RESEARCH ARTICLE Oxidative stress status, antioxidant metabolism and polypeptide patterns in Juncus maritimus shoots exhibiting differential mercury burdens in Ria de Aveiro coastal lagoon (Portugal) Naser A. Anjum & Armando C. Duarte & Eduarda Pereira & Iqbal Ahmad Received: 22 October 2013 /Accepted: 20 January 2014 # Springer-Verlag Berlin Heidelberg 2014 Abstract This study assessed the oxidative stress status, an- tioxidant metabolism and polypeptide patterns in salt marsh macrophyte Juncus maritimus shoots exhibiting differential mercury burdens in Ria de Aveiro coastal lagoon at reference and the sites with highest, moderate and the lowest mercury contamination. In order to achieve these goals, shoot-mercury burden and the responses of representative oxidative stress indices, and the components of both non-glutathione- and glutathione-based H 2 O 2 -metabolizing systems were analyzed and cross-talked with shoot-polypeptide patterns. Compared to the reference site, significant elevations in J. maritimus shoot mercury and the oxidative stress indices such as H 2 O 2 , lipid peroxidation, electrolyte leakage and reactive carbonyls were maximum at the site with highest followed by moderate and the lowest mercury contamination. Significantly elevated activity of non-glutathione-based H 2 O 2 -metabolizing en- zymes such as ascorbate peroxidase and catalase accompanied the studied damage-endpoint responses, whereas the activity of glutathione-based H 2 O 2 -scavenging enzymes glutathione peroxidase and glutathione sulfo-transferase was inhibited. Concomitantly, significantly enhanced glutathione reductase activity and the contents of both reduced and oxidized gluta- thione were perceptible in high mercury-exhibiting shoots. It is inferred that high mercury-accrued elevations in oxidative stress indices were obvious, where non-glutathione-based H 2 O 2 -decomposing enzyme system was dominant over the glutathione-based H 2 O 2 -scavenging enzyme system. In par- ticular, the glutathione-based H 2 O 2 -scavenging system failed to coordinate with elevated glutathione reductase which in turn resulted into increased pool of oxidized glutathione and the ratio of oxidized glutathione-to-reduced glutathione. The substantiation of the studied oxidative stress indices and anti- oxidant metabolism with approximately 53-kDa polypeptide warrants further studies. Keywords Salt marsh . Mercury . Macrophyte . Juncus maritimus . Oxidative stress . Antioxidant metabolism . Polypeptide patterns Introduction Salt marshes represent marine ecosystems and perform nu- merous essential ecological functions such as the stabilization of coastal shorelines, preventing erosion and sediments from washing offshore; absorption and trapping of potential pollut- ants before they reach estuaries and fragile waterways; and the biogeochemical cycling and storage of varied metal/metalloid contaminants including mercury (Hg) (Pereira et al. 2009; Anjum et al. 2013a). Extensive studies have evidenced salt marsh macrophyte-mediated considerable control of sediment-associated Hg bioavailability to other biota through differential accumulation and allocation in their organs (Weis and Weis 2004; Anjum et al. 2011, 2012a, 2013a, b). The accumulation of Hg by salt marsh macrophytes and its subse- quent allocation in their organs have been reported to be controlled primarily by factors related to plants (species/ge- notypes, time/stage of exposure, growth stage metals sensitiv- ity), metal/metalloid (level, form and type) and sediments (physico-chemical traits including pH, redox potential, organic matter content) (Reboreda and Caçador 2007; Anjum et al. 2011, 2012a; Figueira et al. 2012). Among the Responsible editor: Elena Maestri N. A. Anjum : A. C. Duarte : E. Pereira : I. Ahmad (*) Department of Chemistry and CESAM-Centre for Environmental and Marine Studies, University of Aveiro, 3810-193 Aveiro, Portugal e-mail: [email protected]I. Ahmad Department of Biology and CESAM-Centre for Environmental and Marine Studies, University of Aveiro, 3810-193 Aveiro, Portugal Environ Sci Pollut Res DOI 10.1007/s11356-014-2578-4
Transcript
RESEARCH ARTICLE
Oxidative stress status, antioxidant metabolism and polypeptidepatterns in Juncus maritimus shoots exhibiting differentialmercury burdens in Ria de Aveiro coastal lagoon (Portugal)
Naser A. Anjum & Armando C. Duarte &
Eduarda Pereira & Iqbal Ahmad
Received: 22 October 2013 /Accepted: 20 January 2014# Springer-Verlag Berlin Heidelberg 2014
Abstract This study assessed the oxidative stress status, an-tioxidant metabolism and polypeptide patterns in salt marshmacrophyte Juncus maritimus shoots exhibiting differentialmercury burdens in Ria de Aveiro coastal lagoon at referenceand the sites with highest, moderate and the lowest mercurycontamination. In order to achieve these goals, shoot-mercuryburden and the responses of representative oxidative stressindices, and the components of both non-glutathione- andglutathione-based H2O2-metabolizing systems were analyzedand cross-talked with shoot-polypeptide patterns. Comparedto the reference site, significant elevations in J. maritimusshoot mercury and the oxidative stress indices such as H2O2,lipid peroxidation, electrolyte leakage and reactive carbonylswere maximum at the site with highest followed by moderateand the lowest mercury contamination. Significantly elevatedactivity of non-glutathione-based H2O2-metabolizing en-zymes such as ascorbate peroxidase and catalase accompaniedthe studied damage-endpoint responses, whereas the activityof glutathione-based H2O2-scavenging enzymes glutathioneperoxidase and glutathione sulfo-transferase was inhibited.Concomitantly, significantly enhanced glutathione reductaseactivity and the contents of both reduced and oxidized gluta-thione were perceptible in high mercury-exhibiting shoots. Itis inferred that high mercury-accrued elevations in oxidativestress indices were obvious, where non-glutathione-basedH2O2-decomposing enzyme system was dominant over the
glutathione-based H2O2-scavenging enzyme system. In par-ticular, the glutathione-based H2O2-scavenging system failedto coordinate with elevated glutathione reductase which inturn resulted into increased pool of oxidized glutathione andthe ratio of oxidized glutathione-to-reduced glutathione. Thesubstantiation of the studied oxidative stress indices and anti-oxidant metabolism with approximately 53-kDa polypeptidewarrants further studies.
Salt marshes represent marine ecosystems and perform nu-merous essential ecological functions such as the stabilizationof coastal shorelines, preventing erosion and sediments fromwashing offshore; absorption and trapping of potential pollut-ants before they reach estuaries and fragile waterways; and thebiogeochemical cycling and storage of varied metal/metalloidcontaminants including mercury (Hg) (Pereira et al. 2009;Anjum et al. 2013a). Extensive studies have evidenced saltmarsh macrophyte-mediated considerable control ofsediment-associated Hg bioavailability to other biota throughdifferential accumulation and allocation in their organs (Weisand Weis 2004; Anjum et al. 2011, 2012a, 2013a, b). Theaccumulation of Hg by salt marsh macrophytes and its subse-quent allocation in their organs have been reported to becontrolled primarily by factors related to plants (species/ge-notypes, time/stage of exposure, growth stage metals sensitiv-ity), metal/metalloid (level, form and type) and sediments(physico-chemical traits including pH, redox potential,organic matter content) (Reboreda and Caçador 2007;Anjum et al. 2011, 2012a; Figueira et al. 2012). Among the
Responsible editor: Elena Maestri
N. A. Anjum :A. C. Duarte : E. Pereira : I. Ahmad (*)Department of Chemistry and CESAM-Centre for EnvironmentalandMarine Studies, University of Aveiro, 3810-193 Aveiro, Portugale-mail: [email protected]
I. AhmadDepartment of Biology and CESAM-Centre for Environmental andMarine Studies, University of Aveiro, 3810-193 Aveiro, Portugal
Environ Sci Pollut ResDOI 10.1007/s11356-014-2578-4
plant-related factors, plant capacity to restrict the accumulatedHg at root level in order to protect above-ground photosyn-thetic parts is of utmost significance. Despite the tight controlof plant roots on Hg translocation, a considerable amount ofHg reaches to above-ground plant parts (reviewed by Anjumet al. 2013a). In above-ground plant parts, the translocated Hgmay cause accelerated generation of reactive oxygen species(ROS; including H2O2) which in turn might trigger oxidativedamages to vital bio-molecules including nucleic acids andproteins (as reactive carbonyls, RCs); membrane lipid perox-idation (LPO), membrane integrity weakening, elevated elec-trolyte leakage (EL) and cell and plant death have been widelyreported (Patra and Sharma 2000; Anjum et al. 2012b, c,2013c). Because above-ground plant parts such as leavesperform photosynthesis to provide energy required for plantlife, it may be assumed that plant’s sustenance under Hgburden can be controlled by plant capacity to successfullymetabolize ROS (including H2O2) in leaves generated due toHg burden (Patra and Sharma 2000; Anjum et al. 2012b, c,2013c).
Plants are equipped with antioxidant defense system whereboth glutathione (GSH)-based (such as reduced GSH pool;glutathione peroxidase, GPX; glutathione sulfo-transferase,GST) and non-GSH-based (such as catalase, CAT; ascorbateperoxidase, APX) armory have a direct or indirect tight con-trol over ROS-accrued potential anomalies (Anjum et al.2012c). Another enzyme glutathione reductase (GR) main-tains the GSH pool and GSSG/GSH redox status necessary forboth GPX- and GST-mediated efficient ROS metabolism.Nonetheless, a fine synchronization between GSH- and non-GSH-based defense components and GR has been considerednecessary in order to keep a balance between ROS productionand metabolism (Gill and Tuteja 2010; Anjum et al. 2012a, b,2013c, Gill et al. 2013). To the other, accumulation of pro-line—a proteinogenic amino acid and osmolyte—has beenevidenced to protect plants against Hg-accrued osmotic stressand damages to cellular membrane, structures and proteins(Patra and Sharma 2000; Sharma and Dietz 2006). In addition,variation in leaf photosynthetic pigments (such as chlorophyll,carotenoids) has been reported to provide information on bothleaf- and plant-physiological state (Patra and Sharma 2000;Anjum et al. 2013c). Despite the above facts, exhaustiveliterature search depicted absence of reports highlighting theoxidative stress status and antioxidant metabolism in Juncusmaritimus Lam. shoots exhibiting differential Hg burdensunder environmental Hg exposure.
Given the above, the present study aimed (i) to determineshoot-Hg burden and analyze oxidative stress status, (ii) toinvestigate the modulation of GSH-based and non-GSH-based defense components and (iii) to cross-talk the responsesof oxidative stress traits and GSH and non-GSH-based de-fense endpoints with the modulation of shoot pigments andpolypeptide patterns.
Materials and methods
Study area
Ria de Aveiro is a coastal lagoon (45-km length; maximum10 km wide) located in the northern region of Portugal alongthe Atlantic Ocean (40′38′N, 8′44′W). Laranjo basin is a 1.5-km2 branch of this lagoon which received Hg-laden effluentsfrom a chlor-alkali plant located in a chemical complex indus-try nearby this lagoon from 1950 to 1994. Despite a cessationof Hg discharge since 1994, Hg concentration in the surfacesediments of this bay is still much higher than pre-industriallevels (Pereira et al. 2009). Laranjo basin has been regarded as“field laboratory” for assessing mercury toxicity under realis-tic environmental conditions since no other important sourcesof contaminants other than Hg have been reported in this area(Pereira et al. 2009).
Plant material
The salt marsh macrophyte J. maritimus, chosen as a modelplant system in the present study, is a C3 monocot plant andbelongs to family Juncaceae. J. maritimus is a tall, ratherslender rush, perennial and attains about 3 ft in height, witha sub-glaucous short, creeping rhizome. Leaves present nearthe base only, and stems are both circular in cross section, withsharp points. Stems are produced in clumps which can formdense stands. J. maritimuswidely occurs in salt marshes alongthe Atlantic coast of Europe and plays an important role in thefloristic coverage of the area. J. maritimus has been widelyinvestigated in context with its role for metal/metalloid reme-diation (reviewed by Anjum et al. 2013a).
Samplings
Along transects defined by the distance from the main Hgsource, samplings were performed considering five replicatesduring low tide in the months of October–November 2011 atsites depicting the highest (L1), moderate (L2) and the lowest(L3) Hg contamination. Additionally, samples were also col-lected from the Hg-free site (R) (Fig. 1). The proceduresadopted earlier by Anjum et al. (2011) were followed in thepresent study for the sample collection. In brief, plants weredug out carefully using a shovel, rinsed well with lagoonwater, placed into plastic bags and transported to the labora-tory maintaining the condition of 4 °C in large sample iceboxes. Once in the laboratory, plants were separated intoshoots and roots/rhizomes. Hot-air oven dried and powderedshoots were used for Hg estimation, whereas shoots wereeither used fresh for physiological/biochemical estimationsor dipped into liquid nitrogen and stored at −80 °C forfurther assays.
Environ Sci Pollut Res
Shoot-mercury determination
J. maritimus shoot-Hg concentration was determined byatomic absorption spectroscopy with thermal decompositionthrough an advanced Hg analyzer (LECO 254, USA) (Anjumet al. 2011).
Oxidative stress trait estimations
H2O2 content
Themethods of Loreto and Velikova (2001) and Dipierro et al.(2005) were adapted to determine shoot H2O2 content. Inbrief, fresh shoot tissues (0.25 g) were homogenized in 2 mlof 0.1 % (w/v) TCA. The homogenate was centrifuged at12,000×g for 15 min, and 0.5 ml of the supernatant wasmixed with 0.5 ml of 10 mM K-phosphate buffer pH 7.0and 1 ml of 1 M KI. H2O2 content of the supernatant wasevaluated by comparing its absorbance at 390 nm with astandard calibration curve.
Lipid peroxidation
As a status of membrane lipid peroxidation in fresh shoots, thecontent of thiobarbituric acid reactive substances (TBARS)was determined as per the method adopted and described byAnjum et al. (2013c). In brief, fresh shoot tissues (0.25 g) werehomogenized in liquid nitrogen, mixed with 0.73 % 2-thiobarbituric acid in 12 % trichloroacetic acid (TCA), incu-bated for 30 min in boiling water, ice-cooled, centrifuged at1,000×g for 10min at 4 °C, and the absorbance was measuredin the supernatant at 532 nm. The rate of lipid peroxidationwas expressed as nanomoles of TBARS formed per gram of
fresh weight, using a molar extinction coefficient of 1.55×105 M−1 cm−1.
Electrolyte leakage
Electrolyte leakage (EL) was assessed following the methodas described by Anjum et al. (2013c). In brief, fresh shoottissues (0.25 g) were placed in glass vials containing 10-mldeionized water. The vials were covered with plastic caps andplaced in a shaking incubator at the constant temperature of25 °C for 6 h, and the electrical conductivity (EC) of thesolution was measured (EC1) using an electrical conductivitymeter (WTW Cond 330i/SET, Weilheim, Germany). Subse-quently, the same vials were kept in water bath shaker at 90 °Cfor 2 h, cooled, and EC2 was measured. EL was calculatedfollowing the formula: EL=EC1/EC2×100.
Protein oxidation
Shoot-protein oxidation was estimated based on the reactionof carbonyls with 2,4-dinitrophenylhydrazine (DNPH)(Levine et al. 1994). In brief, fresh shoot tissues (0.25 g) werehomogenized in phosphate buffer (25 mM, pH 7.0) and cen-trifuged at 2,000×g. To 200 μl of supernatant, 300 μl of10 mMDNPH prepared in 2 M HCl was added and incubatedfor 1 h at room temperature. Proteins were precipitated with10% (w/v) TCA, and the pellets were washed three times with500 μl of ethanol/ethyl acetate (1/1, v/v) and subsequentlydissolved in 6 M guanidine hydrochloride in 20 mM K-phosphate buffer (pH 2.3). The reactive carbonyl contentwas calculated by absorbance at 370 nm, using the extinctioncoefficient for aliphatic hydrazones (22.1 mmol−1 cm−1). Allthe chemicals used were of analytical grade and were pur-chased from Sigma-Aldrich (USA).
Fig. 1 Location of samplingstations in the Laranjo basin, Riade Aveiro coastal lagoon,Portugal
Environ Sci Pollut Res
Proline content
Shoot proline content was estimated following the method ofBates et al. (1973). In brief, fresh shoot tissues (0.25 g) werehomogenized with 3 % aqueous sulfosalicylic acid and cen-trifuged at 3,000×g for 10 min. The supernatant was addedwith acid ninhydrin and glacial acetic acid and boiled in hotwater bath for 10 min. The reaction was stopped by keepingthe epperdorfs in an ice bath, and subsequently toluene wasadded to each tube and mixed vigorously on a cyclomixer for10–15 s in order to facilitate quick diffusion/movement ofchromophores from the aqueous phase to non-aqueous phase.The toluene layer (upper) was separated from the mixture, andabsorbance was read at 520 nm in the UV–vis plate reader(Spectra Max 384, Molecular Devices). The correspondingconcentration of proline was determined against the standardcurve processed in the same manner by using L-proline(Sigma).
Assays of non-GSH- and GSH-based H2O2-metabolizingsystem components
Sample preparation
Samples for the enzymatic antioxidants assays wereprepared following the method as adapted and describedby Anjum et al. (2013c). In brief, fresh shoot tissues(0.25 g) were homogenized in K-phosphate buffer(100 mM, pH 7.0; with 0.25 % v/v; Triton X-100;1 % w/v each of isopropanol, phenylmethylsulfonylfluoride and polyvinylpyrrolidone) and centrifuged at16,000×g for 20 min at 4 °C. The obtained supernatantwas used for enzymatic antioxidants (CAT, GPX, GSTand GR) assays, whereas the homogenizing buffer wassupplemented with 2 mM AsA for the assay of APXactivity.
Non-glutathione-based enzymes
The activity of APX was determined following the method ofNakano and Asada (1981). The reaction mixture contained0.5 mM ascorbate, 0.1 mMH2O2, 0.1 mMEDTA and enzymeextract. The reaction was allowed to run for 3 min at 25 °C.APX activity was calculated by using extinction coefficient of2.8 mM−1 cm−1. The activity of CATwas determined follow-ing the methods of Aebi (1984). The reaction mixturecontained buffer with 3 mM EDTA, enzyme extract and3 mM H2O2. The reaction was allowed to run for 3 min.Activity was calculated by using extinction coefficient (ε)0.036 mM−1 cm−1 and expressed as μmol of H2O2 g
−1 freshweight min−1.
Glutathione-based enzymes
The activity of GSH-metabolizing enzymes such as GPX (bymeasuring the oxidation of NADPH at 340 nm for 3 min usingH2O2 as substrate) and GST (by measuring increase in absor-bance at 340 nm for 3 min due to the formation of theconjugate 1-ch loro-2,4-d in i t robenzene (CDNB)(substratum)/GSH, catalyzed by GST) was assayed followingthe methods of Mohandas et al. (1984) and Drotar et al.(1985), as adapted and described by Anjum et al. (2013c).The activity of GR (GSH-regenerating enzyme) was deter-mined by monitoring the GSH-dependent oxidation ofNADPH (Foyer and Halliwell 1976). GR activity was calcu-lated by using extinction coefficient (e), 6.2 mM cm−1, andexpressed in enzyme unit mg−1 protein. One unit of enzymedetermines the amount necessary to decompose 1 mmol ofnicotinamide adenine dinucleotide phosphate (NADPH) permin at 25 °C.
Non-enzymatic antioxidant
Shoot non-enzymatic antioxidant glutathione (GSH; reduced,oxidized and total glutathione) was determined based on theproduction of 5-thio-2-nitrobenzoic acid as a result of GSHreacting with 5,5′-dithiobis(2-nitrobenzoic acid) reaction withthe sulfhydryl group of GSH following the method ofAnderson (1985).
Shoot pigment status
The contents of total chlorophyll and carotenoids in leaveswere estimated following the method of Alan (1994) asadapted and described by Anjum et al. (2013c). In brief, freshshoots (0.25 g) were extracted with 5-ml 80 % acetone andsubsequently centrifuged at 5,000×g for 10 min. Absorbanceof supernatant was measured at 480, 510, 663 and 645 nm inthe UV–vis plate reader. While, total chlorophyll content wascalculated using the equation: CT=20.2A645+8.02A663,whereas the equation CarT=7.6A480−1.49A510 was usedfor calculating total carotenoid content.
Shoot-polypeptide pattern analysis
Shoot protein was extracted by homogenizing shoot tissues(0.25 g) with phosphate buffer (25 mM, pH 7.0) using achilled mortar and pestle. The supernatant obtained after thecentrifugation of the homogenate at 15,000×g for 20 min at4 °C. The supernatant protein concentration was quantified bythe Coomassie Brilliant Blue G-250 assay (Bradford 1976). Inorder to characterize shoot protein, equal amounts (15 μg) ofproteins were denatured, loaded onto each lane and separatedby sodium dodecyl sulphate-polyacrylamide-gel electropho-resis (SDS-PAGE) (5 and 12.5 % acrylamide for stacking and
Environ Sci Pollut Res
resolving gels, respectively) at a constant current of 20 mAusing a vertical slab gel (BioRad Mini Protean II, Richmond,CA, USA) (Laemmli 1970). Separated polypeptides on the gelwere visualized by staining with Coomassie Brilliant BlueR-250 solution and subsequently detained with glacial aceticacid–methanol solution (1:4, v/v). The proteins fractionatedinto bands were scanned using the Molecular Imager Gel DocXR+System (Bio-Rad).
Data analysis
SPSS (PASW statistics 18) for Windows was used for statis-tical analysis. One-way analysis of variance (ANOVA)followed by pairwise multiple comparisons employing theTukey test was executed in order to detect significant dataamong treatments. The data are expressed as mean values±SD of three independent experiments with at least five repli-cates for each. The significance level was set at P≤0.05.
Results
Shoot-mercury burden
J. maritimus shoot displayed significantly increased shoot-Hgconcentration maximally at L1 (0.87 mg kg−1 dry weight)followed by L2 (0.41 mg kg−1 dry weight) and L3(0.2mg kg−1 dry weight) (vs. R), whereas comparisons amongcontaminated sites revealed significantly lower shoot Hg at L2
Table 1 Juncus maritimus shoot-mercury concentration (mg kg−1 dryweight) at reference (R) and sites with highest (L1), moderate (L2) andlowest (L3) mercury contamination in Laranjo basin salt marsh (Ria deAveiro), Portugal. Values are the means of five replicates±standarddeviation. Significant differences between sites (within the same organ):avs. R; bvs. L1; cvs. L3
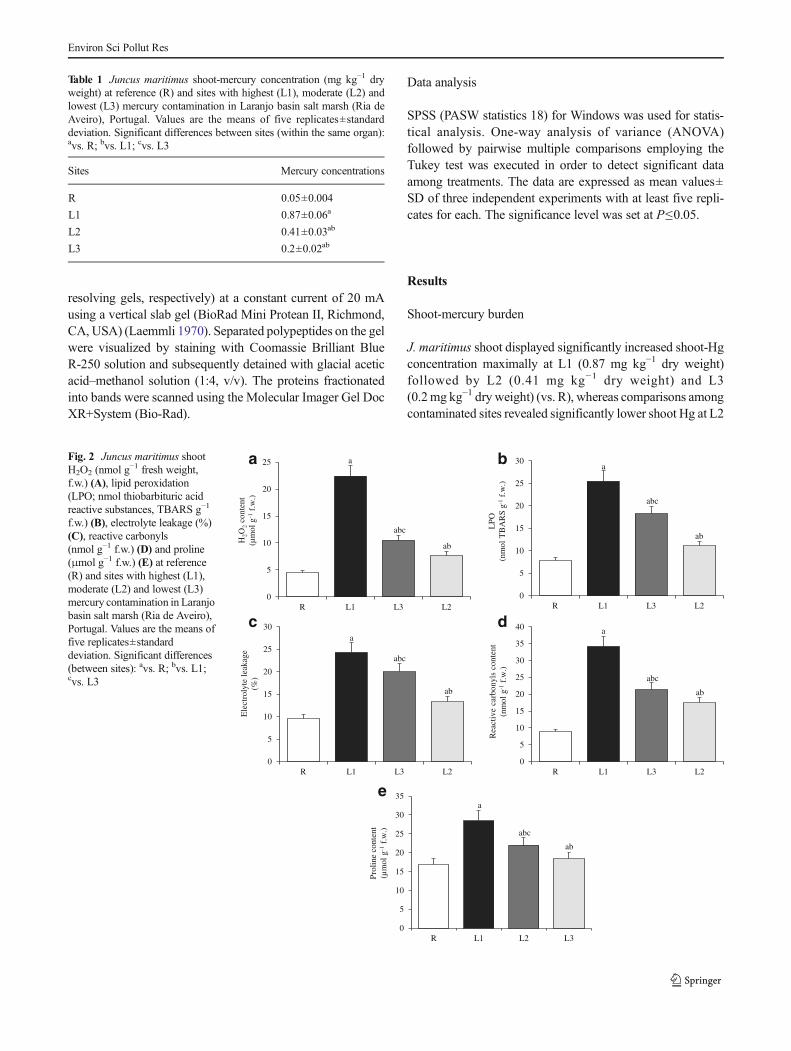
f.w.) (B), electrolyte leakage (%)(C), reactive carbonyls(nmol g−1 f.w.) (D) and proline(μmol g−1 f.w.) (E) at reference(R) and sites with highest (L1),moderate (L2) and lowest (L3)mercury contamination in Laranjobasin salt marsh (Ria de Aveiro),Portugal. Values are the means offive replicates±standarddeviation. Significant differences(between sites): avs. R; bvs. L1;cvs. L3
Environ Sci Pollut Res
(vs. L1) but significantly higher shoot Hg at the same sitewhen compared to L3 (Table 1).
Oxidative stress trait responses
Shoot-EL percent and the contents of H2O2, TBARS, RCs andproline displayed significant increases maximally at L1followed by L2 and L3 (vs. R). Contaminated site compari-sons revealed both significantly lower (vs. L1) and higher (vs.L3) extent of the studied damage indices at L2 (Fig. 2a–e).
Responses of non-GSH- and GSH-based H2O2-metabolizingsystem components
Significantly increased activity of APX, CAT and GR wasdepicted maximum at L1 followed by L2 and L3 (vs. R). Bothsignificantly lower and higher activity of APX, CAT and GRwere displayed at L2 vs. L1 and L3, respectively, whencomparisons among contaminated sites were considered(Fig. 3a–e). Regarding GPX and GST activity responses toHg contamination, significantly decreased GPX and GST
activity was perceptible maximally at L1 followed by L2and L3 (vs. R). Contaminated site comparisons revealed sig-nificantly increased (vs. L1) and decreased (vs. L3) GPX andGST activity at L2 (Fig. 3a–e). On the perspective of GSH,GSSG pools and their ratio (GSH/GSSG), the contents of bothGSH and GSSG displayed significant elevation maximally atL1 followed by L2 and L3 (vs. R). In contrast, GSH/GSSGratio depicted significant decreases maximally at L1 followedby L2 and L3 (vs. R). Comparisons of these traits at contam-inated sites revealed significantly lower contents of both GSHand GSSG at L2 (vs. L1), but at the same site, these traitsdisplayed significantly higher elevations when compared toL3. However, GSH/GSSG ratio comparisons depicted signif-icant increase (vs. L1) and decrease (L3) at L2 (Fig. 4a–c).
Shoot pigment status
Chlorophyll content significantly decreased maximally at L1(depicting highest Hg contamination) followed by L2(depicting moderate Hg contamination) and L3 (depictinglowest Hg contamination) (vs. R); whereas carotenoid content
0
1
2
3
4
5
6
7
R L1 L2 L3
AP
X a
ctiv
ity(n
mol
g- 1
f.w
. m-1
)
CA
T a
ctiv
ity(n
mol
g-1
f.w
. m-1
)
GP
X a
ctiv
ity(n
mol
g-1
f.w
. m- 1
)
GST
act
ivity
(nm
ol g
- 1f.
w. m
-1)
GR
act
ivity
(nm
ol g
-1f.
w. m
-1)
abc
ab
a
0
2
4
6
8
10
12
R L1 L2 L3
0
5
10
15
20
25
R L1 L2 L30
5
10
15
20
25
30
R L1 L2 L3
0
10
20
30
40
50
R L1 L2 L3
abc
ab
a
abab
a
abc
ab
a
abab
a
a b
c d
e
Fig. 3 Juncus maritimus shootactivities of ascorbate peroxidase(APX, nmol g−1 fresh weight,f.w. m−1) (A), catalase (CAT,nmol g−1 f.w. m−1) (B),glutathione peroxidase (GPX,nmol g−1 f.w. m−1) (C),glutathione sulfo-transferase(GST, nmol g−1 f.w. m−1) (D) andglutathione reductase (GR,nmol g−1 f.w. m−1) (E) atreference (R) and sites withhighest (L1), moderate (L2) andlowest (L3) mercurycontamination in Laranjo basinsalt marsh (Ria de Aveiro),Portugal. Values are the means offive replicates±standarddeviation. Significant differences(between sites): avs. R; bvs.L1;cvs. L3
Environ Sci Pollut Res
displayed significant increases maximally at L1 followed byL2 and L3 (vs. R). Comparisons among contaminated sitesrevealed significantly higher chlorophyll content at L3 follow-ed by L2 (vs. L1), while L2 displayed both significantly lowerand higher carotenoid content when compared to L1 and L3,respectively (Table 2).
Polypeptide pattern modulation
J. maritimus shoot SDS-PAGE protein band intensity visual-ization revealed differential responses. A new polypeptide ofapprox. 52 kDa with good intensity was depicted at L1 whichgradually decreased at L2 and L3 (vs. R). However, similarintensity of polypeptide of approx. 43 kDa was displayed at R,L1, L2 and L3 sites (Fig. 5).
Discussion
J. maritimus has been reported to perform the role of Hgphytostabilizer under environmental Hg exposure byrestricting the entry of Hg accumulated at root level; however,a portion of the root-accumulated Hg is transferred to above-ground parts (Anjum et al. 2011, 2012a, 2013b; Marques et al.2011; Figueira et al. 2012). In the present study, J. maritimusshoots exhibited 0.87-mg Hg kg−1 dry weight where, Hgburden altered photosynthetic pigments (such as chlorophyll,carotenoids) and caused a number of anomalies (EL, LPO,RCs) as a result of enhanced ROS generation-mediated oxi-dative stress. These observations are in concurrence with thereports of Patra and Sharma (2000) and Anjum et al. (2013c)where the authors have reported metal/metalloid (includingHg)-accrued alterations in photosynthetic pigments and sig-nificant elevations in several oxidative stress indices includingEL and LPO. As evidenced also herein, the cellular proteinswere earlier reported vulnerable to oxidative stress yieldingultimately to RCs (Davies 2005; Shulaev and Oliver 2006;Anjum et al. 2013c). In context with Hg impact on proline, itsaccumulation in shoots exhibiting the highest Hg burden canbe considered as a symptom of injury because partly of theabsence of the concrete physiological functions of the elevatedproline in metal/metalloid-exposed stressed plants (Sharmaand Dietz 2006; Wang et al. 2009; Szabados and Savoure2010). Additionally, the occurrence of enhanced oxidativestress measured in terms of H2O2, EL, TBARS and RCs inshoots with the same Hg burden also supports the elevatedshoot proline as a symptom of injury. In context with photo-synthetic pigment status, though Hg-burden-mediated
GSH
con
tent
(nm
ol g
-1f.
w.)
GSS
G c
onte
nt(n
mol
g-1
f.w
.)
GSH
/GSS
G r
atio
0
10
20
30
40
50
60
70
80
R L1 L2 L30
5
10
15
20
25
R L1 L2 L3
0.00
1.00
2.00
3.00
4.00
5.00
6.00
7.00
8.00
R L1 L2 L3
abc
ab
a
abc
ab
a
abc
ab
a
a b
c
Fig. 4 Juncus maritimus shootreduced (GSH) (A) and oxidized(GSSG) (B) glutathione contents(nmol g−1 fresh weight, f.w.), andGSSG/GSH ratio (C) at reference(R) and sites with highest (L1),moderate (L2) and lowest (L3)mercury contamination in Laranjobasin salt marsh (Ria de Aveiro),Portugal. Values are the means offive replicates±standard devia-tion. Significant differences (be-tween sites): avs. R; bvs. L1; cvs.L3
Table 2 Juncua maritimus shoot chlorophyll and carotenoid contents(μg g−1 fresh weight) at reference (R) and sites with highest (L1),moderate (L2) and lowest (L3) mercury contamination in Laranjo baysalt marsh (Ria de Aveiro), Portugal. Values are the means of fivereplicates±standard deviation. Significant differences between sites(within the same organ): avs. R, bvs. L1; cvs. L3
Sites Chlorophyll Carotenoids
R 5.3±0.4 4.3±0.3
L1 2.5±0.2a 7.3±0.6a
L2 3.4±0.3abc 5.5±0.4abc
L3 4.2±0.3b 4.6±0.4b
Environ Sci Pollut Res
decrease in chlorophyll level corroborates well with otherreports (Patra and Sharma 2000; Anjum et al. 2013c), thebehavior of the other pigment carotenoids under Hg burdenneeds elaboration. The significance of the elevated carotenoidslevel in J. maritimus shoots exhibiting the highest Hg burdencan be considered protective because carotenoids were reportedearlier as accessory light harvesting pigments and importantquenchers of the singlet state of chlorophyll and singlet oxygen(Saxena and Saiful-Arfeen 2009; Anjum et al. 2013c).
Plant capacity to employ a strong antioxidant defensesystem is one of their characteristics that help them to tolerate
metal/metalloid-accrued potential anomalies (Patra andSharma 2000; Gill and Tuteja 2010; Anjum et al. 2012b, c,2013c). In context with major antioxidant defense systemcomponents studied herein, the role of both GSH- and non-GSH-based ROS (H2O2)-scavenging systems in Hg-harboring J. maritimus shoots stood alone and was also com-parable. Considering GSH-based ROS (H2O2)-scavengingsystem, the activity of both GPX and GST was inhibited,while the activity of GR—a GSH-regenerating enzyme—and the contents of both reduced and oxidized GSH signifi-cantly enhanced. As a major component of the ascorbate(AsA)-GSH pathway, GSH reacts non-enzymatically withROS including H2O2 and gets itself converted into GSSG(Anjum et al. 2012b, c, 2013d; Gill et al. 2013). The observedincreases in GSH pool seem obvious as a result of diminishedactivity of GSH-utilizing enzymes—GPX and GST (Anjumet al. 2012b, c, 2013d). However, the observed increases inGSH oxidation (GSSG) and GSH/GSSG is strange. Thus,elevated GSSG pool and GSH/GSSG ratio reflect insufficien-cy of enhanced GR activity which thoughmaintained elevatedGSH pool failed to completely recycle GSSG back to thereduced GSH pool in order to maintain a high GSH/GSSGnecessary for operation of vital metabolic functions undernormal and stress conditions (Gill and Tuteja 2010; Anjumet al. 2012b, c, 2013c, d; Gill et al. 2013). In context with non-GSH-based H2O2-scavenging system, concomitant to increas-ing shoot-Hg burden, a parallel enhancement in H2O2-metab-olizing enzymes (APX and CAT) was evident. It is known thatboth CAT and APX directly determine the cellular concentra-tion of H2O2 (Asada 1999), where CAT scavenges H2O2
without any substrate, whereas APX requires AsA as a sub-strate during H2O2 metabolism (Gill and Tuteja 2010; Anjum
225
100
75
50
35
M R L1 L2 L3
Fig. 5 Representative gel depicting the Juncus maritimus shoot-polypeptide patterns modulation at reference (R) and sites with highest(L1), moderate (L2) and lowest (L3) mercury contamination in Laranjobasin salt marsh (Ria de Aveiro), Portugal. Arrows indicate significantchanges and extra polypeptides. The gel was stained with CoomassieBrilliant Blue (CBB)-R-250
Substantiation With ShootPolypeptide Expression Patterns
Non-GSH based H2O2-metabolizing
system(APX; CAT)
Com
plet
e C
ontr
ol
GSH based H2O2-metabolizing system
(GPX; GST)
GSH-regenerating system (GR)
Increased GSSG pool and
GSH/GSSG ratio
Partial C
ontrol
Protein oxidation
Lipid peroxidation
Electrolytes leakage
Proline
Mercury-Accrued Anomalies
Control of Photosynthetic Functions, Plant Growth and Development
Fig. 6 Schematic representationof the basic mechanismsunderlying Juncus maritimusshoot physiological/biochemicalresponses and their potentialrelationship with polypeptidepatterns under environmentalmercury exposure.(Abbreviations used: APX,ascorbate peroxidase; CAT,catalase; GPX, glutathioneperoxidase; GST, glutathionesulfo-transferase; GR, glutathionereductase; GSH, reducedglutathione; GSSG, oxidizedglutathione; GSH/GSSG, reducedglutathione-to-oxidizedglutathione ratio). See discussionfor details
Environ Sci Pollut Res
et al. 2012c, 2013c). Thus, the failure of GSH-based enzymes(GPX and GST) to counteract/metabolize Hg-accrued eleva-tions in H2O2 concentrations was compensated by significant-ly elevated activity of non-GSH-based enzymes (APX andCAT) to keepH2O2 level at par with their own activity. Similarresults were reported by Zhou et al. (2008) and Ortega-Villasante et al. (2007) where, APX- and CAT-mediated re-moval of excessive H2O2 was reported in Hg-treated plants.Thus, the dominancy of non-GSH (APX; CAT)-based H2O2-decomposing enzyme system was clearly perceptible overGSH (GPX, GST)-based H2O2-scavenging enzyme system(Fig. 6).
On the perspective of polypeptide pattern responses, earli-er, metal/metalloids have been reported to modulate polypep-tide patterns in different plant species which can easily bevisualized using SDS-PAGE—as a fundamental step towardproteomic studies, plant protein-SDS-PAGE has earlier beenshown to yield vital information significant for understandingthe molecular mechanisms of stress responses (Sobkowiakand Deckert 2006; Ahsan et al. 2007). In the present study,induction of protein synthesis was perceptible in J. maritimusshoots harboring the highest Hg, which increased its intensitymaximally at L1 followed by L2 and L3 sites. This resultcorroborates well with the reports of Gupta (1999),Sobkowiak and Deckert (2006) and Ahsan et al. (2007) wherethe authors reported protein synthesis induction in plantsunder varied metal/metalloids including Hg exposure. Thus,concomitant with the significantly elevated H2O2 metabolism,the 52-kDa polypeptide with increased intensity inJ. maritimus shoots exhibiting the highest Hg burden couldbe stress polypeptides—induced to overcome high Hgburden-accrued potential toxicities.
Conclusions
Cross-talks on the shoot Hg and pigment status and theresponses of both GSH- and non-GSH-based defense compo-nents and GR revealed (i) high Hg-mediated elevated damageindices as an obvious response, (ii) non-GSH (APX; CAT)-based H2O2-decomposing enzyme system dominated overGSH (GPX, GST)-based H2O2-scavenging enzyme system,(iii) GSH-based H2O2-metabolizing system failed to coordi-nate with elevated GR which resulted into increased GSSGpool and GSH/GSSG ratio and (iv) the polypeptide of approx.53 kDa can be a polypeptide of interest which substantiatedHg-led modulation of oxidative stress status and non-GSH-based H2O2-decomposing enzyme system.
Acknowledgments Authors gratefully acknowledge the financial sup-ports received from both FCT (Government of Portugal) through contract(SFRH/BPD/64690/2009; SFRH/BPD/84671/2012) and by the AveiroUniversity Research Institute/CESAM.
References
Aebi H (1984) Catalase in vitro. Method Enzymol 105:121–130Ahsan N, Lee SH, Lee DG, Lee H, Lee SW, Bahk JD, Lee BH (2007)
Physiological and protein profiles alternation of germinating riceseedlings exposed to acute cadmium toxicity. Comp Rend Biol 330:35–746
Alan RW (1994) The spectral determination of chlorophyll a and b, aswell as total carotenoids, using various solvents with spectropho-tometers of different resolution. Plant Physiol 144:307–313
Anderson ME (1985) Determination of glutathione and glutathionedisulfides in biological samples. Method Enzymol 113:548–570
Anjum NA, Ahmad I, Valega M, Pacheco M, Figueira E, Duarte AC,Pereira E (2011) Impact of seasonal fluctuations on the sediment-mercury, its accumulation and partitioning in Halimioneportulacoides and Juncus maritimus collected from Ria de AveiroCoastal Lagoon (Portugal). Water Air Soil Pollut 222:1–15
Anjum NA, Ahmad I, Válega M, Pacheco M, Figueira E, Duarte AC,Pereira E (2012a) Salt marsh macrophyte Phragmites australisstrategies assessment for its dominance in mercury-contaminatedcoastal lagoon (Ria de Aveiro, Portugal). Environ Sci Pollut Res 19:2879–2888
Anjum NA, Ahamd I, Mohmood I, Pacheco M, Duarte AC, Pereira E,Umar S, Ahmad A, Khan NA, Iqbal M, Prasad MNV (2012b)Modulation of glutathione and its related enzymes in plants’ re-sponses to toxic metals and metalloids—a review. Environ Exp Bot75:307–324
AnjumNA, Umar S, Ahmad A (2012c) Oxidative stress in plants: causes,consequences and tolerance. IK International Publishing House,New Delhi
Anjum NA, Ahmad I, Válega M, Mohmood I, Gill SS, Tuteja N, DuarteAC, Pereira E (2013a) Salt marsh halophyte services to metal-metalloid remediation: assessment of the processes and underlyingmechanisms. Crit Rev Environ Sci Technol. doi:10.1080/10643389.2013.828271
Anjum NA, Ahmad I, Válega M, Figueira E, Duarte AC, Pereira E(2013b) Phenological development stages variation versus mercurytolerance, accumulation, and allocation in salt marsh macrophytesTriglochin maritima and Scirpus maritimus prevalent in Ria deAveiro coastal lagoon (Portugal). Environ Sci Pollut Res 20:3910–3922
Anjum NA, Ahmad I, Rodrigues SM, Henriques B, Cruz N, Coelho C,Pacheco M, Duarte AC, Pereira E (2013c) Eriophorumangustifolium and Lolium perenne metabolic adaptations tometals- and metalloids-induced anomalies in the vicinity of a chem-ical industrial complex. Environ Sci Pollut Res 20:568–581
AnjumNA, Singh N, SinghMK, Shah ZA, Duarte AC, Pereira E, AhmadI (2013d) Single-bilayer graphene oxide sheet tolerance and gluta-thione redox system significance assessment in faba bean (Vicia fabaL.). J Nanopart Res 15:1770
Asada K (1999) The water-water cycle in chloroplasts: scavenging ofactive oxygen and dissipation of excess photons. Annu Rev PlantPhysiol Plant Mol Biol 50:601–639
Bates LS, Waldran RP, Teare ID (1973) Rapid determination of freeproline for water stress studies. Plant Soil 39:205–208
BradfordMM (1976) A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding. Anal Biochem 72:248–254
Davies MJ (2005) The oxidative environment and protein damage.Biochim Biophys Acta 1703:93–109
Dipierro N, Mondelli D, Paciolla C, Brunetti G, Dipierro S (2005)Changes in the ascorbate system in the response of pumpkin(Cucurbita pepo L.) roots to aluminium stress. J Plant Physiol162:529–536
Drotar A, Phelphs P, Fall R (1985) Evidence for glutathione peroxidaseactivities in cultured plant cells. Plant Sci 42:35–40
Figueira E, Freitas R, Pereira E, Duarte A (2012) Mercury uptake andallocation in Juncus maritimus: implications for phytoremediationand restoration of a mercury contaminated salt marsh. J EnvironMonit 14:2181–2188
Foyer CH, Halliwell B (1976) The presence of glutathione and glutathi-one reductase in chloroplasts: a proposed role in ascorbic acidmetabolism. Planta 133:21–25
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant ma-chinery in abiotic stress tolerance in crop plants. Plant PhysiolBiochem 48:909–930
Gill SS, Anjum NA, Hasanuzzaman M, Gill R, Trivedi DK, Ahmad I,Pereira E, Tuteja N (2013) Glutathione and glutathione reductase: aboon in disguise for plant abiotic stress defense operations. PlantPhysiol Biochem 70:204–212
Gupta M (1999) Effect of lead and mercury on changes in protein profilein the aquatic macrophytes Hydrilla verticillata (l.f.) royle andVallisneria spiralis L. J Environ Sci Health A Tox Hazard SubstEnviron Eng 34:1093–1104
Laemmli U (1970) Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature 227:680–685
Levine RL,Willians JA, Stadtman ER, Shacter E (1994) Carbonyl assaysfor determination of oxidatively modified proteins. MethodEnzymol 233:346–363
Loreto F, Velikova V (2001) Isoprene produced by leaves protects thephotosynthetic apparatus against ozone damage, quenches ozoneproducts, and reduces lipid peroxidation of cellular membranes.Plant Physiol 127:1781–1787
Marques B, Lillebø AI, Pereira E, Duarte AC (2011)Mercury cycling andsequestration in salt marshes sediments: an ecosystem service pro-vided by Juncus maritimus and Scirpus maritimus. Environ Pollut159:1869–1876
Mohandas J, Marshall JJ, Duggins GG, Horvath JS, Tiller D (1984)Differential distribution of glutathione and glutathione related en-zymes in rabbit kidney. Possible implications in analgesic neuropa-thy. Biochem Pharmacol 33:1801–1807
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged byascorbate-specific peroxidase in spinach chloroplasts. Plant CellPhysiol 22:867–880
Ortega-Villasante C, Herna′ndez LE, Rella′n-A′lvarez R, Del Campo FF,Carpena-Ruiz RO (2007) Rapid alteration of cellular redox homeo-stasis upon exposure to cadmium and mercury in alfalfa seedlings.New Phytol 176:96–107
Patra M, Sharma A (2000) Mercury toxicity in plants. Bot Rev 66:379–422
Pereira ME, Lillebø AI, Pato P, Válega M, Coelho JP et al (2009)Mercury pollution in Ria de Aveiro (Portugal): a review of thesystem assessment. Environ Monit Assess 155:39–49
Reboreda R, Caçador I (2007) Halophyte vegetation influences in saltmarsh retention capacity for heavy metals. Environ Pollut 146:147–154
Saxena DK, Saiful-Arfeen M (2009) Effect of Cu and Cd on oxidativeenzymes and chlorophyll content of moss Racomitrium crispulum.Taiwania 54:365–374
Sharma SS, Dietz KJ (2006) The significance of amino acids and aminoacid-derived molecules in plant responses and adaptation to heavymetal stress. J Exp Bot 57:711–726
Shulaev V, Oliver DJ (2006) Metabolic and proteomic markers foroxidative stress—new tools for reactive oxygen species research.Plant Physiol 141:367–372
Sobkowiak R, Deckert J (2006) Proteins induced by cadmium in soybeancells. J Plant Physiol 163:1203–1206
Szabados L, Savoure A (2010) Proline: a multifunctional amino acid.Trends Plant Sci 15:89–97
Wang F, Zeng B, Sun Z, Zhu C (2009) Relationship between proline andHg2+-induced oxidative stress in a tolerant rice mutant. ArchEnviron Contam Toxicol 56:723–731
Weis J, Weis P (2004) Metal uptake, transport and release by wetlandplants: implications for phytoremediation and restoration. EnvironInt 30:685–700
Zhou ZS,Wang SJ, Yang ZM (2008) Biological detection and analysis ofmercury toxicity to alfalfa (Medicago sativa) plants. Chemosphere70:1500–1509