Copyright q American Museum of Natural History 2003 ISSN 0003-0082 PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3402, 32 pp., 21 figures, 1 table March 27, 2003 Osteology and Relationships of Byronosaurus jaffei (Theropoda: Troodontidae) PETER J. MAKOVICKY, 1 MARK A. NORELL, 2 JAMES M. CLARK, 3 AND TIMOTHY ROWE 4 ABSTRACT The troodontid Byronosaurus jaffei is known from two specimens from adjacent localities in the Nemegt basin, O ¨ mno ¨gov Aimag, Mongolia. These specimens are composed of well- preserved cranial material and fragmentary postcrania. All of these elements are described here. Byronosaurus jaffei is included in a comprehensive phylogenetic analysis of Coeluro- sauria to ascertain its relationships. Several interesting characters of Byronosaurus jaffei have implications both for theropod relationships and for understanding patterns of variation within coelurosaurian theropods. These include the position of a foramen that marks the exit of the supra-alveolar canal (which we suggest is homologous with the subnarial foramen), the flat- tened internarial bar, the unusual interfenestral bar, and the unserrated teeth. Additionally, the well-preserved braincase allows detailed comparison with other troodontid taxa. INTRODUCTION The remains of troodontid dinosaurs are extremely rare, and only nine taxa have been 1 Assistant Curator, Department of Geology, The Field Museum, Roosevelt Rd. at Lake Shore Drive, Chicago IL 60605. Research Associate, Division of Paleontology, American Museum of Natural History. e-mail: pmakovicky@ fieldmuseum.org 2 Chairman, Division of Paleontology, American Museum of Natural History. e-mail: [email protected]3 Ronald S. Weintraub Associate Professor Department of Biological Sciences, George Washington University, Washington, DC 20052. Research Associate, Division of Paleontology, American Museum of Natural History. e-mail: [email protected]4 J. Nalle Gregory Regents Professor of Geology, and Director, Vertebrate Paleontology Laboratory, Department of Geological Sciences, The University of Texas at Austin, TX 78712. e-mail: [email protected]described, most from Central Asia (Osmo ´l- ska and Barsbold, 1990; Russell and Dong, 1993). Here we provide a detailed descrip- tion of Byronosaurus jaffei, a troodontid
Transcript
Copyright q American Museum of Natural History 2003 ISSN 0003-0082
P U B L I S H E D B Y T H E A M E R I C A N M U S E U M O F N AT U R A L H I S T O RY
CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024
Number 3402, 32 pp., 21 figures, 1 table March 27, 2003
Osteology and Relationships ofByronosaurus jaffei (Theropoda: Troodontidae)
PETER J. MAKOVICKY,1 MARK A. NORELL,2 JAMES M. CLARK,3 ANDTIMOTHY ROWE 4
ABSTRACT
The troodontid Byronosaurus jaffei is known from two specimens from adjacent localitiesin the Nemegt basin, Omnogov Aimag, Mongolia. These specimens are composed of well-preserved cranial material and fragmentary postcrania. All of these elements are describedhere. Byronosaurus jaffei is included in a comprehensive phylogenetic analysis of Coeluro-sauria to ascertain its relationships. Several interesting characters of Byronosaurus jaffei haveimplications both for theropod relationships and for understanding patterns of variation withincoelurosaurian theropods. These include the position of a foramen that marks the exit of thesupra-alveolar canal (which we suggest is homologous with the subnarial foramen), the flat-tened internarial bar, the unusual interfenestral bar, and the unserrated teeth. Additionally, thewell-preserved braincase allows detailed comparison with other troodontid taxa.
INTRODUCTION
The remains of troodontid dinosaurs areextremely rare, and only nine taxa have been
1 Assistant Curator, Department of Geology, The Field Museum, Roosevelt Rd. at Lake Shore Drive, Chicago IL60605. Research Associate, Division of Paleontology, American Museum of Natural History. e-mail: [email protected]
2 Chairman, Division of Paleontology, American Museum of Natural History. e-mail: [email protected] Ronald S. Weintraub Associate Professor Department of Biological Sciences, George Washington University,
Washington, DC 20052. Research Associate, Division of Paleontology, American Museum of Natural History. e-mail:[email protected]
4 J. Nalle Gregory Regents Professor of Geology, and Director, Vertebrate Paleontology Laboratory, Departmentof Geological Sciences, The University of Texas at Austin, TX 78712. e-mail: [email protected]
described, most from Central Asia (Osmol-ska and Barsbold, 1990; Russell and Dong,1993). Here we provide a detailed descrip-tion of Byronosaurus jaffei, a troodontid
2 NO. 3402AMERICAN MUSEUM NOVITATES
Fig. 1. The holotype locality looking north. The arrow signifies the point of discovery of IGM 100/983.
from the Djadokhta Formation at Ukhaa Tol-god that was named and briefly described byNorell et al. (2000). Only one other troodon-tid, Saurornithoides mongoliensis (Osborn,1924), has been reported from Djadokhta, orDjadoktha-type, beds in Asia (Currie andDong, 2001a). Although troodontids are rare,well-preserved braincases are known for sev-eral taxa (Osborn, 1924; Barsbold, 1974;Currie, 1985; Currie and Zhao, 1993; Russelland Dong, 1993, Xu et al., 2002), and haveprovided important phylogenetic insightsinto this clade.
Few phylogenetic studies within Troodon-tidae have been undertaken, and broaderstudies that include troodontids and their pur-ported closest relatives (dromaeosaurids[Gauthier, 1986; Sereno, 1999], birds [Holtz,2001]) have not taken into account the vari-ation among troodontid taxa and have beenhindered by fragmentary material and inad-equate descriptions. Here we expand our spe-cies-level phylogenetic analysis of the group(Norell et al., 2000) and examine the evi-dence for monophyly of this group and moreexclusive clades within it. The evidence forthe relationships of troodontids to other
groups will be considered in more detail else-where.
MATERIAL
LOCALITY: The holotype specimen (IGM100/983) was discovered at the sublocalitycalled ‘‘Ankylosaur Flats’’ in 1993 by Mi-chael Novacek (fig. 1). Additional remains ofthis specimen were collected during the 1994and 1995 field seasons. Unlike many of thefossils at Ukhaa Tolgod, the specimen wasnot articulated and was found as a loose as-sociation of bones. Nevertheless, the rostralfragment and the braincase were found inplace adjacent to one another in life position,separated by a zone of weathering. The re-maining parts of the skeleton were collectedas eroded pieces below the skull. Later ex-amination of the specimen revealed that re-mains of a second individual were inter-mixed with IGM 100/983. This individual(IGM 100/987) is significantly larger thanIGM 100/983, and consequently its bonescan easily be separated. This specimen lackstroodontid apomorphies, and is referred to
2003 3MAKOVICKY ET AL.: BYRONOSAURUS JAFFEI
the Ornithomimidae (Makovicky and Norell,1998).
REFERRED SPECIMEN: A second specimen(IGM 100/984) was collected on July 15,1996 at the ‘‘Bolor’s Hill’’ locality approxi-mately 6 km west of the main Ukhaa Tolgodexposure. While extremely fragmentary, thespecimen preserves some important featuresnot present on the type. The specimen con-sists of a rostrum in six pieces. It is referredto Byronosaurus jaffei on the basis of super-numary teeth all lacking serrations, distinc-tive lacrimals with a lateral shelf that over-hangs the anterodorsal corner of the orbit,and a pair of passages through the interfe-nestral bar, which is not recessed from theplane of the rostrum.
DIAGNOSIS
Byronosaurus jaffei exhibits the followingtroodontid synapomorphies (Xu et al., 2002):numerous teeth, close packing of dentarydentition near the rostral tip of the lower jaw,presence of a distinct groove for the neuro-vascular foramina on the dentary, and a dor-soventrally flattened internarial bar.
Byronosaurus jaffei is part of a clade com-prising all troodontids except the basal taxonSinovenator changi. This clade is ambigu-ously diagnosed by: braincase displaying alarge pneumatic sinus (the ‘‘lateral depres-sion’’ ventral and anteroventral to the brain-case; presence of a subotic recess; extensiveanterior process of the lacrimal forming thedorsal border of the antorbital fenestra; andmaxillary participation in the posterior mar-gin of the nares. Ambiguity in the optimi-zation of these characters is due to missingdata and multiple placements of several troo-dontid taxa, including Byronosaurus jaffei,Sinornithoides youngi, and an unnamedMongolian taxon represented by IGM 100/44.
Byronosaurus jaffei can be distinguishedfrom all other troodontids by the possessionof several derived characters, including teethlacking serrations, an interfenestral bar thatis not recessed from the plane of the maxilla,and a shallow groove along the buccal mar-gin of the maxilla.
DESCRIPTION
The skull of IGM 100/983 is preserved intwo parts. The anterior section comprises therostrum and articulated dentaries that havebeen transversely compressed although mostbones appear uncrushed (figs. 2, 3). It wasfound in two pieces that have been glued to-gether, and the glue fills some large gapsalong the contact. The anterior section endsat the anterior margin of the orbits. Thebraincase was preserved separately and hasnot been crushed. The top of the braincase ismissing, exposing an eroded endocast, andsome delicate outer parts of the basicraniumare incomplete.
IGM 100/984 is very fragmentary. It waspreserved as eight separate nodules, onlythree of which contain recognizable ele-ments. One contains the premaxillae and theanterior ends of the maxillae (fig. 4), a sec-ond contains a section of the rostrum at thelevel of the interfenestral bar, and a thirdcontains a section of skull roof at the preor-bital bar preserving the lacrimals and theposterior part of the nasals.
Our description is primarily based on IGM100/983, supplemented by IGM 100/984.Measurements of the holotype IGM 100/983are tabulated in appendix 2. Comparisonswith Saurornithoides junior are based onBarsbold (1974, 1983) and are supplementedby our own observations. Comparisons withTroodon formosus are based on Currie (1985)and Currie and Zhao (1993), and those withSinornithoides youngi are based on Russelland Dong (1993) and our examination of thespecimen. Comparisons with Saurornithoidesmongoliensis are based on our own observa-tions of the type (AMNH 6174). Other com-parisons are referenced herein.
The face and braincase of the type speci-men (IGM 100/983) were scanned separatelyin May 1997 at The University of TexasHigh-resolution X-ray Computed Tomogra-phy Facility. Both were scanned on the high-resolution subsystem, based on a Feinfocusmicrofocal X-ray tube and an image-inten-sifier detector system. The face was scannedwith X-ray energy set at 120 kV, 0.200 mA,at 190% offset in an air wedge, with asource-object distance of 85 mm. Slice thick-ness is 0.25 mm, and 562 consecutive slices
4 NO. 3402AMERICAN MUSEUM NOVITATES
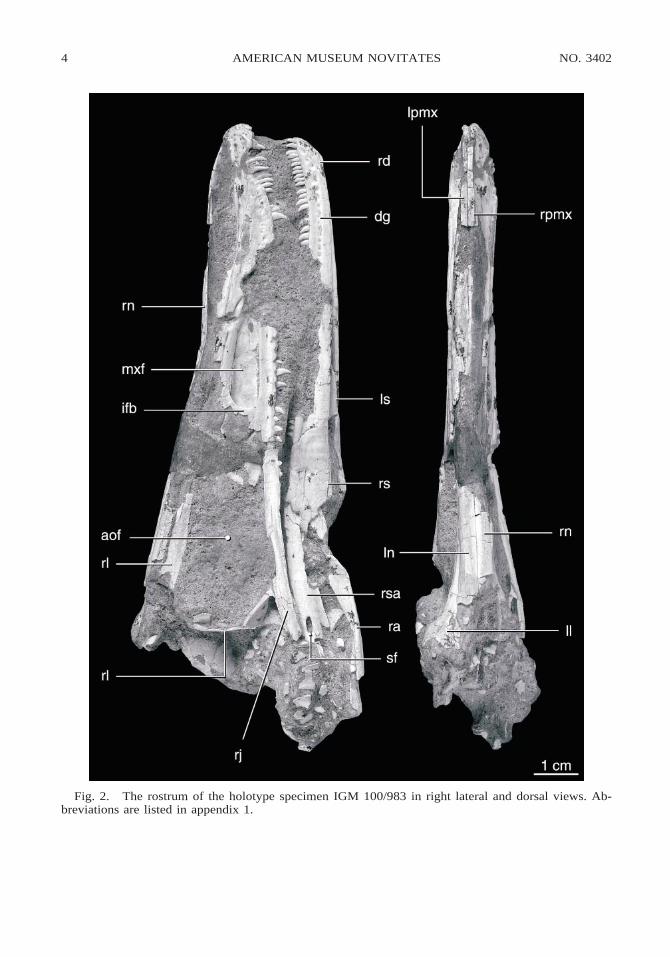
Fig. 2. The rostrum of the holotype specimen IGM 100/983 in right lateral and dorsal views. Ab-breviations are listed in appendix 1.
2003 5MAKOVICKY ET AL.: BYRONOSAURUS JAFFEI
Fig. 3. The rostrum of the holotype specimen IGM 100/983 in left lateral and ventral view. Abbre-viations are listed in appendix 1.
6 NO. 3402AMERICAN MUSEUM NOVITATES
Fig. 4. Stereopair of the ventral surface of the premaxillary area in the referred specimen IGM 100/984.
were generated. For each slice, 1200 viewswere taken with two samples per view. Re-constructed 512 3 512 pixel imagery wasexported as 8-bit TIF files. The braincasewas scanned with X-ray energy set at 120kV, 0.400 mA, at 190% offset in an airwedge, with a source-object distance of 45mm. Slice thickness is 0.20 mm, and 150consectutive slices were generated. For eachslice, 1200 views were taken with two sam-ples per view, and a field of view of approx-imately 36.3 mm. Reconstructed 1024 31024 pixel imagery was exported as 8 bitTIF files. Original CT datasets were archivedon CD-ROM at the University of Texas andthe American Museum of Natural History.Animated serial section movies and three di-mensional volumetric reconstructions arealso viewable on the Internet at www.DigiMorph.org.
THE ROSTRUM
PREMAXILLA: Paired premaxillae form theanterior margin of the skull. Each premaxilla
bears four teeth (fig. 4), as is primitive fortheropods. The nasal process is gently archedand dorsally flattened and triangular in crosssection (fig. 2), and it forms the anterodorsalmargin of the external narial opening. It isthin anteriorly and increases in breadth in thearea where it divides the nasals along themidline, as in Saurornithoides junior. Themaxillary process is short and heavily pittedby small foramina on its small facial surface.Medially, the surface of the maxillary pro-cess arches to form the floor of the naris (fig.2). There is no posterior process as in dro-maeosaurids that excludes the maxilla fromparticipation in the margin of the narial open-ing (figs. 2, 3). Although obscured, the suturewith the maxilla is apparently very simple.
MAXILLA: The maxilla forms most of thelateral surface of the skull. Anteriorly, itforms much of the floor of the large naris.The maxilla contacts the premaxilla along astraight suture anteriorly in this chamber(figs. 4, 5). Just posterior to this suture, andbefore the maxilla rises from the narial floor
2003 7MAKOVICKY ET AL.: BYRONOSAURUS JAFFEI
Fig. 4. Continued.
Fig. 5. Detail of the narial region of the ho-lotype specimen IGM 100/983 in left lateral view.Abbreviations are listed in appendix 1.
to form the posterodorsal margin of the naris,lies a large, anteriorly directed foramen thatis completely enclosed by the maxilla (figs.5, 6). A similar foramen is observed in thealvarezsaurid Shuvuuia deserti (IGM 100/977). This foramen enters into a long supra-alveolar canal that parallels the tooth row(fig. 6). The maxilla rises from the narialfloor in a gentle posterior slope. The lateralsurface below the narial chamber is coveredwith small pits and foramina (fig. 5). Theseare most apparent as a row of large pits thatlie in a groove just dorsal and parallel to thedentigerous margin. This row of pits runsposteriorly below the maxillary fenestra, de-creasing in density posteriorly until they dis-appear below the antorbital fenestra.
The lateral surface of the maxilla is per-forated by two large, elongated antorbitalopenings (figs. 2, 3). The maxillary fenestrais elliptical and extends from below the ex-ternal nares to the midway point between themaxillary-premaxillary suture and the pre-orbital bar. This fenestra has an osseous me-dial wall, which is perforated by a small holein slightly different positions on each side,but these holes may be weathering artifacts.
In Troodon formosus the maxillary fenestrais backed by an unperforated osseous wall.(Brochu, personal commun.). Anteriorly, therostral end of the maxilla overhangs the max-illary fenestra to form a large pocket. Themaxillary fenestra is separated from the ant-orbital fenestra by a large, very flat plate ofmaxilla, here termed the interfenestral bar(figs. 2, 7). Unlike Saurornithoides junior,Saurornithoides mongoliensis, or Sinorni-thoides youngi (Currie and Dong, 2001a), theinterfenestral bar is not inset from the planeof the maxilla.
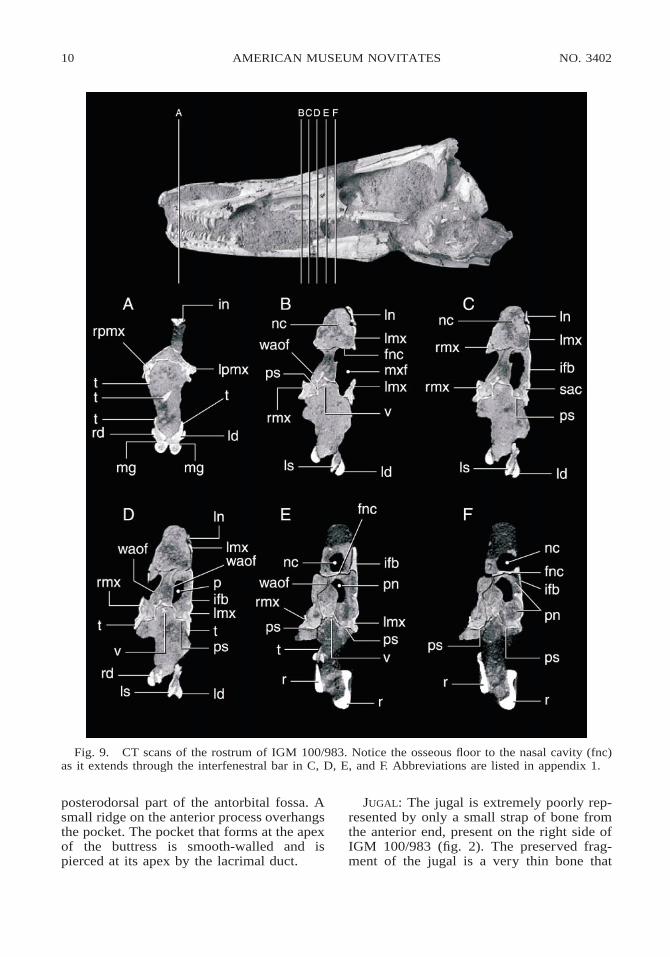
The main antorbital fenestra lacks an os-seous floor, is larger than the anterior fenes-tra, is subtrapezoidal in shape, and narrowsdorsally. Anteriorly, the margin of the fenes-tra is formed by the interfenestral bar, whichis scalloped in two places (fig. 7). Theseemarginations lead to dorsal and ventral pas-sages through the interfenestral bar, whichare separated by an osseous wall (fig. 7). Anerosional cross section of the snout of IGM100/984 exposes the course of the two chan-nels through the interfenestral bar (fig. 8).The dorsal emargination marks the caudalend of the narial passage, which is visiblelaterally as a swollen ridge on the dorsal partof the osseous wall of the maxillary fenestra(figs. 9, 10). The ventral canal connects theantorbital and maxillary fenestrae. The cau-dal entrance into the supra-alveolar canal,which transmits the innervation and bloodsupply of the maxillary dentition, lies on the
8 NO. 3402AMERICAN MUSEUM NOVITATES
Fig. 6. Stereopair of the right dorso-oblique dorsal surface of the rostrum in the referred specimenIGM 100/984. Notice the entrance to the supra-alveolar canal (fsac) which we interpret as the subnarialforamen. Abbreviations are listed in appendix 1.
medial face of the interfenestral bar (fig. 9C).Below the antorbital fenestra, the maxilla isextremely thin, dorsoventrally concave, anddevoid of the small foramina that are so con-spicuous anteriorly.
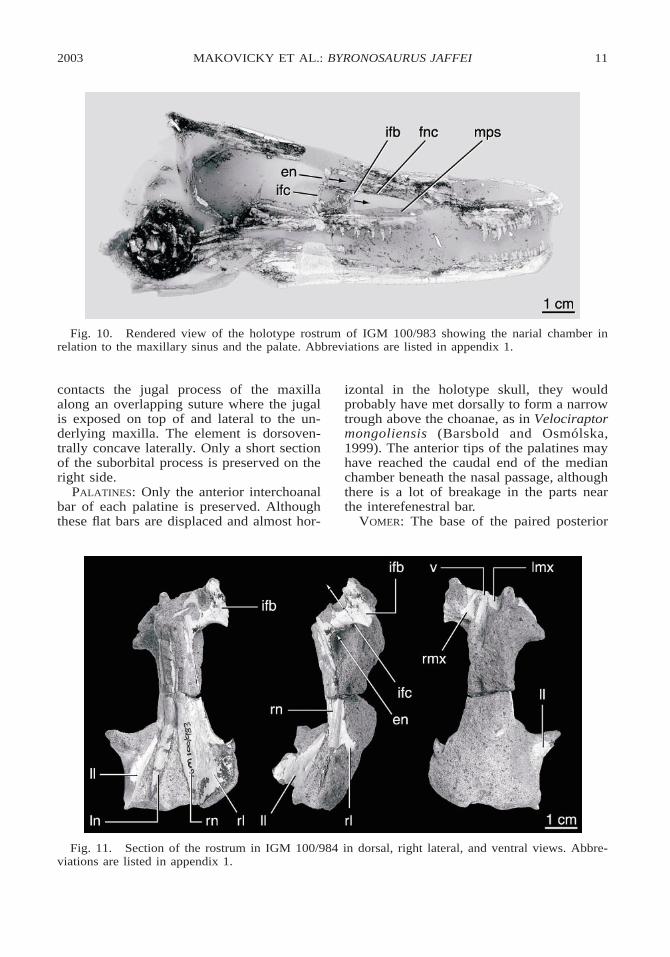
On the palate of IGM 100/984, longitudi-nally extensive palatal shelves of the maxillaform the roof of the mouth (figs. 4, 9, 10).These large palatal flanges extend posteriorlyfrom the contact with the premaxilla, and arejoined by a median vomer at the border ofthe choanae (figs. 11, 12. Together theseform a solid secondary palate on the roof ofthe mouth extending to the choanae, whichlie between the antorbital fenestrae.
NASAL: The nasal is a long bone that cov-ers the top of the rostrum for most of itslength (fig. 2). Anteriorly, the nasal forms theposterolateral boundary of the large externalnaris. The nasal is wider anteriorly than pos-teriorly. As preserved, anteriorly the nasal islaterally convex, forming a continuous tran-sition from a vertical plane anteriorly to ahorizontal plane posteriorly. This transition ismarked by a sharp angle, but the angle maybe influenced by postmortem distortion. Amaxillary process extends posteroventrally,while a long, attenuate premaxillary processextends anteriorly to form the posterodorsalborder of the naris for half its length and con-tact the nasal process of the premaxilla me-dially (fig. 5). This process gives the anteriorsurface of each nasal a concave appearancein lateral view. The lateral edge of the nasalcontacts the maxilla anteriorly and the lacri-mal posteriorly along a straight suture. Par-allel and just medial to this suture lies a row
of tiny foramina within a shallow depression.The posterior contact with the frontal is notpreserved on the type specimen, but is visi-ble on IGM 100/984. The nasals end justposterior to the level of the preorbital bar. Atthe level of the preorbital bar the nasals di-verge along the midline (fig. 13) to accom-modate a missing anterior wedge of the fron-tal, as in Saurornithoides junior. Posterior tothe preorbital bar on the top of the skull thenasals are flat, and together with the lacri-mals they form a flat surface above the an-terior part of the orbits (fig. 13).
LACRIMAL: The lacrimal forms the anteriorand anterodorsal border of the orbit. In lat-eral view the lacrimal is T-shaped, as in dro-maeosaurids; however, the anterior or supra-antorbital fenestral process is much longerthan the supraorbital process (figs. 2, 3). Al-though poorly preserved, several importantfeatures are apparent. The preorbital bar is astraplike process extending from the skullroof to contact the jugal. The preorbital baris oriented nearly vertically; however, thisorientation may be influenced by distortion.Contact with the jugal is preserved only onthe right side, where a posterolaterally open-ing foramen appears to be present, but thisarea is broken (fig. 2). The posterior surfaceis slightly convex in cross section. CT scansreveal that the lacrimal duct exits the orbitjust ventral to the contact between the ante-rior and posterior processes. Currie and Dong(2001a) suggested that a lacrimal duct is ab-sent in troodontids, but in Byronosaurus thelacrimal duct passes through the lacrimal andopens into the posterodorsal corner of the
2003 9MAKOVICKY ET AL.: BYRONOSAURUS JAFFEI
Fig. 6. Continued.
Fig. 8. Antero-ventro oblique view of the re-ferred specimen (IGM 100/984) showing the os-seous wall of the interantorbital canal (iac) sepa-rating it from the nasal chamber just anterior tothe interfenestral bar. Abbreviations are listed inappendix 1.
Fig. 7. Postero-oblique view of the left sideof the rostrum of IGM 100/983 showing the twopassages through the interfenestral bar. The upperpassage (en) connects with the nasal chamber. Thelower passage (iac) connects with the maxillarysinus.
antorbital fossa, as observed in IGM 100/984. The posterior, or supraorbital, process ispreserved only at its base on the type. OnIGM 100/984 the posterior process is appar-ently completely preserved on the right lac-rimal (fig. 13). In this specimen it extends tothe same level posteriorly as the nasals. Theposterior process forms a laterally expandedshelf above the orbit, as in Saurornithoidesjunior, and it apparently meets the frontalalong a posteromedial-anterolateral diagonalsuture (fig. 13). Just posterior to the level ofthe preorbital bar, the dorsal surface of thelacrimal is perforated by a small anteriorlydirected foramen.
The anterior process is extensive andforms most, if not all, of the dorsal border ofthe posterior antorbital fenestra. At the pos-terodorsal corner of the antorbital fenestra alarge pocket is formed by a buttress betweenthe preorbital and anterior processes of thelacrimal. The lateral surface of this buttressis extensive and flat, forming a wall to the
10 NO. 3402AMERICAN MUSEUM NOVITATES
Fig. 9. CT scans of the rostrum of IGM 100/983. Notice the osseous floor to the nasal cavity (fnc)as it extends through the interfenestral bar in C, D, E, and F. Abbreviations are listed in appendix 1.
posterodorsal part of the antorbital fossa. Asmall ridge on the anterior process overhangsthe pocket. The pocket that forms at the apexof the buttress is smooth-walled and ispierced at its apex by the lacrimal duct.
JUGAL: The jugal is extremely poorly rep-resented by only a small strap of bone fromthe anterior end, present on the right side ofIGM 100/983 (fig. 2). The preserved frag-ment of the jugal is a very thin bone that
2003 11MAKOVICKY ET AL.: BYRONOSAURUS JAFFEI
Fig. 10. Rendered view of the holotype rostrum of IGM 100/983 showing the narial chamber inrelation to the maxillary sinus and the palate. Abbreviations are listed in appendix 1.
Fig. 11. Section of the rostrum in IGM 100/984 in dorsal, right lateral, and ventral views. Abbre-viations are listed in appendix 1.
contacts the jugal process of the maxillaalong an overlapping suture where the jugalis exposed on top of and lateral to the un-derlying maxilla. The element is dorsoven-trally concave laterally. Only a short sectionof the suborbital process is preserved on theright side.
PALATINES: Only the anterior interchoanalbar of each palatine is preserved. Althoughthese flat bars are displaced and almost hor-
izontal in the holotype skull, they wouldprobably have met dorsally to form a narrowtrough above the choanae, as in Velociraptormongoliensis (Barsbold and Osmolska,1999). The anterior tips of the palatines mayhave reached the caudal end of the medianchamber beneath the nasal passage, althoughthere is a lot of breakage in the parts nearthe interefenestral bar.
VOMER: The base of the paired posterior
12 NO. 3402AMERICAN MUSEUM NOVITATES
Fig. 12. Stereopair of the palatal region of IGM 100/984 in ventral view. Abbreviations are listedin appendix 1.
Fig. 13. Stereopair of the lacrimal-nasal area of IGM 100/984 in dorsal view. Notice the largeextensions of the lacrimal over the antorbital bar. Abbreviations are listed in appendix 1.
section of the vomer is preserved betweenthe caudal ends of the maxillary palatalshelves in IGM 100/984 (fig. 11). At the lev-el of the interfenestral bar, the posterior pro-cesses conjoin to form a single median ele-ment (fig. 10B, D). In the serial CT sectionsof the holotype skull, the vomer appears toend posterior to the external naris, unlike thecondition in Velociraptor mongoliensis (IGM100/986).
PNEUMATIC SYSTEMS OF THE SNOUT
The snout comprises a complex of threelongitudinal passageways that pass from just
behind the nares back toward the choanae.On either side of the snout, the accessoryantorbital fenestra connects to the antorbitalfenestra through the lower, pneumatic pas-sage through the interfenestral bar (figs. 8,10). A blind, median chamber termed themaxillary antrum by Witmer (1997a) inter-cedes between these paired pneumatic sys-tems (fig. 9C–F). It extends from below thecaudal end of the naris to the interfenestralbar and is roofed by the nasal passage andfloored by the palate. Its lateral walls areformed by the vertical laminae of the maxillathat form the wall of each maxillary fenestra
2003 13MAKOVICKY ET AL.: BYRONOSAURUS JAFFEI
Fig. 12. Continued.
Fig. 13. Continued.
(waof in figs. 2, 3, 9B-F). The chamberopens caudally between the interfenestralbars and just anterior to the level of the cau-dal end of the nasal passage. The palatinescontribute to the ventromedial floors of thelateral passageways, and possibly to the floorof the median passage.
THE BRAINCASE
The braincase was collected in a singleweathered nodule (figs. 14, 15). Althoughseveral of the elements have been lost to ero-sion, many important features are well pre-served and allow additional insight into cra-nial anatomy and its variation in troodontiddinosaurs. The front and top of the braincaseare missing, as is the occipital condyle andthe right paroccipital process. Sutures among
braincase elements are not apparent, indicat-ing that the animal was an adult. Conse-quently, this description relates to morpho-logical features rather than specific cranial el-ements.
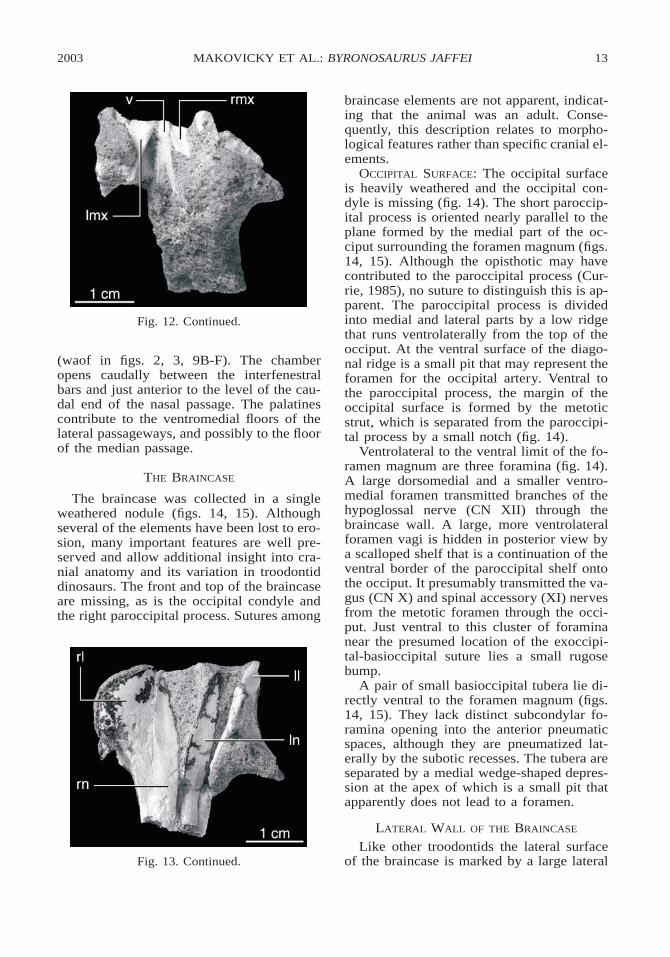
OCCIPITAL SURFACE: The occipital surfaceis heavily weathered and the occipital con-dyle is missing (fig. 14). The short paroccip-ital process is oriented nearly parallel to theplane formed by the medial part of the oc-ciput surrounding the foramen magnum (figs.14, 15). Although the opisthotic may havecontributed to the paroccipital process (Cur-rie, 1985), no suture to distinguish this is ap-parent. The paroccipital process is dividedinto medial and lateral parts by a low ridgethat runs ventrolaterally from the top of theocciput. At the ventral surface of the diago-nal ridge is a small pit that may represent theforamen for the occipital artery. Ventral tothe paroccipital process, the margin of theoccipital surface is formed by the metoticstrut, which is separated from the paroccipi-tal process by a small notch (fig. 14).
Ventrolateral to the ventral limit of the fo-ramen magnum are three foramina (fig. 14).A large dorsomedial and a smaller ventro-medial foramen transmitted branches of thehypoglossal nerve (CN XII) through thebraincase wall. A large, more ventrolateralforamen vagi is hidden in posterior view bya scalloped shelf that is a continuation of theventral border of the paroccipital shelf ontothe occiput. It presumably transmitted the va-gus (CN X) and spinal accessory (XI) nervesfrom the metotic foramen through the occi-put. Just ventral to this cluster of foraminanear the presumed location of the exoccipi-tal-basioccipital suture lies a small rugosebump.
A pair of small basioccipital tubera lie di-rectly ventral to the foramen magnum (figs.14, 15). They lack distinct subcondylar fo-ramina opening into the anterior pneumaticspaces, although they are pneumatized lat-erally by the subotic recesses. The tubera areseparated by a medial wedge-shaped depres-sion at the apex of which is a small pit thatapparently does not lead to a foramen.
LATERAL WALL OF THE BRAINCASE
Like other troodontids the lateral surfaceof the braincase is marked by a large lateral
14 NO. 3402AMERICAN MUSEUM NOVITATES
Fig. 14. Stereopairs of the braincase of the holotype specimen IGM 100/983. Top, posterior; middle,anterior; lower, right lateral views. Abbreviations are listed in appendix 1.
2003 15MAKOVICKY ET AL.: BYRONOSAURUS JAFFEI
Fig. 14. Continued.
depression bounded by an otosphenoidalcrest anteroventral to the middle ear (figs. 14,15).
LATERAL DEPRESSION AND SIDE WALL OF
THE BRAINCASE: A sharp otosphenoidal crestoriginates just ventral to the foramen for thefacial nerve (CN VII) and curves anteroven-trally, bounding the lateral depression ante-riorly and dorsally (fig. 15). The lateral de-pression is bordered posteriorly by a suboticrecess that lies ventral to the metotic strutand invades the basal tuber. Although prom-inent, the lateral depression is not as deep asin Troodon formosus (Currie and Zhao,1993) and Saurornithoides junior (Barsbold,1974).
Ventrally, the left and right lateral depres-sions appear to be separated only by a thinlamina that extends posterior to the hypo-physeal fossa. Although poor preservationmakes it difficult to determine the extent ofthis lamina, connections between the left andright lateral depression may have been pre-sent, as in Troodon formosus (Currie andZhao, 1993).
The wall of the braincase surrounding thehypophyseal fossa defines the anteromedialcorner of each lateral depression (figs. 14,15). Each internal carotid passage descendsanteroventrally through an osseous canal andmeets its counterpart immediately before en-tering the hypophyseal fossa itself in a com-mon carotid opening (fig. 14).
The middle ear is proportionately large inthis taxon. An osseous crista interfenestralissubdivides the middle ear into a smaller, tri-angular anterior aperture (the fenestra ovalis)and a larger fenestra pseudorotunda posteri-orly (figs. 14, 15). The crista interfenestralisis thinner dorsally than ventrally, and a su-ture at the base of the interfenestral crest sug-gests that it is formed entirely from the bonethat forms the dorsal rim of the middle earcavity (the opisthotic in Troodon, accordingto Currie and Zhao, 1993). Anteromedial tothe interfenestral crest, the inner ear forms adeep cochlear recess that is partly dividedinto wider proximal and more constricteddistal parts by a low ridge extending oblique-ly along the medial wall. A foramen locatedin the anterior chamber of the cochlear recessopens anteromedially into the braincase be-low the vestibular pyramid, and probably
16 NO. 3402AMERICAN MUSEUM NOVITATES
Fig. 15. The braincase of the holotype specimen IGM 100/983. Top, left lateral; middle, ventral;lower, dorsal views. Abbreviations are listed in Appendix 1.
2003 17MAKOVICKY ET AL.: BYRONOSAURUS JAFFEI
Fig. 15. Continued.
corresponds to the cochlear branch of CNVIII.
A minute foramen exits the anterior wallof the metotic foramen at mid-height. Itssmall size and proximity to the posterior partof the vestibular pyramid suggest that it isthe opening of the perilymphatic duct (Ma-kovicky and Norell, 1998). A similarly lo-cated foramen is present in ornithomimids(AMNH 5355) and Velociraptor mongolien-sis. The posteromedial part of the inner earis occupied by the metotic foramen. As inTroodon formosus, it is tall and narrow anddisplays an hourglass shape. The constrictionof the metotic foramen is located slightly be-low mid-height. Asymmetry is present be-tween the left and right ears in the region justposterolateral to the metotic foramen. On theright side, the single opening for the vagusand spinal accessory nerves and the dual fo-ramina for the hypoglossal nerve open pos-terolateral to the posterior border of the me-totic fissure (fig. 14). An additional foramenis present in this region on the left side ofthe skull (fig. 15). It opens laterally from thedorsal apex of the metotic foramen into themiddle ear recess, thus opening anterior tothe metotic strut and leading away from theoccipital region of the skull. Another smallforamen is present below the one just de-scribed, connecting the posteroventral cornerof the middle ear to the subotic recess.Again, this feature is absent on the right sideof the skull, although several small depres-sions in the roof of the subotic recess suggestthat some resorption of bone has occurred inthis region. Because it is only present on oneside, this foramen is unlikely to represent thecourse of the stapedial artery.
A pair of foramina open into the base ofthe paroccipital processes from the roof ofthe lateral depression (fig. 15) and suggestthat a posterior tympanic recess was present,albeit dimintive, unlike the condition de-scribed for Troodon formosus (Currie andZhao, 1993). Given the abbreviated length ofthe paroccipital processes, this recess musthave been relatively small in volume. A pitanterodorsal to the base of each paroccipitalprocess is here interpreted as part of the dor-sal tympanic recess. Two additional pits onthe right side suggest that the dorsal tympan-ic recess may have occupied much of the
18 NO. 3402AMERICAN MUSEUM NOVITATES
dorsolateral surface of the posterior region ofthe braincase, although its trace is smallerthan in other coleurosaurs. The left side isdamaged in the region anterodorsal to theparoccipital process, so the presence of thesepits cannot be verified.
CN VII exits the braincase laterally andslightly anteriorly through a deep, funnel-likechannel situated at the posterodorsal termi-nus of the otosphenoidal crest (figs. 14, 15).Only the posterior rim of the opening of CNV is preserved on either side. In relative di-mensions, Byronosaurus jaffei has a propor-tionately larger CN V opening than eitherTroodon formosus (Currie and Zhao, 1993)or Saurornithoides junior, although this isprobably allometric. Both of these foraminaare situated in a wide, shallow trough bor-dered by the otosphenoidal crest below anda horizontal ridge that borders a lateral ex-pansion of the braincase above. The latterstructure markedly overhangs the exit of CNV, and a small pocket opens dorsally into thisoverhang. Other surfaces of the trough arealso scalloped, a feature often associatedwith the presence of pneumatic diverticula.
The braincase is wide just dorsal to theexits of CN V (fig. 15). Dorsal to this lateralexpansion, each side of the braincase curvesmedially, and the lateral surface is concaveboth transversely and parasagittally. The dor-sal sagittal region is not preserved, but thisarea would have been roofed over by theposterior recesses of the parietals anteriorlyand supraoccipital posteriorly.
INTERIOR OF BRAINCASE AND ENDOCAST
Most of the roof and lateral walls of thebraincase are missing, but a well-consolidat-ed natural endocast of the braincase with asmooth surface is preserved (fig. 15). The re-gion of the endocast representing the cere-bellar region lies beneath the supraoccipitaland above the foramen magnum. It is prom-inently domed both sagittally and transverse-ly, and it indicates that the cerebellar regionof the brain was proportionately as large asin Troodon formosus (Currie and Zhao,1993). Beneath the cerebellar area, the floorof the braincase is deeply concave. The com-bination of this deeply concave floor and thelarge foramen magnum suggest a large pons
and medulla. Anterodorsal to the exit for CNV the laterosphenoids widen laterally to ac-commodate the tectal and optic lobes. Theanterior border of the tectal lobes is demar-cated by a deep, oblique groove, which isdorsal to the level of the hypophyseal fossa(figs. 14, 15). Rostral to this groove the rightside of the endocast preserves the base of thecerebral hemisphere, but this structure is in-sufficiently preserved to deduce relative sizecompared with other cerebral structures.
The inner surface of the posterior regionof the braincase has been prepared up to thelevel of the opening of CN VII. The vestib-ular pyramid bulges into the brain cavity op-posite the anterodorsal corner of the base ofthe paroccipital process. The apex of the ves-tibular pyramid is directed anteromedially.Anteroventral to the apex of the pyramid, theauricular fossa containing the foramina forCN VII and the cochlear and vestibularbranches of CN VIII extends laterally fromthe base of the cerebellar portion of the en-docast. On both sides of the braincase, thebony wall of the braincase is shattered in theregion of the floccular recess. Damage to thesurface of both pyramids precludes identifi-cation of the opening of the endolymphaticduct. The vertical vestibular canal is not ex-cessively swollen. The metotic foramen issituated posteroventral to the vestibular pyr-amid, while the exit of CN VII constrains thebase of the vestibular pyramid anteriorly.The exit of CN VII is visible as a larger fo-ramen located anterior to the vestibular pyr-amid, at approximately the same level as theopening for CN VIII.
SCLERAL OSSICLES: Articulated anteriorsegments of both scleral rings are preservedin the anterior part of the orbit on IGM 100/983, two ossicles on the right and three onthe left (figs. 2, 3). Some of the remainingossicles may lie in a mass of fragmentarybones just ventral to the orbit, but most ofthese fragments seem to be from the jugaland mandible. The five ossicles are incom-pletely exposed, but they are generally sim-ilar to those of troodontids (e.g., Saurorni-thoides mongoliensis) and other basal coe-lurosaurs in being rectangular with only agentle lateral convexity.
2003 19MAKOVICKY ET AL.: BYRONOSAURUS JAFFEI
THE MANDIBLE
DENTARY: The dentaries are partly eroded,but from the preserved sections it can be de-termined that they are thin with slightly con-cave dorsal and convex ventral margins (figs.2, 3). Like other troodontids they are tall pos-teriorly and taper anteriorly toward the sym-physis. The lateral surface is scoured by adeep median groove filled with numeroussmall foramina, just below the tooth row.Posteriorly, this groove disappears as the lat-eral surface of the dentary becomes flat; asmall ridge overhangs the lateral surface dor-sally. CT scans show that the groove is levellying with the implanted bases of the toothroots. Small perforating canals that pass be-tween the roots of consecutive teeth connectthe lateral canal and the Meckelian groove,transmitting branches of the mandibularnerve and artery. CT scans also show thatthe Meckelian groove parallels the lateralgroove along the inner surface of the man-dible. The groove terminates dorsal to a fo-ramen
Similar to Sinornithoides youngi (Ruselland Dong, 1993) the dentaries are parallel inventral view, and there is no medial curva-ture of the rami at the mandibular symphysis,as there is in Saurornithoides mongoliensis,Saurornithoides junior, and Troodon formo-sus (fig. 3).
SPLENIAL: Only fragments of the splenialare exposed, in ventral view. The anterior ex-tent of the splenial is hard to determine; how-ever, it reaches at least as far anteriorly asbeneath the accessory antorbital fenestra (fig.3). It is apparently longer than in Deinony-chus antirrhopus and more similar to thecondition in the dromaeosaurids Velociraptormongoliensis and Dromaeosaurus alberten-sis where the splenial is at least half thelength of the dentary. Posteriorly, the splen-ial wraps around the ventral surface of thedentary to become exposed laterally, beneaththe anterior end of the antorbital fenestra.Whether the splenial is significantly exposedlaterally as a triangular surface as in Sauror-nithoides junior, Saurornithoides mongolien-sis (personal obs.), and dromaeosaurids (Cur-rie, 1995) cannot be determined.
SURANGULAR: The surangular is only pre-served as pieces on the right side of the skull,
and little morphology can be ascertained (fig.2). It is transversely convex and apparentlynarrow dorsal to the mandibular fenestra,suggesting that the mandibular fenestra waslarge, unlike the condition in dromaeosaur-ids. Just ventral to the lacrimal buttress, alarge, elliptical surangular foramen perfo-rates the surangular; anterior to this foramenthe bone is slightly concave, but lacks a dis-tinct groove as in some other theropods (e.g.,Ornitholestes hermanni and Dromaeosaurusalbertensis).
ANGULAR: The right angular is also ex-tremely fragmentary (fig. 2). The element iscrescentic, with an expanded lateral surfaceanteriorly. More anteriorly the bone is frag-mented.
DENTITION
The dentition of Byronosaurus jaffei is un-usual for theropod dinosaurs in that no ser-rations are found on the anterior or posteriorcarinae (fig. 16), a feature known only in al-varezsaurids, the ornithomimosaur Pelecani-mimus polyodon, and toothed avialans. Fourpremaxillary teeth are present, but it is dif-ficult to determine the number of maxillaryor dentary teeth because these areas are poor-ly preserved. Nevertheless, the preservedteeth are densely packed, and a conservativeestimate using the spacing of the preservedteeth to estimate the number of missing teethis that at least 30 maxillary teeth and 30 den-tary teeth were present.
All of the teeth are recurved with sym-metrical carinae and somewhat bulbouscrown bases (fig. 16). The tooth bases aremarked by mediolateral, longitudinalgrooves. The premaxillary teeth are smalland closely packed, and they are round incross section. The maxillary teeth are smallcompared with similar-sized theropods. Theyare also heterodont, with larger teeth occu-pying the 11th and 12th tooth positions. Themore anterior teeth are round in cross sec-tion, grading into elliptical and then subrect-angular teeth posteriorly.
POSTCRANIAL SKELETON
Limited postcranial remains were pre-served with the holotype, including severalvertebrae from the presacral part of the ver-
20 NO. 3402AMERICAN MUSEUM NOVITATES
Fig. 16. The teeth of the right maxilla and dentary of the holotype (IGM 100/983). Notice both thelack of serrations and the heterodont condition.
tebral column and tail. Hindlimb elements in-clude a heavily weathered element that rep-resents the proximal end of the right femur,the right knee, a number of broken andweathered sections of hollow limb bones,and a number of pedal phalanges. None ofthe preserved vertebrae are complete.
The preserved cervical elements are iden-tified as parts of the axis and third and fourthcervicals. Two nodules each contain parts oftwo articulated anterior cervicals. Only theposterior half of the neural arch and the pos-teriormost end of the centrum are preservedon the more anterior cervical. The postzy-gapophyses are set far from the midline andare surmounted by small epipophyses. Thebase of the neural spine is preserved and itappears to have been anteroposteriorly shortalong the midline. The following cervical isbetter preserved and comprises the majorityof the centrum and the anterior half of theneural arch (fig. 17). The prezygapophysesdiverge widely from the midline and face an-terodorsally and slightly medially. Small, an-terior pedicular fossae are present just lateralto the floor of the neural canal. Laterally, the
borders of the neural arch arc ventrally andoverhang the centrum. The broken posteriorend of the centrum reveals that the interiorwas hollow, and a small foramen on the sideof the centrum probably marks the pneumaticconnection to the interior. Nearly completecervical ribs are preserved in articulationwith both the centrum and arch of the secondcervical in the nodule. The capitulum and tu-berculum are axially elongate. A short, an-terior process extends a short distance ante-rior to the juncture between the tuberculumand capitulum, and a small pneumatic fora-men is visible on the medial face of the leftrib in this region. Posterior to this juncturethe main shaft of the cervical rib has a long,tapering, transversely flattened shaft.
A second, smaller nodule contains the an-terior portion of the anterior vertebra fromthe nodule described above as well as theposterior part of the preceding element. This,the anteriormost preserved cervical, has asmall but proportionately massive neuralarch. The arch narrows rostrally, suggestingthat it was triangular in dorsal view, as ischaracteristic of the theropod axis. The single
2003 21MAKOVICKY ET AL.: BYRONOSAURUS JAFFEI
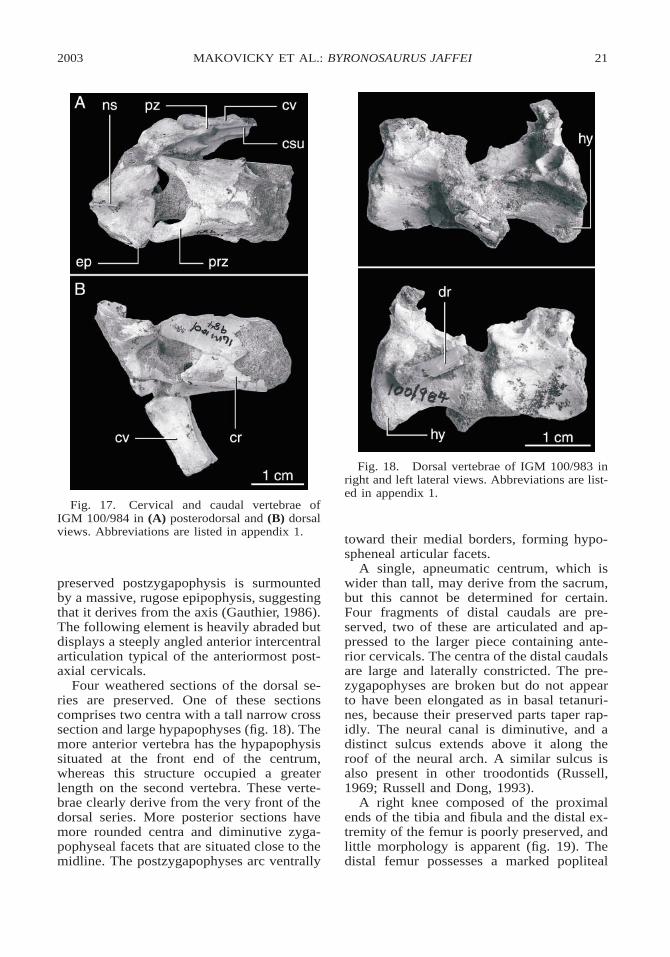
Fig. 17. Cervical and caudal vertebrae ofIGM 100/984 in (A) posterodorsal and (B) dorsalviews. Abbreviations are listed in appendix 1.
Fig. 18. Dorsal vertebrae of IGM 100/983 inright and left lateral views. Abbreviations are list-ed in appendix 1.
preserved postzygapophysis is surmountedby a massive, rugose epipophysis, suggestingthat it derives from the axis (Gauthier, 1986).The following element is heavily abraded butdisplays a steeply angled anterior intercentralarticulation typical of the anteriormost post-axial cervicals.
Four weathered sections of the dorsal se-ries are preserved. One of these sectionscomprises two centra with a tall narrow crosssection and large hypapophyses (fig. 18). Themore anterior vertebra has the hypapophysissituated at the front end of the centrum,whereas this structure occupied a greaterlength on the second vertebra. These verte-brae clearly derive from the very front of thedorsal series. More posterior sections havemore rounded centra and diminutive zyga-pophyseal facets that are situated close to themidline. The postzygapophyses arc ventrally
toward their medial borders, forming hypo-spheneal articular facets.
A single, apneumatic centrum, which iswider than tall, may derive from the sacrum,but this cannot be determined for certain.Four fragments of distal caudals are pre-served, two of these are articulated and ap-pressed to the larger piece containing ante-rior cervicals. The centra of the distal caudalsare large and laterally constricted. The pre-zygapophyses are broken but do not appearto have been elongated as in basal tetanuri-nes, because their preserved parts taper rap-idly. The neural canal is diminutive, and adistinct sulcus extends above it along theroof of the neural arch. A similar sulcus isalso present in other troodontids (Russell,1969; Russell and Dong, 1993).
A right knee composed of the proximalends of the tibia and fibula and the distal ex-tremity of the femur is poorly preserved, andlittle morphology is apparent (fig. 19). Thedistal femur possesses a marked popliteal
22 NO. 3402AMERICAN MUSEUM NOVITATES
Fig. 19. Right knee of the holotype specimenIGM 100/983 in (A) medial and (B) lateral views.Abbreviations are listed in appendix 1.
Fig. 20. Pedal phalanges of the holotype spec-imen IGM 100/983. Abbreviations are listed inappendix 1.
fossa. The distal condyles are narrow and ex-tend far posterior to the femoral shaft. Along, narrow supracondylar crest extendsfrom the lateral condyle toward the proximalend of the femur.
The proximal end of the tibia is subtrian-gular in proximal view. Both the cnemialcrest and the lateral condyle of the tibia arewell developed and project far from theshaft. The cnemial crest is subtrapezoidal inmedial view and curves laterally at its ante-rior rim. A large, suboval tuberosity marksthe proximodorsal extremity of the cnemialcrest, and the anterior border below it isthickened for muscular insertion. No detailsare evident on the short preserved section ofthe fibula. A number of hollow limb bonefragments pertain to the hindlimbs, but theydo not warrant description.
The distal end of right metatarsal II is the
only part of the metatarsus preserved. Thearticulation is narrow and simple, with amore pronounced medial than lateral collat-eral ligament fossa. A pronounced, narrowridge forms the posteromedial edge of the ar-ticulation but is broken a short distanceabove its distal terminus.
Fragments of three pedal phalanges werefound with the holotype of Byronosaurus jaf-fei (fig. 20). One of these can be identifiedas the second phalanx of the second digit be-cause of the presence of an elongated heelproximoventrally. As in other troodontids,but in contrast to derived dromaeosauridslike Velociraptor mongoliensis and Deinon-ychus antirrhopus, the pinched shaft betweenthe articular ends is not foreshortened (Currieand Peng, 1993). The medial face is vertical,whereas the lateral one is convex, a featurethat allows determination that the phalanxderives from the right foot. Erosion has de-stroyed much of the distal ginglymus. In pro-file, the phalanx resembles the corresponding
2003 23MAKOVICKY ET AL.: BYRONOSAURUS JAFFEI
bone in Troodon formosus (Currie and Peng,1993) and an indeterminate troodontid fromMongolia (Barsbold et al., 1987).
The other phalangeal fragments consist ofarticulated distal and proximal ends of twophalanges. Dorsodistally, the articulation ofthe proximal element bears a lappetlike ex-tension of the articular surface. The articu-lated proximal end of one phalanx is trian-gular in cross section and the rest of the el-ement appears to have been relatively slen-der. By comparison with Borogoviagracilicrus (Osmolska, 1987), these frag-ments may represent the phalanges III-2 andIII-3.
DISCUSSION
Byronosaurus jaffei is one of the best pre-served troodontid dinosaurs yet discoveredand extends our knowledge on the diversityof Late Cretaceous troodontids. A number ofimportant characters such as an extensivesecondary palate, the opening here interpret-ed as a subnarial foramen, and the connec-tion between the antorbital and accessory fe-nestrae though the interfenestral bar werepreviously unrecognized in troodontids. Un-usually among troodontids, the teeth are un-serrated. Prior to the discovery of Byrono-saurus jaffei and Sinovenator changii (Xu etal., 2002), most known troodontid teeth werecharacterized by large, apically curved den-ticles with ‘‘blood pits’’ between them (Cur-rie et al., 1990). The only exception is anunnamed troodontid from Mongolia, whichapparently has reduced denticles that extendperpendicular to the axis of the tooth (Bars-bold et al., 1987).
Byronosaurus jaffei provides us with thefirst data on the anatomy of the anterior partof the troodontid palate. The palate is formedanteriorly by the premaxilla and more pos-teriorly by the palatal shelves, which are flatanteriorly and become more vaulted posteri-orly (figs. 4, 12). Premaxillary contributionto the roof of the palate is limited comparedto other coelurosaurs. The vomers appearfused anteriorly and are divided posteriorlyat the level of the interfenestral bar (fig. 12).The vomers apparently lay between the pal-atal shelves posteriorly and lie dorsal to themmore anteriorly. They extend anterior to the
interfenestral bar; however, they apparentlydid not extend along the roof of the palateanteriorly beneath the nares as in Velocirap-tor mongoliensis.
An unusual feature of Byronosaurus jaffei,which is also present in the alvarezsauridShuvuuia deserti, is the presence of a smalltubular foramen on the maxilla just dorsal tothe tooth row on the ventral border of thenarial fossa (fig. 6). We interpret this featureas homologous to the subnarial foramen,which typically lies in the premaxillary-max-illary suture and level with the supraalveolarcanal in theropods and presumably transmit-ted parts of the maxillary branch of CN Vthat innervated the tip of the snout. Thisopening extends into a tubular sinus adjacentto the tooth row (the supra-alveolar canal[Norell et al., 2000]). A very similar config-uration is seen in Shuvuuia deserti (IGM100/977). If our interpretation of this featureis correct, then Sereno’s (2001) assertion thatalvarezsaurids lack a subnarial foramenneeds to be modified.
The internarial bar in most theropod di-nosaurs is an oval or cylindrical rodlikestructure composed of the premaxillae thatmeet the nasals posteriorly. Sereno (2001:93) asserted, in reference to his placement ofalvarezsaurids as the sister group to orni-thomimosaurs, that ‘‘the unusual dorsoven-tral flattening of the internarial bar . . . isunique to these two groups.’’ However, thisconfiguration is also present in the troodontidtaxa Saurornithoides junior and Byronosau-rus jaffei.
The interfenestral bar in Byronosaurus jaf-fei shows some unique features (figs. 7–10).Unlike other troodontids, the interfenestralbar is not inset from the plane of the maxilla,but is flush with the lateral surface of thesnout. As in both species of Saurornithoides,a connection between the antorbital and ac-cessory antorbital fenestrae passes throughthe interfenestral bar (figs. 9, 10). The nasalpassage terminates at the dorsal end of theinterfenestral bars in Byronosaurus jaffei,and it opens into the choanae at the level ofthe antorbital fenestra (fig. 9). Our previousdescription (Norell et al., 2000) stated un-clearly that the nasal passages are connectedto the antorbital fenestra by a connectionthrough the interfenestral bar, but an exposed
24 NO. 3402AMERICAN MUSEUM NOVITATES
cross section of this region in the referredspecimen clearly shows that it is the nasalpassage proper that transits the interfenestralbar.
Although Byronosaurus jaffei shares de-rived dental characters related to tooth spac-ing and shape with other troodontids, it dif-fers in having unserrated teeth (fig. 16). Thishas important implications toward under-standing the phylogenetic position of troo-dontids. Some previous analyses havegrouped the Troodontidae with therizinosau-roids (and the edentulous oviraptorosaurs) inpart because of the enlarged and apicallyhooked serrations seen in members of bothgroups. The presence of small serrations inthe recently described basal troodontid Si-novenator (Xu et al., 2002) and the absenceof serrations in Byronosaurus jaffei clearlyshow that large denticles can no longer beregarded as plesiomorphic within Troodon-tidae, and are diagnostic of a more exclusiveclade within Troodontidae.
PHYLOGENY
In our preliminary description of Byrono-saurus jaffei (Norell et al., 2000), we exam-ined the relationship of Byronosaurus jaffeito other troodontids and found it to be thesister group to a clade of derived large-bod-ied troodontids comprising Troodon formo-sus and both species of Saurornithoides.More recent studies of coelurosaurian rela-tionships (Norell et al., 2001; Xu et al., 2002)have included almost the same sampling oftroodontid taxa as our preliminary study andhave yielded results that are largely congru-ent with it.
Here we have expanded the Xu et al.(2002) study, which included the data of No-rell et al. (2000), by adding an unnamed tax-on from the Early Cretaceous of Mongolia(Barsbold, 1987) and several characters,some of which pertain to troodontid relation-ships. Seven troodontid species are includedalong with 42 other coelurosaurian taxa andthe tetanuran Allosaurus, which was used toroot the most parsimonious trees. The com-plete matrix has 220 characters and 50 taxa,and the character list and data matrix areavailable at http://research.amnh.org/users/norell/index.html. The troodontids Borogo-
via gracilicrus and Tochisaurus nemegtensiswere excluded, because apart from the auta-pomorphic phalangeal characters of the for-mer, the single known specimen of each spe-cies only shows characters that are commonto all troodontids more derived than Sinov-enator, and thus provides no information to-ward elucidating troodontid relationships.Our comprehensive approach of analyzingtroodontid relationships in the broader con-text of coelurosaurian phylogeny is justifiedby the lack of consensus regarding the rela-tionship of Troodontidae to outgroup taxa.Furthermore, the high amounts of homoplasyand missing data that pervade coelurosaurianrelationships complicate the correct estab-lishment of character polarities. In our view,the least assumptive approach to this prob-lem is to analyze a comprehensive range ofoutgroup taxa (i.e., Coelurosauria) to achievean accurate optimization of ancestral states atthe root node of Troodontidae.
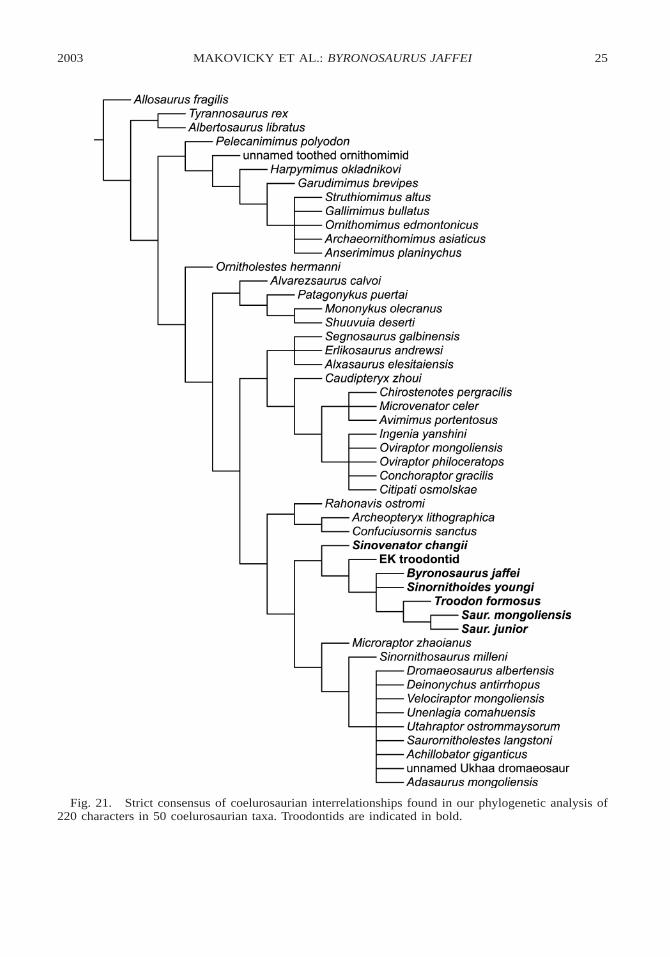
All characters were treated as unordered.Analysis of this dataset was conducted usingNONA version 1.9 (Goloboff, 1999) runthrough the Winclada interface (Nixon,1999). One thousand repetitions of the treebisection regrafting algorithm were used tofind islands of shortest trees, followed bybranch swapping to find all shortest trees.The analysis found 432 trees (TL 5 586, CI5 0.45, RI 5 0.75), and a strict consensustree is presented in fig. 21. Troodontid taxaare boldfaced.
Troodontids are found to be monophyleticand are the sister taxon of the Dromaeosaur-idae within a monophyletic Deinonychosau-ria, which in turn is the sister taxon to Avi-alae. Troodontid monophyly is wellsupportedby the nine characters discussed below.
Dentary nutrient foramina situated in ahorizontal groove on labial face of den-tary: The labial margin of the dentary ispierced by a disparate number of nutrient andnervous foramina in theropods. In Byrono-saurus jaffei (figs. 2, 3), Troodon formosus(Russell, 1969), Saurornithoides mongolien-sis, Saurornithoides junior (Barsbold, 1974),and Sinovenator changii (Xu et al., 2002)these foramina line the floor of a distinct sul-cus on the labial side of the dentary (Currie,1987). In Sinovenator changii this groove isrestricted to the posterior part of the dentary
2003 25MAKOVICKY ET AL.: BYRONOSAURUS JAFFEI
Fig. 21. Strict consensus of coelurosaurian interrelationships found in our phylogenetic analysis of220 characters in 50 coelurosaurian taxa. Troodontids are indicated in bold.
26 NO. 3402AMERICAN MUSEUM NOVITATES
ventral to the caudal end of the tooth row,but in more derived taxa it extends furthertoward, but does not reach, the symphysis. Indromaeosaurids, oviraptorids, and other man-iraptorans the neurovascular foramina arelinearly arranged but do not lie in a groove.
Pneumatic quadrate with pneumato-pore on posterior face: The quadrate bearsa pneumatic foramen posteriorly in the troo-dontid taxa where it is known, includingTroodon formosus (Varrichio, 1997), an un-named Early Cretaceous troodontid (Bars-bold et al., 1987), and Sinovenator changii(Xu et al., 2002). Quadrate pneumaticity isvariably expressed among coelurosauriantheropods. It is observed in tyrannosaurids,advanced oviraptorids (Maryanska and Os-molska, 1997), the compsognathid Sinosau-ropteryx prima (Currie and Dong, 2001b),derived avialans, and possibly some, but notall, ornithomimosaurs (Makovicky and No-rell, 1998). Quadrate pneumaticity is absentin many maniraptorans, including basal ovi-raptorosaurs such as Avimimus, alvarezsaur-ids, therizinosauroids (Clark et al., 1994),and Ornitholestes hermanni. It also appearsto be absent in Archaeopteryx lithographica(Witmer, 1990; Elzanowski and Wellnhofer,1996) and Confuciusornis sanctus (Chiappeet al., 1999).
Basisphenoid recess absent: A blindpocket, derived from the median pharyngealsystem, invades the basisphenoid ventrally inmost theropods and forms a deep pneumaticrecess on the basicranium (Witmer, 1997).This basisphenoid recess is widespread incoelurosaurs and is seen in ornithomimids(Osmolska et al., 1972), oviraptorids (Clarket al., 2002), and dromaeosaurids (e.g., Dro-maeosaurus albertensis AMNH 5356). Thisrecess is secondarily lost in troodontids (Cur-rie, 1985), although the basisphenoid is hol-low and pneumatic in at least Troodon for-mosus, Byronosaurus jaffei (fig. 15), andSaurornithoides junior. Secondary loss ofthe basisphenoid recess is also observed inthe therizinosauroid Erlikosaurus andrewsi(Clark et al., 1994) and in advanced aviali-ans. The condition is unknown for Archae-opteryx lithographica and Confuciusornissanctus.
Dorsoventrally flattened internarialbar: As discussed above, the internarial bar
of troodontids appears to be dorsoventrallycompressed (figs. 2, 5). Each premaxillaryinternarial process is wider than tall and hasa dorsally flat surface, and the short nasalcontribution to the narial bar forms a lateralextension of the base of the bar. This con-trasts with the round internarial bar of dro-maeosaurids such as Velociraptor mongo-liensis (AMNH 6215) and uncrested ovirap-torosaurs. A flattened internarial bar is de-rived independently in ornithomimosaurs andalvarezsaurids (Sereno, 1999). According toour analysis, this feature does not unambig-uously optimize as a synapomorphy of anycombination of these clades.
Closely packed anterior dentition insymphyseal region of dentary: The lowerdentition of troodontids is moderately heter-odont. In the dentary, the anteriormost teethare closely packed, and toothless jaws showthat the alveoli are confluent within a groovein the anterior part of the jaw (Currie, 1987).Posteriorly, the alveoli are separated by wallsof porous interdental bone. In contrast, theteeth of most theropods are more equidis-tantly separated and set in separate alveolithroughout the whole jaw, as in dromaeo-saurids (IGM 100/986, AMNH 5356) andArchaeopteryx lithographica (Elzanowskiand Wellnhofer, 1996). The teeth of alvar-ezsaurids and the basal ornithomimosaur Pe-lecanimimus are set in a groove throughoutthe dentary, but are subequal in size. In ther-izinosauroids, all teeth are set in individualalveoli separated by septa, and the teethgrade toward larger size rostrally (Clark etal., 1994; Xu et al., 2001).
Depression on the ventral surface of thepostorbital process of the laterosphenoid:A ventral pit is visible on the postorbital pro-cess of the laterosphenoid in Troodon for-mosus (Currie, 1985), Saurornithoides junior(IGM 100/1), and Sinovenator changii (Xuet al., 2002). The pit has smooth walls, andmay have been pneumatic. The postorbitalprocess of dromaeosaurids (AMNH 5356,PIN 100/25), Archaeopteryx (Walker, 1985;London specimen), oviraptorosaurs, therizi-nosauroids (Clark et al., 1994), alvarezsaur-ids, and ornithomimosaurs does not bearsuch a pit.
Reduced basal tubera that lie directlyventral to the occipital condyle: The basal
2003 27MAKOVICKY ET AL.: BYRONOSAURUS JAFFEI
tubera are unusual among coelurosaurs in be-ing reduced and situated close to the midline,so that they are directly ventral to the occip-ital condyle in Asian troodontid taxa (fig.14). The tubera are separated by a narrow V-shaped notch in all troodontids. The tuberaare larger in Troodon formosus, but they arestill positioned close to the midline (Russell,1969) and ventral to the condyle. In manyoutgroup taxa, including dromaeosaurids,oviraptorosaurs, and ornithomimosaurs, thetubera are more pronounced and lie in a para-sagittal plane lateral to the occipital condyle.Alvarezsaurids have reduced tubera, butthese are widely separated from each otherby a notch that is wider than the diameter ofthe occipital condyle. The basal tubera of thetherizinosauroid Erlikosaurus andrewsi arevirtually obliterated by the ventral expansionof the pneumatic basisphenoid (Clark et al.,1994).
Large number of teeth: Among advancedtheropods, troodontids are characterized byhaving a very large number of teeth (fig. 2).Although the precise number is unknown inByronosaurus jaffei, a minimum of 4 pre-maxillary and 30 maxillary teeth were pre-sent in the upper jaws, while the dentary held30 or more teeth. In dromaeosaurids thetooth count is substantially smaller— 4 pre-maxillary, and 9 (Dromaeosaurus alberten-sis) to15 (Deinonychus antirrhopus) maxil-lary teeth and 11 (Dromaeosaurus alberten-sis) to 16 (Deinonychus antirrhopus) dentaryteeth (Ostrom, 1969, Currie, 1995). Archae-opteryx lithographica has a dental formula of4 premaxillary plus 8 maxillary and 11 den-tary teeth (Wellnhoffer, 1974). Oviraptoro-saurs and advanced ornithomimids are eden-tulous, but the basal ornithomimid Pelecan-imimus polyodon (Perez-Moreno et al., 1994)has more than 220 teeth in its jaws, and alarge number of teeth are also present in thealvarezsaurid Shuuvuia and in therizinosau-roids. According to our analysis, these taxaderive the large number of teeth independentof troodontids.
Long, slender transverse processes: Thetransverse processes of the trunk vertebraeare relatively long and slender and curveslightly in troodontid taxa that preservethem, including Sinoventor changii andTroodon formosus. The transverse processes
are relatively short in dromaeosaurids suchas Sinornithosaurus millenii (Xu et al., 1999)and Velociraptor mongoliensis (IGM 100/986) and in the basal avialans Archaeopteryxlithographica (Berlin specimen) and Rahon-avis ostromi (Forster et al., 1998). Ovirap-torosaurs have short, wide transverse pro-cesses on the trunk vertebrae (Makovickyand Sues, 1998). Proportionately long trans-verse processes are seen in ornithomimidsand in alvarezsaurids.
In all troodontids for which the tail isknown, the distal caudals have a sulcus onthe dorsal midline in place of a neural spine,but the condition is unknown for the mostbasal troodontid Sinovenator and is thus anambiguous synapomorphy of the clade. Sev-eral characters used to diagnose Troodonti-dae in the past no longer optimize at the baseof Troodontidae, following the discovery ofa number of new deinonychosaurs in recentyears. The troodontid metatarsus is asym-metrical with a slender metatarsal II that ismarkedly shorter than metatarsals III and IV,and a robust metatarsal IV, unlike the moresymmetrical metapodium of most other the-ropods, and this asymmetry has been used todiagnose the clade in the past (Wilson andCurrie, 1985). Somewhat asymmetric meta-podia are also observed in the basal dro-maeosaurids Microraptor zhaoianus (Xu etal., 2000) and, to a lesser degree, in Sinor-nithosaurus millenii (Xu et al., 1999), andthis character may be diagnostic of Deinon-ychosauria rather than of Troodontidae.
Possession of large, apically hooked den-ticles has also been considered a diagnosticcharacter of troodontids (Barsbold, 1974;Currie, 1987). The recently discovered Si-novenator changii has small denticles direct-ed perpendicular to the long axis of the tooth,however, and this character now diagnoses amore exclusive clade within Troodontidae.An inflated bulla at the base of the cultriformprocess has been observed in Saurornithoi-des junior and Troodon formosus, and it waspreviously viewed as a troodontid synapo-morphy. It is, however, absent in the basaltroodontid Sinovenator changii and its pres-ence cannot be ascertained in several othertaxa including Byronosaurus jaffei.
Within troodontids, Sinovenator changii isthe sister taxon to a clade comprising all oth-
28 NO. 3402AMERICAN MUSEUM NOVITATES
er troodontids. Two unambiguous synapo-morphies are present at this node.
Presence of a subotic recess on the sideof the braincase ventral to the middle ear:A pneumatic pocket invades the lateral wallof the basicranium ventral to the middle earin Troodon formosus, Saurornithoides ju-nior, Byronosaurus jaffei (figs. 14, 15), andthe Early Cretaceous troodontid from Kha-mareen Us (Barsbold et al., 1987). Witmer(1997b) termed this feature the subotic re-cess. A subotic recess is absent in dromaeo-saurids, basal avialans, alvarezsaurids, andoviraptorosaurs. It is convergently present inat least some derived ornithomimosauriantaxa (Makovicky and Norell, 1998).
Metatarsal IV oval in cross section,deeper than wide: Basal paravians have amediolaterally expanded metatarsal IV witha lateral flange on the side. This condition isseen in Archaeopteryx lithographica and Ra-honavis ostromi and in dromaeosaurids, in-cluding Microraptor zhaoianus and Veloci-raptor mongoliensis. In higher troodontids,the fourth metatarsal reverses to a more co-lumnar shape with an oval cross section thatis longer along the palmar-plantar axis.
Byronosaurus jaffei and Sinornithoidesyoungi form a clade together with the largerbodied, Late Cretaceous Troodon formosus,Saurornithoides mongoliensis, and Sauror-nithoides junior. Two cranial characters un-ambiguously diagnose this group.
Large otosphenoidal crest defining lat-eral depression on side of braincase: Troo-dontids, with the exceptions of an unnamedEarly Cretaceous form (Barsbold et al.,1987) and Sinovenator changii, bear a well-developed ‘‘lateral depression’’ (Barsbold,1974) on the side of the braincase coveringparts of the laterosphenoid, basisphenoid,prootic, and possibly part of the opisthotic.This lateral depression is delimited anteriorlyand dorsally by the expanded otosphenoidalcrest, which is formed by the laterosphenoidanteriorly and by the prootic and possiblyalso the opisthotic posteriorly. A number ofsmall laminae subdivide the lateral depres-sion into subcompartments, such as the pro-otic recess rostrally. The internal carotids en-ter the hypophyseal fossa near the rostral partof the lateral depression. The middle ear lieswithin the lateral depression in Saurorni-
thoides junior, but is dorsal to it in Byrono-saurus jaffei (figs. 14, 15).
Presence of enlarged, distally orienteddenticles: Theropod teeth are primitively ser-rated. In velociraptorine dromaeosauridsthere are 5–7 denticles per millimeter (Currieet al., 1990), while this ratio is 3–3.2 denti-cles per millimeter in Dromaeosaurus alber-tensis (Currie, 1996). By contrast the numberof denticles is 2.3–3 in Saurornithoides mon-goliensis (Russell, 1969) and less than 2 permm in Troodon formosus (Currie et al.,1990) and Saurornithoides junior (Barsbold,1974). In addition, the denticles of thesetroodontid taxa are long and separated bydeep blood grooves with small proximalblood pits. The teeth of Sinornithoides youn-gi have porportionately smaller serrationsthan do those of the three larger taxa listedabove, but they are still proportionately largerelative to tooth size (Currie and Dong,2001a).
Relationships between Byronosaurus jaf-fei, Sinornithoides youngi, and a clade com-prising the three larger bodied taxa Troodonformosus and both species of Saurornithoi-des cannot be parsed out, partly because thebraincase is undescribed for Sinornithoidesand the dentition is autapomorphic in Byron-osaurus jaffei. The Troodon 1 Saurornithoi-des clade is derived in its possession of amedially deflected symphysis on the dentaryand by the secondary loss of a dorsal tym-panic recess.
ACKNOWLEDGMENTS
We thank the field crews of the 1993,1994, and 1996 field seasons. For help inMongolia, we would especially thank D.Baatar, T. Boldsukh, D. Dashzeveg, and R.Barsbold. Mike Novacek found the typespecimen (IGM 100/983). Martin Kundratcarefully went over the manuscript. Amy Da-vidson and Bill Amaral prepared the speci-mens and Mick Ellison developed the fig-ures. This work was supported by the Divi-sion of Paleontology at the American Mu-seum, the Field Museum Department ofGeology, and the Jaffe Family Foundation.
REFERENCESBarsbold, R. 1974. Saurornithoididae, a new fam-
ily of small theropod dinosaurs from Central
2003 29MAKOVICKY ET AL.: BYRONOSAURUS JAFFEI
Asia and North America. Palaeontologica Po-lonica 30: 5–22.
Barsbold, R. 1983. Carnivorous dinosaurs fromthe Cretaceous of Mongolia. SovmestnayaSovetskogoMongol’skaya Paleontologiches-kaya Ekspeditsya. Trudy 15: 28–39. [in Rus-sian]
Barsbold, R., and H. Osmolska. 1999. The skullof Velociraptor (Theropoda) from the Late Cre-taceous of Mongolia. Acta Palaeontologica Po-lononica 44(2): 189–219.
Barsbold, R., H. Osmolska, and S. Kurzanov.1987. On a new troodontid (Dinosauria, Ther-opoda) from the early Cretaceous of Mongolia.Acta Palaeontologica Polonica 32(1,2): 121–132, pls. 49–52.
Chiappe, L.M., J. Shu’an, J. Qiang, and M.A. No-rell. 1999. Anatomy and systematics of theConfuciusornithidae (Theropoda: Aves) fromthe late Mesozoic of northeastern China. Bul-letin of the American Museum of Natural His-tory 242: 1–89.
Clark, J.M., M. Norell, and T. Rowe. 2002. Cra-nial anatomy of Citipati osmolskae (Theropoda,Oviraptorosauria), and a reinterpretation of theOviraptor philoceratops holotype. AmericanMuseum Novitates 3364: 1–24.
Clark, J., A. Perle, and M.A. Norell. 1994. Theskull of Erlikosaurus [sic] andrewsi, a LateCretaceous ‘‘segnosaur’’ (Theropoda: Therizi-nosauridae) from Mongolia. American MuseumNovitates 3115: 1–39 pp.
Currie, P.J. 1985. Cranial anatomy of Stenonycho-saurus inequalis (Saurischia, Theropoda) andits bearing on the origin of birds. CanadianJournal of Earth Sciences 22: 1643–1658.
Currie, P.J. 1987. Bird-like characteristics of thejaws and teeth of troodontid theropods (Dino-sauria, Saurischia). Journal of Vertebrate Pale-ontology 7: 72–81.
Currie, P.J. 1995. New information on the anato-my and relationships of Dromaeosaurus alber-tensis (Dinosauria: Theropoda). Journal of Ver-tebrate Paleontology 15(3): 576–591.
Currie, P.J., and Z.-M. Dong. 2001a. New infor-mation on Cretaceous troodontids (Dinosauria:Theropoda) from the People’s Republic of Chi-na. Canadian Journal of Earth Sciences 38(12):1753–1766.
Currie P.J., and Z.-M. Dong. 2001b. Anatomy ofSinosauropteryx prima from Liaoning, north-eastern China. Canadian Journal of Earth Sci-ences 38(12): 1705–1727.
Currie, P.J., and J.-H. Peng. 1993. A juvenilespecimen of Saurornithoides mongoliensisfrom the Upper Cretaceous of northern China.Canadian Journal of Earth Sciences 30: 2224–2230.
Currie, P.J., and X.-J. Zhao. 1993. A new troo-dontid (Dinosauria, Theropoda) braincase fromthe Dinosaur Park Formation (Campanian) ofAlberta. Canadian Journal of Earth Sciences30: 2231–2247
Currie, P.J., J.K. Rigby, and R.E. Sloan. 1990.Theropod teeth from the Judith River Forma-tion of southern Alberta. In K. Carpenter andP.J. Currie (editors), Dinosaur systematics—ap-proaches and perspectives: 107–125. Cam-bridge, UK: Cambridge University Press.
Elzanowski, A., and P. Wellnhofer. 1996. Cranialmorphology of Archaeopteryx: evidence formthe seventh skeleton. Journal of Vertebrate Pa-leontology 16: 81–94.
Forster, C.A., S. Sampson, L. Chiappe, and D.Krause. 1998a. The theropod ancestry of birds:new evidence from the Late Cretaceous ofMadagascar. Science 279: 1915–1919.
Gauthier, J.A. 1986. Saurischian monophyly andthe origin of birds. In K. Padian (editor), Theorigin of birds and the evolution of flight.Memoirs of the California Academy of Scienc-es 8: 1–55.
Goloboff, P. 1999. NONA (ver. 1.9). Softwarepublished by the author, S.M. de Tucuman, Ar-gentina. Available on-line at www.cladistics.org.
Holtz, T.R., Jr. 2001. Arctometatarsalia revisited:the problem of homoplasy in reconstructingtheropod phylogeny. In J.A. Gauthier and L.F.Gall (editors), New perspectives on the originand evolution of birds: proceedings of the in-ternational symposium in honor of John H. Os-trom: 99–124. New Haven: Peabody Museumof Natural History, Yale University.
Kurzanov, S., and H. Osmolska. 1991. Tochisau-rus nemegtensis gen. et sp. n., a new troodontid(Dinosauria, Theropoda) from Mongolia. ActaPalaeontologica Polonica 36(1): 69–76, pls. 11,12.
Makovicky, P.J., and M.A. Norell. 1998. A partialornithomimid braincase from Ukhaa Tolgod(Upper Cretaceous, Mongolia). American Mu-seum Novitates 3247: 1–16.
Makovicky, P.J. and H.-D. Sues. 1998. Anatomyand phylogenetic relationships of the theropoddinosaur Microvenator celer from the LowerCretaceous of Montana. American MuseumNovitates 3240: 1–27.
Maryanska, T., and H. Osmolska. 1997. The quad-rate of oviraptorid dinosaurs. Acta Palaeonto-logica Polonica 42: 361–371.
Nixon, K.C. 1999. Winclada (Beta) version 0.9.9.Software published by the author, Ithaca, NY.Available on-line at www.cladistics.org.
Norell, M.A., P.J. Makovicky, and J. M. Clark.2000. A new troodontid theropod from Ukhaa
30 NO. 3402AMERICAN MUSEUM NOVITATES
Tolgod, Mongolia. Journal of Vertebrate Pale-ontology 20(1): 7–11.
Norell, M.A., J.M. Clark, and P.J. Makovicky.2001. Relationships among Maniraptora: prob-lems and prospects. In J.A. Gauthier and L.F.Gall (editors), New perspectives on the originand evolution of birds: proceedings of the in-ternational symposium in honor of John H. Os-trom: 49–68. New Haven: Peabody Museum ofNatural History, Yale University.
Osborn, H.F. 1924. Three new Theropoda, Pro-toceratops zone, central Mongolia. AmericanMuseum Novitates 127: 1–16.
Osmolska, H. 1987. Borogovia gracilicrus gen. etsp. n., a new troodontid dinosaur from the LateCretaceous of Mongolia. Acta PalaeontologicaPolonica 32(1,2): 133–150, pls. 53, 54.
Osmolska, H., and R. Barsbold. 1990. Troodon-tidae. In D.B. Weishampel, P. Dodson, and H.Osmolska (editors): The Dinosauria: 259–268.Berkeley: University of California Press.
Osmolska, H., E. Roniewicz, and R. Barsbold.1972. A new dinosaur, Gallimimus bullatus n.gen., n. sp. (Ornithomimidae) from the UpperCretaceous of Mongolia. Paleontologica Polon-ica 27: 103–143.
Ostrom, J.H. 1969. Osteology of Deinonychus an-tirrhopus, an unusual theropod from the LowerCretaceous of Montana. Bulletin of the Pea-body Museum of Natural History 30: 1–165.
Perez-Moreno, B.P., J.L. Sanz, A. D. Buscalioni,J.J. Moratalla, F. Ortega, and D. Rasskin-Gut-man. 1994. A unique multitoothed ornithomi-mosaur from the Lower Cretaceous of Spain.Nature 370: 363–367.
Russell, D.A. 1969. A new specimen of Stenon-ychosaurus from the Oldman Formation (Cre-taceous) of Alberta. Canadian Journal of EarthSciences 6: 595–612.
Russell, D.A., and Z.-M. Dong. 1993. A nearlycomplete skeleton of a troodontid dinosaurfrom the Early Cretaceous of the Ordos Basin,Inner Mongolia, People’s Republic of China.Canadian Journal of Earth Sciences 30: 2163–2173.
Sereno, P.C. 1999. The evolution of dinosaurs.Science 284: 2137–2147.
Sereno, P.C. 2001. Alvarezsaurids: Birds or orni-thomimosaurs? In J.A. Gauthier and L.F. Gall
(editors), New perspectives on the origin andevolution of birds: proceedings of the interna-tional symposium in honor of John H. Ostrom:69–98. New Haven: Peabody Museum of Nat-ural History, Yale University.
Varrichio, D.J. 1997. Troodontidae. In P.J. Currieand K. Padian (editors), Encyclopedia of di-nosaurs: 749–754. San Diego: Academic Press.
Walker, A.D. 1985. The braincase of Archaeop-tryx. In The beginnings of birds, M.K. Hecht,J.H. Ostrom, H., Viohl, and P. Wellnhofer (ed-itors):123–134. Eichstatt: Freunde des Jura-Museums.
Wellnhoffer, P. 1974. Das funfte Skelettexamplervon Archaeopteryx. Palaeontographica Abtei-lung A 147: 169–216.
Wilson, M.C., and P.J. Currie. 1985. Stenonycho-saurus inequalis (Saurischia: Theropoda) fromthe Judith River (Oldman) Formation of Alber-ta: new findings on metatarsal structure. Cana-dian Journal Earth Sciences 22: 1813–1817.
Witmer, L.M. 1990. The craniofacial air sac sys-tem of Mesozoic birds (Aves). Zoological Jour-nal of the Linnean Society 100: 327–378
Witmer, L.M. 1997a. The evolution of the antor-bital cavity of archosaurs: a case study in soft-tissue reconstruction in the fossil record withan analysis of the function of pneumaticity.Journal of Vertebrate Paleontology 17: 1–73.
Witmer, L.M. 1997b. Craniofacial air sinus sys-tems. In P.J. Currie and K. Padian (editors), En-cyclopedia of dinosaurs: 151–159. San Diego:Academic Press.
Xu, X., X.-l. Wang, and X.-C. Wu. 1999. A dro-maeosaurid dinosaur with a filamentous integ-ument from the Yixian Formation of China.Nature 401: 262–266.
Xu X, Z. Zhou, and X.-l. Wang. 2000. The small-est known non-avian theropod dinosaur. Nature408: 705–708.
Xu, X., X. Zhao, and J. Clark. 2001. A new ther-izinosaur from the Lower Jurassic lower Lu-feng Formation of Yunnan, China. Journal ofVertebrate Paleontology 21: 477–483.
Xu, X., M.A. Norell, X.-L. Wang, P.J. Makovicky,and X.-C. Wu. 2002. A basal troodontid fromthe early Cretaceous of China. Nature 415:780–784.
2003 31MAKOVICKY ET AL.: BYRONOSAURUS JAFFEI
APPENDIX 1
ANATOMICAL ABBREVIATIONS
II-2 pedal phalanx II-2aof antorbital fenestraatr anterior tympanic recessboc base of occipital condylebot basioccipital tubercc common carotid foramencf carotid foramencif interfenestral crestcnc cnemial crestcnV cranial nerve vcnVI foramen for cranial nerve VIcnVII foramen for cranial nerve VIIcn X,XI foramen for cranial nerves X and XIcnXII foramen for cranial nerve XIIcsu caudal vertebral sulcusctr caudal tympanic recessdg dentary groovedr dorsal ribdtr dorsal tympanic recessen entrance to nasal chamberep epipophysisfic fibular crestfm foramen magnumfnc floor of nasal chamberfo foramen ovalefp foramen pseudorotundafsac foramen for supraalveolar canal
(5 subnarial foramen)hf hypophysial fossahy hypapophysisiac interantorbital canalin internarial barifb interfenestral barld left dentaryll left lacrimallmx left maxillaln left nasal
lpmx left premaxillals left splenialmbg groove separating forebrain and mid-
brain of endocastmf metotic foramenmg mandibular groovems metotic strutmxf maxillary fossanar narisnc nasal chamberns neural spineotc otosphenoidal crestpn pneumatic spacepop paroccipital processprz prezygapophysisps palatal shelfpvh posteroventral heelpz postzygapophysisr reconstructedra right angularrd right dentaryrf right fibularj right jugalrl right lacrimalrmx right maxillarn right nasalrpmx right premaxillars right splenialrsa right surangularrt right tibiasac supraalveolar canalsf surangular foramensor subotic recesst toothv vomerwaof wall of antorbital fossa
32 NO. 3402AMERICAN MUSEUM NOVITATES
APPENDIX 2MEASUREMENTS OF THE HOLOTYPE SPECIMEN IGM
100/983 (in mm)
a This paper meets the requirements of ANSI/NISO Z39.48-1992 (Permanence of Paper).