Penetration of Stomata by LiquidsDEPENDENCE ON SURFACE TENSION, WETTABILITY, AND STOMATAL MORPHOLOGY'
Received for publication November 24, 1971
JORG SCHONHERR AND MARTIN J. BUKOVACDepartment of Horticulture, Michigan State University, East Lansing, Michigan 48823
ABSTRACT
Wettability of the leaf surface, surface tension of the liquid,and stomatal morphology control penetration of stomata byliquids. The critical surface tension of the lower leaf surfaceof Zebrina purpusii Bruckn. was estimated to be 25 to 30dyne cm-'. Liquids having a surface tension less than 30 dynecm'i gave zero contact angle on the leaf surface and infiltratedstomata spontaneously while liquids having a surface tensiongreater than 30 dyne cm-' did not wet the leaf surface andfailed to infiltrate stomata. Considering stomata as conicalcapillaries, we were able to show that with liquids giving afinite contact angle, infiltration depended solely on the rela-tionship between the magnitude of the contact angle and thewall angle of the aperture. Generally, spontaneous infiltra-tion of stomata will take place when the contact angle issmaller than the wall angle of the aperture wall. The degree ofstomatal opening (4, 6, 8, or 10 /m) was of little importance.Cuticular ledges present at the entrance to the outer vestibuleand between the inner vestibule and substomatal chamber re-sulted in very small if not zero wall angles, and thus played amajor role in excluding water from the intercellular space ofleaves. We show why the degree of stomatal opening cannotbe assessed by observing spontaneous infiltration of stomataby organic liquids of low surface tension.
tionship between any of these parameters and penetration hasbeen generally poor. In this paper we present a systematicassessment of stomatal penetration by liquids based on thetheory of capillary rise.
THEORY
When a liquid enters a capillary of circular cross-section thepressure difference (P) across the liquid meniscus is representedby equation 1,
P = (R + R (1)
where '/L represents the surface tension of the liquid and R,and RI the principal radii of curvature of the liquid meniscus(1). In a narrow cylindrical capillary the meniscus is a segmentof a sphere; R, and R, are equal and can be expressed in termsof the advancing contact angle (09) and the radius of the capil-lary (r):
R= rcos OA (2)
The advancing contact angle is defined as the contact angleformed by a liquid advancing over a dry surface. Substitutingequation 2 into 1 one obtains:
p= 27L COS OAr (3)
The penetration of liquids into the intercellular air space ofleaves through open stomata has received considerable atten-tion. There is general consensus that pure water does not enterinto the intercellular space through open stomata (4, 22, 27)unless external pressure is applied (6, 7). Some organic liquids,however, are known to penetrate readily, which led to attemptsto estimate the degree of stomatal opening from substomatalpenetration of certain organic liquids (18, 22, 25). Reportsconcerned with the effect of surfactants on promoting penetra-tion into stomata and the substomatal space by aqueous solu-tions are contradictory (3, 4. 11, 14). Surface tension, viscosity,contact angle, diameter of the stomatal aperture, final heightand initial velocity of capillary rise have been implicated incontrolling penetration of liquids into the stomatal pore andsubstomatal chamber (4, 6, 7, 18, 26, 27); however, the rela-
'This study was suported by Public Health Service Grant CC00246 from the National Communicable Disease Center, Atlanta,Georgia, and Food and Drug Administration Grant FD 00223.Michigan Agricultural Experiment Station Journal Article No.5744.
A liquid will rise in the capillary spontaneously only if P ispositive. The sign of P is determined by the term cos O.,; Pbeing positive when OA < 90', zero when 9A = 900 and negativewhen O., > 900. The magnitude of P increases as r decreases.
For a conical capillary (Fig. 1) the angle of the capillary wall(4) must be considered as an additional variable. For a con-verging capillary equation 2 becomes
R =r
sin (0t + OA)
and for a diverging capillary
R=r
sin (02 - 0A)
(4)
(5)
Substituting equation 4 or 5 into 1 one obtains for a conicalconverging capillary
FIG. 1. Schematic drawing of a conical capillary. The wall an-gles (.p and '02) in the converging and diverging portion are identicaland equal to 45°. The menisci of three hypothetical liquids formingcontact angles of 250 (OA'), 450 (OA") or 90° (OA..), respectively, areindicated. Note that in the converging portion (,p + OA) is alwaysless than 1800, the penetrating pressure therefore is positive as in-dicated by the positive curvature of all three menisci, and all threeliquids will rise in this portion of the capillary. At the constrictionthe capillary diverges and only the liquid for which OA'=256 willrise in this portion of the capillary because 02 > 6A' and the pene-trating pressure remains positive. The other two liquids will notpass the constriction as the penetrating pressure is either zero (P2=4A) or negative (q52 < OA.).
OUTER LEDGE
ZONEI
xINNER LEDGESUBSTOMATA L CHAMBER
FIG. 2. Schematic drawing of an Aniaryllis-type stoma. The di-rection of penetration is indicated by the arrow7.
YL sin (-2- OA) (7)
as previously derived by Adam (1).The important difference compared to the cylindrical capil-
lary (equation 3) is that the sign of P is now determined bybothSA and 0. In a converging capillary P will be positive when(01, + OA) < 180° and negative when (sb, +OA) > 180°. In adiverging capillary P will be positive only when 0 < (5b2-OA)< 180° or 02 > OA, negative when 0. < O., and zero when 02=OA4, as shown in Figure 1.
Stomata may be viewed as narrow capillaries having inclinedwalls. In a typical Amaryllis-type stoma (12, 20) (Fig. 2), azone of increasing diameter (zone 1) formed by the outer ledgesis followed by a zone of decreasing diameter approaching aminimum in the pore (zone 2). Towards the substomatalchamber the radius again increases and the wall angle ap-proaches zero at the boundary of the inner cavity and the sub-stomatal chamber (zone 3). Therefore, equation 6 is applicableto zone 2 and equation 7 to zones 1 and 3, while equation 3 isnot applicable. Since stomata are not generally circular conesbut rather elliptic cones or elliptic paraboloids (12, 20, 26),2 r,r2/ (r, + r2) the penetrating pressure can be approximated bysubstituting for r in equations 6 and 7, where r1 and r2 representthe semiminor and semimajor axes, respectively.Any liquid will enter and penetrate a stoma only if P > 0.
Stomatal dimensions generally favor a finite pressure; r1 or r2will never be so large as to make P infinitely small. Thus, instomata the relationship between wall and contact angles be-comes the sole determinant of the sign of P. Entry of liquidsinto stomata and substomatal chambers will be determined bythe conditions prevailing in those zones where the radii in-crease in direction of penetration, that is, in zones 1 and 3(Fig. 2). Here, capillary rise will occur only if OA < 02. In zone2 of stomata, representing a converging capillary, P will bepositive as long as (01 + OA) < 180°, a condition which will bemet by most, if not all, stomata and liquids except mercury.From these considerations the following predictions can be
made and experimentally tested. (a) Any liquid forming a zerocontact angle with the wall of the stomatal aperture will enterand penetrate stomata and substomatal chambers spontane-ously. (b) Any liquid forming a contact angle less than theminimum wall angle in zones 1 and 3 will penetrate into thesubstomatal chamber. (c) Any liquid forming a contact anglegreater than the wall angle will not enter and not penetrate thesubstomatal chamber spontaneously, but can be forced to enterby application of external pressure (Pe,x). The magnitude ofP,X, can be calculated from equation 7 and depends on thecontact angle and stomatal opening, such that (a) when r isconstant P,x, will be proportional to the contact angle and (b)when the contact angle is constant P,, will be inversely propor-tional to stomatal opening.
EXPERIMENTAL
Stomatal Frequency, Distribution, and Morphology. Fre-quency and distribution of stomata on leaves of Zebriniaplurpulsii Briuckn. were determined by scanning electron micros-copy.Whole leaves with open stomata were frozen quickly on Dry
Ice and lyophilized. Small segments (3 X 5 mm) were removedfrom the freeze dried leaves and mounted on a carbon slurryon carbon specimen holders. The leaf tissue was coated withcarbon and gold-palladium (about 200 A) before viewing withan electron microprobe x-ray analyzer (EMX-SM, Applied Re-search Laboratory, Inc.).
For transmission electron microscopy whole leaves withopen stomata were fixed in 10% acrolein at 0 C for 24 hr. Seg-ments (1 x 10 mm) were cut. postfixed with 2% OsO4 in 0.1 Mpotassium phosphate buffer, dehydrated in a graded series ofhexylene glycol, and embedded in a vinylcyclohexane-basedresin according to Spurr (24). Thin sections were cut with aPorter-Blum ultramicrotome (Sorvall MT 2) using a glass knife.A Phillips EM 100 B electron microscope was employed.
Surface Tension Determinations. Solutions differing in sur-face tension were prepared by adding selected surfactants todeionized distilled water having a surface tension of 72.6 dynecmn1. Tween 20 (polyoxyethylene sorbitan monolaurate), Triton
X-100 (octyl phenoxy polyethoxy ethanol), Multi-Film X-77(compounded product of alkyl aryl polyethoxy ethanol and freefatty acids), and Vatsol OT (dioctyl ester of sodium sulfosuc-cinic acid) were used. The surface tension of water, ethanol,butanol, 1, 4-dioxane, and aqueous surfactant solutions wasdetermined from the maximum pull on a platinum ring sus-pended from the left arm of a Chainomatic balance. Measuredvalues were corrected using Harkin's and Jordan's correctiontables (13). The platinum ring was cleaned by flaming betweendeterminations and all glassware was cleaned with concentratedsulfuric acid containing 40 g/liter sodium dichromate. Sincethe surface tension of dilute surfactant solutions changesmarkedly with time, sufficient time (up to 1 hr) was permittedto establish equilibrium. Three determinations were made foreach solution at 21 C. Reproducibility was 0.5 dyne cm-'.
Contact Angle Measurements. Advancing contact angles ofwater and surfactant solutions were determined on the lowersurface of leaves of Z. purpusii. The youngest fully expandedleaves free of surface contamination and defects of greenhouse-grown plants were harvested and placed in the dark to ensurestomatal closure. Leaf strips 5 mm in width were cut perpen-dicular to the midrib. One leaf strip from each of five differentleaves was selected for each concentration and surfactant. Fivecontact angle determinations were made on each strip. Thestrip was placed lower side up on a moist filter paper positionedon the stage of a horizontal microscope. A 2-,ul drop wasformed on the tip of a microsyringe and carefully placed onthe leaf. The drop was slowly built up by adding small amountsof liquid until a constant maximum contact angle was obtained.The height (h) and base width (x) of the drop were measuredwith a horizontal microscope fitted with an eyepiece microme-ter. Measurements were made at 21 C. The contact angle wascalculated using Mack's formula (16), OA = 2 tan-1 (hix). Thisprocedure has been shown to be very accurate for dropletswhich are segments of a sphere, this being the case for smalldroplets where the contact angle is not larger than 900 (5. 16).The largest contact angle observed in this study was 88.1 .
Since contact angles were measured immediately after thedroplet had been built up, the angles reported herein are nottruly equilibrium angles. However, the error was small, as con-tact angles on similar surfaces were found to decrease onlyslightly (2-30) over 1 hr (2). The error due to surface rough-ness was negligible. The roughness factor (of). defined asCOS 0.4 =- COS OA (28). where G'A is measured on a rough and04 on a smooth surface of identical composition, was estimatedto be 1.01. O'A was measured on a positive silicon rubber replicaof the leaf surface and G on a smooth silicon rubber surface.
Determination of Infiltrating Pressure (Pest). To obtainmaximum stomatal opening consistently, leaves of Z. purpusiiwere placed in Plexiglas chambers at 100% relative humidityand CO2-free air with approximately one air change per minute.Temperature was 25 C and light intensity 800 ft-c fromfluorescent (cool white) lamps. Leaves were pretreated for 30min to effect opening, then quickly transferred into the infiltra-tion chamber (Fig. 3), attached to the leaf holder lower surfaceup, the chamber was sealed and pretreatment continued for 10min. During this equilibration time the chamber was partiallyfilled with test solution and submerged in a water bath at 25 C.Following equilibration, the chamber was filled with the testsolution, and pressure was applied to the solution reservoir.Near the point of infiltration the rate of increase of pressurewas approximately 5000 dyne cm-2 min-'. Infiltration occurredalmost instantaneously and was macroscopically apparent asinfiltrated areas appeared dark in incident and bright in trans-mitted light (Fig. 4A). The presence of the test solution in thesubstomatal chamber rather than in the stomatal aperture per
AIR
SOLUTIONPRESSURE
FIG. 3. Infiltration chamber. The leaves were affixed lower sur-face up to the support approximately 1 cm below the cover. Thus,without applied pressure the leaf surface was subjected to a hydro-static pressure of 1 cm of water. This value was taken into accountin determining Pext .
quired to effect infiltration of 10 to 20% of the stomata wasrecorded for each leaf and will be noted as the infiltrating pres-sure (Pest). The pressure required to effect infiltration of only10 to 20% of the stomata was selected to minimize a build upof pressure in the intercellular space resulting from the dis-placement of air by the pentrating solution. This error was con-
sidered small, as the infiltrated solution was not expelled afterthe pressure was released. Three determinations on four leaveseach were made for each test solution. In the experiment toestablish the effect of stomatal opening on Pe,t, the pretreat-ment was conducted in the infiltration chamber and Pest de-termined after prescribed time intervals. Stomatal openingwas estimated following infiltration using a light microscope.
RESULTS
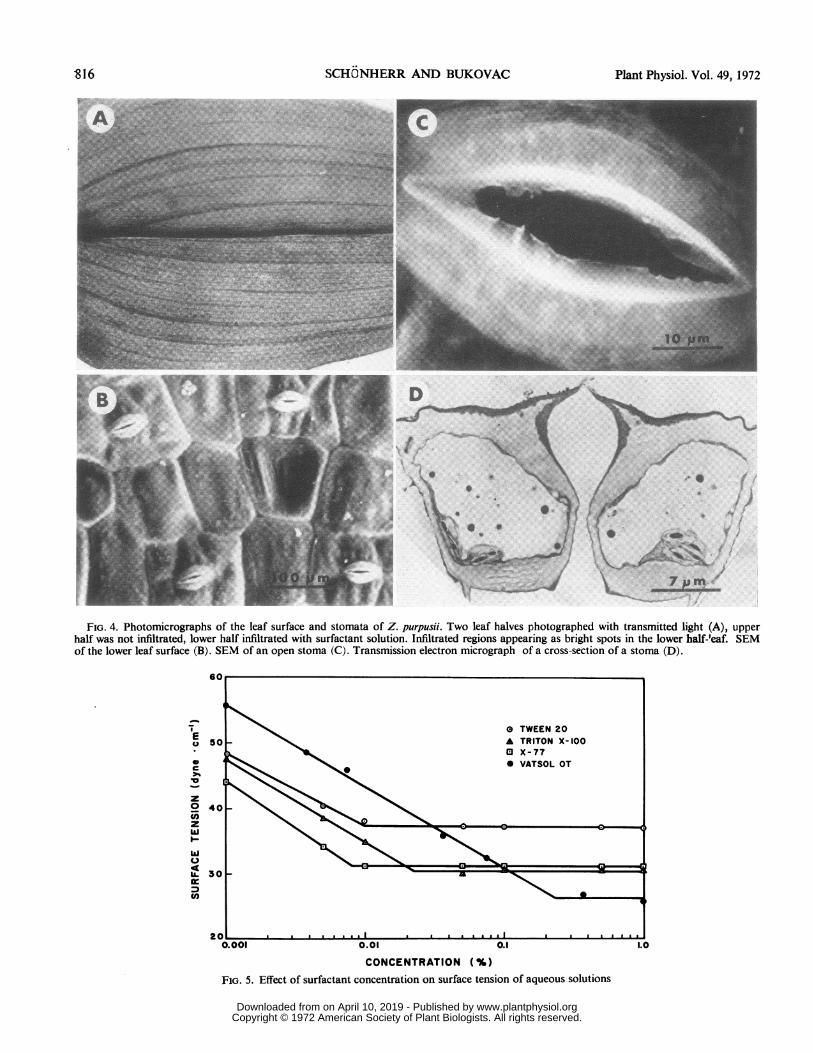
Frequency and Morphology of Stomata. Leaves of Z. pur-
pusii are hypostomatous with stomata arranged in rows parallelto the veins (Fig. 4B). Frequency and size of stomata variedlittle among leaves. There were 625 stomata per cm2 (si =0.25). Stomatal opening between the outer ledges as determinedfrom SEM2 was 2 to 5 ftm (r1) and 14 ,um (r2), respectively, thedimensions of r2 being affected very little by degree of opening.In the SEM the outer ledges appear to project down into theouter cavity (Fig. 4C), however, in cross-section finite wallangles of up to 300 are apparent (Fig. 4D). Small but well de-fined inner ledges are present at the juncture of the inner cavityand the substomatal chamber (Fig. 4D). These estimates ofstomatal opening and wall angles are only approximations sincechanges owing to specimen preparation cannot be precluded.
Surface Tension. The effect of concentration of various sur-
factants on surface tension is illustrated in Figure 5. The lowestsurface tensions obtained for Tween 20, X-77, Triton X-100,and Vatsol OT were 36.6, 30.9, 30.4, and 25.5 dyne cm-',respectively. The point of inflection of each curve coincideswith the critical micelle concentration (19).
Contact Angles. The advancing contact angle of distilled,deionized water on the lower leaf surface of Zebrina was 880.Surfactant solutions of lower surface tension produced smallercontact angles (Fig. 6). Extrapolation of the curves to cos 0.4 =
1.0 gives the critical surface tension (ye) for the lower surface
se was responsible for these infiltration spots. The pressture re -
815Plant Physiol. Vol. 49, 1972
2Abbreviation: SEM: scanning electron micrograph. www.plantphysiol.orgon April 10, 2019 - Published by Downloaded from
FIG. 4. Photomicrographs of the leaf surface and stomata of Z. purpusii. Two leaf halves photographed with transmitted light (A), upperhalf was not infiltrated, lower half infiltrated with surfactant solution. Infiltrated regions appearing as bright spots in the lower half-leaf. SEMof the lower leaf surface (B). SEM of an open stoma (C). Transmission electron micrograph of a cross-section of a stoma (D).
60
*E G\ z TWEEN 20u 50 A TRITON X- 100
O X-77* VATSOL OT
240~~~~
0)ZI.-
U)
CONCENTRATION (%)FIG. 5. Effect of surfactant concentration on surface tension of aqueous solutions
of the Zebrina leaf of 25 to 30 dyne cm-'. The critical surfacetension of a solid surface is defined as that liquid surface ten-sion above which all liquids show finite contact angles (2).Thus, only liquids with a surface tension less than 30 dynecm'i will wet (O., = 0) the lower surface of Zebrina leaves. Ofthe test solutions used only Vatsol OT at concentrations greaterthan 0.1% resulted in zero contact angle (Fig. 5).
Infiltration. Solutions having 5/L > 30 dyne cm-' did notpenetrate through open stomata and into the substomatalchamber spontaneously. Only Vatsol OT solutions having YL <30 dyne cm' infiltrated leaves spontaneously as did ethanol(21.4 dyne cm-') and butanol (23.7 dyne cm-'), while 1 ,4-di-oxane (32.4 dyne cm-') did not.
Solutions having _YL > 30 dyne cm-' could be forced intothe substomatal chamber by applying external pressure. Peltwas directly proportional to the contact angle formed on thelower leaf surface (Fig. 7). A straight line relationship was ob-tained in the range where the Zisman plot was linear (30-80').The line fitted by inspection extrapolates to zero pressure atapproximately 170, indicating that solutions forming O.A = 170would theoretically penetrate open stomata of Zebrina leaves.These data can be interpreted to indicate a limiting wall anglein either zone 1 or 3 of 17°. However, an insufficient numberof determinations at small contact angles makes this extrapola-tion only tentative.
The pressure required to effect infiltration of leaves with asolution forming a contact angle of 630 decreased with increas-ing periods of illumination, that is with increasing stomatalopening (Fig. 8). A minimum value for P,et was observed after30 to 40 min when Pe,,t became time independent.
DISCUSSION
Good qualitative agreement between theoretical predictionsand experimental results was obtained. Only liquids having asurface tension less than the critical surface tension as meas-ured on the lower leaf surface of Z. purpusii penetrated intostomata and the substomatal chamber spontaneously. Thecritical surface tension of 25 to 30 dyne cm-' observed forthe surface of Zebrina leaf cuticle indicates that mainly -CH.and -CH2- groups are exposed at the surface. The criticalsurface tension of pure -CH, and -CH2--surfaces has beenreported to be 25 and 31 dyne cm-', respectively (23). Thesedata are in good agreement with our current knowledge of thecomposition of cutin and epicuticular waxes (14). The failureof liquids with a surface tension greater than 30 dyne cm-' topenetrate stomata may be interpreted to mean that the chemi-
-J
0
z4
I.-
49
z0
w
z
0
20 30 40 50 60 70SURFACE TENSION (dyne-cm-')
0
0
z-c
0
-
a.
06
10
FIG. 6. Contact angles formed on the lower surface of Z. pur-
pusii leaves by surfactant solutions of different surface tensions(Zisman plot).
zso £ RTO -0* 00_ VIGIIV"lX-77 To t * VATSOL OT30E
U)
( 40 -z
520
0 lo 20 30 40 50 60 70 6o 90
CONTACT ANGLE (degrees)
FIG. 7. Relationship between contact angle and pressure re-quired to infiltrate stomata of Z. purpusii.
Z 100
E 90Oto
:"IJ(c 80coQCl
60
; 50
40I0 20 30 40
TIME ( min)50 60
FIG. 8. Effect of illumination period (800 ft-c) of Z. purpusiileaves on pressure required to infiltrate stomata with a Tween 20solution forming a contact angle of 63°. Microscopic examinationof the leaves established that stomatal aperture increased duringthe first 20 to 30 min after which no further change was observed.
cal characteristics of the surface of the cuticle inside thestomatal aperture are similar to those on the leaf surface, sincethe critical surface tension is determined almost entirely by thenature of the chemical groups at the surface of the solid. Re-ports of a more polar nature of the cuticle over guard cells ad-jacent to the stomatal pore as indicated by staining behavior(17, 26) do not necessarily contradict our conclusion because ofthe limited resolution of the light microscope. From our datathere would appear to be very few hydroxyl groups exposed atthe surface, as -CHg--CHOH- surfaces have been reportedto have a critical surface tension between 35 and 42 dyne cm'
(21).The failure of liquids having surface tensions considerably
larger than 30 dyne cm-' to penetrate indicates that the wall
FIG. 9. Maximum infiltrating pressures calculated from theequation P = yL(rl + r2) (r, r2)'1 sin (02 - OA). The value for r2 (14,um) was selected based on measurements of the semimajor axis(see text). The values for r, (4, 6, 8, and 10 ,um) were selected torepresent stomata of varying degrees of opening. Contact anglevalues corresponding to varying surface tension values were takenfrom Figure 6.
angles in zones 1 and 3 must be quite small or zero. This was
confirmed by microscopic observations where a very small orzero wall angle can be estimated in zone 3, largely because ofthe presence of the inner ledges. For zone 1 the microscopicevidence is less conclusive, for in cross-section small wall anglesare evident, while SEM would indicate that the ledges are pro-jected downward resulting in a negative wall angle. This dis-crepancy may be due to unknown effects of sample prepara-tion. The infiltration data do not clarify this point, since theydo not discriminate between entry of liquids into the stomatalaperture and into the substomatal chamber.
Liquids having a surface tension greater than 30 dyne cm-'and, thus, a finite contact angle on the leaf surface could beforced into the intercellular space by application of pressure.The dependence of P,xt on contact angle (Fig. 7) and stomatalopening (Fig. 8) was as predicted; the Pe,, was directly propor-tional to 0A (at constant stomatal opening) and inversely pro-portional to the mean stomatal radius (at constant contactangle). A semiquanitative comparison can be made. A solutionhaving a finite contact angle will enter a stoma up to thepoint where the wall angle is equal to the contact angle andwhere P equals zero. This is most likely to occur near the outeror inner ledges. External pressure must then be applied to forcethe liquid through that portion of the stomatal aperture whereP is negative, that is, where the wall angle is smaller than thecontact angle, and up to that point where P is again positive.Assuming limiting wall angles of zero and 10'. P,., valueswere calculated for various values of r, and r2 and plotted as afunction of 04 (Fig. 9). For this purpose corresponding valuesfor -, and OA were taken from Figure 6. There was a good fitbetween the observed P,,, and calculated values when r,equaled 10 ,tm and r, 14 f,m. The estimate for r2 appears good;however, an estimate of 10 ,pm for r, appears slightly larger
than that observed in the morphological studies. This is not toosurprising when taking into account the approximations madein making these comparisons. The qualitative agreement be-tween theory and experimental results was remarkably close.The data in Figure 7 demonstrate that stomatal pores cannot beassumed to be cylindrical and, therefore, earlier attempts toexplain penetration of liquids on the basis of equation 3 werenot successful (22, 26, 27).The biological significance of the geometry of stomata and
in particular the presence of outer and inner ledges must berecognized. The sudden increase in stomatal aperture withdepth in zones 1 and 3 results in extremely small if not zerowall angles and, therefore, this factor becomes limiting in pene-tration of liquids into stomata and substomatal chambers. Thelow free energy surface of the cuticle resulting in large contactangles contributes to this effect. It must be stressed, however,that the large contact angle alone would not prevent infiltrationof the intercellular space if stomatal apertures were cylindrical,for the contact angle of water on most leaf surfaces is smallerthan 900 (5, 8, 9, 15). With stomata, the effects of small wallangles and a low energy surface combine to present a formida-ble barrier to penetration of liquids with high surface tensionsuch as water or aqueous solutions.Our findings also provide an explanation why in earlier at-
tempts to estimate stomatal opening from stomatal infiltrationby organic liquids, only liquids with very low surface tension,such as, xylene (29.0 dyne cm-'), benzene (28.8), ethanol (22.3),or di(n-alkyl)ethers (25-28.4 dyne cm-') penetrated (18, 2',25). A binary mixture of isobutanol and glycerol (22.6-44.1dyne cm-1) was used by Schorn (22) who found that stomatalpenetration was rarely observed with a mixture of 29.7 dynecm-' and a mixture having 32.9 dyne cm-1 failed to infiltrateany of the leaves tested. This observation is in excellent agree-ment with ours on Zebrina leaves where the critical surfacetension was 25 to 30 dyne cm-'. Therefore, in these earlier in-filtration experiments, most likely only liquids forming zerocontact angle penetrated, which explains why the infiltrationmethod indicated only whether or not stomata were closed, butdid not indicate the degree of opening (22). This is evidentfrom equation 7 showing that occurrence or nonoccurrence ofstomatal penetration is determined by the term sin (02- 0.4)and not by r (within the limits of stomatal dimensions). On theother hand, the pressure required to force a liquid into stomatahaving 0. > 02 is related to stomatal opening (equations 6, 7,Fig. 9) as observed by Froeschel (6, 7). Thus, it is theoreticallypossible but cumbersome to determine degree of stomatal open-ing from Pext measurements, but not by observation of spon-taneous penetration by liquids having zero contact angle.
Entry of foliar sprays into stomatA c;-n also be assessed. Gen-erally, penetration into stomata and substomatal chambers willoccur when the surface tension of the spray liquid is equal orless than the critical surface tension of the leaf surface. Thecritical surface tension of most CutiCular surfaces will be in theneighborhood of 30 dyne cm-' which is the value for a-CH.- surface. Few commercially available surfactants willreduce the surface tension of aqueous solutions below 30 dynecm-n. Vatsol OT. Tergitol 7 (sodium sulfate derivative of 3.9-diethyl tridecanol-6), and Aerosol AY (diamyl ester of sodiumsLIlfosuccinic acid) being among the most effective. There isgood evidence that penetration of stomata and substomatalchambers is achieved by Vatsol OT solutions (4. 1 1) and thesedata are in agreement with our observations. Solutions formingsmall contact angles (<30°) require very little pressure to affectpenetration of the substomatal chamber, a few cm of waterwill suffice (Fig. 7). Thus, care must be exercised in interpret-ing data collected by submerging leaves into solutions whenstudying stomatal penetration (4).
Acknowledgments-We thank Dr. Klaus Raschke and Dr. R. H. Schwende-man, Michigan State University, for helpful discussions and for reviewing themanuscript.
LITERATURE CITED
1. ADAM, N. K. 1948. Principles of penetration of liquids into solids. Disc.Faraday Soc. 3: 5-11.
2. BERNETT, M. K. AND W. A. ZissleN. 1959. Relation of wettability byaqueous solutions to the surface constitution of low energy solids. J.Phys. Chem. 63: 1241-1246.
3. CURRIER, H. B., E. R. PICKERING, AND C. L. Foy. 1964. The relation ofstomatal penetration to herbicidal effects using fluorescent dyes as tracers.Weeds 12: 301-303.
4. DYBING, C. D. AND H. B. CURRIER. 1961. Foliar penetration by chemicals.Plant Physiol. 36: 169-174.
5. EBELING, W. 1939. The role of surface tension and contact angle in theperformance of spray liquids. Hilgardia 12: 665-698.
6. FROESCHEL, P. AND P. CHAPMAN. 1951. A new method of measuring thesize of the stomatal apertures. Celiule 54: 233-250.
7. FROESCHEL, P. 1953. Das Druckstomatometer, ein neuer pflanzenphysiologi-scher Apparat zur Messung der Apertur der Stomata. Cellule 56: 63-70.
8. FOGG, G. E. 1948. Adhesion of water to the external surface of leaves. Disc.Faraday Soc. 3: 162-166.
9. FOGG, G. E. 1947. Quantitative studies on the wetting of leaves by water.Proc. Royal Soc. B. 134: 503-522.
10. Foy, C. L. 1964. Review of herbicide penetration through plant surfaces.J. Agr. Food Chem. 12: 473-477.
11. GREENE, D. W. 1969. Factors influencing the foliar penetration of naph-thaleneacetic acid and naphthaleneacetamide into leaves of pear (Pyruscommunis L.). Ph.D. thesis. Michigan State University, East Lansing.
12. GUTTENBERG, H. V. 1959. Die physiologische Anatomie der Spaltoffnungen.In: W. Ruhland, ed., Handbuch d. Pflanzenphysiologie, Bd. XVII/1.Springer Verlag, Berlin. pp. 399-414.
13. HARKINS, W. D. AND H. F. JORDAN. 1930. A method for the determinationof surface and interfacial tension from the maximum pull on a ring. J.Amer. Chem. Soc. 52: 1751-1772.
OMATA BY LIQUIDS 819
14. HULL, H. M. 1970. Leaf structure as related to absorption of pesticides andother compounds. Residue Rev. 31: 1-155.
15. LINSKENS, H. F. 1950. Quantitative Bestimmung der Benetzbarkeit vonBlattoberflachen. Planta 38: 591-600.
16. MACK, G. L. 1935. The determination of contact angles from measurementsof the dimensions of small bubbles and drops. J. Phys. Chem. 40: 159-167.
17. MAERCKER, U. 1965. Beitrage zur Histochemie der Schliesszellen. Proto-plasma 60: 173-191.
18. MOLIscH, H. 1912. Das Offen- und Geschlossensein der Spaltoffnungenveranschaulicht durch eine neue Methode (Infiltrationsmethode). Z. Bot.4: 106-122.
19. Osipow, L. I. 1964. Surface Chemistry. Reinhold Publishing Corp., NewYork. pp. 163-231.
20. PISEK, A., H. KNAPP, AND J. DITTERSTORFER. 1970. Maximale 6ffnungsweiteund Bau der Stomata, mit Angaben uiber ihre Grosse und Zahl. Flora 159:459-479.
21. RAY, B. R., J. R. ANDERSON, AND J. J. SCHOLZ. 1958. Wetting of polymersurfaces. I. Contact angles of liquids on starch, amylose, amylopectin,cellulose and polyvinyl alcohol. J. Phys. Chem. 62: 1220-1227.
22. SCHOR-X, M. 1929. Untersuchungen ilber die Verwendbarkeit der Alkohol-fixierungs- und der Infiltrationsmethode zur Messung von Spaltoff-nungsweiten. Jahrb. Wiss. Bot. 71: 783-840.
23. SHAFRIaN, E. G. AND W. A. ZISMAN. 1960. Constitutive relations in thewetting of low energy surfaces and the theory of the retraction methodof preparing monolayers. J. Phys. Chem. 64: 519-524.
24. SPURR, A. R. 1969. A low viscosity epoxy resin embedding medium forelectron microscopy. J. Ultrastruct. Res. 26: 31-43.
25. STALFELT, M. G. 1916. tJber die Wirkungsweise der Infiltrationsmethodevon Molisch und einige andere Versuche mit Derselben. Svensk. Bot.Tidskr. 10: 37-46.
26. TURRELL, F. M. 1947. Citrus leaf stomata. Structure, composition andpore size in relation to penetration of liquids. Bot. Gaz. 108: 476-483.
27. URSPRUNG, A. 1925. Uber das Eindringen von Wasser und anderen Fliussig-keiten in Interzellularen. Beih. Bot. Zentralbl. 41: 15-40.
28. WENZEL, R. N. 1936. Resistance of solid surfaces to wetting by water. Ind.Eng. Chem. 28: 988-994.