On the Mechanism of Cardiac Glycoside Action STIMULATION OF MYOSIN B SUPERPRECIPITATION BY OUABAIN AND DIGOXIN By Linda Stowring, M.S., William J. Bowen, Ph.D., Patrick Mattingly, B.A., and Manuel Morales, Ph.D. ABSTRACT A study has been made of the effect of ouabain and other cardiac glyco- sides on a faithful model of contraction, viz. the "superprecipitation" which occurs when ATP is added to a suspension of purified myosin B. Structural changes during superprecipitation were followed by a turbidity method, and rates of ATP utilization with a pH stat. At "optimal" concentrations (ca. 10~ 6 M and 10" 3 M) these substances approximately double the structural and the ATP utilization rates of the myosin B system, but at no concentration do they affect the ATPase of myosin A. In the one case studied, cardioactivity of the substance appeared to parallel its effect in our test system. ADDITIONAL KEY WORDS myosin A ATP utilization actin-myosin junctions • Skou 1 and Post et al. 2 have shown that the cardiac glycoside, ouabain, affects the "membrane ATPase" of crab nerve and human erythrocyte, respectively. Since these ATPases are thought to be involved in "active" ion transport, it has been natural to conjecture that the inotropic effect of glycosides is exerted via the electrical properties of cardiac cells; this may indeed be so, but it is note- worthy that in the model systems the cardiac glycoside concentration is of the order of 10" 5 M, while in reinforcing papillary muscle contraction it often acts at l(r" 8 M (Hoffman, personal communication). In this paper it will be shown that ouabain and digoxin affect the "in vitro contractile sys- tem" (superprecipitation of myosin B) at concentrations as low as 10~ u M. Hints that cardiac glycosides affect contractile proteins From the Cardiovascular Research Institute, Uni- versity of California Medical Center, San Francisco, California. Supported by Grant 60-CI-8 from the American Heart Association, Grant G-19442 from the Na- tional Science Foundation, and Grant HE-06285 from the U. S. Public Health Service. Patrick Mattingly was a Predoctoral Trainee of the U. S. Public Health Service, supported through Grant HTS-5251. Manuel Morales is a Career Investigator of the American Heart Association. Accepted for publication May 27, 1966. directly have emerged in the past (see, for example, Bowen s and Robb and Mallov 4 ), but lack of a sufficiently sensitive method has delayed definition of a direct effect. In this work the structural aspect of superprecipita- tion was followed by the turbidity method of Ebashi, 6 as modified by Yasui and Watan- abe, 6 and the fast ATP utilization accompany- ing superprecipitation was followed in a pH stat. Structural change and ATP utilization seem to be "coupled," and both are sensitive to cardiac glycosides. The test material is a fine, sonicated suspension of thrice-purified myosin B; the possibility of contamination with cellular debris or with particulate relax- ing factor is remote. The fact that cardiac glycosides potentiate the ATP response of myosin B does not mean that they exert their physiological effect on the contractile system, any more than the fact that cardiac glycosides affect membrane ATP- ase means that their physiological effect is exerted on membranes. A restrained statement at this juncture is that cardiac glycosides seem to affect at least two systems of energy-trans- ducing ATPases. Materials and Methods The bulk of this work was done with crystalline ouabain (Nutritional Biochemicals); when in a 496 Circalttion Rtlmrcb, Vol. XIX, Stpttmitr 1966 by guest on May 24, 2018 http://circres.ahajournals.org/ Downloaded from
Transcript
On the Mechanism of Cardiac Glycoside Action
STIMULATION OF MYOSIN B SUPERPRECIPITATION BY OUABAIN AND DIGOXIN
By Linda Stowring, M.S., William J. Bowen, Ph.D.,
Patrick Mattingly, B.A., and Manuel Morales, Ph.D.
ABSTRACTA study has been made of the effect of ouabain and other cardiac glyco-
sides on a faithful model of contraction, viz. the "superprecipitation" whichoccurs when ATP is added to a suspension of purified myosin B. Structuralchanges during superprecipitation were followed by a turbidity method, andrates of ATP utilization with a pH stat. At "optimal" concentrations (ca. 10~6
M and 10"3 M) these substances approximately double the structural and theATP utilization rates of the myosin B system, but at no concentration do theyaffect the ATPase of myosin A. In the one case studied, cardioactivity of thesubstance appeared to parallel its effect in our test system.
ADDITIONAL KEY WORDS myosin AATP utilization
actin-myosin junctions
• Skou1 and Post et al.2 have shown thatthe cardiac glycoside, ouabain, affects the"membrane ATPase" of crab nerve and humanerythrocyte, respectively. Since these ATPasesare thought to be involved in "active" iontransport, it has been natural to conjecturethat the inotropic effect of glycosides isexerted via the electrical properties of cardiaccells; this may indeed be so, but it is note-worthy that in the model systems the cardiacglycoside concentration is of the order of10"5 M, while in reinforcing papillary musclecontraction it often acts at l(r"8 M (Hoffman,personal communication).
In this paper it will be shown that ouabainand digoxin affect the "in vitro contractile sys-tem" (superprecipitation of myosin B) atconcentrations as low as 10~u M. Hints thatcardiac glycosides affect contractile proteins
From the Cardiovascular Research Institute, Uni-versity of California Medical Center, San Francisco,California.
Supported by Grant 60-CI-8 from the AmericanHeart Association, Grant G-19442 from the Na-tional Science Foundation, and Grant HE-06285 fromthe U. S. Public Health Service.
Patrick Mattingly was a Predoctoral Trainee of theU. S. Public Health Service, supported through GrantHTS-5251.
Manuel Morales is a Career Investigator of theAmerican Heart Association.
Accepted for publication May 27, 1966.
directly have emerged in the past (see, forexample, Bowens and Robb and Mallov4),but lack of a sufficiently sensitive method hasdelayed definition of a direct effect. In thiswork the structural aspect of superprecipita-tion was followed by the turbidity method ofEbashi,6 as modified by Yasui and Watan-abe,6 and the fast ATP utilization accompany-ing superprecipitation was followed in a pHstat. Structural change and ATP utilizationseem to be "coupled," and both are sensitiveto cardiac glycosides. The test material is afine, sonicated suspension of thrice-purifiedmyosin B; the possibility of contaminationwith cellular debris or with particulate relax-ing factor is remote.
The fact that cardiac glycosides potentiatethe ATP response of myosin B does not meanthat they exert their physiological effect onthe contractile system, any more than the factthat cardiac glycosides affect membrane ATP-ase means that their physiological effect isexerted on membranes. A restrained statementat this juncture is that cardiac glycosides seemto affect at least two systems of energy-trans-ducing ATPases.
Materials and MethodsThe bulk of this work was done with crystalline
polar solvent this material moved as a singlespot in thin-layer chromatography; contaminationwith Ca was less than 3 parts per million and withMg less than 2 parts per million using the methodof atomic absorption. Digoxin was supplied byBurroughs-Wellcome, and strophanthidinic acidwas kindly donated by Dr. Maximillian Ehren-stein. Naja naja venom (lyophilized was ob-tained from Ross Allen's Reptile Institute, SilverSprings, Florida.
Disodium dihydrogen ATP, Tris (tris-hydroxy-methyl aminomethane) and repurified maleicacid were obtained from Sigma Chemical Co.Inorganic salts were J. T. Baker, reagent grade.Double-distilled (the second time from a glassstill) water was used throughout.
The preparation and characterization of myosinA has been recently described by us.7 Brei forthe extraction of myosin B was made from (1)(Waring) blended beef hearts, chilled on exci-sion, dissected free of connective tissue and fat,and used within 2 hr of sacrifice, or (2) (Latapie)minced bled-rabbit dorsal muscles immediatelychilled, dissected free of fascia and fat, and usedwithin 30 min of sacrifice. Breis were extractedfor 20-22 hr at 0° C in "Weber-Edsall solution"(0.60 M KCl, 0.04 M KHCO.,, and 0.01 MKoCO8). Following extraction, preparations werecentrifuged at 8,000 g (this and subsequent cen-trifugations were performed in a Spinco ModelL-2 preparative ultracentrifuge with rotor temper-ature of 0° C), filtered through clean fine gauze(to remove lipid scum), and diluted so that [KCl]became 0.25 M. The pH was adjusted to 6.80with dilute acetic acid, and the preparation wasallowed to stand. It was then centrifuged for 20min at 8,000 g. The supernatant was dis-carded and the precipitate was collected andweighed, and enough neutralized concentratedKCl solution was added to make [KCl] = 0.60 Mand thereby redissolve the precipitate. The solu-tion was now centrifuged at 46,000 g for 30min, employing centrifuge tubes with nylon caps.The central portions of the supernatants werecollected. Starting with dilution to [KCl] = 0.25 M,the foregoing procedure was repeated two moretimes, except that the final solution was centri-fuged for a full hour. The physical nature of themyosin B suspension obtained on dilution to lowionic strength depends on the schedule of dilu-tion. Except when otherwise specified, the fol-lowing was done at 0° C, using prechilled con-tainers. Stock myosin B solution (usually 1-2%)was diluted by addition of neutral 0.60 M KCl,first to 0.5%, then to 0.1*. Then the myosin Bwas precipitated by adding 0.06 M KCl and 0.02M Tris-maleate (pH 7.0); in this operation theconcentrations of protein, KCl, and Tris-maleate
Circulation Rtjmrcb, Vol. XIX, Stpumbtr 1966
fall, respectively, to 0.02*, 0.17 M and 0.016 M(ionic strength, ca. 0.185 M ) . The resulting sus-pension was allowed to stand overnight. Of thissuspension, 5 ml was pipetted (after the pipettehad been rinsed several times with suspension)into a beaker, and the system was allowed towarm to 25° C in a constant temperature bath(ca. 15 min), was sonicated for 10 sec (at 20Kcps in a Brownwill Sonicator), and the contentswere poured gently into a Zeiss 2 cm X 2 cmquartz cuvette. Then 5 ml of 25° C solvent (0.06M KCl, 0.02 M Tris-maleate, pH 7.0), or of 25°C solvent containing twice the desired cardiacglycoside concentration were pipetted and mixedwith a gently turning magnetic stirrer for a con-stant length of time. In this final solution, [KCl] =0.12 M, [Tris] = 0.018 M and ionic strength isca. 0.13 M. The drug thus interacted with theprotein for 90-120 sec; much longer incubationsgave the same result. Superprecipitation wasinitiated by adding 250 //.liter of the desired ATPconcentration. Again, the stirrer frequency wasreproduced with a Variac and maintained through-out the measurements. The apparent opticaldensity at 550 mfj, of the superprecipitating sus-pension was compared with the same suspensionprior to ATP addition. The difference betweenthese two readings (when these are converted tooptical density units) is taken as the "extent"of structural change, and the rate at which extentvaries with time will often be called the "struc-tural" rate to distinguish it from the rate of ATPhydrolysis. The contents of the cuvette were heldat 25°C in the constant temperature jacket(equipped also with stirrer) of a Zeiss PMQ IIspectrophotometer. The output of the spectro-photometer was applied to a Zeiss log converterand, finally, was linearly displayed in OD unitson a Minneapolis-Honeywell recorder (t1/2 = 0.05sec; accuracy, 0.005 OD units).
Measurements of ATP utilization or of ATPaseactivity were sometimes made with our variantof the Fiske-Subbarow orthophosphate assay,employing SnCI2 as the reducing agent. Rateswere calculated from the zero order phase of thereaction (first 200 sec) by drawing a line through3 or 4 points. In such cases the conditions for theassay were exactly those in which superprecipita-tion was observed. More often ATPase activitywas recorded in a Radiometer pH stat, again in0.12 M KCl, pH 7.0, 25° C, except that the Triswas omitted. In separate measurements it wasascertained that rates of proton production mul-tiplied by 1.59 equalled rates of ATP hydrolysis.In pH stat measurements, stirring and electrodeartifacts subsided in less than 20 sec; normally thelinear phase of superprecipitation lasted for atleast 150 sec.
The conditions of our assay are such that0.1 m a ATP "saturates" the myosin B system;i.e., at this or higher concentrations the finalextent and the rate of superprecipitation andthe ATPase activity are maximal and inde-pendent of the ATP concentration. As didRainford et al.8 we found concentrations of0.025 or 0.05 mM ATP to be "undersaturat-ing"; as in their work also, we found indica-tions that, for a given preparation, the sen-sitivity to ATP varies with the age of thepreparation.
In this work we studied the effects of bothouabain and digoxin, on both skeletal (14different preparations) and cardiac (4 differ-ent preparations) myosin B, but the systemouabain-skeletal myosin B was studied mostextensively. Nonetheless, the data permit usto say that the systems ouabain-cardiac, digox-in-skeletal, and digoxin-cardiac were studiedenough to ensure that the basic findings onouabain-skeletal apply as well to them. Earlyon, we examined for the possibility that pro-gressive purification of myosin B progressivelyeliminated sensitivity to ouabain, and also forthe possibility that increased length of in-cubation of myosin B with cardiac glycosideleads to increased effect; neither phenomenonwas found.
EFFECT OF OUABAIN ON SUPERPRECIPITATIONPARAMETERS AT DIFFERENT CONCENTRATIONS OF ATP
In a typical record of superprecipitationextent following ATP addition, the extent firstrises rapidly and then more slowly approachesa horizontal asymptote; in such cases the ini-tial "structural rate" is easily measured fromthe plot. Occasionally, the plot has a slightsigmoid character; then we have taken themaximum rate (at the inflection) to representthe structural rate. If the [ATP] is saturating,then an "optimal" (see below) concentra-tion of cardiac glycoside approximately dou-bles the structural rate, but has little or noeffect on the asymptotic, or final, value of theextent (Fig. 1). On the other hand, if [ATP]is chosen to be undersaturating, both rateand final extent are elevated (Fig. 2). Suchresults with ouabain are very reproducible
(=
g
—1|
A
=
m
—i
d
2
- +- ^ 1
I1
—
•5-
1 1
f-Ht-H
H
ot-H
-v,
i—r
i—1
(71
1 i
ft
r
&
CO
—
l»|
FIGURE 1
The effect of ouabain on the rate of superprecipttationof skeletal myosin B with saturating [ATP], Actual re-corder tracing. A, 6.7 X 10-* M ouabain. B, control.0.01% myosin B, 0.12 M KCl, 0.018 M tris maleate, pH7.0, lmM ATP, 2S'C. FuU vertical scale, 0.5 ODunits. Smallest horizontal division, 25 sec.
on every preparation tested. Analogous re-sults are obtainable with digoxin or strophan-thidinic acid with saturating [ATP], but theother glycosides were not studied with under-saturating [ATP].
THE INFLUENCE OF OUABAIN CONCENTRATION ONSTRUCTURAL EFFECTS
In Figure 3 we show results which are verytypical of ouabain when this substance is add-ed to systems in which [ATP] is undersatu-rating (when saturating [ATP] is employed,optima of effect on rate are obtained at thesame concentration of ouabain, but then ofcourse effects on final extent cannot be ob-
The effect of ouabain on the rate and extent of superprecipUation of skeletal myosin B withundersaturating [ATP]. Actual recorder tracing. A, 10-" M ouabain. B, control. Myosin B 0.01%,0.12 M KCl, 0.018 M Ms maleate, pH 7.0, ATP, 0.05 nut, 25'C. Full vertical scale, 0.5 ODunits. Smallest horizontal division, 25 sec.
served). However, it must be said here thatwhile all preparations show two optima, thecardiac glycoside concentrations at whichthese optima occur vary with age of a prepara-tion, and from one preparation to another, butnot within a single stock suspension; thus,sometimes optima may occur at IO^M and10̂ * M, and in another instance at 10~nM and10"° M. Most frequently the low concentra-tion optimum occurs at 10~9 M with eitherouabain or digoxin, and the high concentra-tion optimum with ouabain occurs at 10"" M(insolubility of digoxin precludes definition ofa high concentration optimum, but at 10"° Mdigoxin effect is increasing with concentra-tion).
THE INFLUENCE OF OUABAIN CONCENTRATION ONRATE OF ATP UTILIZATION
For reasons to be discussed, we studied theeffect of ouabain at various concentrations ontwo rates of ATP utilization. In order to defineprecise rates, saturating [ATP] was used. Forvarious concentrations of ouabain, Figure 4,curve I, shows the dependence of the fast
CircuUuon Rtitmnb, Vol. XIX, Stpitmbtr 1966
ATPase of myosin B (the activity concurrentwith structural events of superprecipitation);curve II shows the dependence of the ATPaseof myosin A dissolved in precisely the samesolvent. For reference, structural rate (curveIV) and extent (curve III) measurements,made at the same hour on the same myosin Bpreparation, are also shown. Clearly, ouabainhas no effect on the activity of myosin A.Also it has no effect on the ATPase of myosinB observed after superprecipitation is over.Moreover, it is evident that in concentrationsbetween 10~18 M and 10~7 M, ouabain affectsthe fast myosin B ATP utilization in much thesame way that it affects the structural param-eters. Between lCH M and lO"3 M the struc-tural parameters often decreased while theATP utilization rate increased.
OTHER SYSTEMS AND OTHER CARDIAC GLYCOSIDES
Figure 5 shows the effect of digoxin on the"structural rate" of cardiac myosin B. At thesame molarity range, digoxin has roughlythe same effect as ouabain. On the other hand,strophanthidinic acid must be 100 times more
Effect of different ouabain concentrations on extent and rate of skeletal myosin B super-precipitation (Supptn.) with undersaturating [ATP]. [ATP], 0.025 TUM. 0.01% myosin B, 0.12,M KCl, 0.016 M iris maleate, pH 7.0, 25°C.
concentrated in order to elicit the same effectas ouabain (Fig. 6).
POSSIBLE COMPLICATIONS BY SALTS
Because superprecipitation under our con-ditions is very sensitive to Mg2+ we studiedthe effect of Mg-* addition (10"6 M MgCl2)and subtraction (10"" M EDTA) on experi-ments such as shown in Figure 3. In both in-stances, at all ouabain concentrations, theouabain effect was additive with the salteffect. A concentration of IO"6 M CaCl̂ .,thought to be capable of inactivating particu-late relaxing factor, was also additive withthe ouabain effect.
POSSIBLE COMPLICATIONS BY PARTICIPATE"RELAXING FACTOR"
It is well known that particulate relaxingfactor is destroyed by phospholipase.9 To testwhether the ouabain effect might be merelyan inhibition of membranous factor,10 we pre-incubated the myosin B preparation withphospholipases before testing with ouabain.
We studied phospholipase A (unheated snakevenom); this enzyme was very active (94 unitsmg-1 to-1, pH 7.0, 25°C, no Ca2* added)toward lecithin when in superprecipitation sol-vent. The incubations were carried out insuperprecipitation solvent for 40 min at 25°C. The weight concentration of phospholipasewas about equal to that of myosin B. In allexperiments the ouabain effect per se wasunchanged following the incubations. The rateand extent of superprecipitation in the ab-sence as well as in the presence of ouabainwere reduced by these high concentrations ofphospholipase, but this reduction was inde-pendent of incubation time and therefore notdue to enzymatic action of the phospho-lipase.
DISPERSION OF DATA
In the present state of knowledge, the de-gree to which myosin is directly bonded toF-actin, and the degree to which the myosinB floe is dispersed are essentially uncontrolled.
Effect of increasing ouabain concentration on: III, the extent of superprecipitation (Supptn.)of myosin B (A A); (1) rate of utilization of ATP by myosin B (0 • ) ;IV, rate of superprecipitation of myosin B (x X); and II, myosin A ATPase(O O). Superprecipitation data, 0.01% myosin B, 0.12 M KCl, 0.018 M tris maleate,pH 7.0, 0.05 mix ATP. ATP utilization by myosin B (pH stat), 0.01% myosin B, 0.12 M KCl,0.1 m« ATP, 5 mw KOH as titrant, pH 7.0. Myosin A ATPase, 0.02% myosin A, 0.12 MKCl, 0.2 mi* ATP, 5 nut KOH as titrant, pH 7.0. Myosin A ATPase data, mean of three assays;all other data, mean of two assays. All experiments done at 25°C.
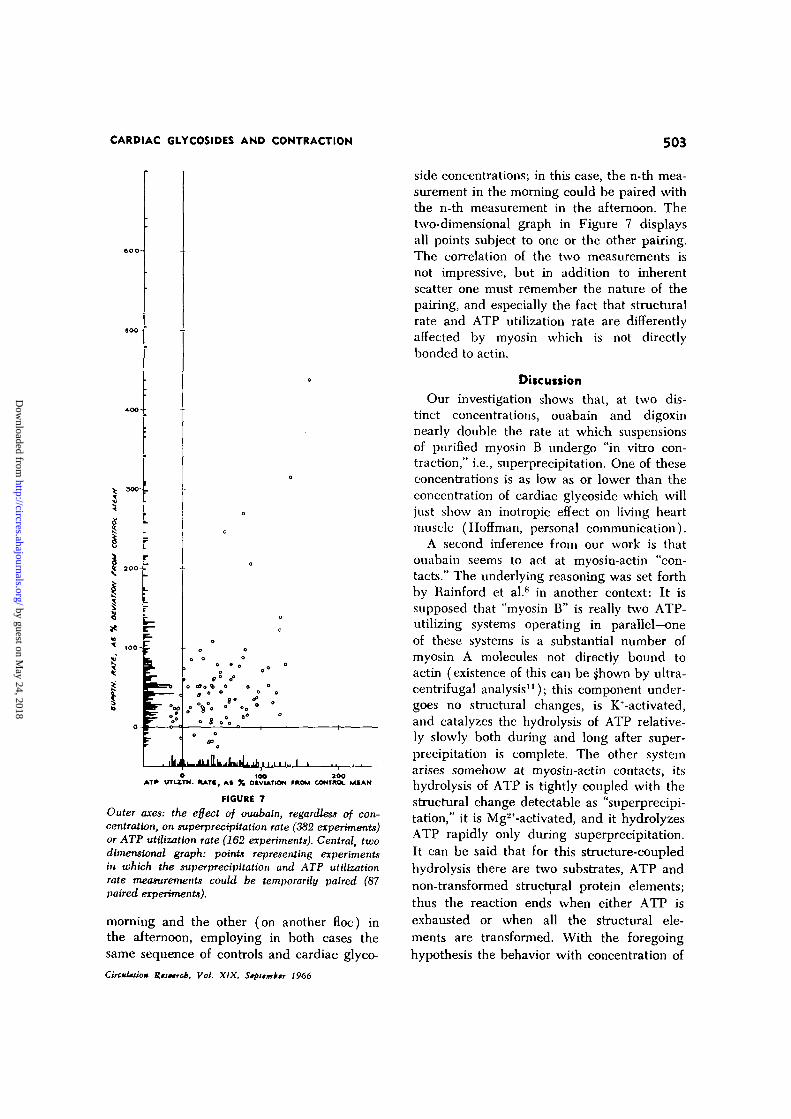
Additionally (see Discussion), myosin (A)not directly bonded to actin contributes sub-stantially to ATP hydrolysis [for example,the magnitude of the effect which a givencardiac glycoside concentration has on therate of ATP utilization can be reduced tozero by experimental addition of myosin(A)], but relatively little to optical changeson ATP addition. From these circumstancesarises a pronounced variability among mea-surements (of either structural or enzymaticparameters) made on different preparations,or on the same preparation at different ages.Our more detailed conclusions are based onmore reproducible results (for example, thosedisplayed in Figs. 3, 4, and 5) obtained inspecial paired experiments (see below). How-ever legitimate, this is a form of data selection,so it is necessary to show that also by usingall recent data (382 experiments on super-
CircnUium Ruetrcb. Vol. XIX, Stpttmbtr 1966
precipitation rate, 162 experiments on ATPutilization rate, on a total of 5 separate myo-sin B preparations) obtained in this labora-tory, the major conclusion of this paper isfirmly upheld. On the one-dimensional graphof Figure 7 are shown all the responses (aspercent deviations from the mean of severalcontrols) ever obtained to ouabain, regardlessof concentration. If ouabain had no effect,then so large a number of rate measurementswould tend to a distribution symmetricalabout the origin; however, the overwhelmingmajority of points deviate positively; that is,ouabain accelerates both the rate of structuralchange and the rate at which ATP is hy-drolyzed during the structural change. Withour present technique it is not possible tomeasure optical density and proton releaseon the very same aliquot of floe, hence tomake a strict pairing of the two measurements.
The effect of different concentrations of digoxin on the rate of superprecipUation of cardiacmyosin B using saturating [ATP]. 0.01% myosin B, 0.12 M KCl, 0.018 M tris maleate, pll 7.0, 1nut ATP, 25°C.
0.0 1 O r
10-12
Cone of Drug, moles • L"FIGURE 6
Comparison of the effect of ouabain ( • ) and strophanthidinic acid (Q-
\0~
-Q) on the rateof superprecipitation of skeletal myosin B using saturating [ATP], 0.01% myosin B, 0.12 H KCl,0.018 M tris maleate, pH 7.0, 1 nun ATP, 25'C.
In -many instances, however, it was possiblefor two operators to draw aliquots and makethe two measurements at very nearly the same
time (circumstances of Fig. 4). Still morefrequently it was possible for a single oper-ator to make one type of measurement in the
Outer axes: the effect of ouabain, regardless of con-centration, on superprectpUation rate (382 experiments)or ATP utilization rate (162 experiments). Central, twodimensional graph: points representing experimentsin which the superpredpitation and ATP utilizationrate measurements could be temporarily paired (87paired experiments).
morning and the other (on another floe) inthe afternoon, employing in both cases thesame sequence of controls and cardiac glyco-CircuUtio* Rtstrcb, Vol. XIX, Stpiimk; 1966
side concentrations; in this case, the n-th mea-surement in the morning could be paired withthe n-th measurement in the afternoon. Thetwo-dimensional graph in Figure 7 displaysall points subject to one or the other pairing.The correlation of the two measurements isnot impressive, but in addition to inherentscatter one must remember the nature of thepairing, and especially the fact that structuralrate and ATP utilization rate are differentlyaffected by myosin which is not directlybonded to actin.
Discussion
Our investigation shows that, at two dis-tinct concentrations, ouabain and digoxinnearly double the rate at which suspensionsof purified myosin B undergo "in vitro con-traction," i.e., superprecipitation. One of theseconcentrations is as low as or lower than theconcentration of cardiac glycoside which willjust show an inotropic effect on living heartmuscle (Hoffman, personal communication).
A second inference from our work is thatouabain seems to act at myosin-actin "con-tacts." The underlying reasoning was set forthby Rainford et al.8 in another context: It issupposed that "myosin B" is really two ATP-utilizing systems operating in parallel—oneof these systems is a substantial number ofmyosin A molecules not directly bound toactin (existence of this can be jhown by ultra-centrifugal analysis11); this component under-goes no structural changes, is Reactivated,and catalyzes the hydrolysis of ATP relative-ly slowly both during and long after super-precipitation is complete. The other systemarises somehow at myosin-actin contacts, itshydrolysis of ATP is tightly coupled with thestructural change detectable as "superprecipi-tation," it is Mg2+-activated, and it hydrolyzesATP rapidly only during superprecipitation.It can be said that for this structure-coupledhydrolysis there are two substrates, ATP andnon-transformed structural protein elements;thus the reaction ends when either ATP isexhausted or when all the structural ele-ments are transformed. With the foregoinghypothesis the behavior with concentration of
an agent which affects only the structure-coupled system is predictable. For Rainfordet al., the agent was hydrostatic pressure, andit caused inhibition; here it is ouabain, and itcauses activation. If ATP is presented to thesystems in unlimited amount, the observedextent of superprecipitation will be governedby the amount of non-transformed protein.Addition of an activator like ouabain will in-crease the rate of the structural change, butthe extent will be the same because it is stillprotein limited. When ATP is presented inlimited quantity, it will be parceled out tothe two enzymes according to their zero-orderrate constants; the reactions will stop whenATP is exhausted, and the observed extentwill depend on the fraction of the ATP whichthe structure-coupled system won in the com-petition. Under these conditions, addition ofan activator like ouabain gives the structure-coupled system an advantage, i.e., aided byouabain, it will win a larger fraction of theATP and the observed extent will therefore belarger, not only the observed rate. It is interms of the foregoing that we explain whyouabain (in an effective concentration) in-creases rate (ATPase or structural) only if[ATP] is saturating, and both rate and extentif [ATP] is undersaturating; also, why oua-bain does not affect the ATPase of myosin A,or the ATPase which continues long aftersuperprecipitation is over. In these terms, the"concentration" of the "structure-coupled sys-tem" is not defined alone by the concentrationof, say, myosin or actin, but depends markedlyon the manner in which the actin and myosinare co-precipitated in the course of preparingthe myosin B suspension; furthermore, the"concentration" is considerably less than themolar concentration of the lesser component(myosin A).
Finally, more of a suggestion than a con-clusion, is that, in the one case tried, therewas a correlation between the effectiveness oftwo cardiac glycosides (ouabain and stro-phanthidinic acid) in their ability to kill catsand in our in vitro tests, and it is conceivablethat superprecipitation could be used for assayor screening.
While we consider the data which we havecited reliable, and the interpretations whichwe have offered reasonable, we would beremiss to slur over some obvious difficulties.It is already perplexing that the effect of thecardiac glycosides passes through an "opti-mum," indicating the existence of an inhibi-tory factor which we do not understand; tomake matters worse, there are actually twooptima, even though we seem to be workingwith a pure cardiac glycoside. Because a car-diac glycoside inhibition of "membraneATPase" has been established, because a hy-drophobic steroid molecule might be expectedto interact with lipids, because the optimalconcentrations are erratic, and because theconcentration of "receptor substance" in oursystem would have to be about 1/100 of theconcentration of myosin, one must seriouslyconsider the hypothesis that the cardiac gly-coside is neutralizing a contamination of par-ticulate relaxing factor. Against this hypothe-sis we have the experimental observations thatthe cardiac glycoside effect is unaffected bytreating the myosin B with either Ca2+ orphospholipases; also we can argue that theconcentration of "actin-myosin contacts" canvery well be 1/100 the concentration ofmyosin, and that it is bound to be erratic be-cause it is almost impossible to co-precipitatetwo proteins reproducibly.
As already remarked, the argument thatcardiac glycosides act physiologically on mem-branes is no better than the argument thatthese substances act in the region of thick-thin filament interaction (at this stage bothare weak arguments). Indeed, that the con-tractile system is affected by physiologicallyplausible (i.e., low enough) concentrationswhile membranes aren't, and that tritiateddigoxin locates in the A-band (see ref. 13),are facts which undermine the "membranetheory." To be sure, in a 1961 symposiumRepke12 reported a 30-50* activation of "car-diac muscle membrane ATPase" by 10"° Mto 10~8 M k-strophanthoside, but activation ofrelaxing factor is not equivalent to activationof contraction—indeed it is just the reverse.Also, in a recent communication, Fozzard
and Smith13 report a correlation of tritium-labeled cardiac glycoside with properties usu-ally ascribed to membrane fragments—easysedimentation and "Na-K ATPase"; becauseof this correlation they appear to surrendertheir original hypothesis,14 and accept morethe views of Sonnenblick et al.,15 who feelthat the tritium label is in the sarcoplasmicreticulum. This later report of Fozzard andSmith18 gives no details concerning ultracen-trifugal fractionation, but we may remark thatin a sucrose density gradient it is essentiallyimpossible to separate thick and thin filamentsfrom other cellular debris, and that activationby mixtures of K+ and Na+ would normallynot discriminate between the ATPase of re-laxing factor and that of, say, myosin.
AddendumAfter the present work was presented in
abstracted lectures at the Universities of To-kyo and Washington, and at the College ofPhysicians and Surgeons of Columbia Univer-sity, Dr. Arnold Katz (Fed. Proc. 25: 643,1966) reported on work ostensibly designedto examine the main conclusions of this paper.Dr. Katz confirmed the insensitivity of myosinA ATPase to ouabain, but was otherwise ledto conclusions very divergent from ours. Inorder to facilitate authentic examination andextension of our present work we list usefulprecautions culled from our experience. (1)Factors affecting the degree of direct actin-myosin bonding are certain to be important.Choice of system may be such a factor; forexample, a "reconstituted" actomyosin systemis mechanically weaker (as a thread) andgives a much smaller percent turbidity changeon ATP addition than the myosin B systemused here. (2) Because of the intrinsic vari-ability in both types of measurement it is high-ly desirable to record OD and H+ liberation(ATP hydrolysis) continuously, but even so,the measurements are unavoidably erratic, sothat a very large number of data have to begathered to establish a conclusion. (3) TheATPase of myosin which is not directly bond-ed to actin is insensitive to ouabain. Accord-ingly, care must be taken to minimize the
CircuUtioH Rtittrcb, Vol. XIX, Stpltrnktr 1966
contribution of this component. For example,if a system contains much Mg2* (e.g. DIM)superprecipitation is preceded by an extreme-ly long "clearing phase," during which ATPis hydrolyzed by unbound, thus insensitive,myosin; likewise after superprecipitation isover, the persisting myosin A ATPase is in-sensitive; it is only the high ATP utilizationrate actually accompanying superprecipitationwhich is activated by ouabain and digoxin.(4) Other general features and useful pre-cautions pertaining to the techniques usedhere will be found in references 6, 8, and 16.
AcknowledgmentWe are very indebted to Dr. M. Honjoh of this
Medical Center, and to Dr. Lynn Anspaugh of theLawrence Radiation Laboratory, for performing thepurity assays on the ouabain which we have used.Professor Brian F. Hoffman of Columbia Universitymade available to us important results in advance ofpublication, and generally encouraged and counselledus in this work. Dr. Bernard Engel of this MedicalCenter gave us valuable advice on data display.
References1. SKOU, J. C : Further investigations on a Mg**
+ Na+-activated adenosintriphosphatase, possi-bly related to the active linked transport ofNa+ and K+ across the nerve membrane. Bio-chim. Biophys. Acta 42: 6, 1960.
2. POST, R. L., MERRITT, C. R., KINSOLVINC, C. R.,
AND ALBRICHT, C. D.: Membrane adenosine
triphosphatase as a participant in the activetransport of sodium and potassium in the hu-man erythrocyte. J. Biol. Chem. 235: 1796,1960.
3. BOWEN, W. J.: Effect of digoxin upon rate ofshortening of myosin B threads. FederationProc. 11: 16, 1952.
4. ROBB, J. S., AND MALLOV, S.: Effect of ouabain
on actomyosin threads. J. Pharmacol. Exptl.Therap. 108: 251, 1953.
5. EBASHI, S.: Calcium binding activity of vesicu-lar relaxing factor. J. Biochem. (Tokyo) 50:236, 1961.
6. YASUI, T., AND WATANABE, S.: Study of "super-
precipitation" of myosin B by the change inturbidity. J. Biol. Chem. 240: 98, 1965.
7. TONOMURA, Y., APPEL, P., AND MORALES, M. F.:
On the molecular weight of myosin, II. Bio-chemistry 5: 515, 1966.
8. RAINFORD, P., NOCUCHJ, H., AND MORALES, M.:
Hydrostatic pressure and the contractile sys-tem. Biochemistry 4: 1958, 1965.
9. KJELLEY, W. W., AND MEYERHOF, O.I Studies onATPase of muscle: III. Lipoprotein nature ofthe Mg-activated ATPase. J. Biol. Chem. 183:391, 1950.
10. SCHWARTZ, A.: Possible mechanism of action ofcardiac glycosides on an enzymatic level. Car-diovascular Res. Center Bull. 2: 73, 1964.See also Lee, K. S., Yu, D. H., and Struthers,J. J.: Study on the effect of cardiac glycosideson the syneresis of myofibrils in the presenceof relaxing factor. J. Pharmacol. Exptl. Therap.148: 277, 1965.
11. GELLERT, M. F., VON HIPPEL, P. H., SCHACH-
MAN, H. K., AND MORALES, M. F.: Studies on
the contractile proteins of muscle: I. ATP-myo-sin B interaction. J. Am. Chem. Soc. 81:1384, 1959.
12. REPKE, K.: Metabolism of cardiac glycosides. InNew Aspects of Cardiac Glycosides, Proceed-
ings of the First International Pharmacologi-cal Meeting, Berne. New York, PergamonPress, 1963, p. 68.
13. FOZZARD, H. A., Aj.T> SMITH, J. R.: Observationson the localization of tritiated digoxin in myo-cardial cells by autoradiography and ultra-microscopy. Am. Heart J. 69: 245, 1965.
14. SMITH, J. R., AND FOZZARD, H. A.: Localizationof tritiated "digoxin" in the myocardial cellsof frogs and dogs by autoradiography com-bined with electron microscopy. Nature 197:562, 1963.
15. SONNENBUCK, E. H., SPOTNITZ, H. M., ANDSPIRO, D.: Role of the sarcomere in ventricu-lar function and the mechanism of heart fail-ure. Circulation Res. 15 (suppl. 2) : 70, 1964.
16. RICE, R. V., ASAI, H., AND MORALES, M. F.: Con-formational changes in myosin B and acto-myosin induced by ATP. Proc. Nat. Acad. Sci.50: 549, 1963.
http://circres.ahajournals.org/content/19/3/496World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Research Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions: