Phylogeography and genetic structure of the threatenedCanarian Juniperus cedrus (Cupressaceae)
BEATRIZ RUMEU1,2*, PABLO VARGAS2, RUTH JAÉN-MOLINA3, MANUEL NOGALES1
and JULI CAUJAPÉ-CASTELLS3
1Island Ecology and Evolution Research Group (IPNA-CSIC), La Laguna, Tenerife, Canary Islands,Spain 382062Real Jardín Botánico (RJB-CSIC), Madrid, Spain 280143Department of Molecular Biodiversity & DNA Bank, Jardín Botánico Canario ‘Viera y Clavijo’ –Unidad Asociada CSIC, Cabildo de Gran Canaria, Las Palmas de Gran Canaria, Spain 35017
Received 9 July 2013; revised 14 January 2014; accepted for publication 27 March 2014
Isolation is a crucial process in plant evolution, andoceanic archipelagos offer an ideal spatio-temporalframework in which to study the patterns of geneticvariation associated with speciation (Carlquist, 1965;Francisco-Ortega, Jansen & Santos-Guerra, 1996;Emerson, 2002; Whittaker & Fernández-Palacios,2007). The Macaronesian archipelagos harbour alarge number of plant endemics, and the CanaryIslands are particularly well studied in terms of his-torical plant evolution and high levels of diversifica-
tion (see Francisco-Ortega et al., 2000; Vargas, 2007;Caujapé-Castells, 2011 and references therein).
The biota of oceanic islands is certainly unique, butalso susceptible to sudden external changes, as shownby many worldwide examples of plant extinctionsprimarily driven by the impact of human activities(e.g. Cronk, 1997; Caujapé-Castells et al., 2010;Triantis et al., 2010). Continental populations mayshow higher levels of genetic diversity than theirisland congeners (but see Fernández-Mazuecos &Vargas, 2011), feasibly as a result of bottlenecksrelated to founder effects (e.g. Frankham, 1997;Crawford et al., 2001). Insular populations also tendto be small; thus, they are more prone to the adverse*Corresponding author. E-mail: [email protected]
bs_bs_banner
Botanical Journal of the Linnean Society, 2014, 175, 376–394. With 5 figures
effects of genetic drift and, presumably, inbreedingdepression (Ellstrand & Elam, 1993). Overall, thesecircumstances imply a higher genetic risk in islandpopulations than in continental ones (Frankham,1998).
The Macaronesian archipelagos harbour three ofthe 11 juniper species of Juniperus L. section Juni-perus (syn: Juniperus section Oxycedrus Spach)(Cupressaceae) (Adams, 2011). Juniperus brevifolia(Seub.) Antoine is endemic to the Azores, J. cedrusWebb & Berth. to the Canarian archipelago andJ. maderensis (Menezes) R.P.Adams to Madeira(Adams et al., 2010; Adams & Schwarzbach, 2012). Arecent plastid DNA phylogenetic analysis of Junipe-rus section Juniperus suggests that the establishmentof J. brevifolia in the Azores followed a single intro-duction event, probably from Europe (Rumeu et al.,2011a). However, the phylogenetic relationshipsbetween J. cedrus and other juniper species fromMorocco and the western Mediterranean remainelusive.
Located between 27°N and 29°N and 13°W and18°W, the Canaries lie only 96 km off the Africancoast and comprise seven main volcanic islands ofdifferent geological ages (22.0–1.2 Ma; Carracedo &Day, 2002). The current populations of J. cedrus arefragmented and mainly restricted to craggy and inac-cessible areas on La Palma, La Gomera, Tenerife andGran Canaria (Fig. 1) . With an estimated occupancyarea of <30 km2, the total population of this species isc. 600 mature trees (La Palma, N = 250; La Gomera,N = 100; Tenerife, N = 200; Gran Canaria, N = 12;Rumeu et al., 2011b). The species is catalogued asendangered (EN) by IUCN (2013), as near threatened
(NT) in the latest edition of the Spanish Red List ofVascular Flora (Moreno, 2008) and as vulnerable (VU)in the Spanish National Catalogue of endangeredspecies (Real Decreto 139/2011, BOE 46 of 23/02/2011). In the last publication, the conservation strat-egies proposed are seed conservation in germplasmbanks, ex-situ culture, control of herbivory andadequate management of protected areas in La Palma(National Park of Caldera de Taburiente), La Gomera(National Park of Garajonay), Tenerife (National Parkof El Teide) and Gran Canaria (Güigüí SpecialNatural Reserve).
The Canarian juniper is a dioecious, anemophilousand endozoochorous taxon. The fleshy female conesare available to seed dispersers throughout the year(Rumeu et al., 2009a) and are consumed mainly bybirds (Rumeu, Padilla & Nogales, 2009b; Rumeuet al., 2011c), as also reported for its continental con-geners (e.g. Livingston, 1972; Zamora, 1990; Jordano,1993; García, 2001). Despite the lack of informationabout the longevity of J. cedrus, studies on someother congeners indicate that junipers are long-livedtrees that can have life spans of more than 200 years(Ward, 1982). Unlike Pinus canariensis Sweet exSpreng. (the endemic pine species from the CanaryIslands), J. cedrus does not appear to be a resprouter(B. Rumeu, pers. observ.).
Some references indicate a wide distribution ofJ. cedrus in the past, at least in Tenerife and GranCanaria, the islands of occurrence with greater areaand environmental heterogeneity (Höllermann, 1978;Leuschner, 1996; Francisco-Ortega, Santos-Guerra &Bacallado, 2009). Although J. cedrus now occurs inprotected areas, and some of its populations have
Figure 1. The Canary Islands and their maximum geological ages according to Carracedo & Day (2002). Sampledpopulations of Juniperus cedrus are indicated with circles. The number of samples collected in each island is indicatedin parentheses.
attained a moderately large number of individuals, itsvaluable wood has had many industrial uses in thepast, and the currently narrow distribution of thisspecies reflects, to a large extent, the strong pressureexerted by humans after the European colonization ofthe islands in the 15th century. This demographichistory leads us to expect an ailing genetic situation forJ. cedrus; the dramatic reductions in population sizesmay have created severe genetic bottlenecks, wherebythe remaining individuals contain only a small fractionof the original gene pool as a result of genetic drift,which is further accentuated by increasing isolationand decreasing population sizes (Ellstrand & Elam,1993; Young, Boyle & Brown, 1996).
Following studies of the basic reproductive andecological traits of J. cedrus (Rumeu et al., 2009a, b,2011c), we used plastid DNA sequences and nuclearamplified fragment length polymorphism (AFLP) datato: (1) infer the temporal and spatial origin ofJ. cedrus; (2) assess the patterns of genetic diversityand differentiation on each island; and (3) offer usefulguidelines for the implementation of the conservationand management strategies planned for this endan-gered juniper.
MATERIAL AND METHODSPLANT SAMPLING AND DNA EXTRACTION
The sampling was fairly representative of the distri-bution range of J. cedrus and needles were collectedfrom trees on all the islands of occurrence. As a resultof the severe fragmentation in many of its distributionareas and the difficulty of accessing most trees, thenumber of individuals sampled varied widely betweenthe islands: 13 for La Palma, 27 for La Gomera, 27 forTenerife and five for Gran Canaria (Fig. 1). Thesesample sizes are close to the number of accessibletrees in each population targeted and represent 5.2%,27%, 13.5% and 41.6% of all sexually mature individu-als estimated on these islands, respectively (seeRumeu et al., 2011b). All the individuals known on LaPalma, La Gomera, Tenerife and Gran Canaria areconfined to narrow areas (see the ‘Population map’at http://www.demiurge-project.org/matrix_digests/43); therefore, in all cases, our samples can safely beascribed to the same population within their islands ofdistribution. Total DNA samples were extracted fromsilica gel-dried needles using the 2 × cetyltrimethyl-ammonium bromide (CTAB) method (Doyle & Doyle,1987; Palmer et al., 1988). The concentration of totalDNA was measured in an Eppendorf biophotometerand its quality was verified in 1% agarose gels.Aliquots of the DNA extracts have been depositedin the DNA Bank of the Canarian Flora at theJBCVCSIC.
PLASTID DNA MARKERS
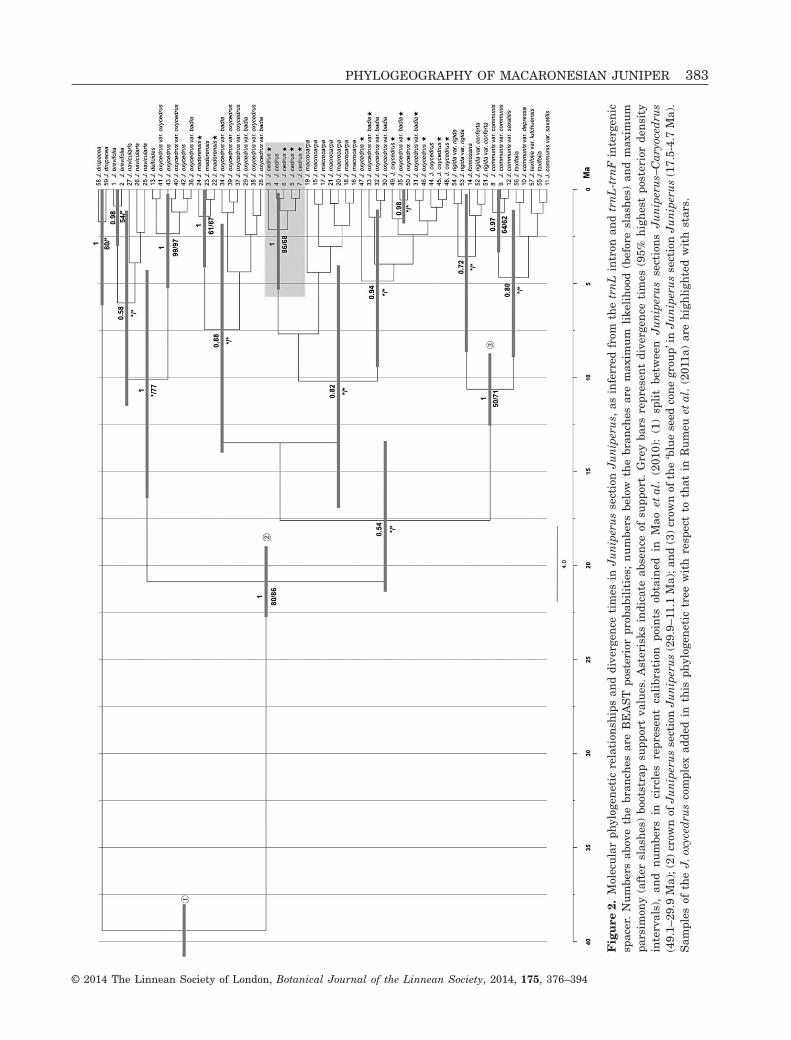
To infer the temporal and spatial origin of J. cedrus,we performed a phylogenetic analysis of Juniperussection Juniperus based on two plastid DNAsequences (trnL intron and trnL-trnF intergenicspacer; Taberlet et al., 1991) which had been used inprevious studies (Mao et al., 2010; Rumeu et al.,2011a). This data matrix was substantially extendedwith four newly sequenced samples of J. cedrus plus11 samples of geographically close species: nine ofJ. oxycedrus L. from Morocco and two of J. maderen-sis from Madeira. Based on previous phylogeneticanalyses of Juniperus (Mao et al., 2010), we alsoadded sequences of two outgroup samples (J. drupa-cea Labill., Juniperus section Caryocedrus Endl.).Table 1 shows the geographical origins of the ingroupand outgroup samples (for details, see SupportingInformation, Table S1). Amplification and sequencingprocedures followed Rumeu et al. (2011a).
To further investigate the putative origin and dif-ferentiation of J. cedrus in the Canary Islandsthrough a phylogeographical analysis, we tested twoto four individuals from different islands for 19plastid DNA regions in an exploratory study based onprevious phylogenetic and phylogeographical analy-ses (Taberlet et al., 1991; Sang, Crawford & Stuessy,1997; Hamilton, 1999; Hwang, Hwang & Lin, 2000;Shaw et al., 2005, 2007; Kress & Erickson, 2007;Fazekas et al., 2008; Provan et al., 2008; Adams,Bartel & Price, 2009). As a result of the lack ofvariation retrieved, we finally sequenced the threeregions previously employed in a study of the Azoreanendemic J. brevifolia (Rumeu et al., 2011a): trnT-trnL(Taberlet et al., 1991), petN-psbM (Adams et al., 2009)and trnS-trnG (Hamilton, 1999). We assembled amatrix with 5–17 individuals per island (dependingon sample availability), plus two samples of J. mac-rocarpa Sibth. & Sm. from Spain and Italy, and 18samples of J. oxycedrus from Morocco and thewestern Mediterranean basin. To investigate thegenetic relationship between J. cedrus and J. mader-ensis, we also included 14 samples from the lattertaxon [Tables 2 and S2 (Supporting Information)].The trnS-trnG region was finally discarded, as it didnot reveal any variability among our samples. Poly-merase chain reaction (PCR) protocols and sequenc-ing followed Rumeu et al. (2011a). All new sequenceshave been deposited in GenBank (see Tables S1 andS2 for accession numbers).
AFLP PROCEDURE
To assess intraspecific genetic diversity in all thesamples of J. cedrus collected, we used AFLP finger-prints, based on their broad application in plantgenetics (Nybom, 2004; Meudt & Clarke, 2007). We
Table 1. Taxon names and origins of the Juniperus samples included in the phylogenetic analysis based on trnL andtrnL-trnF plastid sequences. ‘No’ is the code for sample identification in Figure 2. Taxonomy follows Adams (2011). Varietyis indicated only for the J. oxycedrus specimens determined to that level by the corresponding expert. See Table S1 forvouchers source and GenBank accession numbers
No Taxon Geographical area/locality
1 J. brevifolia Portugal, Azores, Santa Maria, Almagreira2 J. brevifolia Portugal, Azores, Terceira, Malha Grande3 J. cedrus Spain, Canary Islands, La Palma, Pared de Roberto4 J. cedrus Spain, Canary Islands, La Palma, Pared de Roberto5 J. cedrus Spain, Canary Islands, La Gomera, P. N. Garajonay6 J. cedrus Spain, Canary Islands, Tenerife, P. N. del Teide7 J. cedrus Spain, Canary Islands, Gran Canaria, Mña. del Cedro8 J. communis var. communis France9 J. communis var. communis Spain, Granada, Pico Trevenque
10 J. communis var. depressa USA, New Mexico11 J. communis var. saxatilis Pakistan12 J. communis var. saxatilis Georgia, Caucasus13 J. deltoides Turkey14 J. formosana var. mairei China, Gansu15 J. macrocarpa Spain, Cádiz, Barbate, Trafalgar Cape16 J. macrocarpa Spain, Cádiz, Tarifa17 J. macrocarpa Spain, Valencia, El Saler18 J. macrocarpa Spain, Cádiz, Chiclana19 J. macrocarpa Italy, Sardinia, Santa Teresa Gallura20 J. macrocarpa Italy, Sicily, Ragusa21 J. macrocarpa Tunisia, Cap Bon, Sidi Daoud22 J. maderensis Portugal, Madeira, Fajã da Nogueira23 J. maderensis Portugal, Madeira, road Ribeira Brava-Paul da Serra (cultivated)24 J. maderensis Portugal, Madeira, Jardim Botânico (cultivated)25 J. navicularis Portugal, Apostiça26 J. navicularis Portugal, Apostiça27 J. navicularis Portugal, Estremadura28 J. oxycedrus var. badia Spain, Huesca, Sierra de Guara29 J. oxycedrus var. badia Spain, Jaén30 J. oxycedrus var. badia Spain, Jaén31 J. oxycedrus var. badia Morocco, El Ksiba32 J. oxycedrus var. badia Morocco, Kjbel Kelti33 J. oxycedrus var. badia Morocco, Kjbel Kelti34 J. oxycedrus var. badia Morocco, Kjbel Kelti35 J. oxycedrus var. badia Morocco, Marrakech36 J. oxycedrus var. badia Turkey, Gümüshane, Torul37 J. oxycedrus var. oxycedrus Spain, El Penon38 J. oxycedrus var. oxycedrus Spain, El Penon39 J. oxycedrus var. oxycedrus Spain, Balearic Islands, Menorca40 J. oxycedrus var. oxycedrus Greece, Lemos41 J. oxycedrus var. oxycedrus Greece, Lemos42 J. oxycedrus Turkey, Istanbul, Kartal43 J. oxycedrus Greece, Kalavryta, Diakoftó44 J. oxycedrus France45 J. oxycedrus Morocco, Jebel Lekst46 J. oxycedrus Morocco, Jebel Imzi47 J. oxycedrus Morocco, Beni-Mellal, Taghzour48 J. oxycedrus Morocco, High Atlas mountains, Aït el Qâq – Oukaïmedene49 J. oxycedrus Morocco, Asni, Tizi n’Test50 J. oxycedrus Morocco, Agadir, Imouzzer Ida Ou Tane51 J. rigida var. conferta Cultivated (Spain, Pontevedra, Lourizán)52 J. rigida var. conferta Japan53 J. rigida var. rigida Japan54 J. rigida var. rigida Japan55 J. taxifolia Japan56 J. taxifolia Japan, Bonin Island57 J. taxifolia var. lutchuensis Japan
OUTGROUP58 J. drupacea Greece59 J. drupacea Greece
used standard AFLP protocols following Vos et al.(1995), with modifications by García-Verdugo et al.(2009). No explicit data on the genome size ofJ. cedrus are available, but 1C-values (i.e. the amountof DNA in an unreplicated gametic nuclear genome) ofother Juniper spp. are in the range 10.82–12.15 pg(medium-sized genome) (Hizume et al., 2001). Accord-ing to Fay, Cowan & Leitch (2005), this genome sizeis adequate for the application of AFLP and supportsthe expectation of strongly amplified bands combinedwith noisy baselines. Following the recommendationson the number of selective bases in Fay et al. (2005)and the primer combinations used in J. thurifera L.(Terrab et al., 2008), we performed an initial screen-ing of selective primers using 18 primer pairs withthree and four selective nucleotides to test for theirpotential to produce scorable fragments. Of these,four primer pairs (EcoRI/MseI plus ACA/CTCG, ACT/
CTCG, AGG/CTGA, AGG/CTCG) were chosen accord-ing to their reproducibility, levels of polymorphismand previous analyses (Terrab et al., 2008). Two tofour replicates from the restriction-ligation phasewere included in all the subsequent reactions to testreproducibility. The minimum size of the fragmentsscored was 100 bp to minimize the occurrence of frag-ment size homoplasy, which is more likely withshorter fragments (Vekemans et al., 2002). Themaximum size of the scored fragments was 500 bp.The reactions were separated using an ABI 3730automated sequencer. GeneMapper v4.0 (Applied Bio-systems) was used for fragment scoring. The criteriafor choosing AFLP bands followed Terrab et al. (2008).Bands that did not fulfil these criteria were regardedas potentially unreliable and were therefore discardedfrom the final presence/absence matrix for the sake ofdata quality.
Table 2. Plant material of Juniperus used for petN-psbM and trnT-trnL sequencing. N is the number of individualssampled in each locality. H is the corresponding haplotype according to Figure 3. Taxonomy follows Adams (2011). Varietyis indicated only for the J. oxycedrus specimens determined to that level by the corresponding expert. See Table S2 forvoucher source and collection numbers, haplotype number of each tree analysed and the GenBank accession numbers ofeach haplotype found
Taxon Geographical area/locality N H
J. cedrus Spain, Canary Islands, La Palma, Piedra Llana 4 5, 6J. cedrus Spain, Canary Islands, La Palma, Pared de Roberto 9 5, 6, 7J. cedrus Spain, Canary Islands, La Gomera, P. N. Garajonay 17 5, 6J. cedrus Spain, Canary Islands, Tenerife, Riscos de La Fortaleza 6 6, 7J. cedrus Spain, Canary Islands, Tenerife, Siete Cañadas 6 5, 6,J. cedrus Spain, Canary Islands, Tenerife, Mña. del Cedro 3 6J. cedrus Spain, Canary Islands, Tenerife, Ucanca 2 6J. cedrus Spain, Canary Islands, Gran Canaria, Mña. del Cedro 5 6J. macrocarpa Spain, Cádiz, Tarifa 1 2J. macrocarpa Italy, Sardinia, Santa Teresa Gallura 1 2J. macrocarpa Tunisia, Cape Bon, Sidi Daoud 1 2J. maderensis Portugal, Madeira, road Ribeira Brava–Paul da Serra (cultivated) 3 3J. maderensis Portugal, Madeira, road to Pico Arieiro (cultivated) 1 3J. maderensis Portugal, Madeira, Fajã da Nogueira 1 3J. maderensis Portugal, Madeira, Jardim Botânico (cultivated) 5 3, 4J. maderensis Portugal, Madeira, road Santa-Achadas da Cruz (cultivated) 4 3, 4J. oxycedrus Morocco, Anti-Atlas 2 1J. oxycedrus Morocco, Beni-Mellal, Taghzour 1 1J. oxycedrus Morocco, High Atlas mountains, Aït el Qâq – Oukaïmedene 1 1J. oxycedrus Morocco, Asni, Tizi n’Test 1 1J. oxycedrus Morocco, Agadir, Imouzzer Ida Ou Tane 1 1J. oxycedrus var. badia Morocco, Marrakech 1 1J. oxycedrus var. badia Morocco, Jebel Kelti 3 1, 2J. oxycedrus var. badia Morocco, El Ksiba 1 1J. oxycedrus var. badia Spain, Jaén 2 1, 2J. oxycedrus var. badia Spain, Huesca, Sierra de Guara 1 2J. oxycedrus var. oxycedrus Spain, El Penon 2 2J. oxycedrus var. oxycedrus Spain, Balearic Islands, Menorca 1 2
Phylogenetic inference using plastid DNAThe phylogenetic analysis of Juniperus section Juni-perus with the trnL intron and trnL-trnF dataset usedmaximum likelihood (ML), maximum parsimony (MP)and Bayesian inference (BI) approaches. Large indelsfound in the trnL-trnF spacer were manually codedfollowing the ‘simple indel coding’ method describedin Simmons & Ochoterena (2000) and included in theanalyses. The Akaike information criterion (AIC)(Akaike, 1979) was implemented in this dataset usingjModelTest 0.1.1 (Posada, 2008) to determine themodel of sequence evolution that best fits thesequence data (HKY + I, AIC = 1982.7). ML analyseswere performed using RAxML v7.0.3 (Stamatakis,2006) with 100 bootstrap replicates. As this softwareonly implements the GTR evolutionary model, weused the GTRGAMMAI model as the closest to thatsuggested by AIC. Parsimony analyses were run inTNT (Goloboff, Farris & Nixon, 2008) under tradi-tional heuristic search and considering gaps (previ-ously coded as simple indel coding) as a fifthcharacter. We first used the tree bisection–reconnection (TBR) branch swapping algorithm with10 000 replicates (saving two most-parsimonioustrees per replicate). Subsequently, the trees obtainedin the first search were used to start a second heu-ristic search that retained all the best trees. Branchsupport was evaluated using 1000 bootstrap repli-cates and collapsing groups with branch supportvalues of less than 50%.
The BI analysis was used to estimate divergencetimes in Juniperus section Juniperus, including thedate of origin of J. cedrus. We implemented a relaxedmolecular clock approach in BEAST v.1.6.0 to esti-mate phylogenetic relationships and node ages(Drummond et al., 2006; Drummond & Rambaut,2007). The molecular clock analysis was carried outwith two data partitions: (1) the trnL intron andtrnL-trnF intergenic spacer dataset using theHKY + I model following the AIC criterion; and (2)simple indels from partition 1 coded as binary data(binary simple substitution model) using the softwareSeqState 1.4.1 (Müller, 2006). For the temporal cali-bration, we used three divergence times previouslyobtained by Mao et al. (2010) modelled as normaldistributions with standard deviation = 1: (1) the splitbetween Juniperus sections Juniperus–Caryocedrus(49.1–29.9 Ma); (2) the crown of Juniperus sectionJuniperus (29.9–11.1 Ma); and (3) the crown of the‘blue seed cone group’ (BSG) in Juniperus sectionJuniperus (17.5–4.7 Ma). The substitution rate vari-ation was modelled using an uncorrelated log-normaldistribution and a birth–death process (Gernhard,2008) was employed as tree prior. Two Markov chain
Monte Carlo (MCMC) analyses were run for 10million generations, with a sample frequency of 1000and discarding the first 10% generations as burn-in.Analysis with Tracer 1.4 (Rambaut & Drummond,2007) confirmed adequate sample size, with an effec-tive sample size (ESS) of greater than 200. Bothanalyses were combined using LogCombiner 1.4.8,and the trees were then summarized in a maximumclade credibility tree obtained in TreeAnnotator 1.4.8.
Phylogeographical analysis of plastid DNAsequencesTo elucidate the putative origin of J. cedrus and todetect the ancestral haplotypes, the petN-psbM andtrnT-trnL sequences were concatenated into a singlematrix. We then performed a phylogeographicalanalysis based on coalescence theory (Avise, 2000) toinfer the degree of connectivity between the Canarianjuniper and its closest relatives, plus J. maderensisfrom the nearest archipelago (Madeira). The genea-logical relationships among haplotypes were inferredthrough a statistical parsimony method (Templeton,Crandall & Sing, 1992) implemented in TCS 1.21software (Clement, Posada & Crandall, 2000). Themaximum number of differences resulting from singlesubstitutions among haplotypes was calculated with95% confidence limits, treating gaps as missing data.In addition, the analysis of haplotypic diversity wasperformed by treating gaps as the fifth state. Wecalculated the genetic differentiation value (FST)among J. cedrus populations using DnaSP v5(Librado & Rozas, 2009).
AFLP data analysesIntraspecific genetic variability of J. cedrus wasinferred from the AFLP matrix, which was importedfrom a .xlsx file to the software Transformer-4(Caujapé-Castells et al., 2011). This software auto-matically generated the input files needed to run allthe population genetic software used and producedreports with the exclusive fragments per population(Fragexcl) and the fragments exclusively shared perpopulation pair. Transformer-4 was also used tosubmit the AFLP matrix and other relevant items tothe Demiurge information system, where it was peerreviewed and published in the ‘genetic diversitydigest’ coded D-AFLPS-43 (http://www.demiurge-project.org/matrix_digests/D-AFLPS-43).
For each population, the total number of AFLPfragments (Fragtot), usable loci (with <5% missingdata) (Fragusable), polymorphic fragments (Fragpoly) andthe average gene diversity (HD) were estimated usingArlequin 3.5.1.2 (Excoffier & Lischer, 2010), whichwas also used to compute exact tests of pairwisepopulation differentiation. Following Coart et al.(2005), as a result of the variation in sample sizes per
population (5–27), we performed a rarefactionapproach implemented in AFLPdiv V. 1.1 to computethe phenotypic band richness (Br henceforth, with1 = band present and 0 = band absent) expected ateach scored AFLP fragment and the percentage ofpolymorphic loci (PLP) in a standardized sample sizeof five individuals (equal to the smallest populationsample size, on Gran Canaria). Following theseauthors, a locus is considered to be polymorphic at the1% level if 1.01 < Br < 1.99. The proportion of totalgenetic variation attributable to inter-population dif-ferentiation (GST) and total genetic diversity (HT) werecalculated by POPGENE 32 (Yeh et al., 1997). Thenumber of migrants per generation (Nm) was esti-mated using Arlequin 3.5.1.2 in order to determinethe magnitude of gene flow among islands (i.e. popu-lations). To assess the relationship between geo-graphical and genetic distance matrices betweenpopulation pairs, a Mantel test (Mantel, 1967) with9999 permutations was performed using GenAlex6.41 (Peakall & Smouse, 2006).
To investigate the spatial genetic structure of popu-lations, we performed a spatial autocorrelation analy-sis (Sokal & Oden, 1978) which allowed us to assessthe relationship between AFLP genetic distances as afunction of the geographical distances among indi-viduals. We excluded 100 fragments showing frequen-cies of < 0.1 or > 0.9 and seven fragments for whichdata for > 20% individuals were missing. We calcu-lated Moran’s I statistics for the remaining 84 frag-ments using the software PASSaGE V2.0 (Rosenberg& Anderson, 2011), with which we built a preliminarycorrelogram based on 999 non-parametric permuta-tions for ten distance classes, each containingapproximately equal numbers of pairwise compari-sons. From these results, we selected the fragmentsdisplaying significant spatial autocorrelation (N = 45)and constructed a new matrix that was analysed withthe software SPAGeDI 1.3 (Hardy & Vekemans, 2002)to calculate the average values of Moran’s I acrossdistance classes and their standard errors (SEs).
A population assignment test was conducted usingthe software Structure 2.3.2 (Pritchard, Stephens &Donnelly, 2000) to cluster individuals into subpopu-lations. This software assumes a model with K popu-lations (K unknown), each characterized by a set offragment frequencies at each locus. Using the Bayes-ian approach, individuals are probabilisticallyassigned to either a single population or jointly to twoor more populations (if their genotypes indicateadmixture). Following Evanno, Regnaut & Goudet(2005), the presence of a band was coded as 1/(−9),where ‘1’ is the presence of the dominant allele and‘−9’ is a missing value. The absence of a band wascoded as 2/2, indicating that the recessive allele (2)was homozygous. We set the admixture model and the
option of correlated allele frequencies between popu-lations, as this configuration is considered to be themost appropriate in cases of subtle population struc-ture (Falush, Stephens & Pritchard, 2003). Similarly,the program inferred the degree of admixture (alpha)from the data. After several independent runs foreach value of K in a pilot study, we chose a burn-inperiod of 50 000 iterations and 500 000 steps for thelength of the MCMC. We ran ten independent simu-lations of this length for each K to ensure consistentestimates across runs. The possible values of Kranged from ‘1’ to the actual number of sampledpopulations plus ‘3’. We then followed the methoddescribed in Evanno et al. (2005) to calculate the truenumber of populations (K).
RESULTSPHYLOGENETIC RECONSTRUCTION OF JUNIPERUS
SECTION JUNIPERUS AND LINEAGE
DIVERGENCE TIMES
All three analyses using sequences of the trnL intronand the trnL-trnF region supported the monophyly ofJ. cedrus (Fig. 2). The diversification window of theCanarian juniper was estimated at between 5.30 and0.18 Ma, although its probable geographical originwas not consistently supported and only the Bayesian(BEAST) approach weakly pointed (with a posteriorprobability of 0.82) to a closer relationship with abroad sister group formed by J. macrocarpa andJ. oxycedrus.
Our three analyses also supported the monophylyof J. maderensis. The topology of the phylogenetictree did not suggest unequivocally that this taxonoriginated and evolved independently from J. cedrus,because of the low support values. However, the hap-lotype network is congruent with two different originsfrom the continent (see below).
J. CEDRUS PLASTID DIFFERENTIATION AND
HAPLOTYPE NETWORK
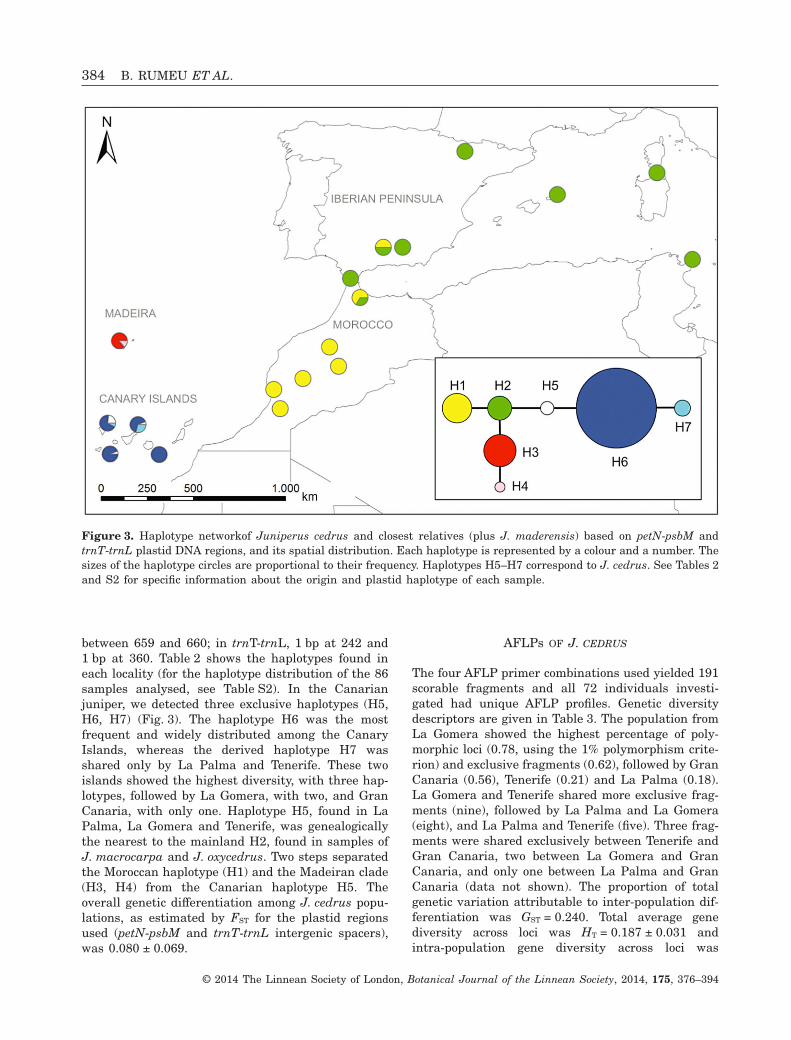
We detected seven haplotypes in J. cedrus and itsrelatives J. maderensis (Madeira), J. macrocarpa andJ. oxycedrus (western Mediterranean basin andMorocco) [Fig. 3, but see Fig. S1 (Supporting Informa-tion) showing the results of the network analysisconsidering gaps as a fifth state, in which 12 haplo-types were retrieved]. Variation in the petN-psbM andtrnT-trnL intergenic spacers was the result of sevennucleotide substitutions and ten gaps, the lengthsand distribution of which in the alignment were asfollows: in petN-psbM, 35 bp between 145 and 179,6 bp between 265 and 270, 19 bp between 280 and298, 5 bp between 431 and 435, 1 bp at 598, 7 bpbetween 620 and 626, 1 bp at position 660 and 2 bp
between 659 and 660; in trnT-trnL, 1 bp at 242 and1 bp at 360. Table 2 shows the haplotypes found ineach locality (for the haplotype distribution of the 86samples analysed, see Table S2). In the Canarianjuniper, we detected three exclusive haplotypes (H5,H6, H7) (Fig. 3). The haplotype H6 was the mostfrequent and widely distributed among the CanaryIslands, whereas the derived haplotype H7 wasshared only by La Palma and Tenerife. These twoislands showed the highest diversity, with three hap-lotypes, followed by La Gomera, with two, and GranCanaria, with only one. Haplotype H5, found in LaPalma, La Gomera and Tenerife, was genealogicallythe nearest to the mainland H2, found in samples ofJ. macrocarpa and J. oxycedrus. Two steps separatedthe Moroccan haplotype (H1) and the Madeiran clade(H3, H4) from the Canarian haplotype H5. Theoverall genetic differentiation among J. cedrus popu-lations, as estimated by FST for the plastid regionsused (petN-psbM and trnT-trnL intergenic spacers),was 0.080 ± 0.069.
AFLPs OF J. CEDRUS
The four AFLP primer combinations used yielded 191scorable fragments and all 72 individuals investi-gated had unique AFLP profiles. Genetic diversitydescriptors are given in Table 3. The population fromLa Gomera showed the highest percentage of poly-morphic loci (0.78, using the 1% polymorphism crite-rion) and exclusive fragments (0.62), followed by GranCanaria (0.56), Tenerife (0.21) and La Palma (0.18).La Gomera and Tenerife shared more exclusive frag-ments (nine), followed by La Palma and La Gomera(eight), and La Palma and Tenerife (five). Three frag-ments were shared exclusively between Tenerife andGran Canaria, two between La Gomera and GranCanaria, and only one between La Palma and GranCanaria (data not shown). The proportion of totalgenetic variation attributable to inter-population dif-ferentiation was GST = 0.240. Total average genediversity across loci was HT = 0.187 ± 0.031 andintra-population gene diversity across loci was
Figure 3. Haplotype networkof Juniperus cedrus and closest relatives (plus J. maderensis) based on petN-psbM andtrnT-trnL plastid DNA regions, and its spatial distribution. Each haplotype is represented by a colour and a number. Thesizes of the haplotype circles are proportional to their frequency. Haplotypes H5–H7 correspond to J. cedrus. See Tables 2and S2 for specific information about the origin and plastid haplotype of each sample.
HD = 0.145 ± 0.026. Average among-population differ-entiation was FST = 0.234 ± 0.080.
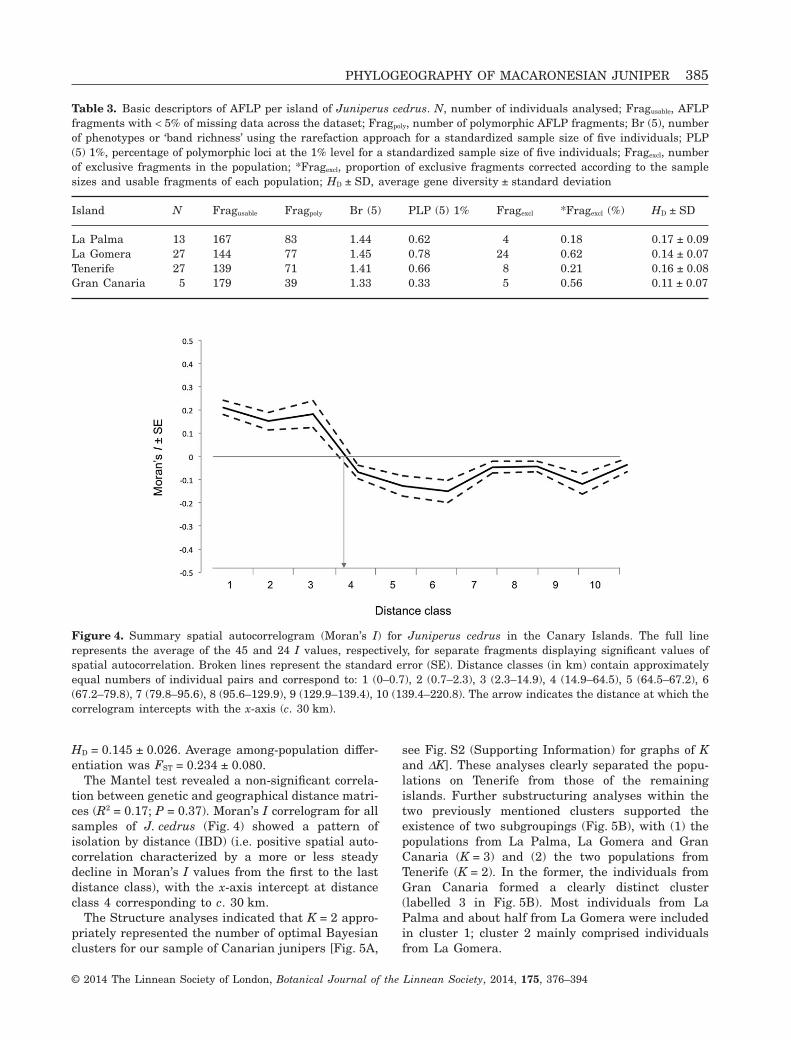
The Mantel test revealed a non-significant correla-tion between genetic and geographical distance matri-ces (R2 = 0.17; P = 0.37). Moran’s I correlogram for allsamples of J. cedrus (Fig. 4) showed a pattern ofisolation by distance (IBD) (i.e. positive spatial auto-correlation characterized by a more or less steadydecline in Moran’s I values from the first to the lastdistance class), with the x-axis intercept at distanceclass 4 corresponding to c. 30 km.
The Structure analyses indicated that K = 2 appro-priately represented the number of optimal Bayesianclusters for our sample of Canarian junipers [Fig. 5A,
see Fig. S2 (Supporting Information) for graphs of Kand ΔK]. These analyses clearly separated the popu-lations on Tenerife from those of the remainingislands. Further substructuring analyses within thetwo previously mentioned clusters supported theexistence of two subgroupings (Fig. 5B), with (1) thepopulations from La Palma, La Gomera and GranCanaria (K = 3) and (2) the two populations fromTenerife (K = 2). In the former, the individuals fromGran Canaria formed a clearly distinct cluster(labelled 3 in Fig. 5B). Most individuals from LaPalma and about half from La Gomera were includedin cluster 1; cluster 2 mainly comprised individualsfrom La Gomera.
Table 3. Basic descriptors of AFLP per island of Juniperus cedrus. N, number of individuals analysed; Fragusable, AFLPfragments with < 5% of missing data across the dataset; Fragpoly, number of polymorphic AFLP fragments; Br (5), numberof phenotypes or ‘band richness’ using the rarefaction approach for a standardized sample size of five individuals; PLP(5) 1%, percentage of polymorphic loci at the 1% level for a standardized sample size of five individuals; Fragexcl, numberof exclusive fragments in the population; *Fragexcl, proportion of exclusive fragments corrected according to the samplesizes and usable fragments of each population; HD ± SD, average gene diversity ± standard deviation
Island N Fragusable Fragpoly Br (5) PLP (5) 1% Fragexcl *Fragexcl (%) HD ± SD
Figure 4. Summary spatial autocorrelogram (Moran’s I) for Juniperus cedrus in the Canary Islands. The full linerepresents the average of the 45 and 24 I values, respectively, for separate fragments displaying significant values ofspatial autocorrelation. Broken lines represent the standard error (SE). Distance classes (in km) contain approximatelyequal numbers of individual pairs and correspond to: 1 (0–0.7), 2 (0.7–2.3), 3 (2.3–14.9), 4 (14.9–64.5), 5 (64.5–67.2), 6(67.2–79.8), 7 (79.8–95.6), 8 (95.6–129.9), 9 (129.9–139.4), 10 (139.4–220.8). The arrow indicates the distance at which thecorrelogram intercepts with the x-axis (c. 30 km).
All phylogenetic reconstructions of Macaronesianjunipers (Rumeu et al., 2011a; this study) suggest,albeit with moderate support (Fig. 2), that the threeMacaronesian junipers represent independent line-ages in the J. oxycedrus complex, pointing to a dis-tinct evolutionary history of J. cedrus relative to theAzorean and Madeiran junipers. This result agreeswith Adams et al. (2010) and Adams & Schwarzbach(2012), who proposed the Madeiran juniper as a dis-tinct species from J. cedrus, and strengthens the pre-diction of multiple colonization events for plantsdisplaying an endozoochorous syndrome (e.g. Olea L.,Hedera L.; Vargas, 2007).
Our phylogenetic analyses, based on a matrixextended with 15 samples of the J. oxycedrus complexwith respect to that in Rumeu et al. (2011a), con-firmed the monophyly of J. cedrus, suggesting a
single seed-mediated colonization event of the Canar-ian archipelago, followed by diversification. The diver-sification period was estimated to be between 5.30and 0.18 Ma, a period relatively recent in comparisonwith the origin of the oldest island of occurrence(Gran Canaria, c. 14.5 Ma). This dating agrees withthe intervals estimated in the Pliocene and Pleisto-cene for many other Canarian endemic lineages, suchas Echium L. (c. 4 Ma, García-Maroto et al., 2009),Sideritis L. (3.3 Ma, Kim et al., 2008), AndrocymbiumWilld. (< 3.5 Ma, del Hoyo, García-Marín &Pedrola-Monfort, 2009) and Lotus L. (2.4 Ma, Ojedaet al., 2012). Nonetheless, the DNA sequence datafailed to suggest a clear sister group relationshiphypothesis for J. cedrus, despite the inclusion of aconsiderable number of juniper samples from thewestern Mediterranean and Morocco in the analyses.The plastid phylogenetic topology (Fig. 2) indicates acloser relationship with populations of J. macrocarpaand J. oxycedrus, mainly distributed in the westernMediterranean. Although the tree in Figure 2 does
Figure 5. Bayesian estimation of the genetic population structure of Juniperus cedrus based on the AFLP dataset. Eachindividual is represented by a thin vertical bar, which is portioned into K coloured segments that represent the estimatedmembership fractions of an individual in K clusters. Horizontal lines on the bottom separate individuals according topopulations (labelled below the figure: P, La Palma; G, La Gomera; C, Gran Canaria; T, Tenerife). A, Juniperus cedrusindividuals from all populations sampled. B, Estimated population substructure based on clusters obtained in (A).Individuals from La Palma, La Gomera and Gran Canaria in the left plot. Individuals from Tenerife in the right plot. Thefigure shown for a given K is based on the highest probability run at that K.
not allow us to hypothesize the ancestral relation-ships between the clades, a Mediterranean origin ofthe Canarian juniper would be consistent with manycolonization histories of Macaronesian island endemicplants (Vargas, 2007; Carine et al., 2010;Caujapé-Castells, 2011). Nevertheless, our resultsindicate that further research is needed to betterunderstand the complex temporal and geographicalorigins of the endemic J. cedrus.
GENETIC DIVERSITY OF THE CANARIAN JUNIPER
With only three plastid DNA haplotypes (four whenconsidering gaps as a fifth state, see Fig. S1), thegenetic diversity of J. cedrus is low in comparison withthat found in the Azorean J. brevifolia using the sameplastid regions (16 haplotypes, Rumeu et al., 2011a) orthe mainland maritime juniper J. macrocarpa [14 hap-lotypes based on three different plastid regions (petG-trnP, trnD-trnT and trnL-trnF), Juan et al., 2012],even though these two species have also sufferedsubstantial reductions in population sizes. A recentdiversification could be responsible for the low haplo-type diversity detected (three haplotypes and nomissing ones). Moreover, the influence of fragmenta-tion has been especially severe in the Canary Islands(and probably much more pronounced than in the caseof J. brevifolia and J. macrocarpa), and the consequentdemographic bottlenecks could also have caused agreater loss of variation (Ellstrand & Elam, 1993).
Our AFLP results showed that the highest percent-age of polymorphic loci occurs on La Gomera, theisland of distribution with the smallest area andwhere only two plastid DNA haplotypes weredetected. Although the haplotypic diversity forJ. cedrus is small overall, the low haplotypic values inLa Gomera are possibly related to a more recentphylogeographical origin of the populations on thisisland, whereas the high AFLP genetic diversityrefers to the much lower impact of anthropogenicactivities and other factors discussed below.
La Gomera also has the highest number of exclu-sive AFLP fragments (Table 3) and shares more frag-ments exclusively with the rest of the islands ofoccurrence. These findings suggest three non-exclusive explanations. First, La Gomera may haverepresented a significant source of genetic variationfor the other islands, given its intermediate geo-graphical position in the range of distribution ofJ. cedrus. Second, felling could have more dramati-cally affected the much larger islands of GranCanaria and Tenerife, so that the genetic distinctnessof La Gomera may be attributed to the preservation ofmostly relictual variation that could have beenpresent in the large populations that previouslyexisted on these two islands (Höllermann, 1978;
Leuschner, 1996; Francisco-Ortega et al., 2009).Third, the old age of La Gomera (c. 11 Ma) and itsgreater geological stability (the latest volcanic activityis dated in the Pliocene, Carracedo & Day, 2002) mayhave allowed for longer diversification periods. Thelack of precise historical information prevents us fromassessing further the relative importance of thesepossibilities on the current genetic diversity ofJ. cedrus.
La Palma, the second smallest island of occurrence(and the most recent geologically), hosts the threehaplotypes detected with plastid sequences and showsthe highest genetic diversity levels (HD) (Table 3). Inthis case, we believe that the most feasible explana-tion of these results is a much lower incidence offelling (Parsons, 1981), allowing genetic diversity tobe maintained as a result of a less dramatic influenceof human-induced genetic bottlenecks.
THE EFFECT OF ISOLATION ON THE GENETIC
STRUCTURE OF J. CEDRUS
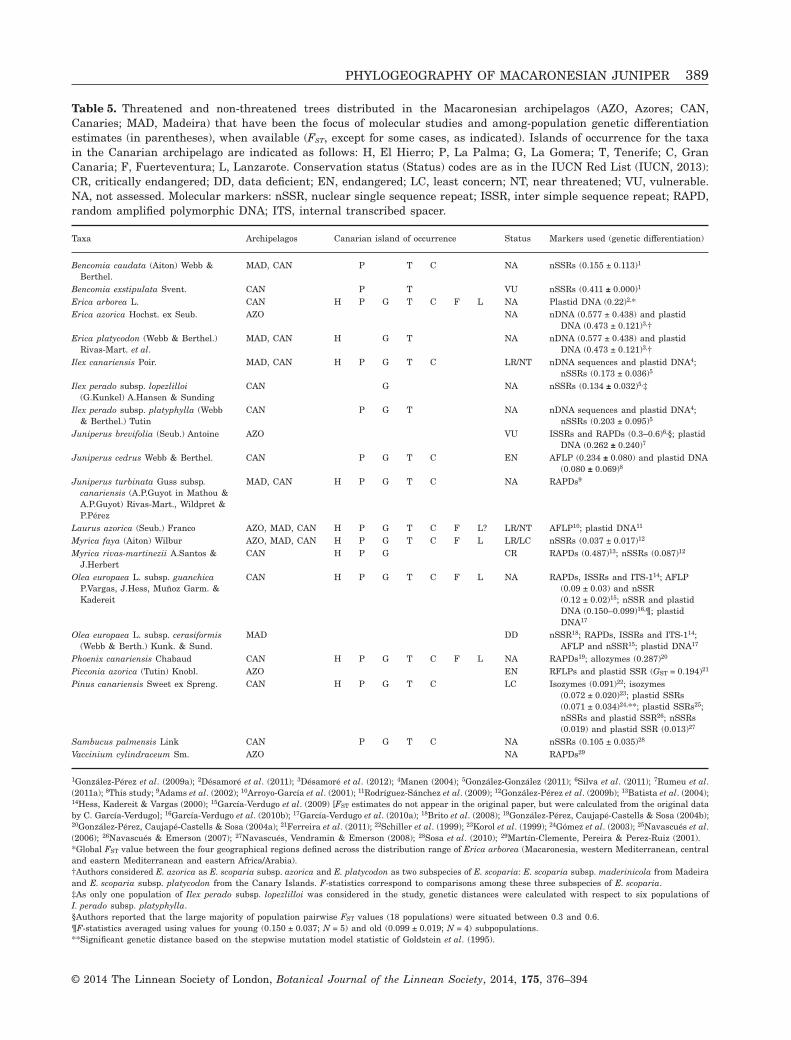
Recent anthropogenic fragmentation may have con-tributed to the relatively high levels of genetic differ-entiation among the populations of J. cedrus. Theaverage among-population differentiation estimatedwith AFLP (FST = 0.234 ± 0.080) is much higher thanthat found for: (1) most Macaronesian trees for whichsimilar data are available (Table 5); and (2) non-endangered trees with a widespread distribution inthe Canaries, such as Pinus canariensis C.Sm. ex DC.(Pinaceae, FST = 0.091, at most) or Myrica faya Aiton(Myricaceae, FST = 0.037). Possibly, the heterogeneityof markers used in the different literature sources ofTable 5 accounts for part of the FST differencesbetween J. cedrus and other Macaronesian trees com-mented above. However, these data highlight thatJ. cedrus still maintains considerable genetic varia-tion levels, despite the forces that have acted toreduce population genetic variation and foster frag-mentation. Furthermore, the AFLP genetic differen-tiation between island pairs (Table 4) is also large(especially for the comparisons involving Gran
Table 4. Population pairwise FST estimates based on the191 AFLP fragments scored for Juniperus cedrus. Valuesin bold indicate significance after Bonferroni correction(critical P = 0.0125)
Canaria) and highly significant in all cases. With thecaution imposed by the possible influence of extinc-tion in the current distribution of J. cedrus, this sug-gests that one of the factors influencing the observeddifferentiation pattern is insularity itself.
The dispersal attributes of J. cedrus (particularlyanemophily and ornithochory) have been described asmost favourable for long-distance dispersal in Maca-ronesia (Vargas, 2007), and several studies have pro-vided robust evidence that plant species displayingthese features are capable of maintaining substantialinter-island gene flow (e.g. García-Verdugo et al.,2010b; Ferreira et al., 2011). Consistently, FST calcu-lated with the plastid DNA data is considerably lowerin J. cedrus (FST = 0.080 ± 0.069) than in any otherMacaronesian tree for which FST estimates based onplastid DNA sequences are available (Table 5). Thesecomparisons and the weak IBD detected by spatialautocorrelation suggest the existence of moderatelevels of gene flow among the islands of distribution ofJ. cedrus. In agreement with this hypothesis, theoverall average number of migrants per generationwas Nm = 1.6 (data not shown), a value which wouldhave sufficed to maintain the genetic cohesion of thespecies, according to Slatkin (1985, 1987).
The dramatic reduction in population sizes after thehuman colonization of the islands from Europe(Höllermann, 1978; Leuschner, 1996; Francisco-Ortega et al., 2009) seems to have influenced thegenetic structure of the Canarian juniper. In contrastwith the plastid results, FST for J. cedrus estimatedwith AFLP (0.235 ± 0.080, Table 5) is considerablyhigher than that found in Olea europaea L. subsp.guanchica P.Vargas, J.Hess, Muñoz Garm. & Kadereit(0.09, recalculated from the original data inGarcía-Verdugo et al., 2009), which is the only otherCanarian tree species with long-distance dispersalabilities and fragmented natural populations for whichsuch data are available. Furthermore, interceptiondistances in the AFLP correlograms (Fig. 4) do notexceed c. 30 km, which is lower than any inter-islanddistance.
Substantial Bayesian genetic structure differencesbetween Tenerife and the other islands of occurrencewere also underscored at a first level of analysis(Fig. 5A) and between Gran Canaria and La Palmaand La Gomera at a second level (Fig. 5B). On thewhole, these results indicate that, in J. cedrus, genedispersal via pollen and seed interchange betweenislands may have been much more restricted thanlevels of effective gene flow within islands. In con-trast, a recent study of its congener J. macrocarpa(Juan et al., 2012), based on AFLP and plastidmarkers, shows that this mainland juniper has notlost much of its genetic variation despite severe popu-lation fragmentation.
Another factor that may have substantially influ-enced the current genetic structuring of J. cedrus ishuman disturbance, which seems to have been espe-cially dramatic on the larger islands of Gran Canariaand Tenerife (Höllermann, 1978; Leuschner, 1996;Francisco-Ortega et al., 2009). Thus, intense felling,human-induced fire (to which the Canarian juniper isintolerant; B. Rumeu, unpubl. data) and the conse-quent reduction in population sizes could also havecontributed to the low genetic diversity levels andmoderate population structure in J. cedrus.
CONSERVATION GUIDELINES
This study clearly points to the small population ofGran Canaria as the least genetically diverse, aggra-vating its critical conservation status and highlight-ing the need for immediate conservation actions. Atthe same time, the smaller islands in the distributionof J. cedrus (La Palma and La Gomera) were geneti-cally the most diverse, and thus management effortsprioritizing the conservation of these populations arenecessary to preserve the evolutionary potential ofthe species. Notably, genetic structure estimatesderived from AFLP (Table 4, Fig. 5) reveal an incipi-ent, but significant, differentiation among the islandsof occurrence, highlighting the importance of preserv-ing the trees from all islands and their habitats.Ex-situ and in-situ management must take this resultinto account, with trees from Tenerife and GranCanaria showing independent genetic structure(higher in the first case), meaning that, wheneverpossible, it would be convenient to avoid the use ofseeds, plants or young trees from other islands ineventual reinforcements. Given the detection of posi-tive spatial autocorrelation (Fig. 4), the first x-axisintercept of the correlogram is an operational esti-mate of the average length of the shortest side of truegenetic patches that contain individuals related bydescent (Sokal, 1979). Although Epperson (1990) criti-cized this calculation because it is liable to slightstochastic and statistical biases, it is useful for con-servation because it gives an indication of the size ofthe areas in which individuals are more likely to beinbred. In the case of J. cedrus (see Fig. 4), spatialautocorrelation indicates that seed collections to fulfilthe conservation strategies proposed by the SpanishNational Catalogue of endangered species (RealDecreto 139/2011, BOE 46 of 23/02/2011) should beseparated by more than 30 km in order to preservemost of the species genetic diversity. In practice, thismeans that seed collection should be carried out on allislands in the distribution, encompassing the largestpossible number of trees without interfering with thereproductive turnover of the species.
The conservation of J. cedrus in the Canary Islandsshould also encompass ecological interactions. We
Table 5. Threatened and non-threatened trees distributed in the Macaronesian archipelagos (AZO, Azores; CAN,Canaries; MAD, Madeira) that have been the focus of molecular studies and among-population genetic differentiationestimates (in parentheses), when available (FST, except for some cases, as indicated). Islands of occurrence for the taxain the Canarian archipelago are indicated as follows: H, El Hierro; P, La Palma; G, La Gomera; T, Tenerife; C, GranCanaria; F, Fuerteventura; L, Lanzarote. Conservation status (Status) codes are as in the IUCN Red List (IUCN, 2013):CR, critically endangered; DD, data deficient; EN, endangered; LC, least concern; NT, near threatened; VU, vulnerable.NA, not assessed. Molecular markers: nSSR, nuclear single sequence repeat; ISSR, inter simple sequence repeat; RAPD,random amplified polymorphic DNA; ITS, internal transcribed spacer.
Taxa Archipelagos Canarian island of occurrence Status Markers used (genetic differentiation)
Bencomia caudata (Aiton) Webb &Berthel.
MAD, CAN P T C NA nSSRs (0.155 ± 0.113)1
Bencomia exstipulata Svent. CAN P T VU nSSRs (0.411 ± 0.000)1
Erica arborea L. CAN H P G T C F L NA Plastid DNA (0.22)2,*Erica azorica Hochst. ex Seub. AZO NA nDNA (0.577 ± 0.438) and plastid
DNA (0.473 ± 0.121)3,†Erica platycodon (Webb & Berthel.)
Rivas-Mart. et al.MAD, CAN H G T NA nDNA (0.577 ± 0.438) and plastid
DNA (0.473 ± 0.121)3,†Ilex canariensis Poir. MAD, CAN H P G T C LR/NT nDNA sequences and plastid DNA4;
Laurus azorica (Seub.) Franco AZO, MAD, CAN H P G T C F L? LR/NT AFLP10; plastid DNA11
Myrica faya (Aiton) Wilbur AZO, MAD, CAN H P G T C F L LR/LC nSSRs (0.037 ± 0.017)12
Myrica rivas-martinezii A.Santos &J.Herbert
CAN H P G CR RAPDs (0.487)13; nSSRs (0.087)12
Olea europaea L. subsp. guanchicaP.Vargas, J.Hess, Muñoz Garm. &Kadereit
CAN H P G T C F L NA RAPDs, ISSRs and ITS-114; AFLP(0.09 ± 0.03) and nSSR(0.12 ± 0.02)15; nSSR and plastidDNA (0.150–0.099)16,¶; plastidDNA17
Olea europaea L. subsp. cerasiformis(Webb & Berth.) Kunk. & Sund.
MAD DD nSSR18; RAPDs, ISSRs and ITS-114;AFLP and nSSR15; plastid DNA17
Phoenix canariensis Chabaud CAN H P G T C F L NA RAPDs19; allozymes (0.287)20
Picconia azorica (Tutin) Knobl. AZO EN RFLPs and plastid SSR (GST = 0.194)21
Pinus canariensis Sweet ex Spreng. CAN H P G T C LC Isozymes (0.091)22; isozymes(0.072 ± 0.020)23; plastid SSRs(0.071 ± 0.034)24,**; plastid SSRs25;nSSRs and plastid SSR26; nSSRs(0.019) and plastid SSR (0.013)27
Sambucus palmensis Link CAN P G T C NA nSSRs (0.105 ± 0.035)28
Vaccinium cylindraceum Sm. AZO NA RAPDs29
1González-Pérez et al. (2009a); 2Désamoré et al. (2011); 3Désamoré et al. (2012); 4Manen (2004); 5González-González (2011); 6Silva et al. (2011); 7Rumeu et al.(2011a); 8This study; 9Adams et al. (2002); 10Arroyo-García et al. (2001); 11Rodríguez-Sánchez et al. (2009); 12González-Pérez et al. (2009b); 13Batista et al. (2004);14Hess, Kadereit & Vargas (2000); 15García-Verdugo et al. (2009) [FST estimates do not appear in the original paper, but were calculated from the original databy C. García-Verdugo]; 16García-Verdugo et al. (2010b); 17García-Verdugo et al. (2010a); 18Brito et al. (2008); 19González-Pérez, Caujapé-Castells & Sosa (2004b);20González-Pérez, Caujapé-Castells & Sosa (2004a); 21Ferreira et al. (2011); 22Schiller et al. (1999); 23Korol et al. (1999); 24Gómez et al. (2003); 25Navascués et al.(2006); 26Navascués & Emerson (2007); 27Navascués, Vendramin & Emerson (2008); 28Sosa et al. (2010); 29Martín-Clemente, Pereira & Perez-Ruiz (2001).*Global FST value between the four geographical regions defined across the distribution range of Erica arborea (Macaronesia, western Mediterranean, centraland eastern Mediterranean and eastern Africa/Arabia).†Authors considered E. azorica as E. scoparia subsp. azorica and E. platycodon as two subspecies of E. scoparia: E. scoparia subsp. maderinicola from Madeiraand E. scoparia subsp. platycodon from the Canary Islands. F-statistics correspond to comparisons among these three subspecies of E. scoparia.‡As only one population of Ilex perado subsp. lopezlilloi was considered in the study, genetic distances were calculated with respect to six populations ofI. perado subsp. platyphylla.§Authors reported that the large majority of population pairwise FST values (18 populations) were situated between 0.3 and 0.6.¶F-statistics averaged using values for young (0.150 ± 0.037; N = 5) and old (0.099 ± 0.019; N = 4) subpopulations.**Significant genetic distance based on the stepwise mutation model statistic of Goldstein et al. (1995).
therefore recommend complementary measures thatpromote the recovery of natural and effective long-range seed dispersal agents, such as ravens [Corvuscorax (Nogales, Hernández & Valdés, 1999; Rumeuet al., 2011c), which are in dramatic decline in theCanaries (Siverio, González & Siverio, 2010)] andwintering ring ouzels [Turdus torquatus (Rumeuet al., 2009b, 2011c), already listed as a speciesdeserving high conservation concern in Britain (Simet al., 2007)]. Finally, these conservation guidelinesshould be combined with actions to enforce habitatprotection and to avoid grazing by introducedmammals. The mitigation of the impacts of herbivores(especially goats and rabbits) would probably improvepopulation turnover.
ACKNOWLEDGEMENTS
We thank F. M. Medina, A. Fernández, D. P. Padilla,H. Fernandes, M. Sequeira, M. Soto and R.Domínguez for their help with sample collection. The‘Organismo Autónomo de Parques Nacionales’ gave uspermission to carry out the sampling on La Gomeraand Tenerife, and the Consejería de Medio Ambientedel Cabildo de La Palma and the Consejería de MedioAmbiente y Ordenación Territorial del Gobierno deCanarias permitted us to sample on La Palma andGran Canaria, respectively. We are especially gratefulto E. Cano for his support in the laboratory and to J.Martínez for essential plant material. J. L. Blanco-Pastor helped us with phylogenetic data analyses. B.Rumeu was funded by a Spanish National ResearchCouncil grant (CSIC: JAE-PRE). This work was sup-ported by projects co-funded by the Spanish ‘Ministe-rio de Ciencia e Innovación’ (CGL2010-18759), the‘Gobierno de Canarias’ (PI2007/05) and the EuropeanUnion. The ‘Organismo Autónomo de Parques Nacion-ales’ (project 051/2010) also financed part of thiswork.
REFERENCES
Adams RP. 2011. Junipers of the world: the genus Juniperus,3rd edn. Bloomington, IN: Trafford Publishing.
Adams RP, Bartel JA, Price RA. 2009. A new genus,Hesperocyparis, for the cypresses of the Western Hemi-sphere (Cupressaceae). Phytologia 91: 160–185.
Adams RP, Fontinha SS, Rumeu B, Nogales M. 2010.Speciation of Juniperus cedrus and J. maderensis in thearchipelagos of Canaries and Madeira based on terpenoidsand nrDNA and petN-psbM sequences. Phytologia 92:44–55.
Adams RP, Pandey N, Rezzi S, Casanova J. 2002. Geo-graphic variation in the random amplified polymorphicDNAs (RAPDs) of Juniperus phoenicea, J. p. var. canarien-
sis, J. p. subsp. eu-mediterranea, and J. p. var. turbinata.Biochemical Systematics and Ecology 30: 223–229.
Adams RP, Schwarzbach AE. 2012. Taxonomy of Junipe-rus: sequence analysis of nrDNA and five cpDNA regions.Phytologia 94: 280–297.
Akaike H. 1979. A Bayesian extension of the minimum AICprocedure of autoregressive model fitting. Biometrika 66:237–242.
Arroyo-García R, Martínez-Zapater JM, FernándezPrieto JA, Álvarez-Arbesú R. 2001. AFLP evaluation ofgenetic similarity among laurel populations (Laurus L.).Euphytica 122: 155–164.
Avise J. 2000. Phylogeography: the history and formation ofspecies. Cambridge, MA: Harvard University Press.
Batista F, Bouza N, González-Pérez MA, Caujapé-Castells J, Sosa PA. 2004. Genetic variation within andbetween populations of two endangered endemic species ofthe laurel forest from the Canary Islands, Myrica rivas-martinezii (Myricaceae) and Sideritis discolor (Lamiaceae).Australian Journal of Botany 52: 471–480.
BOE. 2011. Real Decreto 139/2011, de 4 de febrero, para eldesarrollo del Listado de Especies Silvestres en Régimen deProtección Especial y del Catálogo Español de EspeciesAmenazadas. Boletin Oficial del Estado (BOE) 46: 20 912–20 951.
Brito G, Loureiro J, Lopes T, Rodriguez E, Santos C.2008. Genetic characterisation of olive trees from Madeiraarchipelago using flow cytometry and microsatellitemarkers. Genetic Resources and Crop Evolution 55: 657–664.
Carine MA, Santos-Guerra A, Guma IR, Reyes-Betancort JA. 2010. Endemism and evolution of the Maca-ronesian flora. In: Williams DM, Knapp S, eds. Beyondcladistics: the branching of a paradigm. Berkeley, CA: Uni-versity of California Press, 101–124.
Carlquist S. 1965. Island life: a natural history of the islandsof the world, 1st edn. Garden City, NY: Natural HistoryPress.
Carracedo JC, Day S. 2002. Canary Islands. Harpenden,Hertfordshire: Terra Publishing.
Caujapé-Castells J. 2011. Jesters, red queens, boomerangsand surfers: a molecular outlook on the diversity of theCanarian endemic flora. In: Bramwell D, Caujapé-CastellsJ, eds. The biology of islands floras. Cambridge: CambridgeUniversity Press, 284–324.
Caujapé-Castells J, Castellano JJ, Henríquez V, RamosR, Sabbagh I, Quintana-Trujillo FM, Rodríguez JF.2011. Transformer-4: a genotype matrix format translator.Las Palmas de Gran Canaria, Spain. Available at: http://demiurge-project.org/download_t4: Jardín Botánico Canario‘Viera y Clavijo’-Unidad Asociada CSIC, Instituto Tec-nológico de Canarias, Jablesoft & Inventiaplus.
Caujapé-Castells J, Tye A, Crawford DJ, Santos-GuerraA, Sakai A, Beaver K, Lobin W, Vincent Florens FB,Moura M, Jardim R, Gómes I, Kueffer C. 2010. Conser-vation of oceanic island floras: present and future globalchallenges. Perspectives in Plant Ecology, Evolution andSystematics 12: 107–129.
Clement M, Posada D, Crandall KA. 2000. TCS: a com-puter program to estimate gene genealogies. MolecularEcology 9: 1657–1659.
Coart E, Glabeke SV, Petit RJ, Bockstaele EV,Roldán-Ruiz I. 2005. Range wide versus local patterns ofgenetic diversity in hornbeam (Carpinus betulus L.). Con-servation Genetics 6: 259–273.
Crawford DJ, Ruiz E, Stuessy TF, Tepe E, Aqeveque P,Gonzalez F, Jensen RJ, Anderson GJ, Bernardello G,Baeza CM, Swenson U, Silva OM. 2001. Allozyme diver-sity in endemic flowering plant species of the Juan Fernan-dez archipelago, Chile: ecological and historical factors withimplications for conservation. American Journal of Botany88: 2195–2203.
Désamoré A, Laenen B, Devos N, Popp M,González-Mancebo JM, Carine MA, Vanderpoorten A.2011. Out of Africa: north-westwards Pleistocene expan-sions of the heather Erica arborea. Journal of Biogeography38: 164–176.
Désamoré A, Laenen B, González-Mancebo JM,Jaén Molina R, Bystriakova N, Martinez-Klimova E,Carine MA, Vanderpoorten A. 2012. Inverted patterns ofgenetic diversity in continental and island populations ofthe heather Erica scoparia s.l. Journal of Biogeography 39:574–584.
Doyle JJ, Doyle JL. 1987. A rapid DNA isolation procedurefor small quantities of fresh leaf tissue. Phytochemical Bul-letin, Botanical Society of America 19: 11–15.
Drummond A, Rambaut A. 2007. BEAST: Bayesian evolu-tionary analysis by sampling trees. BMC EvolutionaryBiology 7: 214.
Drummond AJ, Ho SYW, Phillips MJ, Rambaut A. 2006.Relaxed phylogenetics and dating with confidence. PLoSBiology 4: 699–710.
Ellstrand NC, Elam DR. 1993. Population genetic conse-quences of small population size: implications for plantconservation. Annual Review of Ecology and Systematics 24:217–242.
Emerson BC. 2002. Evolution on oceanic islands: molecularphylogenetic approaches to understanding pattern andprocess. Molecular Ecology 11: 951–966.
Epperson BK. 1990. Spatial patterns of genetic variationwithin plant populations. In: Brown AHD, Clegg MT,Kahler AL, Weir BS, eds. Plant population genetics, breed-ing and genetic resources. Sunderland, MA: Sinauer Associ-ates, Inc., 229–253.
Evanno G, Regnaut S, Goudet J. 2005. Detecting thenumber of clusters of individuals using the software Struc-ture: a simulation study. Molecular Ecology 14: 2611–2620.
Excoffier L, Lischer HEL. 2010. Arlequin suite ver 3.5: anew series of programs to perform population geneticsanalyses under Linux and Windows. Molecular EcologyResources 10: 564–567.
Falush D, Stephens M, Pritchard JK. 2003. Inference ofpopulation structure using multilocus genotype data: linked
loci and correlated allele frequencies. Genetics 164: 1567–1587.
Fay MF, Cowan RS, Leitch IJ. 2005. The effects of nuclearDNA content (C-value) on the quality and utility of AFLPfingerprints. Annals of Botany 95: 237–246.
Fazekas AJ, Burgess KS, Kesanakurti PR, Graham SW,Newmaster SG, Husband BC, Percy DM, HajibabaeiM, Barrett SCH. 2008. Multiple multilocus DNA barcodesfrom the plastid genome discriminate plant species equallywell. PLOS ONE 3: e2802.
Fernández-Mazuecos M, Vargas P. 2011. Genetically dep-auperate in the continent but rich in oceanic islands: Cistusmonspeliensis (Cistaceae) in the Canary Islands. PLOSONE 6: e17172.
Ferreira RC, Piredda R, Bagnoli F, Bellarosa R,Attimonelli M, Fineschi S, Schirone B, Simeone MC.2011. Phylogeography and conservation perspectives of anendangered Macaronesian endemic: Picconia azorica (Tutin)Knobl. (Oleaceae). European Journal of Forest Research130: 181–195.
Francisco-Ortega J, Jansen RK, Santos-Guerra A. 1996.Chloroplast DNA evidence of colonization, adaptive radia-tion, and hybridization in the evolution of the Macaronesianflora. Proceedings of the National Academy of Sciences 93:4085–4090.
Francisco-Ortega J, Santos-Guerra A, Bacallado JJ.2009. Canary Islands, biology. In: Gillespie RG, Clague DA,eds. Encyclopedia of Islands. Berkeley, CA: University ofCalifornia Press, 127–133.
Francisco-Ortega J, Santos-Guerra A, Kim S-C,Crawford DJ. 2000. Plant genetic diversity in the CanaryIslands: a conservation perspective. American Journal ofBotany 87: 909–919.
Frankham R. 1997. Do island populations have less geneticvariation than mainland populations? Heredity 78: 311–327.
Frankham R. 1998. Inbreeding and extinction: island popu-lations. Conservation Biology 12: 665–675.
García D. 2001. Effects of seed dispersal on Juniperus com-munis recruitment on a Mediterranean mountain. Journalof Vegetation Science 12: 839–848.
García-Maroto F, Mañas-Fernández A, Garrido-Cárdenas JA, López Alonso D, Guil-Guerrero JL,Guzmán B, Vargas P. 2009. Δ6-desaturase sequence evi-dence for explosive Pliocene radiations within the adaptiveradiation of Macaronesian Echium (Boraginaceae). Molecu-lar Phylogenetics and Evolution 52: 563–574.
García-Verdugo C, Fay MF, Granado-Yela C,Rubio de Casas RR, Balaguer L, Besnard G, Vargas P.2009. Genetic diversity and differentiation processes in theploidy series of Olea europaea L.: a multiscale approachfrom subspecies to insular populations. Molecular Ecology18: 454–467.
García-Verdugo C, Forrest AD, Balaguer L, Fay MF,Vargas P. 2010a. Parallel evolution of insular Oleaeuropaea subspecies based on geographical structuring ofplastid DNA variation and phenotypic similarity in leaftraits. Botanical Journal of the Linnean Society 162: 54–63.
García-Verdugo C, Forrest AD, Fay MF, Vargas P. 2010b.The relevance of gene flow in metapopulation dynamics ofan oceanic island endemic, Olea europaea subsp. guanchica.Evolution 64: 3525–3536.
Gernhard T. 2008. The conditioned reconstructed process.Journal of Theoretical Biology 253: 769–778.
Goldstein DB, Linares AR, Cavalli-Sforza LL, FeldmanMW. 1995. An evaluation of genetic distances for use withmicrosatellite loci. Genetics 139: 463–471.
Goloboff PA, Farris JS, Nixon KC. 2008. TNT, a freeprogram for phylogenetic analysis. Cladistics 24: 774–786.
Gómez A, González-Martínez SC, Collada C, Climent J,Gil L. 2003. Complex population genetic structure in theendemic Canary Island pine revealed using chloroplastmicrosatellite markers. Theoretical and Applied Genetics107: 1123–1131.
González-González EA. 2011. Análisis de la diversidadgenética en poblaciones naturales de especies vegetalesamenazadas: Ilex perado ssp. lopezlilloi (Aquifoliaceae),Silene nocteolens (Caryophyllaceae) y Sorbus aria(Rosaceae). DPhil Thesis, University of Las Palmas de GranCanaria, Spain.
González-Pérez MA, Caujapé-Castells J, Sosa PA. 2004a.Allozyme variation and structure of the Canarian endemicpalm tree Phoenix canariensis (Arecaceae): implications forconservation. Heredity 93: 307–315.
González-Pérez MA, Caujapé-Castells J, Sosa PA. 2004b.Molecular evidence of hybridisation between the endemicPhoenix canariensis and the widespread P. dactylifera withrandom amplified polymorphic DNA (RAPD) markers. PlantSystematics and Evolution 247: 165–175.
González-Pérez MA, Lledó MD, Lexer C, Fay MF,Marrero M, Bañares-Baudet A, Carqué E, Sosa PA.2009a. Genetic diversity and differentiation in natural andreintroduced populations of Bencomia exstipulata and com-parisons with B. caudata (Rosaceae) in the Canary Islands:an analysis using microsatellites. Botanical Journal of theLinnean Society 160: 429–441.
González-Pérez MA, Sosa PA, Rivero E,González-González EA, Naranjo A. 2009b. Molecularmarkers reveal no genetic differentiation between Myricarivas-martinezii and M. faya (Myricaceae). Annals of Botany103: 79–86.
Hamilton MB. 1999. Four primer pairs for the amplificationof chloroplast intergenic regions with intraspecific variation.Molecular Ecology 8: 521–523.
Hardy OJ, Vekemans X. 2002. Spagedi: a versatile computerprogram to analyse spatial genetic structure at the indi-vidual or population levels. Molecular Ecology Notes 2:618–620.
Hess J, Kadereit JW, Vargas P. 2000. The colonizationhistory of Olea europaea L. in Macaronesia based on inter-nal transcribed spacer 1 (ITS-1) sequences, randomly ampli-fied polymorphic DNAs (RAPD), and intersimple sequencerepeats (ISSR). Molecular Ecology 9: 857–868.
Hizume M, Kondo T, Shibata F, Ishizuka R. 2001. Flowcytometric determination of genome size in the Taxodiaceae,
Cupressaceae sensu stricto and Sciadopityaceae. Cytologia66: 307–311.
Höllermann PW. 1978. Geological aspects of the upper tim-berline in Tenerife, Canary Islands. Arctic and AlpineResearch 10: 365–382.
del Hoyo A, García-Marín JL, Pedrola-Monfort J. 2009.Temporal and spatial diversification of the African disjunctgenus Androcymbium (Colchicaceae). Molecular Phylogenet-ics and Evolution 53: 848–861.
Hwang LH, Hwang SY, Lin TP. 2000. Low chloroplast DNAvariation and population differentiation of Chamaecyparisformosensis and Chamaecyparis taiwanensis. TaiwanJournal of Forest Science 15: 229–236.
IUCN. 2013. IUCN red list of threatened species. Version2013.1. Available at: http://www.iucnredlist.org [accessed 30October 2013].
Jordano P. 1993. Geographical ecology and variation ofplant–seed disperser interactions: southern Spanish juni-pers and frugivorous thrushes. Vegetatio 107/108: 85–104.
Juan A, Fay MF, Pastor J, Juan R, Fernández I, CrespoMB. 2012. Genetic structure and phylogeography in Juni-perus oxycedrus subsp. macrocarpa around the Mediterra-nean and Atlantic coasts of the Iberian Peninsula, based onAFLP and plastid markers. European Journal of ForestResearch 131: 845–856.
Kim S-C, McGowen MR, Lubinsky P, Barber JC, MortME, Santos-Guerra A. 2008. Timing and tempo of earlyand successive adaptive radiations in Macaronesia. PLoSONE 3: e2139.
Korol L, Gil L, Climent J, Zehavi A, Schiller G. 1999.Canary Islands pine (Pinus canariensis Chr. Sm. ex DC.) 2.Gene flow among native populations. Forest Genetics 6:277–282.
Kress WJ, Erickson DL. 2007. A two-locus global DNAbarcode for land plants: the coding rbcL gene complementsthe non-coding trnH-psbA spacer region. PLoS ONE 2: e508.
Leuschner C. 1996. Timberline and alpine vegetation on thetropical and warm-temperate oceanic islands of the world:elevation, structure and floristics. Vegetatio 123: 193–206.
Librado P, Rozas J. 2009. DnaSP v5: a software for com-prehensive analysis of DNA polymorphism data. Bioinfor-matics 25: 1451–1452.
Livingston RB. 1972. Influence of birds, stones and soil onthe establishment of pasture juniper, Juniperus communis,and red cedar, J. virginiana in New England pastures.Ecology 53: 1141–1147.
Manen J-F. 2004. Are both sympatric species Ilex perado andIlex canariensis secretly hybridizing? Indication fromnuclear markers collected in Tenerife. BMC EvolutionaryBiology 4: 46.
Mantel N. 1967. Detection of disease clustering and a gen-eralized regression approach. Cancer Research 27: 209–220.
Mao K, Hao G, Liu J, Adams RP, Milne RI. 2010. Diver-sification and biogeography of Juniperus (Cupressaceae):variable diversification rates and multiple intercontinentaldispersals. New Phytologist 188: 254–272.
Martín-Clemente J, Pereira MJ, Perez-Ruiz C. 2001.DNA extraction from leaves of Vaccinium cylindraceum
Smith (Ericaceae). The use of RAPD markers to detectgenetic variation. Preliminary results. Arquipélago. Lifeand Marine Sciences Supl. 2 (B): 105–109.
Meudt HM, Clarke AC. 2007. Almost forgotten or latestpractice? AFLP applications, analyses and advances. Trendsin Plant Science 12: 106–117.
Moreno JC, coord. 2008. Lista Roja 2008 de la flora vascularespañola. Madrid: Dirección General de Medio Natural yPolítica Forestal (Ministerio de Medio Ambiente, y MedioRural y Marino, y Sociedad Española de Biología de laConservación de Plantas).
Müller K. 2006. Incorporating information from length–mutational events into phylogenetic analysis. MolecularPhylogenetics and Evolution 38: 667–676.
Navascués M, Emerson BC. 2007. Natural recovery ofgenetic diversity by gene flow in reforested areas of theendemic Canary Island pine, Pinus canariensis. ForestEcology and Management 244: 122–128.
Navascués M, Vaxevanidou Z, González-Martínez SC,Climent J, Gil L, Emerson BC. 2006. Chloroplast micro-satellites reveal colonization and metapopulation dynamicsin the Canary Island pine. Molecular Ecology 15: 2691–2698.
Navascués M, Vendramin GG, Emerson BC. 2008. Theeffect of altitude on patterns of gene flow in the endemicCanary Island pine, Pinus canariensis. Silvae Genetica 57:357–363.
Nogales M, Hernández EC, Valdés F. 1999. Seed dispersalby common ravens Corvus corax among island habitats(Canarian Archipelago). Ecoscience 6: 56–61.
Nybom H. 2004. Comparison of different nuclear DNAmarkers for estimating intraspecific genetic diversity inplants. Molecular Ecology 13: 1143–1155.
Ojeda I, Santos-Guerra A, Jaén-Molina R, Oliva-TejeraF, Caujapé-Castells J, Cronk Q. 2012. The origin of birdpollination in Macaronesian Lotus (Loteae, Leguminosae).Molecular Phylogenetics and Evolution 62: 306–318.
Palmer JD, Jansen RK, Michaels HJ, Chase MW,Manhart JR. 1988. Chloroplast DNA variation and plantphylogeny. Annals of the Missouri Botanical Garden 75:1180–1206.
Parsons JJ. 1981. Human influences on the pine and laurelforests of the Canary Islands. The Geographical Review 71:253–271.
Posada D. 2008. jModelTest: phylogenetic model averaging.Molecular Biology and Evolution 25: 1253–1256.
Pritchard JK, Stephens M, Donnelly P. 2000. Inference ofpopulation structure using multilocus genotype data. Genet-ics 155: 945–959.
Provan J, Beatty GE, Hunter AM, McDonald RA,McLaughlin E, Preston SJ, Wilson S. 2008. Restrictedgene flow in fragmented populations of a wind-pollinatedtree. Conservation Genetics 9: 1521–1532.
Rambaut A, Drummond AJ. 2007. Tracer v1.4. Available at:http://beast.bio.ed.ac.uk/Tracer
Rodríguez-Sánchez F, Guzmán B, Valido A, Vargas P,Arroyo J. 2009. Late Neogene history of the laurel tree(Laurus L., Lauraceae) based on phylogeographical analysesof Mediterranean and Macaronesian populations. Journal ofBiogeography 36: 1270–1281.
Rosenberg MS, Anderson CD. 2011. PASSaGE: patternanalysis, spatial statistics and geographic exegesis. Version2. Methods in Ecology and Evolution 2: 229–232.
Rumeu B, Caujapé-Castells J, Blanco-Pastor JL,Jaén-Molina R, Nogales M, Elias RB, Vargas P. 2011a.The colonization history of Juniperus brevifolia (Cupres-saceae) in the Azores islands. PLoS ONE 6: e27697.
Rumeu B, Elias RB, Padilla D, Melo C, Nogales M. 2011c.Differential seed dispersal systems of endemic junipers intwo oceanic Macaronesian archipelagos: the influence ofbiogeographic and biological characteristics. Plant Ecology212: 911–921.
Rumeu B, Nogales M, Elias RB, Padilla DP, Resendes T,Rodríguez A, Valdés F, Dias E. 2009a. Contrasting phe-nology and female cone characteristics of the two Macaro-nesian island endemic cedars (Juniperus cedrus andJ. brevifolia). European Journal of Forest Research 128:567–574.
Rumeu B, Padilla DP, Nogales M. 2009b. The key role ofa ring ouzel Turdus torquatus wintering population in seeddispersal of the endangered endemic Juniperus cedrus in aninsular environment. Acta Ornithologica 44: 199–204.
Rumeu B, de Sequeira M, Elliot M, Gardner M. 2011b.Juniperus cedrus. In: IUCN 2012. IUCN red list of threat-ened species. Version 2012.2. Available at: http://www.iucnredlist.org [accessed 27 June 2013].
Sang T, Crawford DJ, Stuessy TF. 1997. Chloroplast DNAphylogeny, reticulate evolution, and biogeography ofPaeonia (Paeoniaceae). American Journal of Botany 84:1120–1136.
Schiller G, Korol L, Ungar ED, Zehavi A, Gil L, ClimentJ. 1999. Canary Islands pine (Pinus canariensis Chr. Sm. exDc.). 1. Differentiation among native populations in theirisoenzymes. Forest Genetics 6: 257–276.
Shaw J, Lickey EB, Beck JT, Farmer SB, Liu W, MillerJ, Siripun KC, Winder CT, Schilling EE, Small RL.2005. The tortoise and the hare II: relative utility of 21noncoding chloroplast DNA sequences for phylogeneticanalysis. American Journal of Botany 92: 142–166.
Shaw J, Lickey EB, Schilling EE, Small RL. 2007. Com-parison of whole chloroplast genome sequences to choosenoncoding regions for phylogenetic studies in angiosperms:the tortoise and the hare III. American Journal of Botany94: 275–288.
Silva L, Elias RB, Moura M, Meimberg H, Dias E. 2011.Genetic variability and differentiation among populations ofthe Azorean endemic gymnosperm Juniperus brevifolia:baseline information for a conservation and restoration per-spective. Biochemical Genetics 49: 715–734.
Sim IMW, Burfield IJ, Grant MC, Pearce-Higgins JW,Brooke M de L. 2007. The role of habitat composition indetermining breeding site occupancy in a declining ringouzel Turdus torquatus population. Ibis 149: 374–385.
Simmons MP, Ochoterena H. 2000. Gaps as characters insequence-based phylogenetic analyses. Systematic Biology49: 369–381.
Siverio M, González EI, Siverio F. 2010. Population sizeand status of common raven (Corvus corax) on the central-western islands of the Canarian archipelago. Vieraea 38:123–132.
Slatkin M. 1985. Gene flow in natural populations. AnnualReview of Ecology and Systematics 16: 393–430.
Slatkin M. 1987. Gene flow and the geographic structure ofnatural populations. Science 236: 787–792.
Sokal RR, Oden NL. 1978. Spatial autocorrelation inbiology: 1. Methodology. Biological Journal of the LinneanSociety 10: 199–228.
Sosa PA, González-Pérez MA, Moreno C, Clarke J. 2010.Conservation genetics of the endangered endemic Sambucuspalmensis Link (Sambucaceae) from the Canary Islands.Conservation Genetics 11: 2357–2368.
Stamatakis A. 2006. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa andmixed models. Bioinformatics 22: 2688–2690.
Taberlet P, Gielly L, Pautou G, Bouvet J. 1991. Universalprimers for amplification of three non-coding regions ofchloroplast DNA. Plant Molecular Biology 17: 1105–1109.
Templeton AR, Crandall KA, Sing CF. 1992. A cladisticanalysis of phenotypic associations with haplotypes inferredfrom restriction endonuclease mapping and DNA sequencedata. III. Cladogram estimation. Genetics 132: 619–633.
Terrab A, Schönswetter P, Talavera S, Vela E, StuessyTF. 2008. Range-wide phylogeography of Juniperus thurif-era L., a presumptive keystone species of western Mediter-ranean vegetation during cold stages of the Pleistocene.Molecular Phylogenetics and Evolution 48: 94–102.
Triantis KA, Borges PAV, Ladle RJ, Hortal J, Cardoso P,Gaspar C, Dinis F, Mendonça E, Silveira LMA, GabrielR, Melo C, Santos AMC, Amorim IR, Ribeiro SP,Serrano ARM, Quartau JA, Whittaker RJ. 2010. Extinc-tion debt on oceanic islands. Ecography 33: 285–294.
Vargas P. 2007. Are Macaronesian islands refugia of relictplant lineages?: a molecular survey. In: Weiss S, Ferrand N,eds. Phylogeography of southern European refugia: evolu-tionary perspectives on the origins and conservation ofEuropean biodiversity. Berlin: Springer, Dordrecht, 297–314.
Vekemans X, Beauwens T, Lemaire M, Roldán-Ruiz I.2002. Data from amplified fragment length polymorphism(AFLP) markers show indication of size homoplasy and of arelationship between degree of homoplasy and fragmentsize. Molecular Ecology 11: 139–151.
Vos P, Hogers R, Bleeker M, Reijans M, van de Lee T,Hornes M, Friters A, Pot J, Paleman J, Kuiper M,Zabeau M. 1995. AFLP: a new technique for DNA finger-printing. Nucleic Acids Research 23: 4407–4414.
Ward LK. 1982. The conservation of juniper: longevity andold age. Journal of Applied Ecology 19: 917–928.
Whittaker RJ, Fernández-Palacios JM. 2007. Island bio-geography: ecology, evolution, and conservation, 2nd edn.New York: Oxford University Press.
Yeh FC, Yang R-C, Boyle TBJ, Ye Z-H, Mao JX. 1997.POPGENE, the user-friendly shareware for populationgenetic analysis. Edmonton, AB: Molecular Biology and Bio-technology Centre, University of Alberta.
Young A, Boyle T, Brown T. 1996. The population geneticconsequences of habitat fragmentation for plants. Trends inEcology & Evolution 11: 413–418.
Zamora R. 1990. The fruit diet of ring-ouzels (Turdus torqua-tus) wintering in Sierra Nevada (south-east Spain). Alauda58: 67–70.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the online version of this article at the publisher’s web-site:
Figure S1 Haplotype network and its spatial distribution based on petN-psbM and trnT-trnL plastid DNAregions and calculated by treating gaps as a fifth state.Figure S2 ΔK calculated as described in Evanno et al. (2005) for each Structure analysis plotted in Figure 5.Table S1 GenBank accession numbers, locality and voucher information of the samples included in thephylogenetic reconstruction of Juniperus section Juniperus, inferred from trnL and trnL-trnF plastid sequences.Table S2 The 86 samples of Juniperus used for petN-psbM and trnT-trnL sequencing and GenBank accessionnumbers for each haplotype detected.