Pigeons can discriminate group mates from strangers using the concept of familiarity Anna Wilkinson * , Hanna L. Specht, Ludwig Huber Department of Cognitive Biology, University of Vienna article info Article history: Received 4 February 2010 Initial acceptance 23 March 2010 Final acceptance 7 April 2010 Available online 21 May 2010 MS. number: 10-00082 Keywords: categorization Columba livia concept familiarity individual recognition pigeon It is widely accepted that group-living animals alter their behaviour towards a conspecific depending on whether it is known or unknown. To distinguish between group members and strangers in an efficient way, it would be adaptive for an animal to categorize conspecifics on the basis of familiarity. We investigated whether pigeons, Columba livia, are able to learn this categorization in a concept-like manner when trained with 2D images of conspecifics. Two of the six experimental pigeons were able correctly to classify test photographs of aviary mates which they had never seen as photographic stimuli before. Control animals, which had no real-life experience of the pictured conspecifics, all failed. This is a cognitively demanding task; not only are there no consistent visual features among group members, but photographic stimuli also present a much degraded version of the depicted pigeons. Further tests revealed that the familiarity concept could be transferred to nonconspecifics and that 24 h of exposure was enough for another pigeon to be classed as familiar. In sum, the findings show that pigeons (1) interpreted computer stimuli as representations of real entities, (2) possessed the ability to form a concept of familiarity and (3) were able to distinguish between social partners and strangers in nature. Ó 2010 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved. The primary function of the brain is to compute dynamic, predictive models of the environment. Being able to identify objects as members of known categories allows an organism to respond to them in appropriate ways. This allows a reduction in the vast amount of information that is perceived, and takes advantage of the fact that objects within the same category share many properties and behaviours. As such it plays a crucial role in a variety of natural contexts, serving as a prerequisite for recognizing food, predators and also conspecifics. Recognizing that food should be eaten and a predator avoided represents appropriate behaviour for that category; however, recognizing that an animal is a conspecific does not allow appropriate behaviour towards that individual. Conspecifics differ in their intentions making recognition of familiar animals critical for territorial behaviour, parental care and pair bond formation. To categorize a conspecific on the basis of familiarity cannot be accomplished using perceptual features but requires the use of the concept of familiarity. The use of a concept for discrimination goes beyond classifying the stimulus on the basis of visual features and allows correct classification of exemplars that bear no physical similarity to each other (Lea 1984; Huber, in press). This is considered higher order learning. Furthermore, the use of the familiarity concept requires the animal to recognize that the stimuli (e.g. photographs) represent real-life conspecifics, identify the specific individual and then classify the individual on the basis of familiarity. Pigeons, Columba livia, are good at categorizing photographic stimuli on the basis of perceptual similarity (e.g. Herrnstein & Loveland 1964). They have even been shown to discriminate between photographs of unfamiliar conspecifics (Nakamura et al. 2003) and could generalize this discrimination to novel angles of the pictured birds, although transfer was partial as they pecked significantly less in the positive test trials than in the positive training trials. However, this task did not require them to recognize that the image represented a specific individual or even a real entity. To see that a picture represents a real-life object is not a simple task. It requires dual representation; that is, an organism must mentally represent both the symbol itself and its relation to the referent (DeLoache 2000). This form of representational insight has been shown in a small number of mammals (e.g. Dasser 1987; Pokorny & de Waal 2009) and recently also in pigeons (Aust & Huber 2006), although the nature of this ability is still debated (Delius et al. 2000). Examination of the pigeons’ ability to discriminate photo- graphic images on the basis of familiarity has produced varying results. This research has largely focused on recognition of landscapes because of the pigeons’ extraordinary homing ability. * Correspondence: A. Wilkinson, Department of Cognitive Biology, University of Vienna, Althanstrasse 14, Vienna 1090, Austria. E-mail address: [email protected](A. Wilkinson). Contents lists available at ScienceDirect Animal Behaviour journal homepage: www.elsevier.com/locate/anbehav 0003-3472/$38.00 Ó 2010 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.anbehav.2010.04.006 Animal Behaviour 80 (2010) 109e115
0003-3472/$38.00 � 2010 The Association for the Studoi:10.1016/j.anbehav.2010.04.006
It is widely accepted that group-living animals alter their behaviour towards a conspecific depending onwhether it is known or unknown. To distinguish between group members and strangers in an efficientway, it would be adaptive for an animal to categorize conspecifics on the basis of familiarity. Weinvestigated whether pigeons, Columba livia, are able to learn this categorization in a concept-likemanner when trained with 2D images of conspecifics. Two of the six experimental pigeons were ablecorrectly to classify test photographs of aviary mates which they had never seen as photographic stimulibefore. Control animals, which had no real-life experience of the pictured conspecifics, all failed. This isa cognitively demanding task; not only are there no consistent visual features among group members,but photographic stimuli also present a much degraded version of the depicted pigeons. Further testsrevealed that the familiarity concept could be transferred to nonconspecifics and that 24 h of exposurewas enough for another pigeon to be classed as familiar. In sum, the findings show that pigeons (1)interpreted computer stimuli as representations of real entities, (2) possessed the ability to forma concept of familiarity and (3) were able to distinguish between social partners and strangers in nature.� 2010 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.
The primary function of the brain is to compute dynamic,predictive models of the environment. Being able to identifyobjects as members of known categories allows an organism torespond to them in appropriate ways. This allows a reduction in thevast amount of information that is perceived, and takes advantageof the fact that objects within the same category share manyproperties and behaviours. As such it plays a crucial role in a varietyof natural contexts, serving as a prerequisite for recognizing food,predators and also conspecifics. Recognizing that food should beeaten and a predator avoided represents appropriate behaviour forthat category; however, recognizing that an animal is a conspecificdoes not allow appropriate behaviour towards that individual.Conspecifics differ in their intentions making recognition offamiliar animals critical for territorial behaviour, parental care andpair bond formation.
To categorize a conspecific on the basis of familiarity cannot beaccomplished using perceptual features but requires the use of theconcept of familiarity. The use of a concept for discrimination goesbeyond classifying the stimulus on the basis of visual features andallows correct classification of exemplars that bear no physicalsimilarity to each other (Lea 1984; Huber, in press). This is
gnitive Biology, University of
ilkinson).
dy of Animal Behaviour. Published
considered higher order learning. Furthermore, the use of thefamiliarity concept requires the animal to recognize that the stimuli(e.g. photographs) represent real-life conspecifics, identify thespecific individual and then classify the individual on the basis offamiliarity.
Pigeons, Columba livia, are good at categorizing photographicstimuli on the basis of perceptual similarity (e.g. Herrnstein &Loveland 1964). They have even been shown to discriminatebetween photographs of unfamiliar conspecifics (Nakamura et al.2003) and could generalize this discrimination to novel angles ofthe pictured birds, although transfer was partial as they peckedsignificantly less in the positive test trials than in the positivetraining trials. However, this task did not require them to recognizethat the image represented a specific individual or even a realentity. To see that a picture represents a real-life object is nota simple task. It requires dual representation; that is, an organismmust mentally represent both the symbol itself and its relation tothe referent (DeLoache 2000). This form of representational insighthas been shown in a small number of mammals (e.g. Dasser 1987;Pokorny & de Waal 2009) and recently also in pigeons (Aust &Huber 2006), although the nature of this ability is still debated(Delius et al. 2000).
Examination of the pigeons’ ability to discriminate photo-graphic images on the basis of familiarity has produced varyingresults. This research has largely focused on recognition oflandscapes because of the pigeons’ extraordinary homing ability.
Figure 1. Housing conditions of the pigeon groups. Each rectangle represents a pigeonaviary. Black rectangle: aviary of the control group (3 � 1.1 m � 3 m high). Stripedrectangle: aviary of the experimental group (2 � 2 � 2 m); birds living in this aviaryserved as experimental and stimulus pigeons. Chequered rectangle: visually separatedaviary; the pigeons in this aviary were used as the unfamiliar stimulus birds. Stippledrectangle: aviary of the semifamiliar stimulus pigeons.
A. Wilkinson et al. / Animal Behaviour 80 (2010) 109e115110
Wilkie et al. (1989) and Kendrick (1992) found evidence forfamiliarity effects when pigeons were asked to discriminatebetween familiar and unfamiliar landscapes. However, thesestudies lacked critical controls concerning the breed of pigeonused (all of the landscape-familiar birds were homing pigeonswhereas three of the four landscape-unfamiliar birds were not),the visual experience of the birds (Wilkie et al. 1989) and thetime period over which the experiments were run (Kendrick1992). Later experiments (Dawkins et al. 1996) found no effectof previous experience with the pictured landscape on discrim-inative performance; however, the pigeons were given very littleexposure to their familiar landscape. Given the varying reports ofthis ability using landscape-type stimuli and the pigeons’ socialnature it is possible that social stimuli are more salient.
Pigeons form loosely structured social groups, based onmonogamous pair bonds between the adults. Within a breedingcolony a clear dominance hierarchy exists, with males competingfor optimal breeding sites, and showing pronounced territorialbehaviour (Vriends 1988). Thus, recognition of familiar individualsis essential for pigeon social life. Studies on social recognition inthe chicken, Gallus gallus domesticus, have shown that they tend toaggregate more with familiar conspecifics than unfamiliar ones(Bradshaw 1992). Furthermore, they choose flockmates ratherthan unfamiliar birds as feeding companions, but this preferencefell to chance levels when photographs of the birds were used(Dawkins 1996). When birds were trained to discriminatebetween either photographs of familiar hens or photographs ofunfamiliar hens (Bradshaw & Dawkins 1993), there was nodifference in speed of learning between groups. Furthermore,transfer to novel views was no better for those discriminatingfamiliar birds than for those discriminating unfamiliar ones.Attempts using video stimuli also failed (D’Eath & Dawkins 1996).Thus it appears that hens are attracted to familiar individuals, butcannot discriminate familiar and unfamiliar individuals whenpresented as photographic stimuli. In contrast to this, there isevidence that some mammals are able to perceive a correspon-dence between photographs of a conspecific and the real animal(Macaca fascicularis: Dasser 1987; Pan troglodytes: Boysen &Berntson 1989; Parr & de Waal 1999; Ovis aries: Kendrick et al.1996). Furthermore, recent work with capuchin monkeys, Cebusapella, has revealed that they are able to categorize photographs ofconspecific faces on the basis of whether they are a group memberor an outsider (Pokorny & de Waal 2009).
We examined whether pigeons could use their real-life socialexperience with conspecifics when discriminating photographs onthe basis of familiarity. In a two-choice touch-screen set-up, birds ofthe experimental groupwere presented with two photographs, onedepicting a familiar pigeon and the other an unfamiliar pigeon. Thecontrol group were unfamiliar to all pictured individuals. Weexpected both groups to learn the discrimination and transfer tonovel views of the trained stimuli (thus replicating the results ofNakamura et al. 2003). Those birds that passed the generalizationtest were then given the critical familiarity test that presentedphotographs of pigeons that had never been used as trainingstimuli. Two were aviary mates of the experimental group and twowere entirely unfamiliar. Correct discrimination of these stimuli ispossible only if the pigeons are able to access their real-life expe-rience with these individuals.
METHODS
Subjects
We assigned 12 adult homing pigeons to either the controlgroup (N ¼ 6) or the experimental group (N ¼ 6) based on their
real-life experience. The experimental subjects were kept in anoutdoor aviary (2 � 2 � 2 m) in a flock of 11 pigeons. All experi-mental subjects had shared their aviary for at least 2 years. Thesepigeons had daily visual, auditory and social contact with eachother and group members were highly familiar. The controlsubjects were housed in a group of 13 pigeons in an aviary(3 � 1.1 m and 3 m high) located 5.6 m from the aviary of theexperimental group (Fig. 1). Opaque barriers surrounding theaviaries resulted in severely restricted visual contact and no socialcontact between members of the control and experimental group.The aviaries contained perches and nestboxes that provided shelterand nesting opportunities for the birds. The aviaries were alsoroofed and some of the side sections were covered with plastic toprovide further protection from the elements.
On training days, food (mixed grain) was provided in theexperimental sessions. The birds also received some additionalfeeding after they had completed their training. On days when thepigeons did not take part in experiments they were supplied withextra rations of food. The subjects were maintained at or above 90%of their free feeding weight. Water and grit were freely available inthe aviary at all times. The experiment described here did notrequire a licence and complies with the laws of Austria, where itwas carried out.
Both groups of subjects had experience with visual discrimi-nation tasks such as tasks involving a person/nonperson conceptand/or face discrimination. None had participated in experimentsthat required the discrimination of conspecifics or a familiarity rule.
Stimuli
Colour photographs of homing pigeons were used as stimuli.The six subjects of the experimental group were all photographed
A. Wilkinson et al. / Animal Behaviour 80 (2010) 109e115 111
and used as stimulus birds. As we could not present the experi-mental subject with photographs of itself, a seventh pigeon (a birdhoused in the same aviary under the same conditions) wasincluded in the stimulus set. The second set of images pictured sixindividuals that were reared in a visually separated compartment(Fig. 1). None of the subjects had ever seen or interacted with thesepigeons. Thus, the photographed stimulus pigeons provided twocategories of photographs: first, pictures that showed pigeons thatneither the control group nor the experimental group had ever
Figure 2. Examples of the stimulus sets. The picture on the left of each stimulus pair showsconspecific taken from the corresponding perspective.
seen or interacted with. The other stimulus class showed pigeonsthat were highly familiar to the subjects of the experimental groupbut entirely unknown to the control group. The stimuli consisted of12 photographs of each of the 13 birds from at least eight differentperspectives. Each stimulus measured 3.8 � 3.8 cm. To ensure nospurious background information could be used for classification,the natural background of the photograph was removed andreplaced by a uniform green background (Fig. 2 shows someexamples).
a flockmate of the experimental group while that on the right represents an unfamiliar
A. Wilkinson et al. / Animal Behaviour 80 (2010) 109e115112
Apparatus
The pigeons were trained in a ‘Skinner box’, the front wall ofwhich was made up of a 15 inch TFT computer screen that wasmounted behind an infrared touch-frame. Food reward wasadministered using a special feeder, the ‘grain lifter’. It consisted ofan electric motor that lifted a piston with a depression on topthrough a food reservoir. The piston was then lifted through a holein the bottom of the test chamber and the grainwas made availabledirectly below the touch-screen. The Skinner box was controlledusing ‘embedded C-Lab’ computer hardware and the ‘CognitionLab’software package (both developed by M. Steurer, Department ofCognitive Biology, University of Vienna, Austria).
Procedure
The pigeons were trained with a two-alternative forced-choiceprocedure in which two photographs were presented on a touch-screen computer monitor, one positive and one negative. Choice ofthe positive stimulus led to an auditory signal, the screen clearingand 3 s of access to food. Choice of the negative stimulus led toa different auditory signal, the screen flashing red (3 s) anda correction trial (a repeat of the same trial). This continued untilthe positive stimulus was selected. Each trial was separated by anintertrial interval of 6 s; during this time the screen was dark.Reward contingencies were counterbalanced, so half of the exper-imental group were rewarded for choosing the familiar pigeon andhalf for choosing the unfamiliar pigeon (Table 1). To control forpossible perceptual cues in the images each control bird was pre-sented with identical stimuli and contingencies as a correspondingexperimental bird.
Discrimination trainingPictures of four of the six experimental group pigeons and four
of the six entirely unknown pigeons served as training stimuli foreach subject. Only 10 of the 12 views of each pigeon were usedduring training. Thus, a training session consisted of 40 novel trials.Each trial included a picture of an entirely unknown pigeon anda bird familiar to the experimental group. Order of presentation andpairing of the stimuli varied randomly. Subjects received one or twotraining sessions per day. They were trained for a minimum of 20sessions, after which they were tested if they had met a criterion of75% correct in four of five consecutive sessions, and at least 70%correct in the remaining one. This criterion was used to ensure thatthe birds consistently performed at high levels. If a subject did notreach criterion after 100 sessions it was excluded from theexperiment.
Generalization testSubjects that met the learning criterion were given a general-
ization test in which the remaining two (untrained) views of eachstimulus bird were presented. Test sessions consisted of 48 trialswith 40 training trials and eight randomly intermixed test trials.
Table 1Sex and reinforcement contingency (Sþ) of the experimental and control subjects
Experimental group Control group
Subject Sex Sþ Subject Sex SþAgnes Female Unfamiliar Arthur Male UnfamiliarFred Male Unfamiliar Birgit Female UnfamiliarGeorge Female Unfamiliar Ferdinand Male UnfamiliarHeinz Male Familiar Heidrun Male FamiliarJudith Female Familiar Meggie Male FamiliarVanessa Female Familiar Trisha Female Familiar
These contained a novel picture of one of the unknown pigeons anda novel image of a bird familiar to the experimental group (thespecific stimuli used varied randomly across pigeons). Each testphotograph was presented three times (once per session, eachpresentation in combination with a different test stimulus), so thatthe birds received a total of 24 test trials over three sessions.Pecking on test stimuli did not result in differential feedback, so thepigeons could not learn about these stimuli. Subjects were giventwo opportunities to pass this test. Those that failed were excludedfrom the remainder of the experiment.
Familiarity testThe second, critical, test examinedwhether thepigeonswere able
to categorize conspecifics on the basis of familiarity. Subjects werepresented with photographs of two entirely unknown pigeons andtwo birds that were familiar to the experimental group. These birdshad never previously been shown as photographic stimuli. All 12views of each of the four birds were used. The test trials were pre-sented in the same manner as the generalization test. Subjects thatfailed this test were excluded from further testing.
Further testsThe pigeons that passed the familiarity test were given a series
of further tests. These were presented in the same manner as thefamiliarity test.
(1) Semifamiliar test: to examine whether social contact wasnecessary for a pigeon to be classed as familiar, the subject pigeonswerepresentedwithphotographsof twopigeons that livedoppositethem (versus two novel unfamiliar pigeons). They were in constantvisual contact, but had no social contact with these birds (see Fig.1).
(2) Familiar object test: to examine whether the birds hadlearned specifically about conspecifics the subject pigeons werepresented with photographs of familiar and unfamiliar objects. Thefamiliar objects were highly salient to the pigeons (water dispenserand food trough), and the unfamiliar (Wellington boot and spade)were entirely unknown.
(3) Novel pigeon test: to examine how quickly a conspecificwould be classed as familiar the pigeons were introduced to anentirely unknown pigeon and tested initially after 24 h and againafter 1week. Only one newpigeonwas introduced to the aviary andtherefore the 12 different photographs of eight different views ofthis pigeon were presented twice. This resulted in 24 test trials.
Data Analysis
We expected all birds to generalize to the novel instances of thestimulus pigeons used in training (Nakamura et al. 2003); however,if the discrimination were based on familiarity only the experi-mental birds would be able to pass the familiarity test. Thediscrimination performance was assessed by means of a two-tailedbinomial test in all cases except the first generalization test. Thiswas assessed using a one-tailed binomial test as all birds wereexpected to generalize to novel views of the stimulus pigeons, andso the hypothesis was not bidirectional. To reach statistical signif-icance in the training trials the birds had to choose the correctstimulus on 26 of 40 trials, for the generalization test they had tochoose 17 of 24 trials correctly and for the remaining tests they hadto choose 18 of 24 trials correctly.
RESULTS
Discrimination Training
There was no difference in performance between those birdsthat were rewarded for pecking on the familiar stimuli and those
0
20
40
60
80
100
Agnes Fred George Judith Vanessa
∗∗∗ ∗∗∗ ∗∗∗ ∗∗∗ ∗∗∗∗ ∗∗
0
20
40
60
80
100
Birgit Heidrun Trisha
Subjects
% C
orre
ct c
hoi
ces
∗∗∗ ∗∗∗ ∗∗∗
(a)
(b)
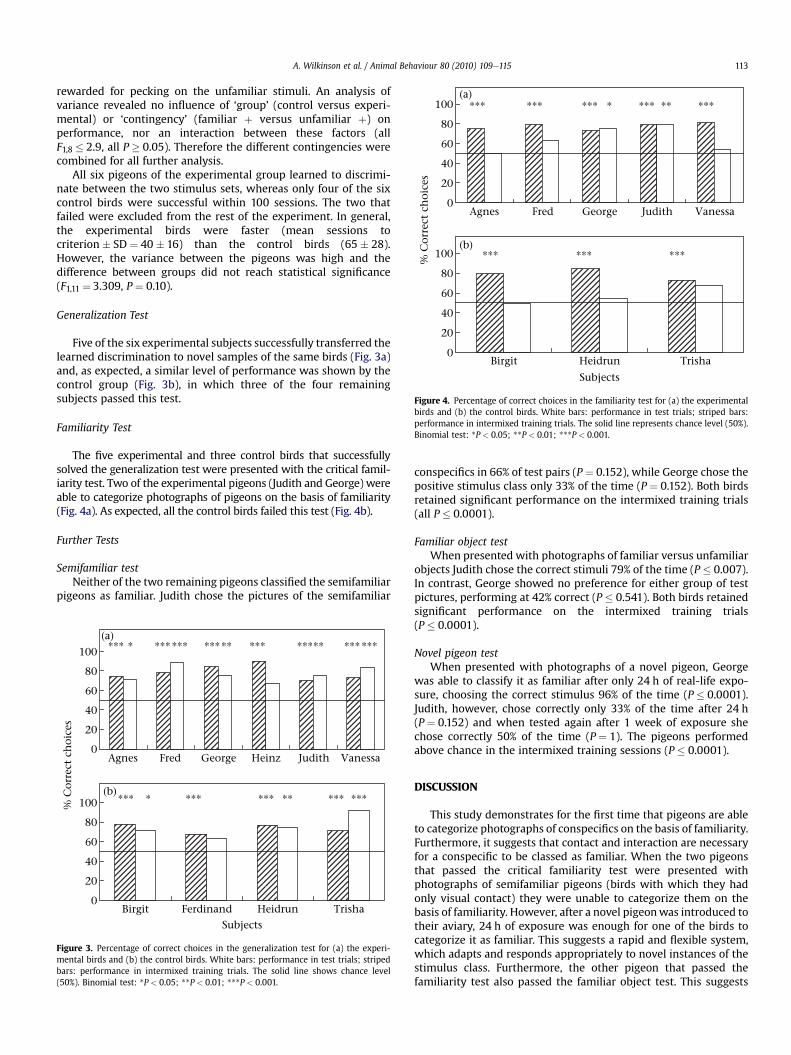
Figure 4. Percentage of correct choices in the familiarity test for (a) the experimentalbirds and (b) the control birds. White bars: performance in test trials; striped bars:performance in intermixed training trials. The solid line represents chance level (50%).Binomial test: *P < 0.05; **P < 0.01; ***P < 0.001.
A. Wilkinson et al. / Animal Behaviour 80 (2010) 109e115 113
rewarded for pecking on the unfamiliar stimuli. An analysis ofvariance revealed no influence of ‘group’ (control versus experi-mental) or ‘contingency’ (familiar þ versus unfamiliar þ) onperformance, nor an interaction between these factors (allF1,8 � 2.9, all P � 0.05). Therefore the different contingencies werecombined for all further analysis.
All six pigeons of the experimental group learned to discrimi-nate between the two stimulus sets, whereas only four of the sixcontrol birds were successful within 100 sessions. The two thatfailed were excluded from the rest of the experiment. In general,the experimental birds were faster (mean sessions tocriterion � SD ¼ 40 � 16) than the control birds (65 � 28).However, the variance between the pigeons was high and thedifference between groups did not reach statistical significance(F1,11 ¼ 3.309, P ¼ 0.10).
Generalization Test
Five of the six experimental subjects successfully transferred thelearned discrimination to novel samples of the same birds (Fig. 3a)and, as expected, a similar level of performance was shown by thecontrol group (Fig. 3b), in which three of the four remainingsubjects passed this test.
Familiarity Test
The five experimental and three control birds that successfullysolved the generalization test were presented with the critical famil-iarity test. Two of the experimental pigeons (Judith and George) wereable to categorize photographs of pigeons on the basis of familiarity(Fig. 4a). As expected, all the control birds failed this test (Fig. 4b).
Further Tests
Semifamiliar testNeither of the two remaining pigeons classified the semifamiliar
pigeons as familiar. Judith chose the pictures of the semifamiliar
0
20
40
60
80
100
Agnes Fred George Heinz Judith Vanessa
∗∗∗ ∗∗ ∗∗∗∗ ∗∗∗ ∗∗∗∗∗∗ ∗∗∗ ∗∗∗ ∗∗ ∗∗∗
0
20
40
60
80
100
Birgit Ferdinand Heidrun Trisha
Subjects
% C
orre
ct c
hoi
ces
∗∗∗ ∗∗∗ ∗∗∗ ∗∗∗ ∗∗∗∗ ∗∗
(a)
(b)
Figure 3. Percentage of correct choices in the generalization test for (a) the experi-mental birds and (b) the control birds. White bars: performance in test trials; stripedbars: performance in intermixed training trials. The solid line shows chance level(50%). Binomial test: *P < 0.05; **P < 0.01; ***P < 0.001.
conspecifics in 66% of test pairs (P ¼ 0.152), while George chose thepositive stimulus class only 33% of the time (P ¼ 0.152). Both birdsretained significant performance on the intermixed training trials(all P � 0.0001).
Familiar object testWhen presented with photographs of familiar versus unfamiliar
objects Judith chose the correct stimuli 79% of the time (P � 0.007).In contrast, George showed no preference for either group of testpictures, performing at 42% correct (P � 0.541). Both birds retainedsignificant performance on the intermixed training trials(P � 0.0001).
Novel pigeon testWhen presented with photographs of a novel pigeon, George
was able to classify it as familiar after only 24 h of real-life expo-sure, choosing the correct stimulus 96% of the time (P � 0.0001).Judith, however, chose correctly only 33% of the time after 24 h(P ¼ 0.152) and when tested again after 1 week of exposure shechose correctly 50% of the time (P ¼ 1). The pigeons performedabove chance in the intermixed training sessions (P � 0.0001).
DISCUSSION
This study demonstrates for the first time that pigeons are ableto categorize photographs of conspecifics on the basis of familiarity.Furthermore, it suggests that contact and interaction are necessaryfor a conspecific to be classed as familiar. When the two pigeonsthat passed the critical familiarity test were presented withphotographs of semifamiliar pigeons (birds with which they hadonly visual contact) they were unable to categorize them on thebasis of familiarity. However, after a novel pigeonwas introduced totheir aviary, 24 h of exposure was enough for one of the birds tocategorize it as familiar. This suggests a rapid and flexible system,which adapts and responds appropriately to novel instances of thestimulus class. Furthermore, the other pigeon that passed thefamiliarity test also passed the familiar object test. This suggests
A. Wilkinson et al. / Animal Behaviour 80 (2010) 109e115114
that she possessed a broad understanding of familiarity that wasnot restricted to conspecifics.
The use of a concept for categorization is considered higherorder learning as it goes beyond categorization on the basis ofperceptual similarities (Lea 1984). Two subjects in this studyresponded to novel instances of familiar and unfamiliar conspe-cifics in the same way as they did to training stimuli. As such thepigeons did not use perceptual cues tomake the discrimination, butbased their categorization on the rule of familiarity. This could beconsidered abstract categorization (Lazareva & Wasserman 2008)as the pigeon not only has to understand the relation between itselfand the stimuli, but also the relations between the stimuli. The useof the familiarity concept is further demonstrated by the flexibilityof one of the pigeon’s discriminative behaviour (transferring therule of familiarity from conspecifics to objects), and the failure ofthe control group on the critical familiarity test.
The stimuli were clearly difficult to discriminate: two of thecontrol pigeons failed to make the discrimination after 100sessions. However, the difference in performance between the twogroups was not based on differences in learning ability. Prior to thestart of the experiment both groups were pretrained on a set ofpictorial stimuli which they had to rote learn. Analysis comparingthe pretraining performance of the two groups revealed no differ-ence between them (t10 ¼ 1.28, P ¼ 0.23). As such it seems likelythat the differences in performance during the experiment weredue to the real-life experience of the experimental group. The twobirds that passed the familiarity test appeared to use their real-lifeexperience in different ways. The bird that passed the familiarobject test did not pass the novel pigeon test. It is possible that oneof the birds categorized the stimuli on the basis of highly familiarobjects within the aviary and the other on the basis of conspecificswith which it had interacted. This fact nicely demonstrates thepigeons’ flexibility in finding a solution to a complex learningproblem.
Only two of the six experimental animals passed the familiaritytest. They were both female and both middle ranking in thedominance hierarchy (with Judith being slightly higher thanGeorge). As there were other females in the experimental group itseems more likely that their rank, rather than their sex, influencedtheir concept formation ability. A middle-ranking bird would haveto adapt its behaviour depending on which pigeon it was inter-acting with; thus it is possible that the two birds that passed thetest monitored the other pigeons more carefully than those thatwere either top or bottom of the dominance hierarchy.
It could be argued that the pigeons’ impressive discriminativebehaviour was the result of simple priming effects; however, this isunlikely for a number of reasons. Half of the birds were rewardedfor choosing the familiar pigeon and the other half for choosing theunfamiliar one. No differences in learning were observed betweenthese two groups (of the two birds that passed the familiarity testone was rewarded for choosing the familiar conspecific and theother was rewarded for choosing the unfamiliar one). Furthermore,all birds produced an ordinary acquisition curve, indicating thatthey had to learn the discrimination and did not rely on automaticperceptual cues for classification. Thus, our results suggest thatpigeons possess the ability to classify items on the basis offamiliarity.
Studies examining the effect of brain lesions on the visualcategorization abilities of pigeons have found a dissociationbetween biologically relevant stimuli (food and conspecifics) andirrelevant stimuli (artificial patterns and different species;Watanabe 1996). It is possible that the biological relevance orfunctional importance of the familiar individuals and objects mayhave controlled our pigeons’ discriminative behaviour. The subjectsof the experimental group interacted with the pictured conspecifics
in a variety of real-life social contexts (mating, competing forterritory, etc.), and as a result these animals were functionallyimportant to them. Moreover, the familiar objects used in theexperiment (food trough and water dispenser) were also biologi-cally important to the pigeons. If the birds used such a method fordiscrimination it is not surprising that they behaved differently onthe tests as it is likely that different things were functionallyimportant to each bird. It is possible that the novel pigeon was ofgreat biological relevance to George, which resulted in a highlysignificant performance on this test, but not to Judith. In contrast,the semifamiliar conspecifics had no biological importance toeither of the subjects; they could not mate with them, and did notneed to defend territory from them, etc. Therefore it is notsurprising that they could not make the discrimination. The aviaryof the semifamiliar pigeons was located 148 cm from that of theexperimental birds (Fig. 1). As such it is highly implausible that thefailure of discrimination was due to the birds being unable to seeeach other. Indirect support for this comes from the fact that thebirds of the experimental group showed a marked change inbehaviour when the birds of the semifamiliar group were fed.
The results of the familiarity test in addition to the novel pigeontest provide evidence for visually based individual recognition inthe pigeon. Recognition of a familiar animal is extremely importantas it allows differential and appropriate responding to a flockmateversus a stranger. On its own this is considered a relatively complextask (Tibbetts & Dale 2007). However, our pigeons not only had torecognize conspecifics in real life, but also had to transfer thatknowledge to photographic stimuli in a touch-screen situation. Tosee that a picture represents a real-life object requires an animal toabstract the relation between the picture and the item that itrepresents. In addition, two-dimensional photographic stimulirepresent a much degraded version of the real-life pigeon. Theyonly provide static visual information; furthermore, that informa-tion is created for the human visual system which differs enor-mously from that of the pigeon (e.g. Varela et al. 1993). As such thestimuli may vary a great deal fromwhat pigeons actually see in reallife. There are contradictory reports of the pigeon’s ability torecognize objects depicted in photographs (Delius et al. 2000). Thefindings of this experiment corroborate earlier reports that pigeonssee a correspondence between a picture on the screen and the real-life object that it represents (Aust & Huber 2006).
In summary, this study shows that pigeons are able todiscriminate photographs of familiar and unfamiliar conspecifics,and can generalize this behaviour to novel views of the trainingstimuli. Critically, two birds correctly classified pictures of entirelynovel stimulus birds on the basis of familiarity. These birds werenever shown during training, but they were part of the experi-mental birds’ social group; the control group failed this test. Thus,the results provide evidence for three important aspects of visualcognition. They suggest that pigeons (1) recognize some corre-spondence between computer stimuli and real items, (2) possessthe ability to form a concept of familiarity, and (3) are able todistinguish between social partners and strangers in nature.
Acknowledgments
We thank the pigeon group of the University of Vienna for theirinput into the paper. This work was supported by funding from theAustrian Science Fund (FWF; to L.H.) under contract numberP19574.
References
Aust, U. & Huber, L. 2006. Pictureeobject recognition in pigeons: evidence ofrepresentational insight in a visual categorization task using a complementary
A. Wilkinson et al. / Animal Behaviour 80 (2010) 109e115 115
information procedure. Journal of Experimental Psychology: Animal BehaviourProcesses, 32, 190e195.
Boysen, S. T. & Berntson, G. G. 1989. Conspecific recognition in the chimpanzee(Pan troglodytes): cardiac responses to significant others. Journal of ComparativePsychology, 103, 215e220.
Bradshaw, R. H.1992. Conspecific discrimination and social preference in the layinghen. Applied Animal Behavior Science, 33, 69e75.
Bradshaw, R.H. &Dawkins,M. S.1993. Slides of conspecifics as representatives of realanimals in laying hens (Gallus domesticus). Behavioural Processes, 28, 165e172.
Dasser, V. 1987. Slides of group members as representations of the real animals(Macaca fascicularis). Ethology, 76, 65e73.
Dawkins, M. S. 1996. Distance and social recognition in hens: implications for theuse of photographs as social stimuli. Behaviour, 33, 663e680.
Dawkins, M. S., Guilford, T., Braithwaite, V. A. & Krebs, J. R. 1996. Discriminationand recognition of photographs of places by homing pigeons. BehaviouralProcesses, 36, 27e38.
D’Eath, R. B. & Dawkins, M. S. 1996. Laying hens do not discriminate betweenvideo images of conspecifics. Animal Behaviour, 52, 903e912.
Delius, J. D., Emmerton, J., Hörster, W., Jäger, R. & Ostheim, J. 2000. Pictureeobjectrecognition in pigeons. In: Picture Perception in Animals (Ed. by J. Fagot), pp.1e36.Hove: Psychology Press.
DeLoache, J. S. 2000. Dual representation and young children’s use of scale models.Child Development, 71, 329e338.
Herrnstein, R. J. & Loveland, D. H. 1964. Complex visual concept in the pigeon.Science, 146, 549e551.
Huber, L. In press. Categories and concepts: language-related competences in non-linguistic species. In: Encyclopedia of Animal Behavior (Ed. by J. Moore & M.Breed). Oxford: Elsevier.
Kendrick, D. F. 1992. Pigeon’s concept of experienced and nonexperienced real-world locations: discrimination and generalization across seasonal variation.
In: Cognitive Aspects of Stimulus Control (Ed. by W. K. Honig & J. G. Fetterman),pp. 113e134. Hillsdale, New Jersey: L. Erlbaum.
Kendrick, K. M., Atkins, K., Hinton, M. R., Heavens, P. & Keverne, B. 1996. Arefaces special for sheep? Evidence from facial and object discrimination learningtests showing effects of inversion and social familiarity. Behavioural Processes,38, 19e35.
Lazareva, O. F. & Wasserman, E. A. 2008. Categories and concepts in animals. In:Learning Theory and Behavior. Vol. 1 of Learning and Memory: A ComprehensiveReference (Ed. by R. Menzel & J. H. Byrne), pp. 197e226. Oxford: Elsevier.
Lea, S. E. G. 1984. In what sense do pigeons learn concepts? In: Animal Cognition(Ed. by H. L. Roitblat, T. G. Bever & H. S. Terrace), pp. 263e276. Hillsdale, NewJersey: L. Erlbaum.
Nakamura, T., Croft, D. B. & Westbrook, R. F. 2003. Domestic pigeons (Columbalivia) discriminate between photographs of individual pigeons. Learning &Behavior, 31, 307e317.
Parr, L. A. & de Waal, F. D. M. 1999. Visual kin recognition in chimpanzees. Nature,399, 647e648.
Pokorny, J. J. & de Waal, F. D. M. 2009. Monkeys recognize the faces of group matesin photographs. Proceedings of the National Academy of Sciences, U.S.A., 106,21539e21543.
Tibbetts, E. A. & Dale, J. 2007. Individual recognition: it is good to be different.Trends in Ecology and Evolution, 22, 529e537.
Varela, F. J., Palacios, A. G. & Goldsmith, T. H. 1993. Color vision of birds. In: Vision,Brain and Behavior in Birds (Ed. by H. P. Zeigler & H-J Bischof), pp. 77e98.Cambridge: Massachusetts: MIT Press.
Vriends, M. M. 1988. Pigeons. Hauppauge, New York: Barrons.Watanabe, S. 1996. Effects of ectostriatal lesions on discriminations of conspecific,
species and familiar objects in pigeons. Behavioural Brain Research, 81, 183e188.Wilkie, D. M., Willson, R. J. & Kardal, S. 1989. Pigeons discriminate pictures of
a geographic location. Animal Learning & Behavior, 17, 163e171.