This article was downloaded by: [Syracuse University Library] On: 21 April 2013, At: 16:29 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Soil and Sediment Contamination: An International Journal Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/bssc20 Phytoextraction of Zinc: Physiological and Molecular Mechanism Rajesh Dhankhar a , Poonam Ahlawat Sainger a & Manish Sainger b a Evironmental Sciences Department, Maharshi Dayanand University, Rohtak, India b Centre for Biotechnology, Maharshi Dayanand University, Rohtak, India Version of record first published: 31 Jan 2012. To cite this article: Rajesh Dhankhar , Poonam Ahlawat Sainger & Manish Sainger (2012): Phytoextraction of Zinc: Physiological and Molecular Mechanism, Soil and Sediment Contamination: An International Journal, 21:1, 115-133 To link to this article: http://dx.doi.org/10.1080/15320383.2012.636983 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae, and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand, or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
Transcript
This article was downloaded by: [Syracuse University Library]On: 21 April 2013, At: 16:29Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Soil and Sediment Contamination: AnInternational JournalPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/bssc20
Phytoextraction of Zinc: Physiologicaland Molecular MechanismRajesh Dhankhar a , Poonam Ahlawat Sainger a & Manish Sainger ba Evironmental Sciences Department, Maharshi Dayanand University,Rohtak, Indiab Centre for Biotechnology, Maharshi Dayanand University, Rohtak,IndiaVersion of record first published: 31 Jan 2012.
To cite this article: Rajesh Dhankhar , Poonam Ahlawat Sainger & Manish Sainger (2012):Phytoextraction of Zinc: Physiological and Molecular Mechanism, Soil and Sediment Contamination: AnInternational Journal, 21:1, 115-133
To link to this article: http://dx.doi.org/10.1080/15320383.2012.636983
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representationthat the contents will be complete or accurate or up to date. The accuracy of anyinstructions, formulae, and drug doses should be independently verified with primarysources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand, or costs or damages whatsoever or howsoever caused arising directly orindirectly in connection with or arising out of the use of this material.
Phytoextraction of Zinc: Physiologicaland Molecular Mechanism
RAJESH DHANKHAR,1 POONAM AHLAWAT SAINGER,1
AND MANISH SAINGER2
1Evironmental Sciences Department, Maharshi Dayanand University,Rohtak, India2Centre for Biotechnology, Maharshi Dayanand University, Rohtak, India
Zinc is an essential trace element, necessary for plants, animals, and microorganisms.Zn is required for many enzymes as a catalytic cofactor, for photosynthetic CO2 fixa-tion, and in maintaining the integrity of bio-membranes. However, Zn is potentially toxicwhen accumulated beyond cellular needs. Phytoextraction technique, which is a part ofphytoremediation, has opened new avenues for remediation of Zn-contaminated places.Hyperaccumulators like Thlaspi caerulescens and Arabidopsis halleri have been iden-tified, which can accumulate up to 40,000 mg kg−1 Zn in the aerial parts of the plantbody. Carboxylic acids, primarily malate, citrate, and oxalate, and amino acids arefound to play an important role in Zn hyperaccumulation. Transmembrane metal trans-porters are assumed to play a key role in Zn metal uptake, xylem loading, and vacuolarsequestration. Members of CDF (cation diffusion facilitator) and ZIP (zinc-regulatedtransporter, iron-regulated transporter like protein) family have been implicated inZn-metal-tolerance mechanisms. A potential metal-binding motif, containing multiplehistidine residues, is found in the variable regions of almost all of the ZIP family,including ZIP1, ZIP2, ZIP4, ZRT1, and ZRT2. Overexpression of some Zn metal trans-porter genes like TcZNT1 (Thlaspi caerulescens Zn transporter1), TcHMA4 (Thlaspicaerulescens heavy metal ATPase) in Thlaspi caerulescens, AhMTP1;3 (Arabidopsishalleri metal transporter1;3) in Arabidopsis halleri, and PtdMTP1(Poplar metal trans-porter1) from a hybrid poplar confer Zn hypertolerance in Thlaspi, Arabidopsis, andPoplar plant species.
Keywords Hyperaccumulators, phytoremediation, hypertolerance, Zn, metal trans-porters
Introduction
Heavy metal pollution of the biosphere has accumulated rapidly since the onset of the in-dustrial revolution. There are various sources of anthropogenic metal contamination, whichinclude smelting of metalliferous ore, electroplating, gas exhaust, energy and fuel produc-tion, application of fertilizers, municipal sludges, and industrial manufacturing (Blaylockand Huang, 2000; Cunningham et al., 1997). Including these anthropogenic activities,substantial amounts of aqueous effluents discharge toxic elements and heavy metals into
Address correspondence to R. Dhankhar, Environmental Bioremediation Lab (37), Envi-ronmental Sciences Department, Maharshi Dayanand University, Rohtak 124001, India. E-mail:[email protected]
115
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
116 R. Dhankhar et al.
the environment. Although it may be possible to reduce the toxicity of certain metals byinfluencing their speciation, they do not degrade and still persist in the environment (NRC,1997). Heavy metals’ persistence in the environment will lead to the loss of biodiversity,amenity, and economic wealth (Bradshaw, 1993). Heavy metals cause serious damage thatis difficult to cure as metals cannot be chemically degraded (Salt et al., 1995). The mostcommon heavy metals (Cu, Cd, Zn, Mn, and Fe) are essential for plant growth in lowconcentration (Reeves and Baker, 2000). Beyond certain threshold concentration, theseelements become toxic for most of the plant species (Blaylock and Huang, 2000). The en-vironmental hazards of these metals depend upon geochemical and biochemical propertiesof a given metal and are related to several processes (Gupta and Sinha, 2006). Excessiveheavy metals in human nutrition can cause acute and chronic diseases. Elevated heavy metalconcentrations in the soil can also negatively affect crop growth. At higher concentrations,they interfere with metabolic processes, inhibit growth, and sometimes lead to plant death(Hoffmann, 1983).
In this review, we have confined our discussion mainly to phytoextraction of Zn metals,and physiological and molecular aspects of Zn hyperaccumulation. We have also discussedZn concentration levels in different environmental components and their importance forhuman beings.
Zinc: An Important Metal
Zinc is an essential trace element, necessary for plants, animals, and microorganisms. Zincis found in nearly 100 specific enzymes, serves as a structural ion in transcription factors,and is transferred in metallothioneins (Webb, 1992). It is the only metal represented in all sixenzyme classes (Webb, 1992). Around 10% of human proteins (2,800) potentially bind zincin addition to hundreds which transport and traffic zinc. In Arabidopsis thaliana 2,367 zinc-related proteins have been found (Broadley et al., 2007). The largest group of zinc-bindingproteins in Arabidopsis thaliana are Zn finger domains, assigned to transcription regulatoractivity and binding functional subcategories (Maret, 2005). Zn serves a purely structuralrole in zinc fingers, twists, and clusters and forms parts of some transcription factors(Cotton, 1997). Zn plays an important role in photosynthetic CO2 fixation in maintainingthe integrity of biomembranes (Brown et al., 1993; Jyung et al., 1972). Zn is an essentialtrace element for plants, but becomes toxic beyond a soil threshold range of 100 to 400 mgkg−1 (Kabata-Pendias and Pendias, 1984). Most rocks and many minerals contain zinc invarying amounts and zinc exists naturally in air, water, and soil (Van Assche et al., 1996).The average natural level of zinc in the earth’s crust is 70 mg kg−1 (dry weight), rangingbetween 10 and 300 mg kg−1 (Malle, 1992). Total Zn concentration varies among differentnatural components of the environment, as depicted in Table 1.
Zn: An Environment Contaminant
The world’s Zn production is continuously rising day by day. Sulfidic Zn and smelterslag produce a large amount of heavy metals in industries. After increasing 10-fold from1850, zinc emissions peaked at 3.4 megatonnes per year in the 1980s (Ensley, 2000).Anthropogenic and natural emissions occur at the ratio of 20:1 (Broadley et al., 2007). Totalemission of Zn into the atmosphere during smelting in USA during 1969 was estimatedat 50,000 tons. Levels of zinc in rivers flowing through industrial or mining areas canbe as high as 20 mg kg−1 (Ensley, 2000). Levels of zinc in excess of 500 mg kg−1 in
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
Phytoextraction of Zinc 117
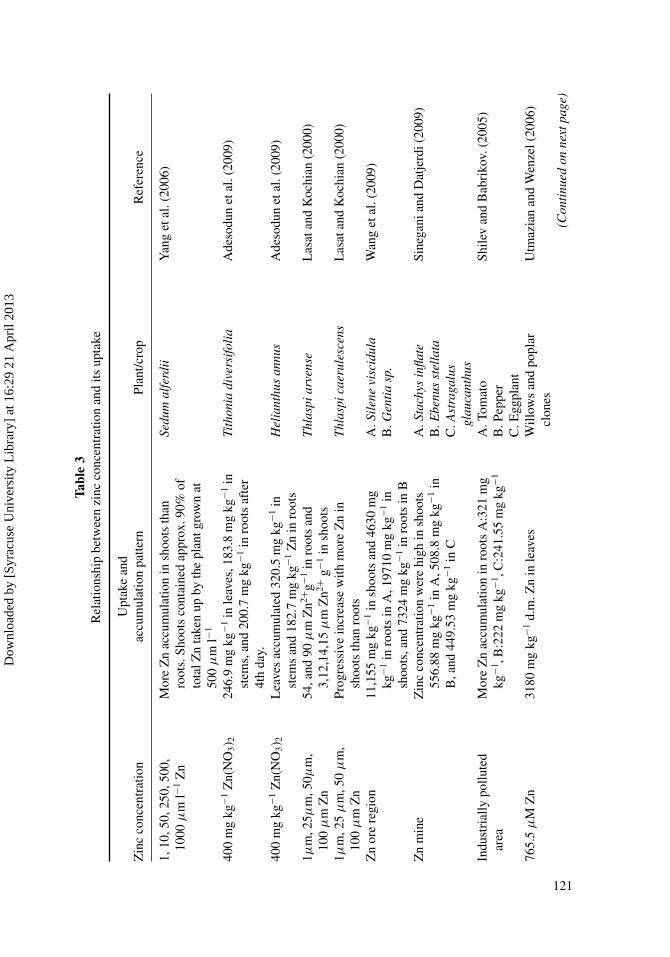
Table 1Natural zinc concentration in various components of environment
Natural zinc levels (total zinc) in the environment Range
a. Open ocean (surface) 0.001–0.06b. Coastal seas/inland seas 0.5–1c. Alluvial lowland rivers rich in nutrients and oligo-elements
(e.g. European lowland)5–40
d. Mountain rivers from old, strongly leached geologicalformations (e.g. Rocky Mountains)
<10
c. Large lakes (e.g. Great Lakes) 0.09–0.3d. Zinc-enriched streams flowing through mineralization areas >200
(From: Van Assche et al., 1996).
soil interfere with the ability of plants to absorb other essential metals, such as iron andmanganese (Broadley et al., 2007). Ensley (2000) showed that zinc levels of 2000 mg kg−1
to 180,000 mg kg−1 (18%) have been recorded in some soil samples and about 50 timeshigher Zn concentration (5000 mg kg−1) was found in soil close to smelter compared withcontrol area. Metal-contaminated soil can be remediated by various chemical and physicaltechniques such as thermal treatment, electrokinetical treatment, chemical oxidation, orreduction acid leaching, etc. Most of these conventional remediation technologies arecostly to implement and cause further disturbance to the already damaged environment.
Phytoremediation
A promising, relatively new technology for heavy-metal-contaminated sites is phytore-mediation. This technology has attracted attention in recent years for the low cost ofimplementation and environmental benefits. Moreover, the technology is likely to be moreacceptable to the public than other traditional methods such as excavation, land fill, leach-ing and virtification, etc. (Chaney et al., 1983; Baker et al., 1994). Phytoremediation andother bioremediation techniques are not only significantly cost-effective over physical andchemical means of soil, water, or air remediation; they also reduce the risk from exposureto the hazardous constituents at the waste and spill sites (Cunningham and Ow, 1996).Phytoextraction is an emerging technology where living plants are used to remove tracemetals from contaminated sites (Haque et al., 2008) and are then removed by harvesting theplants for subsequent volume reduction (i.e., ashing) and storage. In addition, plants needto survive in several odd environments, and hence they possess more flexible metabolic
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
118 R. Dhankhar et al.
Rhizofiltration: Root exudates complex with metals, thus bioavailability of metals decreases in soil/ground water
Phytoimmobilization:Fungi (myccorrhiza) immobilize metals in rhizosphere
Phytostabilization:The use of plants to reduce the bioavailability of pollutants in environment
Phytovolatilization:Selenium and mercury in soil and ground water are removed by evaporation
Phytoextraction:Uptake and bioconcentration of metals in abovegroundplant tissues from soils
PHYTOREMEDIATION TECHNOLOGY
Figure 1. Phytoremediation technology with various approaches.
systems that evolved genetically or adapted physiologically to avoid, partition, degrade,store, or exclude various undesired and toxic substances (Singh et al., 2007). Phytoremedi-ation is currently divided into different areas, which are shown in Figure 1.
Phytoextraction is the most commonly recognized of all phytoremediation technolo-gies. This technology uses pollutant-accumulating plants to remove metals from soil byconcentrating them in the harvestable parts (Salt et al., 1998). It is the best approach toremove contamination from soil and isolate it without destroying the soil structure and fer-tility (Ghosh and Singh, 2005), and best suited for the contamination of diffusely pollutedareas. The ideal plant for phytoextraction should grow rapidly, produce a high amount ofbiomass, and be able to tolerate and accumulate high concentrations of metals in shoots(Xiao et al., 2007). For phytoextraction, hyperaccumulators may be one of the best choicesfor this application because they can accumulate excessively high amounts of metals in theirharvestable parts, which are then easy to dispose (Zhao et al., 2003b; Wang et al., 2009).Hyperaccumulators are defined as higher plant species whose shoots contain >100 mg Cdkg−1, >1000 mg Ni, Pb, and Cu kg−1or >10000 mg Zn and Mn kg−1(dry wt.) when grownin metal-rich soils (Baker and Brooks, 1989). Approximately 400 taxa of terrestrial plantsfrom at least 45 families have been identified as hyperaccumulators of various heavy metals(Baker and Brooks, 1989). Only 16 species of Zn hyperaccumulators, which are definedas being able to accumulate more than 10,000 mg Zn kg−1 in the aboveground parts on adry weight basis in their natural habitat, have been reported (Kupper et al., 1999). Some ofthe Zn hyperaccumulators are listed in Table 2. The first hyperaccumulators characterizedwere members of the Brassicaceae and Fabaceae families (Salt et al., 1998).
Phytoextraction of Zn Metal
Thlaspi caerulescens J. and C. Presl. from the Brassicaeae family is the best known exampleof a Zn hyperaccumulator (Brown et al., 1995). Under hydroponic culture conditionsThlaspi caerulescens can accumulate up to 25,000 to 30,000 mg Zn kg−1 dry weight inshoots without showing any toxicity symptoms or reduction in growth, although Thlaspicaerulescens can accumulate a maximum dry weight of 40,000 mg kg−1 Zn in shoots (Penceet al., 2000). Lasat et al. (1996) reported that Thlaspi caerulescens exhibited a10-fold greaterZn translocation to the shoot then Thlaspi arvense (a non-hyperaccumulator of Zn). Similarresults were obtained when growth and uptake of Thlaspi. caerulescens (hyperaccumulator
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
Phytoextraction of Zinc 119
Table 2List of some zinc hyperaccumulator plants
Scientific name Common name Reference
Agrostis castellana Highland Bent Grass Cutcheon and Schnoor (2003)Arabidopsis halleri Wall cress; mouse-ear cress Bert et al. (2000)Athyrium yokoscense Japanese false spleenwort Cutcheon and Schnoor (2003)Brassica juncea Indian Mustard Schmidt (2003)Eichhornia crassipes Water hyacinth weed Cutcheon and Schnoor (2003)Helianthus annus Sunflower Schmidt (2003)
Cutcheon and Schnoor (2003)Salvinia molesta Kariba weeds or water ferns Cutcheon and Schnoor (2003)Salix viminalis Common Osier Schmidt (2003)Sedum alfredii Stonecrop Yang et al. (2004)Silene vulgaris Bladder campion Ernst et al. (1990)Spirodela polyrhiza Giant duck weed Cutcheon and Schnoor (2003)
Shrivastava et al. (1994)Thlaspi caerulescens Alpine pennycress Lombi et al. (2001)
Baker and Brooks (1989)Thlaspi rotundifolium Round-leaved Pennycress Reeves and Brooks (1983)Trifolium pratense Red clover Baker and Brooks (1989)
sp.) and Thlaspi ochroleucum Bioss and Heldr (a non-hyperaccumulator) were comparedin solution culture experiments. Thlaspi caerulescens was able to tolerate 500 mmol m−3
(32.5 g m−3) Zn in solution without growth reduction and up to 1000 mmol m−3 (65g m−3)Zn without showing visible toxic symptoms. Up to 28,000 mg kg−1 of Zn in shoots (drywt.) were obtained in healthy plants of Thlaspi caerulescens. A higher concentration of Znaccumulated in shoots of Thlaspi caerulescens, whereas Thlaspi ochroleucum accumulatedhigher concentrations of Zn in roots (Shen et al., 1997). Meerts and Isacker (1997) comparedZn uptake and tolerance of two subspecies of T. caerulescens, subsp. calaminiare frommetalliferous sites and subsp. caerulescens from non-metalliferous sites. Though subsp.calamine was more tolerant than subsp. caerulescens, the latter accumulated more Zn thanthe former (Zhao et al., 2003a). The bioaccumulation factor in accumulator species is oftengreater than one (Baker, 1981). Thlaspi caerulescens showed a bioaccumulation factorfor Zn as large as 40. Kupper et al. (1999) studies on sap samples of Thlaspi caerulescensused SEM-EDXMA (scanning electron microscopy-energy dispersive X-ray microanalysis)techniques and showed that the epidermis of leaves contained much greater Zn than themesophylls cells. In Thlaspi caerulescens, Zn hyperaccumulation and hypertolerance areconstitutive species-level traits, though inter- and intra-population variations in Zn toleranceoccur (Assuncao et al., 2006; Molitor et al., 2005). Thlaspi caerulescens does indeed havean enhanced requirement for this micronutrient due to the operation of a constitutive metalsequestration mechanism (Lloyd-Thomas, 1995). Thus active foraging for Zn by Thlaspicaerulescens roots is crucial for enhanced Zn accumulation that is necessary to provideadequate Zn availability in the cytosol of the species. According to Bert et al. (2000),Arabidopsis halleri is a well-known hyperaccumulator of Zn. It is one of the closest relativesof Arabidopsis thaliana, whose genome is entirely sequenced (Koch and Mummenhoff,2001). Arabidopsis halleri is a pseudo-metallophyte, which means that it is found in both
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
120 R. Dhankhar et al.
polluted and non-polluted areas (Sarret et al., 2002). Arabidopsis halleri can accumulate32,000 mg kg−1 DW Zn in the shoots with normal growth at 1000 µM Zn in nutrient solution(Zhao et al., 2000) and under field conditions, 15,000 mg kg−1 Zn in shoots (Bert et al.,2000). Studies of Arabidopsis halleri showed that Zn tolerance and Zn hyperaccumulationare two genetically independent characters (Macnair et al., 1999). Bert et al. (2000) showedthat like Thlaspi caerulescens both characters are constitutive properties of Arabodopsishalleri. Energy dispersive X-ray microanalysis studies revealed that Zn is sequestered inthe base of trichomes and mesophyll cells of Arabidopsis halleri grown in hydroponics(Kupper et al., 2000; Zhao et al., 2000). A plant species native to China, Sedum alferdiiHance has been identified as a new Zn hyperaccumulator. It has all the characteristicsof a hyperaccumulator, i.e. large biomass, fast growth, and asexual propagation (Yanget al., 2002). Sedum alferdii showed no visible symptoms of metal-induced toxicity whenexposed to 3671 µmol Zn l−1; the highest Zn concentration in shoots was 2% (Yang et al.,2002). Field experiments with a Zn-contaminated soil (2000 mg kg−1) showed that Sedumalferdii can concentrate Zn in shoots up to 10 times greater than that in soil (Yang et al.,2002). Sedum alferdii has elevated root foraging mechanisms associated with its greaterrequirement for Zn and allocates 90% of its root biomass to Zn-rich patches (Liu et al.,2009).
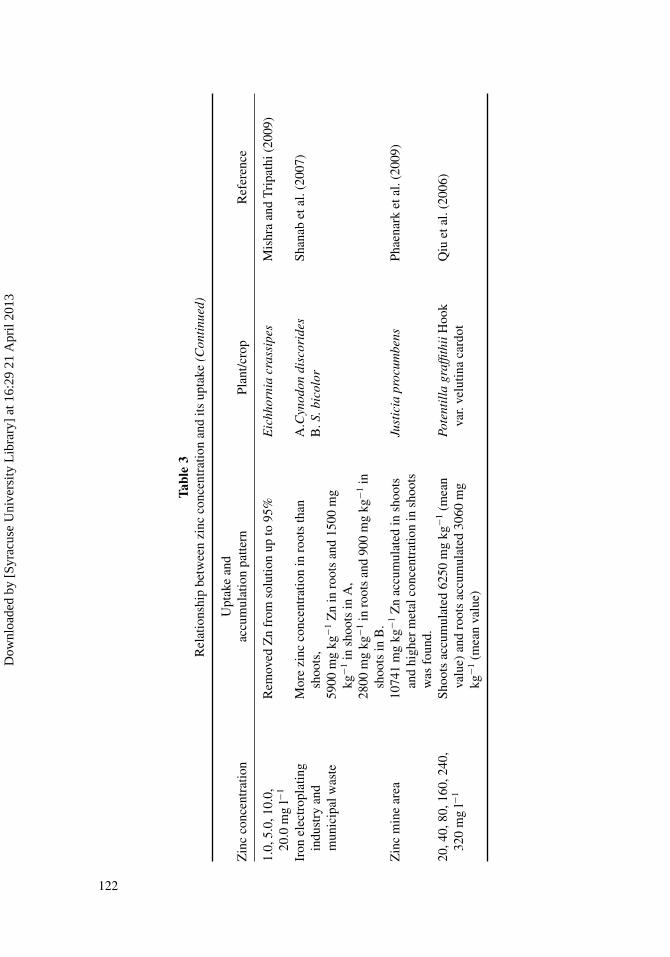
Many studies indicated that Sedum alferdii plant showed enhanced metal transportacross the root membrane, higher xylem loading, and increased metal sequestration in shoots(Li et al., 2005; Tian et al., 2009). Ebbs and Kochian (1998) investigated the toxicity of Zn inthree species from Brassica (B. juncea, B. rapa, and B. napus) and found that Brassica spp.are most effective in removing Zn from contaminated soil due to high biomass productionin comparison to Thlaspi caerulescens. Results from Coolang and Randle (2003) studiesshowed that Zn accumulation in the shoots of Brassica rapa increased linearly to increasingZn concentration up to 200 mg l−1 in the solution culture. Mild Zn toxicity symptoms wereexpressed with Zn concentration beginning at 5 mg l−1. Ariyakanon et al. (2003) concludedfrom their studies that Brassica species can accumulate Zn differently depending on theconcentration of Zn in the contaminated soil. Zn accumulating efficiency between Brassicajuncea Coss. and Brassica chinensis Linn. was compared. Brassica juncea accumulatedmore Zn (4825 mg kg−1 dry wt. plant) when 1000 mg of Zn was added, while Brassicachinensis accumulated more Zn (4766 mg kg−1 dry wt. plant) when 100 mg of Zn wasadded. The above plants could be used as effective hyperaccumulators for Zn-contaminatedsoil pollution. A hydroponic experiments comparing two kinds of grasses (Avena sativa andHordeum vulgare) to Indian mustard (Brassica juncea) indicated that although shoot Znconcentrations were greater for Indian mustard, the grasses were considerably tolerant to Zn(Ebbs and Kochian, 1998). It was demonstrated recently that poplar plants have the potentialto be used for plantations in Zn-contaminated soils (Di Baccio et al., 2003). Phaenark et al.(2009) studied many species in Thailand from a Zn mine area in Tak province. He foundthat Justicia procumbens was the only Zn hyperaccumulator in that area and was suitablefor phytoremediation of Zn-contaminated soil. Moreover, Justica procumbens had highaboveground and high propagation rates. Various plants with different Zn uptake ratio intheir tissues are summarized in Table 3.
Physiological Aspect of Zn Hyperaccumulation
Zn plays multiple roles in plant biochemical and physiological processes, therefore it isessential to understand the physiological mechanism of the plants’ uptake, translocation,
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
Tabl
e3
Rel
atio
nshi
pbe
twee
nzi
ncco
ncen
trat
ion
and
itsup
take
Upt
ake
and
Zin
cco
ncen
trat
ion
accu
mul
atio
npa
ttern
Plan
t/cro
pR
efer
ence
1,10
,50,
250,
500,
1000
µm
l−1Z
nM
ore
Zn
accu
mul
atio
nin
shoo
tsth
anro
ots.
Shoo
tsco
ntai
ned
appr
ox.9
0%of
tota
lZn
take
nup
byth
epl
antg
row
nat
500
µm
l−1
Sedu
mal
ferd
iiY
ang
etal
.(20
06)
400
mg
kg−1
Zn(
NO
3) 2
246.
9m
gkg
−1in
leav
es,1
83.8
mg
kg−1
inst
ems,
and
200.
7m
gkg
−1in
root
saf
ter
4th
day.
Tith
onia
dive
rsif
olia
Ade
sodu
net
al.(
2009
)
400
mg
kg−1
Zn(
NO
3) 2
Lea
ves
accu
mul
ated
320.
5m
gkg
−1in
stem
san
d18
2.7
mg
kg−1
Zn
inro
ots
Hel
iant
hus
annu
sA
deso
dun
etal
.(20
09)
1µm
,25µ
m,5
0µm
,10
0µ
mZ
n54
,and
90µ
mZ
n2+g−1
inro
ots
and
3,12
,14,
15µ
mZ
n2+g−1
insh
oots
Thl
aspi
arve
nse
Las
atan
dK
ochi
an(2
000)
1µm
,25
µm
,50
µm
,10
0µ
mZ
nPr
ogre
ssiv
ein
crea
sew
ithm
ore
Zn
insh
oots
than
root
sT
hlas
pica
erul
esce
nsL
asat
and
Koc
hian
(200
0)
Zn
ore
regi
on11
,155
mg
kg−1
insh
oots
and
4630
mg
kg−1
inro
ots
inA
,197
10m
gkg
−1in
shoo
ts,a
nd73
24m
gkg
−1in
root
sin
B
A.S
ilen
evi
scid
ula
B.G
enti
asp
.W
ang
etal
.(20
09)
Zn
min
eZ
inc
conc
entr
atio
nw
ere
high
insh
oots
556.
88m
gkg
−1in
A,5
08.8
mg
kg−1
inB
,and
449.
53m
gkg
−1in
C
A.S
tach
ysin
flate
B.E
benu
sst
ella
taC
.Ast
raga
lus
glau
cant
hus
Sine
gani
and
Dat
jerd
i(20
09)
Indu
stri
ally
pollu
ted
area
Mor
eZ
nac
cum
ulat
ion
inro
ots
A:3
21m
gkg
−1,B
:222
mg
kg−1
,C:2
41.5
5m
gkg
−1A
.Tom
ato
B.P
eppe
rC
.Egg
plan
t
Shile
van
dB
abri
kov.
(200
5)
765.
5µ
MZ
n31
80m
gkg
−1d.
m.Z
nin
leav
esW
illow
san
dpo
plar
clon
esU
tmaz
ian
and
Wen
zel(
2006
)
(Con
tinu
edon
next
page
)
121
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
Tabl
e3
Rel
atio
nshi
pbe
twee
nzi
ncco
ncen
trat
ion
and
itsup
take
(Con
tinu
ed)
Upt
ake
and
Zin
cco
ncen
trat
ion
accu
mul
atio
npa
ttern
Plan
t/cro
pR
efer
ence
1.0,
5.0,
10.0
,20
.0m
gl−1
Rem
oved
Zn
from
solu
tion
upto
95%
Eic
hhor
nia
cras
sipe
sM
ishr
aan
dT
ripa
thi(
2009
)
Iron
elec
trop
latin
gin
dust
ryan
dm
unic
ipal
was
te
Mor
ezi
ncco
ncen
trat
ion
inro
ots
than
shoo
ts,
5900
mg
kg−1
Zn
inro
ots
and
1500
mg
kg−1
insh
oots
inA
,28
00m
gkg
−1in
root
san
d90
0m
gkg
−1in
shoo
tsin
B.
A.C
ynod
ondi
scor
ides
B.S
.bic
olor
Shan
abet
al.(
2007
)
Zin
cm
ine
area
1074
1m
gkg
−1Z
nac
cum
ulat
edin
shoo
tsan
dhi
gher
met
alco
ncen
trat
ion
insh
oots
was
foun
d.
Just
icia
proc
umbe
nsPh
aena
rket
al.(
2009
)
20,4
0,80
,160
,240
,32
0m
gl−1
Shoo
tsac
cum
ulat
ed62
50m
gkg
−1(m
ean
valu
e)an
dro
ots
accu
mul
ated
3060
mg
kg−1
(mea
nva
lue)
Pote
ntil
lagr
affit
hiiH
ook
var.
velu
tina
card
otQ
iuet
al.(
2006
)
122
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
Phytoextraction of Zinc 123
and storage of Zn. A hypertolerance mechanism is required in metal hyperaccumulatorsto resist the potentially acute cytotoxic effects of the accumulated metals. The Zn metalis compartmentalized in the vacuole of hyperaccumulator species (Ma et al., 2005). Leafvacuoles are found to be primary sites for Zn sequestration in Thlaspi caerulescens (Kupperet al., 2000). Earlier studies showed that the meristematic cells of Festuca rubra roots showincreased vacuolation on treatment with Zn (Davies et al., 1991), while uptake analysisusing Zn65 with barley leaves suggested that rapid compartmentalization of Zn into thevacuole was an important mechanism for dealing with high levels of Zn (Brune et al.,1994). Analysis of transport systems at the tonoplast has added support to a vacuolarmechanism of tolerance. Verkleij et al. (1998) isolated tonoplast vesicles from roots ofZn-tolerant and sensitive ecotypes of Silene vulgaris (Verkleij et al., 1998; Hall, 2002).They showed that at high Zn concentrations Zn transport was 2.5 times higher into vesiclesfrom the tolerant lines than from the sensitive ones, suggesting that the tonoplast plays animportant role in naturally selected Zn tolerance.
Thlaspi caerulescens has been used primarily as a model system for the investigation ofunderlying physiological mechanisms of hyperaccumulation. Milner and Kochian (2008)suggested that the metal transport component of hyperaccumulation in Thlaspi caerulescensappears to involve at least four physiological processes. The first is a stimulated metalinflux across the root cell plasma membrane, the second reduced metal sequestration inthe root vacuole, the third increased loading into the xylem for transport to the shoots and,finally, the fourth involves stimulated metal influx across the leaf cell plasma membraneand sequestration in the leaf vacuole. In the root-cell plasma membrane, the density ofZn transporters was much higher in Zn hyperaccumulators than non-hyperaccumulators.Moreover, Zn uptake in roots appears to be regulated by Zn concentration in plant cells(Lasat et al., 1996). It was found that, along with much higher rates of Zn entry into theroot, the hyperaccumulator species also maintains the root Zn in a more mobile pool thatis moved to the xylem much more readily (Lasat et al., 1998). Recently, Klein et al. (2008)investigated in Thlaspi caerulescens leaf epidermal and mesophyll cells the ability to takeup the heavy metals from the soil and transport them to storage cells. A great numberof intermediary cells in the transport pathway from the root to the leaf epidermis do notretain the Zn, but rather keep it in a mobile pool to facilitate the very efficient root-to-shootmetal transport. Non-accumulator species such as Arabidopsis typically sequester heavymetals in the root to prevent the excess heavy metal from reaching the shoot and leaves.Studies by Kupper et al. (1999) indicated that highest concentrations of Zn are found inleaf epidermal cells, with concentrations four times that of mesophyll cells. This preferredstorage in leaf epidermal cells may be associated with avoidance of heavy metal damageto photosynthesis, although 65–70% of the total Zn is found in leaves’ mesophyll (Ma etal., 2005; Milner and Kochian, 2008). About 80% of shoot Zn is soluble in water or weakacids (Ma et al., 2005).
Until now it has not been clear whether the metals hyperaccumulated inside the cellsor bound to specific ligands or if they were only loosely associated with organic acids thatare abundant in the cell vacuole (Kramer et al., 1996). Kupper et al. (2004) reported thattotal sulfur (S) concentration in cells is lower than Zn and there is no correlation betweenthe concentrations of Zn and sulfur and, further, it was correlated that if Zn is supplied tothe plant, phytochelatins will not be expressed. Thus it is clear that major ligands for Znin hyperaccumulating plants must be different from phytochelatins (Satofuka et al., 2001;Kupper et al., 2000). The role of metallothioneins, which can bind to Zn, is not yet known(Callahan et al., 2006). The role of carboxylic acid, primarily malate, citrate, and oxalate,and amino acids in Zn hyperaccumulation has been widely studied by many authors (Tolra
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
124 R. Dhankhar et al.
et al., 1996; Kupper et al., 2004; Wojcik et al., 2006). Zn was mostly found coordinatedto malate in Arabidopsis halleri leaves (Sarret et al., 2002). In Thlaspi caerluescens, Znwas bound to form a Zn-histidine complex and in xylem sap most of the Zn exists as thefree hydrated Zn2+ cation with ∼20% as Zn citrate. In the leaves, all four forms are found,with citrate being the most common (Peer et al., 2003). In shoots of Thlaspi cearulescens,high concentrations of malate citrate, succinate, aconitate, and formate were detected onlyat high Zn concentrations (Tolra et al., 1996). In xylem saps, 21% of Zn was found to beassociated with Zn citrate. In the roots of T. caerulescens, organic acid concentrations aresubstantially lower than in shoots (Broadley et al., 2007).
Milner and Kochian (2008) also found that xylem loading is an important step inthe hyperaccumulation process; the enhanced xylem loading, triggered by exposure tohigh heavy metal levels for extended periods, may translate into improved heavy metaltolerance. Though the pathways of all these mechanisms are not clear, it might be expectedthat hyperaccumulation is due to the higher expression of several different transportersalong the metals transport pathway from the soil to the shoots.
Molecular Aspect of Zn Hyperaccumulation
Recent advances in our understanding of molecular aspects of plant Zn transport haveopened up new avenues for examining the plant mechanism for accumulation of Zn inhypertolerants. Zinc is taken up from soils by root membrane transport mechanisms. Theselectivity of these transporters determines whether other divalent cations are importedat the same time as zinc (Maser et al., 2001). The ionic selectivity of ion transportersis a very important characteristic for plants because it impinges on the plant’s toleranceand uptake of toxic minerals (Christianson, 1991; Alberts et al., 1998; Zhao et al., 2001).Though the molecular identity of specific zinc transporters has become evident in plants,we know little about how the structures of these proteins interact to create differencesin functional characteristics, such as ionic selectivity (Moreau et al., 2002; Vert et al.,2001; Ramesh et al., 2003). Plants have many kinds of zinc transporters and zinc channels(Vander Zaal et al., 1999). Typical zinc transporters include metal tolerance protein MTP2(Kramer, 2005; Blaudez et al., 2003) ZIP (ZRT, IRT like protein; ZRT, zinc regulatedprotein; IRT, iron regulated protein) and HMA (Heavy Metal ATPase) families (Williamset al., 2000). The ZIP family of zinc transporters is found in plants, bacteria, fungi, andhumans (Gaither and Eide, 2001). In plants, ZIP transporters have been described mainlyfrom dicots, including pea (Pisum sativa), Arabidopsis, and Thalaspi caerulescens (Eideet al., 1996; Grotz et al., 1998). ZIPs are likely to mediate cellular Zn uptake and HMAshave Zn transport functions throughout the cell. (Papayan and Kochian, 2004; Colangeloand Guerinor, 2006). ZIP family members play prominent roles in cytosolic zinc import,transporting zinc from outside the cell (Pence et al., 2000) or from within an intracellularcompartment (Gaither and Eide, 2001). Hyperaccumulators have found to be a higherexpression of some plant defensins (PDFs), which confer Zn tolerance and accumulationin heterologous systems and may act as blockers of Zn-permeable channels (Mirouze et al.,2006).
The first Zn transporter gene cloned from Thlaspi caerulescens was ZNT1 by Penceet al. (2000). TcZNT1 is a member of the ZIP family of transporter of proteins. The firstmembers of this family were identified based on homology to the high affinity Zn-uptaketransporter in yeast, ZRT1, and the Arabidopsis transporter IRT1 (Milner and Kochian,2008) and ZIP4 (Grotz et al., 1998). The predicted amino acid sequence of the ZNT1protein is 36% identical to IRT1 and 88% identical to ZIP4. Pence et al. (2000) found that
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
Phytoextraction of Zinc 125
TcZNT1 might be a major transporter mediating root Zn uptake from the soil. When Thlaspicaerulescens plants were grown at high Zn levels (50–1000 µM), expression was signifi-cantly higher and this hyperexpression of TcZNT1 may play a role in hyperaccumulation(Pence et al., 2000; Letham et al., 2005). In T. caerulescens roots, it was demonstrated thatthis increased gene expression is the basis for the increased Zn2+ uptake from the soil, and itis likely that the same mechanism underlies the enhanced Zn2+ uptake into leaf cells (Penceet al., 2000). ZNT1 expression was higher in the hyperaccumulator Thlaspi caerulescensthan in the non-accumulator Thlaspi arvense, possibly leading to a higher density ofZn transporters in the root cell plasma membrane (Pence et al., 2000). This difference inthe transporter concentration could account for the observation that the hyperaccumulatorand the non-accumulator have the same affinity for Zn but the hyperaccumulator has ahigher rate of uptake (Peer et al., 2003). Based on the 4- to 5-fold increase in Vmax forZn transport observed in Thlaspi caerulescens roots compared with Thlaspi arvense, theauthors have speculated that the enhanced Zn uptake in roots of T. caerulescens is due to anincreased abundance of transporters in root cells (Lasat and Kochian, 2000). The ZNT1 andZNT2 genes are clearly part of the ZIP-like gene family. They encode proteins with eightpredicted transmembrane domains, as found in the Arabidopsis ZIP4 protein and otherrelated ZIP proteins. The predicted proteins contain a histidine-rich region between trans-membrane domains III and IV, which is proposed to be the heavy metal binding sequence(Eng et al., 1998). Two other metal transporters that have received significant attentionin T. caerulescens are ZTP1/MTP1 and TcHMA4. ZTP1 is a homologue to Arabidopsistransporter ZAT1, which is thought to be involved in loading Zn into a vacuole (Maseret al., 2001; Persans et al., 2001). TcHMA4 was found to be expressed primarily in rootsand its expression is induced by both Zn-deficiency and high-Zn treatments (Papoyan andKochian, 2004). As efficient translocation of metals from the root to shoot is an importantpart of metal hyperaccumulators, it has been suggested that TcHMA4 may play a criticalrole in heavy metal transport to shoot during hyperaccumulation (Papoyan and Kochian,2004). In Arabidopsis, overexpression of HMA4 can increase transport of Zn to aerialtissues (Verret et al., 2004), and it has been proposed that HMA2 and HMA4 function,at least partially redundantly, in loading and/or unloading of Zn in vascular tissues forlong-distance transport (Haydon and Cobbett, 2007). AhHMA3 and AhHMA4 have beenidentified as transcripts expressed at elevated levels in Arabidopsis halleri compared toArabidopsis thaliana, and both confer Zn tolerance when expressed in yeast (Becker etal., 2004; Talke et al., 2006). In Arabidopsis halleri, high constitutive expression of theAhHMA4 gene was found to be necessary for both Zn hyperaccumulation and the fullextent of Zn hypertolerance (Barabasz et al., 2010; Mirouze et al., 2006). Vander Zaalet al. (1999) suggested that increased Zn transport into the vacuoles of Zn-tolerant speciessuch as Silene vulgaris might be attributable to a ZAT-like (Zn transporter of Arabidopsisthaliana) protein, although data on ZAT localization were lacking. ZRC1 in Saccharomycescerevisiae is localized to the vacuole membrane and thought to contribute to the storageof Zn ions (Li and Kaplan, 1998; MacDiarmid et al., 2002; Miyabe et al., 2001). MSC2,a third CDF member from S. cerevisiae, was shown to affect the cellular distribution ofzinc, particularly the zinc content of nuclei (Li and Kaplan, 2001). Grotz et al. (1998) haveisolated and functionally characterized three Zn transporter genes from Arabidopsis, calledZIP1, ZIP2, and ZIP3 (ZIP: ZRT, IRT-like protein), by functional complementation of ayeast mutant defective in Zn uptake. They also identified a related genomic DNA sequencepredicted to encode the ZIP4 protein. The proteins encoded by ZIP genes have a high degreeof similarity with the yeast ZRT1 and ZRT2 proteins that are involved in the high- andlow-affinity Zn uptake system (Zhao and Eide 1996), ZIP gene expression is Zn-regulated.
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
126 R. Dhankhar et al.
ZIP1 and ZIP3 are induced in roots and ZIP4 in both roots and shoots of Zn-limited plants.ZIP4 also contains a potential chloroplast targeting sequence to mediate transport of Zninto plastids (Grotz et al., 1998).
In Arabidopsis halleri, an AhMTP1;3 transporter is found that is localized on themembrane of central vacuoles (Drager et al., 2004). Becker et al. (2004) suggested that Zntolerance of A. halleri is due to an increased copy number of the MTP1 gene and enhancedlevel of transcription AhMTP1;3 has a great similarity with AtMTP1 and AtMTP3 (Ara-bidopsis thaliana) metal transporters. Both AtMTP1and AtMTP3 belong to a ubiquitousfamily of transition metal transporter proteins called the cation diffusion facilitator (CDF)protein family (Becker et al., 2004). MTP1 and MTP3 are predominantly expressed inroots and are induced by high Zn, manganese (Mn), and cobalt (Co), as well as iron (Fe)deficiency. It was suggested that MTP3 is important for sequestration of Zn in root vacuolesunder elevated Zn, as well as Fe deficiency (Arrivault et al., 2006). AhMTP1 is localizedto the tonoplast in A. thaliana protoplasts and Zn tolerance and elevated MTP1 transcriptco-segregated with duplicated copies of MTP1 in backcrossed populations, indicating animportant role for MTP1 in Zn tolerance in the hyperaccumulator A. halleri (Drager et al.,2004). In leaves of both A. halleri and T. caerulescens, genes encoding CDF transportersare more highly expressed than in nonaccumulator species (Becker et al., 2004; Hammondet al., 2006). These CDF protein families have been identified in bacteria, archea, andeukaryotes and have been demonstrated to transport Zn, Co, Cd (Paulsen and Saier, 1997;Kawachi et al., 2008). CDFs appear to mediate vacuolar sequestration of Zn efflux fromcytoplasm (Drager et al., 2004; Kramer, 2005; Colangelo and Guerinot, 2006). Zn bindsto many different types of proteins (Christianson, 1991; Alberts et al., 1998) and differentamino acid motifs that may function as zinc-binding sites and have been identified mainlyin soluble proteins (Alberts et al., 1998). In the majority of cases, histidine and cysteine arekey residues in binding Zn. It has been predicted that A. thaliana Zn transporter AtMTP1plays a key role in Zn tolerance, has six transmembrane domains, long N- and L- terminaltails, and a long histidine-rich (His-rich) hydrophilic region (Williams et al., 2000; Kawachiet al., 2008). Studies of Kawachi et al. (2008) indicated that AtMTP1 might function via aZn2+/H+ antiporter mechanism with a relatively high selectivity to Zn both in vitro and invivo. His-rich loop of AtMTP1 is richest in histidine residues among Zn transporters. TheHis-rich region functions as a concentration sensor and buffering pocket of cytoplasmicZn2+ to collect the excess ion in the vacuole. Thus the His-rich region has a physiologicalmerit in plant cells to keep the cytoplasmic concentration of Zn2+ at a low level and protectthe cells from Zn toxicity (Kawachi et al., 2008). Most of the heavy metals are not boundby strong ligands. Study of Kupper et al. (1999) found that from an energetic point ofview it is cheaper for the plant to pump the metals into the epidermal vacuoles and storethem there weakly bound by organic acids that also act as counterions rather than investingenergy for synthesizing the large amounts of strong ligands that would be required forbinding the metal. Kupper et al. (1999) also suggested that decrease in the proportion ofstrong ligands was found with increasing leaf age and it could be due to the increasingproportion of the total metal that is sequestered in the epidermal vacuoles. Recently, a newZn transporter, PtdMTP1, has been recognized. It is a member of the CDF family from thehybrid poplar Populus trichocarpa × Populus deltoides. PtdMTP1 is found to be expressedconstitutively and ubiquitously, although at low levels (Blaudez et al., 2003), and it wasidentified in a poplar EST database (Kohler et al., 2003). We found that PtdMTP1 possesseskey biochemical features: the expected CDF signature and Leucine zipper motifs, both ofwhich are necessary for its functional activity (Blaudez et al., 2003).
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
Phytoextraction of Zinc 127
Conclusion
In the past few years, phytoremediation has emerged as a suitable, inexpensive technologyand a viable alternative to conventional remediation methods. Phytoremediation is consid-ered the best approach to remove heavy metal pollution using hyperaccumulator plants.Phytoextraction research has focused on finding the ideal metal-accumulating plants andmetal uptake and translocation mechanisms. At present, Thlaspi caerulescens is amongthe most viable candidates for the phytoextraction of Zn. Many other plants, such as Ara-bidopsis halleri, Sedum alferdii, and Brassica spp., are also well-known hyperaccumulatorsof Zn. Heavy-metal hyperaccumulation in plants is an intriguing and poorly understoodphenomenon. More research is required for selection of suitable Zn hyperaccumulators.Progress is ongoing in understanding the role of genes involved in Zn hyperaccumulation,but still less is known about the biochemical processes responsible for Zn uptake, translo-cation, and storage. Therefore there is a need to isolate the genes and introduce these genesinto the high biomass plants to produce a plant that will extract large quantities of metalfrom soils.
References
Adesodun, J. K., Atayese, M. O., Agbaje, T. A., Osadiaye, B. A., Mafe, O. F., and Soretire, A. A.2010. Phytoremediation potentials of sunflowers (Tithonia diversifolia and Helianthus annus)for metals in soils contaminated with zinc and lead nitrates. Water, Air, and Soil Pollution 207,195–201.
Alberts, I. L., Nadassy, K., and Wodak, S. J. 1998. Analysis of zinc binding sites in protein crystalstructures. Protein Sci. 7, 1700–1716.
Ariyakanon, N., Soratana K., and Chunkrua T. 2003. Comparison of zinc accumulating efficiencybetween Brassica juncea Coss. and Brassica chinensis Linn. J. Sci. Res. 28, Special Issue I(NRC-EHWM).
Arrivault, S., Senger, T., and Kramer, U. 2006. The Arabidopsis metal tolerance protein AtMTP3maintains metal homeostasis by mediating Zn exclusion from the shoot under Fe deficiency andZn oversupply. Plant J. 46, 861–879.
Assuncao, A. G. L., Pieper, B., Vromans, J., Lindhout, P., Aarts, M. G. M., and Schat, H. 2006.Construction of a genetic linkage map of Thlaspi caerulescens and quantitative trait loci analysisof zinc accumulation. New Phytologist. 170, 21–32.
Baker, A. J. M. 1981. Accumulators and excluders—strategies in the response of plants to heavymetals. Journal of Plant Nutrition. 3, 643–654.
Baker, A. J. M., and Brooks, R. R. 1989. Terrestrial higher plants which hyperaccumulate metallicelements. A review of their distribution, ecology and phytochemistry. Biorecov. 1, 81–126.
Baker, A. J. M., McGrath, S. P., Sidoli, C. M.D., and Reeves, R. D. 1994. The possibility of in situheavy metal decontamination of polluted soils using crops of metal-accumulating plants. Resour.Conserv. Recyc. 11, 41–49.
Barabasz, A., Kramer, U., Hanikenne, M., Rudzka, J., and Antosiewicz, D. M. 2010. Metal accu-mulation in tobacco expressing Arabidopsis halleri metal hyperaccumulation gene depends onexternal supply. Journal of Experimental Botany. 61, 3057–3067.
Becker, M., Talke, I. N., Krall, L., and Kramer, U. 2004. Cross-species microarray transcript pro-filing reveals high constitutive expression of metal homeostasis genes in shoots of the zinchyperaccumulator Arabidopsis halleri. Plant Journal. 37, 251–268.
Bert, V., MacNair, M. R., Delaguerie, P., Saumitou-Laprade, P., and Petit, D. 2000. Zinc toleranceand accumulation in metallicolous and nonmetallicolous populations of Arabidopsis halleri(Brassicaceae). New Phytologist. 146, 225–233.
Blaudez, D., Kohler, A., Martin, F., Sanders, D., and Chalot, M. 2003. Poplar Metal Tolerance Protein1 confers zinc tolerance and is an oligomeric vacuolar zinc transporter with an essential leucinezipper motif. Plant Cell. 15, 2911–2928.
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
128 R. Dhankhar et al.
Blaylock, M. J., and Huang, J. W. 2000. Phytoextraction of metals. In: Raskin, I. and Ensley, B.D. (Ed.), Phytoremediation of Toxic Metals: Using Plants to Clean Up the Environment, JohnWiley & Sons, Inc., New York, pp. 53–70.
Bradshaw, A. D. 1993. Understanding the fundamentals of succession. In: Miles, J., Walton, D. H.(Eds.), Primary Succession on Land, Blackwell, Oxford.
Broadley, M. R., Philip, J., Hammond, J. P., Zelko, I. and Lux, A. 2007. Zinc in plants. NewPhytologist. 173, 677–702.
Brown, P. H., Cakmak, I., and Zhang, Q. 1993. Form and function of zinc within plants. In: Robson,A. D. (Ed.), Zinc in Soils and Plants, Kluwer Academic Publishers, Boston, pp. 93–106.
Brown, S. L., Chaney, R. L., Angle, J. S., and Baker, A. J. M. 1995. Zinc and cadmium uptakeby hyperaccumulator Thlaspi caerulescens and metal tolerant Silene vugaris grown on sludge-amended soils. Environ Sci Technol. 29, 1581–1585.
Brune, A., Urbach, W., and Dietz, K. J. 1994. Compartmentation and transport of zinc in barleyprimary leaves as basic mechanisms involved in zinc tolerance. Plant, Cell and Environment.17, 153–162.
Callahan, D. L., Baker, J. M., Kolev, S. P., and Wedd, A. G. 2006. Metal ion ligands in hyperaccu-mulating plants. Journal of Biological Inorganic Chemistry. 11, 2–12.
Chaney, R. L. 1983. Plant uptake of inorganic waste constitutes. In: Parr et al. (Eds.), Land Treatmentof Hazardous Wastes, Noyes Data Corporation, Park Ridge, NJ, pp. 50–76.
Christianson, D. W. 1991. Structural biology of zinc. Adv Protein Chem. 42, 281–355.Colangelo, E. P., and Guerinot, M. L. 2006. Put the metal to the petal: Metal uptake and transport
throughout plants. Current Opinion in Plant Biology. 9, 322–330.Coolang, T. W., and Randle, W. M. 2003. Zinc concentration in hydroponic solution culture influences
zinc and sulfur accumulation in Brassica rapa L. Journal of Plant Nutrition. 26, 949–959.Cotton, F., Wilkinson, A., Murillo, G., Carlos, A., and Manfred, B. 1997. Advanced Inorganic
Chemistry, (6th ed.), John Wiley & Sons, New York.Cunningham, S. D., and Ow, D. W. 1996. Promises and prospects of phytoremediation of contaminated
soils. Plants Physiol. 110, 715–719.Cunningham, S. D., Shann, J. R. Crowley, D. E. and Anderson, T. A. 1997. Phytoremediation
of contaminated water and soil. In: Kruger, E. L., Anderson, T. A., and Coats, J. R. (Eds.),Phytoremediation of Soil and Water Contaminants, ACS symposium series 664, AmericanChemical Society, Washington, DC, pp. 2–19.
Cutcheon, S. C., and Schnoor, J. L. (Eds.). 2003. Phytoremediation: Transformation and Control ofContaminants, Wiley-Interscience, Hoboken, NJ.
Davies, K. L., Davies, M. S., and Francis, D. 1991. Zinc-induced vacuolation in root meristematiccells of Festuca rubra L. Plant, Cell and Environment. 14, 399–406.
Di Baccio, D., Tognetti, R., Sebastiani, L., and Vitagliano, C. 2003. Responses of Populus deltoides ×Populus nigra (Populus × euramericana) clone I-214 to high zinc concentrations. New Phytol.159, 443–452.
Drager, D. B., Desbrosses-Fonrouge, A. G., Krach, C., Chardonnens, A. N., Meyer, R. C., Saumitou-Laprade, P., and Kramer, U. 2004. Two genes encoding Arabidopsis halleri MTP1 metal transportproteins co-segregate with zinc tolerance and account for high MTP1 transcript levels. Plant J.39, 425–439.
Ebbs, S. D., and Kochian, L. V. 1998. Phytoextraction of zinc by oat (Avena sativa), barley (Hordeumvulgare), and Indian mustard (Brassica juncea). Environ. Sci. Technol. 32, 802–806.
Eide, D., Broderius, M., and Fett, J., Guerinot, M. 1996. A novel iron-regulated metal transporter fromplants identified by functional expression in yeast. Proc Natl Acad Sci USA. 93, 5624–5628.
Eng, B. H., Guerinot, M. L., Eide, D., and Saier, M. H. Jr. 1998. Sequence analyses and phylogeneticcharacterization of the ZIP family of metal ion transport proteins. Journal of Membrane Biology.166, 1–7.
Ensley, B. D. 2000. Rational for use of phytoremediation. In: Raskin, I. and Ensley, B. D. (Eds.),Phytoremediation of Toxic Metals: Using Plants to Clean Up the Environment, John Wiley &Sons, Inc., New York, pp. 3–12.
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
Phytoextraction of Zinc 129
Ernst, W. H. O., Schat, H., and Verkleij, J. A. C. 1990. Evolutionary biology of metal resistance inSilene vulgaris. Evolutionary Trends in Plants. 4, 45–51.
Gaither, L. A., and Eide, D. J. 2001. Eukaryotic zinc transporters and their regulation. Biometals 14,251–270.
Ghosh, M. and Singh, S. P. 2005. A review on phytoremediation of heavy metals and utilization ofits byproducts. Applied Ecology and Environmental Research. 3, 1–18.
Grotz, N., Fox, T., Connolly, E., Park, W., Guerinot, M. L., and Eide, D. 1998. Identification of afamily of zinc transporter genes from Arabidopsis that respond to zinc deficiency. Proc NatlAcad Sci USA. 95, 7220–7224.
Gupta, A. K., and Sinha, S. 2006. Chemical fraction and heavy metal accumulation in the plant ofSesamum indicum (L.) var. T55 grown on soil amended with tannery sludge selection of singleextractants. Chemosphere. 64, 161–173.
Hall, J. E. 2002. Cellular mechanisms for heavy metal detoxification and tolerance. Journal ofExperimental Botany. 53, 1–11.
Hammond, J. P., Bowen, H. C., White, P. J., Mills, V., Pyke, K. A., Baker, A. J. M., Whiting, S. N.,May, S. T., and Broadley, M. R. 2006. A comparison of the Thlaspi caerulescens and Thlaspiarvense shoot transcriptomes. New Phytol. 170, 239–260.
Haque, N., Peralta-Videa, J. R., Jones, G. L., Gill, T. E., and Gardea-Torresdey, J. L. 2008. Screeningthe phytoremediation potential of desert broom (Braccharis sarathoides Gray) growing on minetailings in Arizona, USA. Environmental Pollution. 153, 362–368.
Haydon, J. M., and Cobbett, C. S. 2007. A novel major facilitator super family protein at thetonoplast influences zinc tolerance and accumulation Arabidopsis. Plant Physiology. 143, 1705–1719.
Hoffmann, G. 1983. Relationships between critical levels of pollutants in soils, fodder, and crops (inGerman.) Landwirtsch. Forsch. Sonderh. 39, 130–152.
Jyung, W. H., Camp, M. E., Polson, D. E., Afams, M. W., and Witter, S. H. 1972. Differential responseof two bean varieties to zinc as revealed by electrophoretic protein pattern. Crop Sci. 12, 26–29.
Kabata-Pendias, A., and Pendias, H. 1984. Trace Elements in Soils and Plants, CRC Press, BocaRaton, FL.
Kawachi, M., Kobae, Y., Mimura, T., and Maeshima, M. 2008. Deletion of a histidine-rich loopof AtMTP1, a vacuolar Zn2+/ H+ antiporter of Arabidopsis thaliana, stimulates the transportactivity. J. Biol. Chem. 283, 8374–8383.
Klein, M. A., Sekimoto, H., Milner, M. J., and Kochian, L. V. 2008. Investigation of heavy metal hy-peraccumulation at the cellular level: Development and characterization of Thlaspi caerulescenssuspension cell lines. Plant Physiology. 147, 2006–2016.
Koch, M., and Mummenhoff, K. 2001. Thlaspi s.str. (Brassicaceae) versus Thlaspi s.l. morphologicaland anatomical characters in the light of ITS nrDNA sequence data. Plant Systematics andEvolution. 227, 209–225.
Kohler, A., Delaruelle, C., Martin, D., Encelot, N., and Martin, F. 2003. The poplar root transcriptome:Analysis of 7000 expressed sequence tags. FEBS Lett. 542, 37–41.
Kramer, U., CottonHowells, J. D., Charnock, J. M., Baker, A. J. M., and Smith, J. A.C. 1996. Freehistidine as a metal chelator in plants that accumulate nickel. Nature. 379, 635–638.
Kramer, U. 2005. MTP1 mops up excess zinc in Arabidopsis cells. Trends in Plant Science. 10,313–315.
Kupper, H., Lombi, E., Zhao, F. J., and McGrath, S. P. 2000. Cellular compartmentation of cadmiumand zinc in relation to other elements in the hyperaccumulator Arabidopsis halleri. Planta. 212,75–84.
Kupper, H., Mijovilovich, A., Meyer-Klaucke, W., and Kroneck, P. M.H. 2004. Tissue- and age-dependent differences in the complexation of cadmium and zinc in the cadmium/zinc hyperac-cumulator Thlaspi caerulescens (Ganges ecotype) revealed by X-ray absorption spectroscopy.Plant Physiology. 134, 748–757.
Kupper, H., Zhao, F. J., and McGrath, S. P. 1999. Cellular compartmentation of zinc in leaves of thehyperaccumulator Thlaspi caerulescens. Plant Physiol. 119, 305–312.
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
130 R. Dhankhar et al.
Lasat, M. M., and Kochian, L. V. 2000. Physiology of Zn hyperaccumulation in Thlaspi caerulescens.In: Terry, N., Banuelos, G. (Eds.), Phytoremediation of Contaminated Soil and Water, CRC Press,Boca Raton, FL, pp. 159–169.
Lasat, M. M., Baker, A. J. M., and Kochian, L. V. 1998. Altered Zn compartmentation in the rootsymplasm and stimulated Zn absorption into the leaf as mechanisms involved in Zn hyperaccu-mulation in Thlaspi caerulescens. Plant Physiology. 118, 875–883.
Lasat, M. M., Baker, A. J. M., and Kochian, L. V. 1996. Physiological characterization of root Zn2+
absorption and translocation to shoots in Zn hyperaccumulator and non-accumulator species ofThlaspi. Plant Physiol. 112, 1715–1722.
Letham, D. L. D., Pence, N. S., Lasat, M. M., and Kochian, L. V. 2005. Molecular and physiologicalinvestigations of Thlaspi caerulescens, a Zn/Cd hyperaccumulator. In: Zobel, R. F. and Wright,S. F. (Eds.), Roots and Soil Management: Interactions Between Roots and the Soil, AgronomySociety of America, Madison, WI, pp. 95–106.
Li, L., and Kaplan, J. 1998. Defects in the yeast high affinity iron transport system result in increasedmetal sensitivity because of the increased expression of transporters with a broad transition metalspecificity. J. Biol. Chem. 273, 22181–22187.
Li, L., and Kaplan, J. 2001. The yeast gene MSC2, a member of the cation diffusion facilitator family,affects the cellular distribution of zinc. J. Biol. Chem. 276, 5036–5043.
Li, T. Q., Yang, X. E., Jin, X. F., He, Z. L., Stoffella, P. J., and Hu, Q. H. 2005. Root responses andmetal accumulation in two contrasting ecotypes of Sedum alfredii Hance under lead and zinctoxic stress. J Environ Sci Heal A. 40, 1081–1096.
Liu, F., Tang, Y., Du, R., Yang, H., Wu, Q., and Qiu, R. 2010. Root foraging for zinc and cadmiumrequirement in the Zn/Cd hyperaccumulator plant Sedum alfredii. Plant and Soil 327, 365–375.
Lloyd-Thomas, D. H. 1995. Heavy metal hyperaccumulation by Thlaspi caerulescens J. and C. Presl.PhD thesis, University of Sheffield, Sheffield, UK.
Lombi, E., Zhao, F. J., McGrath, S. P., Young, S. D., and Sacchi G. A. 2001. Physiological evidencefor a high-affinity cadmium transporter highly expressed in a Thlaspi caerulescens ecotype. NewPhytologist. 149, 53–60.
Ma, J. F., Ueno, D., Zhao, F. J., and McGrath, S. P. 2005. Subcellular localization of Cd and Zn inthe leaves of a Cd-hyperaccumulating ecotype of Thlaspi caerulescens. Planta. 220, 731–736.
MacDiarmid, C. W., Milanick, M. A., and Eide, D. J. 2002. Biochemical properties of vacuolar zinctransport systems of Saccharomyces cerevisiae. J. Biol. Chem. 277, 39187–39194.
Macnair, M., Bert, V., Huitson, S. B., Saumitou-Laprade, P., and Petit, D. 1999. Zinc tolerance andhyperaccumulation are genetically independent characters. Proc Royal Soc of London SeriesB-Biol Sci. 266, 2175–2179.
Malle, K. G. 1992. Zinc in the environment. Acta Hydrochim. Hydrobiol. 20, 196–204.Maret, W. 2005. Zinc coordination environments in proteins determine zinc functions. Journal of
Trace Elements in Medicine and Biology. 19, 7–12.Maser, P., Thomine, S., Schroeder, J. I., Ward, J. M., Hirschi, K., Sze, H., Amtmann, A., Maathuis,
F. J. M., Sanders, D., Harper, J. F., Tchieu, J., Gribskov, M., Persans, M. W., Salt, D. E., Kim,S. A., and Guerinot, M. L. 2001. Phylogenetic relationships within cation transporter familiesof Arabidopsis. Plant Physiology. 126, 1646–1667.
Meerts, P., and Van Isacker, N. 1997. Heavy metal tolerance and accumulation in metallicolous andnon-metallicolous populations of Thlaspi caerulescens from continental Europe. Plant Ecol.133, 221–231.
Milner, M. J., and Kochian, L. V. 2008. Investigating heavy-metal hyperaccumulation using Thlaspicaerulescens as a model system. Annals of Botany. 102, 3–13.
Mirouze, M., Sels, J., Richard, O., Czernic, P., Loubet, S., Jacquier, A., Francois, I. E. J. A., Cammue,B. P. A., Lebrun, M., Berthomieu, P., and Marques, L. 2006. A putative novel role for plantdefensins: A defensin from the zinc hyper-accumulating plant, Arabidopsis halleri, confers zinctolerance. Plant Journal. 47, 329–342.
Mishra, V. K., and Tripathi, B. D. 2009. Accumulation of chromium and zinc from aqueous solutionsusing water hyacinth (Eichhornia crassipes). J. Hazard Mater. 30, 1059–1063.
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
Phytoextraction of Zinc 131
Miyabe, S., Izawa, S., and Inoue, Y. 2001. The Zrc1 is involved in zinc transport system betweenvacuole and cytosol in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 282, 79–83.
Molitor, M., Dechamps, C., Gruber, W., and Meerts, P. 2005. Thlaspi caerulescens on nonmetallifer-ous soil in Luxembourg: Ecological niche and genetic variation in mineral element composition.New Phytologist. 165, 503–512.
Moreau, S., Thomson, R. M., Kaiser, B. N., Trevaskis, B., Guerinot, M. L., Udvardi, M. K., Puppo,A., and Day, D. A. 2002. GmZIP1 encodes a symbiosis-specific zinc transporter in soybean. JBiol Chem. 277, 4738–4746.
National Research Council. 1997. Challenges of groundwater and soil cleanup. In: Innovations inGroundwater and Soil Cleanup, National Academy Press, Washington, DC, pp. 18–41.
Papayan, A., and Kochian, L. N. 2004. Identification of Thlaspi caerulescens genes that may beinvolved in heavy metal hyperaccumulation and tolerance: Characterization of a novel heavymetal transporting ATPase. Plant Physiol. 136, 3814–3823.
Paulsen, I. T., and Saier, M. H. 1997. A novel family of ubiquitous heavy metal ion transport proteins.J. Membr. Biol. 156, 99–103.
Peer, W. A., Mamoudian, M., Lahner, B., Reeves, R. D., Murphy, A. S., and Salt, D. E. 2003.Identifying model metal hypetaccumulating plants: germplasm Analysis of 20 Brassicaceaeaccessions from a wide geographical area. New Phytologist. 159, 421–430.
Pence, N. S., Larsen, P. B., Ebbs, S. D., Letham, D. L., Lasat, M. M., Garvin, D. F., Eide, D.,and Kochian, L. V. 2000. The molecular physiology of heavy metal transport in the Zn/Cdhyperaccumulator Thlaspi caerulescens. Proceedings of the National Academy of Sciences. 97,4956–4960.
Persans, M. W., Nieman, K., and Salt, D. E. 2001. Functional activity and role of cation-effluxfamily members in Ni hyperaccumulation in Thlaspi goesingense. Proceedings of the NationalAcademy of Sciences, USA 98, 9995–10000.
Phaenark, C., Pokethitiyook, P., Kruatrachue, M., and Ngrnsansaruay, C. 2009. Cd and Zn accumula-tion in plants from the Padaeng zinc mine area. International Journal of Phytoremediation. 11,479–495.
Qiu, R., Fang, X., Tang, Y., Du, S., and Zeng, X. 2006. Zinc hyperaccumulation and uptake byPotentilla Griffithii Hook. International Journal of Phytoremediation. 8, 299–310.
Ramesh, S. A., Shin, R., Eide, D. J., and Schachtman, D. P. 2003. Differential metal selectivity andgene expression of two zinc transporters from rice. Plant Physiology. 133, 126–134.
Reeves, R. D, and Baker, A. J. M. 2000. Metal- accumulating plants. In Raskin, I. and Ensley, B.D. (Eds.), Phytoremediation of Toxic Metals: Using Plants to Clean Up the Environment, JohnWiley and Sons, New York, pp. 193–230.
Reeves, R. D., and Brooks, R. R. 1983. European species of Thlaspi L. (Cruciferae) as indicators ofnickel and zinc. Journal of Geobotanical Exploration. 18, 275–283.
Salt, D. E., Blaylock, M., Kumar, N. P.B.A., Dushenkov, V., Ensley, B. D. and Chet, I. 1995.Phytoremediation: A novel strategy for the removal of toxic metals from the environment usingplants. Biotechnology. 13, 468–474.
Salt, D. E., Smith, R. D., and Raskin, I. 1998. Phytoremediation. Annual Review of Plant Physiologyand Plant Molecular Biology 49, 643–668.
Sarret, G., Saumitou-Laprade, P., Bert, V., Proux, O., Hazemann, J. L., Traverse, A., Marcus, M. A.,and Manceau, A. 2002. Forms of zinc accumulated in the hyperaccumulator Arabidopsis halleri.Plant Physiol 130, 1815–1826.
Satofuka, H., Fukui, T., Takagi, M., Atomi, H., and Imanaka, T. 2001. Metal-binding properties ofphytochelatin-related peptides. J. Inorg. Biochem. 86, 595–602.
Schmidt, U. 2003. Enhancing phytoextraction: The effect of chemical soil manipulations on mobility,plant accumulation, and leaching of heavy metals. J. Environ. Qual. 32, 1939–1954.
Shanab, R. A., Ghenem, N., Ghanem, K., and Kolaiabe, A. 2007. Phytoremediation potential ofcrop and wild plants for multi-metal contaminanted soils. Research Journal of Agriculture andBiological Sciences. 3, 370–376.
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
132 R. Dhankhar et al.
Shen, Z. G., Zhao, F. J., and McGrath, S. P. 1997. Uptake and transport of zinc in the hyperaccumulatorThlaspi caerulescens and the non-hyperaccumulator Thlaspi ochroleucum. Plant Cell Environ.20, 898–906.
Shilev, S., and Babrikov, T. 2005. Heavy metal accumulation in Solanaceae-plants grown at con-taminated area. In: Gruev, B., Nikolova, M. and Donev, A. (Eds.), Proceedings of the BalkanScientific Conference of Biology in Plovdiv (Bulgaria), May 19–21, 2005, pp. 452–460, Bulgaria:John Hopkins University Press.
Shrivastava, R. K., Gupta S. K., Nygam, K. D. P., and Vasudevan P. 1994. Treatment of chromiumand nickel in wastewater by using aquatic plants. Water Resources. 28, 1631–1638.
Sinegani, A. S. and Datjerdi, F. S. 2009. The accumulation of zinc and nickel in Irankoh indigenousplant species on a contaminated land. Soil and Sediment Contamination. 18, 525–534.
Singh, R. P., Dhania, G., Sharma, A. and Jaiwal, P. K. 2007. Biotechnological approaches to im-prove phytoremediation efficiency for environment contaminants. In: Tripathi, S. N. and Tri-pathi, R. D. (Eds.), Environmental Bioremediation Technologies, Springer, Heidelberg, pp. 223–257.
Talke, I. N., Hanikenne, M., and Kramer, U. 2006. Zinc-dependent global transcriptional control,transcriptional deregulation, and higher gene copy number for genes in metal homeostasis of thehyperaccumulator Arabidopsis halleri. Plant Physiol. 142, 148–167.
Tian, S. K., Lu, L. L., Yang, X. E., Labavitch, J. M., Huang, Y. Y., and Brown, P. 2009. Stem and leafsequestration of zinc at the cellular level in the hyperaccumulator Sedum alfredii. New Phytol.182, 116–126.
Tolra, R. P., Poschenrieder, C., and Barcelo, J. 1996. Zinc hyperaccumulation in Thlaspi caerulescens.II. Influence on organic acids. J Plant Nutr. 19, 1541–1550.
Utmazian, M. N. D., and Wenzel, W. W. 2006. Phytoextraction of metal polluted soils in LatinAmerica. In: Proceedings of Environmental Applications of Poplar and Willow Working Party,May 18–20, 2006, Northern Ireland. Northern Ireland: International Poplar Commission,pp. 1–7.
Van Assche, F., Tilborg, W., and Waeterschoot, H. 1996. Environmental risk assessment for essentialelements: Case study zinc. In: Report of the International Workshop on Risk Assessment ofMetals and their Inorganic Compounds, ICME, Ottawa, pp. 171–180.
Verkleij, J. A. C., Koevoets, P. L. M., Mechteld, M. A., Blake-Kalff, M. M. A., and Chardonnens, A.N. 1998. Evidence for an important role of the tonoplast in the mechanism of naturally selectedzinc tolerance in Silene vulgaris. Journal of Plant Physiology. 153, 188–191.
Verret, F., Gravot, A., Auroy, P., Leonhardt, N., David, P., Nussaume, L., Vavasseur, A., and Richaud,P. 2004. Overexpression of AtHMA4 enhances root-to-shoot translocation of zinc and cadmiumand plant metal tolerance. FEBS Lett. 576, 306–312.
Vert, G., Briat, J. F., and Curie, C. 2001. Arabidopsis IRT2 gene encodes a root-periphery irontransporter. Plant J. 26, 181–189.
Wang, S. L., Liao, W. B., Yu, F. Q., Liao, B., and Shu, W. S. 2009. Hyperaccumulation of lead,zinc, and cadmium in plants growing on a lead/zinc outcrop in Yunnan Province, China. EnvironGeol. 58, 471–476.
Webb, E. C. 1992. Enzyme Nomenclature, Recommendations of the Nomenclature Committee of theInternational Union of Biochemistry and Molecular Biology, Academic Press, New York.
Williams, L. E., Pittman, J. K., and Hall, J. L. 2000. Emerging mechanisms for heavy metal transportin plants. Biochimica et Biophysica Acta- Biomembranea. 1465, 104–126.
Wojcik, M., Skorzynska-Polit, E., and Tukiendorf, A. 2006. Organic acids accumulation and antioxi-dant enzyme activities in Thlaspi caerulescens under Zn and Cd stress. Plant Growth Regulation48, 145–155.
Xiao, Y., Yan, J., and Zhen, H. 2007. Role of soil rhizobacteria in phytoremediation of heavy metalcontaminated soils. J. Zhejiang Univ. Sci. B. 8, 192–207.
Yang, X. E., Li, T. Q., Long, X. X., Xiong, Y. H., He, L. Z. and Stoffella, P. J. 2006. Dynamics ofzinc uptake and accumulation in the hyperaccumulating and non-hyperaccumulating ecotypesof Sedum alferdii Hance. Plant and Soil. 284, 109–119.
Dow
nloa
ded
by [
Syra
cuse
Uni
vers
ity L
ibra
ry]
at 1
6:29
21
Apr
il 20
13
Phytoextraction of Zinc 133
Yang, X. E., Long, X. X., Ni, W. Z., and Fu, C. X. 2002. Sedum alfredi H: A new zinc hyperaccumu-lating plant species native to China. Chinese Sci. Bull. 47, 1003–1006.
Yang, X. E., Ye, H. B., Long, X. X., He, B., He, Z. L., Stoffella, P. J., and Calvert, D. V. 2004.Uptake and accumulation of cadmium and zinc by Sedum alferdii Hance at different Cd/Znsupply levels. J. Plant Nutri. 27, 1963–1977.
Vander Zaal, B. J., Neuteboom, L. W., Pinas, J. E., Chardonnes, A. N., Schat, H., and Verkleji, J. A.C., et al 1999. Overexpression of a novel Arabidopsis gene related to putative zinc-transportergenes from animals can lead to enhanced zinc resistance and accumulation. Plant Physiology.119, 1047–1055.
Zhao, F., Wang, J., Barker, J. H. A., Schat, H., Bleeker, P. M., and McGrath, S. P. 2003a. The roleof phytochelatins in arsenic tolerance in the hyperaccumulator Pteris vittata. New Phytol, 159,403–410.
Zhao, F. J., Hamon, R. E., Lombi, E., McLaughlin, M. J., and McGrath, S. P. 2001. Characteristicsof cadmium uptake in two contrasting ecotypes of the hyper-accumulator Thlaspi caerulescens.J Exp Bot. 53, 535–543.
Zhao, F. J., Lombi E, Breedon, T., and McGrath, S. P. 2000. Zinc hyperaccumulation and cellulardistribution in Arabidopsis halleri. Plant Cell Environ. 23, 507–514.
Zhao, F. J., Lombi, E., and McGrath, S. P. 2003b. Assessing the potential for zinc and cadmiumphytoremediation with the hyperaccumulator Thlaspi caerulescens. Plant Soil. 249, 37–43.
Zhao, H. and Eide, D. 1996. The ZRT2 gene encodes the low affinity zinc transporter in Saccharomycescerevisiae. Journal of Biological Chemistry 271, 23203–23210.