Norwitz, L6pez Bernal, and Starkey Turnbull AC. Prostaglandins, chorioamnionitis and pre- term labour. Br J Obstet Gynaecol 1987;94: 1156-8. 19. Romero R, Wu YK, Brody D, Oyarzun E, DuffGW, Du- rum SK. Human decidua: a source of interleukin-1. Ob- stet Gynecol 1989;73:31-4. 20. Santhanam U, Avila C, Romero R, et al. Cytokines in normal and abnormal parturition: elevated amniotic fluid September 1992 Am J Obstet Gynecol interleukin-6 levels in women with premature rupture of membranes associated with intrauterine infection. Cyto- kines 1991;3:155-63. 21. Norwitz ER, Starkey PM, L6pez Bernal A. Prostaglandin D. production by term human decidua: cellular origins defined using flow cytometry. Obstet Gynecol 1992;80 [In press]. Physiologic baroreceptor activity in the fetal lamb Akihiko Wakatsuki, MD, Yuji Murata, MD. Yuko Ninomiya, MD, Naoki Masaoka, MD, James G. Tyner, and Krishna K. Kutty Orange, California OBJECTIVE: We investigated the influences of gestational age and behavioral state on fetal baroreceptor activity. STUDY DESIGN: Two protocols were used to analyze jow-voltage fast activity and high-voltage slow activity in eight chronically instrumented ovine fetuses at. days 127 to 142. In protocol 1 correlation of fetal heart rate to mean arterial blood pressure was assessed under physiologic conditions, and the linear regression slope of fetal heart rate on mean arterial pressure was calculated. In protocol 2 an injection of 7.5 I1g of norepinephrine was given during consecutive behavioral cycles. The ratio of fetal heart rate decrease to mean arterial pressure increase was determined as representing baroreceptor activity. RESULTS: Under protocol 1 the slope was steeper in high-voltage slow activity (2.33 ± 1.03) than in low-voltage fast activity (0.89 ± 0.37) (n = 21, P < 0.001), indicating that fetal heart rate changes are sensitive to blood pressure changes during high-voltage slow activity. Under protocol 2 inverse relationships were found between the ratio of fetal heart rate decrease to mean arterial pressure increase and gestational age in low-voltage fast activity and high-voltage slow activity. CONCLUSION: Fetal baroreceptor activity increased during high-voltage slow activity and decreased with gestational age. (AM J OaSTET GVNECOL 1992;167:820-7.) Key words: Baroreceptor reflex activity, fetal lamb, behavioral state, gestational age Heart rate decelerations have been frequently ob- served in human fetuses before 30 weeks' gestation,I.3 in animal fetuses born prematurely,. and in human neonates. 5 The mechanism of these decelerations may be reflex vagal nerve mediated! It has been reported that the increase in cholinesterase activity during ges- tation may help protect the mature fetus from severe bradycardia. 4 The physiologic implications of fetal From the Division of Maternal-Fetal Medicine, Department of Ob- stetrics and Gynecology, University of California, Irvine, College of Medicine, UCI Medical Center. Presented in part at the Thirty-eighth Annual Meeting of the Society for Gynecologic Investigation, San Antonio, Texas, March 20-23, 1991. Received for publication June 25, 1991, revised April 27, 1992; accepted May 4, 1992. Reprint requests: Yuji Murata, MD, Division of Maternal-Fetal M ed- icine, Department of Obstetrics and Gynecology, University of Cali- fornia, Irvine College of Medicine, UCI Medical Center, PO Box 14091, Orange, CA 92613-1491. 6/1/39052 820 heart rate (FHR) decelerations at early gestational ages and during the quiet behavioral state are not under- stood well enough to apply in clinical practice in peri- natal medicine. The purpose of this study was to quantitatively eval- uate baroreceptor reflex activities at differing gesta- tional ages and behavioral states in chronically instru- mented fetal lambs. Material and methods Surgical preparation. The fetal lambs of eight time- mated Columbia-Rambouillet cross-bred sheep were chronically instrumented between 127 and 142 days' gestation (term 145 days). All animals were maintained in a facility approved by the American Association for the Accreditation of Laboratory Animal Care and in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Before implementation research
19. Romero R, Wu YK, Brody D, Oyarzun E, DuffGW, Durum SK. Human decidua: a source of interleukin-1. Obstet Gynecol 1989;73:31-4.
20. Santhanam U, Avila C, Romero R, et al. Cytokines in normal and abnormal parturition: elevated amniotic fluid
September 1992 Am J Obstet Gynecol
interleukin-6 levels in women with premature rupture of membranes associated with intrauterine infection. Cytokines 1991;3:155-63.
21. Norwitz ER, Starkey PM, L6pez Bernal A. Prostaglandin D. production by term human decidua: cellular origins defined using flow cytometry. Obstet Gynecol 1992;80 [In press].
Physiologic baroreceptor activity in the fetal lamb
Akihiko Wakatsuki, MD, Yuji Murata, MD. Yuko Ninomiya, MD, Naoki Masaoka, MD, James G. Tyner, and Krishna K. Kutty
Orange, California
OBJECTIVE: We investigated the influences of gestational age and behavioral state on fetal baroreceptor activity.
STUDY DESIGN: Two protocols were used to analyze jow-voltage fast activity and high-voltage slow
activity in eight chronically instrumented ovine fetuses at. days 127 to 142. In protocol 1 correlation of fetal heart rate to mean arterial blood pressure was assessed under physiologic conditions, and the linear
regression slope of fetal heart rate on mean arterial pressure was calculated. In protocol 2 an injection of 7.5 I1g of norepinephrine was given during consecutive behavioral cycles. The ratio of fetal heart rate
decrease to mean arterial pressure increase was determined as representing baroreceptor activity.
RESULTS: Under protocol 1 the slope was steeper in high-voltage slow activity (2.33 ± 1.03) than in
low-voltage fast activity (0.89 ± 0.37) (n = 21, P < 0.001), indicating that fetal heart rate changes are sensitive to blood pressure changes during high-voltage slow activity. Under protocol 2 inverse
relationships were found between the ratio of fetal heart rate decrease to mean arterial pressure increase and gestational age in low-voltage fast activity and high-voltage slow activity.
CONCLUSION: Fetal baroreceptor activity increased during high-voltage slow activity and decreased with
Heart rate decelerations have been frequently observed in human fetuses before 30 weeks' gestation,I.3 in animal fetuses born prematurely,. and in human neonates. 5 The mechanism of these decelerations may be reflex vagal nerve mediated! It has been reported that the increase in cholinesterase activity during gestation may help protect the mature fetus from severe bradycardia.4 The physiologic implications of fetal
From the Division of Maternal-Fetal Medicine, Department of Obstetrics and Gynecology, University of California, Irvine, College of Medicine, UCI Medical Center. Presented in part at the Thirty-eighth Annual Meeting of the Society for Gynecologic Investigation, San Antonio, Texas, March 20-23, 1991. Received for publication June 25, 1991, revised April 27, 1992; accepted May 4, 1992. Reprint requests: Yuji Murata, MD, Division of Maternal-Fetal M edicine, Department of Obstetrics and Gynecology, University of California, Irvine College of Medicine, UCI Medical Center, PO Box 14091, Orange, CA 92613-1491. 6/1/39052
820
heart rate (FHR) decelerations at early gestational ages and during the quiet behavioral state are not understood well enough to apply in clinical practice in perinatal medicine.
The purpose of this study was to quantitatively evaluate baroreceptor reflex activities at differing gestational ages and behavioral states in chronically instrumented fetal lambs.
Material and methods
Surgical preparation. The fetal lambs of eight timemated Columbia-Rambouillet cross-bred sheep were chronically instrumented between 127 and 142 days' gestation (term 145 days).
All animals were maintained in a facility approved by the American Association for the Accreditation of Laboratory Animal Care and in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Before implementation research
Volume 167 Number 3
Baroreceptor activity in fetal lamb 821
Norepinephrine 7.5Jl1 1 min
-~ = E
120
! 60 ~ = ~
o
220
140
60
I I
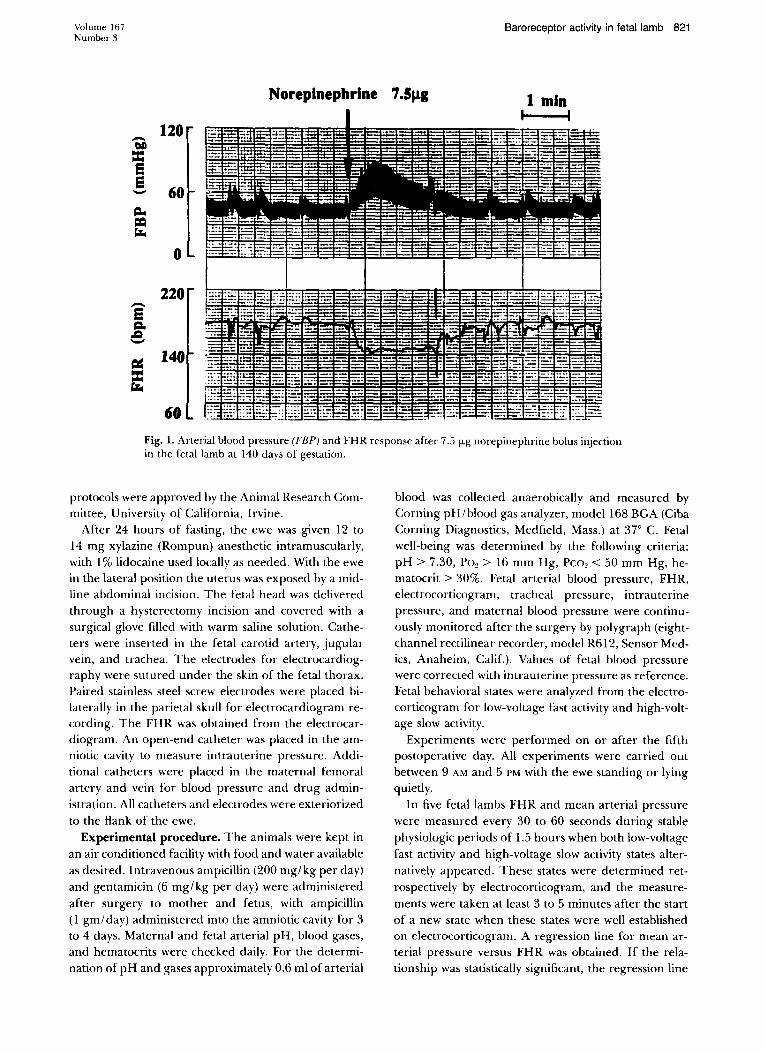
Fig. 1. Arterial blood pressure (FBP) and FHR response after 7.5 flog norepinephrine bolus injection in the fetal lamb at 140 days of gestation.
protocols were approved by the Animal Research Committee, University of California, Irvine.
After 24 hours of fasting, the ewe was given 12 to 14 mg xylazine (Rompun) anesthetic intramuscularly, with 1 % lidocaine used locally as needed. With the ewe in the lateral position the uterus was exposed by a midline abdominal incision. The fetal head was delivered through a hysterectomy incision and covered with a surgical glove filled with warm saline solution. Catheters were inserted in the fetal carotid artery, jugular vein, and trachea. The electrodes for electrocardiography were sutured under the skin of the fetal thorax. Paired stainless steel screw electrodes were placed bilaterally in the parietal skull for electrocardiogram recording. The FHR was obtained from the electrocardiogram. An open-end catheter was placed in the amniotic cavity to measure intrauterine pressure. Additional catheters were placed in the maternal femoral artery and vein for blood pressure and drug administration. All catheters and electrodes were exteriorized to the flank of the ewe.
Experimental procedure. The animals were kept in an air conditioned facility with food and water available as desired. Intravenous ampicillin (200 mg/kg per day) and gentamicin (6 mg/kg per day) were administered after surgery to mother and fetus, with ampicillin (1 gm/day) administered into the amniotic cavity for 3 to 4 days. Maternal and fetal arterial pH, blood gases, and hematocrits were checked daily. For the determination of pH and gases approximately 0.6 ml of arterial
blood was collected anaerobically and measured by Corning pH/blood gas analyzer, model 168 BGA (Ciba Corning Diagnostics, Medfield, Mass.) at 37° C. Fetal well-being was determined by the following criteria: pH> 7.30, POe> 16 mm Hg, Peo. < 50 mm Hg, hematocrit> 30%. Fetal arterial blood pressure, FHR, electrocorticogram, tracheal pressure, intrauterine pressure, and maternal blood pressure were continuously monitored after the surgery by polygraph (eightchannel rectilinear recorder, model R612, Sensor Medics, Anaheim, Calif.). Values of fetal blood pressure were corrected with intrauterine pressure as reference. Fetal behavioral states were analyzed from the electrocorticogram for low-voltage fast activity and high-voltage slow activity.
Experiments were performed on or after the fifth postoperative day. All experiments were carried out between 9 AM and 5 PM with the ewe standing or lying quietly.
In five fetal lambs FHR and mean arterial pressure were measured every 30 to 60 seconds during stable physiologic periods of 1.5 hours when both low-voltage fast activity and high-voltage slow activity states alternatively appeared. These states were determined retrospectively by electrocorticogram, and the measurements were taken at least 3 to 5 minutes after the start of a new state when these states were well established on electrocorticogram. A regression line for mean arterial pressure versus FHR was obtained. If the relationship was statistically significant, the regression line
822 Wakatsuki et al.
200
180
E CI. e 160 0
c::.:: = 0 r-
140
CD 0
0 0
•
o
• •
o LVFA
• HVSA
o
September 1992 Am J Obstet Gynecol
120~~--~--~~~~--~--~~~~--~
30 35 40 45 50 55
MAP (mmHg)
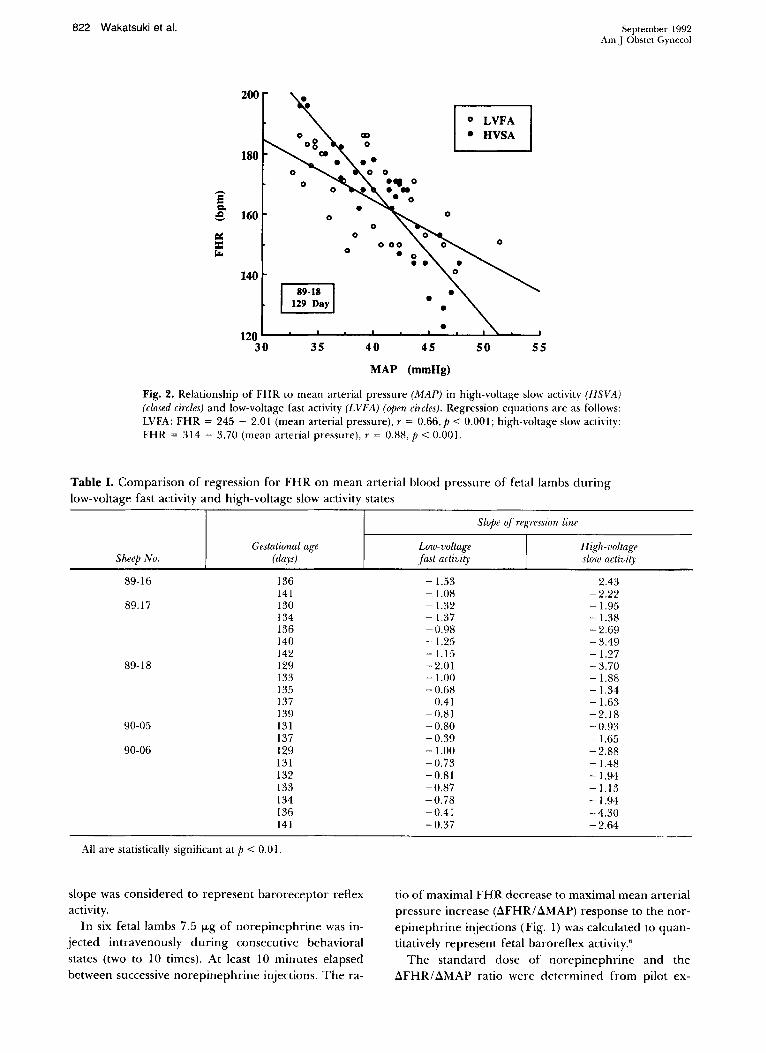
Fig. 2. Relationship of FHR to mean arterial pressure (MAP) in high-voltage slow activity (HSVA) (closed circles) and low-voltage fast activity (LVFA) (open circles). Regression equations are as follows: LVFA: FHR = 245 - 2.01 (mean arterial pressure), r = 0.66, P < 0.00 I; high-voltage slow actiyity: FHR = 314 - 3.70 (mean arterial pressure), r = 0.88, P < 0.00 I.
Table I. Comparison of regression for FHR on mean arterial blood pressure of fetal lambs during low-voltage fast activity and high-voltage slow activity states
slope was considered to represent baroreceptor reflex activity.
In six fetal lambs 7.5 fLg of norepinephrine was injected intravenously during consecutive behavioral states (two to 10 times). At least 10 minutes elapsed between successive norepinephrine injections. The ra-
tio of maximal FHR decrease to maximal mean arterial pressure increase (ilFHRI ilMAP) response to the norepinephrine injections (Fig. 1) was calculated to quantitatively represent fetal baroreAex activity."
The standard dose of norepinephrine and the ilFHRI ilMAP ratio were aett'fminea from pilot ex-
Volume 167 Number 3
periments conducted on three fetal lambs at 127, 129, and 138 days' gestation, respectively. In these norepinephrine was injected at five doses (2.5, 5.0, 7.5, 10.0, and 15.0 J.1g). The relation between AFHR and AMAP and norepinephrine dosage in low-voltage fast activity and high-voltage slow activity states was observed.
Data analysis. The differences between low-voltage fast activity and high-voltage slow activity behavioral states were analyzed by paired t test and found significant. The regression line was analyzed by the leastsquares method, and regression slopes were compared by t test regression analysis. 7 Statistical significance was accepted at p < 0.05. All data were reported as mean ± SD.
Results
Fig. 2 demonstrates an example of the regression analysis of FHR on mean arterial pressure in both behavioral states, with the slope of the regression line steeper during high-voltage slow activity than low-voltage fast activity (p < 0.001, t test). Table I compares both behavioral states for five fetal lambs. All regression lines achieved statistical significance (p < 0.01). The high-voltage slow activity (2.33 + 1.03) slope is significantly steeper than that of low-voltage fast activity (0.89 ± 0.37) (n = 21, P < 0.001).
In the basic norepinephrine dose-response experiment strong positive correlations were obtained between AFHR and norepinephrine dosage (2.5, 5.0, 7.5, 10.0, and 15.0 ILg) in both behavioral states (Fig. 3). Similarly, strong positive correlations were found between AMAP and norepinephrine dosage (Fig. 4). On the basis of these results, 7.5 ILg norepinephrine was selected as a standard dose because AFHR and AMAP at this level achieved approximately 50% of the maximal norepinephrine dosage response. In addition, strong positive correlations were found between AFHR and AMAP in both behavioral states (Fig. 5). From this result the ratio of AFHR and AMAP was also used as an index of baroreceptor reflex function.
To assess tachyphylaxis of the response reflex, AFHR, AMAP, and the ratio of AFHR/AMAP were compared for the first and last norepinephrine injection in both behavioral states. No significant difference (paired t test) was found between the values for the first and last norepinephrine injection (Table II).
Fig. 6 shows the ratio of AFHR/ AMAP in response to sequential standard norepinephrine injections for each animal. In 80 observations the ratio was significantly higher in high-voltage slow activity (2.73 ± 0.74 beats/min per 1 mm Hg) than in low-voltage fast activity state (1.85 ± 0.78 beats/min per I mm Hg) (p < 0.00 I, paired t test). Significant inverse relationships were found between the AFHR/ AMAP
Fig. 3. Relationship between 8FHR and norepinephrine (NE) doses (2.5, 5.0, 7.5, 10.0, and 15.0 f.Lg) in low-voltage fast activity (LVFA) and high-voltage slow activity (HSVA) state for three different fetal lambs.
ratio and gestational age III both behavioral states (Fig. 7).
Comment
Several investigators have confirmed the fall of FHR attributable to baroreceptor mechanisms·olO and that baroreceptor activity was present as early as 85 days of gestation.· Dawes et al. 1I reported that in the human the presence of decelerations during 28 to 41 weeks of gestation was common (18%) in records with normal neonatal outcome.
To determine the baroreceptor reflex function, two techniques were used in this study. First, in observations
824 Wakatsuki et al.
40
30
i'iI 1 20
~ 10
LVFA State
D 138 days: r=O.94 p<O.OI
• 135 days: r=O.96 p<O.OI
o 129 days: r=O.98 p<O.OO5
O~ __ ~ __ ~ __ ~~ __ ~ __ -L __ ~ __ __
0.0 2.5 5.0
40
30
10
7.5 10.0 12.5 15.0
NE Dosage (Ilg)
HVSA State
o
c 138 days: r=O.94 p<O.OI
• 135 days: r=O.97 p<O.OI
o 129 days: r=O.96 p<O.OI
O~ __ ~ __ ~ ____ ~ __ ~ ____ ~ __ ~ __ _
0.0 2.5 5.0 7.5 10.0 12.5 15.0
NE Dosage (Ilg)
Fig. 4. Relationship between ~MAP and norepinephrine (NE) doses (2.5, 5.0, 7.5, 10.0 and 15.0 f.Lg) in low-voltage fast activity (LVFA) and high-voltage slow activity (HVSA) state for three different fetal lambs.
of mean arterial pressure and FHR under physiologic conditions the slope of a linear regression line of FHR on mean arterial pressure was assessed. The slope was steeper in the high-voltage slow activity than in the lowvoltage fast activity state. This result suggests that baroreceptor reflex function is higher in the high-voltage slow activity than in the low-voltage fast activity state.
In 1980 Dawes et al. '2 reported the threshold for baroreceptor response to be above the range of the normal fetal arterial pressure. They suggested that baroreflexes would be elicited only by a large change
100
80
i 60
e cr:: 40 == '-<l
20
o
100
80
E 60 Q, e
cr:: 40 is <l
20
0 0
September 1992 Am J Dbstet Gynecol
LVFA State
c 138 days: r=O.96 p<O.OO5
• 135 days: r=O.99 p&.OO5
0 129 days: r=O.99 p<O.OOI
10 20
dMAP (mmHg)
HVSA State
c 138 days: r=O.98 p<O.OO5
• 135 days: r=O.97 p<O.OI o 129 days: r=O.99 p<O.OOI
10 20
dMAP (mmHg)
30 40
30 40
Fig. 5. Relationship between ~FHR and ~MAP (2.5, 5.0, 7.5, 10.0, and 15.0 f.Lg) in low-voltage fast activity (LVSA) and highvoltage slow activity (HSVA) state for three different fetal lambs.
in pressure and would not be important factors in normal physiologic FHR regulation or in buffering blood pressure in fetal life. In contrast, Itskovitz et al. 13 concluded that arterial baroreceptors play
an important role in regulating fetal circulation even under normal physiologic conditions. In the current study the linear regression between FHR and mean arterial pressure during physiologic conditions demonstrated that there is active baroreceptor regulation of the fetal cardiovascular system under physiologic conditions.
The relationship between cardiac rhythm and arte-
Volume 167 Number 3
en :c E E E
Co e Q.,
< ::; ~ CI: :c .... <l
5
4
3
2
0
Baroreceptor activity in fetal lamb 825
HV HV HV HV HV HV LV LV LV LV LV LV LV
Fig. 6. Ratio of 6.FHR to 6.MAP in response to 7.5 ILg norepinephrine injection to six animals at sequential behavioral states for each animal (high-voltage [HV] high-voltage slow activity; low-voltage [LV] , low-voltage fast activity). Ratio was significantly greater in high-voltage slow activity state (P < 0.001 , paired t test, 80 pairs).
Table II. Values of ~FHR, ~MAP, and ~FHR/ ~MAP ratio at first and last norepinephrine injections during low-voltage fast activity and high-voltage slow activity states
Low-voltage fast actil1it.v High-voltage slow aclivit)'
6.FHR (beats /min) 6.MAP (mm Hg) 6.FHR /6.MAP
(beats /min per 1 mm Hg)
Values are mean ± SD.
n
15 15 15
First norepinephrine
4 1.00 ± 13.77 22.54 ± 6.07
2.07 ± 0.99
rial blood pressure has been examined as an indicator of baroreceptor reflex functions in human'" 15 and animal6. R. 9. 12 . ... 16·19 studies. A number of workers have
studied baroreceptor behavior in the fetal and neonatal lamb.s.9. 12. 13. 16 The ratio of ~FHRI ~MAP after a-sympathetic stimulator injection has been confirmed to indicate baroreceptor reflex activi ty.6 Norepinephrine may exert direct l3-adrenergic effects on the myocardium. In the current study use of norepinephrine to elevate blood pressure resulted in a significant positive correlation between ~FHR and ~MAP at all norepinephrine dose levels. Wang and Borison20 reported that the earliest component influencing heart rate was vagus and that the sympathetic component developed more slowly. According to Coleman," pressor drug injection produced vagal reflex deceleration 1 second after the maximum blood pressure response, and this change in heart rate occurred before any appreciable l3-sympathetic component developed. There appears to be little
I Last
norepineph1'ine
39.67 ± 8.76 21.32 ± 5.24
2.09 ± 0.96
First norepinepln-ine
59.75 ± 17.88 2 1.54 ± 5. 19
3.08 ± 1.63
I Last
nm'epinephrine
57.06 ± 12.37 20.8 1 ± 5.44
3.06 ± 1.29
or no norepinephrine ~-adrenergic contribition in the ratio.
Norepinephrine injection was repeated two to 10 times during sequential behavioral states. To examine for tachyphylaxis , ~FHR, ~MAP, and the ratio of ~FHR/ ~MAP for first and last norepinephrine injections were compared. The absence of significant differences suggests minimal possibility of norepinephrine tachyphylaxis in the current study.
In 80 observations the ratio was significantly higher in the high-voltage slow activity than in the low-voltage fas t activity state. This suggests that the sensitivity of baroreceptor regulator function is higher in the highvoltage slow activity state. Dawes et al. ' 2 estimated baroreceptor sensitivity as the slope of heart period on arterial blood pressure produced by either balloon inflation or pressor drugs. The values they obtained did not differ during the low-voltage fast activity and highvoltage slow activity state. However, Zhu and Szeto"
826 Wakatsuki et al.
5
~ 4 ::c s i Q, 3 e
• I :
130 135 140
LVFA HVSA
145
September 1992 Am J Obstet Gynecol
Gestational Age (Days)
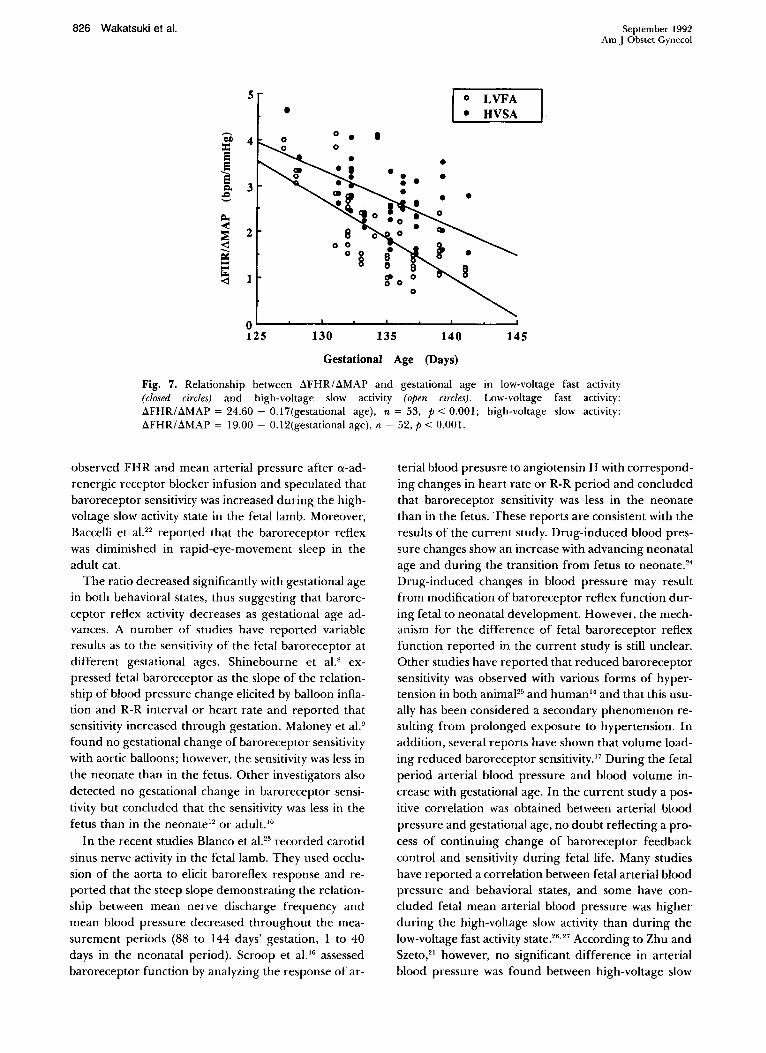
Fig. 7. Relationship between aFHRI aMAP and gestational age in low-voltage fast activity (closed circles) and high-voltage slow activity (open circles). Low-voltage fast activity: aFHRI aMAP = 24.60 - 0.17(gestational age), n = 53, P < 0.001; high-voltage slow activity: aFHR/aMAP = 19.00 - 0.12(gestational age), n = 52, P < 0.001.
observed FHR and mean arterial pressure after a-adrenergic receptor blocker infusion and speculated that baroreceptor sensitivity was increased during the highvoltage slow activity state in the fetal lamb. Moreover, Baccelli et al. 22 reported that the baroreceptor reflex was diminished in rapid-eye-movement sleep in the adult cat.
The ratio decreased significantly with gestational age in both behavioral states, thus suggesting that baroreceptor reflex activity decreases as gestational age advances. A number of studies have reported variable results as to the sensitivity of the fetal baroreceptor at different gestational ages. Shinebourne et al." expressed fetal baroreceptor as the slope of the relationship of blood pressure change elicited by balloon inflation and R-R interval or heart rate and reported that sensitivity increased through gestation. Maloney et al.9
found no gestational change of baroreceptor sensitivity with aortic balloons; however, the sensitivity was less in the neonate than in the fetus. Other investigators also detected no gestational change in baroreceptor sensitivity but concluded that the sensitivity was less in the fetus than in the neonate l2 or adult.!O
In the recent studies Blanco et aJ.23 recorded carotid sinus nerve activity in the fetal lamb. They used occlusion of the aorta to elicit baroreflex response and reported that the steep slope demonstrating the relationship between mean nerve discharge frequency and mean blood pressure decreased throughout the measurement periods (88 to 144 days' gestation, 1 to 40 days in the neonatal period). Scroop et al. 16 assessed baroreceptor function by analyzing the response of ar-
terial blood presusre to angiotensin II with corresponding changes in heart rate or R-R period and concluded that baroreceptor sensitivity was less in the neonate than in the fetus. These reports are consistent with the results of the current study. Drug-induced blood pressure changes show an increase with advancing neonatal age and during the transition from fetus to neonate." Drug-induced changes in blood pressure may result from modification of baroreceptor reflex function during fetal to neonatal development. However, the mechanism for the difference of fetal baroreceptor reflex function reported in the current study is still unclear. Other studies have reported that reduced baroreceptor sensitivity was observed with various forms of hypertension in both animaJ25 and human I' and that this usually has been considered a secondary phenomenon resulting from prolonged exposure to hypertension. In addition, several reports have shown that volume loading reduced baroreceptor sensitivity.17 During the fetal period arterial blood pressure and blood volume increase with gestational age. In the current study a positive correlation was obtained between arterial blood pressure and gestational age, no doubt reflecting a process of continuing change of baroreceptor feedback control and sensitivity during fetal life. Many studies have reported a correlation between fetal arterial blood pressure and behavioral states, and some have concluded fetal mean arterial blood pressure was higher during the high-voltage slow activity than during the low-voltage fast activity state.26,27 According to Zhu and Szeto,21 however, no significant difference in arterial blood pressure was found between high-voltage slow
Volume 167 Number 3
activity and low-voltage fast activity behavioral states. In the current study, similar to their report, no significant difference was observed between these behavioral states. The observed difference of baroreceptor sensitivity in both behavioral states remains to be explained.
REFERENCES
I. Swartwout JR, Campbell WE, Williams LG. Observations on the fetal heart rate. AM J OBSTET GVNECOL 1961;82:301-3.
2. Wheeler T, Murrillis A. Patterns of fetal heart rate during normal pregnancy. BrJ Obstet GynaecoI1978;85:18-27.
3. Sorokin Y, Dierker LJ, Pillay SK, Zador IE, Schreiner ML, Rosen MG. The association between fetal heart rate patterns and fetal movements in pregnancies between 20 and 30 weeks' gestation. AM J OBSTET GVNECOL 1982;143: 243-9.
4. Maurer M Jr. Developmental factors contributing to the susceptibility to bradycardia in isolated, cultured fetal mouse hearts. Pediatr Res 1979;13:1052-7.
5. Kattwinkel J, Fanaroff AA, Klaus MH. Bradycardia in pre term infants: indications and hazards of atropine therapy. Pediatrics 1976;58:494-9.
6. Coleman TG. Arterial baroreflex control of heart rate in the conscious rat. Am J Physiol 1980;238: H515-20.
7. Glantz SA. How to test for trends. In: Barry BK, White J, eds. Primer of biostatistics. New York: McGraw-Hili, 1987.
8. Shinebourne EA, Vapaavouri EK, Williams RL, Heymann MA, Rudolph AM. Development of baroreflex activity in unanesthetized fetal and neonatal lambs. Circ Res 1972;31 :710-8.
9. Maloney JE, Cannata J, Dowling MH, Else W, Ritchie B. Baroreflex activity in conscious fetal and neonatal lambs. Bioi Neonatal 1977;31:340-50.
10. Ismay MJ, Lumbers ER, Stevens AD. The action of angiotensin II on the baroreflex response of the conscious ewe and the conscious fetus. J Physiol (Lond) 1979;288:467-79.
II. Dawes GS, Houghton CR, Redman CW, Visser GH. Pattern of the normal human fetal heart rate. Br J Obstet Gynaecol 1982;89:276-84.
12. Dawes GS, Johnston BM, Walker DW. Relationshiop of arterial pressure and heart rate in fetal, new-born and adult sheep. J Physiol (Lond) 1980;309:405-17.
trol of the circulation in chronically instrumented fetal lamb. Circ Res 1983;52:589-96.
14. Wasserstrum N, Kirshon B, Rossavik IK, Willis RS, Moise KJ Jr, Cotton DB. Implications of sino-aortic baroreceptor reflex dysfunction in severe preeclampsia. Obstet Gynecol 1989;74:34-9.
15. Smyth HS, Sleight P, Pickering GW. Reflex regulations of arterial pressure during sleep in man. Circ Res 1969;24: 109-21.
16. Suoop GC, Marker JD, Stankewytsch-Janusch B, Seamark RF. Angiotensin I and II in the assessment of baroreceptor function in fetal and neonatal sheep. J Dev PhysioI1986;8:123-37.
17. Vatner SF, Boettcher DH, Hendrickx GR, McRitchie RJ. Reduced baroreflex sensitivity with volume loading in conscious dogs. Circ Res 1975;37:236-42.
18. Gardiner SM, Bennett T, Kemp PA. Systemic arterial hypertension in rats exposed to short-term isolation; intraarterial systolic and diastolic blood pressure and baroreflex sensitivity. Med Bioi 1980;58:232-9.
19. McRitchie RJ, Vatner SF, Heyndrickx GR, Braunwald E. The role of arterial baroreceptors in the regulation of arterial pressure in conscious dog. Circ Res 1976;39:666-70.
20. Wang SC, Borison HL. An analysis of the carotid sinus cardiovascular reflex mechanism. Am .I Physiol 1947; 150:712-21.
21. Zhu YS, Szeto HH. Cyclic variation in fetal heart rate and sympathetic activity. AM .I OBSTET GVNECOL 1987; 156:1001-5.
22. Baccelli G, Albertini R, Mancia G, Zanchetti A. Interactions between sino-aortic reflexes and cardiovascular effects of sleep and emotional behavior in the cat. Circ Res 1976;38 (suppl 2):30-4.
23. Blanco CE, Dawes GH, Hanson MA, McCooke HB. Carotid baroreceptor in fetal and newborn sheep. Pediatr Res 1988;24:342-6.
24. Woods JR Jr, Dandavino A, Murayama K, Brinkman CR III, Assali NS. Autonomic control of cardiovascular functions during neonatal development and in adult sheep. Circ Res 1977;40:401-7.
25. Fletcher PJ. Baroreceptor heart rate reflex in rabbits after reversal of renal hypertension. Am J Physiol 1984;246:H261-6.
26 . .lost RG, Quilligan EJ, Yeh SY, Anderson CG. Intrauterine electroencephalogram of the sheep fetus. AM .I OBSTET GV:-.IECOL 1972;114:535-9.
27. Mann LI, Duchin S, Weiss RR. Fetal EEG sleep stages and physiologic variability. AM .I OBSTET GVNECOL 1974; 119:533-8.