrophils subsequently dievia apoptosisrdquo Clinical and Experimental Immunology vol 79no 3 pp 448ndash453 1990
[53] A Benito D Grillot G Nunez and J L Fernandez-Luna ldquoReg-ulation and function of Bcl-2 during differentiation-inducedcell death in HL-60 promyelocytic cellsrdquoThe American Journalof Pathology vol 146 no 2 pp 481ndash490 1995
[54] M Lamkanfi N Festjens W Declercq T V Berghe andP Vandenabeele ldquoCaspases in cell survival proliferation anddifferentiationrdquo Cell Death and Differentiation vol 14 no 1 pp44ndash55 2007
[55] O Sordet C Rebe S Plenchette et al ldquoSpecific involvement ofcaspases in the differentiation of monocytes into macrophagesrdquoBlood vol 100 no 13 pp 4446ndash4453 2002
[56] T-B Kang T Ben-Moshe E E Varfolomeev et al ldquoCaspase-8 serves both apoptotic and nonapoptotic rolesrdquo Journal ofImmunology vol 173 no 5 pp 2976ndash2984 2004
[57] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[58] J Cheng J Yang Y Xia M Karin and B Su ldquoSynergisticinteraction of MEK kinase 2 c-Jun N-terminal kinase (JNK)kinase 2 and JNK1 results in efficient and specific JNK1activationrdquo Molecular and Cellular Biology vol 20 no 7 pp2334ndash2342 2000
[59] J L Blank P Gerwins EM Elliott S Sather andG L JohnsonldquoMolecular cloning of mitogen-activated proteinERK kinasekinases (MEKK) 2 and 3 regulation of sequential phosphoryla-tion pathways involving mitogen-activated protein kinase andc-Jun kinaserdquo Journal of Biological Chemistry vol 271 no 10pp 5361ndash5368 1996
[60] S Papa F Zazzeroni C G Pham C Bubici and G FranzosoldquoLinking JNK signaling to NF-120581B a key to survivalrdquo Journal ofCell Science vol 117 no 22 pp 5197ndash5208 2004
[61] T Kadoya A Khurana M Tcherpakov et al ldquoJAMP a JUB N-terminal kinase 1 (JNK1)-associated membrane protein regu-lates duration of JNK activityrdquo Molecular and Cellular Biologyvol 25 no 19 pp 8619ndash8630 2005
[62] M Qi and E A Elion ldquoMAP kinase pathwaysrdquo Journal of CellScience vol 118 no 16 pp 3569ndash3572 2005
[63] Q Zhao and F S Lee ldquoMitogen-activated protein kinaseERKkinase kinases 2 and 3 activate nuclear factor-120581B through I120581Bkinase-120572 and I120581B kinase-120573rdquoThe Journal of Biological Chemistryvol 274 no 13 pp 8355ndash8358 1999
[64] H Namba V Saenko and S Yamashita ldquoNuclear factor-120581Bin thyroid carcinogenesis and progression a novel therapeutictarget for advanced thyroid cancerrdquo Arquivos Brasileiros deEndocrinologia e Metabologia vol 51 no 5 pp 843ndash851 2007
[65] B Feng S Cheng W S Pear and H-C Liou ldquoNF-kBinhibitor blocks B cell development at two checkpointsrdquoMedi-cal Immunology vol 3 article 1 2004
[66] S Gerondakis and U Siebenlist ldquoRoles of the NF-kappaBpathway in lymphocyte development and functionrdquoCold SpringHarbor Perspectives in Biology vol 2 no 5 Article ID a0001822010
[67] K P Bhat V Balasubramaniyan B Vaillant et al ldquoMesenchy-mal differentiation mediated by NF-kB promotes radiationresistance in glioblastomardquo Cancer Cell vol 24 no 3 pp 331ndash346 2013
Figure 1 HPLC chromatogram of Chemlali Olive Leaf Extract (COLE) at (a) 254 nm and (b) 330 nm Extraction was conducted with 70ethanol Peaks 1 Hydroxytyrosol 2 Verbascoside 3 Luteolin-7-glucoside 4 Apigenin-7-glucoside 5 Oleuropein 6 Luteolin 7 Apigenin
caused a significant decrease of the proliferation of K562 cellsin a dose dependent manner After 72 h of treatment with150 120583gmL of COLE the cell proliferation was inhibited to17 compared with the control cells (Figure 2(a))
Observation of the morphological changes of K562 cellsafter treatment with the different concentrations revealed anincrease in cell size especially in cells treated with 150120583gmLof COLE compared with control cells which kept the samemorphology and cell size (Figure 2(b))
33 COLE Inhibits the Growth Rate of K562 Cells To under-stand if the antiproliferative effect was led by cell death orgrowth inhibition we determined the cell number and cellviability after treatment with COLE by flow cytometry Thenumbers of K562 cells treated with 100 120583gmL and 150 120583gmLwere significantly less than the number of control cellsK562 cells treated with COLE exhibited a slow cell growthcompared to the control (Figure 3(a))
Results (Figure 3(b)) showed that treatment with150 120583gmL of COLE caused a slight decrease of the viabilityof K562 cells during the first 3 days with keeping more than
80 of viable cells Then the decrease became drastic andonly 20 of live cells remain at the 6th day of treatment bythe same concentration The viability of cells treated with100 120583gmL of COLE started to decrease significantly fromthe 4th day while treatment with 50120583gmL did not affect thecell viability during the whole period Taken together theseresults suggested that the antiproliferative effect of COLEat the concentrations of 100 120583gmL and 150 120583gmL mightbe conducted in part by inhibiting the growth rate and to alesser extent by causing cell death
34 COLE Modulates Cell Cycle Progression in K562 CellsSinceCOLE reduced the growth ofK562 cells we investigatedits effect on the cell cycle progression During the 1st and2nd day K562 cells treated with 100 and 150 120583gmL werearrested at G0G1 During the following days results showeda significant increase of G2M population in cell treated with100 and 150 120583gmL of COLE In the case of cells treatedwith 150120583gmL of COLE this increase was associated with asignificant decrease of G0G1 cell population throughout the3rd and 4th days of treatment (Table 2)
Oxidative Medicine and Cellular Longevity 5
0
20
40
60
80
100
120
24 48 72
Cel
l pro
lifer
atio
n (
of co
ntro
l)
Control50120583gmL75120583gmL
100120583gmL125120583gmL150120583gmL
lowastlowastlowast lowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowastlowastlowast
(h)
(a)
Control 50120583gmL
100120583gmL 150120583gmL
(b)
Figure 2 (a) Effect ofChemlaliOlive Leaf Extract (COLE) on the proliferation ofK562 cellsThe cells were treatedwith various concentrations(50 75 100 125 and 150120583gmL) of COLE for 24 48 and 72 h Cell proliferation was measured byMTT assay Control represents cells treatedwith 03 ethanol in medium Results are represented as the mean plusmn SD of three independent experiments lowast lowastlowast significantly differentfrom the control (119875 lt 005 and 119875 lt 001 resp) (b) Morphological observations of K562 cells after treatment with various concentrationsof Chemlali Olive Leaf Extract (COLE) for 72 h Cells were observed under a phase contrast microscope at 100x magnification Scale barsrepresent 100 120583m
0
2
4
6
8
10
12
14
16
18
20
Day 0 Day 1 Day 2 Day 3 Day 4 Day 6
Num
ber o
f via
ble c
ells
(times106)
Control 50120583gmL100 120583gmL 150120583gmL
(a)
0102030405060708090100
Day 0 Day 1 Day 2 Day 3 Day 4 Day 6
Viab
ility
()
Control50120583gmL
100 120583gmL150120583gmL
lowastlowastlowastlowast
lowastlowast
lowastlowast
lowastlowast
lowast lowast
(b)
Figure 3 Effect of Chemlali Olive Leaf Extract (COLE) on the cell number and viability of K562 cells (a) Number of viable cells aftertreatment with COLE after incubation up to 6 days (b) Percentage of viability in K562 cells treated with COLE K562 cells were treated at afinal concentration of 50 100 and 150120583gmL of COLE and incubated for different periods Cell number and cell viability were measured byflow cytometry Control represents cells treated with 03 ethanol in medium Results are represented as the mean plusmn SD of three independentexperiments lowast lowastlowast significantly different from the control (119875 lt 005 and 119875 lt 001 resp)
35 COLE Induces Apoptosis in K562 Cells The decreasein cell viability (Figure 3(b)) caused by COLE treatmentprompted us to investigate if the extract induces apoptosis inK562 cells Apoptosis is an important and active regulatorypathway of cell growth and proliferation resulting in somecharacteristic physiological changes Among these external-ization of phosphatidylserine (PS) is easily detected by flowcytometry after binding to the labeled Annexin VThe resultsindicated an increase in Annexin V positive cells from the1st day of incubation with 150 120583gmL of COLE in comparisonwith control cells (Figure 4) The percentage of Annexin V
cells remained stable during the first 3 days of treatmentwith 20 of total treated cells and interestingly increasedin the 4th and 6th days with 45 and 60 respectivelyTreatment with 100 120583gmL caused the apparition of apoptoticcells from the 4th day of incubation with a proportionof 162 This population increased in the 6th day up to267No significant difference in apoptotic cells amountwasdetected when cells were treated with 50 120583gmL until 6 daysof incubation
6 Oxidative Medicine and Cellular Longevity
Table 2 The distribution of cell cycle in K562 cells treated with Chemlali Olive Leaf Extract (COLE)
Results are represented as the means plusmn SD of three independent experimentslowast and lowastlowast mean that difference between control and treated cells in each phase (G0G1 S and G2M) is statistically significant at 119875 lt 005 and 119875 lt 001respectively
36 Impact of COLE on the Differentiation Capacity of K562Cells The reduction in cell growth as well as the morpho-logical changes observed in COLE treated cells led us tothink about the differentiation assessment K562 cells arepluripotent malignant cells that spontaneously differentiatealong erythroid macrophage and megakaryocyte lineages[22] Treated cells were harvested and analyzed for theexpression of lineage differentiation markers The analysiswas performed by flow cytometry at different days on themonocytemacrophage marker CD14 on the granulocytemarker CD11b on the erythrocyte marker GPA and on themegakaryocyte marker CD41 The results showed a signifi-cant increase in the expression of CD14 marker from day 1until day 6 in the cells treated with 150120583gmL suggestingthe commitment of K562 cells to the monocytemacrophagelineage (Figure 5(a)) A slight increase of the expression ofCD11b was also detected at this concentration (Figure 5(b))Treatment with 100 120583gmL showed a gain in the expression ofCD14 on the 1st and 2nd day and an augmentation of CD11bexpression from the 3rd day of treatment Treatment with50120583gmL did not show any significant effect on the differenti-ationmarkersCD14 andCD11b Even after 6 days of treatmentwith 150 120583gmL of COLE we could not detect the erythroidmarker at all the tested concentrations (Figure 5(d)) How-ever an instant increase in the megakaryocytic marker wasdetected on the 1st day of treatment followed by a drasticdecrease lower than the control during the following days(Figure 5(c))
37 Gene Expression Profile of COLE Treated K562 Cells Tofurther elucidate the mechanism by which COLE inducesapoptosis and differentiation of K562 cells we investigatedthe changes in gene expression profiles in treated cells usingHG219 GeneChip array Microarray analysis was performedon K562 cells at the 3rd day of treatment based on the obser-vation that such timing generated a significant expression ofthe differentiation markers Genes with more than 15-foldchange in expression levels between control and 150 120583gmLofCOLE treated cells were classified into categories accordingto the cell biological processes According to the GO analysis
0
10
20
30
40
50
60
70
Day 1 Day 2 Day 3 Day 4 Day 6
Ann
exin
pos
itive
(
)
Control50120583gmL
100 120583gmL150120583gmL
lowastlowast lowastlowast lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
Figure 4 Induction of apoptosis in K562 cells treatedwith ChemlaliOlive Leaf Extract (COLE) Cells were treated with 50 100 and150 120583gmL of COLE and incubated for different time At the indi-cated time K562 cells were stained with Annexin V and analyzed byflow cytometry Control represents cells treated with 03 ethanolin medium Results are represented as the mean plusmn SD of threeindependent experiments lowast lowastlowast significantly different from thecontrol (119875 lt 005 and 119875 lt 001 resp)
results (Tables S1 and S2 in Supplementary Material avail-able online at httpdxdoiorg1011552014927619) COLEtreatment induced the upregulation of genes involved inhematopoiesis such as CTNNB1 SH2B3 CIAPIN1 RPA1 andgenes implicated in cell differentiation and its regulation suchasMCL1CTNNB1 andCCNE1 Interestingly genes related tomyeloid cell differentiation such as IFI16ACIN1 andCASP8and particularly genes engaged in monocyte differentiationwere represented in the upregulated categories Moreoveran increased expression of genes involved in chemotaxissuch as NUP85 HRAS IL8 CXCL2 and CXCL3 and genesrelated to cytokine production was also detected Amongupregulated genes AP1G1 RABEP1 RAB5C RAB21 EPS15L1and CORO1C were also found involved in the process ofendocytosis and phagocytosis as well as genes related toprotein transport and genes implicated in cell adhesion suchas ICAM3 HSPB11 and PNN
Conversely the erythrocyte differentiation categorieswere represented in the list of the decreased genes (Table S2)
Figure 5 Expression of differentiationmarkers onK562 cells treatedwith Chemlali Olive Leaf Extract (COLE) up to 6 days Cells were treatedwith 50 100 and 150 120583gmL of COLE and incubated for different periods Control cells were treated with ethanol at a final concentration of03 Expression of cell surface markers was detected by flow cytometry (a) Expression of CD14 (monocytemacrophage specific marker)(b) Expression of CD11b (granulocytemonocyte specific marker) (c) Expression of CD41 (megakaryocyte specific marker) (d) Expressionof glycophorin A (GPA) (erythrocyte specific marker) Results are represented as the mean plusmn SD of three independent experiments lowast lowastlowastsignificantly different from the control at 119875 lt 005 and 119875 lt 001 respectively
including GYPA HBE1 FECH and ALAS2 which is consis-tent with the decreased expression of erythrocyte differenti-ation marker in the flow cytometry results (Figure 5(d))
Analysis of the microarray data showed the upregu-lation of several proapoptotic genes and genes involvedin the regulation of mitochondrial membrane permeabilitysuch as CASP6 CASP8 DFFA and BID On the otherhandexpression of genes negatively regulating the apoptosis
and those inhibiting the caspase activity was significantlydecreased such as IGF1R HSPA5 and BCL2
Among the upregulated genes we found those related tothe NF-kappaB cascade as well as to the MAPKKK cascadeand the Wnt receptor signaling pathway
Categories related to cell cycle regulation and cell pro-liferation were represented by both upregulated and down-regulated genes Finally some transcription factors werealso downregulated such as FOXP1 whereas others wereupregulated like EGR1 and NFYA
The list of selected genes is presented in Table 3 Fromthis list it is clear that the differentially expressed genes aftertreatment with 100 and 150120583gmL have mostly the sametendency which could validate the obtained results
12 Oxidative Medicine and Cellular Longevity
4 Discussion
Previous reports had indicated that olive leaf extract inducesapoptosis in the human leukemic cell lines HL-60 and Jurkatcells [13 21] In this study we give evidence that COLEexhibits its antileukemia effect by both inducing apoptosisand promoting differentiation of the multipotent humanleukemia K562 cells
Induction of differentiation as well as apoptosis has beenfrequently reported to be associatedwith a loss of proliferativecapacity of the cells It has been reported that olive leaf extractinhibits the proliferation of some cancer cell lines such asB16 melanoma cells HL-60 leukemia cells the breast cancerMCF-7 cells and the glioblastoma cells [12 25ndash27] Ourresults indicate that COLE inhibits the proliferation of K562cells in a dose dependent manner (Figure 2(a)) Howeverdespite the drastic decrease in cell proliferation the viabilityof cells treated with the highest concentration of COLE(150 120583gmL) remains relatively high (80) for up to 3 daysof treatment (Figure 3(b))
Since it is well established that the arrest of the cellcycle progression allows the cells to follow other processessuch as apoptosis and differentiation we then explored theeffect of COLE on the cell cycle distribution (Table 2) Oliveleaf extract has been shown to cause cell cycle arrest atG0G1 phase in both the breast cancer MCF-7 cells and themelanoma B16 cells [12 25] However the treatment timesapplied in both of these studies were limited to 48 h and 18 hrespectively In this study COLE treated cells were arrested atG0G1 on the 1st and 2ndday of treatment which is consistentwith the reported studies Prolonged incubation with COLEshowed an arrest of cell cycle at G2M phase (3rd and 4thday of incubation) The microarray analysis performed forthe 3rd day of treatment revealed a significant increase ofCHEK2 gene expression in COLE treated cells The encodedprotein regulates the cell cycle checkpoint arrest throughthe inhibition of the activity of CDC25A CDC25B andCDC25C [28] CDC25 proteins trigger the entry into mitosisat different points of the cell cycle by activating theCdk-cyclincomplexes CDC25A acts early in the cell cycle regulatingthe G1S transition whereas CDC25B and CDC25C act atG2M [29] Interestingly our results showed a decrease in theexpression of CDC25C gene in COLE treated cells associatedwith an increase in the expression of CDC25A which mayexplain the cell cycle arrest at G2M phase observed at the3rd day of treatment with COLE
Treatment with COLE increased the expression of CD14on the cell surface of treated cells indicating the differenti-ation of K562 cells toward the mono-macrophage lineage(Figure 5(a)) The increase in the percentage of cells positivefor CD11b (Figure 5(b)) confirms this hypothesis In factCD11b is expressed on both monocytes and granulocytesThus together with the increase of CD14 expression theexpression of CD11b suggests that treatment with COLEpromotes the commitment of K562 cells into the mono-cytemacrophage lineage at the expense of the granulocyticdifferentiationThe instant increase of the expression of CD41
on the 1st day of treatment and its absence during the fol-lowing days (Figure 5(c)) could be consistent with monocytedifferentiation instead of megakaryocyte differentiation sinceit has been reported that 120572IIb promoter which initiates thetranscription of CD41 is transcriptionally active in pluripo-tent myeloid progenitors in early stages of erythropoiesisand all along the megakaryocytic differentiation as well asto a lesser extent in the early stages of myelomonocyticdifferentiation and the late stages of erythropoiesis [30] Thisinstant increase could be also explained by the phenomenonof lineage conversion where some hematopoietic progenitorscan be converted into lineages other than their own bythe ectopic expression of some transcription factors [31]In this regard it was previously reported that commonlymphoid progenitors megakaryocyte erythroid progenitorsearly B cells and early T cells could be converted to thegranulocytemonocyte lineage by the induction of CEBP120572activity [32] Microarray results support the absence ofmegakaryocyte differentiation in the 3rd day of treatment Infact data showed a decrease in the expression of genes relatedtomegakaryopoiesis such as the essential regulator of plateletrelease NFE2 and its main candidate target gene TUBB1(encoding the megakaryocyte and platelet specific isoform of120573-tubulin (1205731)) [33] as well as the BACH2 transcription factorreported to be induced during megakaryocyte differentiation[34]
Interestingly results of microarray analysis (Table 3)showed the upregulation of several genes already reportedas key players in monocytemacrophage differentiation inthe COLE treated cells Among these genes the expressionof IFI16 was markedly increased Constitutively expressedin lymphoid cells IFI16 was shown to be associated withthe differentiation of human myeloid cells in response tointerferon gamma [35] Moreover IFI16 has been selectivelyexpressed during the differentiation of myeloid progenitorcells along monocytic lineage while it is downregulatedduring the commitment toward the granulocytic or erythroidlineage suggesting its potential involvement in the differenti-ation andmaturation of the monocytic lineage [36 37] Geneencoding for EGR-1 a positive modulator of macrophagedifferentiation was also significantly upregulated EGR-1 isknown to dictate development of myeloid progenitors alongthe macrophage lineage at the expense of development alongother lineages [38 39] Dauffy et al [40] reported thatIFI16-enforced expression in myeloid progenitors inducedthe expression of EGR-1 and conducted the macrophagedifferentiation in the absence of the macrophage colonystimulating factor (M-CSF) This suggests that EGR-1 couldbe regulated by the expression of IFI16 Treatment with COLEincreased as well the expression of NFYA encoding the Asubunit of the nuclear transcription factor Y whose synthesiswas previously reported during the terminal differentiationof monocytes to macrophages [38 41] The downregula-tion of FOXP1 in COLE treated K562 cells is consistentwith monocytemacrophage differentiation since it has beendemonstrated that the expression of FOXP1 was markedlydecreased in monocyte-induced differentiation of HL-60cells as well as in human peripheral blood monocytes and
Oxidative Medicine and Cellular Longevity 13
that the overexpression of FOXP1 prevented the morphologicmacrophage-like differentiation [42 43]
Monocytes and macrophages have several functionalcharacteristics including cell adhesion migration chemo-taxis and phagocytic activity Interestingly an enhancedexpression of the chemokines genes CXCL8 (IL8) CXCL2andCXCL3was found in COLE treated cells IL-8 productionhas been observed in vitro in a wide variety of cells includingmonocytes T lymphocytes neutrophils vascular endothe-lial cells dermal fibroblasts keratinocytes hepatocytes andhuman gastric cancer cells while the production of CXCL2andCXCL3was only described inmonocytes fibroblasts andendothelial cells [44 45] Another gene related to chemotaxiswas upregulated NUP85 which is an essential component ofthe nuclear pore complex and was reported to be involved inCCR2-mediated chemotaxis of monocytes [46]
COLE treatment also enhanced the expression of somegenes related to phagocytosis AP1G1 andRab proteins relatedgenes AP1G1 is a subunit of clathrin-associated adaptorprotein complex 1 whose role has been recently highlightedfor efficient phagocytosis at an early stage of phagosomeformation since it participates in the extension of the phago-cytic cup [47 48] The small GTPases Rab are key regulatorsof intracellular membrane trafficking Recent studies havedemonstrated that several RAB proteins play an importantrole in phagocytosis RAB proteins such as RAB11 Rab5 andRab 21 participate in the phagosome formation and matura-tion and are necessary for the phagocytic activity [48ndash51]
Consistently with the increase in Annexin V positivecells (Figure 4) treatment with COLE upregulated someproapoptotic genes such as CASP6 CASP8 DFFA and BIDand downregulated the apoptosis suppressor BCL2 as wellas the caspase inhibitors genes (Table 3) Olive leaf waspreviously shown to induce morphological changes thatare characteristic of apoptosis in leukemia cells but themechanisms underlying this effect were not well investi-gated [13 21] The increase in the proportion of apoptoticcells observed from the 4th day of treatment with COLEcould be explained by the programmed death of the fullydifferentiated cells (Figure 4) In fact this increase wasaccompanied by an important decrease in cell viability aswell as a diminution in CD14 expression (Figures 3(b) and5(a)) Elsewhere it is well established that in hematopoietictissues apoptosis is coupled to terminal differentiation ofmyeloid progenitors even though themechanism responsiblefor the activation of apoptosis during myeloid maturationis still poorly understood [52 53] It is also noteworthy tomention that several reports have highlighted the role ofcaspases in the terminal differentiation of a variety of celltypes [54] One example is the differentiation of human bloodmonocytes into macrophages a process that is blocked bysynthetic caspase inhibitors [55] It has been also reportedthat caspase 8 deletion in bone-marrow cells resulted in arrestof hemopoietic progenitor functioning and that its deletionin cells of the myelomonocytic lineage led to the arrest ofdifferentiation into macrophages and consequently to celldeath [56]
Among the differentially expressed genes in COLEtreated cells (Table 3) genes encoding the mitogen-activated
protein kinase kinase kinases (MAPKKKs) MAP3K2MAP3K5 and MAP3K7 were highly expressed whileMAP2K5 and MAPK14p38-120572 were downregulated TheMAPKKKs act at the upstream of the MAP kinase cascadewhich regulates important cellular processes such as geneexpression cell proliferation differentiation cell survivaland death The MAP3K5 activates the c-Jun N-terminalkinase (JNK) and p38 mitogen-activated protein kinases[57] while the MAP3K2 is reported to preferentially activateJNK [58 59] However the significant decrease in theMAPK14p38-120572 may suggest that the effect of COLE onK562 cells is likely to be mediated by the JNK MAPKsrather than the p38 MAPKs JNK plays an important rolein apoptosis pathways weather by activating proapoptoticor antiapoptotic genes Particularly it was shown thatprolonged but not transient JNK activation promotes thecell death [60] Our results showed that JKAMP gene whichencodes for a membrane-anchored regulator of the durationof JNK1 activity was upregulated (154 plusmn 0051) The elevatedexpression of JKAMP as reported by Kadoya et al [61]results in sustained JNK activity These findings may indicatethe potential role of JNK pathway in the COLE-inducedapoptosis in K562 cells
Thanks to their large regulatory domains some MAP-KKKs can interact with upstream regulators have functionssuch as ubiquitylation and be activated by relief of autoin-hibition and oligomerization [62] In this context MAP3K2has been reported to play an important role in NF-kappaBsignaling pathway by activating the I-kappaB kinases whichin turn phosphorylate the inhibitory factors of the nuclearfactor kappaB (NF-kappaB) resulting in their rapid ubiq-uitination and the liberation of NF-kappaB complex whichtranslocate from the cytoplasm to the nucleus [63] MAP3K7in association with other proteins could also be required forthe activation of NF-kappaB According to the GO analysis(Table S1) COLE treatment upregulated genes implicatedin NF-kappaB-related categories such as positive regulationof NF-kappaB transcription factor activity I-kappaB kinase-NF-kappaB cascade and its positive regulation and I-kappaBphosphorylation Genes encoding the signal transducers inthe NF-kappaB pathway TRAF5 and TRAF6 as well asthe activator of NF-kappaB signaling SNIP1 were upregu-lated Interestingly an increased expression of NFKB1 geneencoding for the precursor of the mature NF-kappaB p50was detected in COLE treated cells compared to untreatedK562 cells NFKB is well recognized as a central activator ofthe antiapoptotic cascades in response to external stimuli orintrinsic immune reactions and its prosurvival activity hasbeen implicated in a variety of biological processes [60 64]In the B cell lineage the activity of NF-120581B is required forthe completion of various developmental stages includingdifferentiation and the response of these cells to antigens [65]NF-kappaB also plays an important role in the survival anddevelopment of T cells [66] and has been recently reported tomediate the differentiation of mesenchymal stem cells [67]
Several studies have demonstrated a crosstalk betweenthe NF-kappaB and JNK pathways seen as an inhibitoryeffect of NF-kappaB on apoptosis through the suppressionof JNK activity and that this suppressive effect may occur
14 Oxidative Medicine and Cellular Longevity
through different mechanisms [60] This suggests that thebalance between JNK and NF-kappaB activities is crucial todetermine the cell fate survival or deathWehypothesize herethat COLE-induced apoptosis or differentiation is likely to bedetermined by the crosstalk between these two pathways inK562 cells Further investigations regarding protein expres-sion and activity are needed to confirm these findings
5 Conclusions
Our study demonstrates for the first time that olive leafextract exhibits an antileukemia effect on the human chronicmyeloid leukemia cells Olive leaf extract was shown toinhibit the proliferation of K562 cells by inducing cell cyclearrest apoptosis and differentiation toward the monocytelineage The induced expression of molecules involved indifferentiation toward the monocytemacrophage lineage aswell asmolecules related to apoptosis and cell cycle regulationconfirmed these findings and provides insights into themechanism by which olive leaf exhibits its antileukemiaeffect
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported by the JICA-JST Science andTechnology Research Partnership for Sustainable Develop-ment (SATREPS) Project ldquoValorization of Bio-Resources inSemi-Arid and Arid Land for Regional Developmentrdquo
References
[1] M Leszczyniecka T Roberts P Dent S Grant and P B FisherldquoDifferentiation therapy of human cancer basic science andclinical applicationsrdquo Pharmacology and Therapeutics vol 90no 2-3 pp 105ndash156 2001
[2] A I Spira and M A Carducci ldquoDifferentiation therapyrdquoCurrent Opinion in Pharmacology vol 3 no 4 pp 338ndash3432003
[3] S Nobili D Lippi E Witort et al ldquoNatural compounds forcancer treatment and preventionrdquo Pharmacological Researchvol 59 no 6 pp 365ndash378 2009
[4] S Ramos ldquoEffects of dietary flavonoids on apoptotic pathwaysrelated to cancer chemopreventionrdquo Journal of NutritionalBiochemistry vol 18 no 7 pp 427ndash442 2007
[5] V Hajhashemi G Vaseghi M Pourfarzam and A AbdollahildquoAre antioxidants helpful for disease preventionrdquo Research inPharmaceutical Sciences vol 5 no 1 pp 1ndash8 2010
[6] M Bouaziz and S Sayadi ldquoIsolation and evaluation of antiox-idants from leaves of a Tunisian cultivar olive treerdquo EuropeanJournal of Lipid Science andTechnology vol 107 no 7-8 pp 497ndash504 2005
[7] A P Pereira I C F R Ferreira F Marcelino et al ldquoPhenoliccompounds and antimicrobial activity of olive (Olea europaeaL Cv Cobrancosa) leavesrdquo Molecules vol 12 no 5 pp 1153ndash1162 2007
[8] L I Somova F O Shode P Ramnanan and A Nadar ldquoAnti-hypertensive antiatherosclerotic and antioxidant activity oftriterpenoids isolated from Olea europaea subspecies africanaleavesrdquo Journal of Ethnopharmacology vol 84 no 2-3 pp 299ndash305 2003
[9] V Micol N Caturla L Perez-Fons V Mas L Perez andA Estepa ldquoThe olive leaf extract exhibits antiviral activityagainst viral haemorrhagic septicaemia rhabdovirus (VHSV)rdquoAntiviral Research vol 66 no 2-3 pp 129ndash136 2005
[10] J Wainstein T Ganz M Boaz et al ldquoOlive leaf extract as ahypoglycemic agent in both human diabetic subjects and inratsrdquo Journal of Medicinal Food vol 15 no 7 pp 605ndash610 2012
[11] L Seddik TM Bah A AouesM Slimani andM BenderdourldquoElucidation of mechanisms underlying the protective effects ofolive leaf extract against lead-induced neurotoxicity in Wistarratsrdquo Journal of Toxicological Sciences vol 36 no 6 pp 797ndash809 2011
[12] Z Bouallagui J Han H Isoda and S Sayadi ldquoHydroxytyrosolrich extract from olive leaves modulates cell cycle progressionin MCF-7 human breast cancer cellsrdquo Food and ChemicalToxicology vol 49 no 1 pp 179ndash184 2011
[13] R Fares S Bazzi S E Baydoun and R M Abdel-Massih ldquoTheantioxidant and anti-proliferative activity of the Lebanese Oleaeuropaea extractrdquo Plant Foods for Human Nutrition vol 66 no1 pp 58ndash63 2011
[14] V Goulas V Exarchou A N Troganis et al ldquoPhytochemicalsin olive-leaf extracts and their antiproliferative activity againstcancer and endothelial cellsrdquo Molecular Nutrition and FoodResearch vol 53 no 5 pp 600ndash608 2009
[15] M de Bock E B Thorstensen J G Derraik H V HendersonP L Hofman and W S Cutfield ldquoHuman absorption andmetabolism of oleuropein and hydroxytyrosol ingested as olive(Olea europaea L) leaf extractrdquo Molecular Nutrition and FoodResearch vol 57 no 11 pp 2079ndash2085 2013
[16] R Briante M Patumi S Terenziani E Bismuto F Febbraioand R Nucci ldquoOlea europaea L leaf extract and derivativesantioxidant propertiesrdquo Journal of Agricultural and Food Chem-istry vol 50 no 17 pp 4934ndash4940 2002
[17] O Benavente-Garcıa J Castillo J Lorente A Ortuno and J ADel Rio ldquoAntioxidant activity of phenolics extracted from Oleaeuropaea L leavesrdquo Food Chemistry vol 68 no 4 pp 457ndash4622000
[18] S H Omar ldquoOleuropein in olive and its pharmacologicaleffectsrdquo Scientia Pharmaceutica vol 78 no 2 pp 133ndash154 2010
[19] H Jemai M Bouaziz I Fki A El Feki and S SayadildquoHypolipidimic and antioxidant activities of oleuropein and itshydrolysis derivative-rich extracts from Chemlali olive leavesrdquoChemico-Biological Interactions vol 176 no 2-3 pp 88ndash982008
[20] R Garcıa-Villalba M Larrosa S Possemiers F A Tomas-Barberan and J C Espın ldquoBioavailability of phenolics from anoleuropein-rich olive (Olea europaea) leaf extract and its acuteeffect on plasma antioxidant status comparison between pre-and postmenopausal womenrdquo European Journal of Nutrition2013
[21] J Anter Z Fernandez-Bedmar M Villatoro-Pulido et al ldquoApilot study on the DNA-protective cytotoxic and apoptosis-inducing properties of olive-leaf extractsrdquo Mutation Researchvol 723 no 2 pp 165ndash170 2011
[22] J A Sutherland A R Turner and P Mannoni ldquoDifferen-tiation of K562 leukemia cells along erythroid macrophage
Oxidative Medicine and Cellular Longevity 15
and megakaryocyte lineagesrdquo Journal of Biological ResponseModifiers vol 5 no 3 pp 250ndash262 1986
[23] K Schwenke H-P Peterson K-H Wangenheim and L EFeinendegen ldquoInduction of differentiation in erythroleukemicK562 cells by 120574-irradiationrdquo Leukemia Research vol 19 no 12pp 955ndash961 1995
[24] V N Enujiugha J Y Talabi S A Malomo and A I OlagunjuldquoDPPH radical scavenging capacity of phenolic extracts fromAfrican Yam Bean (Sphenostylis stenocarpa)rdquo Food and Nutri-tion Sciences vol 3 pp 7ndash13 2012
[25] S A Mijatovic G S Timotijevic D M Miljkovic et alldquoMultiple antimelanoma potential of dry olive leaf extractrdquoInternational Journal of Cancer vol 128 no 8 pp 1955ndash19652011
[26] L Abaza T P N Talorete P Yamada Y KuritaM Zarrouk andH Isoda ldquoInduction of growth inhibition and differentiation ofhuman leukemia HL-60 cells by a Tunisian Gerboui olive leafextractrdquo Bioscience Biotechnology and Biochemistry vol 71 no5 pp 1306ndash1312 2007
[27] B Tunca G Tezcan G Cecener et al ldquoOlea europea leafextract alters microRNA expression in human glioblastomacellsrdquo Journal of Cancer Research and Clinical Oncology vol 138no 11 pp 1831ndash1844 2012
[28] S Matsuoka M Huang and S J Elledge ldquoLinkage of ATM tocell cycle regulation by the Chk2 protein kinaserdquo Science vol282 no 5395 pp 1893ndash1897 1998
[29] C Frazer and P G Young ldquoPhosphorylation mediated regu-lation of Cdc25 activity localization and stabilityrdquo in ProteinPhosphorylation in Human Health C Huang Ed pp 395ndash436InTech 2012
[30] N Debili C Robin V Schiavon et al ldquoDifferent expression ofCD41 on human lymphoid andmyeloid progenitors from adultsand neonatesrdquo Blood vol 97 no 7 pp 2023ndash2030 2001
[31] H Nakajima ldquoRole of transcription factors in differentiationand reprogramming of hematopoietic cellsrdquo Keio Journal ofMedicine vol 60 no 2 pp 47ndash55 2011
[32] Y Fukuchi F Shibata M Ito et al ldquoComprehensive analysis ofmyeloid lineage conversion using mice expressing an inducibleform of CEBP120572rdquoThe EMBO Journal vol 25 no 14 pp 3398ndash3410 2006
[33] R A Shivdasani ldquoMolecular and transcriptional regulation ofmegakaryocyte differentiationrdquo Stem Cells vol 19 no 5 pp397ndash407 2001
[34] K Terui Y Takahashi J Kitazawa T TokiM Yokoyama and EIto ldquoExpression of transcription factors during megakaryocyticdifferentiation of CD34+ cells from human cord blood inducedby thrombopoietinrdquo Tohoku Journal of Experimental Medicinevol 192 no 4 pp 259ndash273 2000
[35] J A Trapani M Dawson V A Apostolidis and K A BrowneldquoGenomic organization of IFI16 an interferon-inducible genewhose expression is associated with human myeloid cell differ-entiation correlation of predicted protein domains with exonorganizationrdquo Immunogenetics vol 40 no 6 pp 415ndash424 1994
[36] M J Dawson N J Elwood R W Johnstone and J A TrapanildquoThe IFN-inducible nucleoprotein IFI 16 is expressed in cellsof the monocyte lineage but is rapidly and markedly down-regulated in other myeloid precursor populationsrdquo Journal ofLeukocyte Biology vol 64 no 4 pp 546ndash554 1998
[37] M Gariglio M De Andrea M Lembo et al ldquoThe murinehomolog of the HIN 200 family Ifi 204 is constitutivelyexpressed in myeloid cells and selectively induced in the
monocytemacrophage lineagerdquo Journal of Leukocyte Biologyvol 64 no 5 pp 608ndash614 1998
[38] A F Valledor F E Borras M Cullell-Young and A CeladaldquoTranscription factors that regulate monocytemacrophage dif-ferentiationrdquo Journal of Leukocyte Biology vol 63 no 4 pp405ndash417 1998
[39] K Krishnaraju B Hoffman and D A Liebermann ldquoEarlygrowth response gene 1 stimulates development of hematopoi-etic progenitor cells along the macrophage lineage at theexpense of the granulocyte and erythroid lineagesrdquo Blood vol97 no 5 pp 1298ndash1305 2001
[40] J Dauffy G Mouchiroud and R P Bourette ldquoThe interferon-inducible gene Ifi204 is transcriptionally activated in responseto M-CSF and its expression favors macrophage differentiationin myeloid progenitor cellsrdquo Journal of Leukocyte Biology vol79 no 1 pp 173ndash183 2006
[41] G Marziali E Perrotti R Ilari U Testa E M Coccia andA Battistini ldquoTranscriptional regulation of the ferritin heavy-chain gene the activity of the CCAAT binding factor NF-Y ismodulated in heme-treated Friend leukemia cells and duringmonocyte-to-macrophage differentiationrdquo Molecular and Cel-lular Biology vol 17 no 3 pp 1387ndash1395 1997
[42] C Shi X Zhang Z Chen et al ldquoIntegrin engagement regulatesmonocyte differentiation through the forkhead transcriptionfactor Foxp1rdquo Journal of Clinical Investigation vol 114 no 3 pp408ndash418 2004
[43] C Shi M Sakuma T Mooroka et al ldquoDown-regulation of theforkhead transcription factor Foxp1 is required for monocytedifferentiation andmacrophage functionrdquoBlood vol 112 no 12pp 4699ndash4711 2008
[44] D F Smith E Galkina K Ley and Y Huo ldquoGRO familychemokines are specialized for monocyte arrest from flowrdquoThe American Journal of Physiology Heart and CirculatoryPhysiology vol 289 no 5 pp H1976ndashH1984 2005
[45] J J Oppenheim O M Z Howard and E Goetzl ChemotacticFactors Neuropeptides and Other Ligands for Seven Transmem-brane Receptors 2000
[46] Y Terashima N Onai M Murai et al ldquoPivotal function forcytoplasmic protein FROUNT in CCR2-mediated monocytechemotaxisrdquo Nature Immunology vol 6 no 8 pp 827ndash8352005
[47] Y Lefkir M Malbouyres D Gotthardt et al ldquoInvolvement ofthe AP-1 adaptor complex in early steps of phagocytosis andmacropinocytosisrdquo Molecular Biology of the Cell vol 15 no 2pp 861ndash869 2004
[48] F Niedergang and P Chavrier ldquoSignaling and membranedynamics during phagocytosis many roads lead to the phago-somerdquo Current Opinion in Cell Biology vol 16 no 4 pp 422ndash428 2004
[49] D Cox D J Lee B M Dale J Calafat and S Green-berg ldquoA Rab11-containing rapidly recycling compartment inmacrophages that promotes phagocytosisrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 97 no 2 pp 680ndash685 2000
[50] O V Vieira C Bucci R E Harrison et al ldquoModulation of Rab5and Rab7 recruitment to phagosomes by phosphatidylinositol3-kinaserdquoMolecular andCellular Biology vol 23 no 7 pp 2501ndash2514 2003
[51] T Khurana J A Brzostowski and A R Kimmel ldquoARab21LIM-onlyCH-LIM complex regulates phagocytosis viaboth activating and inhibitory mechanismsrdquo The EMBO Jour-nal vol 24 no 13 pp 2254ndash2264 2005
16 Oxidative Medicine and Cellular Longevity
[52] S J Martin J G Bradley and T G Cotter ldquoHL-60 cellsinduced to differentiate towards neutrophils subsequently dievia apoptosisrdquo Clinical and Experimental Immunology vol 79no 3 pp 448ndash453 1990
[53] A Benito D Grillot G Nunez and J L Fernandez-Luna ldquoReg-ulation and function of Bcl-2 during differentiation-inducedcell death in HL-60 promyelocytic cellsrdquoThe American Journalof Pathology vol 146 no 2 pp 481ndash490 1995
[54] M Lamkanfi N Festjens W Declercq T V Berghe andP Vandenabeele ldquoCaspases in cell survival proliferation anddifferentiationrdquo Cell Death and Differentiation vol 14 no 1 pp44ndash55 2007
[55] O Sordet C Rebe S Plenchette et al ldquoSpecific involvement ofcaspases in the differentiation of monocytes into macrophagesrdquoBlood vol 100 no 13 pp 4446ndash4453 2002
[56] T-B Kang T Ben-Moshe E E Varfolomeev et al ldquoCaspase-8 serves both apoptotic and nonapoptotic rolesrdquo Journal ofImmunology vol 173 no 5 pp 2976ndash2984 2004
[57] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[58] J Cheng J Yang Y Xia M Karin and B Su ldquoSynergisticinteraction of MEK kinase 2 c-Jun N-terminal kinase (JNK)kinase 2 and JNK1 results in efficient and specific JNK1activationrdquo Molecular and Cellular Biology vol 20 no 7 pp2334ndash2342 2000
[59] J L Blank P Gerwins EM Elliott S Sather andG L JohnsonldquoMolecular cloning of mitogen-activated proteinERK kinasekinases (MEKK) 2 and 3 regulation of sequential phosphoryla-tion pathways involving mitogen-activated protein kinase andc-Jun kinaserdquo Journal of Biological Chemistry vol 271 no 10pp 5361ndash5368 1996
[60] S Papa F Zazzeroni C G Pham C Bubici and G FranzosoldquoLinking JNK signaling to NF-120581B a key to survivalrdquo Journal ofCell Science vol 117 no 22 pp 5197ndash5208 2004
[61] T Kadoya A Khurana M Tcherpakov et al ldquoJAMP a JUB N-terminal kinase 1 (JNK1)-associated membrane protein regu-lates duration of JNK activityrdquo Molecular and Cellular Biologyvol 25 no 19 pp 8619ndash8630 2005
[62] M Qi and E A Elion ldquoMAP kinase pathwaysrdquo Journal of CellScience vol 118 no 16 pp 3569ndash3572 2005
[63] Q Zhao and F S Lee ldquoMitogen-activated protein kinaseERKkinase kinases 2 and 3 activate nuclear factor-120581B through I120581Bkinase-120572 and I120581B kinase-120573rdquoThe Journal of Biological Chemistryvol 274 no 13 pp 8355ndash8358 1999
[64] H Namba V Saenko and S Yamashita ldquoNuclear factor-120581Bin thyroid carcinogenesis and progression a novel therapeutictarget for advanced thyroid cancerrdquo Arquivos Brasileiros deEndocrinologia e Metabologia vol 51 no 5 pp 843ndash851 2007
[65] B Feng S Cheng W S Pear and H-C Liou ldquoNF-kBinhibitor blocks B cell development at two checkpointsrdquoMedi-cal Immunology vol 3 article 1 2004
[66] S Gerondakis and U Siebenlist ldquoRoles of the NF-kappaBpathway in lymphocyte development and functionrdquoCold SpringHarbor Perspectives in Biology vol 2 no 5 Article ID a0001822010
[67] K P Bhat V Balasubramaniyan B Vaillant et al ldquoMesenchy-mal differentiation mediated by NF-kB promotes radiationresistance in glioblastomardquo Cancer Cell vol 24 no 3 pp 331ndash346 2013
Figure 2 (a) Effect ofChemlaliOlive Leaf Extract (COLE) on the proliferation ofK562 cellsThe cells were treatedwith various concentrations(50 75 100 125 and 150120583gmL) of COLE for 24 48 and 72 h Cell proliferation was measured byMTT assay Control represents cells treatedwith 03 ethanol in medium Results are represented as the mean plusmn SD of three independent experiments lowast lowastlowast significantly differentfrom the control (119875 lt 005 and 119875 lt 001 resp) (b) Morphological observations of K562 cells after treatment with various concentrationsof Chemlali Olive Leaf Extract (COLE) for 72 h Cells were observed under a phase contrast microscope at 100x magnification Scale barsrepresent 100 120583m
0
2
4
6
8
10
12
14
16
18
20
Day 0 Day 1 Day 2 Day 3 Day 4 Day 6
Num
ber o
f via
ble c
ells
(times106)
Control 50120583gmL100 120583gmL 150120583gmL
(a)
0102030405060708090100
Day 0 Day 1 Day 2 Day 3 Day 4 Day 6
Viab
ility
()
Control50120583gmL
100 120583gmL150120583gmL
lowastlowastlowastlowast
lowastlowast
lowastlowast
lowastlowast
lowast lowast
(b)
Figure 3 Effect of Chemlali Olive Leaf Extract (COLE) on the cell number and viability of K562 cells (a) Number of viable cells aftertreatment with COLE after incubation up to 6 days (b) Percentage of viability in K562 cells treated with COLE K562 cells were treated at afinal concentration of 50 100 and 150120583gmL of COLE and incubated for different periods Cell number and cell viability were measured byflow cytometry Control represents cells treated with 03 ethanol in medium Results are represented as the mean plusmn SD of three independentexperiments lowast lowastlowast significantly different from the control (119875 lt 005 and 119875 lt 001 resp)
35 COLE Induces Apoptosis in K562 Cells The decreasein cell viability (Figure 3(b)) caused by COLE treatmentprompted us to investigate if the extract induces apoptosis inK562 cells Apoptosis is an important and active regulatorypathway of cell growth and proliferation resulting in somecharacteristic physiological changes Among these external-ization of phosphatidylserine (PS) is easily detected by flowcytometry after binding to the labeled Annexin VThe resultsindicated an increase in Annexin V positive cells from the1st day of incubation with 150 120583gmL of COLE in comparisonwith control cells (Figure 4) The percentage of Annexin V
cells remained stable during the first 3 days of treatmentwith 20 of total treated cells and interestingly increasedin the 4th and 6th days with 45 and 60 respectivelyTreatment with 100 120583gmL caused the apparition of apoptoticcells from the 4th day of incubation with a proportionof 162 This population increased in the 6th day up to267No significant difference in apoptotic cells amountwasdetected when cells were treated with 50 120583gmL until 6 daysof incubation
6 Oxidative Medicine and Cellular Longevity
Table 2 The distribution of cell cycle in K562 cells treated with Chemlali Olive Leaf Extract (COLE)
Results are represented as the means plusmn SD of three independent experimentslowast and lowastlowast mean that difference between control and treated cells in each phase (G0G1 S and G2M) is statistically significant at 119875 lt 005 and 119875 lt 001respectively
36 Impact of COLE on the Differentiation Capacity of K562Cells The reduction in cell growth as well as the morpho-logical changes observed in COLE treated cells led us tothink about the differentiation assessment K562 cells arepluripotent malignant cells that spontaneously differentiatealong erythroid macrophage and megakaryocyte lineages[22] Treated cells were harvested and analyzed for theexpression of lineage differentiation markers The analysiswas performed by flow cytometry at different days on themonocytemacrophage marker CD14 on the granulocytemarker CD11b on the erythrocyte marker GPA and on themegakaryocyte marker CD41 The results showed a signifi-cant increase in the expression of CD14 marker from day 1until day 6 in the cells treated with 150120583gmL suggestingthe commitment of K562 cells to the monocytemacrophagelineage (Figure 5(a)) A slight increase of the expression ofCD11b was also detected at this concentration (Figure 5(b))Treatment with 100 120583gmL showed a gain in the expression ofCD14 on the 1st and 2nd day and an augmentation of CD11bexpression from the 3rd day of treatment Treatment with50120583gmL did not show any significant effect on the differenti-ationmarkersCD14 andCD11b Even after 6 days of treatmentwith 150 120583gmL of COLE we could not detect the erythroidmarker at all the tested concentrations (Figure 5(d)) How-ever an instant increase in the megakaryocytic marker wasdetected on the 1st day of treatment followed by a drasticdecrease lower than the control during the following days(Figure 5(c))
37 Gene Expression Profile of COLE Treated K562 Cells Tofurther elucidate the mechanism by which COLE inducesapoptosis and differentiation of K562 cells we investigatedthe changes in gene expression profiles in treated cells usingHG219 GeneChip array Microarray analysis was performedon K562 cells at the 3rd day of treatment based on the obser-vation that such timing generated a significant expression ofthe differentiation markers Genes with more than 15-foldchange in expression levels between control and 150 120583gmLofCOLE treated cells were classified into categories accordingto the cell biological processes According to the GO analysis
0
10
20
30
40
50
60
70
Day 1 Day 2 Day 3 Day 4 Day 6
Ann
exin
pos
itive
(
)
Control50120583gmL
100 120583gmL150120583gmL
lowastlowast lowastlowast lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
Figure 4 Induction of apoptosis in K562 cells treatedwith ChemlaliOlive Leaf Extract (COLE) Cells were treated with 50 100 and150 120583gmL of COLE and incubated for different time At the indi-cated time K562 cells were stained with Annexin V and analyzed byflow cytometry Control represents cells treated with 03 ethanolin medium Results are represented as the mean plusmn SD of threeindependent experiments lowast lowastlowast significantly different from thecontrol (119875 lt 005 and 119875 lt 001 resp)
results (Tables S1 and S2 in Supplementary Material avail-able online at httpdxdoiorg1011552014927619) COLEtreatment induced the upregulation of genes involved inhematopoiesis such as CTNNB1 SH2B3 CIAPIN1 RPA1 andgenes implicated in cell differentiation and its regulation suchasMCL1CTNNB1 andCCNE1 Interestingly genes related tomyeloid cell differentiation such as IFI16ACIN1 andCASP8and particularly genes engaged in monocyte differentiationwere represented in the upregulated categories Moreoveran increased expression of genes involved in chemotaxissuch as NUP85 HRAS IL8 CXCL2 and CXCL3 and genesrelated to cytokine production was also detected Amongupregulated genes AP1G1 RABEP1 RAB5C RAB21 EPS15L1and CORO1C were also found involved in the process ofendocytosis and phagocytosis as well as genes related toprotein transport and genes implicated in cell adhesion suchas ICAM3 HSPB11 and PNN
Conversely the erythrocyte differentiation categorieswere represented in the list of the decreased genes (Table S2)
Figure 5 Expression of differentiationmarkers onK562 cells treatedwith Chemlali Olive Leaf Extract (COLE) up to 6 days Cells were treatedwith 50 100 and 150 120583gmL of COLE and incubated for different periods Control cells were treated with ethanol at a final concentration of03 Expression of cell surface markers was detected by flow cytometry (a) Expression of CD14 (monocytemacrophage specific marker)(b) Expression of CD11b (granulocytemonocyte specific marker) (c) Expression of CD41 (megakaryocyte specific marker) (d) Expressionof glycophorin A (GPA) (erythrocyte specific marker) Results are represented as the mean plusmn SD of three independent experiments lowast lowastlowastsignificantly different from the control at 119875 lt 005 and 119875 lt 001 respectively
including GYPA HBE1 FECH and ALAS2 which is consis-tent with the decreased expression of erythrocyte differenti-ation marker in the flow cytometry results (Figure 5(d))
Analysis of the microarray data showed the upregu-lation of several proapoptotic genes and genes involvedin the regulation of mitochondrial membrane permeabilitysuch as CASP6 CASP8 DFFA and BID On the otherhandexpression of genes negatively regulating the apoptosis
and those inhibiting the caspase activity was significantlydecreased such as IGF1R HSPA5 and BCL2
Among the upregulated genes we found those related tothe NF-kappaB cascade as well as to the MAPKKK cascadeand the Wnt receptor signaling pathway
Categories related to cell cycle regulation and cell pro-liferation were represented by both upregulated and down-regulated genes Finally some transcription factors werealso downregulated such as FOXP1 whereas others wereupregulated like EGR1 and NFYA
The list of selected genes is presented in Table 3 Fromthis list it is clear that the differentially expressed genes aftertreatment with 100 and 150120583gmL have mostly the sametendency which could validate the obtained results
12 Oxidative Medicine and Cellular Longevity
4 Discussion
Previous reports had indicated that olive leaf extract inducesapoptosis in the human leukemic cell lines HL-60 and Jurkatcells [13 21] In this study we give evidence that COLEexhibits its antileukemia effect by both inducing apoptosisand promoting differentiation of the multipotent humanleukemia K562 cells
Induction of differentiation as well as apoptosis has beenfrequently reported to be associatedwith a loss of proliferativecapacity of the cells It has been reported that olive leaf extractinhibits the proliferation of some cancer cell lines such asB16 melanoma cells HL-60 leukemia cells the breast cancerMCF-7 cells and the glioblastoma cells [12 25ndash27] Ourresults indicate that COLE inhibits the proliferation of K562cells in a dose dependent manner (Figure 2(a)) Howeverdespite the drastic decrease in cell proliferation the viabilityof cells treated with the highest concentration of COLE(150 120583gmL) remains relatively high (80) for up to 3 daysof treatment (Figure 3(b))
Since it is well established that the arrest of the cellcycle progression allows the cells to follow other processessuch as apoptosis and differentiation we then explored theeffect of COLE on the cell cycle distribution (Table 2) Oliveleaf extract has been shown to cause cell cycle arrest atG0G1 phase in both the breast cancer MCF-7 cells and themelanoma B16 cells [12 25] However the treatment timesapplied in both of these studies were limited to 48 h and 18 hrespectively In this study COLE treated cells were arrested atG0G1 on the 1st and 2ndday of treatment which is consistentwith the reported studies Prolonged incubation with COLEshowed an arrest of cell cycle at G2M phase (3rd and 4thday of incubation) The microarray analysis performed forthe 3rd day of treatment revealed a significant increase ofCHEK2 gene expression in COLE treated cells The encodedprotein regulates the cell cycle checkpoint arrest throughthe inhibition of the activity of CDC25A CDC25B andCDC25C [28] CDC25 proteins trigger the entry into mitosisat different points of the cell cycle by activating theCdk-cyclincomplexes CDC25A acts early in the cell cycle regulatingthe G1S transition whereas CDC25B and CDC25C act atG2M [29] Interestingly our results showed a decrease in theexpression of CDC25C gene in COLE treated cells associatedwith an increase in the expression of CDC25A which mayexplain the cell cycle arrest at G2M phase observed at the3rd day of treatment with COLE
Treatment with COLE increased the expression of CD14on the cell surface of treated cells indicating the differenti-ation of K562 cells toward the mono-macrophage lineage(Figure 5(a)) The increase in the percentage of cells positivefor CD11b (Figure 5(b)) confirms this hypothesis In factCD11b is expressed on both monocytes and granulocytesThus together with the increase of CD14 expression theexpression of CD11b suggests that treatment with COLEpromotes the commitment of K562 cells into the mono-cytemacrophage lineage at the expense of the granulocyticdifferentiationThe instant increase of the expression of CD41
on the 1st day of treatment and its absence during the fol-lowing days (Figure 5(c)) could be consistent with monocytedifferentiation instead of megakaryocyte differentiation sinceit has been reported that 120572IIb promoter which initiates thetranscription of CD41 is transcriptionally active in pluripo-tent myeloid progenitors in early stages of erythropoiesisand all along the megakaryocytic differentiation as well asto a lesser extent in the early stages of myelomonocyticdifferentiation and the late stages of erythropoiesis [30] Thisinstant increase could be also explained by the phenomenonof lineage conversion where some hematopoietic progenitorscan be converted into lineages other than their own bythe ectopic expression of some transcription factors [31]In this regard it was previously reported that commonlymphoid progenitors megakaryocyte erythroid progenitorsearly B cells and early T cells could be converted to thegranulocytemonocyte lineage by the induction of CEBP120572activity [32] Microarray results support the absence ofmegakaryocyte differentiation in the 3rd day of treatment Infact data showed a decrease in the expression of genes relatedtomegakaryopoiesis such as the essential regulator of plateletrelease NFE2 and its main candidate target gene TUBB1(encoding the megakaryocyte and platelet specific isoform of120573-tubulin (1205731)) [33] as well as the BACH2 transcription factorreported to be induced during megakaryocyte differentiation[34]
Interestingly results of microarray analysis (Table 3)showed the upregulation of several genes already reportedas key players in monocytemacrophage differentiation inthe COLE treated cells Among these genes the expressionof IFI16 was markedly increased Constitutively expressedin lymphoid cells IFI16 was shown to be associated withthe differentiation of human myeloid cells in response tointerferon gamma [35] Moreover IFI16 has been selectivelyexpressed during the differentiation of myeloid progenitorcells along monocytic lineage while it is downregulatedduring the commitment toward the granulocytic or erythroidlineage suggesting its potential involvement in the differenti-ation andmaturation of the monocytic lineage [36 37] Geneencoding for EGR-1 a positive modulator of macrophagedifferentiation was also significantly upregulated EGR-1 isknown to dictate development of myeloid progenitors alongthe macrophage lineage at the expense of development alongother lineages [38 39] Dauffy et al [40] reported thatIFI16-enforced expression in myeloid progenitors inducedthe expression of EGR-1 and conducted the macrophagedifferentiation in the absence of the macrophage colonystimulating factor (M-CSF) This suggests that EGR-1 couldbe regulated by the expression of IFI16 Treatment with COLEincreased as well the expression of NFYA encoding the Asubunit of the nuclear transcription factor Y whose synthesiswas previously reported during the terminal differentiationof monocytes to macrophages [38 41] The downregula-tion of FOXP1 in COLE treated K562 cells is consistentwith monocytemacrophage differentiation since it has beendemonstrated that the expression of FOXP1 was markedlydecreased in monocyte-induced differentiation of HL-60cells as well as in human peripheral blood monocytes and
Oxidative Medicine and Cellular Longevity 13
that the overexpression of FOXP1 prevented the morphologicmacrophage-like differentiation [42 43]
Monocytes and macrophages have several functionalcharacteristics including cell adhesion migration chemo-taxis and phagocytic activity Interestingly an enhancedexpression of the chemokines genes CXCL8 (IL8) CXCL2andCXCL3was found in COLE treated cells IL-8 productionhas been observed in vitro in a wide variety of cells includingmonocytes T lymphocytes neutrophils vascular endothe-lial cells dermal fibroblasts keratinocytes hepatocytes andhuman gastric cancer cells while the production of CXCL2andCXCL3was only described inmonocytes fibroblasts andendothelial cells [44 45] Another gene related to chemotaxiswas upregulated NUP85 which is an essential component ofthe nuclear pore complex and was reported to be involved inCCR2-mediated chemotaxis of monocytes [46]
COLE treatment also enhanced the expression of somegenes related to phagocytosis AP1G1 andRab proteins relatedgenes AP1G1 is a subunit of clathrin-associated adaptorprotein complex 1 whose role has been recently highlightedfor efficient phagocytosis at an early stage of phagosomeformation since it participates in the extension of the phago-cytic cup [47 48] The small GTPases Rab are key regulatorsof intracellular membrane trafficking Recent studies havedemonstrated that several RAB proteins play an importantrole in phagocytosis RAB proteins such as RAB11 Rab5 andRab 21 participate in the phagosome formation and matura-tion and are necessary for the phagocytic activity [48ndash51]
Consistently with the increase in Annexin V positivecells (Figure 4) treatment with COLE upregulated someproapoptotic genes such as CASP6 CASP8 DFFA and BIDand downregulated the apoptosis suppressor BCL2 as wellas the caspase inhibitors genes (Table 3) Olive leaf waspreviously shown to induce morphological changes thatare characteristic of apoptosis in leukemia cells but themechanisms underlying this effect were not well investi-gated [13 21] The increase in the proportion of apoptoticcells observed from the 4th day of treatment with COLEcould be explained by the programmed death of the fullydifferentiated cells (Figure 4) In fact this increase wasaccompanied by an important decrease in cell viability aswell as a diminution in CD14 expression (Figures 3(b) and5(a)) Elsewhere it is well established that in hematopoietictissues apoptosis is coupled to terminal differentiation ofmyeloid progenitors even though themechanism responsiblefor the activation of apoptosis during myeloid maturationis still poorly understood [52 53] It is also noteworthy tomention that several reports have highlighted the role ofcaspases in the terminal differentiation of a variety of celltypes [54] One example is the differentiation of human bloodmonocytes into macrophages a process that is blocked bysynthetic caspase inhibitors [55] It has been also reportedthat caspase 8 deletion in bone-marrow cells resulted in arrestof hemopoietic progenitor functioning and that its deletionin cells of the myelomonocytic lineage led to the arrest ofdifferentiation into macrophages and consequently to celldeath [56]
Among the differentially expressed genes in COLEtreated cells (Table 3) genes encoding the mitogen-activated
protein kinase kinase kinases (MAPKKKs) MAP3K2MAP3K5 and MAP3K7 were highly expressed whileMAP2K5 and MAPK14p38-120572 were downregulated TheMAPKKKs act at the upstream of the MAP kinase cascadewhich regulates important cellular processes such as geneexpression cell proliferation differentiation cell survivaland death The MAP3K5 activates the c-Jun N-terminalkinase (JNK) and p38 mitogen-activated protein kinases[57] while the MAP3K2 is reported to preferentially activateJNK [58 59] However the significant decrease in theMAPK14p38-120572 may suggest that the effect of COLE onK562 cells is likely to be mediated by the JNK MAPKsrather than the p38 MAPKs JNK plays an important rolein apoptosis pathways weather by activating proapoptoticor antiapoptotic genes Particularly it was shown thatprolonged but not transient JNK activation promotes thecell death [60] Our results showed that JKAMP gene whichencodes for a membrane-anchored regulator of the durationof JNK1 activity was upregulated (154 plusmn 0051) The elevatedexpression of JKAMP as reported by Kadoya et al [61]results in sustained JNK activity These findings may indicatethe potential role of JNK pathway in the COLE-inducedapoptosis in K562 cells
Thanks to their large regulatory domains some MAP-KKKs can interact with upstream regulators have functionssuch as ubiquitylation and be activated by relief of autoin-hibition and oligomerization [62] In this context MAP3K2has been reported to play an important role in NF-kappaBsignaling pathway by activating the I-kappaB kinases whichin turn phosphorylate the inhibitory factors of the nuclearfactor kappaB (NF-kappaB) resulting in their rapid ubiq-uitination and the liberation of NF-kappaB complex whichtranslocate from the cytoplasm to the nucleus [63] MAP3K7in association with other proteins could also be required forthe activation of NF-kappaB According to the GO analysis(Table S1) COLE treatment upregulated genes implicatedin NF-kappaB-related categories such as positive regulationof NF-kappaB transcription factor activity I-kappaB kinase-NF-kappaB cascade and its positive regulation and I-kappaBphosphorylation Genes encoding the signal transducers inthe NF-kappaB pathway TRAF5 and TRAF6 as well asthe activator of NF-kappaB signaling SNIP1 were upregu-lated Interestingly an increased expression of NFKB1 geneencoding for the precursor of the mature NF-kappaB p50was detected in COLE treated cells compared to untreatedK562 cells NFKB is well recognized as a central activator ofthe antiapoptotic cascades in response to external stimuli orintrinsic immune reactions and its prosurvival activity hasbeen implicated in a variety of biological processes [60 64]In the B cell lineage the activity of NF-120581B is required forthe completion of various developmental stages includingdifferentiation and the response of these cells to antigens [65]NF-kappaB also plays an important role in the survival anddevelopment of T cells [66] and has been recently reported tomediate the differentiation of mesenchymal stem cells [67]
Several studies have demonstrated a crosstalk betweenthe NF-kappaB and JNK pathways seen as an inhibitoryeffect of NF-kappaB on apoptosis through the suppressionof JNK activity and that this suppressive effect may occur
14 Oxidative Medicine and Cellular Longevity
through different mechanisms [60] This suggests that thebalance between JNK and NF-kappaB activities is crucial todetermine the cell fate survival or deathWehypothesize herethat COLE-induced apoptosis or differentiation is likely to bedetermined by the crosstalk between these two pathways inK562 cells Further investigations regarding protein expres-sion and activity are needed to confirm these findings
5 Conclusions
Our study demonstrates for the first time that olive leafextract exhibits an antileukemia effect on the human chronicmyeloid leukemia cells Olive leaf extract was shown toinhibit the proliferation of K562 cells by inducing cell cyclearrest apoptosis and differentiation toward the monocytelineage The induced expression of molecules involved indifferentiation toward the monocytemacrophage lineage aswell asmolecules related to apoptosis and cell cycle regulationconfirmed these findings and provides insights into themechanism by which olive leaf exhibits its antileukemiaeffect
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported by the JICA-JST Science andTechnology Research Partnership for Sustainable Develop-ment (SATREPS) Project ldquoValorization of Bio-Resources inSemi-Arid and Arid Land for Regional Developmentrdquo
References
[1] M Leszczyniecka T Roberts P Dent S Grant and P B FisherldquoDifferentiation therapy of human cancer basic science andclinical applicationsrdquo Pharmacology and Therapeutics vol 90no 2-3 pp 105ndash156 2001
[2] A I Spira and M A Carducci ldquoDifferentiation therapyrdquoCurrent Opinion in Pharmacology vol 3 no 4 pp 338ndash3432003
[3] S Nobili D Lippi E Witort et al ldquoNatural compounds forcancer treatment and preventionrdquo Pharmacological Researchvol 59 no 6 pp 365ndash378 2009
[4] S Ramos ldquoEffects of dietary flavonoids on apoptotic pathwaysrelated to cancer chemopreventionrdquo Journal of NutritionalBiochemistry vol 18 no 7 pp 427ndash442 2007
[5] V Hajhashemi G Vaseghi M Pourfarzam and A AbdollahildquoAre antioxidants helpful for disease preventionrdquo Research inPharmaceutical Sciences vol 5 no 1 pp 1ndash8 2010
[6] M Bouaziz and S Sayadi ldquoIsolation and evaluation of antiox-idants from leaves of a Tunisian cultivar olive treerdquo EuropeanJournal of Lipid Science andTechnology vol 107 no 7-8 pp 497ndash504 2005
[7] A P Pereira I C F R Ferreira F Marcelino et al ldquoPhenoliccompounds and antimicrobial activity of olive (Olea europaeaL Cv Cobrancosa) leavesrdquo Molecules vol 12 no 5 pp 1153ndash1162 2007
[8] L I Somova F O Shode P Ramnanan and A Nadar ldquoAnti-hypertensive antiatherosclerotic and antioxidant activity oftriterpenoids isolated from Olea europaea subspecies africanaleavesrdquo Journal of Ethnopharmacology vol 84 no 2-3 pp 299ndash305 2003
[9] V Micol N Caturla L Perez-Fons V Mas L Perez andA Estepa ldquoThe olive leaf extract exhibits antiviral activityagainst viral haemorrhagic septicaemia rhabdovirus (VHSV)rdquoAntiviral Research vol 66 no 2-3 pp 129ndash136 2005
[10] J Wainstein T Ganz M Boaz et al ldquoOlive leaf extract as ahypoglycemic agent in both human diabetic subjects and inratsrdquo Journal of Medicinal Food vol 15 no 7 pp 605ndash610 2012
[11] L Seddik TM Bah A AouesM Slimani andM BenderdourldquoElucidation of mechanisms underlying the protective effects ofolive leaf extract against lead-induced neurotoxicity in Wistarratsrdquo Journal of Toxicological Sciences vol 36 no 6 pp 797ndash809 2011
[12] Z Bouallagui J Han H Isoda and S Sayadi ldquoHydroxytyrosolrich extract from olive leaves modulates cell cycle progressionin MCF-7 human breast cancer cellsrdquo Food and ChemicalToxicology vol 49 no 1 pp 179ndash184 2011
[13] R Fares S Bazzi S E Baydoun and R M Abdel-Massih ldquoTheantioxidant and anti-proliferative activity of the Lebanese Oleaeuropaea extractrdquo Plant Foods for Human Nutrition vol 66 no1 pp 58ndash63 2011
[14] V Goulas V Exarchou A N Troganis et al ldquoPhytochemicalsin olive-leaf extracts and their antiproliferative activity againstcancer and endothelial cellsrdquo Molecular Nutrition and FoodResearch vol 53 no 5 pp 600ndash608 2009
[15] M de Bock E B Thorstensen J G Derraik H V HendersonP L Hofman and W S Cutfield ldquoHuman absorption andmetabolism of oleuropein and hydroxytyrosol ingested as olive(Olea europaea L) leaf extractrdquo Molecular Nutrition and FoodResearch vol 57 no 11 pp 2079ndash2085 2013
[16] R Briante M Patumi S Terenziani E Bismuto F Febbraioand R Nucci ldquoOlea europaea L leaf extract and derivativesantioxidant propertiesrdquo Journal of Agricultural and Food Chem-istry vol 50 no 17 pp 4934ndash4940 2002
[17] O Benavente-Garcıa J Castillo J Lorente A Ortuno and J ADel Rio ldquoAntioxidant activity of phenolics extracted from Oleaeuropaea L leavesrdquo Food Chemistry vol 68 no 4 pp 457ndash4622000
[18] S H Omar ldquoOleuropein in olive and its pharmacologicaleffectsrdquo Scientia Pharmaceutica vol 78 no 2 pp 133ndash154 2010
[19] H Jemai M Bouaziz I Fki A El Feki and S SayadildquoHypolipidimic and antioxidant activities of oleuropein and itshydrolysis derivative-rich extracts from Chemlali olive leavesrdquoChemico-Biological Interactions vol 176 no 2-3 pp 88ndash982008
[20] R Garcıa-Villalba M Larrosa S Possemiers F A Tomas-Barberan and J C Espın ldquoBioavailability of phenolics from anoleuropein-rich olive (Olea europaea) leaf extract and its acuteeffect on plasma antioxidant status comparison between pre-and postmenopausal womenrdquo European Journal of Nutrition2013
[21] J Anter Z Fernandez-Bedmar M Villatoro-Pulido et al ldquoApilot study on the DNA-protective cytotoxic and apoptosis-inducing properties of olive-leaf extractsrdquo Mutation Researchvol 723 no 2 pp 165ndash170 2011
[22] J A Sutherland A R Turner and P Mannoni ldquoDifferen-tiation of K562 leukemia cells along erythroid macrophage
Oxidative Medicine and Cellular Longevity 15
and megakaryocyte lineagesrdquo Journal of Biological ResponseModifiers vol 5 no 3 pp 250ndash262 1986
[23] K Schwenke H-P Peterson K-H Wangenheim and L EFeinendegen ldquoInduction of differentiation in erythroleukemicK562 cells by 120574-irradiationrdquo Leukemia Research vol 19 no 12pp 955ndash961 1995
[24] V N Enujiugha J Y Talabi S A Malomo and A I OlagunjuldquoDPPH radical scavenging capacity of phenolic extracts fromAfrican Yam Bean (Sphenostylis stenocarpa)rdquo Food and Nutri-tion Sciences vol 3 pp 7ndash13 2012
[25] S A Mijatovic G S Timotijevic D M Miljkovic et alldquoMultiple antimelanoma potential of dry olive leaf extractrdquoInternational Journal of Cancer vol 128 no 8 pp 1955ndash19652011
[26] L Abaza T P N Talorete P Yamada Y KuritaM Zarrouk andH Isoda ldquoInduction of growth inhibition and differentiation ofhuman leukemia HL-60 cells by a Tunisian Gerboui olive leafextractrdquo Bioscience Biotechnology and Biochemistry vol 71 no5 pp 1306ndash1312 2007
[27] B Tunca G Tezcan G Cecener et al ldquoOlea europea leafextract alters microRNA expression in human glioblastomacellsrdquo Journal of Cancer Research and Clinical Oncology vol 138no 11 pp 1831ndash1844 2012
[28] S Matsuoka M Huang and S J Elledge ldquoLinkage of ATM tocell cycle regulation by the Chk2 protein kinaserdquo Science vol282 no 5395 pp 1893ndash1897 1998
[29] C Frazer and P G Young ldquoPhosphorylation mediated regu-lation of Cdc25 activity localization and stabilityrdquo in ProteinPhosphorylation in Human Health C Huang Ed pp 395ndash436InTech 2012
[30] N Debili C Robin V Schiavon et al ldquoDifferent expression ofCD41 on human lymphoid andmyeloid progenitors from adultsand neonatesrdquo Blood vol 97 no 7 pp 2023ndash2030 2001
[31] H Nakajima ldquoRole of transcription factors in differentiationand reprogramming of hematopoietic cellsrdquo Keio Journal ofMedicine vol 60 no 2 pp 47ndash55 2011
[32] Y Fukuchi F Shibata M Ito et al ldquoComprehensive analysis ofmyeloid lineage conversion using mice expressing an inducibleform of CEBP120572rdquoThe EMBO Journal vol 25 no 14 pp 3398ndash3410 2006
[33] R A Shivdasani ldquoMolecular and transcriptional regulation ofmegakaryocyte differentiationrdquo Stem Cells vol 19 no 5 pp397ndash407 2001
[34] K Terui Y Takahashi J Kitazawa T TokiM Yokoyama and EIto ldquoExpression of transcription factors during megakaryocyticdifferentiation of CD34+ cells from human cord blood inducedby thrombopoietinrdquo Tohoku Journal of Experimental Medicinevol 192 no 4 pp 259ndash273 2000
[35] J A Trapani M Dawson V A Apostolidis and K A BrowneldquoGenomic organization of IFI16 an interferon-inducible genewhose expression is associated with human myeloid cell differ-entiation correlation of predicted protein domains with exonorganizationrdquo Immunogenetics vol 40 no 6 pp 415ndash424 1994
[36] M J Dawson N J Elwood R W Johnstone and J A TrapanildquoThe IFN-inducible nucleoprotein IFI 16 is expressed in cellsof the monocyte lineage but is rapidly and markedly down-regulated in other myeloid precursor populationsrdquo Journal ofLeukocyte Biology vol 64 no 4 pp 546ndash554 1998
[37] M Gariglio M De Andrea M Lembo et al ldquoThe murinehomolog of the HIN 200 family Ifi 204 is constitutivelyexpressed in myeloid cells and selectively induced in the
monocytemacrophage lineagerdquo Journal of Leukocyte Biologyvol 64 no 5 pp 608ndash614 1998
[38] A F Valledor F E Borras M Cullell-Young and A CeladaldquoTranscription factors that regulate monocytemacrophage dif-ferentiationrdquo Journal of Leukocyte Biology vol 63 no 4 pp405ndash417 1998
[39] K Krishnaraju B Hoffman and D A Liebermann ldquoEarlygrowth response gene 1 stimulates development of hematopoi-etic progenitor cells along the macrophage lineage at theexpense of the granulocyte and erythroid lineagesrdquo Blood vol97 no 5 pp 1298ndash1305 2001
[40] J Dauffy G Mouchiroud and R P Bourette ldquoThe interferon-inducible gene Ifi204 is transcriptionally activated in responseto M-CSF and its expression favors macrophage differentiationin myeloid progenitor cellsrdquo Journal of Leukocyte Biology vol79 no 1 pp 173ndash183 2006
[41] G Marziali E Perrotti R Ilari U Testa E M Coccia andA Battistini ldquoTranscriptional regulation of the ferritin heavy-chain gene the activity of the CCAAT binding factor NF-Y ismodulated in heme-treated Friend leukemia cells and duringmonocyte-to-macrophage differentiationrdquo Molecular and Cel-lular Biology vol 17 no 3 pp 1387ndash1395 1997
[42] C Shi X Zhang Z Chen et al ldquoIntegrin engagement regulatesmonocyte differentiation through the forkhead transcriptionfactor Foxp1rdquo Journal of Clinical Investigation vol 114 no 3 pp408ndash418 2004
[43] C Shi M Sakuma T Mooroka et al ldquoDown-regulation of theforkhead transcription factor Foxp1 is required for monocytedifferentiation andmacrophage functionrdquoBlood vol 112 no 12pp 4699ndash4711 2008
[44] D F Smith E Galkina K Ley and Y Huo ldquoGRO familychemokines are specialized for monocyte arrest from flowrdquoThe American Journal of Physiology Heart and CirculatoryPhysiology vol 289 no 5 pp H1976ndashH1984 2005
[45] J J Oppenheim O M Z Howard and E Goetzl ChemotacticFactors Neuropeptides and Other Ligands for Seven Transmem-brane Receptors 2000
[46] Y Terashima N Onai M Murai et al ldquoPivotal function forcytoplasmic protein FROUNT in CCR2-mediated monocytechemotaxisrdquo Nature Immunology vol 6 no 8 pp 827ndash8352005
[47] Y Lefkir M Malbouyres D Gotthardt et al ldquoInvolvement ofthe AP-1 adaptor complex in early steps of phagocytosis andmacropinocytosisrdquo Molecular Biology of the Cell vol 15 no 2pp 861ndash869 2004
[48] F Niedergang and P Chavrier ldquoSignaling and membranedynamics during phagocytosis many roads lead to the phago-somerdquo Current Opinion in Cell Biology vol 16 no 4 pp 422ndash428 2004
[49] D Cox D J Lee B M Dale J Calafat and S Green-berg ldquoA Rab11-containing rapidly recycling compartment inmacrophages that promotes phagocytosisrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 97 no 2 pp 680ndash685 2000
[50] O V Vieira C Bucci R E Harrison et al ldquoModulation of Rab5and Rab7 recruitment to phagosomes by phosphatidylinositol3-kinaserdquoMolecular andCellular Biology vol 23 no 7 pp 2501ndash2514 2003
[51] T Khurana J A Brzostowski and A R Kimmel ldquoARab21LIM-onlyCH-LIM complex regulates phagocytosis viaboth activating and inhibitory mechanismsrdquo The EMBO Jour-nal vol 24 no 13 pp 2254ndash2264 2005
16 Oxidative Medicine and Cellular Longevity
[52] S J Martin J G Bradley and T G Cotter ldquoHL-60 cellsinduced to differentiate towards neutrophils subsequently dievia apoptosisrdquo Clinical and Experimental Immunology vol 79no 3 pp 448ndash453 1990
[53] A Benito D Grillot G Nunez and J L Fernandez-Luna ldquoReg-ulation and function of Bcl-2 during differentiation-inducedcell death in HL-60 promyelocytic cellsrdquoThe American Journalof Pathology vol 146 no 2 pp 481ndash490 1995
[54] M Lamkanfi N Festjens W Declercq T V Berghe andP Vandenabeele ldquoCaspases in cell survival proliferation anddifferentiationrdquo Cell Death and Differentiation vol 14 no 1 pp44ndash55 2007
[55] O Sordet C Rebe S Plenchette et al ldquoSpecific involvement ofcaspases in the differentiation of monocytes into macrophagesrdquoBlood vol 100 no 13 pp 4446ndash4453 2002
[56] T-B Kang T Ben-Moshe E E Varfolomeev et al ldquoCaspase-8 serves both apoptotic and nonapoptotic rolesrdquo Journal ofImmunology vol 173 no 5 pp 2976ndash2984 2004
[57] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[58] J Cheng J Yang Y Xia M Karin and B Su ldquoSynergisticinteraction of MEK kinase 2 c-Jun N-terminal kinase (JNK)kinase 2 and JNK1 results in efficient and specific JNK1activationrdquo Molecular and Cellular Biology vol 20 no 7 pp2334ndash2342 2000
[59] J L Blank P Gerwins EM Elliott S Sather andG L JohnsonldquoMolecular cloning of mitogen-activated proteinERK kinasekinases (MEKK) 2 and 3 regulation of sequential phosphoryla-tion pathways involving mitogen-activated protein kinase andc-Jun kinaserdquo Journal of Biological Chemistry vol 271 no 10pp 5361ndash5368 1996
[60] S Papa F Zazzeroni C G Pham C Bubici and G FranzosoldquoLinking JNK signaling to NF-120581B a key to survivalrdquo Journal ofCell Science vol 117 no 22 pp 5197ndash5208 2004
[61] T Kadoya A Khurana M Tcherpakov et al ldquoJAMP a JUB N-terminal kinase 1 (JNK1)-associated membrane protein regu-lates duration of JNK activityrdquo Molecular and Cellular Biologyvol 25 no 19 pp 8619ndash8630 2005
[62] M Qi and E A Elion ldquoMAP kinase pathwaysrdquo Journal of CellScience vol 118 no 16 pp 3569ndash3572 2005
[63] Q Zhao and F S Lee ldquoMitogen-activated protein kinaseERKkinase kinases 2 and 3 activate nuclear factor-120581B through I120581Bkinase-120572 and I120581B kinase-120573rdquoThe Journal of Biological Chemistryvol 274 no 13 pp 8355ndash8358 1999
[64] H Namba V Saenko and S Yamashita ldquoNuclear factor-120581Bin thyroid carcinogenesis and progression a novel therapeutictarget for advanced thyroid cancerrdquo Arquivos Brasileiros deEndocrinologia e Metabologia vol 51 no 5 pp 843ndash851 2007
[65] B Feng S Cheng W S Pear and H-C Liou ldquoNF-kBinhibitor blocks B cell development at two checkpointsrdquoMedi-cal Immunology vol 3 article 1 2004
[66] S Gerondakis and U Siebenlist ldquoRoles of the NF-kappaBpathway in lymphocyte development and functionrdquoCold SpringHarbor Perspectives in Biology vol 2 no 5 Article ID a0001822010
[67] K P Bhat V Balasubramaniyan B Vaillant et al ldquoMesenchy-mal differentiation mediated by NF-kB promotes radiationresistance in glioblastomardquo Cancer Cell vol 24 no 3 pp 331ndash346 2013
Results are represented as the means plusmn SD of three independent experimentslowast and lowastlowast mean that difference between control and treated cells in each phase (G0G1 S and G2M) is statistically significant at 119875 lt 005 and 119875 lt 001respectively
36 Impact of COLE on the Differentiation Capacity of K562Cells The reduction in cell growth as well as the morpho-logical changes observed in COLE treated cells led us tothink about the differentiation assessment K562 cells arepluripotent malignant cells that spontaneously differentiatealong erythroid macrophage and megakaryocyte lineages[22] Treated cells were harvested and analyzed for theexpression of lineage differentiation markers The analysiswas performed by flow cytometry at different days on themonocytemacrophage marker CD14 on the granulocytemarker CD11b on the erythrocyte marker GPA and on themegakaryocyte marker CD41 The results showed a signifi-cant increase in the expression of CD14 marker from day 1until day 6 in the cells treated with 150120583gmL suggestingthe commitment of K562 cells to the monocytemacrophagelineage (Figure 5(a)) A slight increase of the expression ofCD11b was also detected at this concentration (Figure 5(b))Treatment with 100 120583gmL showed a gain in the expression ofCD14 on the 1st and 2nd day and an augmentation of CD11bexpression from the 3rd day of treatment Treatment with50120583gmL did not show any significant effect on the differenti-ationmarkersCD14 andCD11b Even after 6 days of treatmentwith 150 120583gmL of COLE we could not detect the erythroidmarker at all the tested concentrations (Figure 5(d)) How-ever an instant increase in the megakaryocytic marker wasdetected on the 1st day of treatment followed by a drasticdecrease lower than the control during the following days(Figure 5(c))
37 Gene Expression Profile of COLE Treated K562 Cells Tofurther elucidate the mechanism by which COLE inducesapoptosis and differentiation of K562 cells we investigatedthe changes in gene expression profiles in treated cells usingHG219 GeneChip array Microarray analysis was performedon K562 cells at the 3rd day of treatment based on the obser-vation that such timing generated a significant expression ofthe differentiation markers Genes with more than 15-foldchange in expression levels between control and 150 120583gmLofCOLE treated cells were classified into categories accordingto the cell biological processes According to the GO analysis
0
10
20
30
40
50
60
70
Day 1 Day 2 Day 3 Day 4 Day 6
Ann
exin
pos
itive
(
)
Control50120583gmL
100 120583gmL150120583gmL
lowastlowast lowastlowast lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
Figure 4 Induction of apoptosis in K562 cells treatedwith ChemlaliOlive Leaf Extract (COLE) Cells were treated with 50 100 and150 120583gmL of COLE and incubated for different time At the indi-cated time K562 cells were stained with Annexin V and analyzed byflow cytometry Control represents cells treated with 03 ethanolin medium Results are represented as the mean plusmn SD of threeindependent experiments lowast lowastlowast significantly different from thecontrol (119875 lt 005 and 119875 lt 001 resp)
results (Tables S1 and S2 in Supplementary Material avail-able online at httpdxdoiorg1011552014927619) COLEtreatment induced the upregulation of genes involved inhematopoiesis such as CTNNB1 SH2B3 CIAPIN1 RPA1 andgenes implicated in cell differentiation and its regulation suchasMCL1CTNNB1 andCCNE1 Interestingly genes related tomyeloid cell differentiation such as IFI16ACIN1 andCASP8and particularly genes engaged in monocyte differentiationwere represented in the upregulated categories Moreoveran increased expression of genes involved in chemotaxissuch as NUP85 HRAS IL8 CXCL2 and CXCL3 and genesrelated to cytokine production was also detected Amongupregulated genes AP1G1 RABEP1 RAB5C RAB21 EPS15L1and CORO1C were also found involved in the process ofendocytosis and phagocytosis as well as genes related toprotein transport and genes implicated in cell adhesion suchas ICAM3 HSPB11 and PNN
Conversely the erythrocyte differentiation categorieswere represented in the list of the decreased genes (Table S2)
Figure 5 Expression of differentiationmarkers onK562 cells treatedwith Chemlali Olive Leaf Extract (COLE) up to 6 days Cells were treatedwith 50 100 and 150 120583gmL of COLE and incubated for different periods Control cells were treated with ethanol at a final concentration of03 Expression of cell surface markers was detected by flow cytometry (a) Expression of CD14 (monocytemacrophage specific marker)(b) Expression of CD11b (granulocytemonocyte specific marker) (c) Expression of CD41 (megakaryocyte specific marker) (d) Expressionof glycophorin A (GPA) (erythrocyte specific marker) Results are represented as the mean plusmn SD of three independent experiments lowast lowastlowastsignificantly different from the control at 119875 lt 005 and 119875 lt 001 respectively
including GYPA HBE1 FECH and ALAS2 which is consis-tent with the decreased expression of erythrocyte differenti-ation marker in the flow cytometry results (Figure 5(d))
Analysis of the microarray data showed the upregu-lation of several proapoptotic genes and genes involvedin the regulation of mitochondrial membrane permeabilitysuch as CASP6 CASP8 DFFA and BID On the otherhandexpression of genes negatively regulating the apoptosis
and those inhibiting the caspase activity was significantlydecreased such as IGF1R HSPA5 and BCL2
Among the upregulated genes we found those related tothe NF-kappaB cascade as well as to the MAPKKK cascadeand the Wnt receptor signaling pathway
Categories related to cell cycle regulation and cell pro-liferation were represented by both upregulated and down-regulated genes Finally some transcription factors werealso downregulated such as FOXP1 whereas others wereupregulated like EGR1 and NFYA
The list of selected genes is presented in Table 3 Fromthis list it is clear that the differentially expressed genes aftertreatment with 100 and 150120583gmL have mostly the sametendency which could validate the obtained results
12 Oxidative Medicine and Cellular Longevity
4 Discussion
Previous reports had indicated that olive leaf extract inducesapoptosis in the human leukemic cell lines HL-60 and Jurkatcells [13 21] In this study we give evidence that COLEexhibits its antileukemia effect by both inducing apoptosisand promoting differentiation of the multipotent humanleukemia K562 cells
Induction of differentiation as well as apoptosis has beenfrequently reported to be associatedwith a loss of proliferativecapacity of the cells It has been reported that olive leaf extractinhibits the proliferation of some cancer cell lines such asB16 melanoma cells HL-60 leukemia cells the breast cancerMCF-7 cells and the glioblastoma cells [12 25ndash27] Ourresults indicate that COLE inhibits the proliferation of K562cells in a dose dependent manner (Figure 2(a)) Howeverdespite the drastic decrease in cell proliferation the viabilityof cells treated with the highest concentration of COLE(150 120583gmL) remains relatively high (80) for up to 3 daysof treatment (Figure 3(b))
Since it is well established that the arrest of the cellcycle progression allows the cells to follow other processessuch as apoptosis and differentiation we then explored theeffect of COLE on the cell cycle distribution (Table 2) Oliveleaf extract has been shown to cause cell cycle arrest atG0G1 phase in both the breast cancer MCF-7 cells and themelanoma B16 cells [12 25] However the treatment timesapplied in both of these studies were limited to 48 h and 18 hrespectively In this study COLE treated cells were arrested atG0G1 on the 1st and 2ndday of treatment which is consistentwith the reported studies Prolonged incubation with COLEshowed an arrest of cell cycle at G2M phase (3rd and 4thday of incubation) The microarray analysis performed forthe 3rd day of treatment revealed a significant increase ofCHEK2 gene expression in COLE treated cells The encodedprotein regulates the cell cycle checkpoint arrest throughthe inhibition of the activity of CDC25A CDC25B andCDC25C [28] CDC25 proteins trigger the entry into mitosisat different points of the cell cycle by activating theCdk-cyclincomplexes CDC25A acts early in the cell cycle regulatingthe G1S transition whereas CDC25B and CDC25C act atG2M [29] Interestingly our results showed a decrease in theexpression of CDC25C gene in COLE treated cells associatedwith an increase in the expression of CDC25A which mayexplain the cell cycle arrest at G2M phase observed at the3rd day of treatment with COLE
Treatment with COLE increased the expression of CD14on the cell surface of treated cells indicating the differenti-ation of K562 cells toward the mono-macrophage lineage(Figure 5(a)) The increase in the percentage of cells positivefor CD11b (Figure 5(b)) confirms this hypothesis In factCD11b is expressed on both monocytes and granulocytesThus together with the increase of CD14 expression theexpression of CD11b suggests that treatment with COLEpromotes the commitment of K562 cells into the mono-cytemacrophage lineage at the expense of the granulocyticdifferentiationThe instant increase of the expression of CD41
on the 1st day of treatment and its absence during the fol-lowing days (Figure 5(c)) could be consistent with monocytedifferentiation instead of megakaryocyte differentiation sinceit has been reported that 120572IIb promoter which initiates thetranscription of CD41 is transcriptionally active in pluripo-tent myeloid progenitors in early stages of erythropoiesisand all along the megakaryocytic differentiation as well asto a lesser extent in the early stages of myelomonocyticdifferentiation and the late stages of erythropoiesis [30] Thisinstant increase could be also explained by the phenomenonof lineage conversion where some hematopoietic progenitorscan be converted into lineages other than their own bythe ectopic expression of some transcription factors [31]In this regard it was previously reported that commonlymphoid progenitors megakaryocyte erythroid progenitorsearly B cells and early T cells could be converted to thegranulocytemonocyte lineage by the induction of CEBP120572activity [32] Microarray results support the absence ofmegakaryocyte differentiation in the 3rd day of treatment Infact data showed a decrease in the expression of genes relatedtomegakaryopoiesis such as the essential regulator of plateletrelease NFE2 and its main candidate target gene TUBB1(encoding the megakaryocyte and platelet specific isoform of120573-tubulin (1205731)) [33] as well as the BACH2 transcription factorreported to be induced during megakaryocyte differentiation[34]
Interestingly results of microarray analysis (Table 3)showed the upregulation of several genes already reportedas key players in monocytemacrophage differentiation inthe COLE treated cells Among these genes the expressionof IFI16 was markedly increased Constitutively expressedin lymphoid cells IFI16 was shown to be associated withthe differentiation of human myeloid cells in response tointerferon gamma [35] Moreover IFI16 has been selectivelyexpressed during the differentiation of myeloid progenitorcells along monocytic lineage while it is downregulatedduring the commitment toward the granulocytic or erythroidlineage suggesting its potential involvement in the differenti-ation andmaturation of the monocytic lineage [36 37] Geneencoding for EGR-1 a positive modulator of macrophagedifferentiation was also significantly upregulated EGR-1 isknown to dictate development of myeloid progenitors alongthe macrophage lineage at the expense of development alongother lineages [38 39] Dauffy et al [40] reported thatIFI16-enforced expression in myeloid progenitors inducedthe expression of EGR-1 and conducted the macrophagedifferentiation in the absence of the macrophage colonystimulating factor (M-CSF) This suggests that EGR-1 couldbe regulated by the expression of IFI16 Treatment with COLEincreased as well the expression of NFYA encoding the Asubunit of the nuclear transcription factor Y whose synthesiswas previously reported during the terminal differentiationof monocytes to macrophages [38 41] The downregula-tion of FOXP1 in COLE treated K562 cells is consistentwith monocytemacrophage differentiation since it has beendemonstrated that the expression of FOXP1 was markedlydecreased in monocyte-induced differentiation of HL-60cells as well as in human peripheral blood monocytes and
Oxidative Medicine and Cellular Longevity 13
that the overexpression of FOXP1 prevented the morphologicmacrophage-like differentiation [42 43]
Monocytes and macrophages have several functionalcharacteristics including cell adhesion migration chemo-taxis and phagocytic activity Interestingly an enhancedexpression of the chemokines genes CXCL8 (IL8) CXCL2andCXCL3was found in COLE treated cells IL-8 productionhas been observed in vitro in a wide variety of cells includingmonocytes T lymphocytes neutrophils vascular endothe-lial cells dermal fibroblasts keratinocytes hepatocytes andhuman gastric cancer cells while the production of CXCL2andCXCL3was only described inmonocytes fibroblasts andendothelial cells [44 45] Another gene related to chemotaxiswas upregulated NUP85 which is an essential component ofthe nuclear pore complex and was reported to be involved inCCR2-mediated chemotaxis of monocytes [46]
COLE treatment also enhanced the expression of somegenes related to phagocytosis AP1G1 andRab proteins relatedgenes AP1G1 is a subunit of clathrin-associated adaptorprotein complex 1 whose role has been recently highlightedfor efficient phagocytosis at an early stage of phagosomeformation since it participates in the extension of the phago-cytic cup [47 48] The small GTPases Rab are key regulatorsof intracellular membrane trafficking Recent studies havedemonstrated that several RAB proteins play an importantrole in phagocytosis RAB proteins such as RAB11 Rab5 andRab 21 participate in the phagosome formation and matura-tion and are necessary for the phagocytic activity [48ndash51]
Consistently with the increase in Annexin V positivecells (Figure 4) treatment with COLE upregulated someproapoptotic genes such as CASP6 CASP8 DFFA and BIDand downregulated the apoptosis suppressor BCL2 as wellas the caspase inhibitors genes (Table 3) Olive leaf waspreviously shown to induce morphological changes thatare characteristic of apoptosis in leukemia cells but themechanisms underlying this effect were not well investi-gated [13 21] The increase in the proportion of apoptoticcells observed from the 4th day of treatment with COLEcould be explained by the programmed death of the fullydifferentiated cells (Figure 4) In fact this increase wasaccompanied by an important decrease in cell viability aswell as a diminution in CD14 expression (Figures 3(b) and5(a)) Elsewhere it is well established that in hematopoietictissues apoptosis is coupled to terminal differentiation ofmyeloid progenitors even though themechanism responsiblefor the activation of apoptosis during myeloid maturationis still poorly understood [52 53] It is also noteworthy tomention that several reports have highlighted the role ofcaspases in the terminal differentiation of a variety of celltypes [54] One example is the differentiation of human bloodmonocytes into macrophages a process that is blocked bysynthetic caspase inhibitors [55] It has been also reportedthat caspase 8 deletion in bone-marrow cells resulted in arrestof hemopoietic progenitor functioning and that its deletionin cells of the myelomonocytic lineage led to the arrest ofdifferentiation into macrophages and consequently to celldeath [56]
Among the differentially expressed genes in COLEtreated cells (Table 3) genes encoding the mitogen-activated
protein kinase kinase kinases (MAPKKKs) MAP3K2MAP3K5 and MAP3K7 were highly expressed whileMAP2K5 and MAPK14p38-120572 were downregulated TheMAPKKKs act at the upstream of the MAP kinase cascadewhich regulates important cellular processes such as geneexpression cell proliferation differentiation cell survivaland death The MAP3K5 activates the c-Jun N-terminalkinase (JNK) and p38 mitogen-activated protein kinases[57] while the MAP3K2 is reported to preferentially activateJNK [58 59] However the significant decrease in theMAPK14p38-120572 may suggest that the effect of COLE onK562 cells is likely to be mediated by the JNK MAPKsrather than the p38 MAPKs JNK plays an important rolein apoptosis pathways weather by activating proapoptoticor antiapoptotic genes Particularly it was shown thatprolonged but not transient JNK activation promotes thecell death [60] Our results showed that JKAMP gene whichencodes for a membrane-anchored regulator of the durationof JNK1 activity was upregulated (154 plusmn 0051) The elevatedexpression of JKAMP as reported by Kadoya et al [61]results in sustained JNK activity These findings may indicatethe potential role of JNK pathway in the COLE-inducedapoptosis in K562 cells
Thanks to their large regulatory domains some MAP-KKKs can interact with upstream regulators have functionssuch as ubiquitylation and be activated by relief of autoin-hibition and oligomerization [62] In this context MAP3K2has been reported to play an important role in NF-kappaBsignaling pathway by activating the I-kappaB kinases whichin turn phosphorylate the inhibitory factors of the nuclearfactor kappaB (NF-kappaB) resulting in their rapid ubiq-uitination and the liberation of NF-kappaB complex whichtranslocate from the cytoplasm to the nucleus [63] MAP3K7in association with other proteins could also be required forthe activation of NF-kappaB According to the GO analysis(Table S1) COLE treatment upregulated genes implicatedin NF-kappaB-related categories such as positive regulationof NF-kappaB transcription factor activity I-kappaB kinase-NF-kappaB cascade and its positive regulation and I-kappaBphosphorylation Genes encoding the signal transducers inthe NF-kappaB pathway TRAF5 and TRAF6 as well asthe activator of NF-kappaB signaling SNIP1 were upregu-lated Interestingly an increased expression of NFKB1 geneencoding for the precursor of the mature NF-kappaB p50was detected in COLE treated cells compared to untreatedK562 cells NFKB is well recognized as a central activator ofthe antiapoptotic cascades in response to external stimuli orintrinsic immune reactions and its prosurvival activity hasbeen implicated in a variety of biological processes [60 64]In the B cell lineage the activity of NF-120581B is required forthe completion of various developmental stages includingdifferentiation and the response of these cells to antigens [65]NF-kappaB also plays an important role in the survival anddevelopment of T cells [66] and has been recently reported tomediate the differentiation of mesenchymal stem cells [67]
Several studies have demonstrated a crosstalk betweenthe NF-kappaB and JNK pathways seen as an inhibitoryeffect of NF-kappaB on apoptosis through the suppressionof JNK activity and that this suppressive effect may occur
14 Oxidative Medicine and Cellular Longevity
through different mechanisms [60] This suggests that thebalance between JNK and NF-kappaB activities is crucial todetermine the cell fate survival or deathWehypothesize herethat COLE-induced apoptosis or differentiation is likely to bedetermined by the crosstalk between these two pathways inK562 cells Further investigations regarding protein expres-sion and activity are needed to confirm these findings
5 Conclusions
Our study demonstrates for the first time that olive leafextract exhibits an antileukemia effect on the human chronicmyeloid leukemia cells Olive leaf extract was shown toinhibit the proliferation of K562 cells by inducing cell cyclearrest apoptosis and differentiation toward the monocytelineage The induced expression of molecules involved indifferentiation toward the monocytemacrophage lineage aswell asmolecules related to apoptosis and cell cycle regulationconfirmed these findings and provides insights into themechanism by which olive leaf exhibits its antileukemiaeffect
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported by the JICA-JST Science andTechnology Research Partnership for Sustainable Develop-ment (SATREPS) Project ldquoValorization of Bio-Resources inSemi-Arid and Arid Land for Regional Developmentrdquo
References
[1] M Leszczyniecka T Roberts P Dent S Grant and P B FisherldquoDifferentiation therapy of human cancer basic science andclinical applicationsrdquo Pharmacology and Therapeutics vol 90no 2-3 pp 105ndash156 2001
[2] A I Spira and M A Carducci ldquoDifferentiation therapyrdquoCurrent Opinion in Pharmacology vol 3 no 4 pp 338ndash3432003
[3] S Nobili D Lippi E Witort et al ldquoNatural compounds forcancer treatment and preventionrdquo Pharmacological Researchvol 59 no 6 pp 365ndash378 2009
[4] S Ramos ldquoEffects of dietary flavonoids on apoptotic pathwaysrelated to cancer chemopreventionrdquo Journal of NutritionalBiochemistry vol 18 no 7 pp 427ndash442 2007
[5] V Hajhashemi G Vaseghi M Pourfarzam and A AbdollahildquoAre antioxidants helpful for disease preventionrdquo Research inPharmaceutical Sciences vol 5 no 1 pp 1ndash8 2010
[6] M Bouaziz and S Sayadi ldquoIsolation and evaluation of antiox-idants from leaves of a Tunisian cultivar olive treerdquo EuropeanJournal of Lipid Science andTechnology vol 107 no 7-8 pp 497ndash504 2005
[7] A P Pereira I C F R Ferreira F Marcelino et al ldquoPhenoliccompounds and antimicrobial activity of olive (Olea europaeaL Cv Cobrancosa) leavesrdquo Molecules vol 12 no 5 pp 1153ndash1162 2007
[8] L I Somova F O Shode P Ramnanan and A Nadar ldquoAnti-hypertensive antiatherosclerotic and antioxidant activity oftriterpenoids isolated from Olea europaea subspecies africanaleavesrdquo Journal of Ethnopharmacology vol 84 no 2-3 pp 299ndash305 2003
[9] V Micol N Caturla L Perez-Fons V Mas L Perez andA Estepa ldquoThe olive leaf extract exhibits antiviral activityagainst viral haemorrhagic septicaemia rhabdovirus (VHSV)rdquoAntiviral Research vol 66 no 2-3 pp 129ndash136 2005
[10] J Wainstein T Ganz M Boaz et al ldquoOlive leaf extract as ahypoglycemic agent in both human diabetic subjects and inratsrdquo Journal of Medicinal Food vol 15 no 7 pp 605ndash610 2012
[11] L Seddik TM Bah A AouesM Slimani andM BenderdourldquoElucidation of mechanisms underlying the protective effects ofolive leaf extract against lead-induced neurotoxicity in Wistarratsrdquo Journal of Toxicological Sciences vol 36 no 6 pp 797ndash809 2011
[12] Z Bouallagui J Han H Isoda and S Sayadi ldquoHydroxytyrosolrich extract from olive leaves modulates cell cycle progressionin MCF-7 human breast cancer cellsrdquo Food and ChemicalToxicology vol 49 no 1 pp 179ndash184 2011
[13] R Fares S Bazzi S E Baydoun and R M Abdel-Massih ldquoTheantioxidant and anti-proliferative activity of the Lebanese Oleaeuropaea extractrdquo Plant Foods for Human Nutrition vol 66 no1 pp 58ndash63 2011
[14] V Goulas V Exarchou A N Troganis et al ldquoPhytochemicalsin olive-leaf extracts and their antiproliferative activity againstcancer and endothelial cellsrdquo Molecular Nutrition and FoodResearch vol 53 no 5 pp 600ndash608 2009
[15] M de Bock E B Thorstensen J G Derraik H V HendersonP L Hofman and W S Cutfield ldquoHuman absorption andmetabolism of oleuropein and hydroxytyrosol ingested as olive(Olea europaea L) leaf extractrdquo Molecular Nutrition and FoodResearch vol 57 no 11 pp 2079ndash2085 2013
[16] R Briante M Patumi S Terenziani E Bismuto F Febbraioand R Nucci ldquoOlea europaea L leaf extract and derivativesantioxidant propertiesrdquo Journal of Agricultural and Food Chem-istry vol 50 no 17 pp 4934ndash4940 2002
[17] O Benavente-Garcıa J Castillo J Lorente A Ortuno and J ADel Rio ldquoAntioxidant activity of phenolics extracted from Oleaeuropaea L leavesrdquo Food Chemistry vol 68 no 4 pp 457ndash4622000
[18] S H Omar ldquoOleuropein in olive and its pharmacologicaleffectsrdquo Scientia Pharmaceutica vol 78 no 2 pp 133ndash154 2010
[19] H Jemai M Bouaziz I Fki A El Feki and S SayadildquoHypolipidimic and antioxidant activities of oleuropein and itshydrolysis derivative-rich extracts from Chemlali olive leavesrdquoChemico-Biological Interactions vol 176 no 2-3 pp 88ndash982008
[20] R Garcıa-Villalba M Larrosa S Possemiers F A Tomas-Barberan and J C Espın ldquoBioavailability of phenolics from anoleuropein-rich olive (Olea europaea) leaf extract and its acuteeffect on plasma antioxidant status comparison between pre-and postmenopausal womenrdquo European Journal of Nutrition2013
[21] J Anter Z Fernandez-Bedmar M Villatoro-Pulido et al ldquoApilot study on the DNA-protective cytotoxic and apoptosis-inducing properties of olive-leaf extractsrdquo Mutation Researchvol 723 no 2 pp 165ndash170 2011
[22] J A Sutherland A R Turner and P Mannoni ldquoDifferen-tiation of K562 leukemia cells along erythroid macrophage
Oxidative Medicine and Cellular Longevity 15
and megakaryocyte lineagesrdquo Journal of Biological ResponseModifiers vol 5 no 3 pp 250ndash262 1986
[23] K Schwenke H-P Peterson K-H Wangenheim and L EFeinendegen ldquoInduction of differentiation in erythroleukemicK562 cells by 120574-irradiationrdquo Leukemia Research vol 19 no 12pp 955ndash961 1995
[24] V N Enujiugha J Y Talabi S A Malomo and A I OlagunjuldquoDPPH radical scavenging capacity of phenolic extracts fromAfrican Yam Bean (Sphenostylis stenocarpa)rdquo Food and Nutri-tion Sciences vol 3 pp 7ndash13 2012
[25] S A Mijatovic G S Timotijevic D M Miljkovic et alldquoMultiple antimelanoma potential of dry olive leaf extractrdquoInternational Journal of Cancer vol 128 no 8 pp 1955ndash19652011
[26] L Abaza T P N Talorete P Yamada Y KuritaM Zarrouk andH Isoda ldquoInduction of growth inhibition and differentiation ofhuman leukemia HL-60 cells by a Tunisian Gerboui olive leafextractrdquo Bioscience Biotechnology and Biochemistry vol 71 no5 pp 1306ndash1312 2007
[27] B Tunca G Tezcan G Cecener et al ldquoOlea europea leafextract alters microRNA expression in human glioblastomacellsrdquo Journal of Cancer Research and Clinical Oncology vol 138no 11 pp 1831ndash1844 2012
[28] S Matsuoka M Huang and S J Elledge ldquoLinkage of ATM tocell cycle regulation by the Chk2 protein kinaserdquo Science vol282 no 5395 pp 1893ndash1897 1998
[29] C Frazer and P G Young ldquoPhosphorylation mediated regu-lation of Cdc25 activity localization and stabilityrdquo in ProteinPhosphorylation in Human Health C Huang Ed pp 395ndash436InTech 2012
[30] N Debili C Robin V Schiavon et al ldquoDifferent expression ofCD41 on human lymphoid andmyeloid progenitors from adultsand neonatesrdquo Blood vol 97 no 7 pp 2023ndash2030 2001
[31] H Nakajima ldquoRole of transcription factors in differentiationand reprogramming of hematopoietic cellsrdquo Keio Journal ofMedicine vol 60 no 2 pp 47ndash55 2011
[32] Y Fukuchi F Shibata M Ito et al ldquoComprehensive analysis ofmyeloid lineage conversion using mice expressing an inducibleform of CEBP120572rdquoThe EMBO Journal vol 25 no 14 pp 3398ndash3410 2006
[33] R A Shivdasani ldquoMolecular and transcriptional regulation ofmegakaryocyte differentiationrdquo Stem Cells vol 19 no 5 pp397ndash407 2001
[34] K Terui Y Takahashi J Kitazawa T TokiM Yokoyama and EIto ldquoExpression of transcription factors during megakaryocyticdifferentiation of CD34+ cells from human cord blood inducedby thrombopoietinrdquo Tohoku Journal of Experimental Medicinevol 192 no 4 pp 259ndash273 2000
[35] J A Trapani M Dawson V A Apostolidis and K A BrowneldquoGenomic organization of IFI16 an interferon-inducible genewhose expression is associated with human myeloid cell differ-entiation correlation of predicted protein domains with exonorganizationrdquo Immunogenetics vol 40 no 6 pp 415ndash424 1994
[36] M J Dawson N J Elwood R W Johnstone and J A TrapanildquoThe IFN-inducible nucleoprotein IFI 16 is expressed in cellsof the monocyte lineage but is rapidly and markedly down-regulated in other myeloid precursor populationsrdquo Journal ofLeukocyte Biology vol 64 no 4 pp 546ndash554 1998
[37] M Gariglio M De Andrea M Lembo et al ldquoThe murinehomolog of the HIN 200 family Ifi 204 is constitutivelyexpressed in myeloid cells and selectively induced in the
monocytemacrophage lineagerdquo Journal of Leukocyte Biologyvol 64 no 5 pp 608ndash614 1998
[38] A F Valledor F E Borras M Cullell-Young and A CeladaldquoTranscription factors that regulate monocytemacrophage dif-ferentiationrdquo Journal of Leukocyte Biology vol 63 no 4 pp405ndash417 1998
[39] K Krishnaraju B Hoffman and D A Liebermann ldquoEarlygrowth response gene 1 stimulates development of hematopoi-etic progenitor cells along the macrophage lineage at theexpense of the granulocyte and erythroid lineagesrdquo Blood vol97 no 5 pp 1298ndash1305 2001
[40] J Dauffy G Mouchiroud and R P Bourette ldquoThe interferon-inducible gene Ifi204 is transcriptionally activated in responseto M-CSF and its expression favors macrophage differentiationin myeloid progenitor cellsrdquo Journal of Leukocyte Biology vol79 no 1 pp 173ndash183 2006
[41] G Marziali E Perrotti R Ilari U Testa E M Coccia andA Battistini ldquoTranscriptional regulation of the ferritin heavy-chain gene the activity of the CCAAT binding factor NF-Y ismodulated in heme-treated Friend leukemia cells and duringmonocyte-to-macrophage differentiationrdquo Molecular and Cel-lular Biology vol 17 no 3 pp 1387ndash1395 1997
[42] C Shi X Zhang Z Chen et al ldquoIntegrin engagement regulatesmonocyte differentiation through the forkhead transcriptionfactor Foxp1rdquo Journal of Clinical Investigation vol 114 no 3 pp408ndash418 2004
[43] C Shi M Sakuma T Mooroka et al ldquoDown-regulation of theforkhead transcription factor Foxp1 is required for monocytedifferentiation andmacrophage functionrdquoBlood vol 112 no 12pp 4699ndash4711 2008
[44] D F Smith E Galkina K Ley and Y Huo ldquoGRO familychemokines are specialized for monocyte arrest from flowrdquoThe American Journal of Physiology Heart and CirculatoryPhysiology vol 289 no 5 pp H1976ndashH1984 2005
[45] J J Oppenheim O M Z Howard and E Goetzl ChemotacticFactors Neuropeptides and Other Ligands for Seven Transmem-brane Receptors 2000
[46] Y Terashima N Onai M Murai et al ldquoPivotal function forcytoplasmic protein FROUNT in CCR2-mediated monocytechemotaxisrdquo Nature Immunology vol 6 no 8 pp 827ndash8352005
[47] Y Lefkir M Malbouyres D Gotthardt et al ldquoInvolvement ofthe AP-1 adaptor complex in early steps of phagocytosis andmacropinocytosisrdquo Molecular Biology of the Cell vol 15 no 2pp 861ndash869 2004
[48] F Niedergang and P Chavrier ldquoSignaling and membranedynamics during phagocytosis many roads lead to the phago-somerdquo Current Opinion in Cell Biology vol 16 no 4 pp 422ndash428 2004
[49] D Cox D J Lee B M Dale J Calafat and S Green-berg ldquoA Rab11-containing rapidly recycling compartment inmacrophages that promotes phagocytosisrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 97 no 2 pp 680ndash685 2000
[50] O V Vieira C Bucci R E Harrison et al ldquoModulation of Rab5and Rab7 recruitment to phagosomes by phosphatidylinositol3-kinaserdquoMolecular andCellular Biology vol 23 no 7 pp 2501ndash2514 2003
[51] T Khurana J A Brzostowski and A R Kimmel ldquoARab21LIM-onlyCH-LIM complex regulates phagocytosis viaboth activating and inhibitory mechanismsrdquo The EMBO Jour-nal vol 24 no 13 pp 2254ndash2264 2005
16 Oxidative Medicine and Cellular Longevity
[52] S J Martin J G Bradley and T G Cotter ldquoHL-60 cellsinduced to differentiate towards neutrophils subsequently dievia apoptosisrdquo Clinical and Experimental Immunology vol 79no 3 pp 448ndash453 1990
[53] A Benito D Grillot G Nunez and J L Fernandez-Luna ldquoReg-ulation and function of Bcl-2 during differentiation-inducedcell death in HL-60 promyelocytic cellsrdquoThe American Journalof Pathology vol 146 no 2 pp 481ndash490 1995
[54] M Lamkanfi N Festjens W Declercq T V Berghe andP Vandenabeele ldquoCaspases in cell survival proliferation anddifferentiationrdquo Cell Death and Differentiation vol 14 no 1 pp44ndash55 2007
[55] O Sordet C Rebe S Plenchette et al ldquoSpecific involvement ofcaspases in the differentiation of monocytes into macrophagesrdquoBlood vol 100 no 13 pp 4446ndash4453 2002
[56] T-B Kang T Ben-Moshe E E Varfolomeev et al ldquoCaspase-8 serves both apoptotic and nonapoptotic rolesrdquo Journal ofImmunology vol 173 no 5 pp 2976ndash2984 2004
[57] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[58] J Cheng J Yang Y Xia M Karin and B Su ldquoSynergisticinteraction of MEK kinase 2 c-Jun N-terminal kinase (JNK)kinase 2 and JNK1 results in efficient and specific JNK1activationrdquo Molecular and Cellular Biology vol 20 no 7 pp2334ndash2342 2000
[59] J L Blank P Gerwins EM Elliott S Sather andG L JohnsonldquoMolecular cloning of mitogen-activated proteinERK kinasekinases (MEKK) 2 and 3 regulation of sequential phosphoryla-tion pathways involving mitogen-activated protein kinase andc-Jun kinaserdquo Journal of Biological Chemistry vol 271 no 10pp 5361ndash5368 1996
[60] S Papa F Zazzeroni C G Pham C Bubici and G FranzosoldquoLinking JNK signaling to NF-120581B a key to survivalrdquo Journal ofCell Science vol 117 no 22 pp 5197ndash5208 2004
[61] T Kadoya A Khurana M Tcherpakov et al ldquoJAMP a JUB N-terminal kinase 1 (JNK1)-associated membrane protein regu-lates duration of JNK activityrdquo Molecular and Cellular Biologyvol 25 no 19 pp 8619ndash8630 2005
[62] M Qi and E A Elion ldquoMAP kinase pathwaysrdquo Journal of CellScience vol 118 no 16 pp 3569ndash3572 2005
[63] Q Zhao and F S Lee ldquoMitogen-activated protein kinaseERKkinase kinases 2 and 3 activate nuclear factor-120581B through I120581Bkinase-120572 and I120581B kinase-120573rdquoThe Journal of Biological Chemistryvol 274 no 13 pp 8355ndash8358 1999
[64] H Namba V Saenko and S Yamashita ldquoNuclear factor-120581Bin thyroid carcinogenesis and progression a novel therapeutictarget for advanced thyroid cancerrdquo Arquivos Brasileiros deEndocrinologia e Metabologia vol 51 no 5 pp 843ndash851 2007
[65] B Feng S Cheng W S Pear and H-C Liou ldquoNF-kBinhibitor blocks B cell development at two checkpointsrdquoMedi-cal Immunology vol 3 article 1 2004
[66] S Gerondakis and U Siebenlist ldquoRoles of the NF-kappaBpathway in lymphocyte development and functionrdquoCold SpringHarbor Perspectives in Biology vol 2 no 5 Article ID a0001822010
[67] K P Bhat V Balasubramaniyan B Vaillant et al ldquoMesenchy-mal differentiation mediated by NF-kB promotes radiationresistance in glioblastomardquo Cancer Cell vol 24 no 3 pp 331ndash346 2013
Figure 5 Expression of differentiationmarkers onK562 cells treatedwith Chemlali Olive Leaf Extract (COLE) up to 6 days Cells were treatedwith 50 100 and 150 120583gmL of COLE and incubated for different periods Control cells were treated with ethanol at a final concentration of03 Expression of cell surface markers was detected by flow cytometry (a) Expression of CD14 (monocytemacrophage specific marker)(b) Expression of CD11b (granulocytemonocyte specific marker) (c) Expression of CD41 (megakaryocyte specific marker) (d) Expressionof glycophorin A (GPA) (erythrocyte specific marker) Results are represented as the mean plusmn SD of three independent experiments lowast lowastlowastsignificantly different from the control at 119875 lt 005 and 119875 lt 001 respectively
including GYPA HBE1 FECH and ALAS2 which is consis-tent with the decreased expression of erythrocyte differenti-ation marker in the flow cytometry results (Figure 5(d))
Analysis of the microarray data showed the upregu-lation of several proapoptotic genes and genes involvedin the regulation of mitochondrial membrane permeabilitysuch as CASP6 CASP8 DFFA and BID On the otherhandexpression of genes negatively regulating the apoptosis
and those inhibiting the caspase activity was significantlydecreased such as IGF1R HSPA5 and BCL2
Among the upregulated genes we found those related tothe NF-kappaB cascade as well as to the MAPKKK cascadeand the Wnt receptor signaling pathway
Categories related to cell cycle regulation and cell pro-liferation were represented by both upregulated and down-regulated genes Finally some transcription factors werealso downregulated such as FOXP1 whereas others wereupregulated like EGR1 and NFYA
The list of selected genes is presented in Table 3 Fromthis list it is clear that the differentially expressed genes aftertreatment with 100 and 150120583gmL have mostly the sametendency which could validate the obtained results
12 Oxidative Medicine and Cellular Longevity
4 Discussion
Previous reports had indicated that olive leaf extract inducesapoptosis in the human leukemic cell lines HL-60 and Jurkatcells [13 21] In this study we give evidence that COLEexhibits its antileukemia effect by both inducing apoptosisand promoting differentiation of the multipotent humanleukemia K562 cells
Induction of differentiation as well as apoptosis has beenfrequently reported to be associatedwith a loss of proliferativecapacity of the cells It has been reported that olive leaf extractinhibits the proliferation of some cancer cell lines such asB16 melanoma cells HL-60 leukemia cells the breast cancerMCF-7 cells and the glioblastoma cells [12 25ndash27] Ourresults indicate that COLE inhibits the proliferation of K562cells in a dose dependent manner (Figure 2(a)) Howeverdespite the drastic decrease in cell proliferation the viabilityof cells treated with the highest concentration of COLE(150 120583gmL) remains relatively high (80) for up to 3 daysof treatment (Figure 3(b))
Since it is well established that the arrest of the cellcycle progression allows the cells to follow other processessuch as apoptosis and differentiation we then explored theeffect of COLE on the cell cycle distribution (Table 2) Oliveleaf extract has been shown to cause cell cycle arrest atG0G1 phase in both the breast cancer MCF-7 cells and themelanoma B16 cells [12 25] However the treatment timesapplied in both of these studies were limited to 48 h and 18 hrespectively In this study COLE treated cells were arrested atG0G1 on the 1st and 2ndday of treatment which is consistentwith the reported studies Prolonged incubation with COLEshowed an arrest of cell cycle at G2M phase (3rd and 4thday of incubation) The microarray analysis performed forthe 3rd day of treatment revealed a significant increase ofCHEK2 gene expression in COLE treated cells The encodedprotein regulates the cell cycle checkpoint arrest throughthe inhibition of the activity of CDC25A CDC25B andCDC25C [28] CDC25 proteins trigger the entry into mitosisat different points of the cell cycle by activating theCdk-cyclincomplexes CDC25A acts early in the cell cycle regulatingthe G1S transition whereas CDC25B and CDC25C act atG2M [29] Interestingly our results showed a decrease in theexpression of CDC25C gene in COLE treated cells associatedwith an increase in the expression of CDC25A which mayexplain the cell cycle arrest at G2M phase observed at the3rd day of treatment with COLE
Treatment with COLE increased the expression of CD14on the cell surface of treated cells indicating the differenti-ation of K562 cells toward the mono-macrophage lineage(Figure 5(a)) The increase in the percentage of cells positivefor CD11b (Figure 5(b)) confirms this hypothesis In factCD11b is expressed on both monocytes and granulocytesThus together with the increase of CD14 expression theexpression of CD11b suggests that treatment with COLEpromotes the commitment of K562 cells into the mono-cytemacrophage lineage at the expense of the granulocyticdifferentiationThe instant increase of the expression of CD41
on the 1st day of treatment and its absence during the fol-lowing days (Figure 5(c)) could be consistent with monocytedifferentiation instead of megakaryocyte differentiation sinceit has been reported that 120572IIb promoter which initiates thetranscription of CD41 is transcriptionally active in pluripo-tent myeloid progenitors in early stages of erythropoiesisand all along the megakaryocytic differentiation as well asto a lesser extent in the early stages of myelomonocyticdifferentiation and the late stages of erythropoiesis [30] Thisinstant increase could be also explained by the phenomenonof lineage conversion where some hematopoietic progenitorscan be converted into lineages other than their own bythe ectopic expression of some transcription factors [31]In this regard it was previously reported that commonlymphoid progenitors megakaryocyte erythroid progenitorsearly B cells and early T cells could be converted to thegranulocytemonocyte lineage by the induction of CEBP120572activity [32] Microarray results support the absence ofmegakaryocyte differentiation in the 3rd day of treatment Infact data showed a decrease in the expression of genes relatedtomegakaryopoiesis such as the essential regulator of plateletrelease NFE2 and its main candidate target gene TUBB1(encoding the megakaryocyte and platelet specific isoform of120573-tubulin (1205731)) [33] as well as the BACH2 transcription factorreported to be induced during megakaryocyte differentiation[34]
Interestingly results of microarray analysis (Table 3)showed the upregulation of several genes already reportedas key players in monocytemacrophage differentiation inthe COLE treated cells Among these genes the expressionof IFI16 was markedly increased Constitutively expressedin lymphoid cells IFI16 was shown to be associated withthe differentiation of human myeloid cells in response tointerferon gamma [35] Moreover IFI16 has been selectivelyexpressed during the differentiation of myeloid progenitorcells along monocytic lineage while it is downregulatedduring the commitment toward the granulocytic or erythroidlineage suggesting its potential involvement in the differenti-ation andmaturation of the monocytic lineage [36 37] Geneencoding for EGR-1 a positive modulator of macrophagedifferentiation was also significantly upregulated EGR-1 isknown to dictate development of myeloid progenitors alongthe macrophage lineage at the expense of development alongother lineages [38 39] Dauffy et al [40] reported thatIFI16-enforced expression in myeloid progenitors inducedthe expression of EGR-1 and conducted the macrophagedifferentiation in the absence of the macrophage colonystimulating factor (M-CSF) This suggests that EGR-1 couldbe regulated by the expression of IFI16 Treatment with COLEincreased as well the expression of NFYA encoding the Asubunit of the nuclear transcription factor Y whose synthesiswas previously reported during the terminal differentiationof monocytes to macrophages [38 41] The downregula-tion of FOXP1 in COLE treated K562 cells is consistentwith monocytemacrophage differentiation since it has beendemonstrated that the expression of FOXP1 was markedlydecreased in monocyte-induced differentiation of HL-60cells as well as in human peripheral blood monocytes and
Oxidative Medicine and Cellular Longevity 13
that the overexpression of FOXP1 prevented the morphologicmacrophage-like differentiation [42 43]
Monocytes and macrophages have several functionalcharacteristics including cell adhesion migration chemo-taxis and phagocytic activity Interestingly an enhancedexpression of the chemokines genes CXCL8 (IL8) CXCL2andCXCL3was found in COLE treated cells IL-8 productionhas been observed in vitro in a wide variety of cells includingmonocytes T lymphocytes neutrophils vascular endothe-lial cells dermal fibroblasts keratinocytes hepatocytes andhuman gastric cancer cells while the production of CXCL2andCXCL3was only described inmonocytes fibroblasts andendothelial cells [44 45] Another gene related to chemotaxiswas upregulated NUP85 which is an essential component ofthe nuclear pore complex and was reported to be involved inCCR2-mediated chemotaxis of monocytes [46]
COLE treatment also enhanced the expression of somegenes related to phagocytosis AP1G1 andRab proteins relatedgenes AP1G1 is a subunit of clathrin-associated adaptorprotein complex 1 whose role has been recently highlightedfor efficient phagocytosis at an early stage of phagosomeformation since it participates in the extension of the phago-cytic cup [47 48] The small GTPases Rab are key regulatorsof intracellular membrane trafficking Recent studies havedemonstrated that several RAB proteins play an importantrole in phagocytosis RAB proteins such as RAB11 Rab5 andRab 21 participate in the phagosome formation and matura-tion and are necessary for the phagocytic activity [48ndash51]
Consistently with the increase in Annexin V positivecells (Figure 4) treatment with COLE upregulated someproapoptotic genes such as CASP6 CASP8 DFFA and BIDand downregulated the apoptosis suppressor BCL2 as wellas the caspase inhibitors genes (Table 3) Olive leaf waspreviously shown to induce morphological changes thatare characteristic of apoptosis in leukemia cells but themechanisms underlying this effect were not well investi-gated [13 21] The increase in the proportion of apoptoticcells observed from the 4th day of treatment with COLEcould be explained by the programmed death of the fullydifferentiated cells (Figure 4) In fact this increase wasaccompanied by an important decrease in cell viability aswell as a diminution in CD14 expression (Figures 3(b) and5(a)) Elsewhere it is well established that in hematopoietictissues apoptosis is coupled to terminal differentiation ofmyeloid progenitors even though themechanism responsiblefor the activation of apoptosis during myeloid maturationis still poorly understood [52 53] It is also noteworthy tomention that several reports have highlighted the role ofcaspases in the terminal differentiation of a variety of celltypes [54] One example is the differentiation of human bloodmonocytes into macrophages a process that is blocked bysynthetic caspase inhibitors [55] It has been also reportedthat caspase 8 deletion in bone-marrow cells resulted in arrestof hemopoietic progenitor functioning and that its deletionin cells of the myelomonocytic lineage led to the arrest ofdifferentiation into macrophages and consequently to celldeath [56]
Among the differentially expressed genes in COLEtreated cells (Table 3) genes encoding the mitogen-activated
protein kinase kinase kinases (MAPKKKs) MAP3K2MAP3K5 and MAP3K7 were highly expressed whileMAP2K5 and MAPK14p38-120572 were downregulated TheMAPKKKs act at the upstream of the MAP kinase cascadewhich regulates important cellular processes such as geneexpression cell proliferation differentiation cell survivaland death The MAP3K5 activates the c-Jun N-terminalkinase (JNK) and p38 mitogen-activated protein kinases[57] while the MAP3K2 is reported to preferentially activateJNK [58 59] However the significant decrease in theMAPK14p38-120572 may suggest that the effect of COLE onK562 cells is likely to be mediated by the JNK MAPKsrather than the p38 MAPKs JNK plays an important rolein apoptosis pathways weather by activating proapoptoticor antiapoptotic genes Particularly it was shown thatprolonged but not transient JNK activation promotes thecell death [60] Our results showed that JKAMP gene whichencodes for a membrane-anchored regulator of the durationof JNK1 activity was upregulated (154 plusmn 0051) The elevatedexpression of JKAMP as reported by Kadoya et al [61]results in sustained JNK activity These findings may indicatethe potential role of JNK pathway in the COLE-inducedapoptosis in K562 cells
Thanks to their large regulatory domains some MAP-KKKs can interact with upstream regulators have functionssuch as ubiquitylation and be activated by relief of autoin-hibition and oligomerization [62] In this context MAP3K2has been reported to play an important role in NF-kappaBsignaling pathway by activating the I-kappaB kinases whichin turn phosphorylate the inhibitory factors of the nuclearfactor kappaB (NF-kappaB) resulting in their rapid ubiq-uitination and the liberation of NF-kappaB complex whichtranslocate from the cytoplasm to the nucleus [63] MAP3K7in association with other proteins could also be required forthe activation of NF-kappaB According to the GO analysis(Table S1) COLE treatment upregulated genes implicatedin NF-kappaB-related categories such as positive regulationof NF-kappaB transcription factor activity I-kappaB kinase-NF-kappaB cascade and its positive regulation and I-kappaBphosphorylation Genes encoding the signal transducers inthe NF-kappaB pathway TRAF5 and TRAF6 as well asthe activator of NF-kappaB signaling SNIP1 were upregu-lated Interestingly an increased expression of NFKB1 geneencoding for the precursor of the mature NF-kappaB p50was detected in COLE treated cells compared to untreatedK562 cells NFKB is well recognized as a central activator ofthe antiapoptotic cascades in response to external stimuli orintrinsic immune reactions and its prosurvival activity hasbeen implicated in a variety of biological processes [60 64]In the B cell lineage the activity of NF-120581B is required forthe completion of various developmental stages includingdifferentiation and the response of these cells to antigens [65]NF-kappaB also plays an important role in the survival anddevelopment of T cells [66] and has been recently reported tomediate the differentiation of mesenchymal stem cells [67]
Several studies have demonstrated a crosstalk betweenthe NF-kappaB and JNK pathways seen as an inhibitoryeffect of NF-kappaB on apoptosis through the suppressionof JNK activity and that this suppressive effect may occur
14 Oxidative Medicine and Cellular Longevity
through different mechanisms [60] This suggests that thebalance between JNK and NF-kappaB activities is crucial todetermine the cell fate survival or deathWehypothesize herethat COLE-induced apoptosis or differentiation is likely to bedetermined by the crosstalk between these two pathways inK562 cells Further investigations regarding protein expres-sion and activity are needed to confirm these findings
5 Conclusions
Our study demonstrates for the first time that olive leafextract exhibits an antileukemia effect on the human chronicmyeloid leukemia cells Olive leaf extract was shown toinhibit the proliferation of K562 cells by inducing cell cyclearrest apoptosis and differentiation toward the monocytelineage The induced expression of molecules involved indifferentiation toward the monocytemacrophage lineage aswell asmolecules related to apoptosis and cell cycle regulationconfirmed these findings and provides insights into themechanism by which olive leaf exhibits its antileukemiaeffect
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported by the JICA-JST Science andTechnology Research Partnership for Sustainable Develop-ment (SATREPS) Project ldquoValorization of Bio-Resources inSemi-Arid and Arid Land for Regional Developmentrdquo
References
[1] M Leszczyniecka T Roberts P Dent S Grant and P B FisherldquoDifferentiation therapy of human cancer basic science andclinical applicationsrdquo Pharmacology and Therapeutics vol 90no 2-3 pp 105ndash156 2001
[2] A I Spira and M A Carducci ldquoDifferentiation therapyrdquoCurrent Opinion in Pharmacology vol 3 no 4 pp 338ndash3432003
[3] S Nobili D Lippi E Witort et al ldquoNatural compounds forcancer treatment and preventionrdquo Pharmacological Researchvol 59 no 6 pp 365ndash378 2009
[4] S Ramos ldquoEffects of dietary flavonoids on apoptotic pathwaysrelated to cancer chemopreventionrdquo Journal of NutritionalBiochemistry vol 18 no 7 pp 427ndash442 2007
[5] V Hajhashemi G Vaseghi M Pourfarzam and A AbdollahildquoAre antioxidants helpful for disease preventionrdquo Research inPharmaceutical Sciences vol 5 no 1 pp 1ndash8 2010
[6] M Bouaziz and S Sayadi ldquoIsolation and evaluation of antiox-idants from leaves of a Tunisian cultivar olive treerdquo EuropeanJournal of Lipid Science andTechnology vol 107 no 7-8 pp 497ndash504 2005
[7] A P Pereira I C F R Ferreira F Marcelino et al ldquoPhenoliccompounds and antimicrobial activity of olive (Olea europaeaL Cv Cobrancosa) leavesrdquo Molecules vol 12 no 5 pp 1153ndash1162 2007
[8] L I Somova F O Shode P Ramnanan and A Nadar ldquoAnti-hypertensive antiatherosclerotic and antioxidant activity oftriterpenoids isolated from Olea europaea subspecies africanaleavesrdquo Journal of Ethnopharmacology vol 84 no 2-3 pp 299ndash305 2003
[9] V Micol N Caturla L Perez-Fons V Mas L Perez andA Estepa ldquoThe olive leaf extract exhibits antiviral activityagainst viral haemorrhagic septicaemia rhabdovirus (VHSV)rdquoAntiviral Research vol 66 no 2-3 pp 129ndash136 2005
[10] J Wainstein T Ganz M Boaz et al ldquoOlive leaf extract as ahypoglycemic agent in both human diabetic subjects and inratsrdquo Journal of Medicinal Food vol 15 no 7 pp 605ndash610 2012
[11] L Seddik TM Bah A AouesM Slimani andM BenderdourldquoElucidation of mechanisms underlying the protective effects ofolive leaf extract against lead-induced neurotoxicity in Wistarratsrdquo Journal of Toxicological Sciences vol 36 no 6 pp 797ndash809 2011
[12] Z Bouallagui J Han H Isoda and S Sayadi ldquoHydroxytyrosolrich extract from olive leaves modulates cell cycle progressionin MCF-7 human breast cancer cellsrdquo Food and ChemicalToxicology vol 49 no 1 pp 179ndash184 2011
[13] R Fares S Bazzi S E Baydoun and R M Abdel-Massih ldquoTheantioxidant and anti-proliferative activity of the Lebanese Oleaeuropaea extractrdquo Plant Foods for Human Nutrition vol 66 no1 pp 58ndash63 2011
[14] V Goulas V Exarchou A N Troganis et al ldquoPhytochemicalsin olive-leaf extracts and their antiproliferative activity againstcancer and endothelial cellsrdquo Molecular Nutrition and FoodResearch vol 53 no 5 pp 600ndash608 2009
[15] M de Bock E B Thorstensen J G Derraik H V HendersonP L Hofman and W S Cutfield ldquoHuman absorption andmetabolism of oleuropein and hydroxytyrosol ingested as olive(Olea europaea L) leaf extractrdquo Molecular Nutrition and FoodResearch vol 57 no 11 pp 2079ndash2085 2013
[16] R Briante M Patumi S Terenziani E Bismuto F Febbraioand R Nucci ldquoOlea europaea L leaf extract and derivativesantioxidant propertiesrdquo Journal of Agricultural and Food Chem-istry vol 50 no 17 pp 4934ndash4940 2002
[17] O Benavente-Garcıa J Castillo J Lorente A Ortuno and J ADel Rio ldquoAntioxidant activity of phenolics extracted from Oleaeuropaea L leavesrdquo Food Chemistry vol 68 no 4 pp 457ndash4622000
[18] S H Omar ldquoOleuropein in olive and its pharmacologicaleffectsrdquo Scientia Pharmaceutica vol 78 no 2 pp 133ndash154 2010
[19] H Jemai M Bouaziz I Fki A El Feki and S SayadildquoHypolipidimic and antioxidant activities of oleuropein and itshydrolysis derivative-rich extracts from Chemlali olive leavesrdquoChemico-Biological Interactions vol 176 no 2-3 pp 88ndash982008
[20] R Garcıa-Villalba M Larrosa S Possemiers F A Tomas-Barberan and J C Espın ldquoBioavailability of phenolics from anoleuropein-rich olive (Olea europaea) leaf extract and its acuteeffect on plasma antioxidant status comparison between pre-and postmenopausal womenrdquo European Journal of Nutrition2013
[21] J Anter Z Fernandez-Bedmar M Villatoro-Pulido et al ldquoApilot study on the DNA-protective cytotoxic and apoptosis-inducing properties of olive-leaf extractsrdquo Mutation Researchvol 723 no 2 pp 165ndash170 2011
[22] J A Sutherland A R Turner and P Mannoni ldquoDifferen-tiation of K562 leukemia cells along erythroid macrophage
Oxidative Medicine and Cellular Longevity 15
and megakaryocyte lineagesrdquo Journal of Biological ResponseModifiers vol 5 no 3 pp 250ndash262 1986
[23] K Schwenke H-P Peterson K-H Wangenheim and L EFeinendegen ldquoInduction of differentiation in erythroleukemicK562 cells by 120574-irradiationrdquo Leukemia Research vol 19 no 12pp 955ndash961 1995
[24] V N Enujiugha J Y Talabi S A Malomo and A I OlagunjuldquoDPPH radical scavenging capacity of phenolic extracts fromAfrican Yam Bean (Sphenostylis stenocarpa)rdquo Food and Nutri-tion Sciences vol 3 pp 7ndash13 2012
[25] S A Mijatovic G S Timotijevic D M Miljkovic et alldquoMultiple antimelanoma potential of dry olive leaf extractrdquoInternational Journal of Cancer vol 128 no 8 pp 1955ndash19652011
[26] L Abaza T P N Talorete P Yamada Y KuritaM Zarrouk andH Isoda ldquoInduction of growth inhibition and differentiation ofhuman leukemia HL-60 cells by a Tunisian Gerboui olive leafextractrdquo Bioscience Biotechnology and Biochemistry vol 71 no5 pp 1306ndash1312 2007
[27] B Tunca G Tezcan G Cecener et al ldquoOlea europea leafextract alters microRNA expression in human glioblastomacellsrdquo Journal of Cancer Research and Clinical Oncology vol 138no 11 pp 1831ndash1844 2012
[28] S Matsuoka M Huang and S J Elledge ldquoLinkage of ATM tocell cycle regulation by the Chk2 protein kinaserdquo Science vol282 no 5395 pp 1893ndash1897 1998
[29] C Frazer and P G Young ldquoPhosphorylation mediated regu-lation of Cdc25 activity localization and stabilityrdquo in ProteinPhosphorylation in Human Health C Huang Ed pp 395ndash436InTech 2012
[30] N Debili C Robin V Schiavon et al ldquoDifferent expression ofCD41 on human lymphoid andmyeloid progenitors from adultsand neonatesrdquo Blood vol 97 no 7 pp 2023ndash2030 2001
[31] H Nakajima ldquoRole of transcription factors in differentiationand reprogramming of hematopoietic cellsrdquo Keio Journal ofMedicine vol 60 no 2 pp 47ndash55 2011
[32] Y Fukuchi F Shibata M Ito et al ldquoComprehensive analysis ofmyeloid lineage conversion using mice expressing an inducibleform of CEBP120572rdquoThe EMBO Journal vol 25 no 14 pp 3398ndash3410 2006
[33] R A Shivdasani ldquoMolecular and transcriptional regulation ofmegakaryocyte differentiationrdquo Stem Cells vol 19 no 5 pp397ndash407 2001
[34] K Terui Y Takahashi J Kitazawa T TokiM Yokoyama and EIto ldquoExpression of transcription factors during megakaryocyticdifferentiation of CD34+ cells from human cord blood inducedby thrombopoietinrdquo Tohoku Journal of Experimental Medicinevol 192 no 4 pp 259ndash273 2000
[35] J A Trapani M Dawson V A Apostolidis and K A BrowneldquoGenomic organization of IFI16 an interferon-inducible genewhose expression is associated with human myeloid cell differ-entiation correlation of predicted protein domains with exonorganizationrdquo Immunogenetics vol 40 no 6 pp 415ndash424 1994
[36] M J Dawson N J Elwood R W Johnstone and J A TrapanildquoThe IFN-inducible nucleoprotein IFI 16 is expressed in cellsof the monocyte lineage but is rapidly and markedly down-regulated in other myeloid precursor populationsrdquo Journal ofLeukocyte Biology vol 64 no 4 pp 546ndash554 1998
[37] M Gariglio M De Andrea M Lembo et al ldquoThe murinehomolog of the HIN 200 family Ifi 204 is constitutivelyexpressed in myeloid cells and selectively induced in the
monocytemacrophage lineagerdquo Journal of Leukocyte Biologyvol 64 no 5 pp 608ndash614 1998
[38] A F Valledor F E Borras M Cullell-Young and A CeladaldquoTranscription factors that regulate monocytemacrophage dif-ferentiationrdquo Journal of Leukocyte Biology vol 63 no 4 pp405ndash417 1998
[39] K Krishnaraju B Hoffman and D A Liebermann ldquoEarlygrowth response gene 1 stimulates development of hematopoi-etic progenitor cells along the macrophage lineage at theexpense of the granulocyte and erythroid lineagesrdquo Blood vol97 no 5 pp 1298ndash1305 2001
[40] J Dauffy G Mouchiroud and R P Bourette ldquoThe interferon-inducible gene Ifi204 is transcriptionally activated in responseto M-CSF and its expression favors macrophage differentiationin myeloid progenitor cellsrdquo Journal of Leukocyte Biology vol79 no 1 pp 173ndash183 2006
[41] G Marziali E Perrotti R Ilari U Testa E M Coccia andA Battistini ldquoTranscriptional regulation of the ferritin heavy-chain gene the activity of the CCAAT binding factor NF-Y ismodulated in heme-treated Friend leukemia cells and duringmonocyte-to-macrophage differentiationrdquo Molecular and Cel-lular Biology vol 17 no 3 pp 1387ndash1395 1997
[42] C Shi X Zhang Z Chen et al ldquoIntegrin engagement regulatesmonocyte differentiation through the forkhead transcriptionfactor Foxp1rdquo Journal of Clinical Investigation vol 114 no 3 pp408ndash418 2004
[43] C Shi M Sakuma T Mooroka et al ldquoDown-regulation of theforkhead transcription factor Foxp1 is required for monocytedifferentiation andmacrophage functionrdquoBlood vol 112 no 12pp 4699ndash4711 2008
[44] D F Smith E Galkina K Ley and Y Huo ldquoGRO familychemokines are specialized for monocyte arrest from flowrdquoThe American Journal of Physiology Heart and CirculatoryPhysiology vol 289 no 5 pp H1976ndashH1984 2005
[45] J J Oppenheim O M Z Howard and E Goetzl ChemotacticFactors Neuropeptides and Other Ligands for Seven Transmem-brane Receptors 2000
[46] Y Terashima N Onai M Murai et al ldquoPivotal function forcytoplasmic protein FROUNT in CCR2-mediated monocytechemotaxisrdquo Nature Immunology vol 6 no 8 pp 827ndash8352005
[47] Y Lefkir M Malbouyres D Gotthardt et al ldquoInvolvement ofthe AP-1 adaptor complex in early steps of phagocytosis andmacropinocytosisrdquo Molecular Biology of the Cell vol 15 no 2pp 861ndash869 2004
[48] F Niedergang and P Chavrier ldquoSignaling and membranedynamics during phagocytosis many roads lead to the phago-somerdquo Current Opinion in Cell Biology vol 16 no 4 pp 422ndash428 2004
[49] D Cox D J Lee B M Dale J Calafat and S Green-berg ldquoA Rab11-containing rapidly recycling compartment inmacrophages that promotes phagocytosisrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 97 no 2 pp 680ndash685 2000
[50] O V Vieira C Bucci R E Harrison et al ldquoModulation of Rab5and Rab7 recruitment to phagosomes by phosphatidylinositol3-kinaserdquoMolecular andCellular Biology vol 23 no 7 pp 2501ndash2514 2003
[51] T Khurana J A Brzostowski and A R Kimmel ldquoARab21LIM-onlyCH-LIM complex regulates phagocytosis viaboth activating and inhibitory mechanismsrdquo The EMBO Jour-nal vol 24 no 13 pp 2254ndash2264 2005
16 Oxidative Medicine and Cellular Longevity
[52] S J Martin J G Bradley and T G Cotter ldquoHL-60 cellsinduced to differentiate towards neutrophils subsequently dievia apoptosisrdquo Clinical and Experimental Immunology vol 79no 3 pp 448ndash453 1990
[53] A Benito D Grillot G Nunez and J L Fernandez-Luna ldquoReg-ulation and function of Bcl-2 during differentiation-inducedcell death in HL-60 promyelocytic cellsrdquoThe American Journalof Pathology vol 146 no 2 pp 481ndash490 1995
[54] M Lamkanfi N Festjens W Declercq T V Berghe andP Vandenabeele ldquoCaspases in cell survival proliferation anddifferentiationrdquo Cell Death and Differentiation vol 14 no 1 pp44ndash55 2007
[55] O Sordet C Rebe S Plenchette et al ldquoSpecific involvement ofcaspases in the differentiation of monocytes into macrophagesrdquoBlood vol 100 no 13 pp 4446ndash4453 2002
[56] T-B Kang T Ben-Moshe E E Varfolomeev et al ldquoCaspase-8 serves both apoptotic and nonapoptotic rolesrdquo Journal ofImmunology vol 173 no 5 pp 2976ndash2984 2004
[57] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[58] J Cheng J Yang Y Xia M Karin and B Su ldquoSynergisticinteraction of MEK kinase 2 c-Jun N-terminal kinase (JNK)kinase 2 and JNK1 results in efficient and specific JNK1activationrdquo Molecular and Cellular Biology vol 20 no 7 pp2334ndash2342 2000
[59] J L Blank P Gerwins EM Elliott S Sather andG L JohnsonldquoMolecular cloning of mitogen-activated proteinERK kinasekinases (MEKK) 2 and 3 regulation of sequential phosphoryla-tion pathways involving mitogen-activated protein kinase andc-Jun kinaserdquo Journal of Biological Chemistry vol 271 no 10pp 5361ndash5368 1996
[60] S Papa F Zazzeroni C G Pham C Bubici and G FranzosoldquoLinking JNK signaling to NF-120581B a key to survivalrdquo Journal ofCell Science vol 117 no 22 pp 5197ndash5208 2004
[61] T Kadoya A Khurana M Tcherpakov et al ldquoJAMP a JUB N-terminal kinase 1 (JNK1)-associated membrane protein regu-lates duration of JNK activityrdquo Molecular and Cellular Biologyvol 25 no 19 pp 8619ndash8630 2005
[62] M Qi and E A Elion ldquoMAP kinase pathwaysrdquo Journal of CellScience vol 118 no 16 pp 3569ndash3572 2005
[63] Q Zhao and F S Lee ldquoMitogen-activated protein kinaseERKkinase kinases 2 and 3 activate nuclear factor-120581B through I120581Bkinase-120572 and I120581B kinase-120573rdquoThe Journal of Biological Chemistryvol 274 no 13 pp 8355ndash8358 1999
[64] H Namba V Saenko and S Yamashita ldquoNuclear factor-120581Bin thyroid carcinogenesis and progression a novel therapeutictarget for advanced thyroid cancerrdquo Arquivos Brasileiros deEndocrinologia e Metabologia vol 51 no 5 pp 843ndash851 2007
[65] B Feng S Cheng W S Pear and H-C Liou ldquoNF-kBinhibitor blocks B cell development at two checkpointsrdquoMedi-cal Immunology vol 3 article 1 2004
[66] S Gerondakis and U Siebenlist ldquoRoles of the NF-kappaBpathway in lymphocyte development and functionrdquoCold SpringHarbor Perspectives in Biology vol 2 no 5 Article ID a0001822010
[67] K P Bhat V Balasubramaniyan B Vaillant et al ldquoMesenchy-mal differentiation mediated by NF-kB promotes radiationresistance in glioblastomardquo Cancer Cell vol 24 no 3 pp 331ndash346 2013
Figure 5 Expression of differentiationmarkers onK562 cells treatedwith Chemlali Olive Leaf Extract (COLE) up to 6 days Cells were treatedwith 50 100 and 150 120583gmL of COLE and incubated for different periods Control cells were treated with ethanol at a final concentration of03 Expression of cell surface markers was detected by flow cytometry (a) Expression of CD14 (monocytemacrophage specific marker)(b) Expression of CD11b (granulocytemonocyte specific marker) (c) Expression of CD41 (megakaryocyte specific marker) (d) Expressionof glycophorin A (GPA) (erythrocyte specific marker) Results are represented as the mean plusmn SD of three independent experiments lowast lowastlowastsignificantly different from the control at 119875 lt 005 and 119875 lt 001 respectively
including GYPA HBE1 FECH and ALAS2 which is consis-tent with the decreased expression of erythrocyte differenti-ation marker in the flow cytometry results (Figure 5(d))
Analysis of the microarray data showed the upregu-lation of several proapoptotic genes and genes involvedin the regulation of mitochondrial membrane permeabilitysuch as CASP6 CASP8 DFFA and BID On the otherhandexpression of genes negatively regulating the apoptosis
and those inhibiting the caspase activity was significantlydecreased such as IGF1R HSPA5 and BCL2
Among the upregulated genes we found those related tothe NF-kappaB cascade as well as to the MAPKKK cascadeand the Wnt receptor signaling pathway
Categories related to cell cycle regulation and cell pro-liferation were represented by both upregulated and down-regulated genes Finally some transcription factors werealso downregulated such as FOXP1 whereas others wereupregulated like EGR1 and NFYA
The list of selected genes is presented in Table 3 Fromthis list it is clear that the differentially expressed genes aftertreatment with 100 and 150120583gmL have mostly the sametendency which could validate the obtained results
12 Oxidative Medicine and Cellular Longevity
4 Discussion
Previous reports had indicated that olive leaf extract inducesapoptosis in the human leukemic cell lines HL-60 and Jurkatcells [13 21] In this study we give evidence that COLEexhibits its antileukemia effect by both inducing apoptosisand promoting differentiation of the multipotent humanleukemia K562 cells
Induction of differentiation as well as apoptosis has beenfrequently reported to be associatedwith a loss of proliferativecapacity of the cells It has been reported that olive leaf extractinhibits the proliferation of some cancer cell lines such asB16 melanoma cells HL-60 leukemia cells the breast cancerMCF-7 cells and the glioblastoma cells [12 25ndash27] Ourresults indicate that COLE inhibits the proliferation of K562cells in a dose dependent manner (Figure 2(a)) Howeverdespite the drastic decrease in cell proliferation the viabilityof cells treated with the highest concentration of COLE(150 120583gmL) remains relatively high (80) for up to 3 daysof treatment (Figure 3(b))
Since it is well established that the arrest of the cellcycle progression allows the cells to follow other processessuch as apoptosis and differentiation we then explored theeffect of COLE on the cell cycle distribution (Table 2) Oliveleaf extract has been shown to cause cell cycle arrest atG0G1 phase in both the breast cancer MCF-7 cells and themelanoma B16 cells [12 25] However the treatment timesapplied in both of these studies were limited to 48 h and 18 hrespectively In this study COLE treated cells were arrested atG0G1 on the 1st and 2ndday of treatment which is consistentwith the reported studies Prolonged incubation with COLEshowed an arrest of cell cycle at G2M phase (3rd and 4thday of incubation) The microarray analysis performed forthe 3rd day of treatment revealed a significant increase ofCHEK2 gene expression in COLE treated cells The encodedprotein regulates the cell cycle checkpoint arrest throughthe inhibition of the activity of CDC25A CDC25B andCDC25C [28] CDC25 proteins trigger the entry into mitosisat different points of the cell cycle by activating theCdk-cyclincomplexes CDC25A acts early in the cell cycle regulatingthe G1S transition whereas CDC25B and CDC25C act atG2M [29] Interestingly our results showed a decrease in theexpression of CDC25C gene in COLE treated cells associatedwith an increase in the expression of CDC25A which mayexplain the cell cycle arrest at G2M phase observed at the3rd day of treatment with COLE
Treatment with COLE increased the expression of CD14on the cell surface of treated cells indicating the differenti-ation of K562 cells toward the mono-macrophage lineage(Figure 5(a)) The increase in the percentage of cells positivefor CD11b (Figure 5(b)) confirms this hypothesis In factCD11b is expressed on both monocytes and granulocytesThus together with the increase of CD14 expression theexpression of CD11b suggests that treatment with COLEpromotes the commitment of K562 cells into the mono-cytemacrophage lineage at the expense of the granulocyticdifferentiationThe instant increase of the expression of CD41
on the 1st day of treatment and its absence during the fol-lowing days (Figure 5(c)) could be consistent with monocytedifferentiation instead of megakaryocyte differentiation sinceit has been reported that 120572IIb promoter which initiates thetranscription of CD41 is transcriptionally active in pluripo-tent myeloid progenitors in early stages of erythropoiesisand all along the megakaryocytic differentiation as well asto a lesser extent in the early stages of myelomonocyticdifferentiation and the late stages of erythropoiesis [30] Thisinstant increase could be also explained by the phenomenonof lineage conversion where some hematopoietic progenitorscan be converted into lineages other than their own bythe ectopic expression of some transcription factors [31]In this regard it was previously reported that commonlymphoid progenitors megakaryocyte erythroid progenitorsearly B cells and early T cells could be converted to thegranulocytemonocyte lineage by the induction of CEBP120572activity [32] Microarray results support the absence ofmegakaryocyte differentiation in the 3rd day of treatment Infact data showed a decrease in the expression of genes relatedtomegakaryopoiesis such as the essential regulator of plateletrelease NFE2 and its main candidate target gene TUBB1(encoding the megakaryocyte and platelet specific isoform of120573-tubulin (1205731)) [33] as well as the BACH2 transcription factorreported to be induced during megakaryocyte differentiation[34]
Interestingly results of microarray analysis (Table 3)showed the upregulation of several genes already reportedas key players in monocytemacrophage differentiation inthe COLE treated cells Among these genes the expressionof IFI16 was markedly increased Constitutively expressedin lymphoid cells IFI16 was shown to be associated withthe differentiation of human myeloid cells in response tointerferon gamma [35] Moreover IFI16 has been selectivelyexpressed during the differentiation of myeloid progenitorcells along monocytic lineage while it is downregulatedduring the commitment toward the granulocytic or erythroidlineage suggesting its potential involvement in the differenti-ation andmaturation of the monocytic lineage [36 37] Geneencoding for EGR-1 a positive modulator of macrophagedifferentiation was also significantly upregulated EGR-1 isknown to dictate development of myeloid progenitors alongthe macrophage lineage at the expense of development alongother lineages [38 39] Dauffy et al [40] reported thatIFI16-enforced expression in myeloid progenitors inducedthe expression of EGR-1 and conducted the macrophagedifferentiation in the absence of the macrophage colonystimulating factor (M-CSF) This suggests that EGR-1 couldbe regulated by the expression of IFI16 Treatment with COLEincreased as well the expression of NFYA encoding the Asubunit of the nuclear transcription factor Y whose synthesiswas previously reported during the terminal differentiationof monocytes to macrophages [38 41] The downregula-tion of FOXP1 in COLE treated K562 cells is consistentwith monocytemacrophage differentiation since it has beendemonstrated that the expression of FOXP1 was markedlydecreased in monocyte-induced differentiation of HL-60cells as well as in human peripheral blood monocytes and
Oxidative Medicine and Cellular Longevity 13
that the overexpression of FOXP1 prevented the morphologicmacrophage-like differentiation [42 43]
Monocytes and macrophages have several functionalcharacteristics including cell adhesion migration chemo-taxis and phagocytic activity Interestingly an enhancedexpression of the chemokines genes CXCL8 (IL8) CXCL2andCXCL3was found in COLE treated cells IL-8 productionhas been observed in vitro in a wide variety of cells includingmonocytes T lymphocytes neutrophils vascular endothe-lial cells dermal fibroblasts keratinocytes hepatocytes andhuman gastric cancer cells while the production of CXCL2andCXCL3was only described inmonocytes fibroblasts andendothelial cells [44 45] Another gene related to chemotaxiswas upregulated NUP85 which is an essential component ofthe nuclear pore complex and was reported to be involved inCCR2-mediated chemotaxis of monocytes [46]
COLE treatment also enhanced the expression of somegenes related to phagocytosis AP1G1 andRab proteins relatedgenes AP1G1 is a subunit of clathrin-associated adaptorprotein complex 1 whose role has been recently highlightedfor efficient phagocytosis at an early stage of phagosomeformation since it participates in the extension of the phago-cytic cup [47 48] The small GTPases Rab are key regulatorsof intracellular membrane trafficking Recent studies havedemonstrated that several RAB proteins play an importantrole in phagocytosis RAB proteins such as RAB11 Rab5 andRab 21 participate in the phagosome formation and matura-tion and are necessary for the phagocytic activity [48ndash51]
Consistently with the increase in Annexin V positivecells (Figure 4) treatment with COLE upregulated someproapoptotic genes such as CASP6 CASP8 DFFA and BIDand downregulated the apoptosis suppressor BCL2 as wellas the caspase inhibitors genes (Table 3) Olive leaf waspreviously shown to induce morphological changes thatare characteristic of apoptosis in leukemia cells but themechanisms underlying this effect were not well investi-gated [13 21] The increase in the proportion of apoptoticcells observed from the 4th day of treatment with COLEcould be explained by the programmed death of the fullydifferentiated cells (Figure 4) In fact this increase wasaccompanied by an important decrease in cell viability aswell as a diminution in CD14 expression (Figures 3(b) and5(a)) Elsewhere it is well established that in hematopoietictissues apoptosis is coupled to terminal differentiation ofmyeloid progenitors even though themechanism responsiblefor the activation of apoptosis during myeloid maturationis still poorly understood [52 53] It is also noteworthy tomention that several reports have highlighted the role ofcaspases in the terminal differentiation of a variety of celltypes [54] One example is the differentiation of human bloodmonocytes into macrophages a process that is blocked bysynthetic caspase inhibitors [55] It has been also reportedthat caspase 8 deletion in bone-marrow cells resulted in arrestof hemopoietic progenitor functioning and that its deletionin cells of the myelomonocytic lineage led to the arrest ofdifferentiation into macrophages and consequently to celldeath [56]
Among the differentially expressed genes in COLEtreated cells (Table 3) genes encoding the mitogen-activated
protein kinase kinase kinases (MAPKKKs) MAP3K2MAP3K5 and MAP3K7 were highly expressed whileMAP2K5 and MAPK14p38-120572 were downregulated TheMAPKKKs act at the upstream of the MAP kinase cascadewhich regulates important cellular processes such as geneexpression cell proliferation differentiation cell survivaland death The MAP3K5 activates the c-Jun N-terminalkinase (JNK) and p38 mitogen-activated protein kinases[57] while the MAP3K2 is reported to preferentially activateJNK [58 59] However the significant decrease in theMAPK14p38-120572 may suggest that the effect of COLE onK562 cells is likely to be mediated by the JNK MAPKsrather than the p38 MAPKs JNK plays an important rolein apoptosis pathways weather by activating proapoptoticor antiapoptotic genes Particularly it was shown thatprolonged but not transient JNK activation promotes thecell death [60] Our results showed that JKAMP gene whichencodes for a membrane-anchored regulator of the durationof JNK1 activity was upregulated (154 plusmn 0051) The elevatedexpression of JKAMP as reported by Kadoya et al [61]results in sustained JNK activity These findings may indicatethe potential role of JNK pathway in the COLE-inducedapoptosis in K562 cells
Thanks to their large regulatory domains some MAP-KKKs can interact with upstream regulators have functionssuch as ubiquitylation and be activated by relief of autoin-hibition and oligomerization [62] In this context MAP3K2has been reported to play an important role in NF-kappaBsignaling pathway by activating the I-kappaB kinases whichin turn phosphorylate the inhibitory factors of the nuclearfactor kappaB (NF-kappaB) resulting in their rapid ubiq-uitination and the liberation of NF-kappaB complex whichtranslocate from the cytoplasm to the nucleus [63] MAP3K7in association with other proteins could also be required forthe activation of NF-kappaB According to the GO analysis(Table S1) COLE treatment upregulated genes implicatedin NF-kappaB-related categories such as positive regulationof NF-kappaB transcription factor activity I-kappaB kinase-NF-kappaB cascade and its positive regulation and I-kappaBphosphorylation Genes encoding the signal transducers inthe NF-kappaB pathway TRAF5 and TRAF6 as well asthe activator of NF-kappaB signaling SNIP1 were upregu-lated Interestingly an increased expression of NFKB1 geneencoding for the precursor of the mature NF-kappaB p50was detected in COLE treated cells compared to untreatedK562 cells NFKB is well recognized as a central activator ofthe antiapoptotic cascades in response to external stimuli orintrinsic immune reactions and its prosurvival activity hasbeen implicated in a variety of biological processes [60 64]In the B cell lineage the activity of NF-120581B is required forthe completion of various developmental stages includingdifferentiation and the response of these cells to antigens [65]NF-kappaB also plays an important role in the survival anddevelopment of T cells [66] and has been recently reported tomediate the differentiation of mesenchymal stem cells [67]
Several studies have demonstrated a crosstalk betweenthe NF-kappaB and JNK pathways seen as an inhibitoryeffect of NF-kappaB on apoptosis through the suppressionof JNK activity and that this suppressive effect may occur
14 Oxidative Medicine and Cellular Longevity
through different mechanisms [60] This suggests that thebalance between JNK and NF-kappaB activities is crucial todetermine the cell fate survival or deathWehypothesize herethat COLE-induced apoptosis or differentiation is likely to bedetermined by the crosstalk between these two pathways inK562 cells Further investigations regarding protein expres-sion and activity are needed to confirm these findings
5 Conclusions
Our study demonstrates for the first time that olive leafextract exhibits an antileukemia effect on the human chronicmyeloid leukemia cells Olive leaf extract was shown toinhibit the proliferation of K562 cells by inducing cell cyclearrest apoptosis and differentiation toward the monocytelineage The induced expression of molecules involved indifferentiation toward the monocytemacrophage lineage aswell asmolecules related to apoptosis and cell cycle regulationconfirmed these findings and provides insights into themechanism by which olive leaf exhibits its antileukemiaeffect
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported by the JICA-JST Science andTechnology Research Partnership for Sustainable Develop-ment (SATREPS) Project ldquoValorization of Bio-Resources inSemi-Arid and Arid Land for Regional Developmentrdquo
References
[1] M Leszczyniecka T Roberts P Dent S Grant and P B FisherldquoDifferentiation therapy of human cancer basic science andclinical applicationsrdquo Pharmacology and Therapeutics vol 90no 2-3 pp 105ndash156 2001
[2] A I Spira and M A Carducci ldquoDifferentiation therapyrdquoCurrent Opinion in Pharmacology vol 3 no 4 pp 338ndash3432003
[3] S Nobili D Lippi E Witort et al ldquoNatural compounds forcancer treatment and preventionrdquo Pharmacological Researchvol 59 no 6 pp 365ndash378 2009
[4] S Ramos ldquoEffects of dietary flavonoids on apoptotic pathwaysrelated to cancer chemopreventionrdquo Journal of NutritionalBiochemistry vol 18 no 7 pp 427ndash442 2007
[5] V Hajhashemi G Vaseghi M Pourfarzam and A AbdollahildquoAre antioxidants helpful for disease preventionrdquo Research inPharmaceutical Sciences vol 5 no 1 pp 1ndash8 2010
[6] M Bouaziz and S Sayadi ldquoIsolation and evaluation of antiox-idants from leaves of a Tunisian cultivar olive treerdquo EuropeanJournal of Lipid Science andTechnology vol 107 no 7-8 pp 497ndash504 2005
[7] A P Pereira I C F R Ferreira F Marcelino et al ldquoPhenoliccompounds and antimicrobial activity of olive (Olea europaeaL Cv Cobrancosa) leavesrdquo Molecules vol 12 no 5 pp 1153ndash1162 2007
[8] L I Somova F O Shode P Ramnanan and A Nadar ldquoAnti-hypertensive antiatherosclerotic and antioxidant activity oftriterpenoids isolated from Olea europaea subspecies africanaleavesrdquo Journal of Ethnopharmacology vol 84 no 2-3 pp 299ndash305 2003
[9] V Micol N Caturla L Perez-Fons V Mas L Perez andA Estepa ldquoThe olive leaf extract exhibits antiviral activityagainst viral haemorrhagic septicaemia rhabdovirus (VHSV)rdquoAntiviral Research vol 66 no 2-3 pp 129ndash136 2005
[10] J Wainstein T Ganz M Boaz et al ldquoOlive leaf extract as ahypoglycemic agent in both human diabetic subjects and inratsrdquo Journal of Medicinal Food vol 15 no 7 pp 605ndash610 2012
[11] L Seddik TM Bah A AouesM Slimani andM BenderdourldquoElucidation of mechanisms underlying the protective effects ofolive leaf extract against lead-induced neurotoxicity in Wistarratsrdquo Journal of Toxicological Sciences vol 36 no 6 pp 797ndash809 2011
[12] Z Bouallagui J Han H Isoda and S Sayadi ldquoHydroxytyrosolrich extract from olive leaves modulates cell cycle progressionin MCF-7 human breast cancer cellsrdquo Food and ChemicalToxicology vol 49 no 1 pp 179ndash184 2011
[13] R Fares S Bazzi S E Baydoun and R M Abdel-Massih ldquoTheantioxidant and anti-proliferative activity of the Lebanese Oleaeuropaea extractrdquo Plant Foods for Human Nutrition vol 66 no1 pp 58ndash63 2011
[14] V Goulas V Exarchou A N Troganis et al ldquoPhytochemicalsin olive-leaf extracts and their antiproliferative activity againstcancer and endothelial cellsrdquo Molecular Nutrition and FoodResearch vol 53 no 5 pp 600ndash608 2009
[15] M de Bock E B Thorstensen J G Derraik H V HendersonP L Hofman and W S Cutfield ldquoHuman absorption andmetabolism of oleuropein and hydroxytyrosol ingested as olive(Olea europaea L) leaf extractrdquo Molecular Nutrition and FoodResearch vol 57 no 11 pp 2079ndash2085 2013
[16] R Briante M Patumi S Terenziani E Bismuto F Febbraioand R Nucci ldquoOlea europaea L leaf extract and derivativesantioxidant propertiesrdquo Journal of Agricultural and Food Chem-istry vol 50 no 17 pp 4934ndash4940 2002
[17] O Benavente-Garcıa J Castillo J Lorente A Ortuno and J ADel Rio ldquoAntioxidant activity of phenolics extracted from Oleaeuropaea L leavesrdquo Food Chemistry vol 68 no 4 pp 457ndash4622000
[18] S H Omar ldquoOleuropein in olive and its pharmacologicaleffectsrdquo Scientia Pharmaceutica vol 78 no 2 pp 133ndash154 2010
[19] H Jemai M Bouaziz I Fki A El Feki and S SayadildquoHypolipidimic and antioxidant activities of oleuropein and itshydrolysis derivative-rich extracts from Chemlali olive leavesrdquoChemico-Biological Interactions vol 176 no 2-3 pp 88ndash982008
[20] R Garcıa-Villalba M Larrosa S Possemiers F A Tomas-Barberan and J C Espın ldquoBioavailability of phenolics from anoleuropein-rich olive (Olea europaea) leaf extract and its acuteeffect on plasma antioxidant status comparison between pre-and postmenopausal womenrdquo European Journal of Nutrition2013
[21] J Anter Z Fernandez-Bedmar M Villatoro-Pulido et al ldquoApilot study on the DNA-protective cytotoxic and apoptosis-inducing properties of olive-leaf extractsrdquo Mutation Researchvol 723 no 2 pp 165ndash170 2011
[22] J A Sutherland A R Turner and P Mannoni ldquoDifferen-tiation of K562 leukemia cells along erythroid macrophage
Oxidative Medicine and Cellular Longevity 15
and megakaryocyte lineagesrdquo Journal of Biological ResponseModifiers vol 5 no 3 pp 250ndash262 1986
[23] K Schwenke H-P Peterson K-H Wangenheim and L EFeinendegen ldquoInduction of differentiation in erythroleukemicK562 cells by 120574-irradiationrdquo Leukemia Research vol 19 no 12pp 955ndash961 1995
[24] V N Enujiugha J Y Talabi S A Malomo and A I OlagunjuldquoDPPH radical scavenging capacity of phenolic extracts fromAfrican Yam Bean (Sphenostylis stenocarpa)rdquo Food and Nutri-tion Sciences vol 3 pp 7ndash13 2012
[25] S A Mijatovic G S Timotijevic D M Miljkovic et alldquoMultiple antimelanoma potential of dry olive leaf extractrdquoInternational Journal of Cancer vol 128 no 8 pp 1955ndash19652011
[26] L Abaza T P N Talorete P Yamada Y KuritaM Zarrouk andH Isoda ldquoInduction of growth inhibition and differentiation ofhuman leukemia HL-60 cells by a Tunisian Gerboui olive leafextractrdquo Bioscience Biotechnology and Biochemistry vol 71 no5 pp 1306ndash1312 2007
[27] B Tunca G Tezcan G Cecener et al ldquoOlea europea leafextract alters microRNA expression in human glioblastomacellsrdquo Journal of Cancer Research and Clinical Oncology vol 138no 11 pp 1831ndash1844 2012
[28] S Matsuoka M Huang and S J Elledge ldquoLinkage of ATM tocell cycle regulation by the Chk2 protein kinaserdquo Science vol282 no 5395 pp 1893ndash1897 1998
[29] C Frazer and P G Young ldquoPhosphorylation mediated regu-lation of Cdc25 activity localization and stabilityrdquo in ProteinPhosphorylation in Human Health C Huang Ed pp 395ndash436InTech 2012
[30] N Debili C Robin V Schiavon et al ldquoDifferent expression ofCD41 on human lymphoid andmyeloid progenitors from adultsand neonatesrdquo Blood vol 97 no 7 pp 2023ndash2030 2001
[31] H Nakajima ldquoRole of transcription factors in differentiationand reprogramming of hematopoietic cellsrdquo Keio Journal ofMedicine vol 60 no 2 pp 47ndash55 2011
[32] Y Fukuchi F Shibata M Ito et al ldquoComprehensive analysis ofmyeloid lineage conversion using mice expressing an inducibleform of CEBP120572rdquoThe EMBO Journal vol 25 no 14 pp 3398ndash3410 2006
[33] R A Shivdasani ldquoMolecular and transcriptional regulation ofmegakaryocyte differentiationrdquo Stem Cells vol 19 no 5 pp397ndash407 2001
[34] K Terui Y Takahashi J Kitazawa T TokiM Yokoyama and EIto ldquoExpression of transcription factors during megakaryocyticdifferentiation of CD34+ cells from human cord blood inducedby thrombopoietinrdquo Tohoku Journal of Experimental Medicinevol 192 no 4 pp 259ndash273 2000
[35] J A Trapani M Dawson V A Apostolidis and K A BrowneldquoGenomic organization of IFI16 an interferon-inducible genewhose expression is associated with human myeloid cell differ-entiation correlation of predicted protein domains with exonorganizationrdquo Immunogenetics vol 40 no 6 pp 415ndash424 1994
[36] M J Dawson N J Elwood R W Johnstone and J A TrapanildquoThe IFN-inducible nucleoprotein IFI 16 is expressed in cellsof the monocyte lineage but is rapidly and markedly down-regulated in other myeloid precursor populationsrdquo Journal ofLeukocyte Biology vol 64 no 4 pp 546ndash554 1998
[37] M Gariglio M De Andrea M Lembo et al ldquoThe murinehomolog of the HIN 200 family Ifi 204 is constitutivelyexpressed in myeloid cells and selectively induced in the
monocytemacrophage lineagerdquo Journal of Leukocyte Biologyvol 64 no 5 pp 608ndash614 1998
[38] A F Valledor F E Borras M Cullell-Young and A CeladaldquoTranscription factors that regulate monocytemacrophage dif-ferentiationrdquo Journal of Leukocyte Biology vol 63 no 4 pp405ndash417 1998
[39] K Krishnaraju B Hoffman and D A Liebermann ldquoEarlygrowth response gene 1 stimulates development of hematopoi-etic progenitor cells along the macrophage lineage at theexpense of the granulocyte and erythroid lineagesrdquo Blood vol97 no 5 pp 1298ndash1305 2001
[40] J Dauffy G Mouchiroud and R P Bourette ldquoThe interferon-inducible gene Ifi204 is transcriptionally activated in responseto M-CSF and its expression favors macrophage differentiationin myeloid progenitor cellsrdquo Journal of Leukocyte Biology vol79 no 1 pp 173ndash183 2006
[41] G Marziali E Perrotti R Ilari U Testa E M Coccia andA Battistini ldquoTranscriptional regulation of the ferritin heavy-chain gene the activity of the CCAAT binding factor NF-Y ismodulated in heme-treated Friend leukemia cells and duringmonocyte-to-macrophage differentiationrdquo Molecular and Cel-lular Biology vol 17 no 3 pp 1387ndash1395 1997
[42] C Shi X Zhang Z Chen et al ldquoIntegrin engagement regulatesmonocyte differentiation through the forkhead transcriptionfactor Foxp1rdquo Journal of Clinical Investigation vol 114 no 3 pp408ndash418 2004
[43] C Shi M Sakuma T Mooroka et al ldquoDown-regulation of theforkhead transcription factor Foxp1 is required for monocytedifferentiation andmacrophage functionrdquoBlood vol 112 no 12pp 4699ndash4711 2008
[44] D F Smith E Galkina K Ley and Y Huo ldquoGRO familychemokines are specialized for monocyte arrest from flowrdquoThe American Journal of Physiology Heart and CirculatoryPhysiology vol 289 no 5 pp H1976ndashH1984 2005
[45] J J Oppenheim O M Z Howard and E Goetzl ChemotacticFactors Neuropeptides and Other Ligands for Seven Transmem-brane Receptors 2000
[46] Y Terashima N Onai M Murai et al ldquoPivotal function forcytoplasmic protein FROUNT in CCR2-mediated monocytechemotaxisrdquo Nature Immunology vol 6 no 8 pp 827ndash8352005
[47] Y Lefkir M Malbouyres D Gotthardt et al ldquoInvolvement ofthe AP-1 adaptor complex in early steps of phagocytosis andmacropinocytosisrdquo Molecular Biology of the Cell vol 15 no 2pp 861ndash869 2004
[48] F Niedergang and P Chavrier ldquoSignaling and membranedynamics during phagocytosis many roads lead to the phago-somerdquo Current Opinion in Cell Biology vol 16 no 4 pp 422ndash428 2004
[49] D Cox D J Lee B M Dale J Calafat and S Green-berg ldquoA Rab11-containing rapidly recycling compartment inmacrophages that promotes phagocytosisrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 97 no 2 pp 680ndash685 2000
[50] O V Vieira C Bucci R E Harrison et al ldquoModulation of Rab5and Rab7 recruitment to phagosomes by phosphatidylinositol3-kinaserdquoMolecular andCellular Biology vol 23 no 7 pp 2501ndash2514 2003
[51] T Khurana J A Brzostowski and A R Kimmel ldquoARab21LIM-onlyCH-LIM complex regulates phagocytosis viaboth activating and inhibitory mechanismsrdquo The EMBO Jour-nal vol 24 no 13 pp 2254ndash2264 2005
16 Oxidative Medicine and Cellular Longevity
[52] S J Martin J G Bradley and T G Cotter ldquoHL-60 cellsinduced to differentiate towards neutrophils subsequently dievia apoptosisrdquo Clinical and Experimental Immunology vol 79no 3 pp 448ndash453 1990
[53] A Benito D Grillot G Nunez and J L Fernandez-Luna ldquoReg-ulation and function of Bcl-2 during differentiation-inducedcell death in HL-60 promyelocytic cellsrdquoThe American Journalof Pathology vol 146 no 2 pp 481ndash490 1995
[54] M Lamkanfi N Festjens W Declercq T V Berghe andP Vandenabeele ldquoCaspases in cell survival proliferation anddifferentiationrdquo Cell Death and Differentiation vol 14 no 1 pp44ndash55 2007
[55] O Sordet C Rebe S Plenchette et al ldquoSpecific involvement ofcaspases in the differentiation of monocytes into macrophagesrdquoBlood vol 100 no 13 pp 4446ndash4453 2002
[56] T-B Kang T Ben-Moshe E E Varfolomeev et al ldquoCaspase-8 serves both apoptotic and nonapoptotic rolesrdquo Journal ofImmunology vol 173 no 5 pp 2976ndash2984 2004
[57] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[58] J Cheng J Yang Y Xia M Karin and B Su ldquoSynergisticinteraction of MEK kinase 2 c-Jun N-terminal kinase (JNK)kinase 2 and JNK1 results in efficient and specific JNK1activationrdquo Molecular and Cellular Biology vol 20 no 7 pp2334ndash2342 2000
[59] J L Blank P Gerwins EM Elliott S Sather andG L JohnsonldquoMolecular cloning of mitogen-activated proteinERK kinasekinases (MEKK) 2 and 3 regulation of sequential phosphoryla-tion pathways involving mitogen-activated protein kinase andc-Jun kinaserdquo Journal of Biological Chemistry vol 271 no 10pp 5361ndash5368 1996
[60] S Papa F Zazzeroni C G Pham C Bubici and G FranzosoldquoLinking JNK signaling to NF-120581B a key to survivalrdquo Journal ofCell Science vol 117 no 22 pp 5197ndash5208 2004
[61] T Kadoya A Khurana M Tcherpakov et al ldquoJAMP a JUB N-terminal kinase 1 (JNK1)-associated membrane protein regu-lates duration of JNK activityrdquo Molecular and Cellular Biologyvol 25 no 19 pp 8619ndash8630 2005
[62] M Qi and E A Elion ldquoMAP kinase pathwaysrdquo Journal of CellScience vol 118 no 16 pp 3569ndash3572 2005
[63] Q Zhao and F S Lee ldquoMitogen-activated protein kinaseERKkinase kinases 2 and 3 activate nuclear factor-120581B through I120581Bkinase-120572 and I120581B kinase-120573rdquoThe Journal of Biological Chemistryvol 274 no 13 pp 8355ndash8358 1999
[64] H Namba V Saenko and S Yamashita ldquoNuclear factor-120581Bin thyroid carcinogenesis and progression a novel therapeutictarget for advanced thyroid cancerrdquo Arquivos Brasileiros deEndocrinologia e Metabologia vol 51 no 5 pp 843ndash851 2007
[65] B Feng S Cheng W S Pear and H-C Liou ldquoNF-kBinhibitor blocks B cell development at two checkpointsrdquoMedi-cal Immunology vol 3 article 1 2004
[66] S Gerondakis and U Siebenlist ldquoRoles of the NF-kappaBpathway in lymphocyte development and functionrdquoCold SpringHarbor Perspectives in Biology vol 2 no 5 Article ID a0001822010
[67] K P Bhat V Balasubramaniyan B Vaillant et al ldquoMesenchy-mal differentiation mediated by NF-kB promotes radiationresistance in glioblastomardquo Cancer Cell vol 24 no 3 pp 331ndash346 2013
Figure 5 Expression of differentiationmarkers onK562 cells treatedwith Chemlali Olive Leaf Extract (COLE) up to 6 days Cells were treatedwith 50 100 and 150 120583gmL of COLE and incubated for different periods Control cells were treated with ethanol at a final concentration of03 Expression of cell surface markers was detected by flow cytometry (a) Expression of CD14 (monocytemacrophage specific marker)(b) Expression of CD11b (granulocytemonocyte specific marker) (c) Expression of CD41 (megakaryocyte specific marker) (d) Expressionof glycophorin A (GPA) (erythrocyte specific marker) Results are represented as the mean plusmn SD of three independent experiments lowast lowastlowastsignificantly different from the control at 119875 lt 005 and 119875 lt 001 respectively
including GYPA HBE1 FECH and ALAS2 which is consis-tent with the decreased expression of erythrocyte differenti-ation marker in the flow cytometry results (Figure 5(d))
Analysis of the microarray data showed the upregu-lation of several proapoptotic genes and genes involvedin the regulation of mitochondrial membrane permeabilitysuch as CASP6 CASP8 DFFA and BID On the otherhandexpression of genes negatively regulating the apoptosis
and those inhibiting the caspase activity was significantlydecreased such as IGF1R HSPA5 and BCL2
Among the upregulated genes we found those related tothe NF-kappaB cascade as well as to the MAPKKK cascadeand the Wnt receptor signaling pathway
Categories related to cell cycle regulation and cell pro-liferation were represented by both upregulated and down-regulated genes Finally some transcription factors werealso downregulated such as FOXP1 whereas others wereupregulated like EGR1 and NFYA
The list of selected genes is presented in Table 3 Fromthis list it is clear that the differentially expressed genes aftertreatment with 100 and 150120583gmL have mostly the sametendency which could validate the obtained results
12 Oxidative Medicine and Cellular Longevity
4 Discussion
Previous reports had indicated that olive leaf extract inducesapoptosis in the human leukemic cell lines HL-60 and Jurkatcells [13 21] In this study we give evidence that COLEexhibits its antileukemia effect by both inducing apoptosisand promoting differentiation of the multipotent humanleukemia K562 cells
Induction of differentiation as well as apoptosis has beenfrequently reported to be associatedwith a loss of proliferativecapacity of the cells It has been reported that olive leaf extractinhibits the proliferation of some cancer cell lines such asB16 melanoma cells HL-60 leukemia cells the breast cancerMCF-7 cells and the glioblastoma cells [12 25ndash27] Ourresults indicate that COLE inhibits the proliferation of K562cells in a dose dependent manner (Figure 2(a)) Howeverdespite the drastic decrease in cell proliferation the viabilityof cells treated with the highest concentration of COLE(150 120583gmL) remains relatively high (80) for up to 3 daysof treatment (Figure 3(b))
Since it is well established that the arrest of the cellcycle progression allows the cells to follow other processessuch as apoptosis and differentiation we then explored theeffect of COLE on the cell cycle distribution (Table 2) Oliveleaf extract has been shown to cause cell cycle arrest atG0G1 phase in both the breast cancer MCF-7 cells and themelanoma B16 cells [12 25] However the treatment timesapplied in both of these studies were limited to 48 h and 18 hrespectively In this study COLE treated cells were arrested atG0G1 on the 1st and 2ndday of treatment which is consistentwith the reported studies Prolonged incubation with COLEshowed an arrest of cell cycle at G2M phase (3rd and 4thday of incubation) The microarray analysis performed forthe 3rd day of treatment revealed a significant increase ofCHEK2 gene expression in COLE treated cells The encodedprotein regulates the cell cycle checkpoint arrest throughthe inhibition of the activity of CDC25A CDC25B andCDC25C [28] CDC25 proteins trigger the entry into mitosisat different points of the cell cycle by activating theCdk-cyclincomplexes CDC25A acts early in the cell cycle regulatingthe G1S transition whereas CDC25B and CDC25C act atG2M [29] Interestingly our results showed a decrease in theexpression of CDC25C gene in COLE treated cells associatedwith an increase in the expression of CDC25A which mayexplain the cell cycle arrest at G2M phase observed at the3rd day of treatment with COLE
Treatment with COLE increased the expression of CD14on the cell surface of treated cells indicating the differenti-ation of K562 cells toward the mono-macrophage lineage(Figure 5(a)) The increase in the percentage of cells positivefor CD11b (Figure 5(b)) confirms this hypothesis In factCD11b is expressed on both monocytes and granulocytesThus together with the increase of CD14 expression theexpression of CD11b suggests that treatment with COLEpromotes the commitment of K562 cells into the mono-cytemacrophage lineage at the expense of the granulocyticdifferentiationThe instant increase of the expression of CD41
on the 1st day of treatment and its absence during the fol-lowing days (Figure 5(c)) could be consistent with monocytedifferentiation instead of megakaryocyte differentiation sinceit has been reported that 120572IIb promoter which initiates thetranscription of CD41 is transcriptionally active in pluripo-tent myeloid progenitors in early stages of erythropoiesisand all along the megakaryocytic differentiation as well asto a lesser extent in the early stages of myelomonocyticdifferentiation and the late stages of erythropoiesis [30] Thisinstant increase could be also explained by the phenomenonof lineage conversion where some hematopoietic progenitorscan be converted into lineages other than their own bythe ectopic expression of some transcription factors [31]In this regard it was previously reported that commonlymphoid progenitors megakaryocyte erythroid progenitorsearly B cells and early T cells could be converted to thegranulocytemonocyte lineage by the induction of CEBP120572activity [32] Microarray results support the absence ofmegakaryocyte differentiation in the 3rd day of treatment Infact data showed a decrease in the expression of genes relatedtomegakaryopoiesis such as the essential regulator of plateletrelease NFE2 and its main candidate target gene TUBB1(encoding the megakaryocyte and platelet specific isoform of120573-tubulin (1205731)) [33] as well as the BACH2 transcription factorreported to be induced during megakaryocyte differentiation[34]
Interestingly results of microarray analysis (Table 3)showed the upregulation of several genes already reportedas key players in monocytemacrophage differentiation inthe COLE treated cells Among these genes the expressionof IFI16 was markedly increased Constitutively expressedin lymphoid cells IFI16 was shown to be associated withthe differentiation of human myeloid cells in response tointerferon gamma [35] Moreover IFI16 has been selectivelyexpressed during the differentiation of myeloid progenitorcells along monocytic lineage while it is downregulatedduring the commitment toward the granulocytic or erythroidlineage suggesting its potential involvement in the differenti-ation andmaturation of the monocytic lineage [36 37] Geneencoding for EGR-1 a positive modulator of macrophagedifferentiation was also significantly upregulated EGR-1 isknown to dictate development of myeloid progenitors alongthe macrophage lineage at the expense of development alongother lineages [38 39] Dauffy et al [40] reported thatIFI16-enforced expression in myeloid progenitors inducedthe expression of EGR-1 and conducted the macrophagedifferentiation in the absence of the macrophage colonystimulating factor (M-CSF) This suggests that EGR-1 couldbe regulated by the expression of IFI16 Treatment with COLEincreased as well the expression of NFYA encoding the Asubunit of the nuclear transcription factor Y whose synthesiswas previously reported during the terminal differentiationof monocytes to macrophages [38 41] The downregula-tion of FOXP1 in COLE treated K562 cells is consistentwith monocytemacrophage differentiation since it has beendemonstrated that the expression of FOXP1 was markedlydecreased in monocyte-induced differentiation of HL-60cells as well as in human peripheral blood monocytes and
Oxidative Medicine and Cellular Longevity 13
that the overexpression of FOXP1 prevented the morphologicmacrophage-like differentiation [42 43]
Monocytes and macrophages have several functionalcharacteristics including cell adhesion migration chemo-taxis and phagocytic activity Interestingly an enhancedexpression of the chemokines genes CXCL8 (IL8) CXCL2andCXCL3was found in COLE treated cells IL-8 productionhas been observed in vitro in a wide variety of cells includingmonocytes T lymphocytes neutrophils vascular endothe-lial cells dermal fibroblasts keratinocytes hepatocytes andhuman gastric cancer cells while the production of CXCL2andCXCL3was only described inmonocytes fibroblasts andendothelial cells [44 45] Another gene related to chemotaxiswas upregulated NUP85 which is an essential component ofthe nuclear pore complex and was reported to be involved inCCR2-mediated chemotaxis of monocytes [46]
COLE treatment also enhanced the expression of somegenes related to phagocytosis AP1G1 andRab proteins relatedgenes AP1G1 is a subunit of clathrin-associated adaptorprotein complex 1 whose role has been recently highlightedfor efficient phagocytosis at an early stage of phagosomeformation since it participates in the extension of the phago-cytic cup [47 48] The small GTPases Rab are key regulatorsof intracellular membrane trafficking Recent studies havedemonstrated that several RAB proteins play an importantrole in phagocytosis RAB proteins such as RAB11 Rab5 andRab 21 participate in the phagosome formation and matura-tion and are necessary for the phagocytic activity [48ndash51]
Consistently with the increase in Annexin V positivecells (Figure 4) treatment with COLE upregulated someproapoptotic genes such as CASP6 CASP8 DFFA and BIDand downregulated the apoptosis suppressor BCL2 as wellas the caspase inhibitors genes (Table 3) Olive leaf waspreviously shown to induce morphological changes thatare characteristic of apoptosis in leukemia cells but themechanisms underlying this effect were not well investi-gated [13 21] The increase in the proportion of apoptoticcells observed from the 4th day of treatment with COLEcould be explained by the programmed death of the fullydifferentiated cells (Figure 4) In fact this increase wasaccompanied by an important decrease in cell viability aswell as a diminution in CD14 expression (Figures 3(b) and5(a)) Elsewhere it is well established that in hematopoietictissues apoptosis is coupled to terminal differentiation ofmyeloid progenitors even though themechanism responsiblefor the activation of apoptosis during myeloid maturationis still poorly understood [52 53] It is also noteworthy tomention that several reports have highlighted the role ofcaspases in the terminal differentiation of a variety of celltypes [54] One example is the differentiation of human bloodmonocytes into macrophages a process that is blocked bysynthetic caspase inhibitors [55] It has been also reportedthat caspase 8 deletion in bone-marrow cells resulted in arrestof hemopoietic progenitor functioning and that its deletionin cells of the myelomonocytic lineage led to the arrest ofdifferentiation into macrophages and consequently to celldeath [56]
Among the differentially expressed genes in COLEtreated cells (Table 3) genes encoding the mitogen-activated
protein kinase kinase kinases (MAPKKKs) MAP3K2MAP3K5 and MAP3K7 were highly expressed whileMAP2K5 and MAPK14p38-120572 were downregulated TheMAPKKKs act at the upstream of the MAP kinase cascadewhich regulates important cellular processes such as geneexpression cell proliferation differentiation cell survivaland death The MAP3K5 activates the c-Jun N-terminalkinase (JNK) and p38 mitogen-activated protein kinases[57] while the MAP3K2 is reported to preferentially activateJNK [58 59] However the significant decrease in theMAPK14p38-120572 may suggest that the effect of COLE onK562 cells is likely to be mediated by the JNK MAPKsrather than the p38 MAPKs JNK plays an important rolein apoptosis pathways weather by activating proapoptoticor antiapoptotic genes Particularly it was shown thatprolonged but not transient JNK activation promotes thecell death [60] Our results showed that JKAMP gene whichencodes for a membrane-anchored regulator of the durationof JNK1 activity was upregulated (154 plusmn 0051) The elevatedexpression of JKAMP as reported by Kadoya et al [61]results in sustained JNK activity These findings may indicatethe potential role of JNK pathway in the COLE-inducedapoptosis in K562 cells
Thanks to their large regulatory domains some MAP-KKKs can interact with upstream regulators have functionssuch as ubiquitylation and be activated by relief of autoin-hibition and oligomerization [62] In this context MAP3K2has been reported to play an important role in NF-kappaBsignaling pathway by activating the I-kappaB kinases whichin turn phosphorylate the inhibitory factors of the nuclearfactor kappaB (NF-kappaB) resulting in their rapid ubiq-uitination and the liberation of NF-kappaB complex whichtranslocate from the cytoplasm to the nucleus [63] MAP3K7in association with other proteins could also be required forthe activation of NF-kappaB According to the GO analysis(Table S1) COLE treatment upregulated genes implicatedin NF-kappaB-related categories such as positive regulationof NF-kappaB transcription factor activity I-kappaB kinase-NF-kappaB cascade and its positive regulation and I-kappaBphosphorylation Genes encoding the signal transducers inthe NF-kappaB pathway TRAF5 and TRAF6 as well asthe activator of NF-kappaB signaling SNIP1 were upregu-lated Interestingly an increased expression of NFKB1 geneencoding for the precursor of the mature NF-kappaB p50was detected in COLE treated cells compared to untreatedK562 cells NFKB is well recognized as a central activator ofthe antiapoptotic cascades in response to external stimuli orintrinsic immune reactions and its prosurvival activity hasbeen implicated in a variety of biological processes [60 64]In the B cell lineage the activity of NF-120581B is required forthe completion of various developmental stages includingdifferentiation and the response of these cells to antigens [65]NF-kappaB also plays an important role in the survival anddevelopment of T cells [66] and has been recently reported tomediate the differentiation of mesenchymal stem cells [67]
Several studies have demonstrated a crosstalk betweenthe NF-kappaB and JNK pathways seen as an inhibitoryeffect of NF-kappaB on apoptosis through the suppressionof JNK activity and that this suppressive effect may occur
14 Oxidative Medicine and Cellular Longevity
through different mechanisms [60] This suggests that thebalance between JNK and NF-kappaB activities is crucial todetermine the cell fate survival or deathWehypothesize herethat COLE-induced apoptosis or differentiation is likely to bedetermined by the crosstalk between these two pathways inK562 cells Further investigations regarding protein expres-sion and activity are needed to confirm these findings
5 Conclusions
Our study demonstrates for the first time that olive leafextract exhibits an antileukemia effect on the human chronicmyeloid leukemia cells Olive leaf extract was shown toinhibit the proliferation of K562 cells by inducing cell cyclearrest apoptosis and differentiation toward the monocytelineage The induced expression of molecules involved indifferentiation toward the monocytemacrophage lineage aswell asmolecules related to apoptosis and cell cycle regulationconfirmed these findings and provides insights into themechanism by which olive leaf exhibits its antileukemiaeffect
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported by the JICA-JST Science andTechnology Research Partnership for Sustainable Develop-ment (SATREPS) Project ldquoValorization of Bio-Resources inSemi-Arid and Arid Land for Regional Developmentrdquo
References
[1] M Leszczyniecka T Roberts P Dent S Grant and P B FisherldquoDifferentiation therapy of human cancer basic science andclinical applicationsrdquo Pharmacology and Therapeutics vol 90no 2-3 pp 105ndash156 2001
[2] A I Spira and M A Carducci ldquoDifferentiation therapyrdquoCurrent Opinion in Pharmacology vol 3 no 4 pp 338ndash3432003
[3] S Nobili D Lippi E Witort et al ldquoNatural compounds forcancer treatment and preventionrdquo Pharmacological Researchvol 59 no 6 pp 365ndash378 2009
[4] S Ramos ldquoEffects of dietary flavonoids on apoptotic pathwaysrelated to cancer chemopreventionrdquo Journal of NutritionalBiochemistry vol 18 no 7 pp 427ndash442 2007
[5] V Hajhashemi G Vaseghi M Pourfarzam and A AbdollahildquoAre antioxidants helpful for disease preventionrdquo Research inPharmaceutical Sciences vol 5 no 1 pp 1ndash8 2010
[6] M Bouaziz and S Sayadi ldquoIsolation and evaluation of antiox-idants from leaves of a Tunisian cultivar olive treerdquo EuropeanJournal of Lipid Science andTechnology vol 107 no 7-8 pp 497ndash504 2005
[7] A P Pereira I C F R Ferreira F Marcelino et al ldquoPhenoliccompounds and antimicrobial activity of olive (Olea europaeaL Cv Cobrancosa) leavesrdquo Molecules vol 12 no 5 pp 1153ndash1162 2007
[8] L I Somova F O Shode P Ramnanan and A Nadar ldquoAnti-hypertensive antiatherosclerotic and antioxidant activity oftriterpenoids isolated from Olea europaea subspecies africanaleavesrdquo Journal of Ethnopharmacology vol 84 no 2-3 pp 299ndash305 2003
[9] V Micol N Caturla L Perez-Fons V Mas L Perez andA Estepa ldquoThe olive leaf extract exhibits antiviral activityagainst viral haemorrhagic septicaemia rhabdovirus (VHSV)rdquoAntiviral Research vol 66 no 2-3 pp 129ndash136 2005
[10] J Wainstein T Ganz M Boaz et al ldquoOlive leaf extract as ahypoglycemic agent in both human diabetic subjects and inratsrdquo Journal of Medicinal Food vol 15 no 7 pp 605ndash610 2012
[11] L Seddik TM Bah A AouesM Slimani andM BenderdourldquoElucidation of mechanisms underlying the protective effects ofolive leaf extract against lead-induced neurotoxicity in Wistarratsrdquo Journal of Toxicological Sciences vol 36 no 6 pp 797ndash809 2011
[12] Z Bouallagui J Han H Isoda and S Sayadi ldquoHydroxytyrosolrich extract from olive leaves modulates cell cycle progressionin MCF-7 human breast cancer cellsrdquo Food and ChemicalToxicology vol 49 no 1 pp 179ndash184 2011
[13] R Fares S Bazzi S E Baydoun and R M Abdel-Massih ldquoTheantioxidant and anti-proliferative activity of the Lebanese Oleaeuropaea extractrdquo Plant Foods for Human Nutrition vol 66 no1 pp 58ndash63 2011
[14] V Goulas V Exarchou A N Troganis et al ldquoPhytochemicalsin olive-leaf extracts and their antiproliferative activity againstcancer and endothelial cellsrdquo Molecular Nutrition and FoodResearch vol 53 no 5 pp 600ndash608 2009
[15] M de Bock E B Thorstensen J G Derraik H V HendersonP L Hofman and W S Cutfield ldquoHuman absorption andmetabolism of oleuropein and hydroxytyrosol ingested as olive(Olea europaea L) leaf extractrdquo Molecular Nutrition and FoodResearch vol 57 no 11 pp 2079ndash2085 2013
[16] R Briante M Patumi S Terenziani E Bismuto F Febbraioand R Nucci ldquoOlea europaea L leaf extract and derivativesantioxidant propertiesrdquo Journal of Agricultural and Food Chem-istry vol 50 no 17 pp 4934ndash4940 2002
[17] O Benavente-Garcıa J Castillo J Lorente A Ortuno and J ADel Rio ldquoAntioxidant activity of phenolics extracted from Oleaeuropaea L leavesrdquo Food Chemistry vol 68 no 4 pp 457ndash4622000
[18] S H Omar ldquoOleuropein in olive and its pharmacologicaleffectsrdquo Scientia Pharmaceutica vol 78 no 2 pp 133ndash154 2010
[19] H Jemai M Bouaziz I Fki A El Feki and S SayadildquoHypolipidimic and antioxidant activities of oleuropein and itshydrolysis derivative-rich extracts from Chemlali olive leavesrdquoChemico-Biological Interactions vol 176 no 2-3 pp 88ndash982008
[20] R Garcıa-Villalba M Larrosa S Possemiers F A Tomas-Barberan and J C Espın ldquoBioavailability of phenolics from anoleuropein-rich olive (Olea europaea) leaf extract and its acuteeffect on plasma antioxidant status comparison between pre-and postmenopausal womenrdquo European Journal of Nutrition2013
[21] J Anter Z Fernandez-Bedmar M Villatoro-Pulido et al ldquoApilot study on the DNA-protective cytotoxic and apoptosis-inducing properties of olive-leaf extractsrdquo Mutation Researchvol 723 no 2 pp 165ndash170 2011
[22] J A Sutherland A R Turner and P Mannoni ldquoDifferen-tiation of K562 leukemia cells along erythroid macrophage
Oxidative Medicine and Cellular Longevity 15
and megakaryocyte lineagesrdquo Journal of Biological ResponseModifiers vol 5 no 3 pp 250ndash262 1986
[23] K Schwenke H-P Peterson K-H Wangenheim and L EFeinendegen ldquoInduction of differentiation in erythroleukemicK562 cells by 120574-irradiationrdquo Leukemia Research vol 19 no 12pp 955ndash961 1995
[24] V N Enujiugha J Y Talabi S A Malomo and A I OlagunjuldquoDPPH radical scavenging capacity of phenolic extracts fromAfrican Yam Bean (Sphenostylis stenocarpa)rdquo Food and Nutri-tion Sciences vol 3 pp 7ndash13 2012
[25] S A Mijatovic G S Timotijevic D M Miljkovic et alldquoMultiple antimelanoma potential of dry olive leaf extractrdquoInternational Journal of Cancer vol 128 no 8 pp 1955ndash19652011
[26] L Abaza T P N Talorete P Yamada Y KuritaM Zarrouk andH Isoda ldquoInduction of growth inhibition and differentiation ofhuman leukemia HL-60 cells by a Tunisian Gerboui olive leafextractrdquo Bioscience Biotechnology and Biochemistry vol 71 no5 pp 1306ndash1312 2007
[27] B Tunca G Tezcan G Cecener et al ldquoOlea europea leafextract alters microRNA expression in human glioblastomacellsrdquo Journal of Cancer Research and Clinical Oncology vol 138no 11 pp 1831ndash1844 2012
[28] S Matsuoka M Huang and S J Elledge ldquoLinkage of ATM tocell cycle regulation by the Chk2 protein kinaserdquo Science vol282 no 5395 pp 1893ndash1897 1998
[29] C Frazer and P G Young ldquoPhosphorylation mediated regu-lation of Cdc25 activity localization and stabilityrdquo in ProteinPhosphorylation in Human Health C Huang Ed pp 395ndash436InTech 2012
[30] N Debili C Robin V Schiavon et al ldquoDifferent expression ofCD41 on human lymphoid andmyeloid progenitors from adultsand neonatesrdquo Blood vol 97 no 7 pp 2023ndash2030 2001
[31] H Nakajima ldquoRole of transcription factors in differentiationand reprogramming of hematopoietic cellsrdquo Keio Journal ofMedicine vol 60 no 2 pp 47ndash55 2011
[32] Y Fukuchi F Shibata M Ito et al ldquoComprehensive analysis ofmyeloid lineage conversion using mice expressing an inducibleform of CEBP120572rdquoThe EMBO Journal vol 25 no 14 pp 3398ndash3410 2006
[33] R A Shivdasani ldquoMolecular and transcriptional regulation ofmegakaryocyte differentiationrdquo Stem Cells vol 19 no 5 pp397ndash407 2001
[34] K Terui Y Takahashi J Kitazawa T TokiM Yokoyama and EIto ldquoExpression of transcription factors during megakaryocyticdifferentiation of CD34+ cells from human cord blood inducedby thrombopoietinrdquo Tohoku Journal of Experimental Medicinevol 192 no 4 pp 259ndash273 2000
[35] J A Trapani M Dawson V A Apostolidis and K A BrowneldquoGenomic organization of IFI16 an interferon-inducible genewhose expression is associated with human myeloid cell differ-entiation correlation of predicted protein domains with exonorganizationrdquo Immunogenetics vol 40 no 6 pp 415ndash424 1994
[36] M J Dawson N J Elwood R W Johnstone and J A TrapanildquoThe IFN-inducible nucleoprotein IFI 16 is expressed in cellsof the monocyte lineage but is rapidly and markedly down-regulated in other myeloid precursor populationsrdquo Journal ofLeukocyte Biology vol 64 no 4 pp 546ndash554 1998
[37] M Gariglio M De Andrea M Lembo et al ldquoThe murinehomolog of the HIN 200 family Ifi 204 is constitutivelyexpressed in myeloid cells and selectively induced in the
monocytemacrophage lineagerdquo Journal of Leukocyte Biologyvol 64 no 5 pp 608ndash614 1998
[38] A F Valledor F E Borras M Cullell-Young and A CeladaldquoTranscription factors that regulate monocytemacrophage dif-ferentiationrdquo Journal of Leukocyte Biology vol 63 no 4 pp405ndash417 1998
[39] K Krishnaraju B Hoffman and D A Liebermann ldquoEarlygrowth response gene 1 stimulates development of hematopoi-etic progenitor cells along the macrophage lineage at theexpense of the granulocyte and erythroid lineagesrdquo Blood vol97 no 5 pp 1298ndash1305 2001
[40] J Dauffy G Mouchiroud and R P Bourette ldquoThe interferon-inducible gene Ifi204 is transcriptionally activated in responseto M-CSF and its expression favors macrophage differentiationin myeloid progenitor cellsrdquo Journal of Leukocyte Biology vol79 no 1 pp 173ndash183 2006
[41] G Marziali E Perrotti R Ilari U Testa E M Coccia andA Battistini ldquoTranscriptional regulation of the ferritin heavy-chain gene the activity of the CCAAT binding factor NF-Y ismodulated in heme-treated Friend leukemia cells and duringmonocyte-to-macrophage differentiationrdquo Molecular and Cel-lular Biology vol 17 no 3 pp 1387ndash1395 1997
[42] C Shi X Zhang Z Chen et al ldquoIntegrin engagement regulatesmonocyte differentiation through the forkhead transcriptionfactor Foxp1rdquo Journal of Clinical Investigation vol 114 no 3 pp408ndash418 2004
[43] C Shi M Sakuma T Mooroka et al ldquoDown-regulation of theforkhead transcription factor Foxp1 is required for monocytedifferentiation andmacrophage functionrdquoBlood vol 112 no 12pp 4699ndash4711 2008
[44] D F Smith E Galkina K Ley and Y Huo ldquoGRO familychemokines are specialized for monocyte arrest from flowrdquoThe American Journal of Physiology Heart and CirculatoryPhysiology vol 289 no 5 pp H1976ndashH1984 2005
[45] J J Oppenheim O M Z Howard and E Goetzl ChemotacticFactors Neuropeptides and Other Ligands for Seven Transmem-brane Receptors 2000
[46] Y Terashima N Onai M Murai et al ldquoPivotal function forcytoplasmic protein FROUNT in CCR2-mediated monocytechemotaxisrdquo Nature Immunology vol 6 no 8 pp 827ndash8352005
[47] Y Lefkir M Malbouyres D Gotthardt et al ldquoInvolvement ofthe AP-1 adaptor complex in early steps of phagocytosis andmacropinocytosisrdquo Molecular Biology of the Cell vol 15 no 2pp 861ndash869 2004
[48] F Niedergang and P Chavrier ldquoSignaling and membranedynamics during phagocytosis many roads lead to the phago-somerdquo Current Opinion in Cell Biology vol 16 no 4 pp 422ndash428 2004
[49] D Cox D J Lee B M Dale J Calafat and S Green-berg ldquoA Rab11-containing rapidly recycling compartment inmacrophages that promotes phagocytosisrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 97 no 2 pp 680ndash685 2000
[50] O V Vieira C Bucci R E Harrison et al ldquoModulation of Rab5and Rab7 recruitment to phagosomes by phosphatidylinositol3-kinaserdquoMolecular andCellular Biology vol 23 no 7 pp 2501ndash2514 2003
[51] T Khurana J A Brzostowski and A R Kimmel ldquoARab21LIM-onlyCH-LIM complex regulates phagocytosis viaboth activating and inhibitory mechanismsrdquo The EMBO Jour-nal vol 24 no 13 pp 2254ndash2264 2005
16 Oxidative Medicine and Cellular Longevity
[52] S J Martin J G Bradley and T G Cotter ldquoHL-60 cellsinduced to differentiate towards neutrophils subsequently dievia apoptosisrdquo Clinical and Experimental Immunology vol 79no 3 pp 448ndash453 1990
[53] A Benito D Grillot G Nunez and J L Fernandez-Luna ldquoReg-ulation and function of Bcl-2 during differentiation-inducedcell death in HL-60 promyelocytic cellsrdquoThe American Journalof Pathology vol 146 no 2 pp 481ndash490 1995
[54] M Lamkanfi N Festjens W Declercq T V Berghe andP Vandenabeele ldquoCaspases in cell survival proliferation anddifferentiationrdquo Cell Death and Differentiation vol 14 no 1 pp44ndash55 2007
[55] O Sordet C Rebe S Plenchette et al ldquoSpecific involvement ofcaspases in the differentiation of monocytes into macrophagesrdquoBlood vol 100 no 13 pp 4446ndash4453 2002
[56] T-B Kang T Ben-Moshe E E Varfolomeev et al ldquoCaspase-8 serves both apoptotic and nonapoptotic rolesrdquo Journal ofImmunology vol 173 no 5 pp 2976ndash2984 2004
[57] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[58] J Cheng J Yang Y Xia M Karin and B Su ldquoSynergisticinteraction of MEK kinase 2 c-Jun N-terminal kinase (JNK)kinase 2 and JNK1 results in efficient and specific JNK1activationrdquo Molecular and Cellular Biology vol 20 no 7 pp2334ndash2342 2000
[59] J L Blank P Gerwins EM Elliott S Sather andG L JohnsonldquoMolecular cloning of mitogen-activated proteinERK kinasekinases (MEKK) 2 and 3 regulation of sequential phosphoryla-tion pathways involving mitogen-activated protein kinase andc-Jun kinaserdquo Journal of Biological Chemistry vol 271 no 10pp 5361ndash5368 1996
[60] S Papa F Zazzeroni C G Pham C Bubici and G FranzosoldquoLinking JNK signaling to NF-120581B a key to survivalrdquo Journal ofCell Science vol 117 no 22 pp 5197ndash5208 2004
[61] T Kadoya A Khurana M Tcherpakov et al ldquoJAMP a JUB N-terminal kinase 1 (JNK1)-associated membrane protein regu-lates duration of JNK activityrdquo Molecular and Cellular Biologyvol 25 no 19 pp 8619ndash8630 2005
[62] M Qi and E A Elion ldquoMAP kinase pathwaysrdquo Journal of CellScience vol 118 no 16 pp 3569ndash3572 2005
[63] Q Zhao and F S Lee ldquoMitogen-activated protein kinaseERKkinase kinases 2 and 3 activate nuclear factor-120581B through I120581Bkinase-120572 and I120581B kinase-120573rdquoThe Journal of Biological Chemistryvol 274 no 13 pp 8355ndash8358 1999
[64] H Namba V Saenko and S Yamashita ldquoNuclear factor-120581Bin thyroid carcinogenesis and progression a novel therapeutictarget for advanced thyroid cancerrdquo Arquivos Brasileiros deEndocrinologia e Metabologia vol 51 no 5 pp 843ndash851 2007
[65] B Feng S Cheng W S Pear and H-C Liou ldquoNF-kBinhibitor blocks B cell development at two checkpointsrdquoMedi-cal Immunology vol 3 article 1 2004
[66] S Gerondakis and U Siebenlist ldquoRoles of the NF-kappaBpathway in lymphocyte development and functionrdquoCold SpringHarbor Perspectives in Biology vol 2 no 5 Article ID a0001822010
[67] K P Bhat V Balasubramaniyan B Vaillant et al ldquoMesenchy-mal differentiation mediated by NF-kB promotes radiationresistance in glioblastomardquo Cancer Cell vol 24 no 3 pp 331ndash346 2013
Figure 5 Expression of differentiationmarkers onK562 cells treatedwith Chemlali Olive Leaf Extract (COLE) up to 6 days Cells were treatedwith 50 100 and 150 120583gmL of COLE and incubated for different periods Control cells were treated with ethanol at a final concentration of03 Expression of cell surface markers was detected by flow cytometry (a) Expression of CD14 (monocytemacrophage specific marker)(b) Expression of CD11b (granulocytemonocyte specific marker) (c) Expression of CD41 (megakaryocyte specific marker) (d) Expressionof glycophorin A (GPA) (erythrocyte specific marker) Results are represented as the mean plusmn SD of three independent experiments lowast lowastlowastsignificantly different from the control at 119875 lt 005 and 119875 lt 001 respectively
including GYPA HBE1 FECH and ALAS2 which is consis-tent with the decreased expression of erythrocyte differenti-ation marker in the flow cytometry results (Figure 5(d))
Analysis of the microarray data showed the upregu-lation of several proapoptotic genes and genes involvedin the regulation of mitochondrial membrane permeabilitysuch as CASP6 CASP8 DFFA and BID On the otherhandexpression of genes negatively regulating the apoptosis
and those inhibiting the caspase activity was significantlydecreased such as IGF1R HSPA5 and BCL2
Among the upregulated genes we found those related tothe NF-kappaB cascade as well as to the MAPKKK cascadeand the Wnt receptor signaling pathway
Categories related to cell cycle regulation and cell pro-liferation were represented by both upregulated and down-regulated genes Finally some transcription factors werealso downregulated such as FOXP1 whereas others wereupregulated like EGR1 and NFYA
The list of selected genes is presented in Table 3 Fromthis list it is clear that the differentially expressed genes aftertreatment with 100 and 150120583gmL have mostly the sametendency which could validate the obtained results
12 Oxidative Medicine and Cellular Longevity
4 Discussion
Previous reports had indicated that olive leaf extract inducesapoptosis in the human leukemic cell lines HL-60 and Jurkatcells [13 21] In this study we give evidence that COLEexhibits its antileukemia effect by both inducing apoptosisand promoting differentiation of the multipotent humanleukemia K562 cells
Induction of differentiation as well as apoptosis has beenfrequently reported to be associatedwith a loss of proliferativecapacity of the cells It has been reported that olive leaf extractinhibits the proliferation of some cancer cell lines such asB16 melanoma cells HL-60 leukemia cells the breast cancerMCF-7 cells and the glioblastoma cells [12 25ndash27] Ourresults indicate that COLE inhibits the proliferation of K562cells in a dose dependent manner (Figure 2(a)) Howeverdespite the drastic decrease in cell proliferation the viabilityof cells treated with the highest concentration of COLE(150 120583gmL) remains relatively high (80) for up to 3 daysof treatment (Figure 3(b))
Since it is well established that the arrest of the cellcycle progression allows the cells to follow other processessuch as apoptosis and differentiation we then explored theeffect of COLE on the cell cycle distribution (Table 2) Oliveleaf extract has been shown to cause cell cycle arrest atG0G1 phase in both the breast cancer MCF-7 cells and themelanoma B16 cells [12 25] However the treatment timesapplied in both of these studies were limited to 48 h and 18 hrespectively In this study COLE treated cells were arrested atG0G1 on the 1st and 2ndday of treatment which is consistentwith the reported studies Prolonged incubation with COLEshowed an arrest of cell cycle at G2M phase (3rd and 4thday of incubation) The microarray analysis performed forthe 3rd day of treatment revealed a significant increase ofCHEK2 gene expression in COLE treated cells The encodedprotein regulates the cell cycle checkpoint arrest throughthe inhibition of the activity of CDC25A CDC25B andCDC25C [28] CDC25 proteins trigger the entry into mitosisat different points of the cell cycle by activating theCdk-cyclincomplexes CDC25A acts early in the cell cycle regulatingthe G1S transition whereas CDC25B and CDC25C act atG2M [29] Interestingly our results showed a decrease in theexpression of CDC25C gene in COLE treated cells associatedwith an increase in the expression of CDC25A which mayexplain the cell cycle arrest at G2M phase observed at the3rd day of treatment with COLE
Treatment with COLE increased the expression of CD14on the cell surface of treated cells indicating the differenti-ation of K562 cells toward the mono-macrophage lineage(Figure 5(a)) The increase in the percentage of cells positivefor CD11b (Figure 5(b)) confirms this hypothesis In factCD11b is expressed on both monocytes and granulocytesThus together with the increase of CD14 expression theexpression of CD11b suggests that treatment with COLEpromotes the commitment of K562 cells into the mono-cytemacrophage lineage at the expense of the granulocyticdifferentiationThe instant increase of the expression of CD41
on the 1st day of treatment and its absence during the fol-lowing days (Figure 5(c)) could be consistent with monocytedifferentiation instead of megakaryocyte differentiation sinceit has been reported that 120572IIb promoter which initiates thetranscription of CD41 is transcriptionally active in pluripo-tent myeloid progenitors in early stages of erythropoiesisand all along the megakaryocytic differentiation as well asto a lesser extent in the early stages of myelomonocyticdifferentiation and the late stages of erythropoiesis [30] Thisinstant increase could be also explained by the phenomenonof lineage conversion where some hematopoietic progenitorscan be converted into lineages other than their own bythe ectopic expression of some transcription factors [31]In this regard it was previously reported that commonlymphoid progenitors megakaryocyte erythroid progenitorsearly B cells and early T cells could be converted to thegranulocytemonocyte lineage by the induction of CEBP120572activity [32] Microarray results support the absence ofmegakaryocyte differentiation in the 3rd day of treatment Infact data showed a decrease in the expression of genes relatedtomegakaryopoiesis such as the essential regulator of plateletrelease NFE2 and its main candidate target gene TUBB1(encoding the megakaryocyte and platelet specific isoform of120573-tubulin (1205731)) [33] as well as the BACH2 transcription factorreported to be induced during megakaryocyte differentiation[34]
Interestingly results of microarray analysis (Table 3)showed the upregulation of several genes already reportedas key players in monocytemacrophage differentiation inthe COLE treated cells Among these genes the expressionof IFI16 was markedly increased Constitutively expressedin lymphoid cells IFI16 was shown to be associated withthe differentiation of human myeloid cells in response tointerferon gamma [35] Moreover IFI16 has been selectivelyexpressed during the differentiation of myeloid progenitorcells along monocytic lineage while it is downregulatedduring the commitment toward the granulocytic or erythroidlineage suggesting its potential involvement in the differenti-ation andmaturation of the monocytic lineage [36 37] Geneencoding for EGR-1 a positive modulator of macrophagedifferentiation was also significantly upregulated EGR-1 isknown to dictate development of myeloid progenitors alongthe macrophage lineage at the expense of development alongother lineages [38 39] Dauffy et al [40] reported thatIFI16-enforced expression in myeloid progenitors inducedthe expression of EGR-1 and conducted the macrophagedifferentiation in the absence of the macrophage colonystimulating factor (M-CSF) This suggests that EGR-1 couldbe regulated by the expression of IFI16 Treatment with COLEincreased as well the expression of NFYA encoding the Asubunit of the nuclear transcription factor Y whose synthesiswas previously reported during the terminal differentiationof monocytes to macrophages [38 41] The downregula-tion of FOXP1 in COLE treated K562 cells is consistentwith monocytemacrophage differentiation since it has beendemonstrated that the expression of FOXP1 was markedlydecreased in monocyte-induced differentiation of HL-60cells as well as in human peripheral blood monocytes and
Oxidative Medicine and Cellular Longevity 13
that the overexpression of FOXP1 prevented the morphologicmacrophage-like differentiation [42 43]
Monocytes and macrophages have several functionalcharacteristics including cell adhesion migration chemo-taxis and phagocytic activity Interestingly an enhancedexpression of the chemokines genes CXCL8 (IL8) CXCL2andCXCL3was found in COLE treated cells IL-8 productionhas been observed in vitro in a wide variety of cells includingmonocytes T lymphocytes neutrophils vascular endothe-lial cells dermal fibroblasts keratinocytes hepatocytes andhuman gastric cancer cells while the production of CXCL2andCXCL3was only described inmonocytes fibroblasts andendothelial cells [44 45] Another gene related to chemotaxiswas upregulated NUP85 which is an essential component ofthe nuclear pore complex and was reported to be involved inCCR2-mediated chemotaxis of monocytes [46]
COLE treatment also enhanced the expression of somegenes related to phagocytosis AP1G1 andRab proteins relatedgenes AP1G1 is a subunit of clathrin-associated adaptorprotein complex 1 whose role has been recently highlightedfor efficient phagocytosis at an early stage of phagosomeformation since it participates in the extension of the phago-cytic cup [47 48] The small GTPases Rab are key regulatorsof intracellular membrane trafficking Recent studies havedemonstrated that several RAB proteins play an importantrole in phagocytosis RAB proteins such as RAB11 Rab5 andRab 21 participate in the phagosome formation and matura-tion and are necessary for the phagocytic activity [48ndash51]
Consistently with the increase in Annexin V positivecells (Figure 4) treatment with COLE upregulated someproapoptotic genes such as CASP6 CASP8 DFFA and BIDand downregulated the apoptosis suppressor BCL2 as wellas the caspase inhibitors genes (Table 3) Olive leaf waspreviously shown to induce morphological changes thatare characteristic of apoptosis in leukemia cells but themechanisms underlying this effect were not well investi-gated [13 21] The increase in the proportion of apoptoticcells observed from the 4th day of treatment with COLEcould be explained by the programmed death of the fullydifferentiated cells (Figure 4) In fact this increase wasaccompanied by an important decrease in cell viability aswell as a diminution in CD14 expression (Figures 3(b) and5(a)) Elsewhere it is well established that in hematopoietictissues apoptosis is coupled to terminal differentiation ofmyeloid progenitors even though themechanism responsiblefor the activation of apoptosis during myeloid maturationis still poorly understood [52 53] It is also noteworthy tomention that several reports have highlighted the role ofcaspases in the terminal differentiation of a variety of celltypes [54] One example is the differentiation of human bloodmonocytes into macrophages a process that is blocked bysynthetic caspase inhibitors [55] It has been also reportedthat caspase 8 deletion in bone-marrow cells resulted in arrestof hemopoietic progenitor functioning and that its deletionin cells of the myelomonocytic lineage led to the arrest ofdifferentiation into macrophages and consequently to celldeath [56]
Among the differentially expressed genes in COLEtreated cells (Table 3) genes encoding the mitogen-activated
protein kinase kinase kinases (MAPKKKs) MAP3K2MAP3K5 and MAP3K7 were highly expressed whileMAP2K5 and MAPK14p38-120572 were downregulated TheMAPKKKs act at the upstream of the MAP kinase cascadewhich regulates important cellular processes such as geneexpression cell proliferation differentiation cell survivaland death The MAP3K5 activates the c-Jun N-terminalkinase (JNK) and p38 mitogen-activated protein kinases[57] while the MAP3K2 is reported to preferentially activateJNK [58 59] However the significant decrease in theMAPK14p38-120572 may suggest that the effect of COLE onK562 cells is likely to be mediated by the JNK MAPKsrather than the p38 MAPKs JNK plays an important rolein apoptosis pathways weather by activating proapoptoticor antiapoptotic genes Particularly it was shown thatprolonged but not transient JNK activation promotes thecell death [60] Our results showed that JKAMP gene whichencodes for a membrane-anchored regulator of the durationof JNK1 activity was upregulated (154 plusmn 0051) The elevatedexpression of JKAMP as reported by Kadoya et al [61]results in sustained JNK activity These findings may indicatethe potential role of JNK pathway in the COLE-inducedapoptosis in K562 cells
Thanks to their large regulatory domains some MAP-KKKs can interact with upstream regulators have functionssuch as ubiquitylation and be activated by relief of autoin-hibition and oligomerization [62] In this context MAP3K2has been reported to play an important role in NF-kappaBsignaling pathway by activating the I-kappaB kinases whichin turn phosphorylate the inhibitory factors of the nuclearfactor kappaB (NF-kappaB) resulting in their rapid ubiq-uitination and the liberation of NF-kappaB complex whichtranslocate from the cytoplasm to the nucleus [63] MAP3K7in association with other proteins could also be required forthe activation of NF-kappaB According to the GO analysis(Table S1) COLE treatment upregulated genes implicatedin NF-kappaB-related categories such as positive regulationof NF-kappaB transcription factor activity I-kappaB kinase-NF-kappaB cascade and its positive regulation and I-kappaBphosphorylation Genes encoding the signal transducers inthe NF-kappaB pathway TRAF5 and TRAF6 as well asthe activator of NF-kappaB signaling SNIP1 were upregu-lated Interestingly an increased expression of NFKB1 geneencoding for the precursor of the mature NF-kappaB p50was detected in COLE treated cells compared to untreatedK562 cells NFKB is well recognized as a central activator ofthe antiapoptotic cascades in response to external stimuli orintrinsic immune reactions and its prosurvival activity hasbeen implicated in a variety of biological processes [60 64]In the B cell lineage the activity of NF-120581B is required forthe completion of various developmental stages includingdifferentiation and the response of these cells to antigens [65]NF-kappaB also plays an important role in the survival anddevelopment of T cells [66] and has been recently reported tomediate the differentiation of mesenchymal stem cells [67]
Several studies have demonstrated a crosstalk betweenthe NF-kappaB and JNK pathways seen as an inhibitoryeffect of NF-kappaB on apoptosis through the suppressionof JNK activity and that this suppressive effect may occur
14 Oxidative Medicine and Cellular Longevity
through different mechanisms [60] This suggests that thebalance between JNK and NF-kappaB activities is crucial todetermine the cell fate survival or deathWehypothesize herethat COLE-induced apoptosis or differentiation is likely to bedetermined by the crosstalk between these two pathways inK562 cells Further investigations regarding protein expres-sion and activity are needed to confirm these findings
5 Conclusions
Our study demonstrates for the first time that olive leafextract exhibits an antileukemia effect on the human chronicmyeloid leukemia cells Olive leaf extract was shown toinhibit the proliferation of K562 cells by inducing cell cyclearrest apoptosis and differentiation toward the monocytelineage The induced expression of molecules involved indifferentiation toward the monocytemacrophage lineage aswell asmolecules related to apoptosis and cell cycle regulationconfirmed these findings and provides insights into themechanism by which olive leaf exhibits its antileukemiaeffect
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported by the JICA-JST Science andTechnology Research Partnership for Sustainable Develop-ment (SATREPS) Project ldquoValorization of Bio-Resources inSemi-Arid and Arid Land for Regional Developmentrdquo
References
[1] M Leszczyniecka T Roberts P Dent S Grant and P B FisherldquoDifferentiation therapy of human cancer basic science andclinical applicationsrdquo Pharmacology and Therapeutics vol 90no 2-3 pp 105ndash156 2001
[2] A I Spira and M A Carducci ldquoDifferentiation therapyrdquoCurrent Opinion in Pharmacology vol 3 no 4 pp 338ndash3432003
[3] S Nobili D Lippi E Witort et al ldquoNatural compounds forcancer treatment and preventionrdquo Pharmacological Researchvol 59 no 6 pp 365ndash378 2009
[4] S Ramos ldquoEffects of dietary flavonoids on apoptotic pathwaysrelated to cancer chemopreventionrdquo Journal of NutritionalBiochemistry vol 18 no 7 pp 427ndash442 2007
[5] V Hajhashemi G Vaseghi M Pourfarzam and A AbdollahildquoAre antioxidants helpful for disease preventionrdquo Research inPharmaceutical Sciences vol 5 no 1 pp 1ndash8 2010
[6] M Bouaziz and S Sayadi ldquoIsolation and evaluation of antiox-idants from leaves of a Tunisian cultivar olive treerdquo EuropeanJournal of Lipid Science andTechnology vol 107 no 7-8 pp 497ndash504 2005
[7] A P Pereira I C F R Ferreira F Marcelino et al ldquoPhenoliccompounds and antimicrobial activity of olive (Olea europaeaL Cv Cobrancosa) leavesrdquo Molecules vol 12 no 5 pp 1153ndash1162 2007
[8] L I Somova F O Shode P Ramnanan and A Nadar ldquoAnti-hypertensive antiatherosclerotic and antioxidant activity oftriterpenoids isolated from Olea europaea subspecies africanaleavesrdquo Journal of Ethnopharmacology vol 84 no 2-3 pp 299ndash305 2003
[9] V Micol N Caturla L Perez-Fons V Mas L Perez andA Estepa ldquoThe olive leaf extract exhibits antiviral activityagainst viral haemorrhagic septicaemia rhabdovirus (VHSV)rdquoAntiviral Research vol 66 no 2-3 pp 129ndash136 2005
[10] J Wainstein T Ganz M Boaz et al ldquoOlive leaf extract as ahypoglycemic agent in both human diabetic subjects and inratsrdquo Journal of Medicinal Food vol 15 no 7 pp 605ndash610 2012
[11] L Seddik TM Bah A AouesM Slimani andM BenderdourldquoElucidation of mechanisms underlying the protective effects ofolive leaf extract against lead-induced neurotoxicity in Wistarratsrdquo Journal of Toxicological Sciences vol 36 no 6 pp 797ndash809 2011
[12] Z Bouallagui J Han H Isoda and S Sayadi ldquoHydroxytyrosolrich extract from olive leaves modulates cell cycle progressionin MCF-7 human breast cancer cellsrdquo Food and ChemicalToxicology vol 49 no 1 pp 179ndash184 2011
[13] R Fares S Bazzi S E Baydoun and R M Abdel-Massih ldquoTheantioxidant and anti-proliferative activity of the Lebanese Oleaeuropaea extractrdquo Plant Foods for Human Nutrition vol 66 no1 pp 58ndash63 2011
[14] V Goulas V Exarchou A N Troganis et al ldquoPhytochemicalsin olive-leaf extracts and their antiproliferative activity againstcancer and endothelial cellsrdquo Molecular Nutrition and FoodResearch vol 53 no 5 pp 600ndash608 2009
[15] M de Bock E B Thorstensen J G Derraik H V HendersonP L Hofman and W S Cutfield ldquoHuman absorption andmetabolism of oleuropein and hydroxytyrosol ingested as olive(Olea europaea L) leaf extractrdquo Molecular Nutrition and FoodResearch vol 57 no 11 pp 2079ndash2085 2013
[16] R Briante M Patumi S Terenziani E Bismuto F Febbraioand R Nucci ldquoOlea europaea L leaf extract and derivativesantioxidant propertiesrdquo Journal of Agricultural and Food Chem-istry vol 50 no 17 pp 4934ndash4940 2002
[17] O Benavente-Garcıa J Castillo J Lorente A Ortuno and J ADel Rio ldquoAntioxidant activity of phenolics extracted from Oleaeuropaea L leavesrdquo Food Chemistry vol 68 no 4 pp 457ndash4622000
[18] S H Omar ldquoOleuropein in olive and its pharmacologicaleffectsrdquo Scientia Pharmaceutica vol 78 no 2 pp 133ndash154 2010
[19] H Jemai M Bouaziz I Fki A El Feki and S SayadildquoHypolipidimic and antioxidant activities of oleuropein and itshydrolysis derivative-rich extracts from Chemlali olive leavesrdquoChemico-Biological Interactions vol 176 no 2-3 pp 88ndash982008
[20] R Garcıa-Villalba M Larrosa S Possemiers F A Tomas-Barberan and J C Espın ldquoBioavailability of phenolics from anoleuropein-rich olive (Olea europaea) leaf extract and its acuteeffect on plasma antioxidant status comparison between pre-and postmenopausal womenrdquo European Journal of Nutrition2013
[21] J Anter Z Fernandez-Bedmar M Villatoro-Pulido et al ldquoApilot study on the DNA-protective cytotoxic and apoptosis-inducing properties of olive-leaf extractsrdquo Mutation Researchvol 723 no 2 pp 165ndash170 2011
[22] J A Sutherland A R Turner and P Mannoni ldquoDifferen-tiation of K562 leukemia cells along erythroid macrophage
Oxidative Medicine and Cellular Longevity 15
and megakaryocyte lineagesrdquo Journal of Biological ResponseModifiers vol 5 no 3 pp 250ndash262 1986
[23] K Schwenke H-P Peterson K-H Wangenheim and L EFeinendegen ldquoInduction of differentiation in erythroleukemicK562 cells by 120574-irradiationrdquo Leukemia Research vol 19 no 12pp 955ndash961 1995
[24] V N Enujiugha J Y Talabi S A Malomo and A I OlagunjuldquoDPPH radical scavenging capacity of phenolic extracts fromAfrican Yam Bean (Sphenostylis stenocarpa)rdquo Food and Nutri-tion Sciences vol 3 pp 7ndash13 2012
[25] S A Mijatovic G S Timotijevic D M Miljkovic et alldquoMultiple antimelanoma potential of dry olive leaf extractrdquoInternational Journal of Cancer vol 128 no 8 pp 1955ndash19652011
[26] L Abaza T P N Talorete P Yamada Y KuritaM Zarrouk andH Isoda ldquoInduction of growth inhibition and differentiation ofhuman leukemia HL-60 cells by a Tunisian Gerboui olive leafextractrdquo Bioscience Biotechnology and Biochemistry vol 71 no5 pp 1306ndash1312 2007
[27] B Tunca G Tezcan G Cecener et al ldquoOlea europea leafextract alters microRNA expression in human glioblastomacellsrdquo Journal of Cancer Research and Clinical Oncology vol 138no 11 pp 1831ndash1844 2012
[28] S Matsuoka M Huang and S J Elledge ldquoLinkage of ATM tocell cycle regulation by the Chk2 protein kinaserdquo Science vol282 no 5395 pp 1893ndash1897 1998
[29] C Frazer and P G Young ldquoPhosphorylation mediated regu-lation of Cdc25 activity localization and stabilityrdquo in ProteinPhosphorylation in Human Health C Huang Ed pp 395ndash436InTech 2012
[30] N Debili C Robin V Schiavon et al ldquoDifferent expression ofCD41 on human lymphoid andmyeloid progenitors from adultsand neonatesrdquo Blood vol 97 no 7 pp 2023ndash2030 2001
[31] H Nakajima ldquoRole of transcription factors in differentiationand reprogramming of hematopoietic cellsrdquo Keio Journal ofMedicine vol 60 no 2 pp 47ndash55 2011
[32] Y Fukuchi F Shibata M Ito et al ldquoComprehensive analysis ofmyeloid lineage conversion using mice expressing an inducibleform of CEBP120572rdquoThe EMBO Journal vol 25 no 14 pp 3398ndash3410 2006
[33] R A Shivdasani ldquoMolecular and transcriptional regulation ofmegakaryocyte differentiationrdquo Stem Cells vol 19 no 5 pp397ndash407 2001
[34] K Terui Y Takahashi J Kitazawa T TokiM Yokoyama and EIto ldquoExpression of transcription factors during megakaryocyticdifferentiation of CD34+ cells from human cord blood inducedby thrombopoietinrdquo Tohoku Journal of Experimental Medicinevol 192 no 4 pp 259ndash273 2000
[35] J A Trapani M Dawson V A Apostolidis and K A BrowneldquoGenomic organization of IFI16 an interferon-inducible genewhose expression is associated with human myeloid cell differ-entiation correlation of predicted protein domains with exonorganizationrdquo Immunogenetics vol 40 no 6 pp 415ndash424 1994
[36] M J Dawson N J Elwood R W Johnstone and J A TrapanildquoThe IFN-inducible nucleoprotein IFI 16 is expressed in cellsof the monocyte lineage but is rapidly and markedly down-regulated in other myeloid precursor populationsrdquo Journal ofLeukocyte Biology vol 64 no 4 pp 546ndash554 1998
[37] M Gariglio M De Andrea M Lembo et al ldquoThe murinehomolog of the HIN 200 family Ifi 204 is constitutivelyexpressed in myeloid cells and selectively induced in the
monocytemacrophage lineagerdquo Journal of Leukocyte Biologyvol 64 no 5 pp 608ndash614 1998
[38] A F Valledor F E Borras M Cullell-Young and A CeladaldquoTranscription factors that regulate monocytemacrophage dif-ferentiationrdquo Journal of Leukocyte Biology vol 63 no 4 pp405ndash417 1998
[39] K Krishnaraju B Hoffman and D A Liebermann ldquoEarlygrowth response gene 1 stimulates development of hematopoi-etic progenitor cells along the macrophage lineage at theexpense of the granulocyte and erythroid lineagesrdquo Blood vol97 no 5 pp 1298ndash1305 2001
[40] J Dauffy G Mouchiroud and R P Bourette ldquoThe interferon-inducible gene Ifi204 is transcriptionally activated in responseto M-CSF and its expression favors macrophage differentiationin myeloid progenitor cellsrdquo Journal of Leukocyte Biology vol79 no 1 pp 173ndash183 2006
[41] G Marziali E Perrotti R Ilari U Testa E M Coccia andA Battistini ldquoTranscriptional regulation of the ferritin heavy-chain gene the activity of the CCAAT binding factor NF-Y ismodulated in heme-treated Friend leukemia cells and duringmonocyte-to-macrophage differentiationrdquo Molecular and Cel-lular Biology vol 17 no 3 pp 1387ndash1395 1997
[42] C Shi X Zhang Z Chen et al ldquoIntegrin engagement regulatesmonocyte differentiation through the forkhead transcriptionfactor Foxp1rdquo Journal of Clinical Investigation vol 114 no 3 pp408ndash418 2004
[43] C Shi M Sakuma T Mooroka et al ldquoDown-regulation of theforkhead transcription factor Foxp1 is required for monocytedifferentiation andmacrophage functionrdquoBlood vol 112 no 12pp 4699ndash4711 2008
[44] D F Smith E Galkina K Ley and Y Huo ldquoGRO familychemokines are specialized for monocyte arrest from flowrdquoThe American Journal of Physiology Heart and CirculatoryPhysiology vol 289 no 5 pp H1976ndashH1984 2005
[45] J J Oppenheim O M Z Howard and E Goetzl ChemotacticFactors Neuropeptides and Other Ligands for Seven Transmem-brane Receptors 2000
[46] Y Terashima N Onai M Murai et al ldquoPivotal function forcytoplasmic protein FROUNT in CCR2-mediated monocytechemotaxisrdquo Nature Immunology vol 6 no 8 pp 827ndash8352005
[47] Y Lefkir M Malbouyres D Gotthardt et al ldquoInvolvement ofthe AP-1 adaptor complex in early steps of phagocytosis andmacropinocytosisrdquo Molecular Biology of the Cell vol 15 no 2pp 861ndash869 2004
[48] F Niedergang and P Chavrier ldquoSignaling and membranedynamics during phagocytosis many roads lead to the phago-somerdquo Current Opinion in Cell Biology vol 16 no 4 pp 422ndash428 2004
[49] D Cox D J Lee B M Dale J Calafat and S Green-berg ldquoA Rab11-containing rapidly recycling compartment inmacrophages that promotes phagocytosisrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 97 no 2 pp 680ndash685 2000
[50] O V Vieira C Bucci R E Harrison et al ldquoModulation of Rab5and Rab7 recruitment to phagosomes by phosphatidylinositol3-kinaserdquoMolecular andCellular Biology vol 23 no 7 pp 2501ndash2514 2003
[51] T Khurana J A Brzostowski and A R Kimmel ldquoARab21LIM-onlyCH-LIM complex regulates phagocytosis viaboth activating and inhibitory mechanismsrdquo The EMBO Jour-nal vol 24 no 13 pp 2254ndash2264 2005
16 Oxidative Medicine and Cellular Longevity
[52] S J Martin J G Bradley and T G Cotter ldquoHL-60 cellsinduced to differentiate towards neutrophils subsequently dievia apoptosisrdquo Clinical and Experimental Immunology vol 79no 3 pp 448ndash453 1990
[53] A Benito D Grillot G Nunez and J L Fernandez-Luna ldquoReg-ulation and function of Bcl-2 during differentiation-inducedcell death in HL-60 promyelocytic cellsrdquoThe American Journalof Pathology vol 146 no 2 pp 481ndash490 1995
[54] M Lamkanfi N Festjens W Declercq T V Berghe andP Vandenabeele ldquoCaspases in cell survival proliferation anddifferentiationrdquo Cell Death and Differentiation vol 14 no 1 pp44ndash55 2007
[55] O Sordet C Rebe S Plenchette et al ldquoSpecific involvement ofcaspases in the differentiation of monocytes into macrophagesrdquoBlood vol 100 no 13 pp 4446ndash4453 2002
[56] T-B Kang T Ben-Moshe E E Varfolomeev et al ldquoCaspase-8 serves both apoptotic and nonapoptotic rolesrdquo Journal ofImmunology vol 173 no 5 pp 2976ndash2984 2004
[57] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[58] J Cheng J Yang Y Xia M Karin and B Su ldquoSynergisticinteraction of MEK kinase 2 c-Jun N-terminal kinase (JNK)kinase 2 and JNK1 results in efficient and specific JNK1activationrdquo Molecular and Cellular Biology vol 20 no 7 pp2334ndash2342 2000
[59] J L Blank P Gerwins EM Elliott S Sather andG L JohnsonldquoMolecular cloning of mitogen-activated proteinERK kinasekinases (MEKK) 2 and 3 regulation of sequential phosphoryla-tion pathways involving mitogen-activated protein kinase andc-Jun kinaserdquo Journal of Biological Chemistry vol 271 no 10pp 5361ndash5368 1996
[60] S Papa F Zazzeroni C G Pham C Bubici and G FranzosoldquoLinking JNK signaling to NF-120581B a key to survivalrdquo Journal ofCell Science vol 117 no 22 pp 5197ndash5208 2004
[61] T Kadoya A Khurana M Tcherpakov et al ldquoJAMP a JUB N-terminal kinase 1 (JNK1)-associated membrane protein regu-lates duration of JNK activityrdquo Molecular and Cellular Biologyvol 25 no 19 pp 8619ndash8630 2005
[62] M Qi and E A Elion ldquoMAP kinase pathwaysrdquo Journal of CellScience vol 118 no 16 pp 3569ndash3572 2005
[63] Q Zhao and F S Lee ldquoMitogen-activated protein kinaseERKkinase kinases 2 and 3 activate nuclear factor-120581B through I120581Bkinase-120572 and I120581B kinase-120573rdquoThe Journal of Biological Chemistryvol 274 no 13 pp 8355ndash8358 1999
[64] H Namba V Saenko and S Yamashita ldquoNuclear factor-120581Bin thyroid carcinogenesis and progression a novel therapeutictarget for advanced thyroid cancerrdquo Arquivos Brasileiros deEndocrinologia e Metabologia vol 51 no 5 pp 843ndash851 2007
[65] B Feng S Cheng W S Pear and H-C Liou ldquoNF-kBinhibitor blocks B cell development at two checkpointsrdquoMedi-cal Immunology vol 3 article 1 2004
[66] S Gerondakis and U Siebenlist ldquoRoles of the NF-kappaBpathway in lymphocyte development and functionrdquoCold SpringHarbor Perspectives in Biology vol 2 no 5 Article ID a0001822010
[67] K P Bhat V Balasubramaniyan B Vaillant et al ldquoMesenchy-mal differentiation mediated by NF-kB promotes radiationresistance in glioblastomardquo Cancer Cell vol 24 no 3 pp 331ndash346 2013
Figure 5 Expression of differentiationmarkers onK562 cells treatedwith Chemlali Olive Leaf Extract (COLE) up to 6 days Cells were treatedwith 50 100 and 150 120583gmL of COLE and incubated for different periods Control cells were treated with ethanol at a final concentration of03 Expression of cell surface markers was detected by flow cytometry (a) Expression of CD14 (monocytemacrophage specific marker)(b) Expression of CD11b (granulocytemonocyte specific marker) (c) Expression of CD41 (megakaryocyte specific marker) (d) Expressionof glycophorin A (GPA) (erythrocyte specific marker) Results are represented as the mean plusmn SD of three independent experiments lowast lowastlowastsignificantly different from the control at 119875 lt 005 and 119875 lt 001 respectively
including GYPA HBE1 FECH and ALAS2 which is consis-tent with the decreased expression of erythrocyte differenti-ation marker in the flow cytometry results (Figure 5(d))
Analysis of the microarray data showed the upregu-lation of several proapoptotic genes and genes involvedin the regulation of mitochondrial membrane permeabilitysuch as CASP6 CASP8 DFFA and BID On the otherhandexpression of genes negatively regulating the apoptosis
and those inhibiting the caspase activity was significantlydecreased such as IGF1R HSPA5 and BCL2
Among the upregulated genes we found those related tothe NF-kappaB cascade as well as to the MAPKKK cascadeand the Wnt receptor signaling pathway
Categories related to cell cycle regulation and cell pro-liferation were represented by both upregulated and down-regulated genes Finally some transcription factors werealso downregulated such as FOXP1 whereas others wereupregulated like EGR1 and NFYA
The list of selected genes is presented in Table 3 Fromthis list it is clear that the differentially expressed genes aftertreatment with 100 and 150120583gmL have mostly the sametendency which could validate the obtained results
12 Oxidative Medicine and Cellular Longevity
4 Discussion
Previous reports had indicated that olive leaf extract inducesapoptosis in the human leukemic cell lines HL-60 and Jurkatcells [13 21] In this study we give evidence that COLEexhibits its antileukemia effect by both inducing apoptosisand promoting differentiation of the multipotent humanleukemia K562 cells
Induction of differentiation as well as apoptosis has beenfrequently reported to be associatedwith a loss of proliferativecapacity of the cells It has been reported that olive leaf extractinhibits the proliferation of some cancer cell lines such asB16 melanoma cells HL-60 leukemia cells the breast cancerMCF-7 cells and the glioblastoma cells [12 25ndash27] Ourresults indicate that COLE inhibits the proliferation of K562cells in a dose dependent manner (Figure 2(a)) Howeverdespite the drastic decrease in cell proliferation the viabilityof cells treated with the highest concentration of COLE(150 120583gmL) remains relatively high (80) for up to 3 daysof treatment (Figure 3(b))
Since it is well established that the arrest of the cellcycle progression allows the cells to follow other processessuch as apoptosis and differentiation we then explored theeffect of COLE on the cell cycle distribution (Table 2) Oliveleaf extract has been shown to cause cell cycle arrest atG0G1 phase in both the breast cancer MCF-7 cells and themelanoma B16 cells [12 25] However the treatment timesapplied in both of these studies were limited to 48 h and 18 hrespectively In this study COLE treated cells were arrested atG0G1 on the 1st and 2ndday of treatment which is consistentwith the reported studies Prolonged incubation with COLEshowed an arrest of cell cycle at G2M phase (3rd and 4thday of incubation) The microarray analysis performed forthe 3rd day of treatment revealed a significant increase ofCHEK2 gene expression in COLE treated cells The encodedprotein regulates the cell cycle checkpoint arrest throughthe inhibition of the activity of CDC25A CDC25B andCDC25C [28] CDC25 proteins trigger the entry into mitosisat different points of the cell cycle by activating theCdk-cyclincomplexes CDC25A acts early in the cell cycle regulatingthe G1S transition whereas CDC25B and CDC25C act atG2M [29] Interestingly our results showed a decrease in theexpression of CDC25C gene in COLE treated cells associatedwith an increase in the expression of CDC25A which mayexplain the cell cycle arrest at G2M phase observed at the3rd day of treatment with COLE
Treatment with COLE increased the expression of CD14on the cell surface of treated cells indicating the differenti-ation of K562 cells toward the mono-macrophage lineage(Figure 5(a)) The increase in the percentage of cells positivefor CD11b (Figure 5(b)) confirms this hypothesis In factCD11b is expressed on both monocytes and granulocytesThus together with the increase of CD14 expression theexpression of CD11b suggests that treatment with COLEpromotes the commitment of K562 cells into the mono-cytemacrophage lineage at the expense of the granulocyticdifferentiationThe instant increase of the expression of CD41
on the 1st day of treatment and its absence during the fol-lowing days (Figure 5(c)) could be consistent with monocytedifferentiation instead of megakaryocyte differentiation sinceit has been reported that 120572IIb promoter which initiates thetranscription of CD41 is transcriptionally active in pluripo-tent myeloid progenitors in early stages of erythropoiesisand all along the megakaryocytic differentiation as well asto a lesser extent in the early stages of myelomonocyticdifferentiation and the late stages of erythropoiesis [30] Thisinstant increase could be also explained by the phenomenonof lineage conversion where some hematopoietic progenitorscan be converted into lineages other than their own bythe ectopic expression of some transcription factors [31]In this regard it was previously reported that commonlymphoid progenitors megakaryocyte erythroid progenitorsearly B cells and early T cells could be converted to thegranulocytemonocyte lineage by the induction of CEBP120572activity [32] Microarray results support the absence ofmegakaryocyte differentiation in the 3rd day of treatment Infact data showed a decrease in the expression of genes relatedtomegakaryopoiesis such as the essential regulator of plateletrelease NFE2 and its main candidate target gene TUBB1(encoding the megakaryocyte and platelet specific isoform of120573-tubulin (1205731)) [33] as well as the BACH2 transcription factorreported to be induced during megakaryocyte differentiation[34]
Interestingly results of microarray analysis (Table 3)showed the upregulation of several genes already reportedas key players in monocytemacrophage differentiation inthe COLE treated cells Among these genes the expressionof IFI16 was markedly increased Constitutively expressedin lymphoid cells IFI16 was shown to be associated withthe differentiation of human myeloid cells in response tointerferon gamma [35] Moreover IFI16 has been selectivelyexpressed during the differentiation of myeloid progenitorcells along monocytic lineage while it is downregulatedduring the commitment toward the granulocytic or erythroidlineage suggesting its potential involvement in the differenti-ation andmaturation of the monocytic lineage [36 37] Geneencoding for EGR-1 a positive modulator of macrophagedifferentiation was also significantly upregulated EGR-1 isknown to dictate development of myeloid progenitors alongthe macrophage lineage at the expense of development alongother lineages [38 39] Dauffy et al [40] reported thatIFI16-enforced expression in myeloid progenitors inducedthe expression of EGR-1 and conducted the macrophagedifferentiation in the absence of the macrophage colonystimulating factor (M-CSF) This suggests that EGR-1 couldbe regulated by the expression of IFI16 Treatment with COLEincreased as well the expression of NFYA encoding the Asubunit of the nuclear transcription factor Y whose synthesiswas previously reported during the terminal differentiationof monocytes to macrophages [38 41] The downregula-tion of FOXP1 in COLE treated K562 cells is consistentwith monocytemacrophage differentiation since it has beendemonstrated that the expression of FOXP1 was markedlydecreased in monocyte-induced differentiation of HL-60cells as well as in human peripheral blood monocytes and
Oxidative Medicine and Cellular Longevity 13
that the overexpression of FOXP1 prevented the morphologicmacrophage-like differentiation [42 43]
Monocytes and macrophages have several functionalcharacteristics including cell adhesion migration chemo-taxis and phagocytic activity Interestingly an enhancedexpression of the chemokines genes CXCL8 (IL8) CXCL2andCXCL3was found in COLE treated cells IL-8 productionhas been observed in vitro in a wide variety of cells includingmonocytes T lymphocytes neutrophils vascular endothe-lial cells dermal fibroblasts keratinocytes hepatocytes andhuman gastric cancer cells while the production of CXCL2andCXCL3was only described inmonocytes fibroblasts andendothelial cells [44 45] Another gene related to chemotaxiswas upregulated NUP85 which is an essential component ofthe nuclear pore complex and was reported to be involved inCCR2-mediated chemotaxis of monocytes [46]
COLE treatment also enhanced the expression of somegenes related to phagocytosis AP1G1 andRab proteins relatedgenes AP1G1 is a subunit of clathrin-associated adaptorprotein complex 1 whose role has been recently highlightedfor efficient phagocytosis at an early stage of phagosomeformation since it participates in the extension of the phago-cytic cup [47 48] The small GTPases Rab are key regulatorsof intracellular membrane trafficking Recent studies havedemonstrated that several RAB proteins play an importantrole in phagocytosis RAB proteins such as RAB11 Rab5 andRab 21 participate in the phagosome formation and matura-tion and are necessary for the phagocytic activity [48ndash51]
Consistently with the increase in Annexin V positivecells (Figure 4) treatment with COLE upregulated someproapoptotic genes such as CASP6 CASP8 DFFA and BIDand downregulated the apoptosis suppressor BCL2 as wellas the caspase inhibitors genes (Table 3) Olive leaf waspreviously shown to induce morphological changes thatare characteristic of apoptosis in leukemia cells but themechanisms underlying this effect were not well investi-gated [13 21] The increase in the proportion of apoptoticcells observed from the 4th day of treatment with COLEcould be explained by the programmed death of the fullydifferentiated cells (Figure 4) In fact this increase wasaccompanied by an important decrease in cell viability aswell as a diminution in CD14 expression (Figures 3(b) and5(a)) Elsewhere it is well established that in hematopoietictissues apoptosis is coupled to terminal differentiation ofmyeloid progenitors even though themechanism responsiblefor the activation of apoptosis during myeloid maturationis still poorly understood [52 53] It is also noteworthy tomention that several reports have highlighted the role ofcaspases in the terminal differentiation of a variety of celltypes [54] One example is the differentiation of human bloodmonocytes into macrophages a process that is blocked bysynthetic caspase inhibitors [55] It has been also reportedthat caspase 8 deletion in bone-marrow cells resulted in arrestof hemopoietic progenitor functioning and that its deletionin cells of the myelomonocytic lineage led to the arrest ofdifferentiation into macrophages and consequently to celldeath [56]
Among the differentially expressed genes in COLEtreated cells (Table 3) genes encoding the mitogen-activated
protein kinase kinase kinases (MAPKKKs) MAP3K2MAP3K5 and MAP3K7 were highly expressed whileMAP2K5 and MAPK14p38-120572 were downregulated TheMAPKKKs act at the upstream of the MAP kinase cascadewhich regulates important cellular processes such as geneexpression cell proliferation differentiation cell survivaland death The MAP3K5 activates the c-Jun N-terminalkinase (JNK) and p38 mitogen-activated protein kinases[57] while the MAP3K2 is reported to preferentially activateJNK [58 59] However the significant decrease in theMAPK14p38-120572 may suggest that the effect of COLE onK562 cells is likely to be mediated by the JNK MAPKsrather than the p38 MAPKs JNK plays an important rolein apoptosis pathways weather by activating proapoptoticor antiapoptotic genes Particularly it was shown thatprolonged but not transient JNK activation promotes thecell death [60] Our results showed that JKAMP gene whichencodes for a membrane-anchored regulator of the durationof JNK1 activity was upregulated (154 plusmn 0051) The elevatedexpression of JKAMP as reported by Kadoya et al [61]results in sustained JNK activity These findings may indicatethe potential role of JNK pathway in the COLE-inducedapoptosis in K562 cells
Thanks to their large regulatory domains some MAP-KKKs can interact with upstream regulators have functionssuch as ubiquitylation and be activated by relief of autoin-hibition and oligomerization [62] In this context MAP3K2has been reported to play an important role in NF-kappaBsignaling pathway by activating the I-kappaB kinases whichin turn phosphorylate the inhibitory factors of the nuclearfactor kappaB (NF-kappaB) resulting in their rapid ubiq-uitination and the liberation of NF-kappaB complex whichtranslocate from the cytoplasm to the nucleus [63] MAP3K7in association with other proteins could also be required forthe activation of NF-kappaB According to the GO analysis(Table S1) COLE treatment upregulated genes implicatedin NF-kappaB-related categories such as positive regulationof NF-kappaB transcription factor activity I-kappaB kinase-NF-kappaB cascade and its positive regulation and I-kappaBphosphorylation Genes encoding the signal transducers inthe NF-kappaB pathway TRAF5 and TRAF6 as well asthe activator of NF-kappaB signaling SNIP1 were upregu-lated Interestingly an increased expression of NFKB1 geneencoding for the precursor of the mature NF-kappaB p50was detected in COLE treated cells compared to untreatedK562 cells NFKB is well recognized as a central activator ofthe antiapoptotic cascades in response to external stimuli orintrinsic immune reactions and its prosurvival activity hasbeen implicated in a variety of biological processes [60 64]In the B cell lineage the activity of NF-120581B is required forthe completion of various developmental stages includingdifferentiation and the response of these cells to antigens [65]NF-kappaB also plays an important role in the survival anddevelopment of T cells [66] and has been recently reported tomediate the differentiation of mesenchymal stem cells [67]
Several studies have demonstrated a crosstalk betweenthe NF-kappaB and JNK pathways seen as an inhibitoryeffect of NF-kappaB on apoptosis through the suppressionof JNK activity and that this suppressive effect may occur
14 Oxidative Medicine and Cellular Longevity
through different mechanisms [60] This suggests that thebalance between JNK and NF-kappaB activities is crucial todetermine the cell fate survival or deathWehypothesize herethat COLE-induced apoptosis or differentiation is likely to bedetermined by the crosstalk between these two pathways inK562 cells Further investigations regarding protein expres-sion and activity are needed to confirm these findings
5 Conclusions
Our study demonstrates for the first time that olive leafextract exhibits an antileukemia effect on the human chronicmyeloid leukemia cells Olive leaf extract was shown toinhibit the proliferation of K562 cells by inducing cell cyclearrest apoptosis and differentiation toward the monocytelineage The induced expression of molecules involved indifferentiation toward the monocytemacrophage lineage aswell asmolecules related to apoptosis and cell cycle regulationconfirmed these findings and provides insights into themechanism by which olive leaf exhibits its antileukemiaeffect
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported by the JICA-JST Science andTechnology Research Partnership for Sustainable Develop-ment (SATREPS) Project ldquoValorization of Bio-Resources inSemi-Arid and Arid Land for Regional Developmentrdquo
References
[1] M Leszczyniecka T Roberts P Dent S Grant and P B FisherldquoDifferentiation therapy of human cancer basic science andclinical applicationsrdquo Pharmacology and Therapeutics vol 90no 2-3 pp 105ndash156 2001
[2] A I Spira and M A Carducci ldquoDifferentiation therapyrdquoCurrent Opinion in Pharmacology vol 3 no 4 pp 338ndash3432003
[3] S Nobili D Lippi E Witort et al ldquoNatural compounds forcancer treatment and preventionrdquo Pharmacological Researchvol 59 no 6 pp 365ndash378 2009
[4] S Ramos ldquoEffects of dietary flavonoids on apoptotic pathwaysrelated to cancer chemopreventionrdquo Journal of NutritionalBiochemistry vol 18 no 7 pp 427ndash442 2007
[5] V Hajhashemi G Vaseghi M Pourfarzam and A AbdollahildquoAre antioxidants helpful for disease preventionrdquo Research inPharmaceutical Sciences vol 5 no 1 pp 1ndash8 2010
[6] M Bouaziz and S Sayadi ldquoIsolation and evaluation of antiox-idants from leaves of a Tunisian cultivar olive treerdquo EuropeanJournal of Lipid Science andTechnology vol 107 no 7-8 pp 497ndash504 2005
[7] A P Pereira I C F R Ferreira F Marcelino et al ldquoPhenoliccompounds and antimicrobial activity of olive (Olea europaeaL Cv Cobrancosa) leavesrdquo Molecules vol 12 no 5 pp 1153ndash1162 2007
[8] L I Somova F O Shode P Ramnanan and A Nadar ldquoAnti-hypertensive antiatherosclerotic and antioxidant activity oftriterpenoids isolated from Olea europaea subspecies africanaleavesrdquo Journal of Ethnopharmacology vol 84 no 2-3 pp 299ndash305 2003
[9] V Micol N Caturla L Perez-Fons V Mas L Perez andA Estepa ldquoThe olive leaf extract exhibits antiviral activityagainst viral haemorrhagic septicaemia rhabdovirus (VHSV)rdquoAntiviral Research vol 66 no 2-3 pp 129ndash136 2005
[10] J Wainstein T Ganz M Boaz et al ldquoOlive leaf extract as ahypoglycemic agent in both human diabetic subjects and inratsrdquo Journal of Medicinal Food vol 15 no 7 pp 605ndash610 2012
[11] L Seddik TM Bah A AouesM Slimani andM BenderdourldquoElucidation of mechanisms underlying the protective effects ofolive leaf extract against lead-induced neurotoxicity in Wistarratsrdquo Journal of Toxicological Sciences vol 36 no 6 pp 797ndash809 2011
[12] Z Bouallagui J Han H Isoda and S Sayadi ldquoHydroxytyrosolrich extract from olive leaves modulates cell cycle progressionin MCF-7 human breast cancer cellsrdquo Food and ChemicalToxicology vol 49 no 1 pp 179ndash184 2011
[13] R Fares S Bazzi S E Baydoun and R M Abdel-Massih ldquoTheantioxidant and anti-proliferative activity of the Lebanese Oleaeuropaea extractrdquo Plant Foods for Human Nutrition vol 66 no1 pp 58ndash63 2011
[14] V Goulas V Exarchou A N Troganis et al ldquoPhytochemicalsin olive-leaf extracts and their antiproliferative activity againstcancer and endothelial cellsrdquo Molecular Nutrition and FoodResearch vol 53 no 5 pp 600ndash608 2009
[15] M de Bock E B Thorstensen J G Derraik H V HendersonP L Hofman and W S Cutfield ldquoHuman absorption andmetabolism of oleuropein and hydroxytyrosol ingested as olive(Olea europaea L) leaf extractrdquo Molecular Nutrition and FoodResearch vol 57 no 11 pp 2079ndash2085 2013
[16] R Briante M Patumi S Terenziani E Bismuto F Febbraioand R Nucci ldquoOlea europaea L leaf extract and derivativesantioxidant propertiesrdquo Journal of Agricultural and Food Chem-istry vol 50 no 17 pp 4934ndash4940 2002
[17] O Benavente-Garcıa J Castillo J Lorente A Ortuno and J ADel Rio ldquoAntioxidant activity of phenolics extracted from Oleaeuropaea L leavesrdquo Food Chemistry vol 68 no 4 pp 457ndash4622000
[18] S H Omar ldquoOleuropein in olive and its pharmacologicaleffectsrdquo Scientia Pharmaceutica vol 78 no 2 pp 133ndash154 2010
[19] H Jemai M Bouaziz I Fki A El Feki and S SayadildquoHypolipidimic and antioxidant activities of oleuropein and itshydrolysis derivative-rich extracts from Chemlali olive leavesrdquoChemico-Biological Interactions vol 176 no 2-3 pp 88ndash982008
[20] R Garcıa-Villalba M Larrosa S Possemiers F A Tomas-Barberan and J C Espın ldquoBioavailability of phenolics from anoleuropein-rich olive (Olea europaea) leaf extract and its acuteeffect on plasma antioxidant status comparison between pre-and postmenopausal womenrdquo European Journal of Nutrition2013
[21] J Anter Z Fernandez-Bedmar M Villatoro-Pulido et al ldquoApilot study on the DNA-protective cytotoxic and apoptosis-inducing properties of olive-leaf extractsrdquo Mutation Researchvol 723 no 2 pp 165ndash170 2011
[22] J A Sutherland A R Turner and P Mannoni ldquoDifferen-tiation of K562 leukemia cells along erythroid macrophage
Oxidative Medicine and Cellular Longevity 15
and megakaryocyte lineagesrdquo Journal of Biological ResponseModifiers vol 5 no 3 pp 250ndash262 1986
[23] K Schwenke H-P Peterson K-H Wangenheim and L EFeinendegen ldquoInduction of differentiation in erythroleukemicK562 cells by 120574-irradiationrdquo Leukemia Research vol 19 no 12pp 955ndash961 1995
[24] V N Enujiugha J Y Talabi S A Malomo and A I OlagunjuldquoDPPH radical scavenging capacity of phenolic extracts fromAfrican Yam Bean (Sphenostylis stenocarpa)rdquo Food and Nutri-tion Sciences vol 3 pp 7ndash13 2012
[25] S A Mijatovic G S Timotijevic D M Miljkovic et alldquoMultiple antimelanoma potential of dry olive leaf extractrdquoInternational Journal of Cancer vol 128 no 8 pp 1955ndash19652011
[26] L Abaza T P N Talorete P Yamada Y KuritaM Zarrouk andH Isoda ldquoInduction of growth inhibition and differentiation ofhuman leukemia HL-60 cells by a Tunisian Gerboui olive leafextractrdquo Bioscience Biotechnology and Biochemistry vol 71 no5 pp 1306ndash1312 2007
[27] B Tunca G Tezcan G Cecener et al ldquoOlea europea leafextract alters microRNA expression in human glioblastomacellsrdquo Journal of Cancer Research and Clinical Oncology vol 138no 11 pp 1831ndash1844 2012
[28] S Matsuoka M Huang and S J Elledge ldquoLinkage of ATM tocell cycle regulation by the Chk2 protein kinaserdquo Science vol282 no 5395 pp 1893ndash1897 1998
[29] C Frazer and P G Young ldquoPhosphorylation mediated regu-lation of Cdc25 activity localization and stabilityrdquo in ProteinPhosphorylation in Human Health C Huang Ed pp 395ndash436InTech 2012
[30] N Debili C Robin V Schiavon et al ldquoDifferent expression ofCD41 on human lymphoid andmyeloid progenitors from adultsand neonatesrdquo Blood vol 97 no 7 pp 2023ndash2030 2001
[31] H Nakajima ldquoRole of transcription factors in differentiationand reprogramming of hematopoietic cellsrdquo Keio Journal ofMedicine vol 60 no 2 pp 47ndash55 2011
[32] Y Fukuchi F Shibata M Ito et al ldquoComprehensive analysis ofmyeloid lineage conversion using mice expressing an inducibleform of CEBP120572rdquoThe EMBO Journal vol 25 no 14 pp 3398ndash3410 2006
[33] R A Shivdasani ldquoMolecular and transcriptional regulation ofmegakaryocyte differentiationrdquo Stem Cells vol 19 no 5 pp397ndash407 2001
[34] K Terui Y Takahashi J Kitazawa T TokiM Yokoyama and EIto ldquoExpression of transcription factors during megakaryocyticdifferentiation of CD34+ cells from human cord blood inducedby thrombopoietinrdquo Tohoku Journal of Experimental Medicinevol 192 no 4 pp 259ndash273 2000
[35] J A Trapani M Dawson V A Apostolidis and K A BrowneldquoGenomic organization of IFI16 an interferon-inducible genewhose expression is associated with human myeloid cell differ-entiation correlation of predicted protein domains with exonorganizationrdquo Immunogenetics vol 40 no 6 pp 415ndash424 1994
[36] M J Dawson N J Elwood R W Johnstone and J A TrapanildquoThe IFN-inducible nucleoprotein IFI 16 is expressed in cellsof the monocyte lineage but is rapidly and markedly down-regulated in other myeloid precursor populationsrdquo Journal ofLeukocyte Biology vol 64 no 4 pp 546ndash554 1998
[37] M Gariglio M De Andrea M Lembo et al ldquoThe murinehomolog of the HIN 200 family Ifi 204 is constitutivelyexpressed in myeloid cells and selectively induced in the
monocytemacrophage lineagerdquo Journal of Leukocyte Biologyvol 64 no 5 pp 608ndash614 1998
[38] A F Valledor F E Borras M Cullell-Young and A CeladaldquoTranscription factors that regulate monocytemacrophage dif-ferentiationrdquo Journal of Leukocyte Biology vol 63 no 4 pp405ndash417 1998
[39] K Krishnaraju B Hoffman and D A Liebermann ldquoEarlygrowth response gene 1 stimulates development of hematopoi-etic progenitor cells along the macrophage lineage at theexpense of the granulocyte and erythroid lineagesrdquo Blood vol97 no 5 pp 1298ndash1305 2001
[40] J Dauffy G Mouchiroud and R P Bourette ldquoThe interferon-inducible gene Ifi204 is transcriptionally activated in responseto M-CSF and its expression favors macrophage differentiationin myeloid progenitor cellsrdquo Journal of Leukocyte Biology vol79 no 1 pp 173ndash183 2006
[41] G Marziali E Perrotti R Ilari U Testa E M Coccia andA Battistini ldquoTranscriptional regulation of the ferritin heavy-chain gene the activity of the CCAAT binding factor NF-Y ismodulated in heme-treated Friend leukemia cells and duringmonocyte-to-macrophage differentiationrdquo Molecular and Cel-lular Biology vol 17 no 3 pp 1387ndash1395 1997
[42] C Shi X Zhang Z Chen et al ldquoIntegrin engagement regulatesmonocyte differentiation through the forkhead transcriptionfactor Foxp1rdquo Journal of Clinical Investigation vol 114 no 3 pp408ndash418 2004
[43] C Shi M Sakuma T Mooroka et al ldquoDown-regulation of theforkhead transcription factor Foxp1 is required for monocytedifferentiation andmacrophage functionrdquoBlood vol 112 no 12pp 4699ndash4711 2008
[44] D F Smith E Galkina K Ley and Y Huo ldquoGRO familychemokines are specialized for monocyte arrest from flowrdquoThe American Journal of Physiology Heart and CirculatoryPhysiology vol 289 no 5 pp H1976ndashH1984 2005
[45] J J Oppenheim O M Z Howard and E Goetzl ChemotacticFactors Neuropeptides and Other Ligands for Seven Transmem-brane Receptors 2000
[46] Y Terashima N Onai M Murai et al ldquoPivotal function forcytoplasmic protein FROUNT in CCR2-mediated monocytechemotaxisrdquo Nature Immunology vol 6 no 8 pp 827ndash8352005
[47] Y Lefkir M Malbouyres D Gotthardt et al ldquoInvolvement ofthe AP-1 adaptor complex in early steps of phagocytosis andmacropinocytosisrdquo Molecular Biology of the Cell vol 15 no 2pp 861ndash869 2004
[48] F Niedergang and P Chavrier ldquoSignaling and membranedynamics during phagocytosis many roads lead to the phago-somerdquo Current Opinion in Cell Biology vol 16 no 4 pp 422ndash428 2004
[49] D Cox D J Lee B M Dale J Calafat and S Green-berg ldquoA Rab11-containing rapidly recycling compartment inmacrophages that promotes phagocytosisrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 97 no 2 pp 680ndash685 2000
[50] O V Vieira C Bucci R E Harrison et al ldquoModulation of Rab5and Rab7 recruitment to phagosomes by phosphatidylinositol3-kinaserdquoMolecular andCellular Biology vol 23 no 7 pp 2501ndash2514 2003
[51] T Khurana J A Brzostowski and A R Kimmel ldquoARab21LIM-onlyCH-LIM complex regulates phagocytosis viaboth activating and inhibitory mechanismsrdquo The EMBO Jour-nal vol 24 no 13 pp 2254ndash2264 2005
16 Oxidative Medicine and Cellular Longevity
[52] S J Martin J G Bradley and T G Cotter ldquoHL-60 cellsinduced to differentiate towards neutrophils subsequently dievia apoptosisrdquo Clinical and Experimental Immunology vol 79no 3 pp 448ndash453 1990
[53] A Benito D Grillot G Nunez and J L Fernandez-Luna ldquoReg-ulation and function of Bcl-2 during differentiation-inducedcell death in HL-60 promyelocytic cellsrdquoThe American Journalof Pathology vol 146 no 2 pp 481ndash490 1995
[54] M Lamkanfi N Festjens W Declercq T V Berghe andP Vandenabeele ldquoCaspases in cell survival proliferation anddifferentiationrdquo Cell Death and Differentiation vol 14 no 1 pp44ndash55 2007
[55] O Sordet C Rebe S Plenchette et al ldquoSpecific involvement ofcaspases in the differentiation of monocytes into macrophagesrdquoBlood vol 100 no 13 pp 4446ndash4453 2002
[56] T-B Kang T Ben-Moshe E E Varfolomeev et al ldquoCaspase-8 serves both apoptotic and nonapoptotic rolesrdquo Journal ofImmunology vol 173 no 5 pp 2976ndash2984 2004
[57] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[58] J Cheng J Yang Y Xia M Karin and B Su ldquoSynergisticinteraction of MEK kinase 2 c-Jun N-terminal kinase (JNK)kinase 2 and JNK1 results in efficient and specific JNK1activationrdquo Molecular and Cellular Biology vol 20 no 7 pp2334ndash2342 2000
[59] J L Blank P Gerwins EM Elliott S Sather andG L JohnsonldquoMolecular cloning of mitogen-activated proteinERK kinasekinases (MEKK) 2 and 3 regulation of sequential phosphoryla-tion pathways involving mitogen-activated protein kinase andc-Jun kinaserdquo Journal of Biological Chemistry vol 271 no 10pp 5361ndash5368 1996
[60] S Papa F Zazzeroni C G Pham C Bubici and G FranzosoldquoLinking JNK signaling to NF-120581B a key to survivalrdquo Journal ofCell Science vol 117 no 22 pp 5197ndash5208 2004
[61] T Kadoya A Khurana M Tcherpakov et al ldquoJAMP a JUB N-terminal kinase 1 (JNK1)-associated membrane protein regu-lates duration of JNK activityrdquo Molecular and Cellular Biologyvol 25 no 19 pp 8619ndash8630 2005
[62] M Qi and E A Elion ldquoMAP kinase pathwaysrdquo Journal of CellScience vol 118 no 16 pp 3569ndash3572 2005
[63] Q Zhao and F S Lee ldquoMitogen-activated protein kinaseERKkinase kinases 2 and 3 activate nuclear factor-120581B through I120581Bkinase-120572 and I120581B kinase-120573rdquoThe Journal of Biological Chemistryvol 274 no 13 pp 8355ndash8358 1999
[64] H Namba V Saenko and S Yamashita ldquoNuclear factor-120581Bin thyroid carcinogenesis and progression a novel therapeutictarget for advanced thyroid cancerrdquo Arquivos Brasileiros deEndocrinologia e Metabologia vol 51 no 5 pp 843ndash851 2007
[65] B Feng S Cheng W S Pear and H-C Liou ldquoNF-kBinhibitor blocks B cell development at two checkpointsrdquoMedi-cal Immunology vol 3 article 1 2004
[66] S Gerondakis and U Siebenlist ldquoRoles of the NF-kappaBpathway in lymphocyte development and functionrdquoCold SpringHarbor Perspectives in Biology vol 2 no 5 Article ID a0001822010
[67] K P Bhat V Balasubramaniyan B Vaillant et al ldquoMesenchy-mal differentiation mediated by NF-kB promotes radiationresistance in glioblastomardquo Cancer Cell vol 24 no 3 pp 331ndash346 2013
Previous reports had indicated that olive leaf extract inducesapoptosis in the human leukemic cell lines HL-60 and Jurkatcells [13 21] In this study we give evidence that COLEexhibits its antileukemia effect by both inducing apoptosisand promoting differentiation of the multipotent humanleukemia K562 cells
Induction of differentiation as well as apoptosis has beenfrequently reported to be associatedwith a loss of proliferativecapacity of the cells It has been reported that olive leaf extractinhibits the proliferation of some cancer cell lines such asB16 melanoma cells HL-60 leukemia cells the breast cancerMCF-7 cells and the glioblastoma cells [12 25ndash27] Ourresults indicate that COLE inhibits the proliferation of K562cells in a dose dependent manner (Figure 2(a)) Howeverdespite the drastic decrease in cell proliferation the viabilityof cells treated with the highest concentration of COLE(150 120583gmL) remains relatively high (80) for up to 3 daysof treatment (Figure 3(b))
Since it is well established that the arrest of the cellcycle progression allows the cells to follow other processessuch as apoptosis and differentiation we then explored theeffect of COLE on the cell cycle distribution (Table 2) Oliveleaf extract has been shown to cause cell cycle arrest atG0G1 phase in both the breast cancer MCF-7 cells and themelanoma B16 cells [12 25] However the treatment timesapplied in both of these studies were limited to 48 h and 18 hrespectively In this study COLE treated cells were arrested atG0G1 on the 1st and 2ndday of treatment which is consistentwith the reported studies Prolonged incubation with COLEshowed an arrest of cell cycle at G2M phase (3rd and 4thday of incubation) The microarray analysis performed forthe 3rd day of treatment revealed a significant increase ofCHEK2 gene expression in COLE treated cells The encodedprotein regulates the cell cycle checkpoint arrest throughthe inhibition of the activity of CDC25A CDC25B andCDC25C [28] CDC25 proteins trigger the entry into mitosisat different points of the cell cycle by activating theCdk-cyclincomplexes CDC25A acts early in the cell cycle regulatingthe G1S transition whereas CDC25B and CDC25C act atG2M [29] Interestingly our results showed a decrease in theexpression of CDC25C gene in COLE treated cells associatedwith an increase in the expression of CDC25A which mayexplain the cell cycle arrest at G2M phase observed at the3rd day of treatment with COLE
Treatment with COLE increased the expression of CD14on the cell surface of treated cells indicating the differenti-ation of K562 cells toward the mono-macrophage lineage(Figure 5(a)) The increase in the percentage of cells positivefor CD11b (Figure 5(b)) confirms this hypothesis In factCD11b is expressed on both monocytes and granulocytesThus together with the increase of CD14 expression theexpression of CD11b suggests that treatment with COLEpromotes the commitment of K562 cells into the mono-cytemacrophage lineage at the expense of the granulocyticdifferentiationThe instant increase of the expression of CD41
on the 1st day of treatment and its absence during the fol-lowing days (Figure 5(c)) could be consistent with monocytedifferentiation instead of megakaryocyte differentiation sinceit has been reported that 120572IIb promoter which initiates thetranscription of CD41 is transcriptionally active in pluripo-tent myeloid progenitors in early stages of erythropoiesisand all along the megakaryocytic differentiation as well asto a lesser extent in the early stages of myelomonocyticdifferentiation and the late stages of erythropoiesis [30] Thisinstant increase could be also explained by the phenomenonof lineage conversion where some hematopoietic progenitorscan be converted into lineages other than their own bythe ectopic expression of some transcription factors [31]In this regard it was previously reported that commonlymphoid progenitors megakaryocyte erythroid progenitorsearly B cells and early T cells could be converted to thegranulocytemonocyte lineage by the induction of CEBP120572activity [32] Microarray results support the absence ofmegakaryocyte differentiation in the 3rd day of treatment Infact data showed a decrease in the expression of genes relatedtomegakaryopoiesis such as the essential regulator of plateletrelease NFE2 and its main candidate target gene TUBB1(encoding the megakaryocyte and platelet specific isoform of120573-tubulin (1205731)) [33] as well as the BACH2 transcription factorreported to be induced during megakaryocyte differentiation[34]
Interestingly results of microarray analysis (Table 3)showed the upregulation of several genes already reportedas key players in monocytemacrophage differentiation inthe COLE treated cells Among these genes the expressionof IFI16 was markedly increased Constitutively expressedin lymphoid cells IFI16 was shown to be associated withthe differentiation of human myeloid cells in response tointerferon gamma [35] Moreover IFI16 has been selectivelyexpressed during the differentiation of myeloid progenitorcells along monocytic lineage while it is downregulatedduring the commitment toward the granulocytic or erythroidlineage suggesting its potential involvement in the differenti-ation andmaturation of the monocytic lineage [36 37] Geneencoding for EGR-1 a positive modulator of macrophagedifferentiation was also significantly upregulated EGR-1 isknown to dictate development of myeloid progenitors alongthe macrophage lineage at the expense of development alongother lineages [38 39] Dauffy et al [40] reported thatIFI16-enforced expression in myeloid progenitors inducedthe expression of EGR-1 and conducted the macrophagedifferentiation in the absence of the macrophage colonystimulating factor (M-CSF) This suggests that EGR-1 couldbe regulated by the expression of IFI16 Treatment with COLEincreased as well the expression of NFYA encoding the Asubunit of the nuclear transcription factor Y whose synthesiswas previously reported during the terminal differentiationof monocytes to macrophages [38 41] The downregula-tion of FOXP1 in COLE treated K562 cells is consistentwith monocytemacrophage differentiation since it has beendemonstrated that the expression of FOXP1 was markedlydecreased in monocyte-induced differentiation of HL-60cells as well as in human peripheral blood monocytes and
Oxidative Medicine and Cellular Longevity 13
that the overexpression of FOXP1 prevented the morphologicmacrophage-like differentiation [42 43]
Monocytes and macrophages have several functionalcharacteristics including cell adhesion migration chemo-taxis and phagocytic activity Interestingly an enhancedexpression of the chemokines genes CXCL8 (IL8) CXCL2andCXCL3was found in COLE treated cells IL-8 productionhas been observed in vitro in a wide variety of cells includingmonocytes T lymphocytes neutrophils vascular endothe-lial cells dermal fibroblasts keratinocytes hepatocytes andhuman gastric cancer cells while the production of CXCL2andCXCL3was only described inmonocytes fibroblasts andendothelial cells [44 45] Another gene related to chemotaxiswas upregulated NUP85 which is an essential component ofthe nuclear pore complex and was reported to be involved inCCR2-mediated chemotaxis of monocytes [46]
COLE treatment also enhanced the expression of somegenes related to phagocytosis AP1G1 andRab proteins relatedgenes AP1G1 is a subunit of clathrin-associated adaptorprotein complex 1 whose role has been recently highlightedfor efficient phagocytosis at an early stage of phagosomeformation since it participates in the extension of the phago-cytic cup [47 48] The small GTPases Rab are key regulatorsof intracellular membrane trafficking Recent studies havedemonstrated that several RAB proteins play an importantrole in phagocytosis RAB proteins such as RAB11 Rab5 andRab 21 participate in the phagosome formation and matura-tion and are necessary for the phagocytic activity [48ndash51]
Consistently with the increase in Annexin V positivecells (Figure 4) treatment with COLE upregulated someproapoptotic genes such as CASP6 CASP8 DFFA and BIDand downregulated the apoptosis suppressor BCL2 as wellas the caspase inhibitors genes (Table 3) Olive leaf waspreviously shown to induce morphological changes thatare characteristic of apoptosis in leukemia cells but themechanisms underlying this effect were not well investi-gated [13 21] The increase in the proportion of apoptoticcells observed from the 4th day of treatment with COLEcould be explained by the programmed death of the fullydifferentiated cells (Figure 4) In fact this increase wasaccompanied by an important decrease in cell viability aswell as a diminution in CD14 expression (Figures 3(b) and5(a)) Elsewhere it is well established that in hematopoietictissues apoptosis is coupled to terminal differentiation ofmyeloid progenitors even though themechanism responsiblefor the activation of apoptosis during myeloid maturationis still poorly understood [52 53] It is also noteworthy tomention that several reports have highlighted the role ofcaspases in the terminal differentiation of a variety of celltypes [54] One example is the differentiation of human bloodmonocytes into macrophages a process that is blocked bysynthetic caspase inhibitors [55] It has been also reportedthat caspase 8 deletion in bone-marrow cells resulted in arrestof hemopoietic progenitor functioning and that its deletionin cells of the myelomonocytic lineage led to the arrest ofdifferentiation into macrophages and consequently to celldeath [56]
Among the differentially expressed genes in COLEtreated cells (Table 3) genes encoding the mitogen-activated
protein kinase kinase kinases (MAPKKKs) MAP3K2MAP3K5 and MAP3K7 were highly expressed whileMAP2K5 and MAPK14p38-120572 were downregulated TheMAPKKKs act at the upstream of the MAP kinase cascadewhich regulates important cellular processes such as geneexpression cell proliferation differentiation cell survivaland death The MAP3K5 activates the c-Jun N-terminalkinase (JNK) and p38 mitogen-activated protein kinases[57] while the MAP3K2 is reported to preferentially activateJNK [58 59] However the significant decrease in theMAPK14p38-120572 may suggest that the effect of COLE onK562 cells is likely to be mediated by the JNK MAPKsrather than the p38 MAPKs JNK plays an important rolein apoptosis pathways weather by activating proapoptoticor antiapoptotic genes Particularly it was shown thatprolonged but not transient JNK activation promotes thecell death [60] Our results showed that JKAMP gene whichencodes for a membrane-anchored regulator of the durationof JNK1 activity was upregulated (154 plusmn 0051) The elevatedexpression of JKAMP as reported by Kadoya et al [61]results in sustained JNK activity These findings may indicatethe potential role of JNK pathway in the COLE-inducedapoptosis in K562 cells
Thanks to their large regulatory domains some MAP-KKKs can interact with upstream regulators have functionssuch as ubiquitylation and be activated by relief of autoin-hibition and oligomerization [62] In this context MAP3K2has been reported to play an important role in NF-kappaBsignaling pathway by activating the I-kappaB kinases whichin turn phosphorylate the inhibitory factors of the nuclearfactor kappaB (NF-kappaB) resulting in their rapid ubiq-uitination and the liberation of NF-kappaB complex whichtranslocate from the cytoplasm to the nucleus [63] MAP3K7in association with other proteins could also be required forthe activation of NF-kappaB According to the GO analysis(Table S1) COLE treatment upregulated genes implicatedin NF-kappaB-related categories such as positive regulationof NF-kappaB transcription factor activity I-kappaB kinase-NF-kappaB cascade and its positive regulation and I-kappaBphosphorylation Genes encoding the signal transducers inthe NF-kappaB pathway TRAF5 and TRAF6 as well asthe activator of NF-kappaB signaling SNIP1 were upregu-lated Interestingly an increased expression of NFKB1 geneencoding for the precursor of the mature NF-kappaB p50was detected in COLE treated cells compared to untreatedK562 cells NFKB is well recognized as a central activator ofthe antiapoptotic cascades in response to external stimuli orintrinsic immune reactions and its prosurvival activity hasbeen implicated in a variety of biological processes [60 64]In the B cell lineage the activity of NF-120581B is required forthe completion of various developmental stages includingdifferentiation and the response of these cells to antigens [65]NF-kappaB also plays an important role in the survival anddevelopment of T cells [66] and has been recently reported tomediate the differentiation of mesenchymal stem cells [67]
Several studies have demonstrated a crosstalk betweenthe NF-kappaB and JNK pathways seen as an inhibitoryeffect of NF-kappaB on apoptosis through the suppressionof JNK activity and that this suppressive effect may occur
14 Oxidative Medicine and Cellular Longevity
through different mechanisms [60] This suggests that thebalance between JNK and NF-kappaB activities is crucial todetermine the cell fate survival or deathWehypothesize herethat COLE-induced apoptosis or differentiation is likely to bedetermined by the crosstalk between these two pathways inK562 cells Further investigations regarding protein expres-sion and activity are needed to confirm these findings
5 Conclusions
Our study demonstrates for the first time that olive leafextract exhibits an antileukemia effect on the human chronicmyeloid leukemia cells Olive leaf extract was shown toinhibit the proliferation of K562 cells by inducing cell cyclearrest apoptosis and differentiation toward the monocytelineage The induced expression of molecules involved indifferentiation toward the monocytemacrophage lineage aswell asmolecules related to apoptosis and cell cycle regulationconfirmed these findings and provides insights into themechanism by which olive leaf exhibits its antileukemiaeffect
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported by the JICA-JST Science andTechnology Research Partnership for Sustainable Develop-ment (SATREPS) Project ldquoValorization of Bio-Resources inSemi-Arid and Arid Land for Regional Developmentrdquo
References
[1] M Leszczyniecka T Roberts P Dent S Grant and P B FisherldquoDifferentiation therapy of human cancer basic science andclinical applicationsrdquo Pharmacology and Therapeutics vol 90no 2-3 pp 105ndash156 2001
[2] A I Spira and M A Carducci ldquoDifferentiation therapyrdquoCurrent Opinion in Pharmacology vol 3 no 4 pp 338ndash3432003
[3] S Nobili D Lippi E Witort et al ldquoNatural compounds forcancer treatment and preventionrdquo Pharmacological Researchvol 59 no 6 pp 365ndash378 2009
[4] S Ramos ldquoEffects of dietary flavonoids on apoptotic pathwaysrelated to cancer chemopreventionrdquo Journal of NutritionalBiochemistry vol 18 no 7 pp 427ndash442 2007
[5] V Hajhashemi G Vaseghi M Pourfarzam and A AbdollahildquoAre antioxidants helpful for disease preventionrdquo Research inPharmaceutical Sciences vol 5 no 1 pp 1ndash8 2010
[6] M Bouaziz and S Sayadi ldquoIsolation and evaluation of antiox-idants from leaves of a Tunisian cultivar olive treerdquo EuropeanJournal of Lipid Science andTechnology vol 107 no 7-8 pp 497ndash504 2005
[7] A P Pereira I C F R Ferreira F Marcelino et al ldquoPhenoliccompounds and antimicrobial activity of olive (Olea europaeaL Cv Cobrancosa) leavesrdquo Molecules vol 12 no 5 pp 1153ndash1162 2007
[8] L I Somova F O Shode P Ramnanan and A Nadar ldquoAnti-hypertensive antiatherosclerotic and antioxidant activity oftriterpenoids isolated from Olea europaea subspecies africanaleavesrdquo Journal of Ethnopharmacology vol 84 no 2-3 pp 299ndash305 2003
[9] V Micol N Caturla L Perez-Fons V Mas L Perez andA Estepa ldquoThe olive leaf extract exhibits antiviral activityagainst viral haemorrhagic septicaemia rhabdovirus (VHSV)rdquoAntiviral Research vol 66 no 2-3 pp 129ndash136 2005
[10] J Wainstein T Ganz M Boaz et al ldquoOlive leaf extract as ahypoglycemic agent in both human diabetic subjects and inratsrdquo Journal of Medicinal Food vol 15 no 7 pp 605ndash610 2012
[11] L Seddik TM Bah A AouesM Slimani andM BenderdourldquoElucidation of mechanisms underlying the protective effects ofolive leaf extract against lead-induced neurotoxicity in Wistarratsrdquo Journal of Toxicological Sciences vol 36 no 6 pp 797ndash809 2011
[12] Z Bouallagui J Han H Isoda and S Sayadi ldquoHydroxytyrosolrich extract from olive leaves modulates cell cycle progressionin MCF-7 human breast cancer cellsrdquo Food and ChemicalToxicology vol 49 no 1 pp 179ndash184 2011
[13] R Fares S Bazzi S E Baydoun and R M Abdel-Massih ldquoTheantioxidant and anti-proliferative activity of the Lebanese Oleaeuropaea extractrdquo Plant Foods for Human Nutrition vol 66 no1 pp 58ndash63 2011
[14] V Goulas V Exarchou A N Troganis et al ldquoPhytochemicalsin olive-leaf extracts and their antiproliferative activity againstcancer and endothelial cellsrdquo Molecular Nutrition and FoodResearch vol 53 no 5 pp 600ndash608 2009
[15] M de Bock E B Thorstensen J G Derraik H V HendersonP L Hofman and W S Cutfield ldquoHuman absorption andmetabolism of oleuropein and hydroxytyrosol ingested as olive(Olea europaea L) leaf extractrdquo Molecular Nutrition and FoodResearch vol 57 no 11 pp 2079ndash2085 2013
[16] R Briante M Patumi S Terenziani E Bismuto F Febbraioand R Nucci ldquoOlea europaea L leaf extract and derivativesantioxidant propertiesrdquo Journal of Agricultural and Food Chem-istry vol 50 no 17 pp 4934ndash4940 2002
[17] O Benavente-Garcıa J Castillo J Lorente A Ortuno and J ADel Rio ldquoAntioxidant activity of phenolics extracted from Oleaeuropaea L leavesrdquo Food Chemistry vol 68 no 4 pp 457ndash4622000
[18] S H Omar ldquoOleuropein in olive and its pharmacologicaleffectsrdquo Scientia Pharmaceutica vol 78 no 2 pp 133ndash154 2010
[19] H Jemai M Bouaziz I Fki A El Feki and S SayadildquoHypolipidimic and antioxidant activities of oleuropein and itshydrolysis derivative-rich extracts from Chemlali olive leavesrdquoChemico-Biological Interactions vol 176 no 2-3 pp 88ndash982008
[20] R Garcıa-Villalba M Larrosa S Possemiers F A Tomas-Barberan and J C Espın ldquoBioavailability of phenolics from anoleuropein-rich olive (Olea europaea) leaf extract and its acuteeffect on plasma antioxidant status comparison between pre-and postmenopausal womenrdquo European Journal of Nutrition2013
[21] J Anter Z Fernandez-Bedmar M Villatoro-Pulido et al ldquoApilot study on the DNA-protective cytotoxic and apoptosis-inducing properties of olive-leaf extractsrdquo Mutation Researchvol 723 no 2 pp 165ndash170 2011
[22] J A Sutherland A R Turner and P Mannoni ldquoDifferen-tiation of K562 leukemia cells along erythroid macrophage
Oxidative Medicine and Cellular Longevity 15
and megakaryocyte lineagesrdquo Journal of Biological ResponseModifiers vol 5 no 3 pp 250ndash262 1986
[23] K Schwenke H-P Peterson K-H Wangenheim and L EFeinendegen ldquoInduction of differentiation in erythroleukemicK562 cells by 120574-irradiationrdquo Leukemia Research vol 19 no 12pp 955ndash961 1995
[24] V N Enujiugha J Y Talabi S A Malomo and A I OlagunjuldquoDPPH radical scavenging capacity of phenolic extracts fromAfrican Yam Bean (Sphenostylis stenocarpa)rdquo Food and Nutri-tion Sciences vol 3 pp 7ndash13 2012
[25] S A Mijatovic G S Timotijevic D M Miljkovic et alldquoMultiple antimelanoma potential of dry olive leaf extractrdquoInternational Journal of Cancer vol 128 no 8 pp 1955ndash19652011
[26] L Abaza T P N Talorete P Yamada Y KuritaM Zarrouk andH Isoda ldquoInduction of growth inhibition and differentiation ofhuman leukemia HL-60 cells by a Tunisian Gerboui olive leafextractrdquo Bioscience Biotechnology and Biochemistry vol 71 no5 pp 1306ndash1312 2007
[27] B Tunca G Tezcan G Cecener et al ldquoOlea europea leafextract alters microRNA expression in human glioblastomacellsrdquo Journal of Cancer Research and Clinical Oncology vol 138no 11 pp 1831ndash1844 2012
[28] S Matsuoka M Huang and S J Elledge ldquoLinkage of ATM tocell cycle regulation by the Chk2 protein kinaserdquo Science vol282 no 5395 pp 1893ndash1897 1998
[29] C Frazer and P G Young ldquoPhosphorylation mediated regu-lation of Cdc25 activity localization and stabilityrdquo in ProteinPhosphorylation in Human Health C Huang Ed pp 395ndash436InTech 2012
[30] N Debili C Robin V Schiavon et al ldquoDifferent expression ofCD41 on human lymphoid andmyeloid progenitors from adultsand neonatesrdquo Blood vol 97 no 7 pp 2023ndash2030 2001
[31] H Nakajima ldquoRole of transcription factors in differentiationand reprogramming of hematopoietic cellsrdquo Keio Journal ofMedicine vol 60 no 2 pp 47ndash55 2011
[32] Y Fukuchi F Shibata M Ito et al ldquoComprehensive analysis ofmyeloid lineage conversion using mice expressing an inducibleform of CEBP120572rdquoThe EMBO Journal vol 25 no 14 pp 3398ndash3410 2006
[33] R A Shivdasani ldquoMolecular and transcriptional regulation ofmegakaryocyte differentiationrdquo Stem Cells vol 19 no 5 pp397ndash407 2001
[34] K Terui Y Takahashi J Kitazawa T TokiM Yokoyama and EIto ldquoExpression of transcription factors during megakaryocyticdifferentiation of CD34+ cells from human cord blood inducedby thrombopoietinrdquo Tohoku Journal of Experimental Medicinevol 192 no 4 pp 259ndash273 2000
[35] J A Trapani M Dawson V A Apostolidis and K A BrowneldquoGenomic organization of IFI16 an interferon-inducible genewhose expression is associated with human myeloid cell differ-entiation correlation of predicted protein domains with exonorganizationrdquo Immunogenetics vol 40 no 6 pp 415ndash424 1994
[36] M J Dawson N J Elwood R W Johnstone and J A TrapanildquoThe IFN-inducible nucleoprotein IFI 16 is expressed in cellsof the monocyte lineage but is rapidly and markedly down-regulated in other myeloid precursor populationsrdquo Journal ofLeukocyte Biology vol 64 no 4 pp 546ndash554 1998
[37] M Gariglio M De Andrea M Lembo et al ldquoThe murinehomolog of the HIN 200 family Ifi 204 is constitutivelyexpressed in myeloid cells and selectively induced in the
monocytemacrophage lineagerdquo Journal of Leukocyte Biologyvol 64 no 5 pp 608ndash614 1998
[38] A F Valledor F E Borras M Cullell-Young and A CeladaldquoTranscription factors that regulate monocytemacrophage dif-ferentiationrdquo Journal of Leukocyte Biology vol 63 no 4 pp405ndash417 1998
[39] K Krishnaraju B Hoffman and D A Liebermann ldquoEarlygrowth response gene 1 stimulates development of hematopoi-etic progenitor cells along the macrophage lineage at theexpense of the granulocyte and erythroid lineagesrdquo Blood vol97 no 5 pp 1298ndash1305 2001
[40] J Dauffy G Mouchiroud and R P Bourette ldquoThe interferon-inducible gene Ifi204 is transcriptionally activated in responseto M-CSF and its expression favors macrophage differentiationin myeloid progenitor cellsrdquo Journal of Leukocyte Biology vol79 no 1 pp 173ndash183 2006
[41] G Marziali E Perrotti R Ilari U Testa E M Coccia andA Battistini ldquoTranscriptional regulation of the ferritin heavy-chain gene the activity of the CCAAT binding factor NF-Y ismodulated in heme-treated Friend leukemia cells and duringmonocyte-to-macrophage differentiationrdquo Molecular and Cel-lular Biology vol 17 no 3 pp 1387ndash1395 1997
[42] C Shi X Zhang Z Chen et al ldquoIntegrin engagement regulatesmonocyte differentiation through the forkhead transcriptionfactor Foxp1rdquo Journal of Clinical Investigation vol 114 no 3 pp408ndash418 2004
[43] C Shi M Sakuma T Mooroka et al ldquoDown-regulation of theforkhead transcription factor Foxp1 is required for monocytedifferentiation andmacrophage functionrdquoBlood vol 112 no 12pp 4699ndash4711 2008
[44] D F Smith E Galkina K Ley and Y Huo ldquoGRO familychemokines are specialized for monocyte arrest from flowrdquoThe American Journal of Physiology Heart and CirculatoryPhysiology vol 289 no 5 pp H1976ndashH1984 2005
[45] J J Oppenheim O M Z Howard and E Goetzl ChemotacticFactors Neuropeptides and Other Ligands for Seven Transmem-brane Receptors 2000
[46] Y Terashima N Onai M Murai et al ldquoPivotal function forcytoplasmic protein FROUNT in CCR2-mediated monocytechemotaxisrdquo Nature Immunology vol 6 no 8 pp 827ndash8352005
[47] Y Lefkir M Malbouyres D Gotthardt et al ldquoInvolvement ofthe AP-1 adaptor complex in early steps of phagocytosis andmacropinocytosisrdquo Molecular Biology of the Cell vol 15 no 2pp 861ndash869 2004
[48] F Niedergang and P Chavrier ldquoSignaling and membranedynamics during phagocytosis many roads lead to the phago-somerdquo Current Opinion in Cell Biology vol 16 no 4 pp 422ndash428 2004
[49] D Cox D J Lee B M Dale J Calafat and S Green-berg ldquoA Rab11-containing rapidly recycling compartment inmacrophages that promotes phagocytosisrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 97 no 2 pp 680ndash685 2000
[50] O V Vieira C Bucci R E Harrison et al ldquoModulation of Rab5and Rab7 recruitment to phagosomes by phosphatidylinositol3-kinaserdquoMolecular andCellular Biology vol 23 no 7 pp 2501ndash2514 2003
[51] T Khurana J A Brzostowski and A R Kimmel ldquoARab21LIM-onlyCH-LIM complex regulates phagocytosis viaboth activating and inhibitory mechanismsrdquo The EMBO Jour-nal vol 24 no 13 pp 2254ndash2264 2005
16 Oxidative Medicine and Cellular Longevity
[52] S J Martin J G Bradley and T G Cotter ldquoHL-60 cellsinduced to differentiate towards neutrophils subsequently dievia apoptosisrdquo Clinical and Experimental Immunology vol 79no 3 pp 448ndash453 1990
[53] A Benito D Grillot G Nunez and J L Fernandez-Luna ldquoReg-ulation and function of Bcl-2 during differentiation-inducedcell death in HL-60 promyelocytic cellsrdquoThe American Journalof Pathology vol 146 no 2 pp 481ndash490 1995
[54] M Lamkanfi N Festjens W Declercq T V Berghe andP Vandenabeele ldquoCaspases in cell survival proliferation anddifferentiationrdquo Cell Death and Differentiation vol 14 no 1 pp44ndash55 2007
[55] O Sordet C Rebe S Plenchette et al ldquoSpecific involvement ofcaspases in the differentiation of monocytes into macrophagesrdquoBlood vol 100 no 13 pp 4446ndash4453 2002
[56] T-B Kang T Ben-Moshe E E Varfolomeev et al ldquoCaspase-8 serves both apoptotic and nonapoptotic rolesrdquo Journal ofImmunology vol 173 no 5 pp 2976ndash2984 2004
[57] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[58] J Cheng J Yang Y Xia M Karin and B Su ldquoSynergisticinteraction of MEK kinase 2 c-Jun N-terminal kinase (JNK)kinase 2 and JNK1 results in efficient and specific JNK1activationrdquo Molecular and Cellular Biology vol 20 no 7 pp2334ndash2342 2000
[59] J L Blank P Gerwins EM Elliott S Sather andG L JohnsonldquoMolecular cloning of mitogen-activated proteinERK kinasekinases (MEKK) 2 and 3 regulation of sequential phosphoryla-tion pathways involving mitogen-activated protein kinase andc-Jun kinaserdquo Journal of Biological Chemistry vol 271 no 10pp 5361ndash5368 1996
[60] S Papa F Zazzeroni C G Pham C Bubici and G FranzosoldquoLinking JNK signaling to NF-120581B a key to survivalrdquo Journal ofCell Science vol 117 no 22 pp 5197ndash5208 2004
[61] T Kadoya A Khurana M Tcherpakov et al ldquoJAMP a JUB N-terminal kinase 1 (JNK1)-associated membrane protein regu-lates duration of JNK activityrdquo Molecular and Cellular Biologyvol 25 no 19 pp 8619ndash8630 2005
[62] M Qi and E A Elion ldquoMAP kinase pathwaysrdquo Journal of CellScience vol 118 no 16 pp 3569ndash3572 2005
[63] Q Zhao and F S Lee ldquoMitogen-activated protein kinaseERKkinase kinases 2 and 3 activate nuclear factor-120581B through I120581Bkinase-120572 and I120581B kinase-120573rdquoThe Journal of Biological Chemistryvol 274 no 13 pp 8355ndash8358 1999
[64] H Namba V Saenko and S Yamashita ldquoNuclear factor-120581Bin thyroid carcinogenesis and progression a novel therapeutictarget for advanced thyroid cancerrdquo Arquivos Brasileiros deEndocrinologia e Metabologia vol 51 no 5 pp 843ndash851 2007
[65] B Feng S Cheng W S Pear and H-C Liou ldquoNF-kBinhibitor blocks B cell development at two checkpointsrdquoMedi-cal Immunology vol 3 article 1 2004
[66] S Gerondakis and U Siebenlist ldquoRoles of the NF-kappaBpathway in lymphocyte development and functionrdquoCold SpringHarbor Perspectives in Biology vol 2 no 5 Article ID a0001822010
[67] K P Bhat V Balasubramaniyan B Vaillant et al ldquoMesenchy-mal differentiation mediated by NF-kB promotes radiationresistance in glioblastomardquo Cancer Cell vol 24 no 3 pp 331ndash346 2013
that the overexpression of FOXP1 prevented the morphologicmacrophage-like differentiation [42 43]
Monocytes and macrophages have several functionalcharacteristics including cell adhesion migration chemo-taxis and phagocytic activity Interestingly an enhancedexpression of the chemokines genes CXCL8 (IL8) CXCL2andCXCL3was found in COLE treated cells IL-8 productionhas been observed in vitro in a wide variety of cells includingmonocytes T lymphocytes neutrophils vascular endothe-lial cells dermal fibroblasts keratinocytes hepatocytes andhuman gastric cancer cells while the production of CXCL2andCXCL3was only described inmonocytes fibroblasts andendothelial cells [44 45] Another gene related to chemotaxiswas upregulated NUP85 which is an essential component ofthe nuclear pore complex and was reported to be involved inCCR2-mediated chemotaxis of monocytes [46]
COLE treatment also enhanced the expression of somegenes related to phagocytosis AP1G1 andRab proteins relatedgenes AP1G1 is a subunit of clathrin-associated adaptorprotein complex 1 whose role has been recently highlightedfor efficient phagocytosis at an early stage of phagosomeformation since it participates in the extension of the phago-cytic cup [47 48] The small GTPases Rab are key regulatorsof intracellular membrane trafficking Recent studies havedemonstrated that several RAB proteins play an importantrole in phagocytosis RAB proteins such as RAB11 Rab5 andRab 21 participate in the phagosome formation and matura-tion and are necessary for the phagocytic activity [48ndash51]
Consistently with the increase in Annexin V positivecells (Figure 4) treatment with COLE upregulated someproapoptotic genes such as CASP6 CASP8 DFFA and BIDand downregulated the apoptosis suppressor BCL2 as wellas the caspase inhibitors genes (Table 3) Olive leaf waspreviously shown to induce morphological changes thatare characteristic of apoptosis in leukemia cells but themechanisms underlying this effect were not well investi-gated [13 21] The increase in the proportion of apoptoticcells observed from the 4th day of treatment with COLEcould be explained by the programmed death of the fullydifferentiated cells (Figure 4) In fact this increase wasaccompanied by an important decrease in cell viability aswell as a diminution in CD14 expression (Figures 3(b) and5(a)) Elsewhere it is well established that in hematopoietictissues apoptosis is coupled to terminal differentiation ofmyeloid progenitors even though themechanism responsiblefor the activation of apoptosis during myeloid maturationis still poorly understood [52 53] It is also noteworthy tomention that several reports have highlighted the role ofcaspases in the terminal differentiation of a variety of celltypes [54] One example is the differentiation of human bloodmonocytes into macrophages a process that is blocked bysynthetic caspase inhibitors [55] It has been also reportedthat caspase 8 deletion in bone-marrow cells resulted in arrestof hemopoietic progenitor functioning and that its deletionin cells of the myelomonocytic lineage led to the arrest ofdifferentiation into macrophages and consequently to celldeath [56]
Among the differentially expressed genes in COLEtreated cells (Table 3) genes encoding the mitogen-activated
protein kinase kinase kinases (MAPKKKs) MAP3K2MAP3K5 and MAP3K7 were highly expressed whileMAP2K5 and MAPK14p38-120572 were downregulated TheMAPKKKs act at the upstream of the MAP kinase cascadewhich regulates important cellular processes such as geneexpression cell proliferation differentiation cell survivaland death The MAP3K5 activates the c-Jun N-terminalkinase (JNK) and p38 mitogen-activated protein kinases[57] while the MAP3K2 is reported to preferentially activateJNK [58 59] However the significant decrease in theMAPK14p38-120572 may suggest that the effect of COLE onK562 cells is likely to be mediated by the JNK MAPKsrather than the p38 MAPKs JNK plays an important rolein apoptosis pathways weather by activating proapoptoticor antiapoptotic genes Particularly it was shown thatprolonged but not transient JNK activation promotes thecell death [60] Our results showed that JKAMP gene whichencodes for a membrane-anchored regulator of the durationof JNK1 activity was upregulated (154 plusmn 0051) The elevatedexpression of JKAMP as reported by Kadoya et al [61]results in sustained JNK activity These findings may indicatethe potential role of JNK pathway in the COLE-inducedapoptosis in K562 cells
Thanks to their large regulatory domains some MAP-KKKs can interact with upstream regulators have functionssuch as ubiquitylation and be activated by relief of autoin-hibition and oligomerization [62] In this context MAP3K2has been reported to play an important role in NF-kappaBsignaling pathway by activating the I-kappaB kinases whichin turn phosphorylate the inhibitory factors of the nuclearfactor kappaB (NF-kappaB) resulting in their rapid ubiq-uitination and the liberation of NF-kappaB complex whichtranslocate from the cytoplasm to the nucleus [63] MAP3K7in association with other proteins could also be required forthe activation of NF-kappaB According to the GO analysis(Table S1) COLE treatment upregulated genes implicatedin NF-kappaB-related categories such as positive regulationof NF-kappaB transcription factor activity I-kappaB kinase-NF-kappaB cascade and its positive regulation and I-kappaBphosphorylation Genes encoding the signal transducers inthe NF-kappaB pathway TRAF5 and TRAF6 as well asthe activator of NF-kappaB signaling SNIP1 were upregu-lated Interestingly an increased expression of NFKB1 geneencoding for the precursor of the mature NF-kappaB p50was detected in COLE treated cells compared to untreatedK562 cells NFKB is well recognized as a central activator ofthe antiapoptotic cascades in response to external stimuli orintrinsic immune reactions and its prosurvival activity hasbeen implicated in a variety of biological processes [60 64]In the B cell lineage the activity of NF-120581B is required forthe completion of various developmental stages includingdifferentiation and the response of these cells to antigens [65]NF-kappaB also plays an important role in the survival anddevelopment of T cells [66] and has been recently reported tomediate the differentiation of mesenchymal stem cells [67]
Several studies have demonstrated a crosstalk betweenthe NF-kappaB and JNK pathways seen as an inhibitoryeffect of NF-kappaB on apoptosis through the suppressionof JNK activity and that this suppressive effect may occur
14 Oxidative Medicine and Cellular Longevity
through different mechanisms [60] This suggests that thebalance between JNK and NF-kappaB activities is crucial todetermine the cell fate survival or deathWehypothesize herethat COLE-induced apoptosis or differentiation is likely to bedetermined by the crosstalk between these two pathways inK562 cells Further investigations regarding protein expres-sion and activity are needed to confirm these findings
5 Conclusions
Our study demonstrates for the first time that olive leafextract exhibits an antileukemia effect on the human chronicmyeloid leukemia cells Olive leaf extract was shown toinhibit the proliferation of K562 cells by inducing cell cyclearrest apoptosis and differentiation toward the monocytelineage The induced expression of molecules involved indifferentiation toward the monocytemacrophage lineage aswell asmolecules related to apoptosis and cell cycle regulationconfirmed these findings and provides insights into themechanism by which olive leaf exhibits its antileukemiaeffect
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported by the JICA-JST Science andTechnology Research Partnership for Sustainable Develop-ment (SATREPS) Project ldquoValorization of Bio-Resources inSemi-Arid and Arid Land for Regional Developmentrdquo
References
[1] M Leszczyniecka T Roberts P Dent S Grant and P B FisherldquoDifferentiation therapy of human cancer basic science andclinical applicationsrdquo Pharmacology and Therapeutics vol 90no 2-3 pp 105ndash156 2001
[2] A I Spira and M A Carducci ldquoDifferentiation therapyrdquoCurrent Opinion in Pharmacology vol 3 no 4 pp 338ndash3432003
[3] S Nobili D Lippi E Witort et al ldquoNatural compounds forcancer treatment and preventionrdquo Pharmacological Researchvol 59 no 6 pp 365ndash378 2009
[4] S Ramos ldquoEffects of dietary flavonoids on apoptotic pathwaysrelated to cancer chemopreventionrdquo Journal of NutritionalBiochemistry vol 18 no 7 pp 427ndash442 2007
[5] V Hajhashemi G Vaseghi M Pourfarzam and A AbdollahildquoAre antioxidants helpful for disease preventionrdquo Research inPharmaceutical Sciences vol 5 no 1 pp 1ndash8 2010
[6] M Bouaziz and S Sayadi ldquoIsolation and evaluation of antiox-idants from leaves of a Tunisian cultivar olive treerdquo EuropeanJournal of Lipid Science andTechnology vol 107 no 7-8 pp 497ndash504 2005
[7] A P Pereira I C F R Ferreira F Marcelino et al ldquoPhenoliccompounds and antimicrobial activity of olive (Olea europaeaL Cv Cobrancosa) leavesrdquo Molecules vol 12 no 5 pp 1153ndash1162 2007
[8] L I Somova F O Shode P Ramnanan and A Nadar ldquoAnti-hypertensive antiatherosclerotic and antioxidant activity oftriterpenoids isolated from Olea europaea subspecies africanaleavesrdquo Journal of Ethnopharmacology vol 84 no 2-3 pp 299ndash305 2003
[9] V Micol N Caturla L Perez-Fons V Mas L Perez andA Estepa ldquoThe olive leaf extract exhibits antiviral activityagainst viral haemorrhagic septicaemia rhabdovirus (VHSV)rdquoAntiviral Research vol 66 no 2-3 pp 129ndash136 2005
[10] J Wainstein T Ganz M Boaz et al ldquoOlive leaf extract as ahypoglycemic agent in both human diabetic subjects and inratsrdquo Journal of Medicinal Food vol 15 no 7 pp 605ndash610 2012
[11] L Seddik TM Bah A AouesM Slimani andM BenderdourldquoElucidation of mechanisms underlying the protective effects ofolive leaf extract against lead-induced neurotoxicity in Wistarratsrdquo Journal of Toxicological Sciences vol 36 no 6 pp 797ndash809 2011
[12] Z Bouallagui J Han H Isoda and S Sayadi ldquoHydroxytyrosolrich extract from olive leaves modulates cell cycle progressionin MCF-7 human breast cancer cellsrdquo Food and ChemicalToxicology vol 49 no 1 pp 179ndash184 2011
[13] R Fares S Bazzi S E Baydoun and R M Abdel-Massih ldquoTheantioxidant and anti-proliferative activity of the Lebanese Oleaeuropaea extractrdquo Plant Foods for Human Nutrition vol 66 no1 pp 58ndash63 2011
[14] V Goulas V Exarchou A N Troganis et al ldquoPhytochemicalsin olive-leaf extracts and their antiproliferative activity againstcancer and endothelial cellsrdquo Molecular Nutrition and FoodResearch vol 53 no 5 pp 600ndash608 2009
[15] M de Bock E B Thorstensen J G Derraik H V HendersonP L Hofman and W S Cutfield ldquoHuman absorption andmetabolism of oleuropein and hydroxytyrosol ingested as olive(Olea europaea L) leaf extractrdquo Molecular Nutrition and FoodResearch vol 57 no 11 pp 2079ndash2085 2013
[16] R Briante M Patumi S Terenziani E Bismuto F Febbraioand R Nucci ldquoOlea europaea L leaf extract and derivativesantioxidant propertiesrdquo Journal of Agricultural and Food Chem-istry vol 50 no 17 pp 4934ndash4940 2002
[17] O Benavente-Garcıa J Castillo J Lorente A Ortuno and J ADel Rio ldquoAntioxidant activity of phenolics extracted from Oleaeuropaea L leavesrdquo Food Chemistry vol 68 no 4 pp 457ndash4622000
[18] S H Omar ldquoOleuropein in olive and its pharmacologicaleffectsrdquo Scientia Pharmaceutica vol 78 no 2 pp 133ndash154 2010
[19] H Jemai M Bouaziz I Fki A El Feki and S SayadildquoHypolipidimic and antioxidant activities of oleuropein and itshydrolysis derivative-rich extracts from Chemlali olive leavesrdquoChemico-Biological Interactions vol 176 no 2-3 pp 88ndash982008
[20] R Garcıa-Villalba M Larrosa S Possemiers F A Tomas-Barberan and J C Espın ldquoBioavailability of phenolics from anoleuropein-rich olive (Olea europaea) leaf extract and its acuteeffect on plasma antioxidant status comparison between pre-and postmenopausal womenrdquo European Journal of Nutrition2013
[21] J Anter Z Fernandez-Bedmar M Villatoro-Pulido et al ldquoApilot study on the DNA-protective cytotoxic and apoptosis-inducing properties of olive-leaf extractsrdquo Mutation Researchvol 723 no 2 pp 165ndash170 2011
[22] J A Sutherland A R Turner and P Mannoni ldquoDifferen-tiation of K562 leukemia cells along erythroid macrophage
Oxidative Medicine and Cellular Longevity 15
and megakaryocyte lineagesrdquo Journal of Biological ResponseModifiers vol 5 no 3 pp 250ndash262 1986
[23] K Schwenke H-P Peterson K-H Wangenheim and L EFeinendegen ldquoInduction of differentiation in erythroleukemicK562 cells by 120574-irradiationrdquo Leukemia Research vol 19 no 12pp 955ndash961 1995
[24] V N Enujiugha J Y Talabi S A Malomo and A I OlagunjuldquoDPPH radical scavenging capacity of phenolic extracts fromAfrican Yam Bean (Sphenostylis stenocarpa)rdquo Food and Nutri-tion Sciences vol 3 pp 7ndash13 2012
[25] S A Mijatovic G S Timotijevic D M Miljkovic et alldquoMultiple antimelanoma potential of dry olive leaf extractrdquoInternational Journal of Cancer vol 128 no 8 pp 1955ndash19652011
[26] L Abaza T P N Talorete P Yamada Y KuritaM Zarrouk andH Isoda ldquoInduction of growth inhibition and differentiation ofhuman leukemia HL-60 cells by a Tunisian Gerboui olive leafextractrdquo Bioscience Biotechnology and Biochemistry vol 71 no5 pp 1306ndash1312 2007
[27] B Tunca G Tezcan G Cecener et al ldquoOlea europea leafextract alters microRNA expression in human glioblastomacellsrdquo Journal of Cancer Research and Clinical Oncology vol 138no 11 pp 1831ndash1844 2012
[28] S Matsuoka M Huang and S J Elledge ldquoLinkage of ATM tocell cycle regulation by the Chk2 protein kinaserdquo Science vol282 no 5395 pp 1893ndash1897 1998
[29] C Frazer and P G Young ldquoPhosphorylation mediated regu-lation of Cdc25 activity localization and stabilityrdquo in ProteinPhosphorylation in Human Health C Huang Ed pp 395ndash436InTech 2012
[30] N Debili C Robin V Schiavon et al ldquoDifferent expression ofCD41 on human lymphoid andmyeloid progenitors from adultsand neonatesrdquo Blood vol 97 no 7 pp 2023ndash2030 2001
[31] H Nakajima ldquoRole of transcription factors in differentiationand reprogramming of hematopoietic cellsrdquo Keio Journal ofMedicine vol 60 no 2 pp 47ndash55 2011
[32] Y Fukuchi F Shibata M Ito et al ldquoComprehensive analysis ofmyeloid lineage conversion using mice expressing an inducibleform of CEBP120572rdquoThe EMBO Journal vol 25 no 14 pp 3398ndash3410 2006
[33] R A Shivdasani ldquoMolecular and transcriptional regulation ofmegakaryocyte differentiationrdquo Stem Cells vol 19 no 5 pp397ndash407 2001
[34] K Terui Y Takahashi J Kitazawa T TokiM Yokoyama and EIto ldquoExpression of transcription factors during megakaryocyticdifferentiation of CD34+ cells from human cord blood inducedby thrombopoietinrdquo Tohoku Journal of Experimental Medicinevol 192 no 4 pp 259ndash273 2000
[35] J A Trapani M Dawson V A Apostolidis and K A BrowneldquoGenomic organization of IFI16 an interferon-inducible genewhose expression is associated with human myeloid cell differ-entiation correlation of predicted protein domains with exonorganizationrdquo Immunogenetics vol 40 no 6 pp 415ndash424 1994
[36] M J Dawson N J Elwood R W Johnstone and J A TrapanildquoThe IFN-inducible nucleoprotein IFI 16 is expressed in cellsof the monocyte lineage but is rapidly and markedly down-regulated in other myeloid precursor populationsrdquo Journal ofLeukocyte Biology vol 64 no 4 pp 546ndash554 1998
[37] M Gariglio M De Andrea M Lembo et al ldquoThe murinehomolog of the HIN 200 family Ifi 204 is constitutivelyexpressed in myeloid cells and selectively induced in the
monocytemacrophage lineagerdquo Journal of Leukocyte Biologyvol 64 no 5 pp 608ndash614 1998
[38] A F Valledor F E Borras M Cullell-Young and A CeladaldquoTranscription factors that regulate monocytemacrophage dif-ferentiationrdquo Journal of Leukocyte Biology vol 63 no 4 pp405ndash417 1998
[39] K Krishnaraju B Hoffman and D A Liebermann ldquoEarlygrowth response gene 1 stimulates development of hematopoi-etic progenitor cells along the macrophage lineage at theexpense of the granulocyte and erythroid lineagesrdquo Blood vol97 no 5 pp 1298ndash1305 2001
[40] J Dauffy G Mouchiroud and R P Bourette ldquoThe interferon-inducible gene Ifi204 is transcriptionally activated in responseto M-CSF and its expression favors macrophage differentiationin myeloid progenitor cellsrdquo Journal of Leukocyte Biology vol79 no 1 pp 173ndash183 2006
[41] G Marziali E Perrotti R Ilari U Testa E M Coccia andA Battistini ldquoTranscriptional regulation of the ferritin heavy-chain gene the activity of the CCAAT binding factor NF-Y ismodulated in heme-treated Friend leukemia cells and duringmonocyte-to-macrophage differentiationrdquo Molecular and Cel-lular Biology vol 17 no 3 pp 1387ndash1395 1997
[42] C Shi X Zhang Z Chen et al ldquoIntegrin engagement regulatesmonocyte differentiation through the forkhead transcriptionfactor Foxp1rdquo Journal of Clinical Investigation vol 114 no 3 pp408ndash418 2004
[43] C Shi M Sakuma T Mooroka et al ldquoDown-regulation of theforkhead transcription factor Foxp1 is required for monocytedifferentiation andmacrophage functionrdquoBlood vol 112 no 12pp 4699ndash4711 2008
[44] D F Smith E Galkina K Ley and Y Huo ldquoGRO familychemokines are specialized for monocyte arrest from flowrdquoThe American Journal of Physiology Heart and CirculatoryPhysiology vol 289 no 5 pp H1976ndashH1984 2005
[45] J J Oppenheim O M Z Howard and E Goetzl ChemotacticFactors Neuropeptides and Other Ligands for Seven Transmem-brane Receptors 2000
[46] Y Terashima N Onai M Murai et al ldquoPivotal function forcytoplasmic protein FROUNT in CCR2-mediated monocytechemotaxisrdquo Nature Immunology vol 6 no 8 pp 827ndash8352005
[47] Y Lefkir M Malbouyres D Gotthardt et al ldquoInvolvement ofthe AP-1 adaptor complex in early steps of phagocytosis andmacropinocytosisrdquo Molecular Biology of the Cell vol 15 no 2pp 861ndash869 2004
[48] F Niedergang and P Chavrier ldquoSignaling and membranedynamics during phagocytosis many roads lead to the phago-somerdquo Current Opinion in Cell Biology vol 16 no 4 pp 422ndash428 2004
[49] D Cox D J Lee B M Dale J Calafat and S Green-berg ldquoA Rab11-containing rapidly recycling compartment inmacrophages that promotes phagocytosisrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 97 no 2 pp 680ndash685 2000
[50] O V Vieira C Bucci R E Harrison et al ldquoModulation of Rab5and Rab7 recruitment to phagosomes by phosphatidylinositol3-kinaserdquoMolecular andCellular Biology vol 23 no 7 pp 2501ndash2514 2003
[51] T Khurana J A Brzostowski and A R Kimmel ldquoARab21LIM-onlyCH-LIM complex regulates phagocytosis viaboth activating and inhibitory mechanismsrdquo The EMBO Jour-nal vol 24 no 13 pp 2254ndash2264 2005
16 Oxidative Medicine and Cellular Longevity
[52] S J Martin J G Bradley and T G Cotter ldquoHL-60 cellsinduced to differentiate towards neutrophils subsequently dievia apoptosisrdquo Clinical and Experimental Immunology vol 79no 3 pp 448ndash453 1990
[53] A Benito D Grillot G Nunez and J L Fernandez-Luna ldquoReg-ulation and function of Bcl-2 during differentiation-inducedcell death in HL-60 promyelocytic cellsrdquoThe American Journalof Pathology vol 146 no 2 pp 481ndash490 1995
[54] M Lamkanfi N Festjens W Declercq T V Berghe andP Vandenabeele ldquoCaspases in cell survival proliferation anddifferentiationrdquo Cell Death and Differentiation vol 14 no 1 pp44ndash55 2007
[55] O Sordet C Rebe S Plenchette et al ldquoSpecific involvement ofcaspases in the differentiation of monocytes into macrophagesrdquoBlood vol 100 no 13 pp 4446ndash4453 2002
[56] T-B Kang T Ben-Moshe E E Varfolomeev et al ldquoCaspase-8 serves both apoptotic and nonapoptotic rolesrdquo Journal ofImmunology vol 173 no 5 pp 2976ndash2984 2004
[57] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[58] J Cheng J Yang Y Xia M Karin and B Su ldquoSynergisticinteraction of MEK kinase 2 c-Jun N-terminal kinase (JNK)kinase 2 and JNK1 results in efficient and specific JNK1activationrdquo Molecular and Cellular Biology vol 20 no 7 pp2334ndash2342 2000
[59] J L Blank P Gerwins EM Elliott S Sather andG L JohnsonldquoMolecular cloning of mitogen-activated proteinERK kinasekinases (MEKK) 2 and 3 regulation of sequential phosphoryla-tion pathways involving mitogen-activated protein kinase andc-Jun kinaserdquo Journal of Biological Chemistry vol 271 no 10pp 5361ndash5368 1996
[60] S Papa F Zazzeroni C G Pham C Bubici and G FranzosoldquoLinking JNK signaling to NF-120581B a key to survivalrdquo Journal ofCell Science vol 117 no 22 pp 5197ndash5208 2004
[61] T Kadoya A Khurana M Tcherpakov et al ldquoJAMP a JUB N-terminal kinase 1 (JNK1)-associated membrane protein regu-lates duration of JNK activityrdquo Molecular and Cellular Biologyvol 25 no 19 pp 8619ndash8630 2005
[62] M Qi and E A Elion ldquoMAP kinase pathwaysrdquo Journal of CellScience vol 118 no 16 pp 3569ndash3572 2005
[63] Q Zhao and F S Lee ldquoMitogen-activated protein kinaseERKkinase kinases 2 and 3 activate nuclear factor-120581B through I120581Bkinase-120572 and I120581B kinase-120573rdquoThe Journal of Biological Chemistryvol 274 no 13 pp 8355ndash8358 1999
[64] H Namba V Saenko and S Yamashita ldquoNuclear factor-120581Bin thyroid carcinogenesis and progression a novel therapeutictarget for advanced thyroid cancerrdquo Arquivos Brasileiros deEndocrinologia e Metabologia vol 51 no 5 pp 843ndash851 2007
[65] B Feng S Cheng W S Pear and H-C Liou ldquoNF-kBinhibitor blocks B cell development at two checkpointsrdquoMedi-cal Immunology vol 3 article 1 2004
[66] S Gerondakis and U Siebenlist ldquoRoles of the NF-kappaBpathway in lymphocyte development and functionrdquoCold SpringHarbor Perspectives in Biology vol 2 no 5 Article ID a0001822010
[67] K P Bhat V Balasubramaniyan B Vaillant et al ldquoMesenchy-mal differentiation mediated by NF-kB promotes radiationresistance in glioblastomardquo Cancer Cell vol 24 no 3 pp 331ndash346 2013
through different mechanisms [60] This suggests that thebalance between JNK and NF-kappaB activities is crucial todetermine the cell fate survival or deathWehypothesize herethat COLE-induced apoptosis or differentiation is likely to bedetermined by the crosstalk between these two pathways inK562 cells Further investigations regarding protein expres-sion and activity are needed to confirm these findings
5 Conclusions
Our study demonstrates for the first time that olive leafextract exhibits an antileukemia effect on the human chronicmyeloid leukemia cells Olive leaf extract was shown toinhibit the proliferation of K562 cells by inducing cell cyclearrest apoptosis and differentiation toward the monocytelineage The induced expression of molecules involved indifferentiation toward the monocytemacrophage lineage aswell asmolecules related to apoptosis and cell cycle regulationconfirmed these findings and provides insights into themechanism by which olive leaf exhibits its antileukemiaeffect
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported by the JICA-JST Science andTechnology Research Partnership for Sustainable Develop-ment (SATREPS) Project ldquoValorization of Bio-Resources inSemi-Arid and Arid Land for Regional Developmentrdquo
References
[1] M Leszczyniecka T Roberts P Dent S Grant and P B FisherldquoDifferentiation therapy of human cancer basic science andclinical applicationsrdquo Pharmacology and Therapeutics vol 90no 2-3 pp 105ndash156 2001
[2] A I Spira and M A Carducci ldquoDifferentiation therapyrdquoCurrent Opinion in Pharmacology vol 3 no 4 pp 338ndash3432003
[3] S Nobili D Lippi E Witort et al ldquoNatural compounds forcancer treatment and preventionrdquo Pharmacological Researchvol 59 no 6 pp 365ndash378 2009
[4] S Ramos ldquoEffects of dietary flavonoids on apoptotic pathwaysrelated to cancer chemopreventionrdquo Journal of NutritionalBiochemistry vol 18 no 7 pp 427ndash442 2007
[5] V Hajhashemi G Vaseghi M Pourfarzam and A AbdollahildquoAre antioxidants helpful for disease preventionrdquo Research inPharmaceutical Sciences vol 5 no 1 pp 1ndash8 2010
[6] M Bouaziz and S Sayadi ldquoIsolation and evaluation of antiox-idants from leaves of a Tunisian cultivar olive treerdquo EuropeanJournal of Lipid Science andTechnology vol 107 no 7-8 pp 497ndash504 2005
[7] A P Pereira I C F R Ferreira F Marcelino et al ldquoPhenoliccompounds and antimicrobial activity of olive (Olea europaeaL Cv Cobrancosa) leavesrdquo Molecules vol 12 no 5 pp 1153ndash1162 2007
[8] L I Somova F O Shode P Ramnanan and A Nadar ldquoAnti-hypertensive antiatherosclerotic and antioxidant activity oftriterpenoids isolated from Olea europaea subspecies africanaleavesrdquo Journal of Ethnopharmacology vol 84 no 2-3 pp 299ndash305 2003
[9] V Micol N Caturla L Perez-Fons V Mas L Perez andA Estepa ldquoThe olive leaf extract exhibits antiviral activityagainst viral haemorrhagic septicaemia rhabdovirus (VHSV)rdquoAntiviral Research vol 66 no 2-3 pp 129ndash136 2005
[10] J Wainstein T Ganz M Boaz et al ldquoOlive leaf extract as ahypoglycemic agent in both human diabetic subjects and inratsrdquo Journal of Medicinal Food vol 15 no 7 pp 605ndash610 2012
[11] L Seddik TM Bah A AouesM Slimani andM BenderdourldquoElucidation of mechanisms underlying the protective effects ofolive leaf extract against lead-induced neurotoxicity in Wistarratsrdquo Journal of Toxicological Sciences vol 36 no 6 pp 797ndash809 2011
[12] Z Bouallagui J Han H Isoda and S Sayadi ldquoHydroxytyrosolrich extract from olive leaves modulates cell cycle progressionin MCF-7 human breast cancer cellsrdquo Food and ChemicalToxicology vol 49 no 1 pp 179ndash184 2011
[13] R Fares S Bazzi S E Baydoun and R M Abdel-Massih ldquoTheantioxidant and anti-proliferative activity of the Lebanese Oleaeuropaea extractrdquo Plant Foods for Human Nutrition vol 66 no1 pp 58ndash63 2011
[14] V Goulas V Exarchou A N Troganis et al ldquoPhytochemicalsin olive-leaf extracts and their antiproliferative activity againstcancer and endothelial cellsrdquo Molecular Nutrition and FoodResearch vol 53 no 5 pp 600ndash608 2009
[15] M de Bock E B Thorstensen J G Derraik H V HendersonP L Hofman and W S Cutfield ldquoHuman absorption andmetabolism of oleuropein and hydroxytyrosol ingested as olive(Olea europaea L) leaf extractrdquo Molecular Nutrition and FoodResearch vol 57 no 11 pp 2079ndash2085 2013
[16] R Briante M Patumi S Terenziani E Bismuto F Febbraioand R Nucci ldquoOlea europaea L leaf extract and derivativesantioxidant propertiesrdquo Journal of Agricultural and Food Chem-istry vol 50 no 17 pp 4934ndash4940 2002
[17] O Benavente-Garcıa J Castillo J Lorente A Ortuno and J ADel Rio ldquoAntioxidant activity of phenolics extracted from Oleaeuropaea L leavesrdquo Food Chemistry vol 68 no 4 pp 457ndash4622000
[18] S H Omar ldquoOleuropein in olive and its pharmacologicaleffectsrdquo Scientia Pharmaceutica vol 78 no 2 pp 133ndash154 2010
[19] H Jemai M Bouaziz I Fki A El Feki and S SayadildquoHypolipidimic and antioxidant activities of oleuropein and itshydrolysis derivative-rich extracts from Chemlali olive leavesrdquoChemico-Biological Interactions vol 176 no 2-3 pp 88ndash982008
[20] R Garcıa-Villalba M Larrosa S Possemiers F A Tomas-Barberan and J C Espın ldquoBioavailability of phenolics from anoleuropein-rich olive (Olea europaea) leaf extract and its acuteeffect on plasma antioxidant status comparison between pre-and postmenopausal womenrdquo European Journal of Nutrition2013
[21] J Anter Z Fernandez-Bedmar M Villatoro-Pulido et al ldquoApilot study on the DNA-protective cytotoxic and apoptosis-inducing properties of olive-leaf extractsrdquo Mutation Researchvol 723 no 2 pp 165ndash170 2011
[22] J A Sutherland A R Turner and P Mannoni ldquoDifferen-tiation of K562 leukemia cells along erythroid macrophage
Oxidative Medicine and Cellular Longevity 15
and megakaryocyte lineagesrdquo Journal of Biological ResponseModifiers vol 5 no 3 pp 250ndash262 1986
[23] K Schwenke H-P Peterson K-H Wangenheim and L EFeinendegen ldquoInduction of differentiation in erythroleukemicK562 cells by 120574-irradiationrdquo Leukemia Research vol 19 no 12pp 955ndash961 1995
[24] V N Enujiugha J Y Talabi S A Malomo and A I OlagunjuldquoDPPH radical scavenging capacity of phenolic extracts fromAfrican Yam Bean (Sphenostylis stenocarpa)rdquo Food and Nutri-tion Sciences vol 3 pp 7ndash13 2012
[25] S A Mijatovic G S Timotijevic D M Miljkovic et alldquoMultiple antimelanoma potential of dry olive leaf extractrdquoInternational Journal of Cancer vol 128 no 8 pp 1955ndash19652011
[26] L Abaza T P N Talorete P Yamada Y KuritaM Zarrouk andH Isoda ldquoInduction of growth inhibition and differentiation ofhuman leukemia HL-60 cells by a Tunisian Gerboui olive leafextractrdquo Bioscience Biotechnology and Biochemistry vol 71 no5 pp 1306ndash1312 2007
[27] B Tunca G Tezcan G Cecener et al ldquoOlea europea leafextract alters microRNA expression in human glioblastomacellsrdquo Journal of Cancer Research and Clinical Oncology vol 138no 11 pp 1831ndash1844 2012
[28] S Matsuoka M Huang and S J Elledge ldquoLinkage of ATM tocell cycle regulation by the Chk2 protein kinaserdquo Science vol282 no 5395 pp 1893ndash1897 1998
[29] C Frazer and P G Young ldquoPhosphorylation mediated regu-lation of Cdc25 activity localization and stabilityrdquo in ProteinPhosphorylation in Human Health C Huang Ed pp 395ndash436InTech 2012
[30] N Debili C Robin V Schiavon et al ldquoDifferent expression ofCD41 on human lymphoid andmyeloid progenitors from adultsand neonatesrdquo Blood vol 97 no 7 pp 2023ndash2030 2001
[31] H Nakajima ldquoRole of transcription factors in differentiationand reprogramming of hematopoietic cellsrdquo Keio Journal ofMedicine vol 60 no 2 pp 47ndash55 2011
[32] Y Fukuchi F Shibata M Ito et al ldquoComprehensive analysis ofmyeloid lineage conversion using mice expressing an inducibleform of CEBP120572rdquoThe EMBO Journal vol 25 no 14 pp 3398ndash3410 2006
[33] R A Shivdasani ldquoMolecular and transcriptional regulation ofmegakaryocyte differentiationrdquo Stem Cells vol 19 no 5 pp397ndash407 2001
[34] K Terui Y Takahashi J Kitazawa T TokiM Yokoyama and EIto ldquoExpression of transcription factors during megakaryocyticdifferentiation of CD34+ cells from human cord blood inducedby thrombopoietinrdquo Tohoku Journal of Experimental Medicinevol 192 no 4 pp 259ndash273 2000
[35] J A Trapani M Dawson V A Apostolidis and K A BrowneldquoGenomic organization of IFI16 an interferon-inducible genewhose expression is associated with human myeloid cell differ-entiation correlation of predicted protein domains with exonorganizationrdquo Immunogenetics vol 40 no 6 pp 415ndash424 1994
[36] M J Dawson N J Elwood R W Johnstone and J A TrapanildquoThe IFN-inducible nucleoprotein IFI 16 is expressed in cellsof the monocyte lineage but is rapidly and markedly down-regulated in other myeloid precursor populationsrdquo Journal ofLeukocyte Biology vol 64 no 4 pp 546ndash554 1998
[37] M Gariglio M De Andrea M Lembo et al ldquoThe murinehomolog of the HIN 200 family Ifi 204 is constitutivelyexpressed in myeloid cells and selectively induced in the
monocytemacrophage lineagerdquo Journal of Leukocyte Biologyvol 64 no 5 pp 608ndash614 1998
[38] A F Valledor F E Borras M Cullell-Young and A CeladaldquoTranscription factors that regulate monocytemacrophage dif-ferentiationrdquo Journal of Leukocyte Biology vol 63 no 4 pp405ndash417 1998
[39] K Krishnaraju B Hoffman and D A Liebermann ldquoEarlygrowth response gene 1 stimulates development of hematopoi-etic progenitor cells along the macrophage lineage at theexpense of the granulocyte and erythroid lineagesrdquo Blood vol97 no 5 pp 1298ndash1305 2001
[40] J Dauffy G Mouchiroud and R P Bourette ldquoThe interferon-inducible gene Ifi204 is transcriptionally activated in responseto M-CSF and its expression favors macrophage differentiationin myeloid progenitor cellsrdquo Journal of Leukocyte Biology vol79 no 1 pp 173ndash183 2006
[41] G Marziali E Perrotti R Ilari U Testa E M Coccia andA Battistini ldquoTranscriptional regulation of the ferritin heavy-chain gene the activity of the CCAAT binding factor NF-Y ismodulated in heme-treated Friend leukemia cells and duringmonocyte-to-macrophage differentiationrdquo Molecular and Cel-lular Biology vol 17 no 3 pp 1387ndash1395 1997
[42] C Shi X Zhang Z Chen et al ldquoIntegrin engagement regulatesmonocyte differentiation through the forkhead transcriptionfactor Foxp1rdquo Journal of Clinical Investigation vol 114 no 3 pp408ndash418 2004
[43] C Shi M Sakuma T Mooroka et al ldquoDown-regulation of theforkhead transcription factor Foxp1 is required for monocytedifferentiation andmacrophage functionrdquoBlood vol 112 no 12pp 4699ndash4711 2008
[44] D F Smith E Galkina K Ley and Y Huo ldquoGRO familychemokines are specialized for monocyte arrest from flowrdquoThe American Journal of Physiology Heart and CirculatoryPhysiology vol 289 no 5 pp H1976ndashH1984 2005
[45] J J Oppenheim O M Z Howard and E Goetzl ChemotacticFactors Neuropeptides and Other Ligands for Seven Transmem-brane Receptors 2000
[46] Y Terashima N Onai M Murai et al ldquoPivotal function forcytoplasmic protein FROUNT in CCR2-mediated monocytechemotaxisrdquo Nature Immunology vol 6 no 8 pp 827ndash8352005
[47] Y Lefkir M Malbouyres D Gotthardt et al ldquoInvolvement ofthe AP-1 adaptor complex in early steps of phagocytosis andmacropinocytosisrdquo Molecular Biology of the Cell vol 15 no 2pp 861ndash869 2004
[48] F Niedergang and P Chavrier ldquoSignaling and membranedynamics during phagocytosis many roads lead to the phago-somerdquo Current Opinion in Cell Biology vol 16 no 4 pp 422ndash428 2004
[49] D Cox D J Lee B M Dale J Calafat and S Green-berg ldquoA Rab11-containing rapidly recycling compartment inmacrophages that promotes phagocytosisrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 97 no 2 pp 680ndash685 2000
[50] O V Vieira C Bucci R E Harrison et al ldquoModulation of Rab5and Rab7 recruitment to phagosomes by phosphatidylinositol3-kinaserdquoMolecular andCellular Biology vol 23 no 7 pp 2501ndash2514 2003
[51] T Khurana J A Brzostowski and A R Kimmel ldquoARab21LIM-onlyCH-LIM complex regulates phagocytosis viaboth activating and inhibitory mechanismsrdquo The EMBO Jour-nal vol 24 no 13 pp 2254ndash2264 2005
16 Oxidative Medicine and Cellular Longevity
[52] S J Martin J G Bradley and T G Cotter ldquoHL-60 cellsinduced to differentiate towards neutrophils subsequently dievia apoptosisrdquo Clinical and Experimental Immunology vol 79no 3 pp 448ndash453 1990
[53] A Benito D Grillot G Nunez and J L Fernandez-Luna ldquoReg-ulation and function of Bcl-2 during differentiation-inducedcell death in HL-60 promyelocytic cellsrdquoThe American Journalof Pathology vol 146 no 2 pp 481ndash490 1995
[54] M Lamkanfi N Festjens W Declercq T V Berghe andP Vandenabeele ldquoCaspases in cell survival proliferation anddifferentiationrdquo Cell Death and Differentiation vol 14 no 1 pp44ndash55 2007
[55] O Sordet C Rebe S Plenchette et al ldquoSpecific involvement ofcaspases in the differentiation of monocytes into macrophagesrdquoBlood vol 100 no 13 pp 4446ndash4453 2002
[56] T-B Kang T Ben-Moshe E E Varfolomeev et al ldquoCaspase-8 serves both apoptotic and nonapoptotic rolesrdquo Journal ofImmunology vol 173 no 5 pp 2976ndash2984 2004
[57] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[58] J Cheng J Yang Y Xia M Karin and B Su ldquoSynergisticinteraction of MEK kinase 2 c-Jun N-terminal kinase (JNK)kinase 2 and JNK1 results in efficient and specific JNK1activationrdquo Molecular and Cellular Biology vol 20 no 7 pp2334ndash2342 2000
[59] J L Blank P Gerwins EM Elliott S Sather andG L JohnsonldquoMolecular cloning of mitogen-activated proteinERK kinasekinases (MEKK) 2 and 3 regulation of sequential phosphoryla-tion pathways involving mitogen-activated protein kinase andc-Jun kinaserdquo Journal of Biological Chemistry vol 271 no 10pp 5361ndash5368 1996
[60] S Papa F Zazzeroni C G Pham C Bubici and G FranzosoldquoLinking JNK signaling to NF-120581B a key to survivalrdquo Journal ofCell Science vol 117 no 22 pp 5197ndash5208 2004
[61] T Kadoya A Khurana M Tcherpakov et al ldquoJAMP a JUB N-terminal kinase 1 (JNK1)-associated membrane protein regu-lates duration of JNK activityrdquo Molecular and Cellular Biologyvol 25 no 19 pp 8619ndash8630 2005
[62] M Qi and E A Elion ldquoMAP kinase pathwaysrdquo Journal of CellScience vol 118 no 16 pp 3569ndash3572 2005
[63] Q Zhao and F S Lee ldquoMitogen-activated protein kinaseERKkinase kinases 2 and 3 activate nuclear factor-120581B through I120581Bkinase-120572 and I120581B kinase-120573rdquoThe Journal of Biological Chemistryvol 274 no 13 pp 8355ndash8358 1999
[64] H Namba V Saenko and S Yamashita ldquoNuclear factor-120581Bin thyroid carcinogenesis and progression a novel therapeutictarget for advanced thyroid cancerrdquo Arquivos Brasileiros deEndocrinologia e Metabologia vol 51 no 5 pp 843ndash851 2007
[65] B Feng S Cheng W S Pear and H-C Liou ldquoNF-kBinhibitor blocks B cell development at two checkpointsrdquoMedi-cal Immunology vol 3 article 1 2004
[66] S Gerondakis and U Siebenlist ldquoRoles of the NF-kappaBpathway in lymphocyte development and functionrdquoCold SpringHarbor Perspectives in Biology vol 2 no 5 Article ID a0001822010
[67] K P Bhat V Balasubramaniyan B Vaillant et al ldquoMesenchy-mal differentiation mediated by NF-kB promotes radiationresistance in glioblastomardquo Cancer Cell vol 24 no 3 pp 331ndash346 2013
and megakaryocyte lineagesrdquo Journal of Biological ResponseModifiers vol 5 no 3 pp 250ndash262 1986
[23] K Schwenke H-P Peterson K-H Wangenheim and L EFeinendegen ldquoInduction of differentiation in erythroleukemicK562 cells by 120574-irradiationrdquo Leukemia Research vol 19 no 12pp 955ndash961 1995
[24] V N Enujiugha J Y Talabi S A Malomo and A I OlagunjuldquoDPPH radical scavenging capacity of phenolic extracts fromAfrican Yam Bean (Sphenostylis stenocarpa)rdquo Food and Nutri-tion Sciences vol 3 pp 7ndash13 2012
[25] S A Mijatovic G S Timotijevic D M Miljkovic et alldquoMultiple antimelanoma potential of dry olive leaf extractrdquoInternational Journal of Cancer vol 128 no 8 pp 1955ndash19652011
[26] L Abaza T P N Talorete P Yamada Y KuritaM Zarrouk andH Isoda ldquoInduction of growth inhibition and differentiation ofhuman leukemia HL-60 cells by a Tunisian Gerboui olive leafextractrdquo Bioscience Biotechnology and Biochemistry vol 71 no5 pp 1306ndash1312 2007
[27] B Tunca G Tezcan G Cecener et al ldquoOlea europea leafextract alters microRNA expression in human glioblastomacellsrdquo Journal of Cancer Research and Clinical Oncology vol 138no 11 pp 1831ndash1844 2012
[28] S Matsuoka M Huang and S J Elledge ldquoLinkage of ATM tocell cycle regulation by the Chk2 protein kinaserdquo Science vol282 no 5395 pp 1893ndash1897 1998
[29] C Frazer and P G Young ldquoPhosphorylation mediated regu-lation of Cdc25 activity localization and stabilityrdquo in ProteinPhosphorylation in Human Health C Huang Ed pp 395ndash436InTech 2012
[30] N Debili C Robin V Schiavon et al ldquoDifferent expression ofCD41 on human lymphoid andmyeloid progenitors from adultsand neonatesrdquo Blood vol 97 no 7 pp 2023ndash2030 2001
[31] H Nakajima ldquoRole of transcription factors in differentiationand reprogramming of hematopoietic cellsrdquo Keio Journal ofMedicine vol 60 no 2 pp 47ndash55 2011
[32] Y Fukuchi F Shibata M Ito et al ldquoComprehensive analysis ofmyeloid lineage conversion using mice expressing an inducibleform of CEBP120572rdquoThe EMBO Journal vol 25 no 14 pp 3398ndash3410 2006
[33] R A Shivdasani ldquoMolecular and transcriptional regulation ofmegakaryocyte differentiationrdquo Stem Cells vol 19 no 5 pp397ndash407 2001
[34] K Terui Y Takahashi J Kitazawa T TokiM Yokoyama and EIto ldquoExpression of transcription factors during megakaryocyticdifferentiation of CD34+ cells from human cord blood inducedby thrombopoietinrdquo Tohoku Journal of Experimental Medicinevol 192 no 4 pp 259ndash273 2000
[35] J A Trapani M Dawson V A Apostolidis and K A BrowneldquoGenomic organization of IFI16 an interferon-inducible genewhose expression is associated with human myeloid cell differ-entiation correlation of predicted protein domains with exonorganizationrdquo Immunogenetics vol 40 no 6 pp 415ndash424 1994
[36] M J Dawson N J Elwood R W Johnstone and J A TrapanildquoThe IFN-inducible nucleoprotein IFI 16 is expressed in cellsof the monocyte lineage but is rapidly and markedly down-regulated in other myeloid precursor populationsrdquo Journal ofLeukocyte Biology vol 64 no 4 pp 546ndash554 1998
[37] M Gariglio M De Andrea M Lembo et al ldquoThe murinehomolog of the HIN 200 family Ifi 204 is constitutivelyexpressed in myeloid cells and selectively induced in the
monocytemacrophage lineagerdquo Journal of Leukocyte Biologyvol 64 no 5 pp 608ndash614 1998
[38] A F Valledor F E Borras M Cullell-Young and A CeladaldquoTranscription factors that regulate monocytemacrophage dif-ferentiationrdquo Journal of Leukocyte Biology vol 63 no 4 pp405ndash417 1998
[39] K Krishnaraju B Hoffman and D A Liebermann ldquoEarlygrowth response gene 1 stimulates development of hematopoi-etic progenitor cells along the macrophage lineage at theexpense of the granulocyte and erythroid lineagesrdquo Blood vol97 no 5 pp 1298ndash1305 2001
[40] J Dauffy G Mouchiroud and R P Bourette ldquoThe interferon-inducible gene Ifi204 is transcriptionally activated in responseto M-CSF and its expression favors macrophage differentiationin myeloid progenitor cellsrdquo Journal of Leukocyte Biology vol79 no 1 pp 173ndash183 2006
[41] G Marziali E Perrotti R Ilari U Testa E M Coccia andA Battistini ldquoTranscriptional regulation of the ferritin heavy-chain gene the activity of the CCAAT binding factor NF-Y ismodulated in heme-treated Friend leukemia cells and duringmonocyte-to-macrophage differentiationrdquo Molecular and Cel-lular Biology vol 17 no 3 pp 1387ndash1395 1997
[42] C Shi X Zhang Z Chen et al ldquoIntegrin engagement regulatesmonocyte differentiation through the forkhead transcriptionfactor Foxp1rdquo Journal of Clinical Investigation vol 114 no 3 pp408ndash418 2004
[43] C Shi M Sakuma T Mooroka et al ldquoDown-regulation of theforkhead transcription factor Foxp1 is required for monocytedifferentiation andmacrophage functionrdquoBlood vol 112 no 12pp 4699ndash4711 2008
[44] D F Smith E Galkina K Ley and Y Huo ldquoGRO familychemokines are specialized for monocyte arrest from flowrdquoThe American Journal of Physiology Heart and CirculatoryPhysiology vol 289 no 5 pp H1976ndashH1984 2005
[45] J J Oppenheim O M Z Howard and E Goetzl ChemotacticFactors Neuropeptides and Other Ligands for Seven Transmem-brane Receptors 2000
[46] Y Terashima N Onai M Murai et al ldquoPivotal function forcytoplasmic protein FROUNT in CCR2-mediated monocytechemotaxisrdquo Nature Immunology vol 6 no 8 pp 827ndash8352005
[47] Y Lefkir M Malbouyres D Gotthardt et al ldquoInvolvement ofthe AP-1 adaptor complex in early steps of phagocytosis andmacropinocytosisrdquo Molecular Biology of the Cell vol 15 no 2pp 861ndash869 2004
[48] F Niedergang and P Chavrier ldquoSignaling and membranedynamics during phagocytosis many roads lead to the phago-somerdquo Current Opinion in Cell Biology vol 16 no 4 pp 422ndash428 2004
[49] D Cox D J Lee B M Dale J Calafat and S Green-berg ldquoA Rab11-containing rapidly recycling compartment inmacrophages that promotes phagocytosisrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 97 no 2 pp 680ndash685 2000
[50] O V Vieira C Bucci R E Harrison et al ldquoModulation of Rab5and Rab7 recruitment to phagosomes by phosphatidylinositol3-kinaserdquoMolecular andCellular Biology vol 23 no 7 pp 2501ndash2514 2003
[51] T Khurana J A Brzostowski and A R Kimmel ldquoARab21LIM-onlyCH-LIM complex regulates phagocytosis viaboth activating and inhibitory mechanismsrdquo The EMBO Jour-nal vol 24 no 13 pp 2254ndash2264 2005
16 Oxidative Medicine and Cellular Longevity
[52] S J Martin J G Bradley and T G Cotter ldquoHL-60 cellsinduced to differentiate towards neutrophils subsequently dievia apoptosisrdquo Clinical and Experimental Immunology vol 79no 3 pp 448ndash453 1990
[53] A Benito D Grillot G Nunez and J L Fernandez-Luna ldquoReg-ulation and function of Bcl-2 during differentiation-inducedcell death in HL-60 promyelocytic cellsrdquoThe American Journalof Pathology vol 146 no 2 pp 481ndash490 1995
[54] M Lamkanfi N Festjens W Declercq T V Berghe andP Vandenabeele ldquoCaspases in cell survival proliferation anddifferentiationrdquo Cell Death and Differentiation vol 14 no 1 pp44ndash55 2007
[55] O Sordet C Rebe S Plenchette et al ldquoSpecific involvement ofcaspases in the differentiation of monocytes into macrophagesrdquoBlood vol 100 no 13 pp 4446ndash4453 2002
[56] T-B Kang T Ben-Moshe E E Varfolomeev et al ldquoCaspase-8 serves both apoptotic and nonapoptotic rolesrdquo Journal ofImmunology vol 173 no 5 pp 2976ndash2984 2004
[57] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[58] J Cheng J Yang Y Xia M Karin and B Su ldquoSynergisticinteraction of MEK kinase 2 c-Jun N-terminal kinase (JNK)kinase 2 and JNK1 results in efficient and specific JNK1activationrdquo Molecular and Cellular Biology vol 20 no 7 pp2334ndash2342 2000
[59] J L Blank P Gerwins EM Elliott S Sather andG L JohnsonldquoMolecular cloning of mitogen-activated proteinERK kinasekinases (MEKK) 2 and 3 regulation of sequential phosphoryla-tion pathways involving mitogen-activated protein kinase andc-Jun kinaserdquo Journal of Biological Chemistry vol 271 no 10pp 5361ndash5368 1996
[60] S Papa F Zazzeroni C G Pham C Bubici and G FranzosoldquoLinking JNK signaling to NF-120581B a key to survivalrdquo Journal ofCell Science vol 117 no 22 pp 5197ndash5208 2004
[61] T Kadoya A Khurana M Tcherpakov et al ldquoJAMP a JUB N-terminal kinase 1 (JNK1)-associated membrane protein regu-lates duration of JNK activityrdquo Molecular and Cellular Biologyvol 25 no 19 pp 8619ndash8630 2005
[62] M Qi and E A Elion ldquoMAP kinase pathwaysrdquo Journal of CellScience vol 118 no 16 pp 3569ndash3572 2005
[63] Q Zhao and F S Lee ldquoMitogen-activated protein kinaseERKkinase kinases 2 and 3 activate nuclear factor-120581B through I120581Bkinase-120572 and I120581B kinase-120573rdquoThe Journal of Biological Chemistryvol 274 no 13 pp 8355ndash8358 1999
[64] H Namba V Saenko and S Yamashita ldquoNuclear factor-120581Bin thyroid carcinogenesis and progression a novel therapeutictarget for advanced thyroid cancerrdquo Arquivos Brasileiros deEndocrinologia e Metabologia vol 51 no 5 pp 843ndash851 2007
[65] B Feng S Cheng W S Pear and H-C Liou ldquoNF-kBinhibitor blocks B cell development at two checkpointsrdquoMedi-cal Immunology vol 3 article 1 2004
[66] S Gerondakis and U Siebenlist ldquoRoles of the NF-kappaBpathway in lymphocyte development and functionrdquoCold SpringHarbor Perspectives in Biology vol 2 no 5 Article ID a0001822010
[67] K P Bhat V Balasubramaniyan B Vaillant et al ldquoMesenchy-mal differentiation mediated by NF-kB promotes radiationresistance in glioblastomardquo Cancer Cell vol 24 no 3 pp 331ndash346 2013
[52] S J Martin J G Bradley and T G Cotter ldquoHL-60 cellsinduced to differentiate towards neutrophils subsequently dievia apoptosisrdquo Clinical and Experimental Immunology vol 79no 3 pp 448ndash453 1990
[53] A Benito D Grillot G Nunez and J L Fernandez-Luna ldquoReg-ulation and function of Bcl-2 during differentiation-inducedcell death in HL-60 promyelocytic cellsrdquoThe American Journalof Pathology vol 146 no 2 pp 481ndash490 1995
[54] M Lamkanfi N Festjens W Declercq T V Berghe andP Vandenabeele ldquoCaspases in cell survival proliferation anddifferentiationrdquo Cell Death and Differentiation vol 14 no 1 pp44ndash55 2007
[55] O Sordet C Rebe S Plenchette et al ldquoSpecific involvement ofcaspases in the differentiation of monocytes into macrophagesrdquoBlood vol 100 no 13 pp 4446ndash4453 2002
[56] T-B Kang T Ben-Moshe E E Varfolomeev et al ldquoCaspase-8 serves both apoptotic and nonapoptotic rolesrdquo Journal ofImmunology vol 173 no 5 pp 2976ndash2984 2004
[57] H Ichijo E Nishida K Irie et al ldquoInduction of apoptosis byASK1 a mammalian MAPKKK that activates SAPKJNK andp38 signaling pathwaysrdquo Science vol 275 no 5296 pp 90ndash941997
[58] J Cheng J Yang Y Xia M Karin and B Su ldquoSynergisticinteraction of MEK kinase 2 c-Jun N-terminal kinase (JNK)kinase 2 and JNK1 results in efficient and specific JNK1activationrdquo Molecular and Cellular Biology vol 20 no 7 pp2334ndash2342 2000
[59] J L Blank P Gerwins EM Elliott S Sather andG L JohnsonldquoMolecular cloning of mitogen-activated proteinERK kinasekinases (MEKK) 2 and 3 regulation of sequential phosphoryla-tion pathways involving mitogen-activated protein kinase andc-Jun kinaserdquo Journal of Biological Chemistry vol 271 no 10pp 5361ndash5368 1996
[60] S Papa F Zazzeroni C G Pham C Bubici and G FranzosoldquoLinking JNK signaling to NF-120581B a key to survivalrdquo Journal ofCell Science vol 117 no 22 pp 5197ndash5208 2004
[61] T Kadoya A Khurana M Tcherpakov et al ldquoJAMP a JUB N-terminal kinase 1 (JNK1)-associated membrane protein regu-lates duration of JNK activityrdquo Molecular and Cellular Biologyvol 25 no 19 pp 8619ndash8630 2005
[62] M Qi and E A Elion ldquoMAP kinase pathwaysrdquo Journal of CellScience vol 118 no 16 pp 3569ndash3572 2005
[63] Q Zhao and F S Lee ldquoMitogen-activated protein kinaseERKkinase kinases 2 and 3 activate nuclear factor-120581B through I120581Bkinase-120572 and I120581B kinase-120573rdquoThe Journal of Biological Chemistryvol 274 no 13 pp 8355ndash8358 1999
[64] H Namba V Saenko and S Yamashita ldquoNuclear factor-120581Bin thyroid carcinogenesis and progression a novel therapeutictarget for advanced thyroid cancerrdquo Arquivos Brasileiros deEndocrinologia e Metabologia vol 51 no 5 pp 843ndash851 2007
[65] B Feng S Cheng W S Pear and H-C Liou ldquoNF-kBinhibitor blocks B cell development at two checkpointsrdquoMedi-cal Immunology vol 3 article 1 2004
[66] S Gerondakis and U Siebenlist ldquoRoles of the NF-kappaBpathway in lymphocyte development and functionrdquoCold SpringHarbor Perspectives in Biology vol 2 no 5 Article ID a0001822010
[67] K P Bhat V Balasubramaniyan B Vaillant et al ldquoMesenchy-mal differentiation mediated by NF-kB promotes radiationresistance in glioblastomardquo Cancer Cell vol 24 no 3 pp 331ndash346 2013