CONTEMPORARY REVIEW Optical imaging of arrhythmias in the cardiomyocyte monolayer Herman D. Himel IV, PhD, * Gil Bub, PhD, † Prajapathy Lakireddy, MD, ‡z Nabil El-Sherif, MD, FHRS ‡z From * Research Triangle Institute International, Durham, North Carolina, y Department of Physiology, Oxford University, Oxford, United Kingdom, z Downstate Medical Center, State University of New York, Brooklyn, New York and z VA New York Harbor Healthcare System, Brooklyn, New York. In recent years, cultured cardiac cell monolayers have become a contemporary experimental preparation for the study of fundamental mechanisms that underlie normal and pathologic electrophysiology at the tissue level. Ion channels and gap junctions in the cardiomyocyte monolayer may be modulated using drugs that suppress or enhance certain channels/junctions, or by genetic silencing or overexpression. The cardiomyocyte monolayer is particularly well suited for studies of functional electrophysiologic properties of mixtures of cardiac and noncardiac cells (eg, myofibroblasts), which otherwise would be difficult to investigate. Optical mapping of monolayers has provided insight into mechanisms that can set the stage for arrhythmias, such as unidirectional conduction block, gap junction uncoupling, ische- mia, alternans, and anisotropy, and continues to enhance our under- standing of basic electrophysiologic mechanisms. KEYWORDS Alternans; Anisotropy; Arrhythmias; Myocardial ischemia; Myocyte–myofibroblast interactions; Optical mapping ABBREVIATIONS I/R ¼ ischemia/reperfusion; NRVM ¼ neonatal rat ventricular myocyte (Heart Rhythm 2012;9:2077–2082) I 2012 Elsevier Inc. All rights reserved. Introduction Cultured cardiac cell monolayers have become a contem- porary experimental preparation for the study of fundamental mechanisms that underlie normal and pathologic electro- physiology at the tissue level. 1 Cell monolayers are 2- dimensional sheets of cells that are obtained by seeding cell suspensions onto tissue culture surfaces conducive to cell attachment and spreading. Over the course of a few days, the cells form a highly simplified tissue system under the experimenter’s control. Methods of oriented or patterned growth via microabra- sion or micropatterning enable explicit control of the tissue architecture of the cell monolayer. 2,3 Neonatal rat ventricular myocytes (NRVMs) can be aligned in culture to produce simple or complex anisotropy. 3 Changes in fiber curvature can influence cardiac propagation 4 and arrhythmogenicity. 3 Monolayer patterning to simulate structural heterogeneity can promote triggered activity, reflection, and reentrant activity. 5 Multiparametric optical mapping is commonly used today to record the spread of electrical activity in the monolayer. Recent advances in potentiometric and calcium-sensitive dyes as well as newer detector technology have facilitated simultaneous optical mapping of transmembrane voltage and intracellular calcium in the monolayer, even with the use of a single photodetector. 6 The cardiac monolayer also allows investigators to per- form genetic and/or chemical modifications on the entire tissue with relative ease. The monolayer can be manipulated either by drugs that suppress or enhance certain ion channels/ gap junctions 7,8 or by adenoviral silencing or overexpres- sion. 9,10 These studies have added to our understanding of basic electrophysiologic mechanisms of arrhythmia genera- tion. Last, but not least, the cardiac monolayer is particularly well suited for studies of functional electrophysiologic properties of mixtures of cardiac and noncardiac cells such as myofibroblasts. 11 One limitation of the monolayer is that the cell type is largely limited to embryonic (chick) or neonatal (rat or mouse) heart cells, although this landscape may be changing as we develop our understanding of the induced pluripotent stem cell–derived cardiomyocyte. 12 Formation of the cardiac monolayer In a typical cardiac monolayer preparation, neonatal hearts are excised, minced finely, and then subjected to series of enzymatic digestions, which results in a cell suspension containing separated cardiomyocytes. The details of the digestion procedure vary according to the investigator, with the common goal of simultaneously maximizing cell yield and viability. Cell plating surfaces are prepared by coating with substances that are conducive to cell attachment and spreading (eg, fibronectin, laminin, gelatin). Cell suspensions typically are “preplated” for a short duration (1–2 hours) and then removed for longer-duration plating to allow cardiomyocyte attachment. Preplating steps serve to reduce the fraction of Supported in part by a MERIT grant from the VA Central Office to Dr. El-Sherif. Address reprint requests and correspondence: Dr. Nabil El-Sherif, Cardiology Division, VA NY Harbor Healthcare System, 800 Poly Place, Brooklyn, NY 11203. E-mail address: [email protected]. 1547-5271/$-see front matter B 2012 Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.hrthm.2012.08.035
Transcript
CONTEMPORARY REVIEW
Optical imaging of arrhythmias in the cardiomyocyte monolayer
Herman D. Himel IV, PhD,* Gil Bub, PhD,† Prajapathy Lakireddy, MD,‡z Nabil El-Sherif, MD, FHRS‡z
From *Research Triangle Institute International, Durham, North Carolina, yDepartment of Physiology, Oxford University,Oxford, United Kingdom, zDownstate Medical Center, State University of New York, Brooklyn, New York andzVA New York Harbor Healthcare System, Brooklyn, New York.
In recent years, cultured cardiac cell monolayers have become a
contemporary experimental preparation for the study of fundamental
mechanisms that underlie normal and pathologic electrophysiology at
the tissue level. Ion channels and gap junctions in the cardiomyocyte
monolayer may be modulated using drugs that suppress or enhance
certain channels/junctions, or by genetic silencing or overexpression.
The cardiomyocyte monolayer is particularly well suited for studies of
functional electrophysiologic properties of mixtures of cardiac and
noncardiac cells (eg, myofibroblasts), which otherwise would be
difficult to investigate. Optical mapping of monolayers has provided
insight into mechanisms that can set the stage for arrhythmias, such
Supported in part by a MERIT grant from the VA Central Office toDr. El-Sherif. Address reprint requests and correspondence: Dr. NabilEl-Sherif, Cardiology Division, VA NY Harbor Healthcare System, 800Poly Place, Brooklyn, NY 11203. E-mail address: [email protected].
1547-5271/$-see front matter B 2012 Elsevier Inc. All rights reserved.
as unidirectional conduction block, gap junction uncoupling, ische-
mia, alternans, and anisotropy, and continues to enhance our under-
(Heart Rhythm 2012;9:2077–2082) I 2012 Elsevier Inc. All rightsreserved.
IntroductionCultured cardiac cell monolayers have become a contem-porary experimental preparation for the study of fundamentalmechanisms that underlie normal and pathologic electro-physiology at the tissue level.1 Cell monolayers are 2-dimensional sheets of cells that are obtained by seeding cellsuspensions onto tissue culture surfaces conducive to cellattachment and spreading. Over the course of a few days, thecells form a highly simplified tissue system under theexperimenter’s control.
Methods of oriented or patterned growth via microabra-sion or micropatterning enable explicit control of the tissuearchitecture of the cell monolayer.2,3 Neonatal rat ventricularmyocytes (NRVMs) can be aligned in culture to producesimple or complex anisotropy.3 Changes in fiber curvaturecan influence cardiac propagation4 and arrhythmogenicity.3
Monolayer patterning to simulate structural heterogeneity canpromote triggered activity, reflection, and reentrant activity.5
Multiparametric optical mapping is commonly used todayto record the spread of electrical activity in the monolayer.Recent advances in potentiometric and calcium-sensitivedyes as well as newer detector technology have facilitatedsimultaneous optical mapping of transmembrane voltage andintracellular calcium in the monolayer, even with the use of asingle photodetector.6
The cardiac monolayer also allows investigators to per-form genetic and/or chemical modifications on the entiretissue with relative ease. The monolayer can be manipulatedeither by drugs that suppress or enhance certain ion channels/gap junctions7,8 or by adenoviral silencing or overexpres-sion.9,10 These studies have added to our understanding ofbasic electrophysiologic mechanisms of arrhythmia genera-tion. Last, but not least, the cardiac monolayer is particularlywell suited for studies of functional electrophysiologicproperties of mixtures of cardiac and noncardiac cells suchas myofibroblasts.11 One limitation of the monolayer is thatthe cell type is largely limited to embryonic (chick) orneonatal (rat or mouse) heart cells, although this landscapemay be changing as we develop our understanding of theinduced pluripotent stem cell–derived cardiomyocyte.12
Formation of the cardiac monolayerIn a typical cardiac monolayer preparation, neonatal hearts areexcised, minced finely, and then subjected to series ofenzymatic digestions, which results in a cell suspensioncontaining separated cardiomyocytes. The details of thedigestion procedure vary according to the investigator, withthe common goal of simultaneously maximizing cell yield andviability. Cell plating surfaces are prepared by coating withsubstances that are conducive to cell attachment and spreading(eg, fibronectin, laminin, gelatin). Cell suspensions typicallyare “preplated” for a short duration (1–2 hours) and thenremoved for longer-duration plating to allow cardiomyocyteattachment. Preplating steps serve to reduce the fraction of
Figure 1 Schematic of monolayer mapping apparatus. A: Contactfluorescence imaging system, which functions in a transillumination mode.B: Tandem lens system, which functions in an epi-illumination mode.C: Generic schematic of an apparatus for micromapping, also functioning inan epi-illumination configuration. Dotted lines represent filters in panelsA, B, and C or dichroic mirror in panels B and C. Dashed lines in panel Crepresent 2 positions of an adjustable turning mirror, used to directfluorescence into either the detector or the high-resolution camera, for co-registration of electrophysiologic signals with their corresponding cellularanatomy. Green and red arrows in panel C show paths of excitation andemission, respectively.
2078 Heart Rhythm, Vol 9, No 12, December 2012
fibroblasts/myofibroblasts, as these cells attach more quicklyto the preplating surface, leaving a higher percentage ofcardiomyocytes in suspension. The appropriate number ofcells per unit area must be determined so that there aresufficient cells to form a confluent monolayer while avoidingovercrowding. Cell media may be supplemented with chemi-cal agents that reduce proliferation of fibroblasts/myofibro-blasts, such as 5-bromo-20-deoxyuridine (BrdU), mitomycinC, or paclitaxel.
Imaging electrical activity in the monolayerMacroscopic optical mappingThere are 2 basic configurations for mapping the cardiacmonolayer at the macroscopic scale. The first configuration,known as contact fluorescence imaging, was introduced byEntcheva et al13 in 2000. This configuration uses a photodiodearray connected to a fiberoptic bundle. Each fiber is mapped toa separate photodiode, and the fiber bundle is placed in directproximity to the cardiac monolayer. No additional optics arerequired, and this method is capable of achieving very highsignal-to-noise ratios. In this configuration, there is no spacebetween the fiber bundle and the monolayer; therefore,illumination of the sample typically takes place in a transillu-mination configuration, with the excitation source beingdirected toward a translucent sample (Figure 1A).
The cardiac monolayer may also be mapped in an epi-illumination configuration. This configuration typically usesa tandem lens design, with 1 lens serving as the objectivewhile another serves as the condenser. In the tandem lensconfiguration, a pair of lenses is placed front to front. Thisconfiguration has been shown to have superior light-collecting abilities when observing “large” (1–2 cm)objects.14 Between the 2 lens, where ideally the beam iscollimated, an imaging cube may be placed in order toachieve parallel paths for both illumination and imaging(Figure 1B). Alternatively, 1 or more light sources may bepositioned at an oblique angle relative to the monolayersource, and the lens pair may be directly coupled without anoptical cube. In this imaging mode, no dichroic is necessary,and the excitation and emission filters are placed on the endof the excitation source and between the tandem lens and thedetector, respectively (not shown).
Microscopic optical mappingAlthough rates of 500 Hz are sufficient to track excitation wavesat the macroscopic scale (�1 cm), higher rates of acquisitionbecome necessary when observing microscopic propagation(o1 mm). At the microscopic scale, an excitation wavetraveling at 25 cm/s will traverse a 1-mm field in only 4 ms,necessitating acquisition rates of 2 kHz and preferably higherin order to properly resolve propagation parameters. As themodern fluorescence microscope typically has most compo-nents necessary to perform microscopic optical mapping(micromapping), it can often be achieved with less customwork than is necessary for the macroscopic system. Essentialcomponents include an inverted microscope with a side port for
the detector, a high-powered excitation source, and a heated,perfused chamber to maintain viability of the monolayer culturewhile sitting on the microscope stage. Micromapping may beperformed over a wide range of magnifications, ranging from4� to 60� and higher. At higher magnifications (410�),significant motion artifacts appear because cellular contractiledisplacements become large relative to the mapping field; thus,electromechanical uncouplers or signal processing methodsbecome necessary to remove motion signals. Early pioneeringwork in the lab of Kleber in micromapping showcased the utilityof this method for demonstrating basic principles of conductionand for convincingly illustrating consistency between opticaland electrical recordings at the microscale.15 Figure 1C shows ageneric micromapping apparatus.
Cardiac arrhythmias in the monolayerStudies in the monolayer have expanded our understandingof basic electrophysiologic mechanisms that underliearrhythmia generation. Spontaneously beating monolayersmay show unifocal rhythm typically arising close to theperiphery of the monolayer and conducting in a planardirection (Online Supplemental Material Movie 1). Boththe rate and duration of the focal rhythm varies from 1monolayer to another. Monolayers may show rhythms ofvarying complexity under certain situations, such as
Figure 2 Initiation of spiral loop reentry in themonolayer by programmed S1-S2 stimulation proto-col. Shown are selected frames of contour plots ofactivation times of intracellular calcium fluorescence(FCai) (arranged sequentially from left to right) of theS1 stimulus (top), S2 stimulus that induced a spiralwave reentry (middle), followed by sustained spiralwave reentry (bottom). Asterisks denote the site ofstimulation.
Himel et al Optical Imaging of Arrhythmias in the Cardiomyocyte Monolayer 2079
myocyte/myofibroblast uncoupling, some akin to ventricularfibrillation (Online Supplemental Material Movie 2). Func-tional reentry may be induced in the monolayer by pro-grammed stimulation and typically results in the formation ofa single reentrant spiral wave (Figure 2 and Online Supple-mental Material Movie 3).16 Bub et al17 investigated themechanism of spontaneous onset and termination of repeti-tive patterns (bursts) of spiral wave reentry in the embryonicchick monolayer. In the chick monolayer, reentrant patternswere usually in the form of double spiral waves in a figure-of-eight pattern (Figure 3, frames C and D, and Online
Figure 3 Anatomy of typical bursting activity. Recording of intracellularcalcium fluorescence obtained from a chick embryo ventricular myocytemonolayer. Upper trace shows fluorescence intensity during a single burst.Colored images show contour plots of activation times at several timesduring the burst indicated by the labels. A: Unidirectional block. The waveis initiated at a single site and propagates only to the left. B: Formationof 2 mirror-image rotor waves. C, D: Contour plots of the mirror-imagepair of rotors. E: Destruction of one of the rotors leaves a single rotor.F: Termination of the remaining rotor. (Reproduced with permission fromBub G, Glass L, Publicover NG, Shrier A. Bursting calcium rotorsin cultured cardiac myocyte monolayers. Proc Natl Acad Sci U S A1998;95:10283–10287.)
Supplemental Material Movie 4). Other studies have inves-tigated the electrophysiologic mechanisms of spiral wavebreakup in the 2-dimensional NRVM monolayer,18 andexperimental observations have been complemented bycomputational modeling studies.19
Role of heterogeneity and intercellularcoupling in wave propagation in the monolayerThere is increasing evidence that cardiac myocytes can beelectrotonically coupled to nonmyocytes, both in vitro andin vivo.20 Although they occupy a small portion of themyocardial tissue volume, cardiac fibroblasts account for 50%to 70% of the cells by number in the normal adult mammalianheart and even more in pathologic conditions, where differentia-tion into the myofibroblast phenotype occurs.21
Despite accumulating evidence of potential heterocellularelectrical coupling in the diseased myocardium and itsimplications in arrhythmogenesis, the electrophysiologicinterplay between myofibroblasts and their neighboringmyocytes has not been studied in detail.22 Myofibroblastsare unexcitable cells: their resting membrane potential is lessnegative than that of myocytes, and their membrane resis-tance is higher. These characteristics suggest that myofibro-blasts may function as a sink for electrical charge andfunction as short- and long-range conductors.11 Myofibro-blasts have been shown in vitro to promote arrhythmogenesisby direct modification of cardiomyocyte electrophysiologyfollowing establishment of heterocellular electrical couplingand act as passive electrical conduits of excitability. How-ever, propagation across myofibroblast conduits is substan-tially delayed, thereby promoting arrhythmogenic slow anddiscontinuous conduction.11 Furthermore, moderately polar-ized myofibroblasts were found to induce cell density–dependent depolarization of cardiomyocytes, which causesarrhythmogenic slow conduction due to reduction of fastINa.
11 The degree of heterocellular coupling as well as theratio of cardiomyocytes vs myofibroblasts has an impact onthe complexity of wave propagation.
There are several ways to study the role of heterogeneity andintercellular coupling in the monolayer. Bub et al7 investigated aspatially heterogeneous cardiac cell monolayer preparation whoseproperties can be modified by changing the density of cells and
Figure 4 Recordings of intracellular calcium transients from chickembryo ventricular myocyte monolayers. The monolayers are plated at 3different densities and under 3 different concentrations of the gap junctionalblocker a-glycerrhetinic acid (aGA). The trace above each panel shows theactivity for 50 seconds located at the 4 center pixels. The images arepresented as activation plots, which show the overlaid location of active cellsat 100-ms intervals. Depending on the experimental conditions, the mono-layers displayed target waves generated at a constant rate, stable spiral waves,spiral waves that spontaneously started and stopped, giving rise to a burstingrhythm, or irregular activity that consisted of a mix of target patterns and self-terminating spiral waves. (Reproduced with permission from Bub G, ShrierA, Glass L. Global organization of dynamics in oscillatory heterogeneousexcitable media. Phys Rev Lett J2005;94:028105.)
Figure 5 Intracellular calcium transient (FCai)) amplitude maps (left) andselected FCai tracing (right) from a 6-day-old neonatal rat ventricularmyocyte-myofibroblast coculture. Programmed stimulation induced a com-plex reentrant arrhythmia akin to ventricular fibrillation. Tracing 1 shows thepresence of a fairly regular fast spiral wave activity localized at the topsection of the monolayer (tracing 1); the rest of the monolayer showedmultiple slower incomplete reentrant wave fronts (tracings 2–4).
Figure 6 Immunohistochemical staining of the monolayer from the same
2080 Heart Rhythm, Vol 9, No 12, December 2012
by adding the drug a-glycerrhetinic acid, which uncouplesintercellular connections (Figure 4). Decreases in density andincreases in a-glycerrhetinic acid lead to similar effects.
The myocyte/myofibroblast ratio can be changed system-atically, and their percentage may be evaluated by immuno-histochemistry and correlated with wave propagation patterns.This is shown in Figures 5 and 6. In a study by Zlochiveret al10 using NRVM monolayers, the myocyte/myofibroblastratio was changed systematically and junctional coupling ofthe myofibroblasts was reduced or increased using silencingRNAi or adenoviral overexpression of connexin43, respec-tively. Both reentry frequency and conduction velocitydiminished with larger myocyte/myofibroblast area ratiosand complexity of propagation increased, resulting in wavefractionation and reentry multiplication. It should be noted,however, that even a 50% decrease in connexin43 in strandsof cultured neonatal ventricular myocytes from a heterozy-gous mice showed little to no effect on conduction velocity.23
experiment shown in Fig. 5. The areas occupied by stained cardiomyocytesand nonstained noncardiomyocytes in the coculture were quantified, and themorphologic characteristics of the monolayer were correlated with theelectrophysiologic findings. Areas occupied by myofibroblasts were quanti-fied based on color contrast. Cardiomyocytes stained red while the grayareas represent the noncardiomyocytes. The area occupied by noncardio-myocytes in this monolayer was quantified as 28% of the total.
Studies of ischemia/reperfusion relatedarrhythmias in the monolayerAlthough electrophysiologic consequences of global ische-mia have been characterized extensively, heterogeneous
electrophysiologic alterations during regional ischemiaare more challenging to study because of the complex3-dimensional geometry of the myocardium and itsvascular supply, particularly at the interface between normaland ischemic tissue. The monolayer provides a simple2-dimensional model for studies of the mechanismsof arrhythmias during regional ischemia/reperfusion (I/R).
In an early study, Arutunyan et al24 developed an experi-mental approach to study the effects of regional I/R in culturedNRVM monolayers using recordings of the calcium transient.A small area of injury was created using geometrically definedflows of control and ischemia-like solutions.
In 2004, Pitts and Toombs25 described a “coverslip” modelof regional I/R in NRVM monolayers that reproduced
Figure 7 Recording of intracellular calcium transient fluorescence (FCai)from a neonatal rat ventricular myocyte monolayer showing rate-dependentFCai alternans. The monolayer was perfused with anthopleurin-A, a toxinthat blocks sodium channel inactivation and results in bradycardia-depen-dent prolongation of the plateau of FCai simulating long QT syndrome type 3(shown in top panel when the monolayer was stimulated at 4-secondintervals). In this experiment, as in the clinical setting, FCai alternansdeveloped at relatively slow pacing rates (shown in bottom panel when themonolayer was stimulated at 1-second intervals).
Himel et al Optical Imaging of Arrhythmias in the Cardiomyocyte Monolayer 2081
metabolic and ultrastructural changes leading to I/R injury inintact cardiac muscle. de Diego et al26 used the coverslip modelto characterize the effects of acute regional I/R on electro-physiologic alterations and arrhythmias in NRVM monolayers.They found that regional I/R in this preparation recapitulatedmany of the electrophysiologic alterations previously describedin intact tissue, including conduction slowing, action potentialduration shortening, impaired Cai handling, spontaneous extra-systoles, and reentrant arrhythmias, with the border zoneplaying a critical role in the generation of arrhythmias.
Alternans and arrhythmogenesis in themonolayerT-wave alternans, an important marker of arrhythmia risk,arise from beat-to-beat alternation in the electromechanical
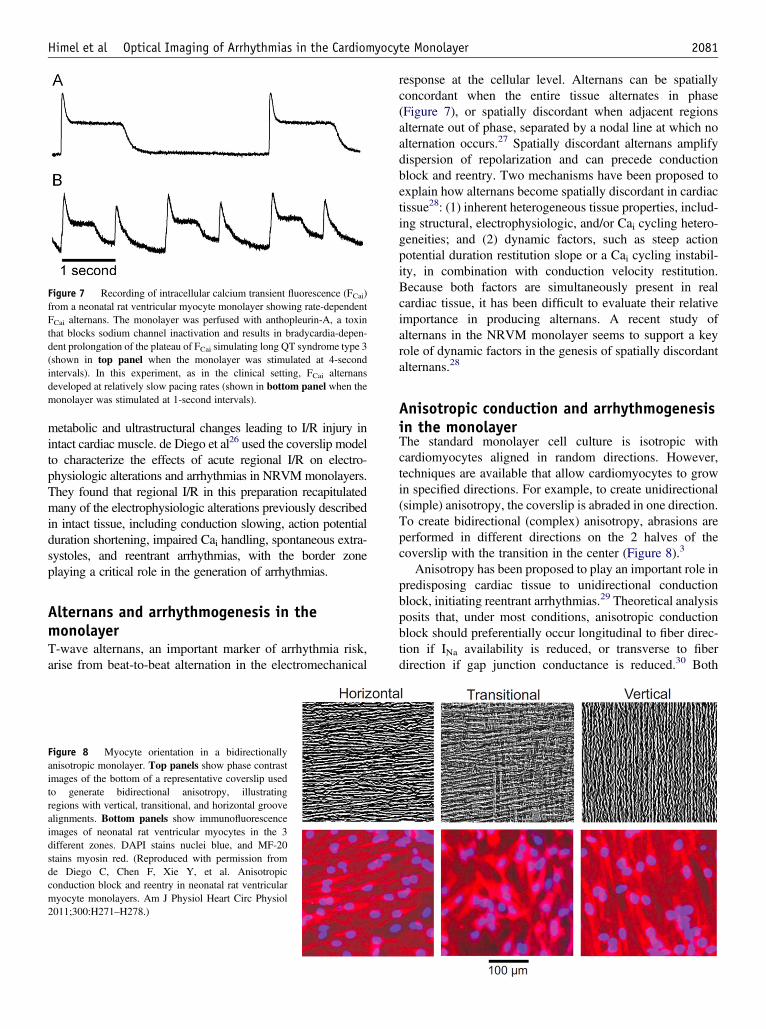
Figure 8 Myocyte orientation in a bidirectionallyanisotropic monolayer. Top panels show phase contrastimages of the bottom of a representative coverslip usedto generate bidirectional anisotropy, illustratingregions with vertical, transitional, and horizontal groovealignments. Bottom panels show immunofluorescenceimages of neonatal rat ventricular myocytes in the 3different zones. DAPI stains nuclei blue, and MF-20stains myosin red. (Reproduced with permission fromde Diego C, Chen F, Xie Y, et al. Anisotropicconduction block and reentry in neonatal rat ventricularmyocyte monolayers. Am J Physiol Heart Circ Physiol2011;300:H271–H278.)
response at the cellular level. Alternans can be spatiallyconcordant when the entire tissue alternates in phase(Figure 7), or spatially discordant when adjacent regionsalternate out of phase, separated by a nodal line at which noalternation occurs.27 Spatially discordant alternans amplifydispersion of repolarization and can precede conductionblock and reentry. Two mechanisms have been proposed toexplain how alternans become spatially discordant in cardiactissue28: (1) inherent heterogeneous tissue properties, includ-ing structural, electrophysiologic, and/or Cai cycling hetero-geneities; and (2) dynamic factors, such as steep actionpotential duration restitution slope or a Cai cycling instabil-ity, in combination with conduction velocity restitution.Because both factors are simultaneously present in realcardiac tissue, it has been difficult to evaluate their relativeimportance in producing alternans. A recent study ofalternans in the NRVM monolayer seems to support a keyrole of dynamic factors in the genesis of spatially discordantalternans.28
Anisotropic conduction and arrhythmogenesisin the monolayerThe standard monolayer cell culture is isotropic withcardiomyocytes aligned in random directions. However,techniques are available that allow cardiomyocytes to growin specified directions. For example, to create unidirectional(simple) anisotropy, the coverslip is abraded in one direction.To create bidirectional (complex) anisotropy, abrasions areperformed in different directions on the 2 halves of thecoverslip with the transition in the center (Figure 8).3
Anisotropy has been proposed to play an important role inpredisposing cardiac tissue to unidirectional conductionblock, initiating reentrant arrhythmias.29 Theoretical analysisposits that, under most conditions, anisotropic conductionblock should preferentially occur longitudinal to fiber direc-tion if INa availability is reduced, or transverse to fiberdirection if gap junction conductance is reduced.30 Both
2082 Heart Rhythm, Vol 9, No 12, December 2012
types of conduction block have been observed experimen-tally in anisotropic cardiac tissue, but factors related to3-dimensional tissue geometry, fiber rotation, nonuniformanisotropy due to tissue fibrosis, variable pacing protocols,and possible influences of unmapped subsurface events havecomplicated the interpretation of these experiments. TheNRVM monolayer provides an optimal 2-dimensionalexperimental model to investigate in more detail the role ofanisotropy on conduction block and arrhythmia generation.In a recent study of anisotropy in the NRVM monolayer,de Diego et al3 showed that in anisotropic monolayers,unidirectional conduction block initiating reentry can occurlongitudinal or transverse to fiber direction, depending onwhether the experimental intervention reduces INa availabil-ity or decreases gap junction conductance. This observationis in agreement with previous theoretical predictions.
Future directionsNovel methods for chemical, genetic, and topologic modula-tion of the cardiomyocyte monolayer will continue to offer abetter understanding of basic electrophysiologic principles.In addition, the availability of human stem cell–derivedcardiomyocytes may lend additional credibility to thecardiomyocyte monolayer as a clinically relevant in vitromodel.12
The recent combination of optical imaging methods withtargeted expression of protein-based fluorescent probes hasalready had great impact on neuroscience.31 Although spear-headed by tools to control membrane voltage, the moregeneral concept of optogenetics includes the use of a varietyof genetically encoded probes of physiologic parameters,ranging from membrane voltage and calcium concentrationto metabolic and cytoskeletal targets. Optogenetics promisesto open a new and exciting paradigm for modulation ofcardiac electrophysiologic function.32
AppendixSupplementary dataSupplementary data associated with this article can be foundin the online version at doi:10.1016/j.hrthm.2012.08.035.
References1. Herron TJ, Lee P, Jalife J. Optical imaging of voltage and calcium in cardiac cells
& tissues. Circ Res 2012;110:609–623.2. Badie N, Bursac N. Novel micropatterned cardiac cell cultures with realistic
ventricular microstructure. Biophys J 2009;96:3873–3885.3. de Diego C, Chen F, Xie Y, et al. Anisotropic conduction block and reentry in
neonatal rat ventricular myocyte monolayers. Am J Physiol Heart Circ Physiol2011;300:H271–H278.
4. Bourgeois EB, Fast VG, Collins RL, Gladden JD, Rogers JM. Change inconduction velocity due to fiber curvature in cultured neonatal rat ventricularmyocytes. IEEE Trans Biomed Eng 2009;56:855–861.
5. Auerbach DS, Grzda KR, Furspan PB, Sato PY, Mironov S, Jalife J. Structuralheterogeneity promotes triggered activity, reflection and arrhythmogenesis incardiomyocyte monolayers. J Physiol 2011;589:2363–2381.
6. Lee P, Bollensdorff C, Quinn TA, Wuskell JP, Loew LM, Kohl P. Single-sensorsystem for spatially-resolved, continuous and multi-parametric optical mappingof cardiac tissue. Heart Rhythm M 2011;8:1482–1491.
7. Bub G, Shrier A, Glass L. Global organization of dynamics in oscillatoryheterogeneous excitable media. Phys Rev Lett J2005;94:028105.
8. Lin JW, Garber L, Qi YR, Chang MG, Cysyk J, Tung L. Region [corrected] ofslowed conduction acts as core for spiral wave reentry in cardiac cell monolayers.Am J Physiol Heart Circ Physiol 2008;294:H58–H65.
9. Hou L, Deo M, Furspan P, et al. A major role for HERG in determining frequencyof reentry in neonatal rat ventricular myocyte monolayer. Circ Res 2010;107:1503–1511.
10. Zlochiver S, Munoz V, Vikstrom KL, Taffet SM, Berenfeld O, Jalife J.Electrotonic myofibroblast-to-myocyte coupling increases propensity to reentrantarrhythmias in two-dimensional cardiac monolayers. Biophys J 2008;95:4469–4480.
11. Rohr S. Myofibroblasts in diseased hearts: new players in cardiac arrhythmias?Heart Rhythm 2009;6:848–856.
12. Lee P, Klos M, Bollensdorff C, et al. Simultaneous voltage and calcium mappingof genetically purified human induced pluripotent stem cell-derived cardiacmyocyte monolayers. Circ Res 2012;110:1556–1563.
13. Entcheva E, Lu SN, Troppman RH, Sharma V, Tung L. Contact fluorescenceimaging of reentry in monolayers of cultured neonatal rat ventricular myocytes.J Cardiovasc Electrophysiol 2000;11:665–676.
15. Fast VG, Kleber AG. Microscopic conduction in cultured strands of neonatal ratheart cells measured with voltage-sensitive dyes. Circ Res 1993;73:914–925.
16. Iravanian S, Nabutovsky Y, Kong CR, Saha S, Bursac N, Tung L. Functionalreentry in cultured monolayers of neonatal rat cardiac cells. Am J Physiol HeartCirc Physiol 2003;285:H449–H456.
17. Bub G, Glass L, Publicover NG, Shrier A. Bursting calcium rotors in culturedcardiac myocyte monolayers. Proc Natl Acad Sci U S A 1998;95:10283–10287.
18. Bursac N, Tung L. Acceleration of functional reentry by rapid pacing inanisotropic cardiac monolayers: formation of multi-wave functional reentries.Cardiovasc Res 2006;69:381–390.
19. Agladze K, Kay MW, Krinsky V, Sarvazyan N. Interaction between spiral andpaced waves in cardiac tissue. Am J Physiol Heart Circ Physiol 2007;293:H503–H513.
20. Kohl P, Camelliti P. Cardiac myocyte-nonmyocyte electrotonic coupling:implications for ventricular arrhythmogenesis. Heart Rhythm 2007;4:233–235.
21. Vasquez C, Mohandas P, Louie KL, Benamer N, Bapat AC, Morley GE.Enhanced fibroblast-myocyte interactions in response to cardiac injury. Circ Res2010;107:1011–1020.
23. Thomas SP, Kucera JP, Bircher-Lehmann L, Rudy Y, Saffitz JE, Kleber AG.Impulse propagation in synthetic strands of neonatal cardiac myocytes withgenetically reduced levels of connexin43. Cir Res 2003;92:1209–1216.
24. Arutunyan A, Webster DR, Swift LM, Sarvazyan N. Localized injury incardiomyocyte network: a new experimental model of ischemia-reperfusionarrhythmias. Am J Physiol Heart Circ Physiol 2001;280:H1905–H1915.
25. Pitts KR, Toombs CF. Coverslip hypoxia: a novel method for studying cardiacmyocyte hypoxia and ischemia in vitro. Am J Physiol Heart Circ Physiol2004;287:H1801–H1812.
26. de Diego C, Pai RK, Chen F, et al. Electrophysiological consequences of acuteregional ischemia/reperfusion in neonatal rat ventricular myocyte monolayers.Circulation 2008;118:2330–2337.
27. Hayashi H, Shiferaw Y, Sato D, et al. Dynamic origin of spatially discordantalternans in cardiac tissue. Biophys J 2007;92:448–460.
28. de Diego C, Pai RK, Dave AS, et al. Spatially discordant alternans incardiomyocyte monolayers. Am J Physiol Heart Circ Physiol 2008;294:H1417–H1425.
29. Spach MS, Miller WT, 3rd, Geselowitz DB, Barr RC, Kootsey JM, Johnson EA.The discontinuous nature of propagation in normal canine cardiac muscle.Evidence for recurrent discontinuities of intracellular resistance that affect themembrane currents. Circ Res 1981;48:39–54.
30. Delmar M, Michaels DC, Johnson T, Jalife J. Effects of increasing intercellularresistance on transverse and longitudinal propagation in sheep epicardial muscle.Circ Res 1987;60:780–785.
31. Dugue GP, Akemann W, Knopfel T. A comprehensive concept of optogenetics.Prog Brain Res 2012;196:1–28.
32. Bruegmann T, Malan D, Hesse M, et al. Optogenetic control of heart musclein vitro and in vivo. Nat Methods 2011;7:897-90