Proc. Nati. Acad. Sci. USA Vol. 89, pp. 10006-10010, November 1992 Immunology Phenotypic and functional immaturity of human umbilical cord blood T lymphocytes (transplantatlon/tolerance/T-celi differentiation) DAVID T. HARRIS*t, MICHAEL J. SCHUMACHERt, JEFFREY LOCASCIO*, FRANCOISE J. BESENCON*, GEORGE B. OLSON*, DOMINICK DELUCA*, LEWIS SHENKER§H, JUDITH BARD*, AND EDWARD A. BOYSE* *Department of Microbiology and Immunology, *Steele Memorial Children's Research Center, Department of Pediatrics, §Department of Obstetrics and Gynecology, University of Arizona Health Sciences Center, Tucson, AZ 85724 Contributed by Edward A. Boyse, June 22, 1992 ABSTRACT Successful implementation of bone marrow transplantation for hematopoletic reconstitution is limited by the lack of suitably IELA-matched donors and by the occur- rence of graft-versus-host disease that frequently accompanies this procedure. Recent clinical reports have implied that the use of umbilical cord blood as a source of transplantable stem cells may solve these problems. To date, definitive experiments have not been performed to assess the immunological potential of T cells found in umbilical cord blood, which could mediate graft-versus-host disease. In the present study we have ob- served that umbilical cord blood contains T lymphocytes that appear to be phenotypically immature. In addition, umbilical cord blood lymphocytes appeared to be functionally immature as shown by minimal responses to stimulation with interleukin 2, phytohemagglutinin, or alloantigens. Thus, umbilical cord blood may be more suitable for allogeneic transplantation than bone marrow in that these cord blood cells may not be as capable of mediating graft-versus-host disease. Bone marrow transplantation plays an integral part in the treatment of various cancers and inherited blood disorders. The ideal donor for bone marrow transplantation is an identical twin, the next best donor being an HLA-matched sibling. As the number of HLA-identical siblings is limited, the use of HLA-matched allogeneic (unrelated) or HLA- haploidentical parental bone marrow donors frequently of- fers the only chance for bone marrow transplantation. Find- ing a suitable unrelated donor has the highest probability of success when the donor population is ethnically matched, due to racial HLA differences, linkage disequilibrium, and haplotype frequencies. Since the current bone marrow donor registries are composed primarily of Western European Cau- casians, it is extremely difficult to find potential donors for ethnic minorities such as Blacks, Hispanics, and Native Americans. Mismatching between donor and recipient (in- evitable to some degree with all except identical twins) frequently leads to complications, the most serious compli- cation being graft-versus-host disease (GVHD). GVHD pa- thology and a requirement for immunosuppressive therapy results in prolonged immunological dysfunction, which con- tributes significantly to posttransplant mortality (1). GVHD can be reduced by the elimination of mature T cells from the bone marrow graft prior to transplantation (2), although this procedure has been implicated in decreased engraftment and an increased incidence of recurrent malignancy (3, 4). Thus, it is apparent that the use of unrelated, HLA-matched bone marrow for transplantation is far from optimal. Recently, considerable interest has arisen as to the use of human umbilical cord blood (HUCB) as a source of stem cells for marrow replacement when familial HLA-matched bone mar- row donors are not available. Several successful HUCB transplants have been performed (5-8). Thus, HUCB ap- pears to be a source of transplantable stem cells suitable for the treatment of diseases that normally require bone marrow transplantation. However, the use of HUCIB for the trans- plantation of HLA-identical, nonrelated (allogeneic) children or adults remains to be established. Previous investigators have reported that HUCB con- tained cells that expressed surface antigens characteristic of immature lymphocytes (9-12). However, the ability to make inferences from these previous data is limited either by the small HUCB sample sizes that have been examined or by the incomplete phenotypic data that have been reported in these studies. Conflicting results have been reported with regard to the functional maturity of T cells isolated from HUCB (13-17). In the current study we have evaluated the immu- nological potential of HUCB cells, as assessed by various phenotypic and functional assays performed on ""200 HUCB samples, 50 of which have been examined in detail. MATERIALS AND METHODS Collection of HUCB and Peripheral Blood Lymphocytes (PBLs). HUCB samples were obtained from normal full-term vaginal deliveries under the auspices of the Department of Obstetrics and Gynecology, University Medical Center, Uni- versity of Arizona. The cord blood was collected with a heparinized 60-ml syringe after delivery of the infant and ligation of the cord, prior to expulsion of the placenta. Heparinized PBL samples were obtained from healthy vol- unteer donors aged 18-35 years. Both HUCB and PBL samples were centrifuged on Ficoll/Hypaque gradients (twice for the HUCB samples) to obtain mononuclear cells. Flow Cytometry. Phenotypic analyses of mononuclear cell populations were performed by one- and two-color flow cytometry using a Becton Dickinson FACStar Plus flow cytometer. A minimum of 20,000 gated events were analyzed for each sample. Mixed Lymphocyte Reactions (MLRs). To assess prolifer- ative capacity, MLRs were set up in 96-well plates. Re- sponder cells (10W) were stimulated as indicated in the figure Abbreviations: GM-CSF, granulocyte/macrophage-colony-stimu- lating factor; GVHD, graft-versus-host disease; GVL, graft-versus- leukemia; HUCB, human umbilical cord blood; IFN, interferon; IL, interleukin; LAK, lymphokine-activated killer; LDCC, lectin- dependent cell-mediated cytotoxicity; MLR, mixed lymphocyte re- action; NK, natural killer; PHA, phytohemagglutinin; RT-PCR, reverse transcriptase-polymerase chain reaction; TCR, T-cell anti- gen receptor; TNF, tumor necrosis factor. tTo whom reprint requests should be addressed. %Deceased, May 9, 1991. 10006 The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Transcript
Proc. Nati. Acad. Sci. USAVol. 89, pp. 10006-10010, November 1992Immunology
Phenotypic and functional immaturity of human umbilical cordblood T lymphocytes
DAVID T. HARRIS*t, MICHAEL J. SCHUMACHERt, JEFFREY LOCASCIO*, FRANCOISE J. BESENCON*,GEORGE B. OLSON*, DOMINICK DELUCA*, LEWIS SHENKER§H, JUDITH BARD*,AND EDWARD A. BOYSE**Department of Microbiology and Immunology, *Steele Memorial Children's Research Center, Department of Pediatrics, §Department of Obstetrics andGynecology, University of Arizona Health Sciences Center, Tucson, AZ 85724
Contributed by Edward A. Boyse, June 22, 1992
ABSTRACT Successful implementation of bone marrowtransplantation for hematopoletic reconstitution is limited bythe lack of suitably IELA-matched donors and by the occur-rence of graft-versus-host disease that frequently accompaniesthis procedure. Recent clinical reports have implied that the useof umbilical cord blood as a source of transplantable stem cellsmay solve these problems. To date, definitive experiments havenot been performed to assess the immunological potential of Tcells found in umbilical cord blood, which could mediategraft-versus-host disease. In the present study we have ob-served that umbilical cord blood contains T lymphocytes thatappear to be phenotypically immature. In addition, umbilicalcord blood lymphocytes appeared to be functionally immatureas shown by minimal responses to stimulation with interleukin2, phytohemagglutinin, or alloantigens. Thus, umbilical cordblood may be more suitable for allogeneic transplantation thanbone marrow in that these cord blood cells may not be ascapable of mediating graft-versus-host disease.
Bone marrow transplantation plays an integral part in thetreatment of various cancers and inherited blood disorders.The ideal donor for bone marrow transplantation is anidentical twin, the next best donor being an HLA-matchedsibling. As the number of HLA-identical siblings is limited,the use of HLA-matched allogeneic (unrelated) or HLA-haploidentical parental bone marrow donors frequently of-fers the only chance for bone marrow transplantation. Find-ing a suitable unrelated donor has the highest probability ofsuccess when the donor population is ethnically matched,due to racial HLA differences, linkage disequilibrium, andhaplotype frequencies. Since the current bone marrow donorregistries are composed primarily of Western European Cau-casians, it is extremely difficult to find potential donors forethnic minorities such as Blacks, Hispanics, and NativeAmericans. Mismatching between donor and recipient (in-evitable to some degree with all except identical twins)frequently leads to complications, the most serious compli-cation being graft-versus-host disease (GVHD). GVHD pa-thology and a requirement for immunosuppressive therapyresults in prolonged immunological dysfunction, which con-tributes significantly to posttransplant mortality (1). GVHDcan be reduced by the elimination of mature T cells from thebone marrow graft prior to transplantation (2), although thisprocedure has been implicated in decreased engraftment andan increased incidence of recurrent malignancy (3, 4). Thus,it is apparent that the use of unrelated, HLA-matched bonemarrow for transplantation is far from optimal. Recently,considerable interest has arisen as to the use of human
umbilical cord blood (HUCB) as a source of stem cells formarrow replacement when familial HLA-matched bone mar-row donors are not available. Several successful HUCBtransplants have been performed (5-8). Thus, HUCB ap-pears to be a source of transplantable stem cells suitable forthe treatment of diseases that normally require bone marrowtransplantation. However, the use of HUCIB for the trans-plantation of HLA-identical, nonrelated (allogeneic) childrenor adults remains to be established.
Previous investigators have reported that HUCB con-tained cells that expressed surface antigens characteristic ofimmature lymphocytes (9-12). However, the ability to makeinferences from these previous data is limited either by thesmall HUCB sample sizes that have been examined or by theincomplete phenotypic data that have been reported in thesestudies. Conflicting results have been reported with regard tothe functional maturity of T cells isolated from HUCB(13-17). In the current study we have evaluated the immu-nological potential of HUCB cells, as assessed by variousphenotypic and functional assays performed on ""200 HUCBsamples, 50 of which have been examined in detail.
MATERIALS AND METHODSCollection of HUCB and Peripheral Blood Lymphocytes
(PBLs). HUCB samples were obtained from normal full-termvaginal deliveries under the auspices of the Department ofObstetrics and Gynecology, University Medical Center, Uni-versity of Arizona. The cord blood was collected with aheparinized 60-ml syringe after delivery of the infant andligation of the cord, prior to expulsion of the placenta.Heparinized PBL samples were obtained from healthy vol-unteer donors aged 18-35 years. Both HUCB and PBLsamples were centrifuged on Ficoll/Hypaque gradients(twice for the HUCB samples) to obtain mononuclear cells.Flow Cytometry. Phenotypic analyses of mononuclear cell
populations were performed by one- and two-color flowcytometry using a Becton Dickinson FACStar Plus flowcytometer. A minimum of 20,000 gated events were analyzedfor each sample.Mixed Lymphocyte Reactions (MLRs). To assess prolifer-
ative capacity, MLRs were set up in 96-well plates. Re-sponder cells (10W) were stimulated as indicated in the figure
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Proc. Natl. Acad. Sci. USA 89 (1992) 10007
legend, and at the end of 72 hr the cultures were incubatedwith [methyl-3H]thymidine [ICN; 1 ,uCi (37 kBq) per well] forthe final 6-8 hr of culture. Incorporation of the radiolabel intoacid-precipitable DNA was determined by harvesting of thecultures with an automated cell harvester and scintillationcounting (Packard). Data were calculated as stimulationindices, determined by (mean cpm of the stimulated culture)/(mean cpm of the unstimulated control culture). Two allo-geneic human tumor cell lines were used as stimuli: IM9, aB-cell lymphoma (HLA class I and II-positive), and HSB, ahuman T-cell leukemia (HLA class I-positive).
Generation of Cytotoxic Cells and Cytotoxicity Assays. Re-sponder lymphocytes were generated in 5- to 7-day bulkcultures with the indicated stimuli. Cytotoxic effector cellresponses were determined in a standard 4-hr 51Cr-releaseassay. Data were calculated as lytic units at the 20%o specificlysis level (LU20) (18). Human erythroleukemic cell lineK562 was used to assess natural killer (NK) cell-mediatedcytotoxicity. NK cell-resistant cell lines IM9 (human B-celllymphoma) and P815 (murine mastocytoma) were used todetermine cytotoxicity mediated by lymphokine-activatedkiller (LAK) cells. Addition of phytohemagglutinin (PHA) tothe cytotoxicity assays was used to measure total cytotox-icity in the cultures by lectin-dependent cell-mediated cyto-toxicity (LDCC).Reverse Transcriptase-Polymerase Chain Reaction (RT-
PCR) and Southern Blot Analysis. Lymphokine productionwas determined by RT-PCR methodology (19). In brief, totalRNA was extracted (20), and 1 ,ug was denatured for 5 minat 65°C and then chilled on ice. First-strand cDNA wassynthesized according to the manufacturer's directions(Clontech) and was either used immediately or stored at 4°Cfor PCR analysis. For these analyses, PCR mixture (Perkin-Elmer/Cetus), 1.25 units of Taq polymerase (Perkin-Elmer/Cetus), and 0.5 AM 3' and 5' PCR primers were added to 5id of first-strand DNA. Lymphokine-specific PCR primerswere obtained from Clontech, as part of the MAPPing kit.Reaction mixtures were incubated for 1 min at 95°C, followedby 30 cycles of 1 min at 94°C, 2 min at 60°C, and 3 min at 72°C,with a final extension step for 7 min at 72°C.PCR products were analyzed by electrophoresing an ali-
quot of the products (one-fourth of the total) in an agarosegels and blotting to a Nylon 66 Plus membrane (Hoefer)according to the Southern method. Blots were hybridizedwith 32P-labeled oligonucleotide probes (40-mers, OncogeneSciences, Manhasset, NY) specific for the internal sequencefor each set of lymphokine primers. Actin was used as aninternal control to assure that equivalent mRNA could bedetected for each condition. Each PCR also was performedwith external positive and negative controls provided byClontech.
RESULTSPhenotypic Analysis of T Lymphocytes from HUCB. Ap-
proximately 200 HUCB samples were examined, 50 ofwhichwere analyzed in detail (Table 1). Overall, there was asignificantly smaller percentage of T cells (CD3+) in HUCBmononuclear cells than was found in PBLs (P < 0.01). Themajority of HUCB T cells expressed the CD3/TCR-a/13phenotype, as assessed by two-color flow cytometric analy-sis, while T cells expressing the TCR-y/6 phenotype wereseen at the same frequency as found in PBLs (1-3%).However, HUCB contained small populations of CD3-8+and CD3-7+ cells that were not found in PBLs. Ifthe CD3-8+cells were subtracted from the total CD8+ percentage, theCD4/CD8 ratio of HUCB was slightly higher than that seenwith adult PBLs (2.4 for HUCB versus 1.7 for PBLs). MostHUCB T cells were of the naive phenotype, as shown by theobservation that =90% of HUCB T cells expressed the
53 ± 20HUCB and PBL mononuclear cells were isolated and analyzed by
flow cytometry. The mean percentage ofcells positive for each oftheindicated antigens is shown, as well as the standard deviation foreach population of cells. The data were derived from independentanalysis of 50 HUCB and 13 PBL samples. Data were analyzed byt test, as reported in the text. TCR, T-cell antigen receptor.
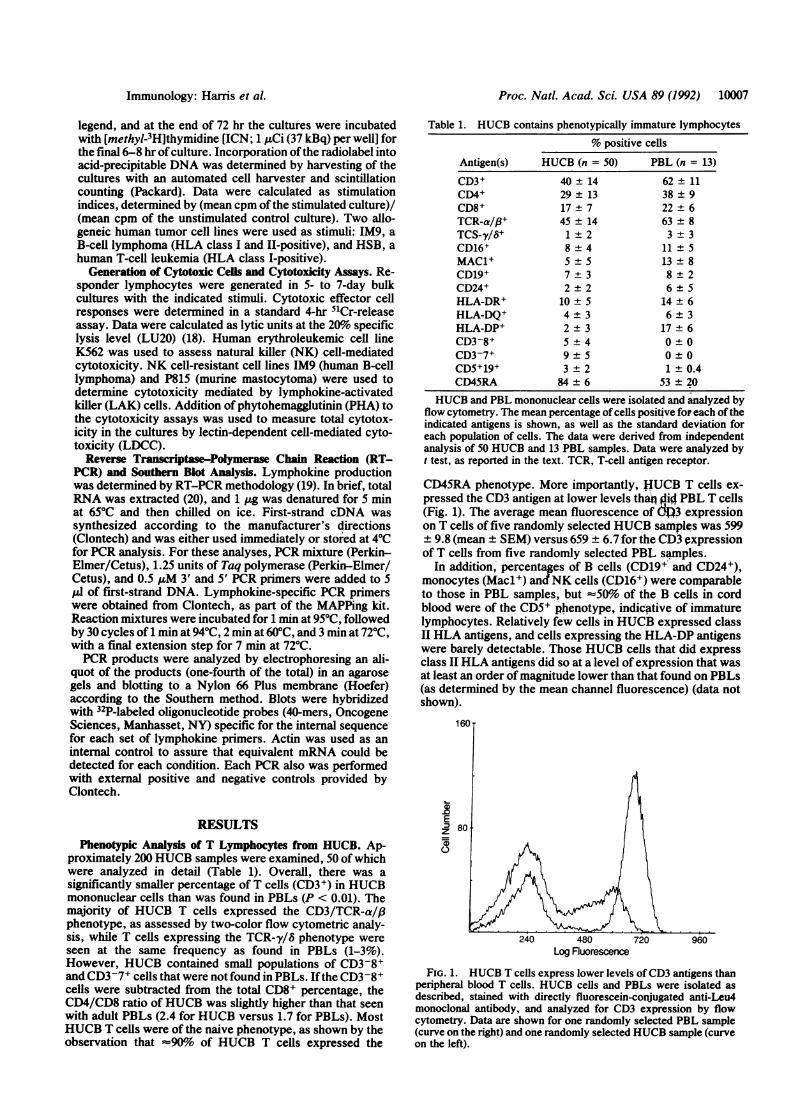
CD45RA phenotype. More importantly, JfUCB T cells ex-pressed the CD3 antigen at lower levels than jlid PBL T cells(Fig. 1). The average mean fluorescence of d>3 expressionon T cells of five randomly selected HUCB samples was 599+ 9.8 (mean ± SEM) versus 659 ± 6.7 for the CD3 expressionof T cells from five randomly selected PBL samples.
In addition, percentages of B cells (CD19+ and CD24+),monocytes (Macl+) andNK cells (CD16+) were comparableto those in PBL samples, but s50% of the B cells in cordblood were of the CD5+ phenotype, indicative of immaturelymphocytes. Relatively few cells in HUCB expressed classII HLA antigens, and cells expressing the HLA-DP antigenswere barely detectable. Those HUCB cells that did expressclass II HLA antigens did so at a level of expression that wasat least an order ofmagnitude lower than that found on PBLs(as determined by the mean channel fluorescence) (data notshown).
160
E:180
240 480 720 960Log Fluorescence
FIG. 1. HUCB T cells express lower levels of CD3 antigens thanperipheral blood T cells. HUCB cells and PBLs were isolated asdescribed, stained with directly fluorescein-conjugated anti-Leu4monoclonal antibody, and analyzed for CD3 expression by flowcytometry. Data are shown for one randomly selected PBL sample(curve on the right) and one randomly selected HUCB sample (curveon the left).
Immunology: Harris et al.
Proc. Natl. Acad. Sci. USA 89 (1992)
;I Il
FIG. 2. HUCB lymphocytes do not generate cytotoxic cells inculture. HUCB lymphocytes were stimulated for 5-7 days in vitrowith either interleukin 2 (IL-2, 1000 units/ml) or PHA (10 jug/ml), or
PHA (2 pg/ml) plus IL-2 (100 units/ml). Freshly isolated HUCBlymphocytes also were analyzed prior to culture. At the end of theculture period the cells were harvested and tested for lytic activity ina standard 4-hr microcytotoxicity assay. The target cells used wereK562 (NK-sensitive; open bars), IM9 or P815 (NK-resistant; solidbars), and IM9 or P815 plus PHA (to determine lectin-dependentcytotoxicity; cross-hatched bars). Data are presented as lytic unitsobtained at the 20% specific lysis level (LU20). Data are shown asmean ± standard deviation of 17 independent experiments utilizing17 different cord blood samples.
Functional Analysis of T Lymphocytes from HUCB. HUCBwas analyzed immediately upon isolation for the ability ofthecells to mediate cytolysis. No lytic activity was detectable infresh HUCB cells, neither NK cell-like lysis (as measuredagainst K562 targets) nor T-cell lytic activity (as measured byLDCC assays using PHA) (Fig. 2). However, upon overnightor 5- to 7-day culture with high doses of IL-2 (1000 units/ml),nonspecific lytic activity rapidly developed. LAK cell activ-ity did not appear to develop, as shown by the inability of theHUCB effector cells to lyse NK-resistant target cells (eitherIM9 or P815 tumor cells) without the addition of lectin.Culture ofHUCB cells with mitogen (PHA) did not result inthe induction of increased levels of lytic activity as demon-strated by the inability of the effector cells to lyse NK-sensitive target cells or to mediate LDCC. However, additionof IL-2 to mitogen-stimulated HUCB cells induced the de-velopment of NK-like lytic effector cells (Fig. 2). Freshlyisolated PBLs expressed NK activity (mean of 30 lytic unitsversus K562 targets) that developed into LAK-like cytolysisupon culture with IL-2 (mean of 192 lytic units versus K562and 26 lytic units versus IM9). The addition of mitogen ormitogen in combination with IL-2 to PBLs resulted in theexpression of both highly lytic NK- and LAK-like effectorcells (data not shown).To better understand the inability ofHUCB to develop into
cytotoxic effector cells, the cultured cells were analyzed byflow cytometry (Table 2). Under each of the culture condi-
tions (IL-2, PHA, or PHA plus IL-2), similar percentages ofCD3+ cells were found at the end of the culture period forboth HUCB and PBL. However, in each instance more CD4+cells than CD8+ cells were found in the HUCB cultures, as
compared with the PBL culture. The percentage of cellsexpressing the TCR-a/(3 phenotype was not significantlydifferent between the two sources of effector cells. Further,the percentage of CD3-CD16+ NK cells in the cultures alsodid not differ greatly between HUCB and PBLs. However,culture of HUCB cells with each of the stimuli (particularlyIL-2) resulted in the expansion of an effector cell populationthat expressed both CD16 and TCR-a/P. The levels ofTCR-8/y+ cells were equivalent in the two populations ofeffector cells under each of the culture conditions, generally<1% of the total cells recovered (data not shown).The functional maturity ofHUCB cells was further tested by
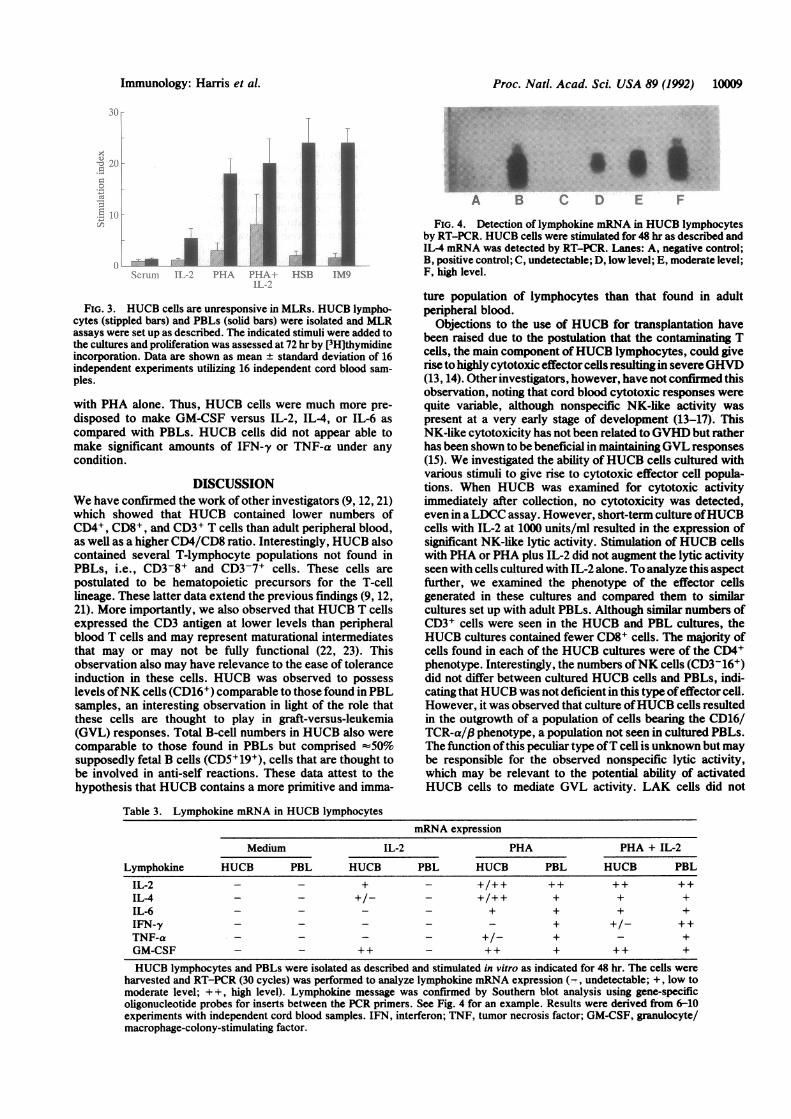
the ability to proliferate in response to stimulation with HLA-,disparate stimulator cells (Fig. 3). Neither HUCB cells norPBLs proliferated in response to autologous serum, indicatingthat neither sera contained growth factors. HUCB cells re-sponded minimally to stimulation with IL-2 or PHA, althoughthe combination of these two stimuli did evoke a somewhatminor proliferative response. Furthermore, stimulation ofHUCB cells with either of two different allogeneic humantumor cell lines expressing HLA class I (HSB, a T-cellleukemia) or class I and II (IM9, a B-cell lymphoma) antigensdid not result in proliferative responses that were abovebackground. PBLs were able to proliferate strongly in re-sponse to these stimuli. Thus, HUCB did not appear to containcells that were capable of expressing mature T-cell functions.The ability of HUCB cells to make lymphokines under
each of the conditions utilized was examined in an attempt tounderstand why HUCa cells appeared to be functionallyimpaired. RT-PCR analysis was employed for this assess-ment and an example is shown in Fig. 4. As shown in Table3, HUCB cells did not spontaneously produce mRNA en-coding lymphokines (stimulation with medium alone). UnlikePBLs, however, the addition of IL-2 to HUCB cells resultedin the production of low levels of IL-4 mRNA, moderatelevels ofIL-2 mRNA, and large amounts ofGM-CSFmRNA.The addition of mitogen to HUCB resulted in moderate levelsofIL-2, IL-4, and IL-6 messages, in addition to large amountsof GM-CSF message. Under these conditions TNF-a mes-sage was on occasion barely detectable in some experiments.Stimulation of PBLs with PHA resulted in moderate to highlevels ofmRNA for each of the lymphokines. Stimulation ofHUCB cells with PHA plus IL-2 did not alter the levels of thelymphokine mRNAs detected after stimulation with PHApone, although infrequently IFN-y mRNA was barely de-tectable in some experiments. Stimulation ofPBLs with PHAplus IL-2, however, resulted in elevated levels of IFN-ymRNA, although the levels of the other lymphokine mes-sages remained comparable to those seen after stimulation
Table 2. Phenotypic characterization of cultured HUCB cells% positive cells
HUCB and PBL mononuclear cells were isolated as described and cultured with either IL-2 (1000 units/ml), or PHA (10hg/ml), or PHA (2 p4g/ml) plus IL-2 (100 units/nil) for 5-7 days. The cells were harvested and analyzed for phenotype byflow cytometry. The mean percentage ofeach cell population is shown as well as the standard deviation for each population.The data were derived from independent analysis of 20 HUCB and 13 PBL samples.
10008 Immunology: Harris et al.
A PI IA I I..
Proc. Natl. Acad. Sci. USA 89 (1992) 10009
4:::
A''A B
N itrm 1-II P PI I I-N tL HSB IM9HI. -:
FIG. 3. HUCB cells are unresponsive in MLRs. HUCB lympho-cytes (stippled bars) and PBLs (solid bars) were isolated and MLRassays were set up as described. The indicated stimuli were added tothe cultures and proliferation was assessed at 72 hr by [3H]thymidineincorporation. Data are shown as mean ± standard deviation of 16independent experiments utilizing 16 independent cord blood sam-
ples.
with PHA alone. Thus, HUCB cells were much more pre-
disposed to make GM-CSF versus IL-2, IL4, or IL-6 as
compared with PBLs. HUCB cells did not appear able tomake significant amounts of IFN-y or TNF-a under anycondition.
DISCUSSIONWe have confirmed the work of other investigators (9, 12, 21)which showed that HUCB contained lower numbers ofCD4+, CD8+, and CD3+ T cells than adult peripheral blood,as well as a higher CD4/CD8 ratio. Interestingly, HUCB alsocontained several T-lymphocyte populations not found inPBLs, i.e., CD3-8+ and CD3-7+ cells. These cells arepostulated to be hematopoietic precursors for the T-celllineage. These latter data extend the previous findings (9, 12,21). More importantly, we also observed that HUCB T cellsexpressed the CD3 antigen at lower levels than peripheralblood T cells and may represent maturational intermediatesthat may or may not be fully functional (22, 23). Thisobservation also may have relevance to the ease of toleranceinduction in these cells. HUCB was observed to possesslevels ofNK cells (CD16+) comparable to those found in PBLsamples, an interesting observation in light of the role thatthese cells are thought to play in graft-versus-leukemia(GVL) responses. Total B-cell numbers in HUCB also werecomparable to those found in PBLs but comprised -50%
supposedly fetal B cells (CD5+19+), cells that are thought tobe involved in anti-self reactions. These data attest to thehypothesis that HUCB contains a more primitive and imma-
*
C D E F
FIG. 4. Detection of lymphokine mRNA in HUCB lymphocytesby RT-PCR. HUCB cells were stimulated for 48 hr as described andIL-4 mRNA was detected by RT-PCR. Lanes: A, negative control;B, positive control; C, undetectable; D, low level; E, moderate level;F, high level.
ture population of lymphocytes than that found in adultperipheral blood.
Objections to the use of HUCB for transplantation havebeen raised due to the postulation that the contaminating Tcells, the main component ofHUCB lymphocytes, could giverise to highly cytotoxic effector cells resulting in severeGHVD(13, 14). Other investigators, however, have not confirmed thisobservation, noting that cord blood cytotoxic responses werequite variable, although nonspecific NK-like activity waspresent at a very early stage of development (13-17). ThisNK-like cytotoxicity has not been related toGVHD but ratherhas been shown to be beneficial in maintainingGVL responses(15). We investigated the ability ofHUCB cells cultured withvarious stimuli to give rise to cytotoxic effector cell popula-tions. When HUCB was examined for cytotoxic activityimmediately after collection, no cytotoxicity was detected,even in aLDCC assay. However, short-term culture ofHUCBcells with IL-2 at 1000 units/ml resulted in the expression ofsignificant NK-like lytic activity. Stimulation ofHUCB cellswith PHA or PHA plus IL-2 did not augment the lytic activityseen with cells cultured with IL-2 alone. To analyze this aspectfurther, we examined the phenotype of the effector cellsgenerated in these cultures and compared them to similarcultures set up with adult PBLs. Although similar numbers ofCD3+ cells were seen in the HUCB and PBL cultures, theHUCB cultures contained fewer CD8+ cells. The majority ofcells found in each of the HUCB cultures were of the CD4+phenotype. Interestingly, the numbers ofNK cells (CD3-16+)did not differ between cultured HUCB cells and PBLs, indi-cating thatHUCB was not deficient in this type ofeffector cell.However, it was observed that culture ofHUCB cells resultedin the outgrowth of a population of cells bearing the CD16/TCR-a/f phenotype, a population not seen in cultured PBLs.The function ofthis peculiar type ofT cell is unknown but maybe responsible for the observed nonspecific lytic activity,which may be relevant to the potential ability of activatedHUCB cells to mediate GVL activity. LAK cells did not
Table 3. Lymphokine mRNA in HUCB lymphocytesmRNA expression
HUCB lymphocytes and PBLs were isolated as described and stimulated in vitro as indicated for 48 hr. The cells wereharvested and RT-PCR (30 cycles) was performed to analyze lymphokine mRNA expression (-, undetectable; +, low tomoderate level; + +, high level). Lymphokine message was confirmed by Southern blot analysis using gene-specificoligonucleotide probes for inserts between the PCR primers. See Fig. 4 for an example. Results were derived from 6-10experiments with independent cord blood samples. IFN, interferon; TNF, tumor necrosis factor; GM-CSF, granulocyte/macrophage-colony-stimulating factor.
Immunology: Haffis et al.
Proc. Natl. Acad. Sci. USA 89 (1992)
develop in the HUCB cultures, as the resulting effector cellswere not able to lyse the NK-resistant IM9 and P815 targetswithout the addition of lectin.Because the functional activities of cell-mediated lymphol-
ysis and MLR are not necessarily correlated in fetal lympho-cytes (15), it was necessary to analyze this aspect of theHUCB immunological potential. Several studies have shownthat HUCB responses in alloantigen-stimulated MLRs wereless than one-quarter of the responses observed with adultPBLs (16, 17). We analyzed HUCB cells for the ability toproliferate in response to a variety of stimuli, includingalloantigen. HUCB T cells responded poorly to mitogen andshowed high levels of autoinduced proliferation (data notshown) in the MLR, which presumably could be a function ofthe increased representation of self-reactive clones in theneonatal pool. Further, HUCB cells did not respond as wellas adult PBLs to stimulation with IL-2 or PHA plus IL-2.Significantly, HUCB cells proliferated minimally in responseto stimulation with HLA-disparate allogeneic tumor cellsdiffering at HLA class I (HSB and IM9 tumor cells) and classII (1M9 tumor cells) antigens. These results, in combinationwith the above data from the cytotoxicity and phenotypingassays, indicate that HUCB cells do not respond well tostimulation and that the cells that are generated by thesestimulations primarily are of the NK or NK-like lymphocyteclass. Stimulation of HUCB cells with HLA-disparate tumorcells in MLR does result in the generation of cells expressinghigh levels of nonspecific lytic activity (data not shown).Whether any of these cytotoxic cells can also mediate anti-gen-specific cytolysis is not known.To understand why HUCB was defective in the generation
of cytotoxic cells, the ability of HUCB cells to make lympho-kines was analyzed by the mRNA-based PCR method. Uponstimulation with mitogen, HUCB cells made moderate to highlevels of IL-2 and IL-4 mRNAs but no IFN-y or TNF-amRNA. HUCB cells made primarily GM-CSF mRNA, whichmay reflect the predisposition of these cells to make hemato-poietic growth factors at this immature stage of development.PBLs, on the other hand, made moderate to high levels of eachof these lymphokine messages. As the production of IFN-yand TNF-a is related to the development of cytotoxicity, thisfinding may explain why HUCB did not generate cytotoxiccells in response to mitogenic stimulation. Why HUCB cellscannot undergo proliferative expansion to the levels seen withPBLs is unclear, since HUCB cells can make IL-2 mRNAwhen stimulated with PHA. Possibly, HUCB cell proliferationwas inhibited by IL-4 that was secreted, which could have hadan antagonistic effect on the growth of the T cells. Interest-ingly, although stimulation of PBLs with IL-2 does not resultin the production of any of the lymphokine mRNAs tested,IL-2 stimulation of HUCB cells resulted in the production ofIL-2, IL-4, and GM-CSF mRNAs, encoding hematopoieticgrowth factors. Thus, activation ofHUCB cells with IL-2 priorto transplantation may serve to activate innate GVL responsesand may aid in hematopoietic reconstitution. Thus, in anotherinstance, HUCB cells appeared to be functionally immature,and the inability to produce certain lymphokines may explainthe other aspects of immunological immaturity that wereobserved.
In conclusion, it appears that HUCB may have the poten-tial to be the graft of choice for transplantation, complement-ing conventional bone marrow usage. Establishment of aHUCB stem-cell bank could open new avenues in the pro-curement of donors for both autologous and allogeneic trans-plantation. A bank of cryopreserved cord bloods couldovercome some ofthe present limitations ofthe bone marrowregistry; e.g., stonring HUCB from the large number ofpotential donors available would ensure that the ethnicbalance in the donor pool was automatically maintained.Significantly,.HUCB may be less able to cause GVHD (ref.
8 and these results), which is often a problem when unrelatedbone marrow is used for transplantation. Due to the seriousproblems associated with both GVHD and immunosuppres-sive therapy [among which are organ/tissue failure, bacterialand viral (herpes simplex virus and cytomegalovirus) infec-tions, reactivation of Epstein-Barr virus, neoplasia (leuke-mia and lymphoma, some of which may be due to Epstein-Barr virus reactivation), and severe growth and developmen-tal defects in children] (24), HUCB could have a significantadvantage over bone marrow for transplantation.
We acknowledge the invaluable technical assistance of PattiParker, Debbie Sakiestewa, and Barb Carolus. Further, the assis-tance of Dr. David Chaffin and Ann Booth, R.N., in obtaining thecord blood specimens is gratefully acknowledged. This work wassupported in part by Grant CA39827 from the National CancerInstitute. E.A.B. is an American Cancer Society Research Professor.
1. Witherspoon, R. P., Storb, R., Ochs, H. D., Flournox, N.,Kopecky, K. J., Sullivan, K. M., Deeg, H. J., Sosa, R., Noel,D. R., Atkinson, K. A. & Thomas, E. D. (1981) Blood 58,360-368.
2. Atkinson, K. (1990) Bone Marrow Transplant. 5, 209-226.3. Apperley, J., Jones, L., Hale, G., Waldmann, H., Hows, J.,
Marcus, R. E., Goolden, A. W. G., Gordon-Smith, E. C.,Catovsky, D., Galton, D. A. G., Goldman, J. M., Rombas, Y.& Tsatalas, C. (1986) Bone Marrow Transplant. 1, 53-66.
4. Slavin, S., Ackerstein, A., Naparstek, E., Weiss, L. & Or, R.(1990) Bone Marrow Transplant. 6, 155-161.
5. Broxmeyer, H. E., Gluckman, E., Auerbach, A., Douglas,G. W., Friedman, H., Cooper, S., Hangoc, G., Kurtzberg, J.,Bard, J. & Boyse, E. A. (1990) Int. J. Cell Cloning 8, Suppl. 1,76-91.
6. Gluckman, E., Broxmeyer, H. E., Auerbach, A., Friedman,H., Douglas, G. W., Devergie, A., Esperou, H., Thierry, D.,Socie, G., Lehn, P., Cooper, S., English, D., Kurtzburg, J.,Bard, J. & Boyse, E. A. (1989) N. Engl. J. Med. 321, 1174-1178.
7. Gluckman, E. (1990) in Peripheral Blood Stem CellAutografts,eds. Henon, P. & Wunder, E. (Springer, New York).
8. Broxmeyer, H. E., Douglas, G. W., Hangoc, G., Cooper, S.,Bard, J., English, D., Arny, M., Thomas, L. & Boyse, E. A.(1989) Proc. Natl. Acad. Sci. USA 86, 3828-3832.
9. Foa, R., Giubellino, M. C., Fierro, M. T., Lusso, P. & Fer-rando, M. L. (1984) Cell. Immunol. 89, 194-201.
10. Yeoman, H. & Mellor, A. (1992) Int. Immunol. 4, 59-65.11. Reinherz, E. L., Kung, P. C., Goldstein, G., Levey, R. H. &
Schlossman, S. F. (1980) Proc. NatI. Acad. Sci. USA 77,1588-1592.
12. Griffiths-Chu, S., Patterson, J. A. K., Berger, C. L., Edelson,R. L. & Chu, A. C. (1984) Blood 64, 296-300.
13. Linch, D. C. & Brent, L. (1989) Nature (London) 340,676-677.14. Rayfield, L. S., Brent, L. & Rodeck, C. H. (1980) Clin. Exp.
Immunol. 42, 561-570.15. Granberg, C. & Hirvonen, T. (1980) Cell. Immunol. 51, 13-22.16. Noh, L. M., Khan, M. M. & Melmon, K. L. (1988) Dev.
Comp. Immunol. 12, 177-187.17. Granberg, C., Mannien, K. & Toivanen, P. (1980) Clin. Immu-
nol. Immunopathol. 6, 187-256.18. Pross, H. F., Baines, M. G., Rubin, P., Shragge, P. & Patter-
son, M. S. (1981) J. Clin. Immunol. 1, 51-62.19. Brenner, C. A., Tam, A. W., Nelson, P. A., Engelman, E. G.,
Suzuki, N., Fry, K. E. & Larrick, J. W. (1989) BioTechniques7, 1096-1103.
20. Maniatis, T., Fritsch, E. F. & Sambrook, J. (1982) MolecularCloning:A Laboratory Manual (Cold Spring Harbor Lab., ColdSpring Harbor, NY).
21. Maccario, R., Nespoli, L., Mingrat, G., Vitiello, A., Ugazio,A. G. & Burgio, G. R. (1983) J. Immunol. 130, 1129-1131.
22. Guidos, C. J., Weissman, I. L. & Adkins, B. (1989) Proc. Natl.Acad. Sci. USA 86, 7542-7546.
23. Ramsdell, F., Jenkins, M., Dinh, Q. & Fowlkes, B. J. (1991) J.Immunol. 147, 1779-1785.
24. Kolb, H. J. & Bender-Gotze, C. (1990) Bone Marrow Trans-plant. 6, 61-72.