37 Plant Defense Systems T o protect crops from diseases and her- bivores, the United Nations Food and Agricultural Organization estimates that the world’s farmers apply over 1.5 million metric tons of the active ingredients found in herbicides, fungicides, and insecticides to their fields each year. When these agents fail, the re- sults can be catastrophic. In 1979 and 1980, a bacterial disease wiped out 60 percent of the rice crop in numerous regions of India; in 1980, a fungus killed the entire wheat crop in many parts of Kazakhstan. Diseases and herbivores constantly threat- en crop plants with annihilation. Yet the world is green. This trivial observation is actu- ally interesting. Most plant tissues are not eaten by herbivores or destroyed by bacteria, fungi, or parasitic nematodes (roundworms). Why? Plants cannot run away from these nemeses; instead, they must stand and fight. Wild plant species do this fighting on their own—they get no assistance from the chemi- cal agents applied by humans. Understanding how plants defend against disease and herbivory is among the most active research areas in all of plant biology. The ex- periments we’ll review are motivated not only by the excitement of understanding the funda- mental biological questions involved, but also by concern about the future of agriculture. Many crops lack desirable disease-fighting traits because such traits have been bred out in an effort to boost productivity. In many cases, massive applications of pesticides are required to make up for the loss of these natural de- fense systems. How do the molecules involved in plant de- fense work? Could alleles that are important for defense be safely introduced into crop vari- eties that are particularly productive and nutri- tious? To begin answering these questions, let’s 709 37.1 Barriers to Entry 37.2 Plant Poisons 37.3 The Cost of Defense 37.4 Responding to Pathogens 37.5 Responding to Herbivores Diseases and insects cause billions of dollars of crop damage each year. What keeps par- asites and herbivores from eating everything?
Transcript
37Plant Defense Systems
To protect crops from diseases and her-bivores, the United Nations Food andAgricultural Organization estimates
that the world’s farmers apply over 1.5 millionmetric tons of the active ingredients found inherbicides, fungicides, and insecticides to theirfields each year. When these agents fail, the re-sults can be catastrophic. In 1979 and 1980, abacterial disease wiped out 60 percent of therice crop in numerous regions of India; in1980, a fungus killed the entire wheat crop inmany parts of Kazakhstan.
Diseases and herbivores constantly threat-en crop plants with annihilation. Yet theworld is green. This trivial observation is actu-ally interesting. Most plant tissues are noteaten by herbivores or destroyed by bacteria,fungi, or parasitic nematodes (roundworms).Why? Plants cannot run away from thesenemeses; instead, they must stand and fight.Wild plant species do this fighting on theirown—they get no assistance from the chemi-cal agents applied by humans.
Understanding how plants defend againstdisease and herbivory is among the most activeresearch areas in all of plant biology. The ex-periments we’ll review are motivated not onlyby the excitement of understanding the funda-mental biological questions involved, but alsoby concern about the future of agriculture.Many crops lack desirable disease-fightingtraits because such traits have been bred out inan effort to boost productivity. In many cases,massive applications of pesticides are requiredto make up for the loss of these natural de-fense systems.
How do the molecules involved in plant de-fense work? Could alleles that are importantfor defense be safely introduced into crop vari-eties that are particularly productive and nutri-tious? To begin answering these questions, let’s
709
37.1 Barriers to Entry
37.2 Plant Poisons
37.3 The Cost of Defense
37.4 Responding to Pathogens
37.5 Responding to Herbivores
Diseases and insects cause billions of dollars of crop damage each year. What keeps par-asites and herbivores from eating everything?
FREEMC37.13081923.709-725.final 11/12/01 10:45 AM Page 709
710 Unit 7 How Plants Work
take a look at defense systems found on the exteriors of plants,and then look at the defenses found inside plants.
37.1 Barriers to EntryThe epidermis of a plant is analogous to the skin of an animal.Because it provides a barrier to entry by disease-causing organ-isms, or pathogens, it serves as an individual’s first line of de-fense. Bacteria, viruses, and fungi must break through the epi-dermal barrier before they can infect cells, reproduce, andcause disease. Thorns or other structures on the epidermis of aplant are important for thwarting animals that eat plant tissues,or herbivores (plant-eaters).
Here we consider two questions about the role of the epider-mis in preventing disease and damage. How does the plant epi-dermis resist entry by pathogens and herbivores? How dopathogens and herbivores overcome these defenses?
Cuticle and CutinaseIn stems and leaves, the outside surface of epidermal cells is cov-ered with a substance called cuticle (Figure 37.1). Cuticle is madeup of a matrix of cross-linked lipid molecules impregnated withthe extremely long-chained lipids called waxes. Both elementsare hydrophobic and function in limiting water loss from the aer-ial parts of the plant. The waxy sheet of cuticle also functions indefense, forming a physical barrier that resists penetration byvirus particles, bacterial cells, and the spores or growing fila-ments of fungi. The epidermal cells of roots lack a waxy coating,but have a tough lipid matrix similar to that found in cuticle.
Pathogens have several ways of circumventing this barrier.Wounds from herbivores and mechanical damage expose cellsto airborne fungal spores, virus particles, and bacterial cells.Bacteria and fungi can also enter stomata when they are open;viruses can be injected directly into plant tissues by aphids,
leafhoppers, and other insects that pierce the plant epidermiswith their mouthparts to suck phloem sap.
Work in Pappachan Kolattukudy’s laboratory establishedthat certain fungi have an equally direct way of entering planttissues: They use enzymes to disrupt the cuticle. Specifically,many pathogenic fungi produce an enzyme called cutinase.Cutinase cleaves the lipid cutin, which forms the cross-linkedmatrix in cuticle. In this way, fungi hack out a hole in cuticle toexpose the cells within.
To appreciate the importance of this enzyme, consider workcarried out by Linda Rogers and her colleagues. They were ableto isolate individuals of the fungus Fusarium solani that con-tain many copies of the cutinase gene, just one copy, or no func-tional copies. When Rogers and co-workers infected pea plantswith spores from each strain, they obtained the data in Figure37.2. The graphs demonstrate a strong correlation between thenumber of copies of the cutinase gene found in a particular fun-gal strain and its virulence, or its ability to cause disease. In thiscase, the ability to break through a plant’s first line of defensedetermines the difference between virulent and benign strainsof a pathogen. Strains with more copies of the gene for cutinaseare more virulent.
WeaponsThe spines, thorns, and prickles produced by some plants canimpede insect and mammal herbivores, just as the cuticlelayer helps to thwart viruses, bacteria, and other microscopicpathogens. In some cases, these structures have other func-tions as well. As Chapter 32 pointed out, the hair-like struc-
Cross-section of epidermis
Cuticle
Cuticle keeps pathogens out
FIGURE 37.1 Cuticle Coats Epidermal CellsCuticle forms a barrier that keeps water in and pathogens out.
Days
1 2 3 4 5 6 7
100
50
75
25
0Per
cent
age
of
pea
s in
fect
ed b
y fu
ngus Fungus with...
many copiesof cutinase gene
one copyof cutinase gene
no copiesof cutinase gene
FIGURE 37.2 Evidence That Cutinase Genes Increase the Virulenceof FungiCutinase is an enzyme that digests plant cuticle. These data indicatea strong correlation between the number of cutinase genes in Fusari-um strains and their ability to infect peas and cause disease.EXERCISE These data show what happened when the researchersapplied a high dosage of spores. They repeated the experiment withmedium and low doses of spores from each fungal strain. Add linesto the graph predicting the results from medium and low doses.
FREEMC37.13081923.709-725.final 11/12/01 10:45 AM Page 710
Chapter 37 Plant Defense Systems 711
tures called trichomes help desert-dwelling plants limit waterloss from stomata.
As the photographs in Figure 37.3 show, hairs and pricklesare weapons analogous to the teeth, pincers, or stingers that an-imals use to defend themselves from predators. In one case, anarmy of mercenaries augments these plant weapons. Some Aca-cia trees native to East Africa have large, bulbous spines thathouse ant colonies (Figure 37.4a). When a browsing mammalbegins to eat leaves or twigs from the tree, the shaking motionstimulates the ants to attack the animal and deliver painfulbites. The relationship between the acacias and ants appears tobe mutually beneficial, with the trees providing a safe nestingsite and the ants providing defense.
The ferocity of the ants made P. G. Willmer and G. N.Stone wonder how acacias are able to attract pollinators totheir flowers. How could a bee spend enough time on an aca-cia flower to pick up and deliver pollen if it is quickly at-tacked by biting ants? To answer this question, the researchersproposed that acacia flowers might produce some sort ofchemical deterrent that affects the ants but not bees. Afterdocumenting that ants spend much more time at old flowersthan at newly opened flowers on the same tree, Willmer andStone were able to formulate a more specific hypothesis. Theyproposed that a component of newly opened flowers, such aspollen, contains a chemical that deters ants but not bees. Totest this idea, they wiped old flowers with newly opened flow-ers and documented how long ants stayed during each visit tothe treated flowers.
159 µm
FIGURE 37.3 Structures ThatProtect PlantsLeft: A close-up of a sepal that pro-tects a rose bud. The large bulbousstructures projecting from the sur-face contain chemicals that deter insects and other small herbivoresfrom attacking the young flower.Dozens of simple hairs are visible aswell. Right: In adult roses, stems areprotected from large herbivores byprickles. QUESTION Why are theleaves of some plants “fuzzy?”
A substance in young flowers deter ants
Ave
rag
e d
urat
ion
of
ant
visi
ts (s
eco
nds)
Youngflowers
Oldflowers
Old flowerswiped with
young flowers
Young flowerswiped withold flowers
(a) Acacia trees are protected from herbivores by biting ants, which live in large bulbs at the base of the thorns.
(b) Does a substance in young flowers keep the ants away from pollinators?
0
10
20
30
40
50
60
70
FIGURE 37.4 Ant-Guarded Acacias(a) In certain species of Acacia, ants in the genus Crematogaster livein large bulbs at the base of spines. (b) These data show the averageamount of time that individual ants spend on acacia flowers of vari-ous ages and experimental treatments. QUESTION What wouldthe data in part (b) look like if young flowers did not produce an ant-deterrent compound? (Your answer is the null hypothesis in thisexperiment.)
The results of their experiment are shown in Figure 37.4b.As predicted, old flowers that had been wiped with newlyopened flowers were just as unattractive to ants as newlyopened flowers. Ant-guarded acacia flowers appear to pro-duce a substance that keeps ants away during the intervalwhen pollination occurs. The study suggests that plants areable to make powerful deterrent compounds, and that in atleast some cases these insecticides are species specific.
FREEMC37.13081923.709-725.final 11/12/01 10:45 AM Page 711
712 Unit 7 How Plants Work
37.2 Plant PoisonsMany plants avoid being eaten by lacing their tissues withpoisons. Some of these chemicals are familiar. The flavorfuloils in peppermint, lemon, basil, and sage have insect repel-lent properties. The pitch that oozes from pines and firs con-tains a molecule called pinene, which is toxic to bark beetles.The pyrethroids produced by Chrysanthemum plants are acommon ingredient in commercial insecticides. Moleculescalled tannins accumulate in the cell vacuoles of many differ-ent species. Because they bind to proteins, tannins inactivatedigestive enzymes in the herbivores that ingest them. As a re-sult, the herbivores get sick. In humans, small doses of tan-nins cause a sharp, astringent sensation in the mouth that isprized in apples, blackberries, teas, and red wine. (Tanninsalso cause structural changes that make proteins less likely todisintegrate; tannins are used to tan animal skins and pro-duce leather.)
These plant poisons are similar to cuticle and thorns in animportant way: They are always present in the plant. Stated an-other way, they are produced constitutively—which means theydo not need a specific stimulus to initiate their production. Thisis in sharp contrast to the defense systems we’ll explore later (insections 37.4 and 37.5), which are activated only after an infec-tion or attack has begun.
Plants that produce poisons constitutively are similar toskunks, stinkbugs, monarch butterflies, and other animals thatavoid being eaten by tasting bad or spraying their enemies withnoxious chemicals. (Plants that sequester poisons also mount arapid response to pathogens and herbivores using systems in-troduced later in the chapter.) Here we ask: Where do plantpoisons come from, and how do these molecules act?
The Role of Secondary MetabolitesIn almost every case, plant defense compounds have structuresand compositions that are closely related to the molecules re-quired for basic cell activities, such as amino acids. To explainthis resemblance, biologists hypothesize that during the evolu-tion of some plant groups, mutations occurred in the genes forenzymes involved in fundamental biosynthetic pathways. Thehypothesis claims that these mutations resulted in the produc-tion of altered enzymes and the synthesis of new or “sec-ondary” compounds. If these compounds happened to be toxicto herbivores or pathogens, they would help the mutant indi-vidual survive better and reproduce more. Over time, then, thealleles responsible for encoding the required enzymes would in-crease in frequency in the population. Based on this logic andtheir chemical similarity to important compounds, plant poi-sons are referred to as secondary metabolites.
To understand how secondary metabolites relate to aminoacids and other “primary” compounds, consider recent workby Monika Frey and colleagues. This research group was inter-ested in a molecule called DIMBOA (2,4-dihydroxy-7-
methoxy-1,4-benzoxazin-3-one) that is found in corn andwheat. DIMBOA is similar in structure to the amino acid tryp-tophan, but is extremely effective at deterring several of the an-imal pests that afflict these crops.
The steps involved in tryptophan synthesis are well known.Is the synthesis of DIMBOA related to these steps? If so, how?Experiments by other researchers had provided an intriguinghint. When corn plants are fed radioactively labeled moleculesthat are precursors in the pathway for tryptophan synthesis, thelabel shows up in DIMBOA as well as in tryptophan. But if thesame plants are fed radioactively labeled tryptophan, the labeldoes not show up in DIMBOA. These observations suggest thatthe synthesis of this secondary metabolite is an “offshoot” oftryptophan synthesis, as illustrated in Figure 37.5a.
To explore this idea, Frey and co-workers analyzed cornplants that have a mutation called Bx1. These individuals can-not synthesize DIMBOA. After an extensive search, the re-searchers were able to locate the Bx1 gene. Nearby on the chro-mosome, they found four related loci that they called Bx2, Bx3,Bx4, and Bx5. To determine whether the protein products ofthese five genes are involved in DIMBOA synthesis, they addedthe five BX enzymes one by one to cells that were growing inculture and analyzed the products that resulted. Each BX en-zyme catalyzed a different reaction, and led to the productionof a different intermediate compound in the pathway for DIM-BOA synthesis. The experimental results are consistent with thepathway illustrated in Figure 37.5b.
These experiments provide strong support for the hypothe-sis that secondary metabolites evolved as offshoots of basicsynthetic pathways. Now that we know where secondarymetabolites come from, let’s take a look at how they poisontheir targets.
How Do Caffeine, Nicotine, and OtherAlkaloids Act as Poisons?Morphine, cocaine, nicotine, and caffeine are plant secondarymetabolites. They are part of a family of chemicals called alka-loids. Members of the alkaloid family are found in about 20percent of all plant species, and over 12,000 different moleculeshave been identified in various plant species to date. Althoughsome researchers propose that alkaloids are merely waste prod-ucts of normal biosynthetic activities, most biologists hypothe-size that they are actively synthesized as defense compounds.
Michael Wink and associates set up a large-scale study todetermine how alkaloids work. They assessed the action of 70different alkaloids and determined whether each of the mole-cules has a detrimental effect on DNA structure, DNA synthe-sis, protein synthesis, membrane permeability, bacterialgrowth, the survival of insects and worms, or the receptors lo-cated in the nerve cells and brains of mammals. The re-searchers’ goals were to test whether alkaloids act as poisons—and if so, to determine how alkaloids affect herbivores andpathogenic bacteria, viruses, and fungi.
FREEMC37.13081923.709-725.final 11/12/01 10:45 AM Page 712
Chapter 37 Plant Defense Systems 713
The study results showed that most of the alkaloids tested ex-hibited strong toxic effects, and that most alkaloids affectedmore than one aspect of cell biology. An alkaloid that poisons amembrane transport protein might also disrupt DNA structure.Quinine, for example, poisons the enzyme called reverse tran-scriptase that is found in pathogenic plant viruses; but it also hasa strong inhibitory effect on protein synthesis. In interpretingthese data, the researchers suggest that natural selection favorsthe synthesis of alkaloids that poison several target enzymes orstructures in the enemy at the same time. The key idea here isthat pathogens and herbivores are unlikely to evolve resistanceto poisons that affect a large number of important enzymes atthe same time. Recall that in Chapter 21, we explored the evo-lution of resistance in detail. Here, it becomes clear that plantsecondary compounds are extremely potent defensive weapons.
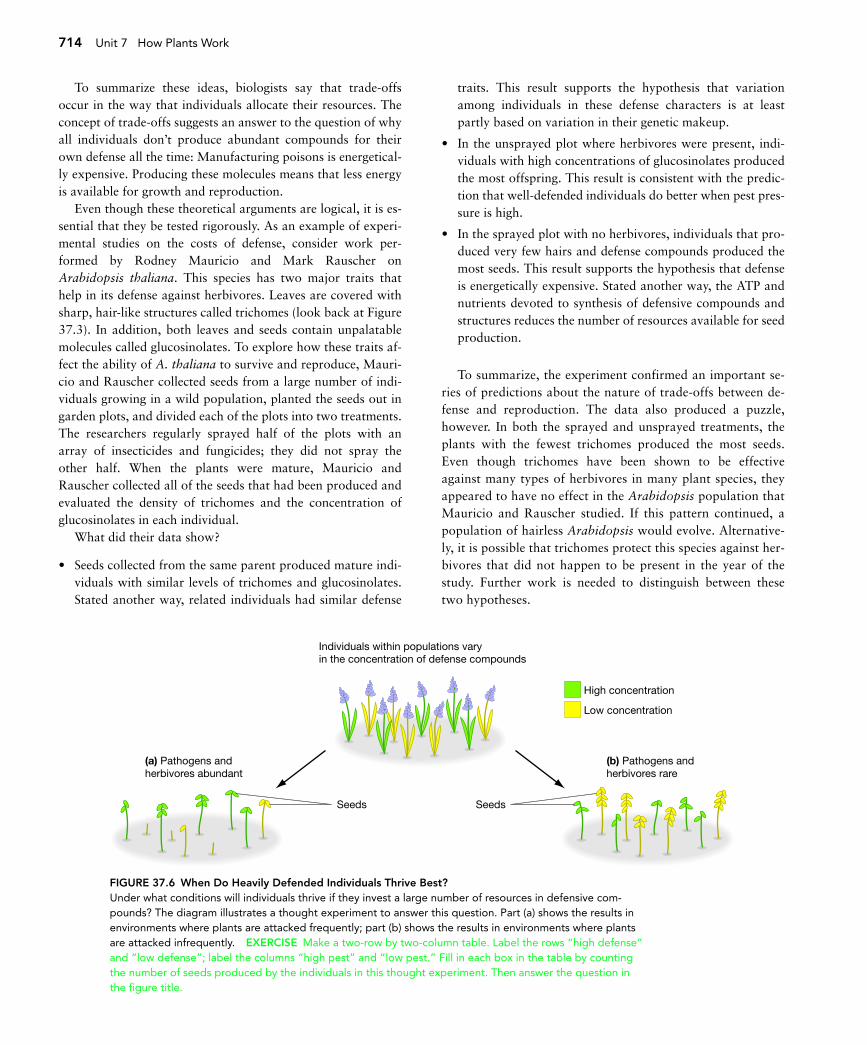
37.3 The Cost of DefenseIf secondary compounds are so effective, why don’t all plantsproduce a lot of them all the time? To address this question, re-searchers rely on a fundamental observation: Every organism hasa finite amount of energy. To understand why this simple fact isimportant, consider a thought experiment on the weedy mustardplant Arabidopsis thaliana. To begin, suppose that every individ-ual in a population manufactures an average of 100 million ATPmolecules in the course of its two-month lifetime. Further, sup-pose that the individuals in the population vary in how they ex-pend this ATP, and that this variation is based on genetic varia-tion among individuals. For example, some individuals mightspend a large percentage of their energy from ATP making defen-sive compounds and a relatively small percentage making seeds.Others might make few secondary metabolites and invest mostof their energy from ATP in reproduction instead. The questionis, which type of individual will survive and reproduce best?
The short answer to this question is “it depends.” Ifpathogens and herbivores are abundant, then most of the poor-ly defended plants will die. In this case, well-defended plantsproduce more offspring than lightly defended plants (Figure37.6a, page 714). As a result, alleles that lead to increased pro-duction of secondary metabolites will increase in frequency inthe population. But if pathogens and herbivores are rare, thenpoorly defended plants will produce the most offspring (Figure37.6b). In this case, alleles that lead to high production of de-fense compounds will decrease in frequency.
(a) Hypothesis: Secondary metabolites evolved as offshoots of basic synthetic pathways.
(b) Experimental support
Precursor compound
Precursor compound
Arrows indicatesteps in biosyntheticpathway, each catalyzed by a different enzyme
The enzymes thattransform the tryptophan precursorto DIMBOA havebeen discovered
Enzyme A
TSABX1
TSB BX2
BX3
BX4
BX5
B
C
D
E
F
TryptophanAmino acid,a primary compound
DIMBOAPest deterrent,a secondary metabolite
Tryptophan
DIMBOA
N
NNH2
N
N
N
N
OH
O
O
OH
O
O
N
OH
OH
CH3O
CO2H
O
O
N
OH
OH
H
H
H
HH
H
O
O
FIGURE 37.5 What Is “Secondary” About Secondary Metabolites?(a) A secondary metabolite is a molecule that is synthesized as an “offshoot” of a synthetic pathway for producing a fundamental, or primary, compound. Experimental data suggested that the defensecompound DIMBOA is manufactured as an offshoot of tryptophansynthesis. (b) In corn, each of the Bx gene products catalyzes a differ-ent reaction involved in the synthesis of DIMBOA. QUESTION Cornseedlings contain over 10 times as much DIMBOA as tryptophan. Inthis light, are the terms primary and secondary used appropriately?
FREEMC37.13081923.709-725.final 11/12/01 10:45 AM Page 713
714 Unit 7 How Plants Work
To summarize these ideas, biologists say that trade-offsoccur in the way that individuals allocate their resources. Theconcept of trade-offs suggests an answer to the question of whyall individuals don’t produce abundant compounds for theirown defense all the time: Manufacturing poisons is energetical-ly expensive. Producing these molecules means that less energyis available for growth and reproduction.
Even though these theoretical arguments are logical, it is es-sential that they be tested rigorously. As an example of experi-mental studies on the costs of defense, consider work per-formed by Rodney Mauricio and Mark Rauscher onArabidopsis thaliana. This species has two major traits thathelp in its defense against herbivores. Leaves are covered withsharp, hair-like structures called trichomes (look back at Figure37.3). In addition, both leaves and seeds contain unpalatablemolecules called glucosinolates. To explore how these traits af-fect the ability of A. thaliana to survive and reproduce, Mauri-cio and Rauscher collected seeds from a large number of indi-viduals growing in a wild population, planted the seeds out ingarden plots, and divided each of the plots into two treatments.The researchers regularly sprayed half of the plots with anarray of insecticides and fungicides; they did not spray theother half. When the plants were mature, Mauricio andRauscher collected all of the seeds that had been produced andevaluated the density of trichomes and the concentration ofglucosinolates in each individual.
What did their data show?
• Seeds collected from the same parent produced mature indi-viduals with similar levels of trichomes and glucosinolates.Stated another way, related individuals had similar defense
traits. This result supports the hypothesis that variationamong individuals in these defense characters is at leastpartly based on variation in their genetic makeup.
• In the unsprayed plot where herbivores were present, indi-viduals with high concentrations of glucosinolates producedthe most offspring. This result is consistent with the predic-tion that well-defended individuals do better when pest pres-sure is high.
• In the sprayed plot with no herbivores, individuals that pro-duced very few hairs and defense compounds produced themost seeds. This result supports the hypothesis that defenseis energetically expensive. Stated another way, the ATP andnutrients devoted to synthesis of defensive compounds andstructures reduces the number of resources available for seedproduction.
To summarize, the experiment confirmed an important se-ries of predictions about the nature of trade-offs between de-fense and reproduction. The data also produced a puzzle,however. In both the sprayed and unsprayed treatments, theplants with the fewest trichomes produced the most seeds.Even though trichomes have been shown to be effectiveagainst many types of herbivores in many plant species, theyappeared to have no effect in the Arabidopsis population thatMauricio and Rauscher studied. If this pattern continued, apopulation of hairless Arabidopsis would evolve. Alternative-ly, it is possible that trichomes protect this species against her-bivores that did not happen to be present in the year of thestudy. Further work is needed to distinguish between thesetwo hypotheses.
Individuals within populations varyin the concentration of defense compounds
SeedsSeeds
High concentration
Low concentration
(a) Pathogens and herbivores abundant
(b) Pathogens and herbivores rare
FIGURE 37.6 When Do Heavily Defended Individuals Thrive Best?Under what conditions will individuals thrive if they invest a large number of resources in defensive com-pounds? The diagram illustrates a thought experiment to answer this question. Part (a) shows the results inenvironments where plants are attacked frequently; part (b) shows the results in environments where plantsare attacked infrequently. EXERCISE Make a two-row by two-column table. Label the rows “high defense”and “low defense”; label the columns “high pest” and “low pest.” Fill in each box in the table by countingthe number of seeds produced by the individuals in this thought experiment. Then answer the question inthe figure title.
FREEMC37.13081923.709-725.final 11/12/01 10:45 AM Page 714
Chapter 37 Plant Defense Systems 715
37.4 Responding to PathogensManufacturing defensive compounds and structures requireslarge expenditures of ATP and limits the ability of individuals togrow and reproduce. In light of these findings, it is not surpris-ing that most plants have systems for responding to pathogensand herbivores only after an infection or attack has begun.
Response systems are currently the focus of intensive re-search because of their importance for agriculture. The produc-tion of constitutive poisons has been bred out of most cropplants, because toxins taste bad and lower productivity. Tofight off diseases and herbivores, then, some crop plants de-pend solely on their response systems and the application ofpesticides. Researchers hope that by gaining a detailed under-standing of how response systems work, they will be able tobreed or genetically engineer crops that defend themselves moreefficiently and are less dependent on pesticides.
Section 37.5 explores how plants respond to attacks by in-sect herbivores; here, we investigate how plants respond to in-fections by disease-causing viruses, bacteria, fungi, and nema-todes. The response systems triggered by these parasites areanalogous to the mammalian immune system introduced inChapter 46. The responses come in two waves: a rapid se-quence of events triggered at the point of infection, and a gen-eral or systemic reaction occurring throughout the body.
The Hypersensitive ResponseWhen a virus, bacterium, fungus, or nematode gets inside aplant and begins to grow, the infected cells respond by dying.The rapid and localized death of one or a few infected cells iscalled the hypersensitive response (HR). If the HR is successful,the pathogen is starved as host cells commit suicide.
In several respects, the hypersensitive response in plants issimilar to the cell-mediated immune response in mammals,which leads to the death of infected cells. The HR is also ex-tremely effective. Plants that mount a hypersensitive responserarely succumb to disease. How does this response get started,and how is it sustained?
What Triggers the Hypersensitive Response? The Gene-for-Gene Hypothesis In the early decades of the twentiethcentury, crop breeders established that plants have disease resis-tance genes that are inherited according to Mendel’s rules.These loci came to be known as resistance (R) genes; many areresponsible for triggering the HR. The same researchers also es-tablished that the fungi found to cause disease in wheat, flax,barley, and other crops have alleles that cause them to be viru-lent or avirulent (not virulent) on certain strains or varieties ofthese crops. The loci associated with virulence or avirulencecame to be known as avirulence (avr) genes.
In 1956, H. H. Flor published data demonstrating a one-to-one correspondence between the resistance alleles found in hostplants and the avirulence alleles found in pathogens. Stated an-
other way, Flor showed that certain R alleles and certain avr al-leles match. If an R allele in the host matches the avr allele in thepathogen, it means that their protein products must also matchin some way. In response to the matching event, an HR occurs.If the R allele in a host and the avr alleles in a pathogen do notmatch, then no HR occurs and the plant succumbs to disease.
The idea that R and avr gene products interact in a specificway came to be called the gene-for-gene hypothesis. What mo-lecular mechanism could be responsible for this pattern? Re-searchers following up on Flor’s work suggested that the HRbegins when proteins produced by host plants bind to proteinsor other molecules produced by the pathogen. The hypothesiswas that R genes produce receptors and avr genes produce lig-ands—meaning molecules that bind to receptors. Figure 37.7provides a general overview of this model.
The first breakthrough in confirming the gene-for-gene hy-pothesis occurred when researchers from a variety of laborato-ries around the world were able to clone and sequence a seriesof R genes from crop plants and avr loci from bacterial and
GENE-FOR-GENE HYPOTHESIS
Virus Bacterium Fungus
1. Pathogens (virus,bacterium, or fungus)enter plant cell viawound or connection with infected cell.
2. Proteins and othermolecules are released from pathogens.
3. R-gene products bind to certain molecules from pathogens (avr gene products).
4. Binding activates Rgene product and triggers protective HypersensitivityResponse (HR).
When R and avr gene products do not match, no HR occurs and plant succumbs to disease.
R
avr avr avr
R R
HR HR
FIGURE 37.7 The HR Begins when R Gene Products Bind to AVRGene ProductsThe gene-for-gene hypothesis predicts that there is a physical inter-action between specific R and avr gene products, and that this interaction initiates the hypersensitive response, which protectsplants from pathogens.
CD
A
CT
IV
IT
Y 3
7.1
Pla
nt D
efe
nse
s
FREEMC37.13081923.709-725.final 11/12/01 10:45 AM Page 715
716 Unit 7 How Plants Work
fungal pathogens. These results confirmed that R and avrgenes exist and code for products that could interact at thestart of an infection.
A second major advance, published in 1996, confirmed thegene-for-gene hypothesis by showing that the R and avr prod-ucts actually do bind to one another. Steven Scofield and co-workers demonstrated this interaction with an R gene in toma-to called Pto and an avr gene from Pseudomonas bacteriacalled avrPto. Figure 37.8 diagrams the researchers’ experimen-tal strategy. Note that the approach relies on a transcriptionfactor from yeast called GAL4, which was introduced inChapter 15. In yeast cells, GAL4 activates the transcription ofthe gene for the �-galactosidase enzyme. This enzyme, in turn,can react with a substrate molecule to produce a bright blueproduct. If the substrate is added to yeast cells in which GAL4is active, the cells turn bright blue.
To confirm that Pto and avrPto actually do bind to one an-other, the researchers fused the two genes to two different seg-ments of the GAL4 gene. As step 3 in Figure 37.8 shows, Ptowas fused to the DNA-binding domain of GAL4, while avrPtowas fused to the transcription activation segment of GAL4.GAL4 activates transcription only when the DNA-binding do-main and the transcription activation domains are physically at-tached to one another. When Scofield and co-workers intro-duced both of the gene constructs into yeast cells that lackGAL4, a complete GAL4 protein was produced, �-galactosidasewas synthesized, and the cells turned blue (step 6 in Figure37.8). This result confirmed that R and avr products physicallyinteract, just as predicted by the gene-for-gene hypothesis.
Resistance Loci A large number of R loci have now beenidentified in Arabidopsis, tomato, flax, tobacco, and otherplants. Two general patterns are emerging as data on thesegenes accumulate. First, R genes that are similar in sequenceand structure tend to be clustered together on the same chro-mosome. Second, within a population of plants, there are usu-ally many different alleles at each R locus. (Stated another way,R loci are highly polymorphic.)
These observations are important because they provide hintsabout the history and function of these genes. For example,clusters of similar loci, or what biologists call gene families, arethought to originate through errors in recombination. AsFigure 37.9 shows, gene duplication events occur when chro-mosomes misalign during crossing over. The result of this mu-tation is a chromosome with an extra copy of a gene. Becausethe organism with this chromosome already has a functioningcopy of the original gene, mutations occurring in the extra copyof the gene do not damage the individual. Instead, the new copymay acquire mutations that make a new function possible. Inthe case of R loci, the hypothesis is that mutations in the new
EVIDENCE SUPPORTING GENE-FOR-GENE HYPOTHESIS
Tomato host R(Pto)
Pathogen avr(avrpto)
?
1. Question: Do theR and avr proteinsinteract as the gene-for-gene hypothesispredicts?
2. Tool to answerquestion: The transcription factorGAL4 will only activate transcriptionwhen the DNA-binding domain and the transcription activation domains are physically attached.When GAL4 is functioning, it transcribes an enzymethat turns a substrateblue.
3. Construct recombinant genes using R and avr and the two parts of GAL4.
4. Insert recombinantgenes into yeast cellsto produce twoproteins: R + GAL4part, avr + GAL4 part.
5. Treat transformedcells with substrate totest for presence of functional GAL4. Cellswill only turn blue if the two parts of GAL4 are attached.
6. Interpretation:R and avr proteins must have bound to one another. When they did, they created a functional GAL4.
R GAL4 DNA-binding
avr GAL4 transcription activation
R protein
GAL4DNA-binding
GAL4transcription activation
β-galactosidase
β-galactosidase
Yeast cell
avr protein
Control cells Experimentalcells
Conclusion: The R and avr proteins DO interact as the gene-for-gene hypothesis predicts.
FIGURE 37.8 Evidence That R and AVR Gene Products Bind to OneAnother
FREEMC37.13081923.709-725.final 11/12/01 10:45 AM Page 716
Chapter 37 Plant Defense Systems 717
copies gave individuals the ability to recognize and respond tonovel avr products and thus new types of pathogens.
Why is it significant that many different alleles exist at eachR locus? The hypothesis here is that different alleles allowplants to recognize different proteins from the same pathogen.Plants are diploid, so they have two copies of each R gene. Ifmany different alleles exist in a population, then each individ-ual is likely to have two different alleles at each locus. The dif-ferent alleles allow the host to recognize different avr products.This is important because new avr products constantly arise inpathogen populations via mutation.
In combination, then, plants that have different alleles ateach of many R loci should be able to recognize and respond toa wide variety of disease-causing agents. The variability ob-served in R genes supports the hypothesis that plants rely ongene-for-gene interactions to recognize and thwart a diverse va-riety of invaders. Stated another way, if gene-for-gene interac-tions underlie some important resistance responses, and if anindividual plant has many different R genes, then it can recog-nize and respond to many different pathogen genes and thusmany different pathogens.
Before moving on to consider other aspects of the HR, it is in-teresting to note that mammals have two different sets of genesthat are responsible for recognizing pathogens and triggering a
response. As we’ll see in Chapter 46, both types of recognitionloci occur in gene families. These loci are found in clusters on thesame chromosome, and they are present in high copy number.The genes, which are called the immunoglobulin and MHC loci,are also highly polymorphic. Within any population, many dif-ferent alleles of each gene exist. The parallels between plant andmammalian disease-response systems are striking.
Reactive Oxygen Intermediates (ROI) The interaction be-tween recognition proteins in plants and avirulence moleculesfrom pathogens triggers a series of events. These responses in-clude the production of hydrogen peroxide (H2O2), superoxide(O2
�), and related molecules that are collectively called reactiveoxygen intermediates (ROI). These compounds trigger reac-tions that help reinforce cell walls. In addition, ROI are ex-tremely unstable molecules. As a result, they may also triggerreactions that are responsible for the death of infected cells orthat kill the pathogen directly.
What steps occur between the R-avr interaction and the endproduct of the HR—cell death? When Massimo Delladonne andco-workers treated plant cells with reactants that lead to theproduction of ROI, they found that some cell death occurred,but nothing like that induced by the actual R-avr interaction.This observation suggested that the R-avr interaction does notsimply lead to the production of compounds required to manu-facture ROI. In interpreting their results, the biologists suggest-ed the R-avr interaction must lead to the production of someother molecule that increases or augments the ROI response.
Delladonne and associates proposed that the missing mole-cule might be nitric oxide, or NO. (Nitric oxide is the active in-gredient in the anesthetic called laughing gas.) Their hypothesiswas inspired by the role that NO plays in the human immunesystem. The immune-system cells in our bodies frequently use alethal combination of NO and ROI to kill bacteria and diseasedhost cells.
To test whether the same killing mechanism occurs in plants,the researchers treated Arabidopsis plants with a drug that in-hibits the enzyme responsible for producing NO. Then theychallenged the plants by spraying the leaves with a bacterialpathogen. Figure 37.10a (page 718) documents the result.Plants that are able to produce normal amounts of NO showthe localized cell death typical of the HR. Plants with abnormalNO production are not able to stop the bacterial infection andhave large diseased areas in their leaves.
The experiment provides strong support for the hypothesisillustrated in Figure 37.10b. Both ROI and NO production ap-pear to be triggered by the R-avr interaction, and both types ofmolecules seem to be required for the HR. The involvement ofNO in the disease response furnishes another parallel betweendisease-fighting systems in plants and animals.
Phytoalexin Production A hallmark of the HR is the pro-duction of antibiotic compounds called phytoalexins at the site
Bars indicatelocation of R loci
Incorrect synapse of chromatids
One of cross-over productshas duplicated R1 locus
R1
R1
R1
R2
R2
R2
R2
R2
R2
R1
R1
R1
FIGURE 37.9 R Gene Families Probably Originated in GeneDuplication EventsWhen biologists find a series of closely related genes clustered together on the same chromosome, as in R loci, they infer that thecluster originated via a series of gene duplication events like the oneillustrated here.
FREEMC37.13081923.709-725.final 11/12/01 10:45 AM Page 717
of infection. A phytoalexin is defined as a small (low molecularweight) plant product that is induced by infection and thatpoisons the disease-causing agent. Because phytoalexins aredefined by their function and not their structure, it is not sur-prising that different plant species produce a diverse array ofthese molecules. The general idea here is that plant cells notonly commit suicide when infected by a pathogen; they pro-duce toxic molecules to poison the pathogen.
To get a better appreciation of how phytoalexins work, con-sider recent research by P. C. Stevenson and colleagues. Theseresearchers were interested in antifungal compounds producedby chickpeas in response to fungi that infect their roots. Chick-
peas (also known as garbanzo beans) are an important food inthe Middle East and south Asia. However, they are often vic-timized by a fungal disease called fusarium wilt. Earlier re-searchers had shown that chickpea roots contain antifungalcompounds called maackiain and medicarpin. What Stevensonand co-workers wanted to know is, are these antibiotics effec-tive against fusarium wilt? If so, are they phytoalexins thatchickpeas produce in response to infection?
To answer the first question, the researchers collected sporesfrom the Fusarium fungus and allowed them to germinate in thepresence of various concentrations of maackiain and medicarpin.As the graph in Figure 37.11 shows, both compounds are effec-tive antifungal agents—especially at high concentration.
Are these phytoalexins produced in response to infection?When the researchers monitored the concentrations of theseantibiotics in chickpea roots over time, they found that strongincreases occurred over the span of a week if they inoculatedthe soil with Fusarium spores. No such increase occurred if thesoil was kept free of fungi. This is strong evidence that maacki-ain and medicarpin are phytoalexins and are part of the re-sponse to infection in chickpeas.
Even more important, Stevenson and co-workers were ableto demonstrate a strong correlation between the ability ofchickpeas to ward off fusarium wilt and their ability to pro-duce phytoalexins. Figure 37.12 shows the results of the ex-periment. Note that the investigators worked with three vari-eties of chickpeas and two strains of Fusarium, and that eachof the three crop varieties shows a different susceptibility tothe Fusarium strains. Some chickpea populations are more re-sistant to certain fungal strains than others. When the experi-menters inoculated individuals from different chickpea vari-eties with different strains of the fungus and monitored theconcentrations of phytoalexins produced, they found that the
718 Unit 7 How Plants Work
(a) Plants must produce NO to stimulate HR
(b) HYPOTHESIS TO EXPLAIN RESULTS
Small lesionsproduced byHR
Large diseasedareas indicate that HR failed
Plant Rprotein
Celldeath
NOproduction
ROIproduction
1. R gene productbinds to pathogenmolecule.
2. Production of both NO and Reactive Oxygen Intermediates(ROI) is stimulated.
3. Both NO and ROIare required to initiatecell death.
Pathogenavr protein
Normal plant Plant treated with drug that inhibits
NO production
FIGURE 37.10 Evidence That Nitric Oxide Is Involved in theHypersensitive Response (HR)(a) If Arabidopsis are challenged with a pathogenic bacterium, the HRoccurs in normal plants and stops the infection. But if plants are treat-ed with a drug that inhibits nitric oxide (NO) production, the bacterialinfection spreads. (b) Researchers suggest that both NO and ROI arerequired for the cell suicide component of the hypersensitive response.
Concentration (µg ml–1)
0 50 100 150 200 250
Both compounds are effectiveanti-fungal agents, especiallyat high concentrations
Maackiain
Medicarpin
100
40
60
80
20
0
Ave
rag
e g
erm
inat
ion
of
fung
al s
po
res
(%)
FIGURE 37.11 Phytoalexins Are Effective PesticidesThis graph shows the percentage of fungal spores that germinatedwhen exposed to various concentrations of two different phytoalexins(a type of poison) produced by chickpeas.
FREEMC37.13081923.709-725.final 11/12/01 10:45 AM Page 718
ability to produce phytoalexins correlated strongly with theobserved level of resistance. Apparently, only certain varietiesof chickpeas are able to mount an effective response to certainstrains of Fusarium.
The data on phytoalexin production in chickpeas are excit-ing because they suggest a molecular mechanism for the geneticdifferences in disease resistance among chickpea strains. Timewill tell whether this knowledge will lead to the development ofnew crop varieties with increased disease resistance.
Systemic Acquired Resistance (SAR)The HR is fast and leads to localized cell death. Phytoalexinproduction also occurs at the point of infection. These respons-es are followed by a slower and more widespread set of eventscalled systemic acquired resistance (SAR). Over the course ofseveral days, SAR primes cells throughout the root or shootsystem for assault by a pathogen—even in cells that have notbeen directly exposed to the disease-causing agent.
How Do the SAR and HR Interact? Figure 37.13 illustrateshow the HR and SAR are thought to interact. A key point hereis that an interaction between R and avr products leads to theproduction of a signal that initiates SAR. This signal acts glob-ally as well as locally (at the point of infection), and results inthe expression of a large suite of genes called the PR (patho-genesis related) loci. The SAR signal qualifies as a hormone be-cause it carries information from one location to another. Whatis this signal, and what effects does it have?
What Molecule Triggers the SAR? When biologists set out tolocate the signaling molecule that triggers the SAR, their atten-tion quickly turned to salicylic acid (SA). Salicylic acid is foundin a wide variety of plants* and was found to increase dramati-cally in concentration after tissues are infected with a pathogen.The SA hypothesis became more convincing when researchers in
Chapter 37 Plant Defense Systems 719
Variety of chickpea tested
The ability of chickpeas to producephytoalexins correlates withthe observed level ofresistance
No exposure to fungus(control)
Susceptible toraces 1 and 2
Low
High
Resistant to race1 and susceptibleto race 2
Resistant toraces 1 and 2
Exposure toFusarium race 2
Exposure toFusarium race 1
600
300
400
500
200
100
0
Mea
n co
ncen
trat
ion
of
phy
toal
exin
(µg
g–1
)
FIGURE 37.12 Phytoalexins and Disease ResistanceTo produce the data in these histograms, researchers analyzed produc-tion of the phytoalexin medicarpin by three chickpea varieties. Onechickpea variety is susceptible to both strains of Fusarium tested, onevariety is resistant to one Fusarium strain but susceptible to the other,and one variety is resistant to both Fusarium strains. As the resultsshow, chickpeas that are resistant to certain fungal strains producephytoalexins only when they are infected with those specific strains.
Protection of plantfrom further infection
Other localizeddefense responses
NO
ROI
R avr
PR gene transcription
SAR signal
SAR signal
HR
Pathogen
FIGURE 37.13 The Hypersensitive Response Produces a SignalThat Induces Systemic Acquired ResistanceThis diagram summarizes the current consensus on how the HR andSAR interact.
*As an aside, it is interesting to note that SA is very closely related to the activeingredient in aspirin. SA is particularly abundant in willow and aspen trees.Long before medical scientists discovered the pain-relieving qualities of aspirin,Native American people used teas made with willow or aspen bark as a remedyfor pain.
FREEMC37.13081923.709-725.final 11/12/01 10:45 AM Page 719
several different laboratories showed that applying SA directlyto tissues triggers the SAR in a variety of plant species.
To confirm SA’s role, Thomas Gaffney and co-workers intro-duced a gene called salicylate hydroxylase into tobacco plants.This gene is not normally found in plants, and it codes for anenzyme that leads to the breakdown of SA. In tobacco plantsthat received the gene, both SA accumulation and the SAR wereabolished. As a result, the transformed plants were susceptibleto infection by a wide variety of pathogens.
It is still controversial, however, whether SA acts as theSAR hormone or whether it is only a local signal that triggersthe expression of genes involved in the SAR. Biologists fromseveral research groups have labeled SA with a radioactiveatom and confirmed that it is transported from infected to un-infected tissues. This observation is consistent with the hy-pothesis that SA actually is a signal that travels throughoutthe plant (Figure 37.14a). But when investigators in a differ-ent laboratory grafted the lower portions of tobacco plantsthat had been transformed with the salicylate hydroxylasegene onto normal shoots and challenged the roots with avirus, the leaves showed a normal SAR. This observation sug-gests that SA is not transported throughout the plant body(Figure 37.14b). As this book goes to press, the issue remainsunresolved. SA is clearly involved in triggering the SAR. It isstill not clear, however, whether SA is a local signal, a globalsignal, or both. Research on the mechanisms of the hypersen-sitive response and the systemic acquired response continues.It is clear, however, that these systems are only activated in re-sponse to a direct attack by bacteria or viruses and thus mini-
mize the cost of defense. Are analogous systems activated inresponse to attacks by herbivores?
37.5 Responding to HerbivoresOver a million species of insects have already been discoveredand named. Most of these species make their living by eatingvegetation, seeds, roots, or pollen. How do plants withstandthis onslaught of herbivores? In addition to fending off would-be predators with thorns and spines and sequestering sec-ondary metabolites that taste bad, plants respond to herbivoreattacks once they have begun. When a grasshopper bites a grassleaf, it sets off a series of carefully orchestrated events.
Here we explore two of the herbivore response systems thathave been researched in some detail. The first response resultsin the synthesis of insecticides at the point of attack and innearby leaves; the second leads to the production of com-pounds that attract enemies of the herbivores.
Proteinase InhibitorsIn the course of studies to determine why some foods are morepalatable and digestible than others, biochemists discoveredthat many seeds and some storage organs, such as potato tu-bers, contain proteins called proteinase inhibitors. Proteinaseinhibitors block the enzymes—found in the mouths and stom-achs of animals—that are responsible for digesting proteins.When an insect or mammalian herbivore ingests a large doseof a proteinase inhibitor, the herbivore gets sick. As a result,herbivores learn to detect proteinase inhibitors by taste, and
720 Unit 7 How Plants Work
(b) EVIDENCE THAT SA DOES NOT INITIATE SAR(a) EVIDENCE THAT SA ACTS AS A HORMONE
Normal shoot
No SA
Grafted root stock that producesenzyme thatdestroys SA
1. Inject leaf withradioactively labeledprecursor to SA.
2. Infect leaf withvirus to trigger SAR.
3. Later, find labeled SA in uninfected leaves.
2. Leaves shownormal SAR even though SA could not have originatedin root to sendmessage.
1. Infect roots withvirus to trigger SAR.
SA*
SA*SA*
SAR
SAR
SAR
SAR
SA*SA*
FIGURE 37.14 Is Salicylic Acid the Hormone That Triggers Systemic Acquired Resistance?(a) Radioactively labeled SA is transported from the site of an infection throughout the plant. This observa-tion suggests SA is the hormone that initiates SAR. (b) When roots that do not produce SA are grafted ontonormal shoot systems and then infected with a virus, the shoots respond with SAR. This observation suggestsSA is not the hormone that initiates SAR.
FREEMC37.13081923.709-725.final 11/12/01 10:45 AM Page 720
avoid plant tissues containing high concentrations of thesemolecules.
Researchers in Clarence Ryan’s laboratory documented thatproteinase inhibitors also occur in the leaves of tomatoes andpotatoes. During preliminary studies on these compounds, theinvestigators noticed that the concentration of compounds var-ied dramatically from plant to plant—sometimes by a factor of10. To explain the variability, the researchers hypothesized thatindividuals might produce proteinase inhibitors in response toattack by herbivores. They confirmed this idea by documentingthat levels of proteinase inhibitors are relatively low in undam-aged tomato leaves, but much higher in leaves that were under-going damage and in leaves at a distance from where damagewas occurring.
T. R. Green and Ryan followed up on this experiment by al-lowing herbivorous beetles to attack one leaf on each of several potato plants. In leaves on the same plant that were notattacked, proteinase inhibitor concentrations averaged 336 �gper mL of liquid from leaves. In leaves of control plants, where noinsect damage had occurred, proteinase inhibitor levels averagedjust 103 �g per mL of leaf juice. This result supports the idea thata hormone produced at the site of damage travels to undamagedtissues and induces the production of proteinase inhibitors.
What is this wound-response hormone? After years of effort,biologists in Ryan’s laboratory succeeded in isolating the mole-cule by purifying the compounds found in tomato leaves andtesting them for the ability to induce proteinase inhibitor pro-duction. The hormone turned out to be a polypeptide, just 18amino acids long, called systemin. It was the first peptide hor-mone ever described in plants. When Gregory Pearce and col-leagues labeled copies of systemin with a radioactive carbonatom, injected the hormone into plants, and then monitored itslocation, they confirmed that systemin moves from damagedsites to undamaged tissues.
Currently, work on systemin and proteinase inhibitor pro-duction is focused on determining each step in the signal trans-duction pathway that alerts undamaged cells to danger. AsFigure 37.15 shows, the data indicate that systemin binds to areceptor on the membrane of an undamaged cell. The activatedreceptor triggers a long series of chemical reactions that eventu-ally result in the synthesis of a molecule called jasmonic acid.Jasmonic acid, in turn, activates the production of at least 15new gene products, including proteinase inhibitors. In this way,plants build potent concentrations of insecticides in tissues thatare in imminent danger of an attack.
Recruiting ParasitoidsIn addition to coping with spines, proteinase inhibitors, andother plant defenses, caterpillars and other herbivorous insectshave enemies of their own. In many cases, these enemies arewasps that lay their eggs in the bodies of the herbivores. Whena wasp egg hatches inside a caterpillar, the wasp larva beginseating its host from the inside out (Figure 37.16). An organism
Chapter 37 Plant Defense Systems 721
FIGURE 37.16 Parasitoids Kill HerbivoresParasitoids lay their eggs in caterpillars and other types of herbivores.As the parasitoid larvae grow, they devour the host. This photoshows wasp larvae emerging from a parasitized caterpillar.
Damaged cell
Undamaged cell
Jasmonic acid
1. When a beetleattacks a potato plant,the damaged cellsproduce the hormonesystemin.
2. Systemin binds to receptors on the membranes of undamaged cells.
3. A series of reactionsproduces jasmonic acid.
4. Jasmonic acid activates transcriptionof proteinase inhibitors that will deter the herbivore from further attack.
Proteinase inhibitors(tastes bad and makes herbivore sick)
Systemin
HOW DOES SYSTEMIN FUNCTION?
FIGURE 37.15 Signals from Damaged Cells Prepare Other Cellsfor AttackSystemin is a hormone produced by herbivore-damaged cells that ini-tiates a protective response in undamaged cells.
FREEMC37.13081923.709-725.final 11/12/01 10:45 AM Page 721
that is free-living as an adult but parasitic as a larva is called aparasitoid. For obvious reasons, parasitoid attacks limit theamount of damage that herbivores do to plants.
The observation that parasitoids seem to be common duringherbivore outbreaks in croplands prompted a question: Dowounded plants release compounds that actively recruit para-sitoids? If so, the distress signal would furnish a novel and ef-fective plant defense system. To explore this idea, T. C. J.Turlings and co-workers collected volatile compounds thatwere released from corn seedlings during attacks by caterpil-lars. (A volatile molecule is one that evaporates rapidly and dif-fuses in the air.) When they analyzed the compounds chemical-ly, the biologists found that insect-damaged leaves produced 11molecules that were not produced by undamaged leaves. Thesevolatile compounds were not produced by leaves that had beencut with a scissors or crushed with a tool; only insect damagetriggered their production.
A key question, though, is whether wasps sense the volatilecompounds and respond to them. To answer this question,Turlings and colleagues set up the experiment diagrammed inFigure 37.17a. The researchers put corn seedlings on either sideof one end of a wind tunnel, put a female wasp at the other end,and recorded which of the two corn plants she visited. As thedata at the bottom of the figure show, the wasps were muchmore likely to visit corn plants that had been damaged by cater-pillars than those that had been artificially damaged. The re-searchers could make artificially damaged plants just as attrac-tive to the wasps, however, by adding caterpillar saliva.
H. T. Alborn and colleagues followed up on this result by pu-rifying a large series of substances from the saliva of beet army-worm caterpillars. The researchers were able to show that one ofthese molecules, called volicitin, induced damaged leaves to emitthe volatile compounds attractive to wasps. The message of thiswork is that plant cells are able to sense a specific molecule thatis present in caterpillar saliva. As a result, they are able to recog-nize when they are being attacked by an herbivore and sound analarm that recruits help (Figure 37.17b).
722 Unit 7 How Plants Work
(a) Wasps can detect herbivore-damaged corn seedlings.
(b) MECHANISM OF COMMUNICATION BETWEEN PLANT AND PARASITOID
Wind tunnel(air blowstoward the wasp)
Wasp
Number of wasp choices
Which side did wasp choose?
1. When a caterpillarstarts eating a corn leaf, volicitin is released in its saliva.
2. Volicitin inducesplant cells to releasevolatile molecules.
3. The wasp followsthe scent to the source and attackscaterpillar.
FIGURE 37.17 Wasps Hone In on Volatile Compounds That PlantsRelease in Response to Volicitin(a) In choice tests, wasps are attracted to corn seedlings that havebeen damaged by caterpillars, or to artificially damaged leaves thathave been treated with caterpillar saliva. (b) When plant cells sensevolicitin, they release the volatile compounds sensed by parasitoidwasps. QUESTION In part (a), why did the experimenters use artifi-cially damaged leaves as a comparison treatment? Why not use un-damaged leaves?
FREEMC37.13081923.709-725.final 11/12/01 10:45 AM Page 722
Chapter 37 Plant Defense Systems 723
Essay Chemical Prospecting
The effort to find naturally occurring compounds that can beused as drugs, fragrances, insecticides, herbicides, or fungicideshas been called chemical prospecting. Traditionally, plant sec-ondary metabolites—and particularly defense compounds—have been a rich source of these molecules. For example, theprominent anticancer drug vincristin/vinblastine was first syn-thesized from a plant native to Madagascar called the rosy peri-
winkle; the anticancer drug taxol was discovered in extractsfrom the Pacific yew tree, which lives only in old-growth forestsof the Pacific Northwest region of North America. These drugsare effective because they kill certain types of rapidly growingcells, but their normal function in periwinkles and Pacific yewmay be to harm herbivores. Similarly, the psychoactive proper-ties of alkaloids such as morphine, cocaine, nicotine, and caf-feine are almost certainly related to their role in plant defense.The natural function of tetrahydrocannabinol, the active ingre-dient in marijuana, is probably to injure the plant’s enemies.
Here we consider two recent twists on traditional efforts topurify plant defense compounds and screen them for medical oragricultural use. In one effort, Ilya Raskin and co-workers havebeen able to stimulate plants that are growing hydroponical-ly—that is, in liquid culture instead of in soil—to churn outlarge quantities of defense chemicals. The researchers do this by
exposing root cells to a fragment of a bacterial cell wall or atoxin from a fungus, and then collecting the defense com-pounds that are produced in response. To date, Raskin’s grouphas been able to collect 5000 samples of materials exuded bythe roots of 700 different plant species. Some of the extractshave been found to kill various types of cancer cells growing inculture. It is still unknown, however, whether this novel strate-gy for producing and purifying defense compounds will resultin the discovery of important new drugs.
The second innovative research program is exemplified byrecent work on a group of fungi called mycoparasites (fungi-parasites). Mycoparasites infect and parasitize other fungi.Matteo Lorito and associates were able to clone a gene from amycoparasite that encodes an enzyme called endochitinase.This enzyme breaks down chitin, which is a major componentof cell walls in many species of pathogenic fungi. The mycopar-asite uses this enzyme to open the cell walls of its hosts so thatit can enter and begin to feed, just as pathogenic fungi use cuti-nase to open the cuticle of plants. When the researchers insert-ed the gene for endochitinase into tobacco and potato plants,the transgenic individuals manufactured the enzyme and werehighly resistant to fungal infection.
The work by Lorito and colleagues illustrates a generaltheme in current research on plant diseases. Chemical prospect-ing has a major new thrust: gene prospecting. If importantgenes—such as the R alleles found in disease-resistant species,genes for particularly effective defense compounds, or se-quences that encode antibiotics such as endochitinase—can beintroduced into crop plants safely, rapid improvements in pestresistance might be possible. If so, then farmers might be ableto reduce their dependence on insecticides, fungicides, andother compounds that are expensive and that can be dangerousto humans and wildlife when improperly handled.
The psychoactive properties ofalkaloids such as morphine,
cocaine, nicotine, and caffeine arealmost certainly related to their
role in plant defense.
Chapter Review
SummaryBy providing a barrier to entry, the epidermis and cuticle ofplants act as the first line of defense against pathogens. Somefungi pierce this barrier by secreting an enzyme called cutinasethat cleaves some of the molecules found in cuticle. Bacteriaand some fungi circumvent the barrier by entering tissuesthrough open stomata or wound sites. In many plants, the epi-dermis contains defensive hairs or spines and serves as the firstline of defense against herbivores as well as pathogens.
Many plants discourage herbivores by lacing their tissueswith poisons. Analyses of the structures of these poisons have
supported the hypothesis that many or most are secondarymetabolites. In both structural and evolutionary terms, manysecondary metabolites are closely related to fundamentally im-portant compounds such as amino acids. When researchers ex-plored the mode of action of the common secondary metabo-lites known as alkaloids, they discovered that many of thesemolecules work by poisoning several different biochemicalpathways or processes at the same time.
Many plants do not produce defense compounds all of thetime, however. Instead, they produce antibiotics and pesticides
FREEMC37.13081923.709-725.final 11/12/01 10:45 AM Page 723
724 Unit 7 How Plants Work
only when an infection or herbivore attack has started. Thispattern is sensible because defense compounds are energeticallyexpensive to produce, and because plants face trade-offs in al-locating resources to defense or to growth and reproduction.The prediction that defense is costly and that trade-offs occurhas been confirmed experimentally. For example, when Ara-bidopsis thaliana plants grow in an environment that lackspathogens and herbivores, lightly defended individuals producethe most seeds. But in environments where pathogens and her-bivores are common, heavily defended individuals produce themost offspring.
When an attack by a pathogen begins, resistant plants re-spond with a rapid series of events called the hypersensitive re-sponse (HR). The HR is believed to be triggered when the prod-uct of a resistance gene (R) located in the host binds to theproduct of an avirulence (avr) gene secreted by the pathogen.Resistance loci are found in large gene families and are highlypolymorphic, meaning that many different alleles exist at eachlocus. Individuals that produce many different R gene productshave the ability to recognize many different pathogen products.As a result, they are immune to a wide array of pathogens.
During the HR, cells at the point of infection commit sui-cide. Cell death results from the combined action of nitrousoxide (NO) and reactive oxygen intermediates (ROI). Cellwalls around the point of infection are also reinforced throughthe action of ROI. In some cases, antibiotic compounds calledphytoalexins are produced as part of the HR as well.
The HR is commonly followed by a slower response thatleads to systemic acquired resistance (SAR). SAR occurs when ahormone, which may be salicylic acid, travels from the infec-tion site to nearby tissues and triggers the expression of a spe-cific set of genes.
When an attack by an herbivore begins, plants respond byproducing proteinase inhibitors. The synthesis of these poisonsoccurs throughout the plant and is triggered by a polypeptidehormone called systemin. Once systemin binds to a receptor onthe membrane of undamaged cells, a series of chemical reac-tions leads to the manufacture of proteinase inhibitors andheightened plant defense. Some plants are also able to sense spe-cific compounds in the saliva of herbivores; in response, they se-crete volatile compounds that attract parasitoids.
b. The HR is not triggered, and the infection spreads.c. The SAR is triggered, and the infection does not spread to sur-
rounding tissue.d. NO and ROI combine to kill cells at the site of infection.
4. What is a phytoalexin?a. any plant secondary metaboliteb. any plant secondary metabolite that is used in defensec. any plant secondary metabolite that is used in defense and pro-
duced in response to a pathogen attackd. any plant secondary metabolite that is used in defense and pro-
duced constitutively
5. What happens after volicitin binds to a receptor in a plant cell?a. SAR is initiated.b. Proteinase inhibitor production is initiated.c. Volatiles are released that attract parasitoids.d. Phytoalexin production is initiated.
QuestionsContent Review
1. Why is it significant that fungi produce the enzyme called cutinase?a. It cleaves molecules in plant cuticle and allows the fungus to
enter.b. It neutralizes R gene products, so the HR does not occur.c. It prevents the release of salicylic acid, so SAR does not occur.d. It prevents the synthesis of secondary metabolites.
2. What does the gene-for-gene hypothesis claim?a. Plant defense systems are heritable, or genetically based.b. R loci occur in families because of gene duplication events.c. Secondary metabolites are the most efficient form of plant de-
fense.d. Plant defense systems activate when an R gene product binds to
an avr gene product.
3. What happens if none of the molecules produced by a pathogenbind to an R gene product?a. The HR is triggered, and the infection is stopped in its tracks.
4. Many of the fragrant molecules given off by plants to attract polli-nators are secondary metabolites. Suggest a scenario for the evolu-tion of these attractant compounds, analogous to the presentationin the text regarding the evolution of defense compounds.
5. Describe some of the similarities and differences in plant defensesagainst pathogens and herbivores.
6. The HR has been called “an efficient suicide program.” Why?
Conceptual Review
1. Summarize the events that occur during the HR and SAR. In whatway are these responses different? How are they complementary?
2. Why is it significant that R loci are usually clustered on the samechromosome? Why is it important that R loci are usually highlypolymorphic?
3. What evidence suggested that a hormone travels from the site of in-sect damage to undamaged tissues? How did investigators discoversystemin and confirm that it is the wound-response hormone?
2. Suppose that you were studying two closely related species of plantsand were able to document that one has much higher concentra-tions of tannins than the other. Which species would you expect togrow more slowly and produce fewer seeds each year? Why?
Applying Ideas
1. Suppose you are walking along a garden path with a friend andnotice a plant leaf with several small, white, dead-looking blotch-es. Nearby, an individual from the same species has leaves that arecompletely yellow and wilted. Explain to your friend what youthink is going on. How could you test your hypothesis?
FREEMC37.13081923.709-725.final 11/12/01 10:45 AM Page 724
Chapter 37 Plant Defense Systems 725
3. Researchers have recently begun studying a secondary metabolitecalled myrosinase in the plant Brassica rapa. (Brassica rapa is animportant source for vegetable oils used in cooking.) Myrosinase isan effective deterrent for herbivorous beetles. In one study, biolo-gists mated individuals with high concentrations of myrosinasewith one another for several generations and individuals with lowconcentrations of myrosinase with one another for several genera-tions. In this way, they created populations with high versus lowconcentrations of myrosinase. Then they documented how mucheach population was visited by pollinators. Predict which popula-tion was more successful in attracting long visits from pollinators.To see if you are correct, check the paper published by SharonStrauss and co-workers in the journal called Evolution, volume 53,pages 1105–1113. In what way does this experiment show a novelcost of defense?
4. Researchers have been able to document that plants undergoinginsect attack release jasmonic acid as a volatile. Other plants that
are growing nearby synthesize increased amounts of proteinase in-hibitors in response. Do these observations surprise you? Why orwhy not?
5. Review the text section that analyzes the experiments on phy-toalexins produced by chickpeas. Note the close correspondencebetween the amount and type of phytoalexins produced by differ-ent chickpea strains in response to different strains of fungi. Sug-gest a hypothesis that explains this observation in terms of differ-ences among the R alleles of chickpea and the avr alleles of fungusstrains. How would you test your hypothesis?
6. The experimental approach diagrammed in Figure 37.8 is widelyused and is called the yeast two-hybrid system. Note that a hybridis produced when two unlike things combine. Explain why thename is appropriate and why the strategy is an effective way tostudy protein-protein interactions in cells.
Plant Defense and Reproductive Energy Budgets This article describes how the energy cost of reproduction must bebalanced by the benefits gained by shunting energy into the devel-opment of plant defenses.
Plant-Produced Pesticides This article describes some of the toxic chemicals that plants can ac-tively produce to fight off herbivory.
CD-ROM and Web ConnectionCD Activity 37.1: Plant Defenses (animation)
(Estimated time for completion � 10 min)How have plants adapted to survive in a world filled with herbivores?
At your Companion Website (http://www.prenhall.com/freeman/biology), you will find self-grading exams and links to the following re-search tools, online resources, and activities:
Gene Silencing This article explains how a plant virus can alter the normal expres-sion of genes in the plant.
Marchand, P. 2001. In the field: riding the witches broom. Natural History 110(May): 40–41.
Stanton, M., and T. Young. (1999). Thorny relationships. Natural History 108(September): 28–30.
Additional ReadingBaker, B., P. Zambryski, B. Staskawicz, and S.P. Dinesh-Kumar. 1997. Signaling
in plant-microbe interactions. Science 276: 726–733. Reviews many of theplant defense systems introduced in this chapter.
Barlow, C. and M. Rothman. 2001. Ghost stories from the ice age. Natural His-tory 110 (September): 62–67.
FREEMC37.13081923.709-725.final 11/12/01 10:45 AM Page 725