CILIARY PROCESS OF FOOD-COLLECTING I N TURRITELLA. 453 26. On a Ciliary Process of Food-collecting in the Gastropod Turritella communis Rieso. By ALASTAIRGRAHAM, M.A., B.Sc. (Depart- ment of Zoology, Birkbeck College, University of London) *. [Received February 19, 1938 : Read May 24, 1938.1 (Text-figures 1-3.) CONTENTS. E. Page Introduction ....................................... 453 Ciliary Currents in tho Mantlo-cavity ................. 453 Ciliary Currents in tho Aliniontary Canal .............. 458 Discussion ........................................ 460 Summary ......................................... 463 References to Literature ............................ 463 INTRODUCTION. It has been known that a crystalline style and style-sac occur in relation with the stomach and intestine of Turritella communis since Randles described it in 1902. Yonge (1932) showed that in all molluscs possessing these struc- tures the feeding mechanism was either ciliary or “ the slow but continuous rasping action of the radula,” both methods giving the approximately con- tinuous stream of food for the digestion of which the continuous liberation of small quantities of enzyme from the style is an adaptation, and he also recorded that Turritella fed by ciliary means. Whilst making a comparative study of the structure of the anterior end of the alimentary canal of prosobranch molluscs I independently made the same observation, and Professor Yonge has kindly left it to me to give the details of the collection and transport of the food by cilia in the mantle-cavity and alimentary tract of that mollusc. I should like to thank Professor H. G. Jackson for his interest in the work and Professor C. M. Yonge for his valuable criticisms and for permitting me to make use of his unpublished work. CILIARY CURRENTS IN THE MANTLE-UAVITY. The mantle of Turritella (m., text-fig. 1) arches over the head and foot of the animal and comes into actual contact with the mid-dorsal region of the head. The aperture leading to the mantle-cavity is thus split into right (e.) and left (i.) portions, that on the animal’s left side being slightly the larger of the two. The anterior tip of the ctenidium (c.) may usually be seen crushed between the animal’s head and the overlying mantle, marking the division clearly. A current enters the mantle-cavity through the left portion of the aperture and an exhalent current passes through that on the right side of the The edge of the mantle bears two series of tentacles : one, set along the complete length of the edge, projecting dorsally and curling over the lip of * Communicated by Prof. H. G. JACKSON, D.Sc., F.Z.S. body.
Transcript
CILIARY PROCESS OF FOOD-COLLECTING IN TURRITELLA. 453
26. On a Ciliary Process of Food-collecting in the Gastropod Turritella communis Rieso. By ALASTAIR GRAHAM, M.A., B.Sc. (Depart- ment of Zoology, Birkbeck College, University of London) *.
INTRODUCTION. It has been known that a crystalline style and style-sac occur in relation
with the stomach and intestine of Turritella communis since Randles described it in 1902. Yonge (1932) showed that in all molluscs possessing these struc- tures the feeding mechanism was either ciliary or “ the slow but continuous rasping action of the radula,” both methods giving the approximately con- tinuous stream of food for the digestion of which the continuous liberation of small quantities of enzyme from the style is an adaptation, and he also recorded that Turritella fed by ciliary means. Whilst making a comparative study of the structure of the anterior end of the alimentary canal of prosobranch molluscs I independently made the same observation, and Professor Yonge has kindly left it to me to give the details of the collection and transport of the food by cilia in the mantle-cavity and alimentary tract of that mollusc. I should like to thank Professor H. G. Jackson for his interest in the work and Professor C . M. Yonge for his valuable criticisms and for permitting me to make use of his unpublished work.
CILIARY CURRENTS IN THE MANTLE-UAVITY. The mantle of Turritella (m., text-fig. 1) arches over the head and foot of
the animal and comes into actual contact with the mid-dorsal region of the head. The aperture leading to the mantle-cavity is thus split into right (e . ) and left (i.) portions, that on the animal’s left side being slightly the larger of the two. The anterior tip of the ctenidium (c.) may usually be seen crushed between the animal’s head and the overlying mantle, marking the division clearly. A current enters the mantle-cavity through the left portion of the aperture and an exhalent current passes through that on the right side of the
The edge of the mantle bears two series of tentacles : one, set along the complete length of the edge, projecting dorsally and curling over the lip of
* Communicated by Prof. H. G. JACKSON, D.Sc., F.Z.S.
body.
454 M R . ALASTAIR GRAHAM ON A CILIARY PROCESS OF
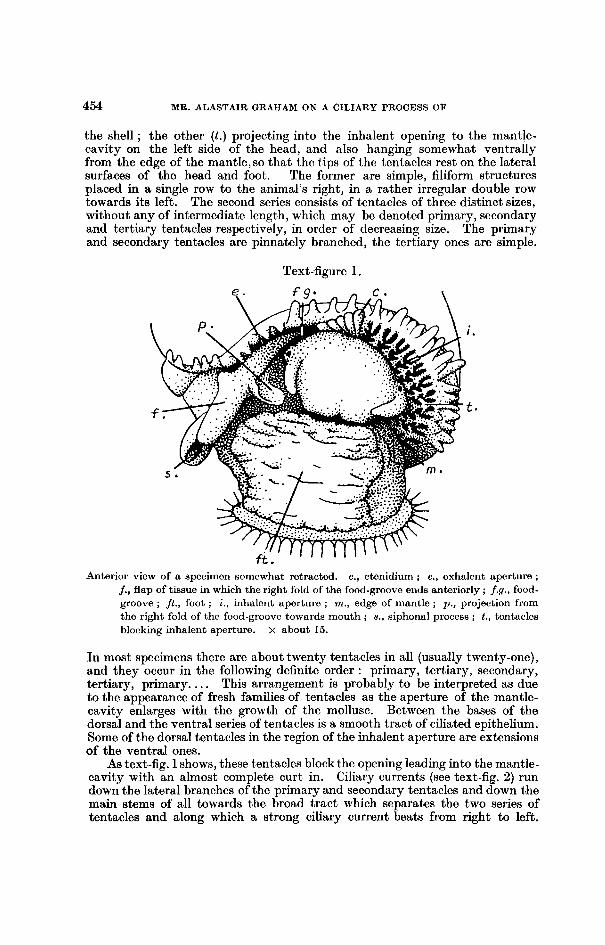
the shell ; the other ( t . ) projecting into the inhalent opening to the mantle- cavity on the left side of the head, and also hanging somewhat ventrally from the edge of the mantle, so that the t.ips of the tentacles rest on the lateral surfaces of the head and foot. The former are simple, filiform structures placed in a single row to the animal’s right, in it rather irregular double row towards its left. The second series consists of tentacles of three distinct sizes, without any of intermediate length, which may be denoted primary, secondary and tertiary tentacles respectively, in order of decreasing size. The primary and secondary tentacles are pinnately branched, the tertiary ones are simple.
Text4igure 1.
Anterior view of a specimen somewhat retracted. c., ctenidium ; e., exhalent aperture ; j., flap of tissue in which the right fold of the food-groove en& anteriorly ; f.g., food- groove ; jt . , foot ; i., inhalcnt aperture ; m., edge of mantle ; p. , projection from the right fold of the food-groove towards mouth ; s., siphonal process ; t., tentacles blocking inhalent aperture. x about 15.
In most specimens there are about twenty tentacles in all (usually twenty-one), and they occur in the following definite order : primary, tertiary, secondary, tertiary, primary. . . . This arrangement is probably to be interpreted as due to the appearance of fresh families of tentacles as the aperture of the mantle- cavity enlarges with the growth of the mollusc. Between the bases of the dorsal and the ventral series of tentacles is a smooth tract of ciliated epithelium. Some of the dorsal tentacles in the region of the inhalent aperture are extensions of the ventral ones.
As text-fig. 1 shows, these t,entacles block the opening leading into the mantle- cavity with an almost complete curt in. Ciliary currents (see text-fig. 2) run down the lateral branches of the primary and secondary tentacles and down the main stems of all towards the broad tract which separates the two series of tentacles and along which a strong ciliary current beats from right to left.
POOD-COLLEUTINO IN TURRITELLA. 455
This arrangement of the ciliary streams shows that the tentacles act as a sieve stretched across the entrance to the mantle-cavity to prevent the ingress of all but small particles. All large particles are caught by the tentacles, carried by the ciliary currents on their surface to the basal tract, along which they travel, to be deposited on the upper or lateral surface of the foot in the neigh- bourhood of the operculum.
The mantle-cavity is very long, and on opening it, by cutting through the mantle along the right side, the appearance seen in text-fig. 2 is presented. On the roof the ctenidium (c.), which is responsible for the creation of the water- currents, runs the whole length of the cavity. Along its base, on the left side, a t,hin, darkly coloured line marks the position of the osphradium (o . ) , which is unusually elongated, and between the ctenidium and the rectum and genital tract (g.), both of which lie in the extreme right of the mantle-cavity, lies the hy obranchial gland (h.), the secretion of which appears not to be mucus bu 1 probably some similar substance. Along the ctenidial axis runs a strip of mucous cells or “ endostyle ” (en.) similar to that described by Orton (1913) in Crepidula. The gill is composed of laterally flattened, finger-shaped filaments each attached to the ctenidial axis by its base and fused to the mantle by the basal third of its dorsal surface. The cilia primarily responsible for the produc- tion of the current are the laterals, strong cilia set in a slight groove which runs along the lateral surface of each filament ; these also sieve particles out of the stream and pass them to the dorsal surface of the filament, upon which an abfrontal tract transports them to the free extremity of the filament. Particles which are caught on the ventral surface of the filaments are passed by a similar frontal tract to the same place. A slight depression occupies the summit of each filament, and here material from the frontal and ahfrontal tracts accumu- lates and is rotated to form a, minute sphere of particles cemented together with mucus. Most of the mucus is provided by the endostyle, but there are also large numbers of mucous cells on the gill-filaments themselves. There are no ciliary currents on the ctenidial axis or on the osphradium.
.
On the floor of the mantle-cavity the most conspicuous object is a deep groove (f.9.) bounded by a pair of high folds. These structures arise at the posterior end of the mantle-cavity slightly left of the middle line, and run along its whole length in a somewhah diagonal fashion to the right side of the “ neck ” region of the animal. Here the left fold ends and the groove continues along the right side of the animal’s head, bounded on the left by a minute ridge and on the right by the right fold. This continues down the side of the head and then passes on to the foot, where it enlarges to form a semi-elliptical flap of tiuslie (f., text-fig. 1) set in a vertical position. Ventral to this extension of the right fold lies a second flap (s., text-figs. 1 & 2), horseshoe-shaped, with the points of the shoe directed backwards and the middle of the curve raised from the general level of the surface of the foot so that it projects into the surrounding water. The apical portions of this fold are kept curled over each other so as to form an incomplete “siphon.” This differs from the true siphon of other prosobranchs in being exhalent, not inhalent. Slightly ventral to the level of the mouth the right fold bears a saucer or spoon-shaped pro- longation (p . , text-figs. 1 & 2) which projects towards the middle line and thus comes to lie immediately below and a little to the right of the mouth.
The folds bounding the groove gradually increase in height as they pass anteriorly in the mantle-cavity.
On the roof of the mantle-cavity, to the left of the osphradium, ciliary currents beat down on to the floor, across which they continue towards the fold
456 MR. ALASTAIR GRAHAM ON A CILIARY PROCESS OF
bounding the groove on its left side. On the outer wall of this fold currents beat towards the summit. Particles which have passed the tentacles guarding tlhe entrance to the mantle-cavity and which fall on to the floor are thus trans- ported into the groove. On the right side of the groove the arrangement of the currents differs in the anterior and posterior regions of the cavity. In the latter the Ciliary currents again beat towards the fold bordering the groove, and on the outer wall of that they beat towards the crest so that particles from the floor of the cavity are carried into the groove ; in the anterior half of the mantle-cavity (in the neighbourhood of the anus) the ciliary currents beat diagonally outwards and towards the mid-line, so that particles are carried
Texbfigure 2.
The mantle-cavity has been exposed by an incision along its right side. Arrows indicato the courso of t,he ciliary currents. n., anus ; en., endostylo ; g . , genital tract (male) ; h., hypobranchial gland ; o. , osphradium ; u., urinary aperture. For other letters see text-fig. 1. x about 8.
towards the base of the right fold. Here there runs a strong exhalent current, and, as the outer wall of the fold in this region appears to be unciliated, all the particles are directed outwards along its base. The enlarged flap in which this fold terminates is normally kept bent over towards the right, and so forms a tunnel along which the rejected material is passed into the space between the limbs of the siphonal process along which it is carried. The mollusc maintains this in a distended condition well out from the side of the body, so that the unwanted or fzecal material which this channel usually bears is deposited some way off the body.
The gill-filaments lie roughly parallel with the roof and floor of the mantle- cavity, with their tips drooping SO as to rest on the latter. The position of
FOOD-COLLECTING IN TURRITELLA. 457
the tips, where the particles gathered on the surfaces of the filanients collect, varies, sometimes lying to the right of the groove, at other times to its left, and occasionally the filaments lie so as to dip into the groove itself. This last condition, however, is not frequent, as the two folds usually lie with their free edges overlapping, so as to convcrt the groove into a tunnel. In the most anterior part of the mantle-cavity, moreover, the ctenidial filaments are too short to reach the groove and always end to its left, thus avoiding the exhalent current on its right. The principal function of the ciliary currents which have been described on the floor of the cavity is thus to direct into the groove those collections of particles which are deposited on it from the tips of tho filaments.
On the exheme riglit of the mantle-cavity lie the rectum and the genital tract. The latter possesses an extremely complex ciliation, especially in the female, but the main effect in both sexes is of an outwardly directed stream with lateral tributary currents ; on the former cilia drive particles to the right and forwards, so that they ultimately get caught in the lateral tributaries of the outward flow along the genital groove. This, theoretically at least, con- stitutes a rejection tract, but it is doubtful whether it ever plays a prominent part in clearing unwanted particles out of the mantle-cavity, as it lies dorsal to and to the right of the.gill-filaments. The urinary aperture (u., text-fig. 2), lying near the proximal end of the rectum, opens directly on to this tract, which i.s thus of considerable importance even when the animal is not breeding.
Within the longitudinal groove running along the mantle-cavity the ciliary currents run transversely from right to l e f t d o w n the inner side of the right fold and up the inner side of the left. Along the summit of the left fold is a strong anteriorly directed tract. On the ctenidial axis runs a strip of mucous gland cells, the so-called endostyle, and the gill-filaments and walls of the groove are richly supplied with mucous cells, with the result that even when the animal is not catching food-particles a considerable quantity of mucus is being steadily produced and transported to the groove, wherein it forms a thick ropy string. Some of this mucus is also probably derived from the hypobranchial gland. Under the action of the ciliary currents of the groove this string is in a state of continual rotation, and is always being slowly passed forwards ; the mecha- nism is identical with the rotation and passage towards the stomach of the crystalline style. Food and other particles which are passed into the groove from the gill become entangled in the mucus and are rotated and carried forwards with it. The collection of particles is made more precise by the fact that the walls of the groove are muscular and are very sensitive to the presence of solid particles on their outer surface. When a large particle, or a cluster of small ones cemented together by mucus, travels up the outer surface of one of the folds, the two part at the summit ?o as to make an opening leading into the groove, and the fold opposite to that on which the particles are travelling straightens out so as to prevent them being shot into the mantle-cavity by the tangential motion they have had imparted to them during their ascent. Having caught the particles the fold rapidly contracts so as to convert the open groove into a closed tunnel once more. As, perhaps, the majority of the particles collected approach the groove from the left side, the ciliary currents beating downwards on the inner side of the right fold are well adapted for driving them into the groove.
Anteriorly, where the left fold ends, the forwardly directed current on its summit passes on to the side of the animal's '' neck ') and runs forward ventral to the right tentacle, gradually extending from the side of the head on to the floor of the groove. It finally ends on the little spoon-shaped projection
458 MR. ALASTAIR GRAHAM ON A CILIARY PROCESS OF
which lies below the mouth. The transverse currents in the groove do not extend further than the level of the base of tho tentacle, and are replaced by the anteriorly directed current on the mesial half, and by obliquely transverse and posteriorly directed currents on the lateral half. The latter have the effect of pushing the anterior end of the mucous string away from the right fold and of forcing it on to the forwardly beating cilia which cover the mesial half of the groove. The result is that the mucous string is carried along the side of the animal's head and collects on the spoon-shaped projection. This forms a platter from which the mollusc feeds : every now and again it may be seen to turn its head to the right, extrude the odontophore, and rake up the mixture of mucu: and food which is waiting for i t at this spot. One stroke of the radula usually pulls a considerable length of mucous string from the platter and out of the groove.
CILIARY CURRENTS IN THE ALIMENTARY CANAL. The buccal cavity is small and contains a very small, even degenerate,
odontophore with a narrow radula bearing fine teeth. Into it open a pair of minute salivary glands. Two folds project from the roof, one on either side of the mid-line dorsally, enclosing a groove : this is the anterior end of the dorsal food-channel, which stretches the lengbh of the esophagus. The two folds and the groove between are strongly ciliated, the remainder of the walls of the buccal cavity being without cilia. From the posterior end of the buccal cavity a narrow esophagus follows an approximately straight course to the stomach. Immediately behind the nerve-ring the esophagus is involved in a twisting due to the torsion of the visceral hump, and it is rotated through 180", so that the dorsal folds and the food-channel curve down the left side of the esophagus to occupy a topographically mid-ventral position. These structures are unpigmented, whilst the morphologically ventral half is dis- tinctly pigmented. The structure of this part of the alimentary canal I hope to deal with in greater detail elsewhere. On all parts of the buccal cavity and the esophagus the ciliary currents beat aborally.
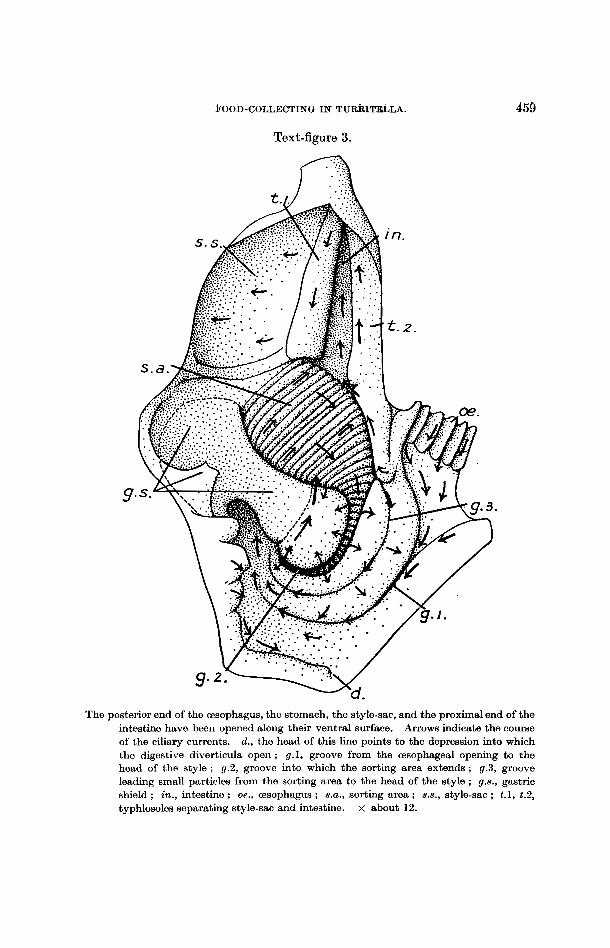
The esophagus (oe., fig. 3) leads to a stomach of complex structure, opening into it about the middle of the left side. The intestine (in.) and a short style- sac (s.s.) lead forwards from the anterior end of the stomach, the two com- municating with one another by a slit-like space (Seshaiya, 1932). A gastric shield (9.8.) covers a considerable part of the right and dorsal walls, and against this abuts the head of the crystalline style, which measures 3 4 mm. in length. The digestive diverticula (d.)-they are in all probability simply organs of absorption, since a style is present-open to the stomach a t the bottom of a slight depression posteriorly. At the opening of the esophagus the longitu- dinal folds within it come to an end and a groove (1) (g.1) bounded by two broad and flat folds leads particles back along the left wall of the stomach and round its posterior end to the region where the head of the style is rotating against the gastric shield. From the anterior border of the shield to the origin of the style-sac and of the intestine lies an area (8.a.) covered with shallow folds running in an obliquely longitudinal direction : these are continued up a groove (2) (9.2) which curves round the left margin of the gastric shield and which is bordered with thick lips. TO the left of this groove, between it and the groove (1) leading back from the esophageal opening, is a third groove (3) (gr.3). The folded area forms a sorting mechanism : small partieles from the region of the head of the style, brought to it by a current along the left margin of the gastric shield, are passed posteriorly across the folds and
BOOD-COLLECTING IN TUrnITEb-4 .
Text-figure 3. n . . ...
469
The posterior end of the Esophagus, the stomach, the style-sac, and the proximal end of the intestine have been opened along their ventral surface. Arrows indicate the course of the ciliary currents. d., the head of this line points to the depression into which the digestive diverticula open ; g.1, groove from the oesophageal opening to the head of the style ; g.2, groove into which the sorting area extends ; g.3, groove leading small particles from the sorting area to the head of the style ; g.8., gastric shield ; in., intestine ; oe., esophagus ; 8.U. , sorting area ; s.8., style-sac ; t.1, t.2, typhlosoles separating style-sac and intestine. x about 12.
460 MR. ALASTAIR GRAHAM ON A CILIARY PROCESS OF
up groove (2), out of which they are flicked by cilia set on the bounding folds. Those which pass to the left of the groove enter groove (3): which leads back to the head of the style and to the region of the duct of the digestive diverticula ; those which pass out of the groove to the right get mixed directly with the style and are carried back to the sorting area. Larger particles are carried along the folds of the sorting area directly into the intestine or into a groove bordering the area on its left side, in which a strong current leads to the intestine. On the walls of the style-sac transverse currents occur, rotating the style, and on one of the typhlosoles which separate style-sac from intestine (t.1) a strong current beats towards the stomach, thus pressing the style against the gastric shield. On the other typhlosole (t.2) and on the intestine itself the cilia beat in the opposite direction, towards the anus.
As usual, it is difficult to see currents in definite relation with the duct of the digestive diverticula. A slight inward current from the region of the gastric shield has been noticed.
The intestine runs from the stomach to the rectum, which opens into the mantle-cavity on the right side well forward(a., text-fig. 2). That part of the alimentary canal which runs across the floor of the cavity may be distinguished as the rectum, the rest as the intestine. In the latter the rejected matter from the stomach is moulded by ciliary and muscular means, which the aid of mucus, into oval faecal pellets ; the rectum merely serves to convey these across the mantle-cavity to a point whence they may be safely expelled.
DISCUSSION. Turritella provides yet another example of a ciliary feeding mechanism in
a gastropod mollusc provided with a crystalline style, and it illustrates once again the fundamental interrelationships between food, mode of feeding, structure of the alimentary canal, and t,he enzyme equipment of the animal, which the researches of Yonge (1930, 1932) have made familiar. As Turritella belongs to a group of prosobranchs far removed from the Calyptraeidae and Capulidae, although fairly closely allied to the Vermetidae, the mode of feeding has probably been independently acquired and, although similar in general principles, presents differences in detail to which attention should be directed.
In the first place it should be emphasized that it is not intended to suggest, on the basis of the present work, that Turritella is an exclusively ciliary feeder. It may well be, but it is also possible that the mechanism just described is one which the mollusc uses in addition to the more ordinary gastropod method of scraping material in with the radula. The structure of the gut, however, is such as to insist that there can be no trituration of material collected in this way-it must already be in a granular condition-and the reduction of the odontophore and of the salivary glands for the lubrication of the radula suggests that the latter is most frequently used for raking in the mixture of food and lubricating material brought to the mouth ready mixed along the ciliated groove and not for the independent collection of food. Hunt (1925) described Tur- ritelh as a selective feeder on bottom deposits, and Yonge (1937) suggests that these are composed partly of diatoms dropped from above and partly of true bottom-living algae.
The principal peculiarities in the arrangement of the ciliary feeding mecha- nism found in Turritella, when compared with those of the other gastropods which have adopted this means of collecting food, are to be correlated with the fact that it lives on a bottom of fine sand or mud, where the water may be somewhat turbid (Steven, 1930, and Yonge, in Zit.), whereaa the others live
FOOD-UOLLECTING IN TURRITELLA. 461
in comparatively clear surroundings. Under such circumstances it would obviously be impossible for any sorting mechanism within the mantle-cavity to deal adequately with the enormous amount of particulate matter which would enter. The only possible method of overcoming this is to prevent un- wanted matter fromentering, and to do this Turritella has evolved the extremely efficient curtain of tentacles close set along the mantle edge to guard the inhalent opening ; in addition, it possesses a sorting mechanism in the series of folds on the stomach wall, where the heterogeneous matter passed down the esophagus is sorted into large and small particles. The barrier of tentacles is so efficient in preventing large particles from entering the mantle-cavity that most ciliary currents within it lead to the food-groove, with the result that all particles that are permitted to enter in the first place in the last make their way to the mouth. It is improbable that Turritella could successfully exploit a ciliary method of collect,ing food in the habitat it has adopted unless it had evolved some such device.
Yonge (1938) has suggested that the currents in the mantle-cavity of prosobranchs which feed by ciliary means are adaptations of the original cleansing and respiratory currents, and has divided them into three principal tracts : A, rejecting large particles via the inhalent aperture ; B, carrying medium sized parGcles across the floor of the cavity for rejection through the exhalent aperture ; and C , carrying fine particles across the roof of the mantle-cavity, where they may be consolidated by the secretion of the hypo- branchial gland prior to ejection by way of the exhalent aperture along with fsecal and excretory matter. If the ciliary currents found in Turritella be made to conform to this scheme it will be seen that the currents on the tentacles presumably represent current A (though they are developed on the mantle itself and not on the floor of the mantle-cavity), those on the floor of the cavity leading into the food-groove B, whilst a relic of current C leads out through the exhalent aperture to the " siphon ') set on the right side of the foot. So far as the transformation of the original currents is concerned Turritella shows about the same degree of modification as Vermetus (Yonge, 1932) and rather less than the CalyptraAda and Capulus. In other respects, in response to the more difficult environment in which it lives, it shows a much higher degree of adaptation. This is well seen in the elaboration of current A by the develop- ment of tentacles whereby the large particles tending to enter the mantle- cavity are actively intercepted instead of being merely removed after they have passively gravitated on to the floor of the mantle-cavity and in the modifi- cation of the current of the food-groove. In place of the simple ciliated tract of Crepidula and its allies and the shallow food-groove of Vermetus there is present the deep channel described above, in which the food is rolled into a firm mucous string, the precise condition in which the animal requires it for transport along the esophagus to the stomach.
Turritella appears to differ from the ciliary feeding types described by Yonge (1938) in possessing numerous mucous cells on the gill-filaments and in having a hypobranchial gland. The conclusion that these prosobranchs had no hypo- branchial gland was reached, Professor Yonge has kindly informed me, on the basis of dissections ; if sections of the mantle be examined, however, it is found that a distinct gland is present between the ctenidial axis and the rectum, though i t no longer projects in folds as it does, for example, in Buccinum. It is the loss of folds, not the complete loss of the gland, that is due, as Yonge has shown, to the increased length of the gill-filaments. The gland is of moderate dimensions in Crepidula, but i s well developed in Calyptrzza and
462 MR. ALASTAIR QRAFIAM ON A CILIARY PROCESS OF
Capulus, secreting large quantities of mucus, though in none of these genera is it as large as in Turritella. Now it will be recalled in the first place that Yonge suggested that the original function of the hypobranchial gland was to produce a secretion to agglutinate the finest particles which entered the mantle-cavity before they were expelled through the exhalent aperture and, in the second, that all these gastropods, having a crystalline style in their stomach and relying on the ingestion of minute particles by the cells of the digestive diverticula for nourishment, select the finest particles €or food. It seems possible in such circumstances that not only the ciliary currents but also the hypobranchial gland may have undergone an alteration in function, and that i t no longer consolidates fine particles for rejection alone, but also for food. This is supported by its greater degree of development in Turritella, where the food undergoes the greatest amount of consolidation before being eaten, and also by the appearance seen in sections of Turritella and of Calyptrza. In the former the secretion of the gland stains brownish with iron haematoxylin, and brown streaks may be seen extending from the gland to the gill-filaments, across the mantle-cavity, and may also be found in the food-groove. I n Calyptrza, where the secretion is mucus, after staining with toluidin blue bright streaks of purple may be seen in corresponding positions, giving the impression that the secretion is being dragged from the surface of the gland by the cilia on the-underlying filaments. Against this supposition must be set the direction of the ciliary currents on the surface of the neighbouring rectum in Turritella, which appear to have retained their original outward beat ; they may, however, be effective only if an excess of secretion is produced on stimula- tion by noxious substances.
The presence of numerous mucous gland cells on the gill-filaments of Tur- ritella is also bound up with the manipulation of food in the mantle-cavity. In Vermetus, where the food receives little treatment on its way to the mouth, mucous cells occur freely on the filaments. In Crepiclula and its allies the number of mucous cells on the gills has been reduced, but this is offset by the appearance of a strip of mucous cells set along the ctenidial axis (Orton’s ‘‘ endostyle ”). In Turritella, where the food-particles are exposed to a con- siderable preliminary manipulation, for which large quantities of mucus are required, there is not only the series of cells on the ctenidial axis, but there are abundant mucous cells on the gill-filaments as well. The size of the “ en- dostyle ” is not such as to suggest that i t is capable, even in the Calyptrseidse, of producing by itself suficient mucus for the entanglement and manipulation of food, which supports the idea that the hypobranchial gland shares this duty.
In view of the preliminary manipulation to which the food is exposed before it enters the alimentary canal, the structure of the anterior part of the gut has been simplified ; the odontophore has become reduced in sue and is less powerful, the radula is modified €or raking in mucous strands or small particles, and the salivary glands, upon which most prosobranchs depend for the lubrication of the radula and of the fore-gut as well rn for the cementing of the individual particles of food into a continuous string, have been replaced by the mucous cells of the “ endostyle ” and of the walls of the pallial food- groove, and have dwindled to minute vestiges.
This clearly suggests the route by which the complete simplification of the gut of lamellibranchs in this region may have been brought about when the animah had become entirely dependent on ciliary currents for the collectioq of their food.
FOOD-COLLECTING I N TURRITELLA. 463
SUMMARY, The mantle-cavity of Turritella communis has an inhalent aperture on the
left side and an exhalent one on the right. The former is guarded by a complex array o f tentacles preventing the ingress of large particles. Ciliary currents collect particles falling on these and transport them to the foot (text-fig. 1).
Particles are collected on the gill-filaments and are transferred to a deep groove bounded by folds which runs along the floor of the mantle-cavity to the right side of the head, ending in a projection below the mouth. Food-particles are built into a ropy string with mucus in this groove, and this is slowly passed forwards to the mouth, into which it is raked.
Exhalent currents pass from the right side of the cavity to a siphonal process on the foot (text-fig. 2).
The mucous string is passed into the buccal cavity, in which lies a reduced odontophore and into which open minute salivary glands, thence down the esophagus, which is straight apart from the twisting caused by torsion, to the stomach. Into this projects a crystalline style, its head bearing against a gastric shield, and a complex ciliary sorting and mixing mechanism is described on its walls. Small particles pass to the digestive diverticula for ingestion, larger ones to the intestine, where they will be elaborated into faxal pellets (text-fig. 3).
In the light of these fact,s the peculiarit,ies of the feeding mechanism of Turritella are discussed, and it is concluded that they have been evolved in relation to its habitat.
REFERENCES TO LITERATURE. HUNT, 0. D. 1925.
ORTON, J. H. 1912.
The Food of the Bottom Fauna of the Plymouth Fishing Grounds J. Mar. Bid. h s . vol. xiii. pp. 560-599.
The Mode of Feeding in Crepidulu, with an Account of the Current- producing Mechanism in the Mantle Cavity, and some Remarks on the Mode of Feeding in Gastropods and Lamellibranchs. J. Mar. Biol. Ass. vol. ix. pp. 444-478.
On Ciliary Mechanisms in Brachiopocts and some Polychaetes, with a Comparison of the Ciliary Mechanisms on the Gills of Molluscs, Proto- chordata, Brachiopods, and Cryptocephalous Polychaetes, and an Account of the Endostyle of Crepidula and its Allies. J. Mar. Biol. Ass. vol. x.
RANDLES, W. B. 1902: On the Presence of a Crystalline Style and Style-sac in Tzirritelkz
SESHAIYA, R. V. 1932. The Style Sacs of some more Gastropods. Rec. Ind. Mus. Calcutta,
STEVEN, G. A. 1930. Bottom Fauna and the Food of Fishes. J. Mar. Biol. Ass. vol. xvi.
YONCE, C. M. 1930. The Crystalline Style of the Mollusca and a Carnivorous Habit cannot Normally Co-exist.
YONCE, C. M. 1932. Notes on Feeding and Digestion in Pteroceru and Vermetus, with a Discussion on the Occurrence of the Crystalline Style in the Gastropoda. Sci. Rpts. Gt. Barrier Reef Exped., 192&29, Brit. MILS. (Nat. Hist.), vol. i. pp. 259-281.
The Biology of Aporrhuis pes-pelecani (L.) and A . serresiunu (Mich.). J. Mar. Biol. Ass. vol. xxi. pp. 687-703.
Evolution of Ciliary Feeding in tho Prosobranchia, with an Account of Feeding in Cupulus unnarirus.
ORTON, J. H. 1913.
pp. 283-311.
communzs.
vol. xxxiv. pp. 171-175.
Anat. Anz. vol. xxi. pp. 200-203.
pp. 677-700.
' Nature,' vol. cxxv. pp. 444-445.
YONCE, C. M. 1937.
YONCE, C. M. 1938. J. Mar. Biol. Ass. vol. xxii. pp. 453-468,