Toxicology and Applied Pharmacology 234 (2009) 192–201
Contents lists available at ScienceDirect
Toxicology and Applied Pharmacology
j ourna l homepage: www.e lsev ie r.com/ locate /ytaap
Patulin causes DNA damage leading to cell cycle arrest and apoptosis throughmodulation of Bax, p53 and p21/WAF1 proteins in skin of mice
Neha Saxena, Kausar M. Ansari, Rahul Kumar, Alok Dhawan, Premendra D. Dwivedi, Mukul Das ⁎Food Toxicology Division, Indian Institute of Toxicology Research (formerly: Industrial Toxicology Research Centre), Council of Scientific and Industrial Research,Mahatma Gandhi Marg, P.O. Box #80, Lucknow-226001, India
Patulin (PAT), a mycotoxin Received 25 June 2008Revised 25 September 2008Accepted 29 September 2008Available online 18 October 2008
Keywords:PatulinApoptosisCell cyclep53p21/WAF1Cytochrome C
found in apples, grapes, oranges, pear and peaches, is a potent genotoxiccompound. WHO has highlighted the need for the study of cutaneous toxicity of PAT as manual labour isemployed during pre and post harvest stages, thereby causing direct exposure to skin. In the present studycutaneous toxicity of PAT was evaluated following topical application to Swiss Albino mice. Dermal exposureof PAT, to mice for 4 h resulted in a dose (40–160 μg/animal) and time (up to 6 h) dependent enhancement ofornithine decarboxylase (ODC), a marker enzyme of cell proliferation. The ODC activity was found to benormal after 12 and 24 h treatment of patulin. Topical application of PAT (160 μg/100 μl acetone) for 24–72 hcaused (a) DNA damage in skin cells showing significant increase (34–63%) in olive tail moment, a parameterof Comet assay (b) significant G 1 and S-phase arrest along with induction of apoptosis (2.8–10 folds) asshown by annexin V and PI staining assay through flow cytometer. Moreover PAT leads to over expression ofp21/WAF1 (3.6–3.9 fold), pro apoptotic protein Bax (1.3–2.6) and tumor suppressor wild type p53 (2.8–3.9 fold)protein. It was also shown that PAT induced apoptosis was mediated through mitochondrial intrinsicpathway as revealed through the release of cytochrome C protein in cytosol leading to enhancement ofcaspase-3 activity in skin cells of mice. These results suggest that PAT has a potential to induce DNA damageleading to p53 mediated cell cycle arrest along with intrinsic pathway mediated apoptosis that may also becorrelated with enhanced polyamine production as evident by induction of ODC activity, which may havedermal toxicological implications.
Patulin (4-hydroxy-4H-furo {2, 3-C} pyron-2 {6H}-1) (clavacin;PAT; Fig. 1), is a mycotoxin produced by several species of Penicillium,Aspergillus and Byssochlamys (Steiman et al., 1989). It is a commoncontaminant of ripe apples, used for the production of juiceconcentrates owing to its solubility in water (Harrison, 1989). PAThas also been determined in pears, apricots, peaches, tomatoes,grapes and oranges and in products derived from these fruits (Prietaet al., 1993; Harrison, 1989). Considering the toxicity of patulin,several countries round the globe have set a maximum permissiblelevel of patulin as 50 ppb in foodstuffs (Codex, 2003; USFDA, 2004;EU, 2003).
PAT has been classified as group-3 carcinogen (IARC, 1987).Several organs including kidney, liver, intestinal tissue and immunesystem have been found to be affected by in vivo administration ofPAT (Speijers et al., 1988; Wichmann et al., 2002). Apart from theacute toxic effects, PAT is reported to be teratogenic and carcino-genic in certain experimental animals (Ciegler et al., 1976; Osswald
tmail.com (M. Das).
ll rights reserved.
et al., 1978). Few studies also reveal PAT induced DNA damage,chromosome aberration, and micronuclei formation in mammaliancells (Alves et al., 2000; Thust et al., 1982; Liu et al., 2003). Aprevious study shows a rapid and persistent activation of extra-cellular signal regulated protein kinases 1 and 2 (ERK 1/2) by PATexposure in certain human cell lines like embryonic kidney(HEK293) cells, peripheral blood mononuclear cells (PBMCs), andMadin–Darby canine kidney (MDCK) cells (Wu et al., 2005). Findingsof similar group have also revealed that PAT leads to rapid activationof two more major mitogen-activated protein kinases (MAPKs), p38kinase and c-Jun N-terminal kinase (JNK) in HEK293 cells (Liu et al.,2006), however only p38 kinase signaling pathway contributes toPAT induced cell death. PAT being electrophilic in nature exertscytotoxic and immunotoxic effects mainly through binding withsulphydryl groups, proteins and amino acids in the plasmamembrane (Riley and Showker, 1991). PAT has been shown tocause effects on cell-cycle distribution, which is responsible for overexpression of a functional p53 protein in V79 cell lines and primaryhuman skin fibroblasts (Lehmann et al., 2003).
The disruption of the equilibrium between cell proliferation andcell death is considered to be an early and important event incarcinogenic process (Di Giovanni, 1992). Recent evidence indicates
Data represent mean±S.E. of 3 animals in each group.Values in parenthesis indicate percent increase (↑).Details of treatment schedule and processing of cells are mentioned in Materials andmethods.⁎ Pb0.05, significant when compared to control.
Fig. 1. Structure of patulin.
193N. Saxena et al. / Toxicology and Applied Pharmacology 234 (2009) 192–201
that polyamines, cell proliferation, and apoptosis are tightlyconnected in a quite complex interplay (Tantini et al., 2006). Itappears that polyamines are Janus-faced regulators of cellular fate,promoting either cell proliferation or cell death, depending on thecell type as well as on the environmental signals (Thomas andThomas, 2001). In several cell types high levels of polyamines maydirectly cause cell death (Poulin et al., 1995; Stefanelli et al. 1998;Erez et al., 2002). Polyamine levels within the cells are regulatedand modulated by the key enzymes that control polyaminebiosynthesis particularly ornithine decarboxylase activity (ODC)that rapidly responds to several stimuli, which is mainly linked tocell growth and/or toxicity (Wallace et al., 2003; Thomas andThomas, 2001). Furthermore, a recent study reveals that elevated
Fig. 2. Effect of topical application of patulin on epidermal ODC activity of mice. (A) Dosedependent effect of topical application of patulin and TPA on epidermal ODC activity ofmice exposed for 4 h. (B) Time dependent effect of patulin on epidermal ODC activity ofmice exposed for 2–24 h. The dose regimen and treatment protocols are described inMaterials andmethods. The dose for time dependent effect of patulinwas 160 μg/0.1 ml.The skin ODC activity was measured in cytosolic fraction using [14C] ornithine assubstrate. Each value represents the mean±SE of three animals. ⁎Pb0.05, significantwith respect to control.
ODC activity and increased biosynthesis of polyamines serve as anovel stimulus to induce the ataxia telangiectasia mutated (ATM)-DNA damage signaling pathway and cell death in normalkeratinocytes (Wei et al., 2008) The tumor suppressor gene p53
mediates activation of programmed cell death, in part by up-regulation of mitochondrial Bax expression (Miyashita et al., 1994).Bax is a key component for apoptosis through mitochondrial stress(Wei et al., 2001). Bax forms oligomers and translocates from thecytosol to the mitochondrial membrane (Jurgensmeier et al., 1998).It increases the membranes permeability through interactions withthe pore proteins on the mitochondrial membrane, which leads tothe release of cytochrome C from mitochondria, and activation ofcaspase 3, a notable effector in apoptosis which is a convergencepoint for two different caspase dependent apoptosis pathway(Narita et al., 1998).
According to WHO guidelines, humans and animals may beexposed to mycotoxins through ingestion, inhalation or skin contact(WHO, 1998). Limited knowledge is available regarding epidermalcarcinogenesis of mycotoxins and WHO has clearly highlighted theneed for toxicological evaluation of mycotoxins through dermalexposure (Anonymous, 1990). This is an important aspect from thepoint of view of developing countries in tropics including Indiawhere manual labour is employed during pre- and post-harveststages in agriculture thus indicating a probable cause of exposurethrough dermal route. In this regard our prior studies haverevealed cutaneous carcinogenic implications of a mycotoxin,Aflatoxin-B1 (AFB1) through dermal exposure and in a sequelstudy it is shown that extract of Ocimum sanctum leaf may cause
Table 2Different phases of skin cells of mice topically treated with patulin
Data represent mean±S.E. of 3 animals per group.Values in parenthesis indicate percent increase (↑) or decrease (↓).Details of treatment schedule and processing of cells are mentioned in Materials andmethods.⁎ Pb0.05, significant when compared to control.
194 N. Saxena et al. / Toxicology and Applied Pharmacology 234 (2009) 192–201
cell cycle arrest in S phase, also suggesting it to have antiproliferative activity (Rastogi et al., 2006, 2007).
On similar lines we have further explored the toxicologicalmechanism of another mycotoxin, patulin in skin cells. Moreoverdue to widespread nature of fungi, mycotoxins act as unavoidablecontaminants in most foods and feeds, therefore it is worthwhile togenerate toxicological data on PAT through dermal exposure.Although, few studies have shown involvement of specific proteinkinases in PAT toxicity in certain in-vitro models (Wu et al., 2005;Liu et al., 2006), however the cellular and molecular mechanisms ofPAT toxicity on exposure to skin are not yet investigated. Thus inthis study, an attempt has been made to establish the role of PAT inactivation of ODC, a marker of cell proliferation. Simultaneously theDNA damaging potential of PAT was also explored that may act as astress signal in up regulation of p53 protein leading to apoptosis.Because of its pivotal role in cell cycle control p21/WAF1 proteinlevels and cell cycle analysis were also investigated in skin cells ofmice topically exposed with PAT. To further explore if themitochondrial pathway is responsible for apoptosis, expression ofBax protein, cytochrome C protein and caspase 3 activity were alsoundertaken.
Fig. 3. Skin cells of mouse with olive tail moment (OTM) after exposure to patulin. (A) Histcontrol animal without comet (400×); (C) Skin cells of patulin (160 μg/mouse) treated animanimals showing comet formation (400×). For preparation of skin cells, the chopped skin piefiltered with muslin cloth and used as single cell suspension. Comet assay was performed a
Materials and methods
Chemicals. Patulin (PAT), dithiothreteiol (DTT), phenylmethyl-sulphonyl fluoride (PMSF), 2-mercaptoethanol, L-ornithine, pyridoxal-5-phosphate, benzo(a)pyrene (BP), 12-O-tetradecanoylphorbol-acetate(TPA), diaminobenzidine, propidium iodide (PI), RNase A, normalmelting agarose, low melting point agarose (LMPA) protease inhibitorcocktail set-I, ethidium bromide (EtBr), ethylenediamine tetraaceticacid (EDTA) disodium salt, Tris buffer, triton X-100 and bovine serumalbumin (BSA) were obtained from Sigma Chemicals Co. (St. Louis, MO).[14C] Ornithine (specific activity, 56 mCi/mol) was purchased fromAmersham Biosciences (Searle, Chicago, IL). Monoclonal antibody(mice) against p21/WAF1 and p53 were procured from BD Biosciences,San Jose, CA, USA while polyclonal rabbit antibody against Bax waspurchased from Cell Signaling, Beverly, MA, USA. Horseradishperoxidase (HRP) and IgG-conjugated secondary antibody waspurchased from Bangalore Genei (Bangalore, India) and HRP-conjugated β-actin was procured from Santa Cruz Biotechnology(Santa Cruz, CA). Hank's balanced salt solution (HBSS) was obtainedfromHiMedia Pvt. Ltd., Mumbai, India. All other chemicals usedwere ofhighest purity commercially available.
ogram showing percent distribution of skin cells with respect to OTM; (B) Skin cells ofals showing comet formation (400×) and (D) Skin cells of BP (160μg/mouse) treated
ces were kept in HBSS buffer containing 10% FBS and stirred at 4 °C. The suspension wass described in Materials and methods.
Fig. 4. Effect of topical application of patulin on the expression of p53, p21/WAF1 and Baxproteins in skin cells. (A) 60 μg proteins were loaded and separated on a 12% SDS-PAGEgel. Proteins were transferred to PVDF membranes and incubated with primarymonoclonal antibodies raised against p53, p21/WAF1 and Bax and a secondary HRP-conjugated antibody as described in Materials and methods. For confirmation of equalprotein loading, the blots were also probed with an antibody specific for β-actin. Lane1:Control; lane 2: acetone (vehicle-treated); lane 3: 160 μg BP, 24 h (positive control); lane4: 160 μg patulin, 24 h; lane 5: 160 μg patulin, 48 h; and lane 6: 160 μg Patulin, 72 h. (B)The relative protein expression levels are mean±S.E for the densitometric analyses ofthree independent experiments normalized by arbitrarily setting the value foruntreated cells as 1. ⁎Pb0.05, significant with respect to control.
195N. Saxena et al. / Toxicology and Applied Pharmacology 234 (2009) 192–201
Estimation of cutaneous ODC activity. Six to seven weeks oldfemale Swiss albino mice (18–22 g) were derived from IndianInstitute of Toxicology Research (IITR) animal breeding colony,raised on commercial pellet diet (Ashirwad, Chandigarh, India) andwater ad libitum were used. The dorsal surfaces of mice wereshaved 3 d prior to treatment. For dose dependent study animalswere divided into four groups containing 3 animals each: (i)Untreated, served as control (ii) acetone (100 μl) topical application,served as vehicle control; (iii) TPA (2.5 μg/0.1 ml acetone) wastopically applied and served as positive control; and (iii) patulin(40, 80 and 160 μg/0.1 ml acetone) was topically applied. Theanimals were maintained in controlled atmosphere of 12 h dark/light, 22±2 °C and 50–60% humidity, fasted overnight after lastdose and euthanized after 4 h (Pandey et al., 2006) by cervicaldislocation with minimal suffering following the rules laid down byAnimal Welfare Committee of IITR.
For time dependent study animals were divided into four groupscontaining 3 animals each: (i) Untreated, served as control (ii)acetone (100 μl) topical application, served as vehicle control; (iii)TPA (2.5 μg/0.1 ml acetone) was topically applied and served aspositive control; and (iv) patulin (160 μg/0.1 ml acetone). Theanimals were maintained in controlled atmosphere of 12 h dark/light, 22±2 °C and 50–60% humidity, fasted overnight after last doseand euthanized after 2, 4, 6, 8 and 12 and 24 h. The skin wasdissected out and cytosolic ODC activity was assayed by the methodof Verma et al. (1979). The specific activity was expressed as pmol14CO2 released/h/mg protein.
Alkaline comet assay. Six to seven weeks old female Swiss albinomice (18–22 g) were divided into four groups. One of the groupsserved as control, which remained untreated. Remaining groupswere treated in the following manner (i) acetone (100 μl) wastopically applied for 24 h, which served as negative control; (ii) BP,(160 μg/100 μl acetone) was topically applied for 24 h and served aspositive control; (iii) PAT (160 μg/100 μl acetone) was topicallyapplied and animals were euthanized after 6, 12, 24, 48, and 72 h.Skin samples from all the groups of animals were dissected out andthe single cell suspension was prepared as described earlier (Das etal., 2005). The alkaline comet assay technique of Singh et al. (1998)and Tice et al. (2000) was utilized for evaluation of DNA damage inmouse skin cells. Slides were prepared and scored with an imageanalysis system (Kinetic Imaging, Liverpool, UK) attached to afluorescence microscope (Leica, Germany) equipped withappropriate filters. The microscope was connected to a computerthrough a charge coupled device (CCD) camera to transport imagesto software (Komet 5.0) for analysis. The final magnification was400×. The parameters undertaken in the study were the olive tailmoment (OTM) (arbitrary units), tail DNA (%), and tail length(migration of the DNA from the nucleus; μm).
Immunoblotting for p53, Bax and p21/WAF1 proteins. The excised skinfrom control, vehicle, BP treated and PAT treated mice for 24–72 hwere homogenized and processed according to the modified methodof Serpi et al. (1999). In brief, the skin sample was homogenized withUltraTurax Polytron (Janke & Kunkel, IKA-Labortechnik, Staufen,Germany) in ice-cold cell lysis buffer (containing 0.02 M HEPES,250 mMNaCl, 1 mM EDTA, 1 mM EGTA, 1% NP-40, 0.2 mM PMSF) withfreshly added protease inhibitor mixture. The homogenate wasincubated for 30 min in an ice bath. The samples were thencentrifuged at 16,000 ×g for 20 min at 4 °C. The clear supernatantobtained (whole cell extract) was used for detection of p53, p21/WAF1
and Bax proteins. Sixty μg proteins were resolved on 12% SDS-PAGE,electro transferred upon a methanol soaked PVDF membrane, for 2 hat 48 mA, with a semidry electroblotting assembly (Amersham LifeSciences, Inc.). Membranes were incubated in blocking solutioncontaining 3% bovine serum albumin (BSA) in PBST (phosphate buffer
saline, pH 7.4 containing 0.1% Tween-20) buffer for 1.5 h at roomtemperature with gentle shaking, followed by incubation overnight at4 °C with a specific primary antibody of p53, p21/WAF1, β-actin and Bax,at the dilutions recommended by the manufacturer. The membraneswere washed three times in PBST buffer each for 25 min, followed byincubation with horseradish peroxidase (HRP)-IgG-conjugatedsecondary antibody for 2 h. Subsequently membranes were washedin TBST buffer for 25 min each and color developed with 3-3-diaminobenzidine tetrahydro-chloride (DAB system). DAB is asubstrate for HRP, which yields a colored deposit. The bands werequantitated with Gene tools software (version 3.02) of GelDocumentation System (Syngene, UK).
Cell cycle analysis. The single cell suspension of skin for the groupsmentioned for immunoblot assay was prepared with Medimachine(Becton-Dickinson, San Jose, CA) as described earlier (Pandey et al.,2006). The suspensionwas centrifuged, washed thrice with 0.5 ml PBSand fixed in ice-cold 70% methanol. The cells were flushed through a21-gauge needle (3–4 times) for uniform dispersion. The fixed cellswere again washed twice with PBS and incubated with Propidiumiodide (20 μg/ml) and RNase A (200 μg/mL) for 60 min at 37 °C. Dataacquisition was performed with an argon laser fluorescence-activatedcell analyzer (FAC Sort, Becton-Dickinson) For each sample, 10,000events were acquired according to the guidelines for the analysis ofDNA content and the coefficient of variance was b8.0% (Shankey et al.
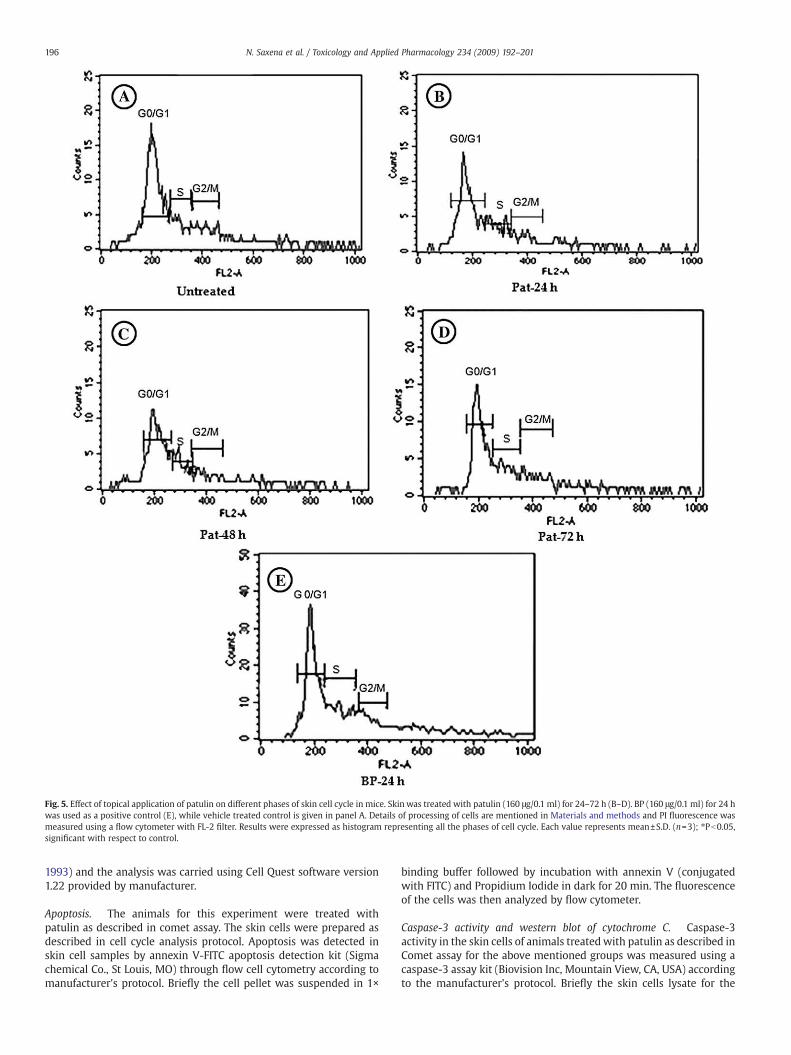
Fig. 5. Effect of topical application of patulin on different phases of skin cell cycle in mice. Skin was treated with patulin (160 μg/0.1 ml) for 24–72 h (B–D). BP (160 μg/0.1 ml) for 24 hwas used as a positive control (E), while vehicle treated control is given in panel A. Details of processing of cells are mentioned in Materials and methods and PI fluorescence wasmeasured using a flow cytometer with FL-2 filter. Results were expressed as histogram representing all the phases of cell cycle. Each value represents mean±S.D. (n=3); ⁎Pb0.05,significant with respect to control.
196 N. Saxena et al. / Toxicology and Applied Pharmacology 234 (2009) 192–201
1993) and the analysis was carried using Cell Quest software version1.22 provided by manufacturer.
Apoptosis. The animals for this experiment were treated withpatulin as described in comet assay. The skin cells were prepared asdescribed in cell cycle analysis protocol. Apoptosis was detected inskin cell samples by annexin V-FITC apoptosis detection kit (Sigmachemical Co., St Louis, MO) through flow cell cytometry according tomanufacturer's protocol. Briefly the cell pellet was suspended in 1×
binding buffer followed by incubation with annexin V (conjugatedwith FITC) and Propidium Iodide in dark for 20 min. The fluorescenceof the cells was then analyzed by flow cytometer.
Caspase-3 activity and western blot of cytochrome C. Caspase-3activity in the skin cells of animals treatedwith patulin as described inComet assay for the above mentioned groups was measured using acaspase-3 assay kit (Biovision Inc, Mountain View, CA, USA) accordingto the manufacturer's protocol. Briefly the skin cells lysate for the
197N. Saxena et al. / Toxicology and Applied Pharmacology 234 (2009) 192–201
above mentioned groups was prepared in the lysis buffer provided bythe manufacturer followed by incubation in ice for 10 min. Thesubstrate was then added followed by reincubation at 37 °C for 1–2 h.
Fig. 6. Effect of topical application of patulin on apoptosis of skin cells inmice. The single cell scytometry. Each dot blot represents 10,000 events (cells).
The fluorescence intensity was measured at an excitation wavelengthof 400 nm and an emission wavelength of 505 nm using microplatereader (Synegy HT Biotech, USA).
uspensionwas prepared and apoptosis was detected by annexin-V FITC kit through flow
Fig. 7. Effect of topical application of patulin on release of cytochrome C and Caspase-3 activity in skin cells of mice. (A) Cytochrome C in the cytosolic fraction of the cellstreated with patulin was analyzed by western blot using anti cytochrome C antibody.For confirmation of equal protein loading, the blot was also reprobed with anantibody specific for β-actin.Lane1: Control; lane 2: 160 μg BP, 24 h (positive control);lane 3: 160 μg Patulin, 6 h; lane 4: 160 μg Patulin, 12 h; lane 5: 160 μg Patulin, 24 h;lane 6: 160 μg Patulin, 48 h; and lane 7:160 μg Patulin, 72 h. (B) The proteinexpression levels are mean±SE for the densitometric analysis of three independentexperiments normalized by arbitrarily setting the value for untreated or control cellsas 1. ⁎Pb0.05, significant with respect to control. (C) Caspase-3 activity in skin cells ofmice treated with patulin was measured using a caspase-3 assay kit in terms ofrelative fluorescence unit from control at an excitation wavelength of 400 nm and anemission wavelength of 505 nm. Each value represents mean±S.E. of four values.⁎Pb0.05, significant with respect to control.
198 N. Saxena et al. / Toxicology and Applied Pharmacology 234 (2009) 192–201
The proteins from cytosolic fraction of skin homogenate preparedfrom animals treated with patulin as described in Comet assayprotocol were taken for cytochrome C detection through western blotanalysis using mouse monoclonal anti cytochrome C antibody(Biovision Inc, Mountain View, CA, USA).
Protein assay. Protein in the samples was determined using bovineserum albumin as standard (Lowry et al., 1951).
Statistical analysis. Values are presented as mean±SE. Statisticalevaluation was carried with one-way ANOVA (Snedecor and Cochran,1967). In all the cases, P values less than 0.05 were consideredsignificant when compared to controls.
Results
The present study was undertaken to determine the effect of PATon ODC to assess its proliferative potential and DNA damage inducedapoptotic signaling pathway. A dose dependent induction profile ofcutaneous ODC activity following single topical application of PAT anda known tumor promoter, TPA, is given in Fig. 2A. Different doses ofPAT were used to measure ODC activity and maximum ODC induction(867%) was found at 160 μg/100 μl acetonewhich was the highest doseof patulin used. TPA, used as positive control, showed 1007% inductionof dermal ODC activity. However, acetone treatment tomice, chosen asvehicle showed no difference in enzyme activity when compared tocontrol. Similarly a time dependent profile of ODC activity (Fig. 2B) at160μg/0.1 ml patulin exposure shows significant enhancement at 4 h,which reached to maximum level at 6 h and then decreased at 8 h andre-established the basal levels at further 12 and 24 h. However, TPAexposure showed the maximum induction of ODC activity at 4 h,which decreased at 6 h and then came to basal level after 8–24 h.
DNA damage in skin cells of mice treated with single topicalapplication of PAT and BP is shown in Table 1. Positive control groupfollowing 24 h exposure of BP showed significant increase in OTM(104%), tail DNA (283%), and tail length (128%). PAT treatment for24–72 h caused significant enhancement of OTM (34–63%), tail DNA(32–151%), and tail length (35–124%) when compared to control.However no DNA damage was observed at 6 and 12 h exposure ofcells with PAT. The frequency of skin cells with OTM followingpatulin treatment to mice is shown in Fig. 3A. In OTM category ofb5 and 10, patulin and BP treatment for 24–72 h showedsignificantly lower percentage of skin cells compared to controls.However, in OTM category of 15 and 20, PAT (24–72 h) and BPtreated animals exhibited significantly higher percentage of skincells as compared to control respectively. PAT treatment for 6 and12 h did not show significant difference in percentage of cell in OTMcategory of 15 and 20. The DNA from skin cells prepared frompatulin treated animals showed typical formation of comet (Fig. 3C),similar to that of positive control treated group (BP) (Fig. 3D) whilecontrol cell showed a well defined circular nucleus (3B).
The effect of single topical application of PAT and BP on expressionof mouse skin p53, Bax and p21/WAF1 proteins in mouse skin are shownin Fig. 4. Dermal wild-type p53 (1.3–2.6-fold), Bax (1.0–1.4-fold) andp21/WAF1 (3.6–3.9-fold) protein expressionwere significantly enhancedfrom 24 to 72 h following treatment with PAT when compared tocontrol. Similarly single topical application of BP (positive controlgroup) also showed an increase in expression of p53 (3.5-fold), Bax(2.0-fold) and p21/WAF1 (4.1-fold) after 24 h of exposure whencompared to control. The animals treatedwith acetone did not indicateany over expression of p53, Bax and p21/WAF1when compared to control(Fig. 4). The respective blots were stripped and probed with HRPconjugated β-actin to ensure equal loading for p53, Bax and p21/WAF1
proteins at 53, 20.5 and 21 kDa, respectively.Fig. 5 and Table 2 shows the effect of single topical application
of PAT and BP on cell-cycle distribution of skin cells. Exposure of
PAT to mice skin resulted in significant increase in the proportionof cells in G0/G1 phase following 24 and 48 h when compared tocontrol. S-phase was also found to be significantly affected in cells(37–103%) exposed to patulin. However, G2/M phase was not foundto be significantly affected by PAT exposure in skin cells for 24–72 h. BP-treated animals exhibited similar response to the cellcycle, by increasing the cell proportion in G0/G1 phase (14%) and S-phase (147%) when compared to control (Table 2).
Apoptosis as revealed through the annexin and Propidiumiodide (PI) staining in skin cells of mice treated with single topicalapplication of PAT and BP is shown in Fig. 6. Mice treated with160 μg of PAT for 24, 48, and 72 h showed enhancement of theapoptosis by 2.8, 6.3, and 10-folds, respectively, when compared tocontrol. The similar trend was observed in group treated with BP,
199N. Saxena et al. / Toxicology and Applied Pharmacology 234 (2009) 192–201
which showed an induction of apoptosis (37-fold) in 24 h. HoweverPAT exposed cells for 6 and 12 h showed almost similar values tothat of controls (Fig. 6). To further explore the PAT inducedmechanism of apoptosis, release of cytochrome C and caspase-3activity were evaluated in cytosol of skin cells (Fig. 7). Patulinshowed a time dependent release of cytochrome C protein in skinlysate for 24, 48 and 72 h indicating the involvement ofmitochondrial pathway during apoptosis, however, at 6 and 12 hof patulin treatment no significant difference was observed as
Fig. 8. A proposedmodel for PAT-induced cascade of events resulting from patulin induced ODleads to activation of several apoptosis related proteins causing arrest of cells leading to apopcell and if it is faulty in nature, the repeated insult may proliferate (ODC↑) the mutated cell
compared to control. BP, a positive control showed over expressionof cytochrome C in skin cytosol after 24 h. Caspase 3 was alsofound to be increased in mice treated with patulin (24–72 h) while6 and 12 h treatment showed no effect (Fig. 7C).
Discussion
Mycotoxins are among the most significant food contaminantwith regard to public health and food security. PAT, one of the
C activity and DNAdamage in skin cells. PATcauses ODC elevation and DNAdamage thattotic pathway activation. The arrest of cells by patulin may repair the DNA damage of thes, which is generally observed in case of tumorigenesis.
200 N. Saxena et al. / Toxicology and Applied Pharmacology 234 (2009) 192–201
mycotoxin produced by Penicillium expansum is often detectable inmouldy fruits and their derivatives. Survey studies round the globehave revealed that PAT content in some commercialized apple andpear products exceeds the safety level (50 ppb) established by theWHO (Lai et al., 2000; Yurdun et al. 2001). Moreover, our previousfinding also indicate the presence of PAT in apple juices and otherfruit juices, which exceed the WHO safety limit as analyzed by HPLCand Mass spectrometry (Saxena et al., in press).
There are several in vitro and in vivo studies suggesting the broadtoxicity spectrum of PAT (Alves et al., 2000; Pfeiffer et al., 1998; Imaidaet al., 1982), however none of these investigations indicate themechanism of toxicity of PAT in skin cells. The present study revealsthat PAT has the ability to cause DNA damage in skin as depicted byalkaline comet assay. In addition to this, topical exposure of PAT to themouse skin led to enhanced expression of p53, Bax and p21/WAF1
proteins that causes apoptosis through mitochondrial mediatedpathway along with concomitant induction of cytochrome C proteinand caspase 3 activity.
It has been shown that the tumor suppressor gene p53 is a keyelement in maintaining a balance between cell growth and celldeath in the living system (Mowat, 1998; Kaelin, 1999; Agarwal etal., 1998). Moreover, in response to DNA damage, p53 triggers avariety of cell cycle-regulatory events to limit the proliferation ofdamaged cells. In addition, p53 controlled cell cycle begins with upregulation of p21 protein, which then arrests the cells in the G1phase or G2/M phase to give time to cell for DNA repair (Yang andHughes, 1998). However, when DNA damage exceeds capacity ofcell for repair, p53 guides the corrupt cell into apoptosis byinducing expression of pro-apoptotic protein Bax (Israels andIsraels, 2000). The present study is in agreement with a recentstudy that suggests the significant role of p53 in the occurrence ofhepatocellular carcinoma (HCC) induced by AFB1, a hepatotoxicmycotoxin (Duan et al., 2005).
PAT treatment also caused induction of ODC activity at earlytime points of 4–6 h which gradually declined to basal levels onfurther exposure time, suggesting enhanced production of poly-amines that may later contribute to significant increase of apoptosisas revealed by annexin V and PI staining method in skin cells up to72 h. In spite of the increasing evidence for involvement ofpolyamines in the activation of the programmed death, the role ofODC in apoptosis is complex and not fully understood. However, insome cell system ODC activity is increased after induction ofapoptosis (Packham and Cleveland, 1994; Tantini et al., 2006).Nonetheless, a recent study for the first time revealed thecorrelation of elevated ODC activity with apoptotic cell death innormal keratinocytes via the induced generation of reactivealdehydes and H2O2 followed by the subsequent activation of themutated ATM-DNA damage response pathway (Wei et al. 2008).Thus it is quite likely that induction of ODC by PAT may lead to thegeneration of reactive oxygen species (ROS) which may beinstrumental in apoptosis apart from proliferation of cells. In thisregard prior studies have shown that PAT has the potential forformation of ROS capable of inducing DNA damage in rat liver cells,human embryonic kidney (HEK293) and human promyelocyticleukemia (HL-60) cells (Barhoumi and Burghardt, 1996; Liu et al.,2007).
Pat exposure showed arrest of skin cells in G1 and S phase ofcell cycle which indicate that the irreparable DNA damaged cellsmay undergo apoptosis or if DNA repair takes place then the cellsmay further be proliferated because of induction of ODC activity.Thus it may be emphasized that induction of ODC by PAT may beinvolved in the regulation of the cell cycle as well as in cell cyclealternation, which may further induce apoptosis. However, earlierin vitro studies have shown that PAT caused G2/M phase arrest ofV79 and cultured mammalian fibroblasts cells (Lehmann et al.,2003; Schumacher et al., 2005) that might allow cells to repair
DNA damage prior to continuing into the S-phase or undergoingmitosis.
The observed induction of apoptosis by PAT is of significancebecause this phenomenon is a physiological process that functions asan essential mechanism of tissue homeostasis and is regarded as thepreferred way to eliminate unwanted cells. On the other hand it isquite likely that PAT exposure could lead to toxicity by enhancingapoptosis in normal skin cells which involves a cascade of eventsincluding cell cycle arrest at G0/G1 phase. The cell cycle arrest bypatulin may pass some of the cells for DNA repair and if it is faulty innature, the repeated insult may proliferate the mutated cells, which isgenerally observed in case of tumorigenesis protocol (Das et al., 2005).A summarized figure (Fig. 8) is shown to describe the probable cellularmechanism of PAT induced dermal toxicity. The present study findssignificance with the rationale that patulin contamination in pearsand apples has been detected at the levels of 288 μg/g on the exteriorside of the fruit (Laidou et al., 2001). Thus, probable human exposurethrough skin may occur at 288 μg during pre- and post-harvest stages,which has got relevance with the present study where 160 μg dosehave been used. Further, the associated risk of multiple exposure toPAT needs to be investigated. In conclusion, our study suggests thatPAT caused a time dependent ODC induction that is transientaccompanied by enhanced levels of p53, Bax, cytochrome C, caspase-3 and p21/WAF1 proteins leading to apoptosis and delay of cells in G0/G1and S-phase thereby indicating the possible dermal carcinogenicpotential of PAT.
Acknowledgments
We are grateful to the Director of our institute, for his keeninterest in this study. Neha Saxena and Rahul Kumar are thankful tothe Council of Scientific and Industrial Research (CSIR)/UniversityGrants Commission, New Delhi for the award of Senior ResearchFellowship and Junior Research Fellowship, respectively. The work issupported by CSIR Network Project NWP-17 and DST project. Wegratefully acknowledge Ms. Kulpreet Kaur for her skilful assistancein providing flow cytometer facility. The manuscript is IITRcommunication # 2684.
References
Agarwal, M.L., Taylor, W.R., Chernov, M.V., Chernova, O.B., Stark, G.R., 1998. The p53network. J. Biol. Chem. 273, 1–4.
Anonymous, 1990. Manuals of Food Quality Control. 10. Training in Mycotoxin Analysis.Food and Agriculture Organization of the United Nations, Rome.
Alves, I., Oliveira, N.G., Laires, A., Rodrigues, A.S., Rueff, J., 2000. Induction of micronucleiand chromosomal aberrations by the mycotoxin patulin in mammalian cells: role ofascorbic acid as a modulator of patulin clastogenicity. Mutagenesis 15, 229–234.
Barhoumi, R., Burghardt, R.C., 1996. Kinetic analysis of the chronology of patulin andgossypol-induced cytotoxicity in vitro. Fundam. Appl. Toxicol. 30, 290–297.
Ciegler, A., Becwith, A.C., Jackson, L.K., 1976. Teratogenicity of patulin and patulinadducts formed with cysteine. Appl. Environ. Microb. 31, 664–667.
Codex Alimentarius commission 2003. Maximum level for patulin in apple juice andapple juice ingredients and other beverages. No: 235, 30 June–5 July, 26th Session,Rome, Italy.
Das, M., Ansari, K.M., Dhawan, A., Shukla, Y., Khanna, S.K., 2005. Correlation of DNAdamage in Epidemic Dropsy patients to carcinogenic potential of argemone oil andisolated sanguinarine alkaloid in mice. Int. J. Cancer 117, 709–717.
Di Giovanni, J., 1992. Multistage carcinogenesis in mouse skin (a review). Pharmac. Ther.54, 63–128.
Duan, X.X., Ou, J.S., Li, Y., Su, J.J., Ou, C., Yang, C., Yue, H.F., Ban, K.C., 2005. Dynamicexpression of apoptosis related genes during development of laboratory hepato-cellular carcinoma and its relation to apoptosis. World J. Gastroenterol. 11,4740–4744.
Erez, O., Goldstaub, D., Friedman, J., Kahana, C., 2002. Putrescine activates oxidativestress dependent apoptotic death in ornithine decarboxylase overproducing mousemyeloma cells. Exp. Cell Res. 281, 148–156.
European Commission (EC), 2003. Regulation No 1425/2003 as regards patulin. OfficialJ. European Union L. 203, 1–3.
Harrison, M.A., 1989. Presence and stability of patulin in apple products: a review. J.Food Safety 9, 147–153.
IARCmonographs on the evaluation of carcinogenic risks to humans; Overall evaluationof carcinogenicity 1987. An updating of IARCmonographs Volumes 1 to Supplement7. IARC, Lyon.
201N. Saxena et al. / Toxicology and Applied Pharmacology 234 (2009) 192–201
Imaida, K., Hirose, M., Ogiso, T., Kurata, Y., Ito, N., 1982. Quantitative analysis of initiatingand promoting activities of five mycotoxins in liver carcinogenesis in rats. CancerLett. 16, 137–143.
directly induces release of cytochrome c from isolated mitochondria. P. Natl. Acad.Sci. U. S. A. 95, 4997–5002.
Kaelin, W.G., 1999. The emerging p53 gene family. J. Natl. Cancer Inst. (Bethesda) 91,594–598.
Lai, C.L., Fu, Y.M., Shih, Y.C., 2000. Determination of mycotoxin patulin in apple juice. J.Food Drug Analysis 8, 85–96.
Laidou, I.A., Thanassoulopoulos, C.C., Liakopoulou-Kyriakides, M., 2001. Diffusion ofpatulin in the flesh of pears inoculated with four post harvest pathogens. J.Phytopathol. 149, 457–461.
Lehmann, L., Franz, U., Metzler, M., 2003. Genotoxic potential of the mycotoxin patulinin cultured mammalial fibroblasts. Naunyn-Schmiedeberg's Arch. Pharmacol. 367,R166.
Liu, B.H., Yu, F.Y., Wu, T.S., Li, S.Y., Su, M.C., Wang, M.C., Shih, S.M., 2003. Evaluation ofgenotoxic risk and oxidative DNA damage in mammalian cells exposed tomycotoxins, patulin and citrinin. Toxicol. Appl. Pharm. 191, 255–263.
Liu, B.H., Wu, T.S., Yu, F.Y., Wang, C.H., 2006. Mycotoxin patulin activates the P38 kinaseand JNK signalling pathways in human embryonik kidney cells. Toxicol. Sci. 89,423–430.
Liu, B.H., Wu, T.S., Yu, F.Y., Su, C.C., 2007. Induction of oxidative stress response by themycotoxin patulin in mammalian cells. Toxicol. Sci. 95, 340–347.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randal, R.J., 1951. Protein measurement withfolin phenol reagent. J. Biol. Chem. 193, 265–275.
Miyashita, T., Krajewski, S., Krajewski,M., Wang, H.G., Lin, H.K., Hoffman, B., 1994. Tumorsuppressor p53 is a regulator of bcl-2 and bax gene expression in vitro and in vivo.Oncogene 9, 1799–1805.
Mowat, M.R., 1998. P53 in tumor progression: life, death, and everything. Adv. CancerRes. 74, 25–48.
Narita, M., Shimizu, S., Ito, T., Chittenden, T., Lutz, R.J., Matsuda, H., Tsujimoto, Y., 1998.Bax interacts with the permeability transition pore to induce permeabilitytransition and cytochrome c release in isolated mitochondria. Proc. Natl. Acad.Sci. U. S. A. 95, 14681–14686.
Osswald, H., Frank, H.K., Komitowski, D., Winter, H., 1978. Long term testing of patulinadminister orally to Sprague-Dawley rats and Swiss mice. Food Cosmet. Toxicol. 16,243–247.
Packham, G., Cleveland, J.L., 1994. Ornithine decarboxylase is a mediator of c-Mycinduced apoptosis. Mol. Cell Biol. 14, 5741–5747.
Pandey, M.K., Dhawan, A., Das, M., 2006. Induction of p53, p21Waf1, ornithinedecarboxylase activity, and DNA damage leading to cell-cycle arrest and apoptosisfollowing topical application of repeated fish fried oil extract to mice. Mol.Carcinogen. 45, 805–813.
Pfeiffer, E., Gross, K., Metzler, M., 1998. Aneuploidogenic and clastogenic potential of themycotoxins citrinin and patulin. Carcinogenesis 19, 1313–1318.
Poulin, R., Pelletier, G., Pegg, A.E., 1995. Induction of apoptosis by excessive polyamineaccumulation in ornithine decarboxylase-overproducing L1210 cells. Biochem. J.311, 723–727.
Prieta, J., Moreno, M.A., Bayo, J., Diaz, S., Suarez, G., Dominguez, L., Canela, R., Sanchis, V.,1993. Determination, of patulin by reversed-phase high-performance liquidchromatography with extraction by diphasic dialysis. Analyst 118, 171–173.
Rastogi, S., Shukla, Y., Paul, B.N., Chowdhuri, D.K., Khanna, S.K., Das, M., 2007. Protectiveeffect of Ocimum sanctum on 3-methylcholanthrene, 7,12-dimethylbenz(a)anthra-cene and aflatoxin B1 induced skin tumorigenesis in mice. Toxicol. Appl. Pharm.224, 228–240.
Riley, R.T., Showker, J.L., 1991. The mechanism of patulin's cytotoxicity and theantioxidant activity of indole tetramic acids. Toxicol. Appl. Pharm. 109, 108–126.
Saxena, N., Dwivedi, P.D., Ansari, K.M., Das, M., in press. Incidence of patulin in apple
juices and its likely intake in Indian population. Food Additive and Contaminants.doi:10.1080/02652030802378848.
Schumacher, D.M., Metzler, M., Lehmann, L., 2005. Mutagenicity of the mycotoxinpatulin in cultured Chinese hamster V79 cells, and its modulation by intracellularglutathione. Arch. Toxicol. 79, 110–121.
Serpi, R., Piispala, J., Jarvilehto, M., Vahakangas, K., 1999. Thapsigargin has similar effecton p53 protein response to benzo (a) pyrene DNA adducts as TPA in mouse skin.Carcinogenesis 20, 1755–1760.
Shankey, T.V., Rabinovitch, P.S., Bagwell, B., Bauer, K.D., Duque, R.E., Hedley, D.W., 1993.Guidelines for implementation of clinical DNA cytometry. The Intl. Society for Anal.Cytol. 14, 472–477.
Singh,N.P.,Mecony,M.T., Tice, R.R., Schneider, E.L.,1998.A simple techniqueofquantitationof low levels of DNA damage in individual cells. Exp. Cell Res. 175, 184–191.
Snedecor, G.W., Cochran, W.G., 1967. One way classification: analysis of variance.Statistical Methods. lowa State University Press, Iowa, pp. 258–296.
Speijers, G.J., Franken, M.A., van Leeuwen, F.X., 1988. Subacute toxicity study of patulinin the rat: effects on the kidney and the gastro-intestinal tract. Food Chem. Toxicol.26, 23–30.
Stefanelli, C., Bonavita, F., Stanic', I., Mignani, M., Facchini, A., Pignatti, C., Tantini, B.,Muscari, C., 1998. Spermine causes caspase activation in leukaemia cells. FEBS Lett.437, 233–266.
Steiman, R., Seigle-Murandi, F., Sage, L., Krivobook, S., 1989. Production of patulin bymicromycetes. Mycopathologia 105, 129–133.
Tantini, B., Fiumana, E., Cetrullo, S., Pignatti, C., Bonavita, F., Shantz, L.M., Giordano, E.,Muscari, C., Flamigni, F., Guarnieri, C., Stefanelli, C., Caldarera, C.M., 2006.Involvement of polyamines in apoptosis of cardiac myoblasts in a model ofsimulated ischemia. J. Mol. Cell Cardiol. 40, 775–782.
Thomas, T., Thomas, T.J., 2001. Polyamines in cell growth and cell death: molecularmechanisms and therapeutic applications. Cell Mol. Life Sci. 58, 244–258.
Thust, R., Kneist, S., Mendel, J., 1982. Patulin, a further clastogenic mycotoxin, isnegative in the SCE assay in Chinese hamster V79-E cells in vitro. Mutat. Res. 103,91–97.
Tice, R.R., Aguvell, E., Anderson, D., Burlinson, B., Hartmann, A., Keboyashi, H., Myamae,Y., Rojas, E., Ryu, J.C., Sasaki, Y.F., 2000. Single cell gel/comet assay: guidelines for invitro and in vivo genetic toxicology testing. Environ. Mol. Mutagen. 35, 206–221.
U.S. Food and Drug Administration [USFDA], 2004. Apple Juice, Apple JuiceConcentrates, and Apple Juice Products Adulteration with Patulin. CompliancePolicy Guidance for FDA Staff. Sec. 510.150.
Verma, A.K., Shapas, B.G., Rice, H.M., Boutwell, R.K., 1979. Correlation of the inhibitionby retinoid of tumor promoter-induced mouse epidermal ornithine decorboxylaseactivity and skin tumor promotion. Cancer Res. 39, 419–425.
Wei, G., DeFeo, K., Hayes, C.S., Woster, P.M., Nayak, L.M., Gilmour, S.K., 2008. Elevatedornithine decarboxylase levels activate ataxia telangiectasia mutated-DNA damagesignalling in normal keratinocytes. Cancer Res. 68, 2214–2222.
WHO Food Additives Series: 47, Safety evaluation of certain mycotoxins in food.Prepared by 49th meeting of JECFA, WHO, Geneva, 1998.
Wichmann, G., Herbarth, O., Lehmann, I., 2002. The mycotoxins citrinin, gliotoxin, andpatulin affect interferon-gamma rather than interleukin-4 production in humanblood cells. Environ. Toxicol. 17, 211–218.
Wallace, H.M., Fraser, A.V., Hughes, A., 2003. A perspective of polyamine metabolism.Biochem. J. 376, 1–14.
Wu, T.S., Yu, F.Y., Su, C.C., Kan, J.C., Chung, C.P., Liu, B.H., 2005. Activation of ERKmitogen-activated protein kinase in human cells by the mycotoxin patulin. Toxicol. Appl.Pharmacol. 207, 103–111.
Yang, J., Hughes, D.P., 1998. A new approach to identifying genotoxic carcinogens: p53induction as an indicator of genotoxic damage. Carcinogenesis 19, 1117–1125.
Yurdun, T., Zehra, Omurtag, G., Ersov, O., 2001. Incidence of patulin in apple juicesmarkets in Turkey. J. Food Protection 64, 1851–1853.