Page 1
ORIGINAL PAPER
Patterning during somatic embryogenesis in Scots pine in relationto polar auxin transport and programmed cell death
Malin Abrahamsson • Silvia Valladares •
Emma Larsson • David Clapham • Sara von Arnold
Received: 23 September 2011 / Accepted: 19 December 2011 / Published online: 30 December 2011
� Springer Science+Business Media B.V. 2011
Abstract Somatic embryogenesis is a useful tool to
propagate conifers vegetatively. However, a major limita-
tion in many pine species is the low quality of cotyledonary
somatic embryos. The aim of this study has been to elu-
cidate the developmental pathway of somatic embryos in
Scots pine (Pinus sylvestris), to identify deviations from
the normal pathway and to identify processes that might
disturb normal development. Initially we compared the
developmental pathway of somatic embryogenesis in rep-
resentative cell lines yielding cotyledonary embryos with
normal and abnormal morphology. Early embryos carrying
suspensor cells in excess of the normal number (supernu-
merary) were more frequent in cell lines giving rise to
abnormal cotyledonary embryos. In this study we show that
the frequency of early somatic embryos with supernumer-
ary suspensor cells increased after treatment with the auxin
transport inhibitor 1-N-naphtylphthalamic acid (NPA).
Furthermore, the yield of developing embryos increased
significantly after treatment with the antiauxin 2-(4-chlo-
rophenoxy)-2-methylpropionic acid (PCIB), but the mor-
phology of the embryos was not affected. The number of
cells undergoing PCD was analyzed using a TUNEL-assay.
The frequency of TUNEL-positive cells was high both in
proliferating cultures and during differentiation of early
somatic embryos. However, the pattern of TUNEL-positive
cells was similar in normal somatic embryos and in
embryos with supernumerary suspensor cells. Together our
results suggest that the presence of supernumerary sus-
pensor cells in early somatic embryos of Scots pine is
caused by disturbed polar auxin transport and results in
aberrant embryo development.
Keywords Embryo patterning � NPA � PCIB � Pinus
sylvestris � Polar auxin transport � Programmed cell death �Scots pine � Somatic embryogenesis
Abbreviations
2,4-D 2,4-dichlorophenoxyacetic acid
ABA Abscisic acid
BA N6-benzyladenine
DCR Basal culture medium (Gupta and Durzan 1985)
DMSO Dimethyl sulfoxide
IAA Indoleacetic acid
NPA 1-N-napthylphthalamic acid
PAT Polar auxin transport
PCD Programmed cell death
PCIB 2-(4-chlorophenoxy)-2-methylpropionic acid
PEG Polyethylene glycol
PEM Proembryogenic mass
PGR Plant growth regulator (auxin and cytokinin)
TUNEL Terminal deoxynucleotidyl transferase (TdT)-
mediated dUTP nick end labelling
Malin Abrahamsson and Silvia Valladares contributed equally to this
work.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11240-011-0103-8) contains supplementarymaterial, which is available to authorized users.
M. Abrahamsson (&) � S. Valladares � E. Larsson �D. Clapham � S. von Arnold
Department of Plant Biology and Forest Genetics, Uppsala
BioCenter, Swedish University of Agricultural Sciences,
P.O. Box 7080, 750 07 Uppsala, Sweden
e-mail: [email protected]
Present Address:S. Valladares
Department of Plant Physiology, Instituto de Investigaciones
Agrobiologicas de Galicia (CSIC), Apartado 122,
15780 Santiago de Compostela, Spain
123
Plant Cell Tiss Organ Cult (2012) 109:391–400
DOI 10.1007/s11240-011-0103-8
Page 2
Introduction
Somatic embryogenesis is an attractive method to propa-
gate conifers vegetatively and is employed both as a tool in
the breeding program and as a method for large scale clonal
propagation. The whole process of regeneration of conifer
plants through somatic embryogenesis comprises a
sequence of steps: initiation and proliferation of embryo-
genic cultures, differentiation of early embryos and
development of late and cotyledonary embryos (von
Arnold and Clapham 2008). Many conifers belonging to
the genus spruce (Picea) can be propagated on a large scale
by somatic embryos (Klimaszewska et al. 2007). However,
for several species belonging to the genus Pinus the initi-
ation frequency of embryogenic cell lines is low and
regeneration of high quality cotyledonary embryos is poor
(Pullman et al. 2008; Bonga et al. 2010).
Development of multiple embryos from a single zygote
by monozygotic polyembryony is common among gym-
nosperms, including pines, but is rare in spruces (Singh
1978). During polyembryony the early embryo goes
through several rounds of cleavage resulting in many
equal-sized embryos. These embryos start to compete for
survival, and ultimately one embryo becomes dominant,
while the subordinate embryos are aborted (Nagmani et al.
1995; Filonova et al. 2002). In most pine species, initiation
of embryogenic cultures is limited to the first weeks of
zygotic embryo development during the cleavage stage
(Keinonen-Mettala et al. 1996; Haggman et al. 1999; Lelu-
Walter et al. 2008) or prior to the emergence of cotyle-
donary embryos (Becwar et al. 1990; Lelu et al. 1999). It
has been suggested that embryogenic cultures initiated
from immature zygotic embryos during the cleavage stage
result from a continuation of the cleavage process (Bozh-
kov et al. 1997; Park et al. 2006). In contrast, embryogenic
cultures of spruces are usually initiated from differentiated
cells in mature zygotic embryos, after these cells have been
stimulated to dedifferentiate (Mo et al. 1996).
In order to efficiently regulate the formation of plants
via somatic embryogenesis, we need to understand how
somatic embryos develop. Ideally, such knowledge should
be gained through the construction of a fate map repre-
senting an adequate number of morphological and molec-
ular markers specifying distinct developmental stages.
Pullman et al. (2003) developed a staging system in lob-
lolly pine (Pinus taeda) to evaluate the morphological
development in zygotic and somatic embryos. However,
our knowledge about how somatic embryos in pine species
develop is still insufficient to allow detailed descriptive
characterization. In contrast, somatic embryogenesis in
Norway spruce (Picea abies) has been thoroughly studied.
A time-lapse tracking technique was used to analyse the
developmental pathway of somatic embryos (Filonova
et al. 2000a). Based on this knowledge the whole process
can be synchronized and controlled (von Arnold and
Clapham 2008).
Auxin plays an essential role in plant development.
Critical to its activity as a developmental regulator is its
polar intercellular transport. Embryo patterning mutants in
Arabidopsis have been used for studying polarization
during early embryo development (Friml et al. 2003).
Auxin-regulated pattern formation has also been studied by
treating embryos with antiauxins such as 2-(4-chloro-
phenoxy)-2-methylpropionic acid (PCIB) or auxin trans-
port inhibitors such as 1-N-napthylphthalamic acid (NPA).
PCIB treatment of embryogenic cultures in different
coniferous species results in reduced proliferation and
increased quality of the mature embryos (Find et al. 2002;
Liao et al. 2008). NPA-treated somatic embryos of Norway
spruce attain an aberrant morphology as a consequence of
blocked polar auxin transport (PAT) (Larsson et al. 2008a).
PAT is crucial for appropriate apical-basal patterning
during early embryogeny in Norway spruce and polariza-
tion proceeds through the establishment of three major cell
types: the meristematic cells of the embryonal mass, the
embryonal tube cells and the terminally differentiated
suspensor cells. The suspensor cells in early somatic
embryos of Norway spruce are eliminated by programmed
cell death (PCD) (Filonova et al. 2000a, b).
The aim of this study was to elucidate the development
of somatic embryos in Scots pine, to identify deviations
from normal development and to identify processes that
might cause the abnormal development. We initially ana-
lyzed the developmental pattern of somatic embryos in cell
lines of Scots pine giving rise to cotyledonary embryos
with normal morphology, or to cotyledonary embryos with
abnormal morphology which do not develop into plants.
During differentiation of early embryos, the formation of
supernumerary suspensor cells, that is cells in excess of the
normal number, was more common in cell lines giving rise
to embryos with abnormal morphology. We treated cul-
tures with PCIB or NPA to examine the effects of reduced
auxin activity or inhibited polar transport on the occurrence
of supernumerary suspensor cells and aberrant embryo
development. Furthermore, the proportion of cells going
through PCD was analyzed using TUNEL-assay.
Materials and methods
Plant material
Embryogenic cell lines of Scots pine (Pinus sylvestris L.)
were initiated in 2000 from immature zygotic embryos
collected from open-pollinated trees growing in a seed
orchard in central Sweden (Burg et al. 2007). The cell lines
392 Plant Cell Tiss Organ Cult (2012) 109:391–400
123
Page 3
were stored in liquid nitrogen and thawed before starting
the experiments. Three embryogenic cell lines were used in
this study: cell line 12:12 which produces cotyledonary
embryos with normal morphology (Fig. 1a) but also some
with abnormal morphology (Fig. 1b) and cell lines 3:6 and
3:10 which give rise to abnormal cotyledonary embryos
(Fig. 1c, d).
DCR medium (Gupta and Durzan 1985) modified as
described previously (Burg et al. 2007) was used as basal
medium. The cultures were proliferated on DCR prolifer-
ation medium, supplemented with 2,4-dichlorophenoxy-
acetic acid (2,4-D) and N6-benzyladenine (BA) at 9.0 and
4.4 lM, respectively, as plant growth regulators (PGRs)
and 1% sucrose. To stimulate embryo maturation, the
cultures were first pretreated on PGR-free DCR medium,
pre-maturation medium, for 2–3 weeks and then trans-
ferred to DCR maturation medium, containing 7.5%
polyethylene glycol (PEG 4000), 60 lM abscisic acid
(ABA), 3% maltose and 0.35% gellan gum (Gelrite�,
Kelco) for 8–11 weeks. The cultures were incubated in
darkness at 22 ± 1�C and sub-cultured every second to
third week.
Histological preparations
Mature embryos from cell line 12:12 were collected for
sectioning after 10 weeks on maturation medium. The
embryos were fixed, embedded and serial-sectioned as
earlier described by Filonova et al. (2000a).
Cell tracking
Cell-tracking experiments were performed with cell line
3:6, 3:10 and 12:12 in thin layers of agarose (Filonova et al.
2000a). For analyzing the developmental pattern, about 50
aggregates per cell line were preselected at the start of the
maturation treatment. Their development was documented
every 3rd to 4th day, over 25 days. The development of
somatic embryos was separated into different stages based
on the developmental pathway of somatic embryos in cell
line 12:12 (Fig. 2).
Antiauxin treatment
Embryogenic cultures from cell lines 3:10 and 12:12 were
exposed to 2-(4-chlorophenoxy)-2-methylpropionic acid
(PCIB) (Sigma-Aldrich) during the pre-maturation treat-
ment. PCIB was dissolved in 95% (v/v) ethanol and added
to the medium after autoclaving at different concentrations
(1–20 lM). The strongest effect was observed at 10 lM
PCIB, while at higher concentrations the survival rate
decreased (data not shown). Therefore, we present data
only from cultures treated with 10 lM PCIB. After the
PCIB treatment the cultures were transferred to maturation
medium.
About 500 mg (f.w.) tissue was transferred to each Petri
dish (diameter 60 mm) and each PCIB treatment included
ten replicates. Differentiation of early somatic embryos
(Fig. 2d) was analyzed directly after 2 weeks on pre-mat-
uration medium. The samples were re-suspended in dH20
and poured into Petri dishes for microscopic examination.
The yield of early somatic embryos differentiated from 100
Fig. 1 Fully matured cotyledonary embryos. a Embryo with 6
developed cotyledons and elongated hypocotyl from cell line 12:12.
b Embryo with 3 developed cotyledons and elongated hypocotyl from
cell line 12:12. c Embryo with 6 developed cotyledons and aborted
hypocotyl from cell line 3:10. d Embryo with 5 developed cotyledons
and without distinct hypocotyl from cell line 3:6. (e and f) Longitu-
dinal sections of embryos from cell line 12.12. (e) Embryo with 5
developed cotyledons and shoot meristem. (f) Embryo with 2
cotyledons and aborted shoot meristem. co cotyledon, hc hypocotyl,
sm shoot meristem. Bars 250 lm
Plant Cell Tiss Organ Cult (2012) 109:391–400 393
123
Page 4
aggregates was determined in three replicates per treat-
ment. The yield and morphology of mature somatic
embryos (Fig. 2j) were recorded in all ten replicates after
8 weeks on maturation medium. The data were analyzed
for significance using ANOVA (analysis of variance,
P B 0.05). All data analysis in these studies was performed
using the SPSS� v. 17.0 statistical software package.
Inhibition of polar auxin transport
The effect of polar auxin transport (PAT) on the differen-
tiation of somatic embryos was assessed by exposing
embryogenic cultures of cell line 3:10 and 12:12 to the
auxin transport inhibitor 1-N-napthylphthalamic acid
(NPA) (Sigma-Aldrich). NPA was dissolved in dimethyl
sulfoxide (DMSO) to obtain a stock solution of 100 mM.
The cultures were treated with 20 and 30 lM NPA during
2 weeks on pre-maturation medium and 2–3 weeks on
maturation medium. The clearest effect was observed after
treatments with 30 lM NPA, therefore we present data
only from this treatment. Media lacking additives as well
as media supplemented with DMSO were used as controls.
Since no effect of DMSO was observed we have not
included data from DMSO controls.
The amount of tissue transferred to each Petri dish
(diameter 60 mm) at the start of the experiments was kept
constant at approximately 500 mg (f.w.). Each NPA
treatment included ten replicates. Differentiation of early
somatic embryos (Fig. 2d) was assessed after 2 weeks on
maturation medium. The samples were re-suspended in
dH20 and poured into Petri dishes for microscopic exami-
nation. The morphology of early somatic embryos differ-
entiated from 200 aggregates was determined in three
replicates per treatment. The yield and the morphology of
the mature somatic embryos (Fig. 2j) were recorded in all
ten replicates after 10–11 weeks on maturation medium.
The data were analyzed for significance using ANOVA
(analysis of variance, P \ 0.1 or P \ 0.05).
In situ detection of DNA fragmentation (TUNEL assay)
The number of cells going through PCD in embryogenic
cultures of Norway spruce has successfully been studied by
using terminal deoxynucleotidyl transferase (TdT)-mediated
Fig. 2 Development of somatic embryos of Scots pine in cell line
12:12. a Proliferating embryogenic cell aggregate in the presence of
plant growth regulators (PGRs) (stage 1a). b Slightly globular
structure with densely packed cells covered by a smooth surface
2–3 weeks after withdrawal of PGRs (stage 1b). c Early embryo after
1–2 weeks exposure to abscisic acid (ABA) (stage 1c). d Early
somatic embryo after 1–3 weeks exposure to ABA (stage 2). e Late
embryo after 2–5 weeks exposure to ABA (stage 3). f Late embryo
before cotyledon differentiation after 5–6 weeks exposure to ABA
(stage 4). g–i Maturing embryos with developing cotyledonary
primordia after 6–8 weeks exposure to ABA (stages 5–7). j Fully
matured cotyledonary embryo after 10 weeks exposure to ABA (stage
8). co cotyledon, em embryonal mass, hc hypocotyl, mc meristematic
cells, s suspensor, vc vacuolated cells. Bars, 100 lm (a–e); 250 lm
(f–j)
394 Plant Cell Tiss Organ Cult (2012) 109:391–400
123
Page 5
dUTP nick end labelling (TUNEL), which is an in situ
method to label fragmented DNA (Filonova et al. 2000b).
Samples for in situ detection of DNA fragmentation were
collected from cell line 3:10 and 12:12: after 2 weeks on
proliferation medium, after 2 weeks on pre-maturation
medium and after 2 weeks on maturation medium. Nuclear
DNA fragmentation was assessed by whole mount TUNEL
(In situ Cell Death Detection Kit, Roche, Penzberg,
Germany) as described earlier by Larsson et al. (2008a). The
frequency of TUNEL-positive cells was based on the total
number of DAPI-stained nuclei in each structure. In total, 15
randomly chosen structures from each sample were ana-
lyzed. The data were analyzed for significance using t test,
P B 0.05.
In order to assess if PCD is suppressed in embryos
carrying supernumerary suspensor cells, TUNEL assays
were performed on somatic embryos at stage 1c, 2, 3 and 4
(Fig. 2c, d, e, f) collected after 2 and 4 weeks on matura-
tion medium. In total 52 embryos from cell line 3:10 and
cell line 12:12 were analyzed.
Results and discussion
Time-lapse tracking
Embryogenic cell lines of Scots pine vary significantly in
their ability to differentiate cotyledonary embryos (Burg
et al. 2007). Furthermore, the morphology of the cotyle-
donary embryos differs among cell lines. Initially we
analyzed the developmental pathway of somatic embryo-
genesis in eight cell lines, of which three representative cell
lines were chosen for more detailed analysis: cell line
12:12 which produces predominantly cotyledonary
embryos with normal morphology (Fig. 1a) and cell lines
3:6 and 3:10 which give rise to abnormal cotyledonary
embryos (Fig. 1c, d).
The development of somatic embryos was separated into
different stages (stage 1–8, Fig. 2). Proliferating embryo-
genic cultures in cell line 12:12 consisted of a mixture of
small meristematic cells and elongated vacuolated cells,
resembling the proembryogenic masses (PEMs) in Norway
spruce (Filonova et al. 2000a). Most of the cells were
clustered into large aggregates, stage 1a (Fig. 2a). At the
periphery of the large aggregates some regions ‘budded
off’, giving rise to new separate aggregates which
increased in size. After withdrawal of PGRs the cultures
continued to proliferate by forming new aggregates at the
same time as parts of the clustered aggregates became more
globular. These globular regions consisted of densely
packed cells covered by a smooth surface, stage 1b
(Fig. 2b). The cultures continued to proliferate after
transfer to maturation medium simultaneously as new
globular structures in the clustered aggregates were formed
and early somatic embryos started to differentiate, stage 1c
(Fig. 2c). Subsequently, early (stage 2, Fig. 2d) and late
(stage 3, Fig. 2e) somatic embryos composed of an
embryonal mass and a suspensor developed. The suspensor
cells started to degrade during stage 4 (Fig. 2f) and were
eliminated around stage 7 and 8 (Fig. 2i, j). The hypocotyl
started to elongate at stage 4 and the differentiation of the
cotyledons along with further elongation of the hypocotyl
took place at stage 5 (Fig. 2g). The number of cotyledons
varied from two to six and about 50% of the somatic
embryos carried four or more cotyledons and most of them
had a shoot meristem (Fig. 1e). Somatic embryos with two
cotyledons lacked a shoot meristem (Fig. 1f).
In cell line 3:6 and 3:10 the somatic embryos usually
started to differentiate after 1 week, and a high proportion
of both early and late somatic embryos carried supernu-
merary suspensor cells, resulting in an unbalanced ratio
between the embryonal mass and the suspensor (Fig. 4c).
Furthermore, the cotyledons started to differentiate already
by stage 4 (Fig. 2f). The number of cotyledons varied from
two to six and about one-third of the embryos carried four
or more cotyledons. The somatic embryos showed radial
growth during the whole process of maturation, resulting in
a high frequency of embryos with aborted or abnormal
hypocotyl (Fig. 1c, d).
There were no fundamental differences between the cell
lines in the developmental pathway leading to cotyledonary
embryos. However, already at the stage of early embry-
ogeny a significantly higher frequency of the somatic
embryos in cell line 3:6 and 3:10 carried supernumerary
suspensor cells compared to in cell line 12:12. About 35%
of the early somatic embryos in cell line 3:10 and only 8%
of the embryos in cell line 12:12 carried supernumerary
suspensor cells. The developmental programs of the
embryo proper and the suspensor are closely coordinated,
and imbalance causes embryo defects and lethality
(Smertenko et al. 2003; Bozhkov et al. 2005).
Decreased endogenous auxin activity stimulates
differentiation of early somatic embryos
In several plant species it has been shown that the endog-
enous indoleacetic acid (IAA) level must be kept low to
ensure normal somatic embryo formation (Korlach and
Zoglaur 1995; Find et al. 2002). It has been proposed that
the auxin antagonist PCIB reduces the activity of endoge-
nous IAA by competitive binding to auxin receptors
(McRae and Bonner 1953). In order to determine whether a
decreased auxin activity could reduce the proliferation of
embryogenic tissue of Scots pine during the pre-maturation
and maturation treatments and promote normal embryo
development, embryogenic cultures of cell line 3:10 and
Plant Cell Tiss Organ Cult (2012) 109:391–400 395
123
Page 6
12:12 were treated with 10 lM PCIB during pre-matura-
tion. In both cell lines the treatment reduced proliferation
of embryogenic cultures on pre-maturation and maturation
media. However, we could not observe any changes in the
morphology of early somatic embryos after treatment with
PCIB. The yield of cotyledonary embryos was significantly
increased after the PCIB treatment (Fig. 3). In both control
cotyledonary somatic embryos and in PCIB-treated
embryos about 33% of the cotyledonary embryos in cell
line 3:10 carried four or more cotyledons, and in cell line
12:12 about 50% of the cotyledonary embryos carried four
or more cotyledons. Therefore, in contrast to what has been
reported previously for other conifers (Find et al. 2002;
Liao et al. 2008), reduced auxin activity does not improve
the morphology either in developing embryos or in coty-
ledonary embryos of Scots pine. This indicates that the
formation of supernumerary suspensor cells is not caused
by too high endogenous auxin content. We cannot, how-
ever, exclude that auxin homeostasis is disturbed in
embryogenic cultures of Scots pine, since the yield of
developing embryos increased.
Inhibition of PAT results in abnormal embryo
morphology
NPA-treatment during development of somatic embryos in
Norway spruce led to an increased endogenous IAA con-
tent, abnormal cell divisions and decreased PCD, resulting
in a changed balance between the embryonal mass and the
suspensor (Larsson et al. 2008a). In order to test if dis-
turbed PAT can be one of the reasons for the formation of
supernumerary suspensor cells in Scots pine somatic
embryos, we analyzed how embryo morphology was
affected when the cultures were treated with an auxin
transport inhibitor.
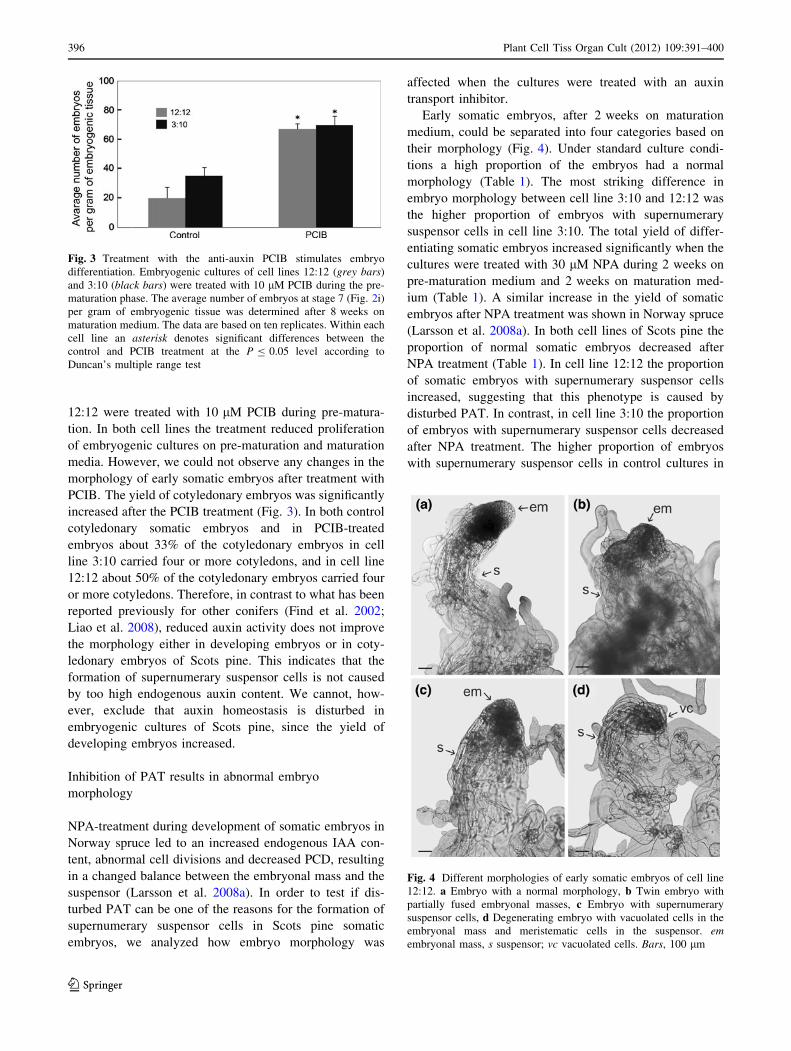
Early somatic embryos, after 2 weeks on maturation
medium, could be separated into four categories based on
their morphology (Fig. 4). Under standard culture condi-
tions a high proportion of the embryos had a normal
morphology (Table 1). The most striking difference in
embryo morphology between cell line 3:10 and 12:12 was
the higher proportion of embryos with supernumerary
suspensor cells in cell line 3:10. The total yield of differ-
entiating somatic embryos increased significantly when the
cultures were treated with 30 lM NPA during 2 weeks on
pre-maturation medium and 2 weeks on maturation med-
ium (Table 1). A similar increase in the yield of somatic
embryos after NPA treatment was shown in Norway spruce
(Larsson et al. 2008a). In both cell lines of Scots pine the
proportion of normal somatic embryos decreased after
NPA treatment (Table 1). In cell line 12:12 the proportion
of somatic embryos with supernumerary suspensor cells
increased, suggesting that this phenotype is caused by
disturbed PAT. In contrast, in cell line 3:10 the proportion
of embryos with supernumerary suspensor cells decreased
after NPA treatment. The higher proportion of embryos
with supernumerary suspensor cells in control cultures in
Fig. 3 Treatment with the anti-auxin PCIB stimulates embryo
differentiation. Embryogenic cultures of cell lines 12:12 (grey bars)
and 3:10 (black bars) were treated with 10 lM PCIB during the pre-
maturation phase. The average number of embryos at stage 7 (Fig. 2i)
per gram of embryogenic tissue was determined after 8 weeks on
maturation medium. The data are based on ten replicates. Within each
cell line an asterisk denotes significant differences between the
control and PCIB treatment at the P B 0.05 level according to
Duncan’s multiple range test
Fig. 4 Different morphologies of early somatic embryos of cell line
12:12. a Embryo with a normal morphology, b Twin embryo with
partially fused embryonal masses, c Embryo with supernumerary
suspensor cells, d Degenerating embryo with vacuolated cells in the
embryonal mass and meristematic cells in the suspensor. emembryonal mass, s suspensor; vc vacuolated cells. Bars, 100 lm
396 Plant Cell Tiss Organ Cult (2012) 109:391–400
123
Page 7
cell line 3:10 than in cell line 12:12 indicates that PAT is
less efficient in somatic embryos in cell line 3:10 and that
further reduction of PAT by treatment with NPA leads to
embryo degeneration. We suggest that the presence of
supernumerary suspensor cells in early somatic embryos of
Scots pine is caused by disturbed PAT. In accordance, in
Norway spruce embryos, blocked PAT during differentia-
tion of the suspensor stimulates cell division of the meri-
stematic cells adjacent to the tube cells and this causes the
formation of supernumerary suspensor cells (Larsson et al.
2008b).
In both cell lines the proportion of somatic embryos with
separated cotyledons decreased and embryos carrying
partially or severely fused cotyledons increased after
treatment with NPA (Supplementary Fig. 1). The propor-
tion of embryos with partially and severely fused cotyle-
dons after NPA treatment was higher in cell line 3:10, but,
owing to a high variation among replicates, the differences
between the cell lines were not significant. The aberrant
morphologies of cotyledonary somatic embryos after
treatment with NPA are comparable with what has previ-
ously been shown in Norway spruce (Larsson et al. 2008a;
Hakman et al. 2009). In Norway spruce, normal cotyle-
donary somatic embryos were formed when the cultures
were exposed to NPA only during the earlier stages, while
embryos treated with NPA during development of late
embryos displayed apical abnormalities including fused or
aborted cotyledons (Larsson et al. 2008a). Based on the
fact that the proportion of Scots pine control embryos with
severely fused cotyledons was low we assume that PAT has
been activated during development of late embryos.
Suspensor cells are degraded by PCD
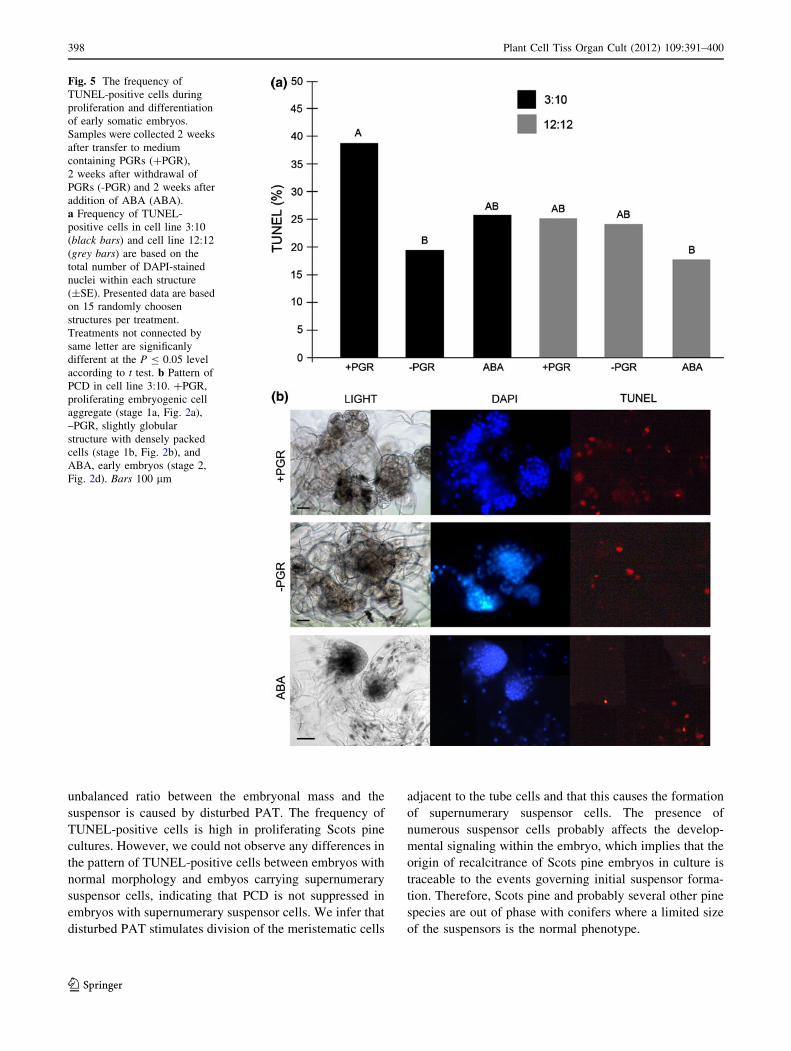
The frequency of TUNEL-positive cells is much higher in
proliferating embryogenic cultures of Scots pine than in
Norway spruce, 25–39% (Fig. 5a) and about 5% (Helmersson
et al. 2008), respectively. Assuming that embryogenic cul-
tures of Scots pine proliferate by a continuous cleavage-like
process, as has been suggested for other pine species
(Bozhkov et al. 1997), it is reasonable to assume that the
mechanism responsible for eliminating subordinate embryos
by PCD (Filonova et al. 2002) is retained in embryogenic
cultures, resulting in a much higher level of PCD in Scots pine
than in Norway spruce.
In cell line 3:10 the frequency of TUNEL-positive cells
decreased significantly during differentiation of early
somatic embryos (Fig. 5), while no significant decrease
was observed in cell line 12:12. An increase in PCD is
crucial during differentiation of early somatic embryos of
Norway spruce (Bozhkov et al. 2002; Helmersson et al.
2004). We assume that the lack of increased PCD during
differentiation of early somatic embryos in Scots pine is a
consequence of the high level of PCD during the prolif-
eration phase.
In order to assess if PCD is suppressed in Scots pine
somatic embryos carrying supernumerary suspensor cells,
we compared the frequency of TUNEL-positive cells in
embryos with normal morphology with that in embyos
carrying supernumerary suspensor cells in both cell line
3:10 and 12:12. We could not observe any clear differences
in the pattern of TUNEL-positive cells either between cell
lines or between normal embryos and embryos carrying
supernumerary suspensor cells (Fig. 6). This suggests that
the unbalanced ratio between the embryonal mass and the
suspensor is due to the formation of extra suspensor cells
rather than suppressed PCD.
Conclusion
The frequency of early somatic embryos with supernu-
merary suspensor cells is higher in Scots pine cell lines
producing cotyledonary embryos with an abnormal
morphology.
The proportion of Scots pine somatic embryos with
supernumerary suspensor cells increases after treatment
with the PAT inhibitor NPA, suggesting that the
Table 1 Effect of NPA treatment on the morphology of early somatic embryos
Cell line Treatment Total number of embryos Embryo morphology (%)
a b c d
12:12 Control 107 58 ± 2.7 2 ± 1.5 10 ± 4.6 31 ± 2.5
NPA 210** 28 ± 1.5** 9 ± 0.8** 21 ± 1.4* 42 ± 2.2*
3:10 Control 173 43 ± 3.8 6 ± 2.4 28 ± 0.9 24 ± 5.9
NPA 366** 29 ± 2.7** 6 ± 1.9 16 ± 3.5** 50 ± 2.3**
Embryogenic cultures of cell lines 12:12 and 3:10 were treated with 30 lM NPA during 2 weeks on pre-maturation medium and 2 weeks on
maturation medium. Presented data are based on scoring 200 aggregates in three independent experiments. The embryo morphology after
2 weeks on maturation was classified as type a, b, c or d as shown in Fig. 4. The frequency of embryos with various morphologies are based on
the total number of embryos ±SE. Within each cell line * denotes significant difference at the P \ 0.1 and ** denotes significant difference at
the P \ 0.05 between control and NPA treatment
Plant Cell Tiss Organ Cult (2012) 109:391–400 397
123
Page 8
unbalanced ratio between the embryonal mass and the
suspensor is caused by disturbed PAT. The frequency of
TUNEL-positive cells is high in proliferating Scots pine
cultures. However, we could not observe any differences in
the pattern of TUNEL-positive cells between embryos with
normal morphology and embyos carrying supernumerary
suspensor cells, indicating that PCD is not suppressed in
embryos with supernumerary suspensor cells. We infer that
disturbed PAT stimulates division of the meristematic cells
adjacent to the tube cells and that this causes the formation
of supernumerary suspensor cells. The presence of
numerous suspensor cells probably affects the develop-
mental signaling within the embryo, which implies that the
origin of recalcitrance of Scots pine embryos in culture is
traceable to the events governing initial suspensor forma-
tion. Therefore, Scots pine and probably several other pine
species are out of phase with conifers where a limited size
of the suspensors is the normal phenotype.
Fig. 5 The frequency of
TUNEL-positive cells during
proliferation and differentiation
of early somatic embryos.
Samples were collected 2 weeks
after transfer to medium
containing PGRs (?PGR),
2 weeks after withdrawal of
PGRs (-PGR) and 2 weeks after
addition of ABA (ABA).
a Frequency of TUNEL-
positive cells in cell line 3:10
(black bars) and cell line 12:12
(grey bars) are based on the
total number of DAPI-stained
nuclei within each structure
(±SE). Presented data are based
on 15 randomly choosen
structures per treatment.
Treatments not connected by
same letter are significanly
different at the P B 0.05 level
according to t test. b Pattern of
PCD in cell line 3:10. ?PGR,
proliferating embryogenic cell
aggregate (stage 1a, Fig. 2a),
–PGR, slightly globular
structure with densely packed
cells (stage 1b, Fig. 2b), and
ABA, early embryos (stage 2,
Fig. 2d). Bars 100 lm
398 Plant Cell Tiss Organ Cult (2012) 109:391–400
123
Page 9
Acknowledgments This work was supported by the Swedish
Research Council for Environment, Agricultural Sciences and Spatial
Planning. Silvia Valladares was supported by an Angeles Alvarino
postdoctoral fellowship from Xunta de Galicia (Spain).
References
Becwar MR, Nagmani R, Wann SR (1990) Initiation of embryogenic
cultures and somatic embryo development in loblolly pine
(Pinus taeda). Can J For Res 20:810–817
Bonga JM, Klimaszewska KK, von Aderkas P (2010) Recalcitrance in
clonal propagation, in particular of conifers. Plant Cell Tissue
Organ Cult 100:241–254
Bozhkov PV, Ahn IS, Park YG (1997) Two alternative pathways of
somatic embryo origin from polyembryonic mature stored seeds
of Pinus koraiensis. Can J Bot 75:509–512
Bozhkov P, Filonova L, von Arnold S (2002) A key developmental
switch during Norway spruce somatic embryogenesis is induced
by withdrawal of growth regulators and is associated with cell
death and extracellular acidification. Biotechnol Bioeng 77:
658–667
Bozhkov PV, Suarez MF, Filonova LH, Daniel G, Zamyatnin AA,
Rodriguez-Nieto S, Zhivotovsky B, Smertenko A (2005) Cysteine
protease mcII-Pa executes programmed cell death during plant
embryogenesis. Proc Natl Acad Sci USA 102:14463–14468
Burg K, Helmersson A, Bozhkov P, von Arnold S (2007) Develop-
mental and genetic variation in nuclear microsatellite stability
during somatic embryogenesis in pine. J Exp Bot 58:687–698
Filonova LH, Bozhkov PV, von Arnold S (2000a) Developmental
pathway of somatic embryogenesis in Picea abies as revealed by
time-lapse tracking. J Exp Bot 51:249–264
Filonova LH, Bozhkov PV, Brukhin VB, Daniel G, Zhivotovsky B,
von Arnold S (2000b) Two waves of programmed cell death
occur during formation and development of somatic embryo in
the gymnosperms, Norway spruce. J Cell Sci 113:4399–4411
Filonova LH, von Arnold S, Daniel G, Bozhkov PV (2002)
Programmed cell death eliminates all but one embryo in a
polyembryonic plant seed. Cell Death Differ 9:1057–1062
Find J, Grace L, Krogstrup P (2002) Effect of anti-auxins on
maturation of embryogenic tissue cultures of Nordmanns fir
(Abies nordmanniana). Physiol Plant 116:231–237
Friml J, Benkova E, Mayer U, Palme K, Muster G (2003) Automated
whole mount localization techniques for plant seedlings. Plant J
34:115–124
Gupta PK, Durzan DJ (1985) Shoot multiplication from mature trees
of Douglas-fir (Pseudotsuga manziesii) and sugar pine (Pinuslambertiana). Plant Cell Rep 4:177–179
Haggman H, Jokela A, Krajnakova J, Kauppi A, Niemi K, Aronen T
(1999) Somatic embryogenesis of Scots pine: cold treatment and
characteristics of explants affecting induction. J Exp Bot 341:
1769–1778
Hakman I, Hallberg H, Palovaara J (2009) The polar auxin transport
inhibitor NPA impairs embryo morphology and increases the
expression of an auxin efflux facilitator protein PIN during Piceaabies somatic embryo development. Tree Physiol 29:483–496
Helmersson A, von Arnold S, Kornel B, Bozhkov P (2004) High
stability of nuclear microsatellite loci during the early stages of
somatic embryogenesis in Norway spruce. Tree Physiol 24:
1181–1186
Helmersson A, Jansson G, Bozhkov PV, von Arnold S (2008) Genetic
variation in microsatellite stability of somatic embryo plants of
Picea abies: a case study using six unrelated full-sib families.
Scand J For Res 23:2–11
Keinonen-Mettala K, Jalonen P, Eurola P, von Arnold S, von
Weissenberg K (1996) Somatic embryogenesis of Pinus sylves-tris. Scand J For Res 11:242–250
Klimaszewska K, Trontin JF, Becwar MR, Devillard C, Park YS,
Lelu-Walter MA (2007) Recent progress on somatic embryo-
genesis of four Pinus ssp. Tree For Sci Biotechnol 1:11–25
Korlach J, Zoglaur K (1995) Developmental patterns during direct
somatic embryogenesis in protoplast cultures of european larch
(Larix decidua Mill.). Plant Cell Rep 15(3–4):242–247
Larsson E, Sitbon F, Ljung K, von Arnold S (2008a) Inhibited polar
auxin transport results in aberrant embryo development in
Norway spruce. New Phytol 177:356–366
Larsson E, Sitbon F, von Arnold S (2008b) Polar auxin transport
controls suspensor fate. Plant Signal behav 3(7):469–470
Lelu MA, Bastien C, Drugeault A, Gouez ML, Klimaszewska K
(1999) Somatic embryogenesis and plantlet development in
Pinus sylvestris and Pinus pinaster on medium with and without
plant growth regulators. Physiol Plant 105:719–728
Lelu-Walter MA, Bernier-Cardou M, Klimaszewska K (2008) Clonal
plant production from self- and cross-pollinated seed families of
Pinus sylvestris (L.) through somatic embryogenesis. Plant Cell
Tiss Organ Cult 92:31–45
Liao YK, Liao CK, Ho YL (2008) Maturation of somatic embryos in
two embryogenic cultures of Picea morrisonicola Hayata as
affected by alteration of endogenous IAA content. Plant Cell
Tiss Organ Cult 93:257–268
McRae DH, Bonner J (1953) Chemical structure and antiauxin
activity. Physiol Plant 6:485–510
Mo LH, Egertsdotter U, von Arnold S (1996) Secretion of specific
extracellular proteins by somatic embryos of Picea abies is
dependent on embryo morphology. Ann Bot 77:143–152
Nagmani R, Diner AM, Garton S, Zipf AE (1995) Anatomical
comparison of somatic and zygotic embryos in conifers. In: Jain
S, Gupta P, Newton R (eds) Somatic embryogenesis in woody
plants. Kluwer Academic Publishers, Dordrecht, pp 23–48
Fig. 6 Pattern of PCD in somatic embryos of cell line 3:10. Whole
mounts of somatic embryos with different morphology were analyzed
by TUNEL. a Embryo with normal morphology at stage 3 (Fig. 2e).
b Abnormal embryo with supernumerary suspensor cells at stage 3/4
(Fig. 2e and f). em, embryonal mass; s, suspensor. Bars 100 lm
Plant Cell Tiss Organ Cult (2012) 109:391–400 399
123
Page 10
Park YS, Lelu-Walter MA, Harvengt L, Trontin JF, MacEacheron I,
Klimaszewska K, Bonga JM (2006) Initiation of somatic
embryogenesis in Pinus banksiana, P. strobus, P. pinaster, and
P. sylvestris at three laboratories in Canada and France. Plant
Cell Tiss Organ Cult 86:87–101
Pullman GS, Buchanan M (2008) Identification and quantitative
analysis of stage-specific sugars and carbohydrates in loblolly
pine (Pinus taeda L.) zygotic embryo and female gametophyte.
Tree Physiol 28:985–996
Pullman G, Johnson S, Peter G, Cairney J, Xu N (2003) Improving
loblolly pine somatic embryo maturation: comparison of somatic
and zygotic embryo morphology, germination, and gene expres-
sion. Plant Cell Rep 21:747–758
Singh H (1978) Embryology of Gymnosperms. In: Zimmermann W,
Carlquist Z, Ozenda P, Wulff HD (eds) Handbuch der Pflan-
zenanatomie. Gebruder Borntrager, Berlin, pp 187–241
Smertenko AP, Bozhkov PV, Filonova LH, von Arnold S, Hussey PJ
(2003) Reorganization of the cytoskeleton during developmental
programmed cell death in Picea abies embryos. Plant J 33:813–824
von Arnold S, Clapham D (2008) Spruce embryogenesis. In: Suarez
MF, Bozhkov PV (eds) Plant embryogenesis: methods in
molecular biology, Human Press, Totowa, NJ, 427:31–47
400 Plant Cell Tiss Organ Cult (2012) 109:391–400
123