Outer Membrane Protein A Gene Sequencing Demonstrates the Polyphyletic Nature of Koala Chlamydia pecorum Isolates
MICHAEL JACKSON, PHILIP GIFFARD and PETER TIMMS
Centre for Molecular Biotechnology, School of Life Science, Queensland University of Technology, Brisbane, Australia
Received November 18, 1996
Summary
Chlamydia are considered to be the most important pathogen of koalas in which they cause ocular and urogenital infections. As recently as 1996 it was realised that koala chlamydial infections do not belong to the species Chlamydia psittaci but instead should be reassigned to the species C. pecorum and C. pneumoniae. We have used DNA sequence analysis of part of the chlamydial major outer membrane protein gene, ompA VD4, to compare 15 koala C. pecorum isolates. Unexpectedly, we found that the koala isolates did not cluster as a single branch in the C. pecorum tree, but instead were represented by five genetically very distinct genotypes. Two of the genotypes (which contained five koala isolates each) were koala-specific whereas one genotype contained a single koala isolate plus three sheep and two cattle isolates. For all five koala genotypes, their nearest relatives were not other koala genotypes, but sheep, cattle or pig isolates. It may be inferred from our data that C. pecorum strains infecting koalas do not form a monophyletic group with respect to other C. pecorum strains, and therefore the model which states that there was a single acquisition of a C. pecorum infection by a koala and that all C. pecorum strains now infecting koalas are descended from that founding strain is unlikely to be correct. The most plausible model is that koalas have obtained C. pecorum infections as a result of a series of cross-species transmission events, possibly from pigs and/or ruminants.
Koalas (Phascolarctos cinereus) are classified as uncommon marsupials which have a restricted range (Australia), no living close relatives, and in some geographical areas are genetically homogeneous due to previous population bottlenecks (HOULDEN et ai., 1996). As a consequence, the conservation of this animal is regarded as being an important issue. Chlamydial infection in koalas can lead to clinical manifestations such as keratoconjunctivitis, rhinitis, urinary tract disease and infertility and has been recognised as an important threat to the long term survival of this species (BROWN et ai., 1987). In some koala populations overt chlamydia I infections are common and can lead to a significant reduction in fertility (MARTIN, 1981) while in other populations the level of clinical disease is low, despite modest level of infection (WEIGLER et ai., 1988). Nevertheless, chlamydial infection is still regarded as the most serious infectious disease affecting this animai.
Previous discussion regarding the longer term impact of chlamydial disease on koalas has been based on opposing views of the history of this disease in koalas. One view is that the chlamydial parasite has reached an equilibrium with its host over a long period of time. It could be regarded as an opportunistic pathogen that only causes significant disease in individuals compromised in some way. The corollary to this is that high levels of reduced fertility and mortality only occur as a result of stress due to habitat disturbance or other human activities, rather than the simple presence of the parasite. The second view is that chlamydial infection is a serious disease in its own right and that the relevant chlamydial strains are not "normal" colonists of the koala's mucosal surfaces. To date, neither scenario has been supported strongly by available data. The aim of the experiments reported here were to use DNA sequencing to determine the genetic diversity and evolu-
188 M. JACKSON et al.
Table 1. Grouping of animal chlamydial strains based on ompA VD4 sequence comparisons (sequences available in the literature plus those generated in this study).
Chlamydial strain Host(s)
• C. trachomatis S45 porcine MoPn murme
• C.psittaci MN avian, bovine, human 6BC avian, human B577 ovine, avian, human BAI bovine OA.S26.3 ovme LW508 bovine Avian type C aVian FEPN feline FEPN/pud feline GV caprine P18 aVian GPIC gumea pig EAE.A22.M unknown
KALTENBOECK et aI., 1989 MOULDER et aI., 1984, EVERETT et aI., 1991 PEREZ-MARTINEZ and STORZ, 19851
GRIFFITHS et aI., Accession No. L39020 HERRING et aI., 1989 PEREZ-MARTINEZ and STORZ, 19851
STOREY, Accession No. L25436 PEREZ-MARTINEZ and STORZ, 19851
JACKSON et al. (this work) JACKSON et al. (this work) JACKSON et al. (this work) ZHANG et aI., 1989 PICKETT et aI., 1988
GIRjES et aI., 19941
STOREY et aI., 1993
PEREZ-MARTINEZ and STORZ, 1985 1
PEREZ-MARTINEZ and STORZ, 19851
PEREZ-MARTINEZ and STORZ, 19851
PEREZ-MARTINEZ and STORZ, 19851
ANDERSON et aI., Accession No. Z18576 JACKSON et al. (this work) JACKSON et al. (this work) JACKSON et al. (this work) JACKSON et al. (this work) JACKSON et al. (this work) JACKSON et al. (this work) JACKSON et al. (this work) JACKSON et al. (this work) JACKSON et al. (this work) JACKSON et al. (this work) JACKSON et al. (this work) JACKSON et al. (this work) JACKSON et al. (this work) JACKSON et al. (this work) JACKSON et al. (this work) JACKSON et al. (this work) JACKSON et al. (this work) JACKSON et al. (this work)
tionary relationships of koala chlamydial isolates to
each other and to other animal chlamydial isolates, and thereby to elucidate details of the natural history of this disease.
have been classed as C. psittaci (GRAYSTON et aI., 1989, FUKUSHI and HIRAI, 1992). C. trachomatis and C. pneumoniae are found mainly in humans while C. psittaci and C. pecorum parasitise a wide range of animals and birds. We have recently characterized a number of koala chlamydial isolates and found that they should all be assigned to one of two species: C. pecorum and C. pneumoniae, and not C. psittaci as previously thought (GLASSICK et ai., 1996). We have also obtained evidence that
The genus Chlamydia is currently divided into four species: C. trachomatis, C. psittaci, C. pneumoniae, and C. pecorum. The taxa C. pneumoniae and C. pecorum have recently been created using primarily genome-based criteria and accommodate isolates that previously would
Outer Membrane Protein A Gene 189
Table 2. Koala isolates from which chlamydia I ompA VD4 sequence data was generated in this study
Genotype Koala isolate Sex TWCl Site CS2 Geographical location
A LPDowell-V female 10 UGT + Lone Pine Koala Sanctuary, QLD A MT113-V female 1 UGT + Mutdapilly, QLD A MT213-V female 6 UGT + Mutdapilly, QLD A RBNar-O female 9 Ocular + Redland Bay, QLD A CS9745-V female 5 UGT + Currumbin Sanctuary, QLD A CS9762-V female 3 UGT + Currumbin Sanctuary, QLD A PMKala-O female 4 Ocular + Port Macquarie, NSW B RI28-V female na UGT + Raymond Island, VIC C FWPBilb-PJ male 10 UGT Featherdale Wildlife Park, NSW D K4 male na na na Australia E K13 na na UGT + Australia E EMDuar-V female na UGT + Emerald, QLD E PMRior-V female 10 UGT + Port Macquarie, NSW E FWPScar-V female mature UGT + Featherdale Wildlife Park, NSW E FWPBoss-P male mature UGT + Featherdale Wildlife Park, NSW
Legend: RBNar-O - population (RB=Redland Bay); animal designator (Nar - Narelle); swab site (0 - ocular); 1 Tooth wear class was used to estimate koala age (GORDON et ai., 1991); 2 Clinical status at the time of sampling; 3 Koala originally from Victoria, but relocated to Featherdale Wildlife Park, Sydney, New South Wales; 4 This koala had ocular discharge at the time of swabbing, but no clinical disease symptoms at the urogenital site.
Table 3. Percent nucleotide dissimilarity of koala C. pecorum ompA VD4 sequences. Sequences from position 192-400 were used in the analysis.
~ ~ v ~ ~ 0 ~ 0-
'"'"' '"'"' .:. V") ~ 'T ... 2:l M M ro 0 0 M N Z I'--~
ro Q G\ -0 f-< f-< v :::8 :::8
[:Q ~ Vl l? ~ ~ .....I U
A MT113-V A MT213-V 0 A RBNar-O 0 0 A LPDoWell-V 0 0 0 A CS9745-V 0 0 0 0 A CS9762-V 0 0 0 0 0 A PMKala-O 0.7 0.7 0.7 0.7 0.7 B RI28-V 17.8 17.8 17.8 17.8 17.8 C FWPBilb-P 20.0 20.0 20.0 20.0 20.0 D K4 24.2 24.2 24.2 24.2 24.2 E EMDuar-V 28.5 28.5 28.5 28.5 28.5 E K13 28.5 28.5 28.5 28.5 28.5 E FWPScar-V 28.5 28.5 28.5 28.5 28.5 E FWPBoss-P 28.5 28.5 28.5 28.5 28.5 E PMRior-V 28.5 28.5 28.5 28.5 28.5
much of the observed clinical symptoms of chlamydia I disease in koalas is due to C. pecorum (unpublished observations). Our identification of koala isolates to the species level was achieved by sequence determination of an ampB fragment. OmpB is a highly conserved cysteinerich protein that is thought to have an important role in maintaining the structural integrity of the elementary body envelope (HATCH et aI., 1984). GLASSICK et al. (1996) found that although ampB sequences from the
two species were approximately 30% diverged, there was zero divergence between sequences from the same species, and so no possibility of determining intra-species relationships. Accordingly, we have now made use of a highly variable fragment of the ampA gene. OmpA is an outer membrane protein that contains four highly variable domains, probably as a consequence of immune selection (STEPHENS et aI., 1987). This gene has previously been used in studies of Chlamydia phylogeny
190 M. JACKSON et al.
,Brisbane
Sydney
13 = unknown
500 kms
Fig. 1. Geographical distribution of koala (_), sheep (e) and cattle (.A.) chlamydial isolates used in this study. Letters A to E indicate Chlamydia pecorum genotypes based on ompA VD4 sequence data.
(KALTENBOECK et al., 1993). In this communication, we report the determination of the relationships between ompA variable domain IV (VD IV) sequences from a number of koala and non-koala C. pecorum isolates.
Materials and Methods
Chlamydial strains studied: A total 25 new chlamydia I isolates (15 from koalas, 10 from other animals) were sequenced at the ompA VD4 locus. Additional ompA sequence data for five strains of C. trachomatis, three strains of C. pneumoniae, 10 strains of C. psittaci and five strains of C. pecorum were obtained from the literature (Table 1) and used for phylogenetic comparison. Fifteen Chlamydia-positive swab samples were collected from captive and free-range koalas from various geographical regions across eastern Australia (Figure 1). Table 2
summarises the geographical location, sex, tooth wear class (an estimate of age) and clinical symptoms of each koala. Based on the results presented in this study, each cluster of koala C. pecorum ompA VD4 sequences has been designated as a genotype (A to E; see Results section). Ten additional Australian Chlamydia isolates from ovine, bovine, caprine, avian and feline hosts were also sequenced as part of this study. B1 = C. pecorum bovine abortion isolate (Townsville, Queensland, Australia); B3 = C. pecorum bovine isolate (Australia); BA8411918 = C. pecorum bovine abortion isolate (Townsville, Queensland, Australia); 511 = C. pecorum ovine polyarthritis isolate (Australia); 517 = C. pecorum ovine liver/kidney isolate (Armidale, N5W, Australia); 519 = C. pecorum ovine ocular isolate (Armidale, N5W, Australia); 521 = C. pecorum ovine polyarthritis isolate (Armidale, N5W, Australia); FEPN/pud = C. psittaci feline pneumonia isolate (Australia); P18 = C. psittaci avian isolate (Australia); GV = C. psittaci caprine vaginitis isolate (Australia).
PCR amplification and sequencing of the chlamydial ompA VD4 region: 5wab and cell culture material was prepared for PCR assay by heating 70 flL of thawed and vortexed sample at 95 °C for 20 minutes, causing lysis of the Chlamydia elementary bodies. Following centrifugation, two microlitres of this supernatant was added to a PCR mixture containing lOx PCR reaction buffer (1.5 mM MgCI2, 50 mM KCI, 10 mM TrisHCI, pH 8.0; Perkin Elmer-Cetus), 0,2 mM deoxynucleotide triphosphates (dATP, dGTP, dCTP, dTTP; Pharmacia, California), primers MJFvd3 (GGITG(CT)GCAACTTTAGGIGC) orMJFvd4(TACCATGAATGGCAAGT) and MJRvd4 (CACAAGCTTTTCTGGACTTC) (1 pmollflL each; Bresatec), 1.2 U Taq polymerase (Perkin Elmer-Cetus), and overlaid with mineral oil. The PCR was performed on a Perkin Elmer-Cetus DNA Thermal Cycler 480. PCR cycling conditions were a single cycle of initial denaturation (94 °c, 5 minutes) and 35 cycles of denaturation (94 °c, 1 minute) primer annealing (54 °c, 1 minute) and primer extension (72 °c, 1 minute). The PCR product was separated by electrophoresis in 1.0% low melting agarose/TBE (45 mM Tris-borate and 1 mM EDTA, pH 8.0) for 60 minutes at 100V. The initial target for PCR and sequencing was the ompA VD4 region. Amplification of this DNA fragment was performed on isolates K4, K13, BA8411918, B1, B3, 517, 519, 521, GV, P18 and FEPN/pud. However, in the second portion of this work, the segment that was amplified and sequenced was expanded to include both the VD3 and VD4 regions (koala isolates MT113-V, MT213-V, RBNar-O, LPDowell-V, C59765-V, PMKala-O, RI28-V. FWPBilb-P, EMDuar-V, FWPBoss-P, PMRior-V and FWP5car-V). For this reason, two sets of primers were used (MJFvd41 MJRvd4 and MJFvd3/MJRvd4) which produced fragments of 270 and 470 base pairs, respectively. The desired fragment was excised from the gel and purified using the WIZARD purification system (Perkin Elmer-Cetus). Approximately 100 ng of amplified PCR product was used for each sequencing reaction. To reduce the risk of PCR contamination, a negative control was performed between each tube containing Chlamydia DNA template. Repeat PCR and repeat sequencing in both directions using forward (MJFvd3 or MJFvd4) and reverse primers (MJR vd4) allowed any anomalies in the amplified sequence to be resolved.
Phylogenetic analysis: 5equence alignment and phylogenetic tree construction were carried out using Clustal W (THOMPSON et aI., 1994) which makes use of the distance based neighbourjoining (NJ) tree building algorithm (5AITOU and NEI, 1987). Confidence limits on branching orders were tested by bootstrap analysis (FELSENSTEIN, 1985) using the NJ and bootstrap application in Clustal W (THOMPSON et aI., 1994).
· .......... T . G .• T •• CT • G ..•..••...••.• G .• C .• T ...•• T . GG ... A .. .
· .•.....•.. T .G .. T .. CT.G ..•.......•..• G .. C .. T ••.•. T .GG ... A .. .
· .•....••.. T .G .. T .. CT.G .•••.••••.••.• G .• C •• T ••... T .GG ... A .. .
· .•..•..•.• T .G .. T .. CT.G .•••.•••..••.. G .• C .• T ••... T .GG •.. A .•.
.•••.•.••.• T.G .. T •. CT.G .••••••••••.....••.• T ••.....•••..•.•.
•.•.••.••....•..•••.. • A •. C •••••.•• G .• G .••••• A .•.. T •.•......•
• •••••••••• T • A •••.••••...••••..•••..• G .• C •• T ••.•. T • GG ••. A •..
· .•..•••••. T • G .. T .• CT • G •••••••••••••••••••. T •..•..•.••..• GG .
.•••••••••••••••••.•• A •• C .•••.••• G •• G •••••.••.••. T •.•••.••.•
.•.•••.•••• T.A •••..••..••.•••..••.••.••.•..•..••.••. G .••••.•
· .••.•........•..•...••...•...•..•.....•... T ......•.•...• GG •
Fig. 2. Nucleotide sequence alignment of ompA VD3ND4 of all chlamydial isolates sequenced in this work. Nucleotide sequences for isolates 66P130, L71, 1710s and LW613 were previously sequenced (KALTENBOECK et aI., 1993). Position 1 ofthe MT113-V sequence corresponds to position 482 of previous ompA sequence data (KAlTENBOECK et aI., 1993). Dots represent nucleotides of aligned sequences which are identical to the MTl13-V sequence. Letters represent variation in nucleotide sequence and solid lines indicate gaps. The variable domain four of all sequences is shaded.
192 M. JACKSON et al.
MT1l3 V
MT213-V
RB ar 0
LPDow V
CS9745 V
CS9762-V
PMKala-O
EMDua -V
FWPBos P
PMRior-V
FWPSc r-V
K 3
K4
.G .. TG .. M.C
.G. .C
.G TG .. M.C
. G TG .. .C •. A ••
CG TGCAG CG GC -----
TC.C ......... AGCA.C ATTC.
TC C ..
.C ..
AGCA CTGATTC
GCA CTG TTC
.G
.G
.G
TC.C ........ AGCA.CTG TTC AA G
120
FWPB11b P .T .... CC.G .. C ........... G ............ G TGC
DNA amplification, sequence determination and sequence alignment
VD4 and/or VD3ND4 containing ompA fragments were successfully amplified by peR from both swab and cell culture material, and all amplified DNA was sucessfully sequenced. The nucleotide sequences were aligned (Fig. 2) and a dissimilarity matrix for the sequences obtained from koala isolates was deduced from this alignment (Table 3).
Designation of koala C. pecorum genotypes
It is clear from Table 3 that the sequences from koala isolates fall into five groups. We have designated these genotypes A-E. The location of the hosts of each koala isolate, together with the deduced genotype is shown in Fig. 1. It may be seen from this that in the case of genotype A, there is some association between the site of koala capture and the genotype of its resident C. pecorum strain, while other genotypes were found over a wide area (eg. genotype E), or were represented by single
A.A.A ••....•..•• C .•...•..•• . C .• C .......•.•.••....•. . G .•...••
A.A.A ....•.•.... C ....•.••.. • C .• C . ••.....••••.....•.. G ...•..•
A.A.A .•.••••.. •• C . ........ .. C .. C . .•••...••••..•.•..• G ......•
A.A.A ....•..... • C . .....••. .• C . • C .•••.• , .••••.••..•.. G •.••.•.
K4
FWPBilb-P
RI28-V
BA84/1918
B1
.•• GT •. A ••...... A .. A ..•.. T •• C .••.•••.••.•.••..••••.. G •....••
B3
66P130
P787
L71
•• A .••• A .•..••.• A .• T •• T •• T •. C •.••..••...•.. C •• C •••.• G .•..•..
ATA.A ••••.••••.. C ..••.....•.....•. A •.••.•. . C •• ..•..• A .••.• A.
••...•....••..••••. C ...•. T .•....•. A •••••......•.••.• A .•..••.
1710s
S17
S19
S21
GV
FEPN/pud
P1S
LW613 ATA.A •........•. C .•..•..••. • C . .•.. A •..•.•.. C .....•.. A •••.• A.
Fig. 2 (3)
strains (eg. genotypes B, C and D). Six genotype A isolates were from South East Queensland (within 100 km of Brisbane) while the seventh isolate in this group (PMKala-O) was from Port Macquarie which is approximately 450 km South of Brisbane. Interestingly, all genotype A sequences were identical except for PMKala-O which differed at two positions.
Also shown in Fig. 1 is the location of isolation of a number of C. pecorum strains from ruminants. The ompA fragments from these strains were also sequenced in the course of this work. These have been designated C. pecorum genotype D as the sequences obtained were identical to that of
koala isolate K4 which has also been designated genotype D. Unfortunately, the history of K4 with respect to location of host capture and clinical disease status is unavailable.
The relative similarities of C. pecorum ompA fragment sequences
In order to more clearly visualize the relationships between the available sequences, and also to estimate the statistical significance of any clustering of sequences, the alignment in Fig. 2 was converted into a tree using the
• C . . • . • . . C. . • • . • CA • C . . T. . T. . T. . AG • G. • . . • . • • G. • • . . • T. . . . T. . • .
• C . • . . • . . C. . • . . • CA • C . • T. . T. . T. . AG • G. • . . . . . • G. . • . . . T. . . . T. . . .
• C . . . . . . . C. . . . . . CA • C . • T. . T. . T. . AG • G. . . . . . • . G. . . . . . T. . . . T. . . .
• C ....•.. C ..•... CA • C .• T .. T .. T .. AG . G ......•. G .•.... T •... T ....
- - - - - - - - - - - . • . . . CA • C . • T. • T. . T. • AG . G. . . . . . • • G. . . . . • T. . . . T. . . .
----------- .•... C ..•.. T ••..•..• TG .•..•..•.•...•...•. T .. T •...
· C. . • . . . • . . . • • . . CA. • . • T. . T. • . . • AG . G. . • . . • • . G. . • . . . . . T . . T. . . .
.••.••.•••••••••••.••..••.••..••.•••..• A ••.••••.•... T •. T ...•
----------- .•... C ..••• T ••.••..• TG .••.•..•.•..••.•.•• T .. T ...•
- - - - - - - - - - -. . . • . CA. . • . T. • T. • . . . AG . G. . • . . • • . G . • . . • . • . T. • T. • . •
- - - - --- -- - - •..•. CA ..•. T •. T ••..• AG • G .•..•..• G •.....•. T .• T ...•
.C .. A ••••.••..•.••.••.••.•...•.. G ..••.••.••......••...•...•.
•..• A .. G ...•... . C •••. • T ..•...•. TG •..•..•.••••••.••.• T .. T ... .
.C .• A .•••..•....•.....•.....••..••..•..•.•..••...•.• T .. T ... .
• .•. G ••.•••••..•.•••.••.•••••••• G ••••.••••••.••.•..•...•.••.
----------- ••... C .•..• T ........ TG •.•...••••..••....• T .. T ... .
- - - - - -- - - - - ••.•• C ••..• T ........ TG .••.•..•.•..•...... T .• T ..•.
----------- ...•. C .•... T ........ TG ...••..•.••.••••.•• T •. T ... .
----------- ...........••. T •• T ••• G.AA.C .•..• A .•..• A .. T .. T ...•
- - - - - -- - - - - •....•..••..•..•. T .• GG • AA . T .•..• A •••...•. T ...•..•
----------- ........•..••. T .. T .•. G.AA.C ..... A ..••. A .. T .. T ..••
••.. A .. G .••..•.• C ..••. T •.•••.•• TG •........••..••.•.• T •• T ....
Neighbour-joining algorithm (SAITOU and NEI, 1987) and the significance of the branching order tested by bootstrapping (FELSENSTEIN, 1985) (Fig. 3). Also included in the analysis were chlamydial ompA sequences previously obtained by others.
(sheep, cattle, pigs). This is most striking in the case of K4 (genotype D), which, as previously indicated, gave a sequence identical to that from isolates of Australian bovine origin (eg. BA8411918). It is most interesting that this sequence is also 100% identical to the sequence obtained from C. pecorum LW613, a European bovine isolate. Koala isolate RI28-V(genotype B) gave a sequence very similar to that of C. pecorum strain L71, a European porcine isolate. The sequence from koala isolate FWPBilb-P (genotype C) is similar to that from the Australian bovine abortion isolates, B1 and B3.
It is clear that the sequence diversity within the koala isolates is quite considerable (up to 29.5%) which is comparable to the sequence diversity within the genus C. pecorum as a whole. Also, there appears to be a specific relationship between several koala isolates (genotypes B, C and D) and several non-koala C. pecorum isolates
· G ................. A .. A .... A . •. G .. C ••. C .. G. TC •. A •• A.G ..•.. A.
.G ................. A .. A .... A . .. G .• C .•. C •. G.TC •. A .. A.G ..••. A.
· G ................. A .. A .... A ... G .. C .•. C .. G. TC .. A .. A.G ..•.. A .
. G ................. A .. A .... A .•• G .. C ... C .. G.TC .. A .. A.G .••.. A.
.G .............. ... A .. A .... A .•• G •• C •.• C .. G. TC •. A .• A.G •.••• A.
· ................... G . A .... A ..••.•••.• C . A •. T •.. A. CT • G .. A •. A •
.... C .. A .. T ........ T ....... A ...•..••.• C .••..••.••.••.••••.••
· . . . . . . . . . . . . . . . . .......... A . .••...•.• C . A •• T ••. A • CT • G •• A •• A •
· . . . . . . . . . . . . ....... G . A .... A .•.....•.. C • A .• T ... A . CT . G .. A .. A .
.... C .. A .. T . ....... T ... . ... A . ••.••.... C ••.••..•.....••.••••.
.... C .. A .. T .... .... T ....... A .••.••..•• C ••..••.••.••.••.••.•.
· .... . . C .. T ........ A ....... A ••.••. A.AG.AG.T .. GA.CT ..•. A •. A.
· ................... G . A .... A .• •....•.. C • A •. T ... A • CT • G .• A .. A •
· . . . . . . . . . . . . . G. . . . . ....... A . C ....••. AC . A •. T ... A . CT • G .• A .. A .
· ...... C .. T ................ A. C . G ...•.. C ...• T •• TAGG .• G ....• A.
· ................... G .A .... A .......... C .A .• T .•. A.CT .G .• A •. A.
· ................... G .A .... A ..•.•.••.• C .A .• T ••. A.CT .G .• A •• A.
· ................... G . A .... A .••••.•..• C • A .• T ..• A. CT . G •. A .. A.
· T .......... GC .. .. . T .. A .. T .A .• .. AAA •.• GAG .• TC. TA.CA. T ..•.• A.
· T . . C . ...... GC ..... T .. A .. T .A ..• G •• C •. AG.A .•. C .AA.CT .... A • • C.
· T .......... GC ..... T .. A .. T.A . ••. AAA ••. GAG .• TC .TA.CA. T •..••.•
· ................... G . A .... A .•...•..•. C . A .. T .•• A . CT . G •. A ••..
If it is assumed that the lineage of the ompA gene reflects the lineage of the Chlamydia strain from which it is derived, and that the analysis described here provides meaningful information as to ompA lineage, it may be inferred from our data that C. pecorum strains that infect koalas do not form a monophyletic group with respect to other C. pecorum strains. In other words, a model which states that there was a single acquisition of a C. pecorum infection by a koala and that all C. pecorum strains now infecting koalas are descended from that founding strain, is unlikely to be correct. The most plau-
sible model consistent with our data is that koalas have obtained C. pecorum infections as a result of a series of cross-species transmission events, possibly from sheep and/or cattle. However, we are not able to formally rule out the notion that C. pecorum originated in koalas and was subsequently transmitted to pigs and ruminants.
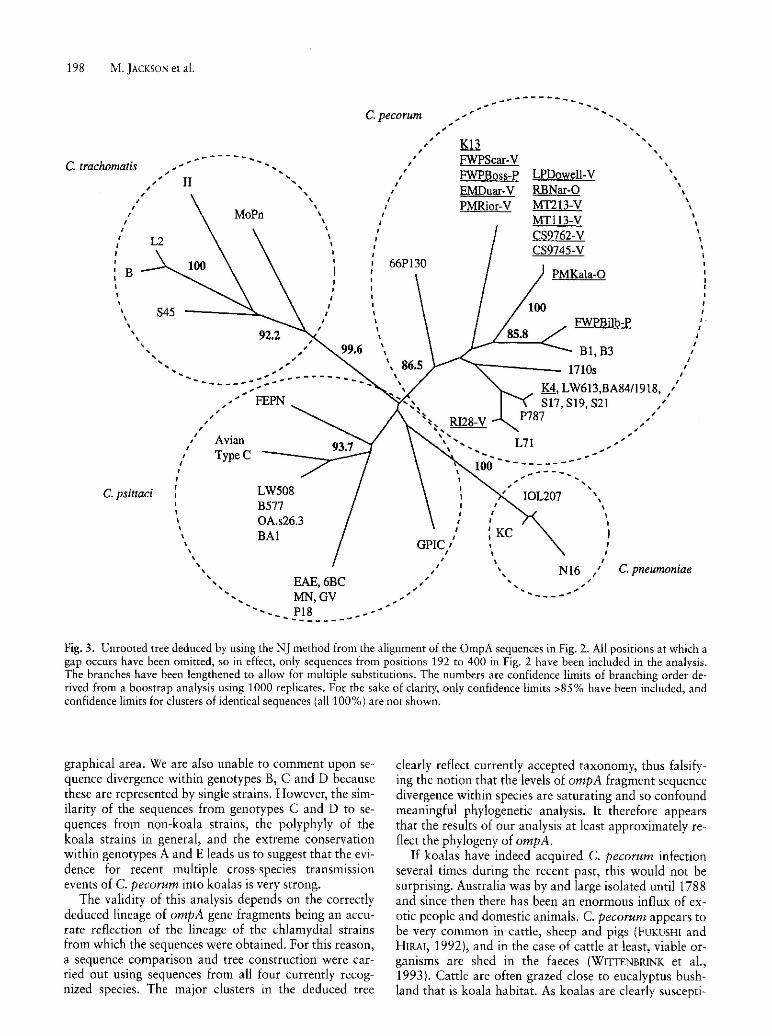
In order to compare the divergence within the ompA gene fragment in koala isolates with that in other Chlamydia species, and also to test whether comparisons using this gene fragment define relationships congruent with that derived from other data, the phylogenetic tree
· ... C .. A .. TC. T .. A •.•. TGGCC ••. A.TTTA------GAC .... C .A .. A •.....
.•.. C .. A.GCC.TA.A ... TC .. CC •. TG.TTTGC---CC.AT.ATAGT.GTA.GG ...
· ...... A ........ A ... C .. GC ... TA .•• ------T.GATGG.ACAA .. A.G. T ..
we constructed included sequences from all four species of Chlamydia (Fig. 3) . It can be seen that the sequences fall into four generally defined clusters each of which is composed of sequences from one of the four currently recognized Chlamydia species. Therefore, analyses using this fragment gives results that are at least broadly consistent with previous data. It can also be seen that the divergence between koala C. pecorum strains is comparable to the total divergence found in C. trachomatis and C. psittaci, and is greater than the divergence observed within C. pneumoniae.
Discussion
At the commencement of this study, we anticipated two possible outcomes to this analysis of ompA fragments from koala Chlamydia strains. The first was that all the sequences would be very similar and form a single cluster in the tree. This would indicate that koalas had acquired C. pecorum relatively recently, or that a single clone had recently become very successful and had risen to prominence. The second was that the sequences were highly diverged, indicating that there had
------ · ................................. . ------ .. . . . . . . . . . . . . ..................... . ------.A ... A. TT ....... A .. G .... G ........ .
------ · ................................ . ------ ..... A. T .. G ..... A ................ .
------.A .. TG. TT ........................ .
------ · ................................ . ------ - ................................. . TTCTAT .... TGT. T .G ....... C ... GA .... G .. C ..
------ .•.... T .A.G ..... C ...... A.G ..... C ..
TTCTAT .... TGT. T.G ....... C ... GA .... G .. C ..
..... _-_ ... ................................ .
been extensive evolution since koalas first acquired this parasite. The result we did obtain was a third unexpected alternative. We found a very high level of divergence in the koala C. pecorum sequences, but two of these sequences clustered with other sequences from non-koala isolates with a high level of statistical significance, and there are no highly significant clusters that contain more than one genotype that do not also contain sequences from non-koala strains. It is therefore highly unlikely that all the koala C. pecorum strains are descended from a single common ancestor that was origi-
nally acquired by koalas in a single cross-species transmission event.
Our analysis yielded several isolates of genotypes A and E. Within these genotypes the sequences obtained were either identical (genotype E) or near identical (genotype A). This is consistent with these genotypes being monophyletic within the context of the strains analysed and being acquired by koalas relatively recently. We are of course unable to rule out such phenomena as single clones within a population becoming dominant or clones circulating through multiple hosts in a particular geo-
Fig. 3. Unrooted tree deduced by using the NJ method from the alignment of the OmpA sequences in Fig. 2. All positions at which a gap occurs have been omitted, so in effect, only sequences from positions 192 to 400 in Fig. 2 have been included in the analysis. The branches have been lengthened to allow for multiple substitutions. The numbers are confidence limits of branching order derived from a boostrap analysis using 1000 replicates. For the sake of clarity, only confidence limits >85% have been included, and confidence limits for clusters of identical sequences (all 100%) are not shown.
graphical area. We are also unable to comment upon sequence divergence within genotypes B, C and D because these are represented by single strains. However, the similarity of the sequences from genotypes C and D to sequences from non-koala strains, the polyphyly of the koala strains in general, and the extreme conservation within genotypes A and E leads us to suggest that the evidence for recent multiple cross-species transmission events of C. pecorum into koalas is very strong.
The validity of this analysis depends on the correctly deduced lineage of ompA gene fragments being an accurate reflection of the lineage of the chlamydial strains from which the sequences were obtained. For this reason, a sequence comparison and tree construction were carried out using sequences from all four currently recognized species. The major clusters in the deduced tree
clearly reflect currently accepted taxonomy, thus falsifying the notion that the levels of ompA fragment sequence divergence within species are saturating and so confound meaningful phylogenetic analysis. It therefore appears that the results of our analysis at least approximately reflect the phylogeny of ompA.
If koalas have indeed acquired C. pecorum infection several times during the recent past, this would not be surprising. Australia was by and large isolated until 1788 and since then there has been an enormous influx of exotic people and domestic animals. C. pecorum appears to be very common in cattle, sheep and pigs (FUKUSHI and HIRAI, 1992), and in the case of cattle at least, viable organisms are shed in the faeces (WITTENBRINK et aI., 1993). Cattle are often grazed close to eucalyptus bushland that is koala habitat. As koalas are clearly suscepti-
ble to a wide phylogenetic range of C. pecorum strains, there would appear to be little physical barrier to their acquiring these parasites.
The species C. pecorum is the most divergent of the four current chlamydia I species, not only from a genetic perspective, but also from the perspective of host range (able to infect sheep, cattle, pigs, koalas) and disease spectrum (inapparent infections, polyarthritis, encephalomyelitis, respiratory, pneumonia, urinary incontinence, infertility, abortion, conjunctivitis, blindness). At this stage, C. pecorum appears to be capable of causing only two main types of disease in the koala, ocular infection (conjunctivitis that may lead to corneal scarring and blindness) and urogenital infection (that may cause urinary incontenance or may lead to cystic ovaries and infertility). To date there have not been any well documented cases in koalas of intestinal, respiratory, encephalomyelitis or polyarthritis due to Chlamydia (as is observed in other animal hosts with C. pecorum). Because the 15 koala C. pecorum isolates studied in this work probably represent a biased sample, it is not possible to confidently relate the C. pecorum genotype infecting koalas to the clinical disease type. Nevertheless, it is interesting to note that genotype A (which was confined to koalas and represented by five isolates) was associated with both ocular and urogenital infections, whereas genotype E (which was also confined to koalas and represented by five isolates) was only associated with urogenital infections. A much more extensive geographical and disease spectrum survey will be necessary to determine if particular genotypes are disease-specific, while others cause a range of clinical disease (as is the case with C. pecorum in other animal hosts).
Acknowledgements Special thanks to P. CANFIELD (University of Sydney), Feath
erdale Wildlife Park (Sydney), ]. MARTIN (Monash University), R. BOOTH and]. McKEE (Currumbin Sanctuary), N. WHITE (University of Queensland), P. O'CALLAGHAN and W. BLANSHARD (Lone Pine Koala Sanctuary) for providing many of the samples used in this study. Financial support for this work was provided by Lone Pine Koala Sanctuary and the Australian Koala Foundation.
References
ANDERSEN, A.A., BAXTER, S.I.F, DUNBAR, S., PHILLIPS, H.L., CLARKSON, M.l, HERRING, A.].: Unpublished. Genomic studies of ruminant and porcine strains of Chlamydia support the creation of the new species designation Chlamydia pecorum. Genebank Accession Number Z18756.
BROWN, A.S., A.A. GIRJES, M.E LAVIN, P. TIMMS, and ].B. WOOLCOCK. Chlamydial disease in koalas. Aus. Vet. ]. 64, 346-349 (1987).
EVERETT, K.D.E., ANDERSEN, A.A., PLAUNT, M., HATCH, T.: Cloning and sequence analysis of the major outer membrane protein gene of Chlamydia psittaci 6BC. Infect. Immun. 59, 2853-2855 (1991).
FELSENSTEIN,].: Confidence limits on phylogenies: an approach u~ing the bootstrap. Evolution 39, 783-791 (1985) .
Outer Membrane Protein A Gene 199
FUKUSHI, H., HIRAI, K.: Proposal of Chlamydia pecorum sp. nov. for Chlamydia strains derived form ruminants. Int. ]. System. Bact. 42. 306-308 (1992).
GIRJES, A.A., CARRICK, EN., LAVIN, M.F.: Remarkable sequence relatedness in the DNA encoding the major outer membrane protein of Chlamydia psittaci (koala type I) and Chlamydia pneumoniae. Gene 138, 139-142 (1994).
GLASSICK, T., GIFFARD, P., TIMMS, P.: Outer membrane protein 2 gene sequences indicate that two chlamydial species, Chlamydia pecorum and Chlamydia pneumoniae, cause infections in koalas. System. Appl. Microbiol. Vol. 19: 456-464 (1996).
GORDON, G.: Estimation of the age of the koala, Phascolarctos cine reus (Marsupialia: Phascolarctidae) from tooth wear and growth. Aust. Mammal. 14,5-12. (1991)
GRIFFITHS, P.c., PLATER, ].M., MARTIN, T.c., HUGHES, S.L., HUGHES, K.l, HEWINSON, R.G., DAWSON, M.: Epizootic bovine abortion in a dairy herd: Characterisation of a Chlamydia psittaci isolate and antibody response. Unpublished. (Accession number L39020).
HATCH, T.P., I. ALLAN, and ].H. PEARCE: Structural and polypeptide differences between envelopes of infective and reproductive life cycles forms of Chlamydia spp. ]. Bacteriol. 157: 13-20 (1984).
HERRING, A.l, TAN, T.W., BAXTER, S., INGLIS, N.E, DUNBAR, S.: Sequence analysis of the major outer membrane protein gene of an ovine abortion strain of Chlamydia psittaci. FEMS Microbiol. Lett. 65, 153-158 (1989).
HOULDEN, B.A., ENGLAND, P.R., TAYLOR, A.C., GREVILLE, W.D., SHERWIN, W.B.: Low genetic variability of the koala Phascolarctos cine reus in south-eastern Australia following a severe population bottleneck. Mol. Ecology 5, 269-281 (1996).
KALTENBOECK, B., KONSTANTIN, K.G., STORZ, J.: Structures of and allelic diversity and relationships among the major outer membrane protein (ompA) genes of the four chlamydial species.]. Bact. 175,487-502 (1993).
MARTIN, R.W.: Age-specific fertility in three populations of the koala, Phascolarctos cine reus, Goldfuss in Victoria. Aust. Wildl. Res. 8: 275-283 (1981).
MOULDER, ].W., HATCH, T.P., Kuo, e-c., SCHACHTER, l, STORZ, J.: Genus Chlamydia. Jones, Rake and Steams, 729-739. In N.R. Krieg and J. Holt (eds.), Bergey's Manual of Systematic Bacteriology, vol. ]. The Williams & Wilkins Co., Baltimore. (1984).
NIGG, c.: Unidentified virus which produces pneumonia and systemic infection in mice. Science. 95, 1254-1260 (1942).
PEREZ-MARTINEZ and STORZ, ].A. STORZ,].: Persistent infection of L cells with an ovine abortion strain of Chlamydia psittaci. Infect. Immun. 50,453-458 (1985).
PICKETT, M.A., EVERSON, ].S., CLARKE, I.N.: Chlamydia psittaci ewe abortion agent: complete nucleotide sequence of the major outer membrane protein gene. FEMS Microbiol. Lett. 55,229-234 (1988).
SAITOU, N., and NEI, M.: The neighbour-joining method: a new method for reconstructing phylogenetic trees. Mol. BioI. Evolution 4, 406-425 (1987).
STEPHENS, R.S., SANCHEZ-PESCADOR, R., WAGAR, E.A., INOUYE, c., URDEA, M.S.: Diversity of Chlamydia trachomatis major outer membrane protein genes.]. Bacteriol. 169,3879-3885 (1987).
STOREY, c., LUSHER, M., YATES, P., RICHMOND, S.: Evidence of Chlamydia pneumoniae of non-human origin. ]. Gen. Microbial. 139,2321-2626 (1993).
200 M. JACKSON et al.
THOMPSON, J.D., HIGGINS, D.J., GIBSON, T.].: CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position specific gap penalties and weight matrix choice. Nucleic Acid Res. 22, 4673-4680 (1994).
WEIGLER, B.]., A.A. GIR]ES, N.A. WHITE, N.D. KUNST, EN. CARRICK, and M .E LAVIN. 1988. Aspects of the epidemiology of Chlamydia psittaci infection in a population of koalas (Phascolarctos cinereus) in southeastern Queensland, Australia. ]. Wildl. Dis. 24: 282-29l.
WITTENBRINK, M.M., BISPING, W., MROZEK, M., HORCHLER, H.: Intestinal Chlamydia psittaci infection in cattle-Incidence and technological aspects of chlamydial isolation. Deutsch Tierarztliche Wochenschrift 100, 195-198 (1993).
ZHANG, Y.-X., MORRISON, S.G., CALDWELL, H.D., BAEHR, W.: Cloning and sequence analysis of the major outer membrane protein genes of two Chlamydia psittaci strains. Infect. Immun. 57, 1621-1625 (1989).
Corresponding author: P. TIMMS, Centre for Molecular Biotechnology, School of Life Science, Queensland University of Technology, GPO Box 2434 Brisbane Australia 400l. Tel.: +61 738642120; Fax +61 738641534; e-mail: [email protected].