Estuarine, Coastal and Shelf Science (1985) 20,593-602 Population Biology of the Portunid Crab Callinectes arcuatus Ordway in the Gulf of Nicoya, Costa Rica, Central America Ana I. Dittel”, C. E. Epifaniob and Juan Bautista Chavarria” “Centro de Investigation en Ciencias Del Mary Limnologia, Universidad de Costa Rica, SunJose, Costa Rica, America Central and bColiege of Marine Studies, University of Delaware, Lewes, DE 19958, U.S.A. Received 29 February 1984 and in revised form 19June 1984 Keywords: crabs; migration; recruitment; seasonal variations; estuaries; Cen- tral AmericanWest Coast; population composition; life cycles Tropical blue crabs Callinectes arcuatus were collected by trawling in the Gulf of Nicoya on the Pacific coast of Costa Rica, Central America. The gulf popu- lation was generallydominated by females. Adult females werecommon in the upper, moreestuarine regions of the gulf during rainy season, but appeared to migrateto the lower gulf during dry season for spawning. Biomass varied with seasonal changes in abundance, but was generally similar to biomass of C. sapidus in Chesapeake Bay. Analysis of size frequency indicated that the population is numerically dominated by adults during Januaryand February and that juveniles arecommon during the remainder of the year. Extrapolation of available datasuggests that female crabs reach maturity in approximately one yearafter hatching. Introduction The Atlantic blue crab Cal2inectes sapidus Rathbun supports important fisheries along the Atlantic and Gulf coastsof the United States. Accordingly, its general biology has been investigated extensively (Van Engle, 1958). However, the genus CaZZinectes is primarily tropical, and only three of its speciesnormally range into temperate waters (Williams, 1974). The life histories of the tropical specieshave been sparsely studied. There have been a few ecological studies of two species of blue crab common in tropical estuariesalong the Pacific coast of South America (Norse 8zEstevez, 1977), and the tax- onomy and known distribution of the genus in the tropics has been reviewed (Rathbun, 1930; Garth & Stevenson, 1966; Williams, 1974). Recently there has been increasing interest in the tropical species Callinectes arcuatus Ordway asthis crab has a commercial potential that is presently unexploited. Paul (1982) studied the abundance, breeding, and growth of C. arcuatus in a lagoonal estuary on the Pacific coast of Mexico. But despite its common occurrence, there has been little ecological study of the species in the estuariesof Central America (DeVries et al., 1984). 593 NY-7714’85’050593i- 10 $03.00/0 0 1985 Academic Press Inc. (London) Ltmited
Transcript
Estuarine, Coastal and Shelf Science (1985) 20,593-602
Population Biology of the Portunid Crab Callinectes arcuatus Ordway in the Gulf of Nicoya, Costa Rica, Central America
Ana I. Dittel”, C. E. Epifaniob and Juan Bautista Chavarria” “Centro de Investigation en Ciencias Del Mary Limnologia, Universidad de Costa Rica, SunJose, Costa Rica, America Central and bColiege of Marine Studies, University of Delaware, Lewes, DE 19958, U.S.A.
Received 29 February 1984 and in revised form 19June 1984
Keywords: crabs; migration; recruitment; seasonal variations; estuaries; Cen- tral American West Coast; population composition; life cycles
Tropical blue crabs Callinectes arcuatus were collected by trawling in the Gulf of Nicoya on the Pacific coast of Costa Rica, Central America. The gulf popu- lation was generally dominated by females. Adult females were common in the upper, more estuarine regions of the gulf during rainy season, but appeared to migrate to the lower gulf during dry season for spawning. Biomass varied with seasonal changes in abundance, but was generally similar to biomass of C. sapidus in Chesapeake Bay. Analysis of size frequency indicated that the population is numerically dominated by adults during January and February and that juveniles are common during the remainder of the year. Extrapolation of available data suggests that female crabs reach maturity in approximately one year after hatching.
Introduction
The Atlantic blue crab Cal2inectes sapidus Rathbun supports important fisheries along the Atlantic and Gulf coasts of the United States. Accordingly, its general biology has been investigated extensively (Van Engle, 1958). However, the genus CaZZinectes is primarily tropical, and only three of its species normally range into temperate waters (Williams, 1974). The life histories of the tropical species have been sparsely studied. There have been a few ecological studies of two species of blue crab common in tropical estuaries along the Pacific coast of South America (Norse 8z Estevez, 1977), and the tax- onomy and known distribution of the genus in the tropics has been reviewed (Rathbun, 1930; Garth & Stevenson, 1966; Williams, 1974).
Recently there has been increasing interest in the tropical species Callinectes arcuatus
Ordway as this crab has a commercial potential that is presently unexploited. Paul (1982) studied the abundance, breeding, and growth of C. arcuatus in a lagoonal estuary on the Pacific coast of Mexico. But despite its common occurrence, there has been little ecological study of the species in the estuaries of Central America (DeVries et al., 1984).
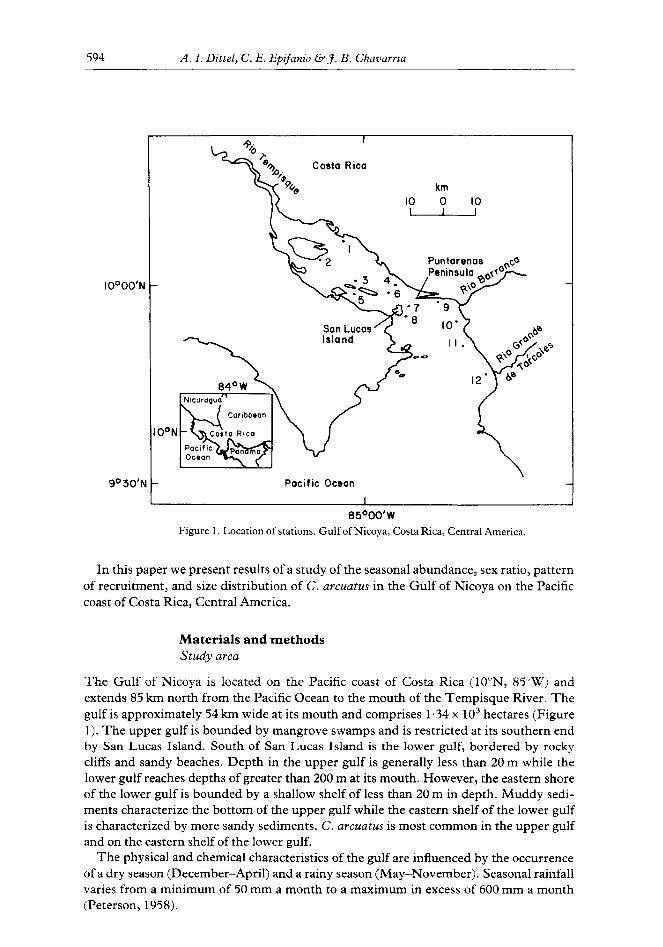
Figure 1. Location of stations. Gulf of Nicoya, Costa Rica, Central America.
In this paper we present results of a study of the seasonal abundance, sex ratio, pattern of recruitment, and size distribution of C. arcuatus in the Gulf of Nicoya on the Pacific coast of Costa Rica, Central America.
Materials and methods Study area
The Gulf of Nicoya is located on the Pacific coast of Costa Rica (lO”N, 85”W) and extends 85 km north from the Pacific Ocean to the mouth of the Tempisque River. The gulf is approximately 54 km wide at its mouth and comprises 1.34 x lo3 hectares (Figure 1). The upper gulf is bounded by mangrove swamps and is restricted at its southern end by San Lucas Island. South of San Lucas Island is the lower gulf, bordered by rocky cliffs and sandy beaches. Depth in the upper gulf is generally less than 20 m while the lower gulf reaches depths of greater than 200 m at its mouth. However, the eastern shore of the lower gulf is bounded by a shallow shelf of less than 20 m in depth. Muddy sedi- ments characterize the bottom of the upper gulf while the eastern shelf of the lower gulf is characterized by more sandy sediments. C. arcuatus is most common in the upper gulf and on the eastern shelf of the lower gulf.
The physical and chemical characteristics of the gulf are influenced by the occurrence of a dry season (December-April) and a rainy season (May-November). Seasonal rainfall varies from a minimum of 50 mm a month to a maximum in excess of 600 mm a month (Peterson, 1958).
Tropical blue crabs 595
TABLE 1. Gulf of Nicoya, Costa Rica, Central America. Mean values for temperature, salinity and dissolved oxygen in bottom waters during dry and rainy seasons, 1981
Season Temperature (“C) Salinity (%o) Oxygen (ppm)
Sampling procedures Crabs (Callinectes arcuatus Ordway) were collected at 12 stations (Figure 1) using a semi-balloon shrimp trawl with a 9.1 m head rope and a 6.5 m mouth opening when fish- ing. Mesh size in the cod end of the net was 3.6 cm when stretched diagonally. The net was towed at an effective speed of 2.5 km h-i against the prevailing current for 10 min. The estimated area of bottom swept per trawl was 2675 m. The six stations in the upper gulf and stations 7 and 8 in the lower gulf were sampled monthly while stations 9-12 in the lower gulf were sampled approximately weekly. Water temperature and salinity were measured during sampling with a Beckman RS5-3 CST meter, and dissolved oxygen was measured with a YSI oxygen electrode. Mean values for the upper and lower gulf are shown in Table 1.
After each trawl all C. arcuatus individuals were sorted from the rest of the catch, and their sex and the state of maturity of the females (as determined by the shape of the abdomen) was recorded. The carapace width (including lateral spines) of each individual was measured to the closest mm.
In order to estimate biomass, initial samples of 25 male and 25 female crabs of various sizes were taken. Carapace widths of the crabs were measured and the crabs were weighed after drying for 48 h at 60 “C. Regressions of body weight (IV) and carapace width (Y) were performed using the results of the measurements:
males: log w=o.22 Y+o.38, r =0.97,
females: log W=O.lB Y+O.Ol, r=0.94.
These equations were used to calculate the mean biomass of crabs collected at each station during each sampling period. Biomass per square metre was calculated by dividing the biomass collected in a given sample by 2675 m, the area of bottom swept per trawl.
Results
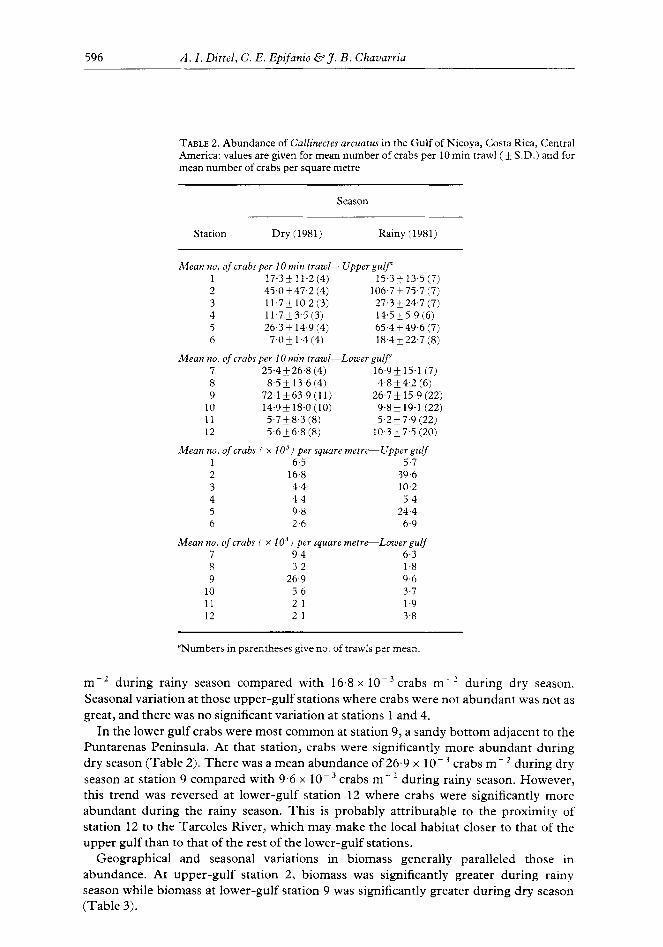
Crabs were more abundant in the upper gulf during rainy season and in the lower gulf during dry season (Table 2). In the upper gulf, crabs were most common at station 2, a shallow (3 m) muddy bottom along the shore of a large island (Isla Chira). Mean abundance during rainy season at station 2 was significantly greater (Mann-Whitney IJ-test; P<O.l) than that during dry season. Mean abundance was 39.6 x 1O-3 crabs
596 A. I. Dittel, C. E. Epifanio &y. B. Chavarria
TABLE 2. Abundance of Callinectes arcuafus in the Gulf of Nicoya, Costa Rica, Central America: values are given for mean number of crabs per 10 min n-awl (+ SD.) and for mean number of crabs per square metre
Season
Station Dry (1981) Rainy (1981)
Mean no. of crabs per 10 min trawl-Upper gulfD 1 17,3&1i.2(4) 15.3 + 13.5 (7) 2 45.0 f 47.2 (4) 106.7 f 75.7 (7) 3 11.7*10.2(3) 27.3k24.7 (7) 4 11.7k3.5 (3) 14.5 +5.9 (6) 5 26.3 + 14.9 (4) 65.4 f 49.6 (7) 6 7.0 f 1.4 (4) 18.4 k22.7 (8)
Mean no. of crabs per 10 min trawl-Lower gulf” 7 25.4k26.8 (4) 16.9* 15.1 17) 8 8.5 & 13-6 (4) 4.8 k4.2 (6) 9 72.1*63-9(11) 26.7& 15.9 (22)
Mean no. of crabs i x 10” I per square metre-Upper gulf 1 6.5 5.7 2 16.8 39.6 3 4.4 10.2 4 4.4 5-4
zJ 2.6 9.8 24.4 6.9
Mean no. of crabs i x 1 O-’ i per square metre-Lower gulf 7 9.4 6.3 8 3.2 1.8 9 26.9 9.6
10 5,6 3.7 11 2.1 1.9 12 2.1 3.8
“Numbers in parentheses give no. of trawls per mean
m -’ during rainy season compared with 16.8 x 1O-3 crabs rn-’ during dry season. Seasonal variation at those upper-gulf stations where crabs were not abundant was not as great, and there was no significant variation at stations 1 and 4.
In the lower gulf crabs were most common at station 9, a sandy bottom adjacent to the Puntarenas Peninsula. At that station, crabs were significantly more abundant during dry season (Table 2). There was a mean abundance of 26.9 x lo- ’ crabs m- ’ during dry season at station 9 compared with 9.6 x lo- 3 crabs m-l during rainy season. However, this trend was reversed at lower-gulf station 12 where crabs were significantly more abundant during the rainy season. This is probably attributable to the proximity of station 12 to the Tarcoles River, which may make the local habitat closer to that of the upper gulf than to that of the rest of the lower-gulf stations.
Geographical and seasonal variations in biomass generally paralleled those in abundance. At upper-gulf station 2, biomass was significantly greater during rainy season while biomass at lower-gulf station 9 was significantly greater during dry season (Table 3).
Tropical blue crabs 597
TABLE 3. Biomass of Callinectes arcuatus in the Gulf of Nicoya, Costa Rica, Central America. Values are given for mean dry weight biomass (g) of crabs per 10 min trawl ( + S.D.) and for mean dry weight biomass (g) per square metre
Season
Station Dry (1981) Rainy (1981)
Mean dry wt biomass ig/ per 10min trawl-Uppergul~ 1 742.0 & 452.8 (4) 542.7 i 537.7 (9) 2 2625.2k2561.8 (4) 5223.8 f 3147.5 (7) 3 598.0 + 675.0 (3) 1273.4 f 1424.0 (7) 4 775.9 k 196.0 (3) 533.3 i 282.8 (6) 5 1551,8* 1238.3 (4) 2950.9 i 1881.9 (7) 6 278.5 f 98.0 (4) 702.8&810-I(8)
Meal2 dr.v wt biomass (g I per square metre-Lower&f 7 0.03 0.20 8 0.15 0.06 9 1.25 0.50
10 0.15 0.05 11 0.10 0.02 12 0.09 0.18
The population in the gulf was numerically dominated by females (Table 4). As pre- vious studies have shown that mature male C. arcuatus are often common in the lower reaches of tidal rivers (Norse & Estevez, 1977), we assume that males are more common in the areas of the Tempisque River north of our stations 1 and 2. Nevertheless the sex ratio in the population sampled in our study varied with season and location. Females were more abundant relative to males during rainy season at upper-gulf station 2 while they showed greater relative abundance during dry season at lower-gulf station 9. This trend was less pronounced (or reversed) at stations where crabs were less abundant.
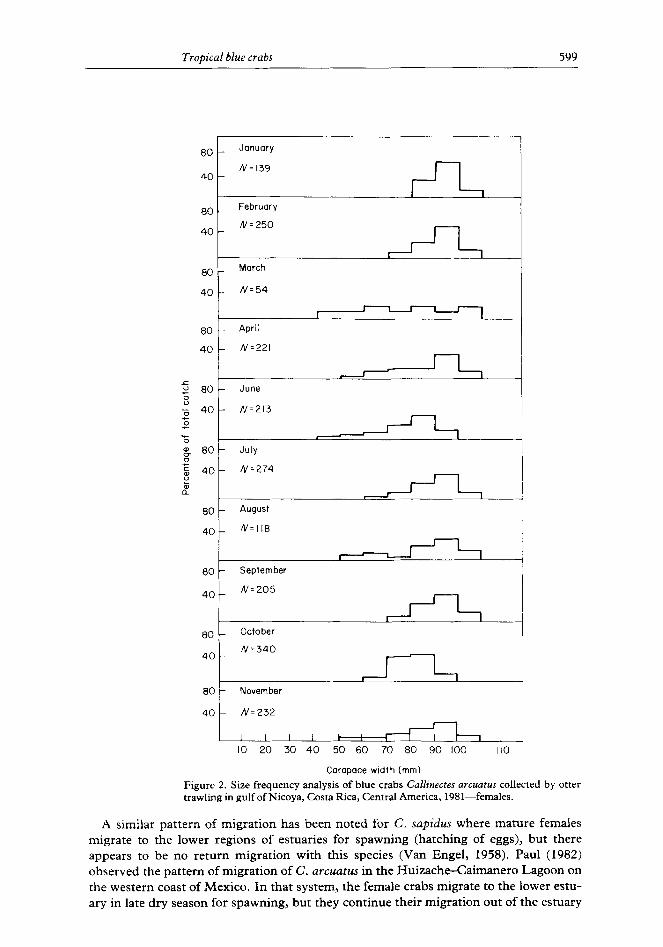
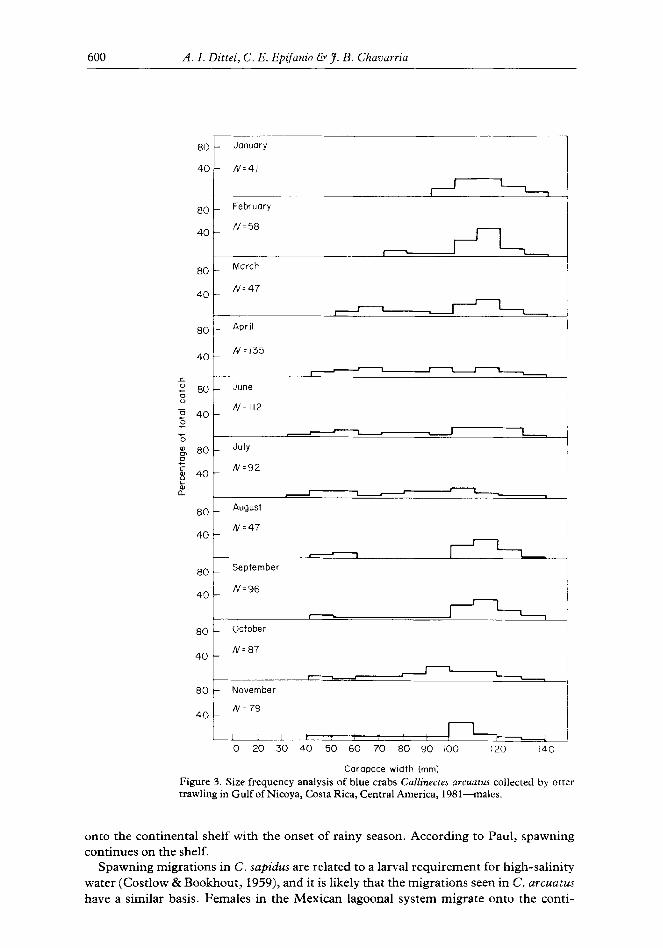
Male crabs reached a larger size than females, and both male and female components of the population were dominated by large individuals at the stations sampled (Figures 2 and 3). Female crabs in the genus Callinectes undergo a change in shape of the abdomen upon reaching maturity, and nearly all females with carapace widths greater than 80 mm were mature. Males do not undergo any external morphological change at maturity, but it was assumed that crabs greater than 90 mm in carapace width were sexually mature (Paul, 1982). Sexually mature crabs made up the largest proportion of the sampled populations during January and February, the period of maximum spawning in the gulf
598 A. I. Dittel, C. E. Epijanio &J. B. Chavarria
TABLE 4. Seasonal comparison of sex ratios in a population of blue crabs Callinecres arcuatus in the Gulf of Nicoya, Costa Rica. Central America. Values are ratio of number of females to males
Season
Station Dry Rainy
2.6 5.0 1.3 2.1 2.2 1.6 2.9 3.6 0.6 3.1 8.3 5.0
4.2 7.1 12.5 25.0 3.3 1.1 4.5 0.8 7.7 1.4 4.2 2.5
(DeVries et al., 1984). Substantial numbers of juveniles were apparent during the remainder of the year, but we were unable to elucidate peaks of recruitment. Crabs smaller than 40 mm were not sampled quantitatively as they were not consistently retained in the net.
Discussion
Results of our investigation have shown that C. arcuatus occurs throughout the upper Gulf of Nicoya and along the eastern shore of the lower Gulf. However, the population was not uniformly distributed in these areas, and there were distinct concentrations of crabs at stations 2 and 5 along the western shore of the upper gulf and at station 9 near the Puntarenas Peninsula in the lower gulf. The population was numerically dominated by females regardless of season or location. However, as earlier studies of C. arcuatus (Norse & Estevez, 1977) have shown that assemblages of mature males extend farther into low-salinity waters than do females, it appears likely that our results concerning sex ratio are biased by the fact that we did not sample in the lower Tempisque River north of our station 1. Nevertheless, the ratio of females to males at station 2 increased during rainy season while a similar ratio at station 9 decreased during rainy season. This was generally the case with the exception of stations 3 and 6 in the upper gulf and stations 7 and 8 in the lower gulf.
[However, if one considers the abundance of mature females and disregards immature females (data from DeVries, 1982), there is a higher ratio of females to males during rainy season in the upper gulf and during dry season in the lower gulf at these stations, as well.] Therefore, our results are consistent with those of DeVries et al., (1984) in that mature females appear to migrate to the lower gulf to spawn during dry season and migrate back to the upper gulf during rainy season. It is not clear how many times a given crab may repeat this cycle.
Tropical blue crabs 599
80
40
80
40
80
40
80
40
80
40
80
40
80
40
, -
/ -
January
February
N=250
March
N=54
April
N=221
June
N=213
July
N-274
August
H118 September
N=205
October
N=340
November
N=232
I I I I t : I
IO 20 30 40 50 60 70 80 90 100 110
Carapace width (mm)
Figure 2. Size frequency analysis of blue crabs CaNinectes arcuatus collected by otter trawling in gulf of Nicoya, Costa Rica, Central America, 1981-females.
A similar pattern of migration has been noted for C. supidus where mature females migrate to the lower regions of estuaries for spawning (hatching of eggs), but there appears to be no return migration with this species (Van Engel, 1958). Paul (1982) observed the pattern of migration of C. arcuatus in the Huizache-Caimanero Lagoon on the western coast of Mexico. In that system, the female crabs migrate to the lower estu- ary in late dry season for spawning, but they continue their migration out of the estuary
600 A. I. Dittel, C. E. Epifanio DJ. B. Chavarria
80 - January
40- N-41
80 _ February
40 - N=50
8. _ March
1
40
t
IV=47
80 t
April
40 - N=l35
L g 80 - June 0
g N=ll2 40-
b L.-l
&!, 80 - J’-“Y 0
F g 40 - N=g2 kJ a 7 - 1
0. _ August 4
40 - N=47
8. _ September
40 N=96 -
80 - October
40 N=87
-
80 - November
4oc N=79
-/ 0 20 30 40 50 60 70 80 90 100 120 140
Carapace width (mm) Figure 3. Size frequency analysis of blue crabs Callinecres U~CUU~US collected by otter trawling in Gulf of Nicoya, Costa Rica, Central America, 1981-males.
onto the continental shelf with the onset of rainy season. According to Paul, spawning
continues on the shelf. Spawning migrations in C. supidus are related to a larval requirement for high-salinity
water (Costlow & Bookhout, 1959), and it is likely that the migrations seen in C. arcuatus have a similar basis. Females in the Mexican lagoonal system migrate onto the conti-
Tropical blue crabs 601
nental shelf because salinities in that estuary fall to nearly zero during rainy season. In contrast, salinities in the region of the Gulf of Nicoya that we studied rarely fall below 25%0 (Voorhis et al., 1983). This apparently allows females to migrate back to the upper estuary after spawning.
Biomass of crabs per square metre at stations 2 and 9 in the Gulf of Nicoya was similar to that reported for C. sapidus in Chesapeake Bay (Miller et al., 1975), but was lower than biomass reported for C. arcuatus in the Huizache-Caimanero Lagoon in Mexico (Paul, 1982). However, Paul’s values appear to be in error as his biomass per trawl, the area swept by the trawl, and the biomass per square metre do not correspond.
Our analysis of size frequency shows great similarity between the C. arcuatus popu- lations in Nicoya and the Huizache-Caimanero system. In both estuaries males reach a larger size than females and both populations are dominated by large individuals during January and February. Juveniles first appear in both estuaries in March, but the larger size classes do not disappear in Nicoya as they do in the Huizache-Caimanero. This can be explained by the fact that adult crabs migrate from the Mexican estuary to the adjacent continental shelf during rainy season while adults remain in the Gulf of Nicoya regardless of season.
The juveniles that appeared in our samples in March were greater than 40mm in carapace width and, hence, were not the result of the maximum spawning activity that occurred in January and February. Based on laboratory growth studies, crabs hatched in January would not reach a size of 40 mm until June (Dittel & Epifanio, 1984). However, we did not observe a large pulse of 40 mm crabs in June, possibly due to the fact that we did not sample the shallow, inshore habitat where 40 mm juveniles may have been concentrated (Van Engle, 1959). We did observe a pulse of 70-90 mm female crabs in October and of 90-l 10 mm males in October and November. Extrapolation of laboratory growth rates suggest that these crabs may have been spawned in January and February.
In any case, the extrapolation of growth rates from size-frequency data is difficult in tropical species with protracted spawning periods. In the case of C. arcuatu~ in the Gulf of Nicoya, there is a period of maximum spawning activity in January and February, but a lower level of spawning continues throughout the year (DeVries et al., 1984). Accord- ingly, juveniles are recruited into the population year-round so that cohorts are difficult to distinguish. Also, crabs in the genus Callinectes do not continue to increase in carapace size throughout their lives, but instead reach a maximum size at their terminal molt. Consequently, there can be considerable overlap in the actual age of crabs in the largest size class. This can result in the largest size class making up a disproportionately large share of the population. An additional complication with crabs of the genus Callinectes is that very small juveniles are often concentrated in shallow water adjacent to the shore while larger crabs are found in deeper water. Therefore, samples obtained by otter trawling would tend to overestimate the importance of the larger size classes.
Paul (1982) constructed growth curves for males and females from his Mexican study site, but the technique employed was limited by a qualitative assessment of modes in the size-frequency analysis and by the small sample size during much of the study period. Regardless, Paul’s growth curve predicts that C. UTCUU~US females grow from a carapace width of 20 mm to a mature size of 90 mm in approximately eight months, and this is in close agreement with the results of laboratory studies of growth in the species (Dittel & Epifanio, 1984).
Based on all available evidence, the life cycle of C. arcuatus in the Gulf of Nicoya appears to include a major spawning period during the middle of the dry season in
602 A. I. Dittel, C. E. Epifanio &J. B. Chavarria
January and February. Zoeal and megalopal development takes approximately 70 days and an additional 60 days are required to grow from a newly metamorphosed juvenile to a carapace width of 20mm (Dittel & Epifanio, 1984). Both laboratory and field data (Dittel & Epifanio, 1984; Paul, 1982) indicate that an additional 240 days are required for females to grow from 20 mm to a mature size of 90 mm, resulting in a total of 370 days required for growth from hatching to maturity. As larger crabs are disproportionately common throughout the year, it seems likely that crabs live for some time after reaching maximum size and that the number of crabs in the larger size classes is continually augmented by new individuals resulting from the low level of spawning that occurs throughout the year in the lower gulf.
Acknowledgements
This study was supported by the International Co-operative Assistance Program of the Office of Sea Grant, the University of Costa Rica, and the Organization of American States. We wish to thank the students and staff of the Universities of Costa Rica and Delaware who assisted in this work. We extend special thanks to Dr Manuel M. Murillo of the University of Costa Rica who provided general supervision of the field work.
References
Costlow Jr, J. D. & Bookhout, C. G. 1959 The larval development of Callinectes sapidus Rathbun reared in the laboratory. Biological Bulletin 115,3X-396.
DeVries, M. C. 1982 Seasonal and lunar spawning of Callinectes arcuatus Ordway in the Gulf of Nicoya, Costa Rica. MS thesis. University of Delaware, Newark, Delaware. 160 pp.
DeVries, M. C., Epifanio, C. E. & Dittel, A. I. 1984 Reproductive periodicity of the tropical crab Callinectes arcuats Ordway in Central America. Estuarine, Coastal and Shelf Science 17,709-716.
Dittel, A. I. & Epifanio, C. E. 1984 Growth and development of the portunid crab Cullinectes urcuutus Ordway: zoeae, megalopae and juveniles. Journal of Crustacean Biology (in press).
Garth, J. S. & Stephenson, W. 1966 Brachyura of the Pacific coast of America, Brachyrhyncha: Portunidae. Allen Hancock Monographs in Marine Biology 1,1-154.
Miller, R. E., Sulkin, S. D. At Lippson, R. L. 1975 Composition and seasonal abundance of the blue crab Callinectes sapidus Rathbun in the Chesapeake and Delaware Canal and adjacent waters. Chesapeake Science 16,27-31.
Norse, E. A. & Estevez, M. 1977 Studies on portunid crabs from the eastern Pacific. I. Zonation along environmental stress gradients from the coast of Colombia. marine BioZogy 40,365-373.
Paul, R. K. 1982 Abundance, breeding and growth of Callinectes arcuatus Ordway and Callinectes toxotes Ordway (Decapoda, Brachyura, Portunidae) in a lagoon system on the Mexican Pacific coast. Estuarine, Coastal and Shelf Science 14,3-26.
Peterson, C. L. 1958 The physical oceanography of the Gulf of Nicoya, Costa Rica, a tropical estuary. Bulletin of the Inter-American Tropical Tuna Commission 3,139-188.
Rathbun, M. J. 1930 The cancroid crabs of America of the families: Eurylidae, Portunidae, Atelecyclidae, Cancridae and Xanthidae. Bulletin of the U.S. National Museum 1.52, l-609.
Van Engle, W. A. 1958 The blue crab and its fishery in Chesapeake Bay. I. Reproduction, early develop- ment, growth, and migration. Commercial Fishery Review 20,617.
Voorhis, A. D., Epifanio, C. E., Maurer, D., Dittel, A. I. & Vargas, J. A. 1983 The estuarine character of the Gulf of Nicoya, an embayment on the Pacific coast of Central America. Hydrobiologia 99,225-237.
Williams, A. B. 1974 The swimming crabs of the genus Callinectes. Fishery Bulletin 72,685-798.