Journal of Insect Physiology 53 (2007) 332–342 Possible evidence for morning and evening oscillators in Drosophila melanogaster populations selected for early and late adult emergence Shailesh Kumar, Dhanya Kumar, V.S. Harish, S. Divya, Vijay Kumar Sharma Chronobiology Laboratory, Evolutionary and Organismal Biology Unit, Jawaharlal Nehru Centre for Advanced Scientific Research, P.O. Box 6436, Jakkur, Bangalore 560064, Karnataka, India Received 6 November 2006; received in revised form 12 December 2006; accepted 13 December 2006 Abstract In this paper, we report the results of our study aimed at a systematic analysis of the circadian phenotypes of fruit flies Drosophila melanogaster selected for early and late adult emergence, in light of the ‘‘morning and evening oscillator’’ (M and E) model for circadian clocks. We monitored adult emergence and activity/rest rhythms in these flies under light/dark (LD) cycles with short (8:16 h), normal (12:12 h) and long (16:8 h) photoperiods, as well as under constant darkness (DD). Across all the three LD cycles, the early populations displayed a morning phenotype with peak of emergence and activity occurring earlier than the controls and greater anticipation to ‘‘lights-on’’ and weak anticipation to ‘‘lights-off’’, while the late populations showed an evening phenotype with peak of emergence and activity occurring later than the controls and greater anticipation to lights-off and weak anticipation to lights-on. The gate of adult emergence and duration of activity in the early populations was narrower than the controls, while those of the late populations were wider than the controls. In addition, the circadian periodicities of adult emergence and activity/rest rhythms of the early flies were significantly shorter than the controls, while those of the late flies were significantly longer than the controls. In summary, the circadian phenotypes indicate that the early populations have evolved a dominant M oscillator, while the late populations have evolved a dominant E oscillator, thus providing an empirical support for the M and E model in Drosophila. r 2007 Elsevier Ltd. All rights reserved. Keywords: Drosophila; Eclosion; Activity; Selection; Photoperiod; Morning and Evening oscillators 1. Introduction Organisms track time in their local environment by entraining their circadian clocks to the light/dark (LD) cycles, a phenomenon that is of paramount importance for their survival under fluctuating environments (Pittendrigh and Daan, 1976; Daan, 1981). Several studies have been carried out in the past to understand how organisms adjust to periodic LD cycles, especially those that mark seasonal changes in the environment (Pittendrigh and Minis, 1971; Pittendrigh, 1981; Pittendrigh and Takamura, 1987; Sumova´ et al., 1995; Jagota et al., 2000; Nuesslein- Hildesheim et al., 2000; Sumova´ et al., 2003). Some of these studies were directly aimed at investigating the functional significance of entrainment and its role in photoperiodic induction. For example, in an early study Pittendrigh and Takamura (1987) assayed entrainment and photoperiodic induction in laboratory populations as well as wild populations of Drosophila auraria and reported that a systematic change in day length is paralleled by consistent changes in the clock’s entrained state, and as a consequence the phase relationship between the circadian rhythm and LD cycle (c) changes. This suggests that circadian clocks are involved in time measurement associated with changing seasons. Although it is generally believed that circadian clocks play an important role in the seasonal time measurement, the mechanism(s) by which such information is deciphered in the organism is still unclear. The working hypothesis proposed by Pittendrigh and Daan (1976) posits that circadian clocks comprising of morning (M) and evening (E) oscillators, with distinctly different circadian periods, and responsiveness to light, track day lengths for seasonal ARTICLE IN PRESS www.elsevier.com/locate/jinsphys 0022-1910/$ - see front matter r 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.jinsphys.2006.12.007 Corresponding author. Tel.: +91 80 22082843; fax: +91 80 22082766. E-mail address: [email protected] (V.K. Sharma).
Transcript
ARTICLE IN PRESS
Journal of Insect Physiology 53 (2007) 332–342
0022-1910/$ - se
doi:10.1016/j.jin
�CorrespondE-mail addr
www.elsevier.com/locate/jinsphys
Possible evidence for morning and evening oscillators in Drosophilamelanogaster populations selected for early and late adult emergence
Chronobiology Laboratory, Evolutionary and Organismal Biology Unit, Jawaharlal Nehru Centre for Advanced Scientific Research, P.O. Box 6436, Jakkur,
Bangalore 560064, Karnataka, India
Received 6 November 2006; received in revised form 12 December 2006; accepted 13 December 2006
Abstract
In this paper, we report the results of our study aimed at a systematic analysis of the circadian phenotypes of fruit flies Drosophila
melanogaster selected for early and late adult emergence, in light of the ‘‘morning and evening oscillator’’ (M and E) model for circadian
clocks. We monitored adult emergence and activity/rest rhythms in these flies under light/dark (LD) cycles with short (8:16 h), normal
(12:12 h) and long (16:8 h) photoperiods, as well as under constant darkness (DD). Across all the three LD cycles, the early populations
displayed a morning phenotype with peak of emergence and activity occurring earlier than the controls and greater anticipation to
‘‘lights-on’’ and weak anticipation to ‘‘lights-off’’, while the late populations showed an evening phenotype with peak of emergence and
activity occurring later than the controls and greater anticipation to lights-off and weak anticipation to lights-on. The gate of adult
emergence and duration of activity in the early populations was narrower than the controls, while those of the late populations were
wider than the controls. In addition, the circadian periodicities of adult emergence and activity/rest rhythms of the early flies were
significantly shorter than the controls, while those of the late flies were significantly longer than the controls. In summary, the circadian
phenotypes indicate that the early populations have evolved a dominant M oscillator, while the late populations have evolved a dominant
E oscillator, thus providing an empirical support for the M and E model in Drosophila.
r 2007 Elsevier Ltd. All rights reserved.
Keywords: Drosophila; Eclosion; Activity; Selection; Photoperiod; Morning and Evening oscillators
1. Introduction
Organisms track time in their local environment byentraining their circadian clocks to the light/dark (LD)cycles, a phenomenon that is of paramount importance fortheir survival under fluctuating environments (Pittendrighand Daan, 1976; Daan, 1981). Several studies have beencarried out in the past to understand how organisms adjustto periodic LD cycles, especially those that mark seasonalchanges in the environment (Pittendrigh and Minis, 1971;Pittendrigh, 1981; Pittendrigh and Takamura, 1987;Sumova et al., 1995; Jagota et al., 2000; Nuesslein-Hildesheim et al., 2000; Sumova et al., 2003). Some ofthese studies were directly aimed at investigating thefunctional significance of entrainment and its role in
e front matter r 2007 Elsevier Ltd. All rights reserved.
sphys.2006.12.007
ing author. Tel.: +9180 22082843; fax: +91 80 22082766.
photoperiodic induction. For example, in an early studyPittendrigh and Takamura (1987) assayed entrainment andphotoperiodic induction in laboratory populations as wellas wild populations of Drosophila auraria and reported thata systematic change in day length is paralleled by consistentchanges in the clock’s entrained state, and as a consequencethe phase relationship between the circadian rhythm andLD cycle (c) changes. This suggests that circadian clocksare involved in time measurement associated with changingseasons.Although it is generally believed that circadian clocks
play an important role in the seasonal time measurement,the mechanism(s) by which such information is decipheredin the organism is still unclear. The working hypothesisproposed by Pittendrigh and Daan (1976) posits thatcircadian clocks comprising of morning (M) and evening(E) oscillators, with distinctly different circadian periods,and responsiveness to light, track day lengths for seasonal
ARTICLE IN PRESSS. Kumar et al. / Journal of Insect Physiology 53 (2007) 332–342 333
functions. It also assumes that the morning and eveningactivity peaks exhibited by several organisms are overtmanifestations of the M and E oscillators (Pittendrigh andDaan, 1976; Daan et al., 2001). Notwithstanding theimportance of this model in a wide variety of biologicalfunctions, it has never been rigorously investigated in wild-type animals that display distinct morning and eveningpreference in behaviour.
Much like many crepuscular organisms, the activitypatterns of several insect species have two distinct bouts,one centred around dawn (morning) and the other arounddusk (evening). Such bimodal activity patterns have beenobserved in cockroaches (Roberts, 1962; Wiedenmann1980, 1983), crickets (Wiedenmann 1983; Wiedenmannand Loher, 1984), mosquitoes (Taylor and Jones, 1969;Chiba, 1971; Chiba et al., 1981; Chiba et al., 1982;Clopton, 1984) and silk moths (Truman, 1974). It hasbeen shown that two bilaterally distributed clocks in thebrain regulate bimodality in activity rhythm of cock-roaches (Wiedenmann, 1977, 1980; Page, 1978; Page et al.,1977), song rhythm of crickets (Wiedenmann 1983;Wiedenmann and Loher, 1984) and ERG rhythm ofbeetles (Koehler and Fleissner, 1978). This suggests thatthe circadian timing systems of several insect species arecomposed of two oscillators, the M and E oscillators.Several species of Drosophila too anticipate dawn and duskas evidenced by initialization of activity few hours beforethe actual point of transition between light and dark(Hardeland and Stange, 1973; Helfrich and Engelmann,1983; Hamblen et al., 1986; Helfrich, 1986, Helfrich-Forster, 2000, 2001). Arrhythmic mutants do not displaysuch anticipatory activity, instead they display a startleresponse, immediately after lights-on and lights-off, sug-gesting a driven periodic behaviour (Wheeler et al., 1993).Recently in a series of studies using a geneticallymanipulated fruit fly Drosophila melanogaster, the M andE oscillators were implicated in the regulation of suchanticipatory activities (Grima et al., 2004; Stoleru et al.,2004). Flies with functional ventro-lateral neurons (LNvs)displayed a morning circadian phenotype, while thosewith dorso-lateral neurons (LNds) showed an eveningcircadian phenotype, suggesting that these cells functionas the putative M and E oscillators in the Drosophila
circadian timing system. Furthermore, the LNv (candidateM) and LNd (candidate E) group of neurons respondeddifferently to light (Rieger et al., 2006), thus fulfilling themajor criteria laid out for the M and E oscillators in themodel.
Studies aimed at investigating the dual oscillator modelin Drosophila suggest that the period (per) gene is anessential component of both M as well as E oscillators(Grima et al., 2004; Stoleru et al., 2004). This is, however,contrary to a previously held view that per is necessary forthe E oscillators but not for the M oscillators (Hamblen-Coyle et al., 1992; Helfrich-Forster, 2001), since per0
mutants of D. melanogaster do not show evening activitypeak. Further, it is only the evening activity which is
affected in the pers and perl mutants. How do we reconcilethese widely divergent findings? Such inconsistencies inresults are quite common in studies on mutant lines thatare created for the purpose of obtaining deviant pheno-types, because these flies are typically inbred (Rose, 1984;Mueller and Joshi, 2000). Therefore, the only logical wayto overcome this problem is to create wild-type populationswith morning and evening circadian preference forbehaviour employing laboratory selection schemes andthen to investigate their circadian phenotypes and theunderlying mechanisms.Strains with morning and evening preference for
behaviour have been created in a few early studies byselecting for flies that emerged under 12:12 h LD cyclesduring the ‘‘lights-on’’ (morning) and ‘‘lights-off’’ (eve-ning), respectively (Drosophila pseudoobscura: Pittendrigh,1966; Pectinophora gossypiella: Pittendrigh and Minis,1971; D. melanogaster: Clayton and Paietta, 1972). As aconsequence, the selected strains evolved morning andevening phenotypes with peak of emergence rhythmoccurring about 6 h apart and circadian period (t) differingby about 2.5 h. Although some clock properties, such as thetiming of emergence peak and circadian period weredifferent among the early and late strains, as predicted inthe M and E oscillator model, their light-induced phaseresponse curves (PRC) bore striking similarity (Pittendrigh,1981). However, these results could not be reproduced in aseparate study when a different species of Drosophila
was used (Pittendrigh and Takamura, 1987). In this study,the early and late populations were raised under a differentLD cycle. Though the selected populations show diver-gence in the peak of emergence rhythm, in a manner similarto previous studies, and phase separation between thepeaks were also maintained across a wide range ofphotoperiods, the early strains had a shorter t than thelate strains, and the photoperiodic response of the twostrains was strikingly similar. Although these studiesclearly demonstrate that the early and late strains evolvemorning and evening timing of adult emergence, thecircadian phenotypes of the selected strains do not upholdthe M and E oscillator model. Moreover, it is difficult toformulate a mechanism based on these studies as they lackdetails of starting populations, selection protocols andpopulation size. The fact that most previous studiesincluding those of Pittendrigh did not use replicatepopulations, makes it difficult to rule out the possibilitythat the results were masked by inbreeding, reduction ingenetic variability and random genetic drift (Prasad andJoshi, 2003; David et al., 2005).Here we report our study aimed at investigating the
circadian phenotypes of the early and late emerging flies inlight of the M and E oscillator model. The early andlate populations were raised by selecting for flies thatemerged during the morning and the evening hours under a12:12 h LD cycle. At the 70th generation, we assayed thetime course and waveform of two well-characterizedbehavioural rhythms (adult emergence and activity/rest)
ARTICLE IN PRESSS. Kumar et al. / Journal of Insect Physiology 53 (2007) 332–342334
under LD cycles with different photoperiods, as well as incontinuous darkness (DD). In an earlier study on the samepopulations, we found that at the 55th generation, thepercentage of flies emerging during the morning andevening hours increased in the early and late populationscompared to the controls, suggesting that D. melanogaster
populations respond to selection on the timing of adultemergence by evolving different temporal preference fortheir behaviour (Kumar et al., 2007). The circadian periodand light pulse PRC of the early and late populationsdiffered from each other as well as from the controls. Sinceit is known that extreme photoperiods modulate circadianrhythms, either by magnifying or reducing circadianwaveforms (Sumova et al., 2003), we chose to evaluatethe circadian phenotypes of the selected and controlpopulations under short (LD 8:16 h), normal (LD12:12 h) and long (LD 16:8 h) photoperiods.
2. Materials and methods
2.1. Fly population maintenance and selection protocol
The early and late populations were initiated from fourancestral baseline populations of D. melanogaster that havebeen maintained in the laboratory for about 75 generationsas separate entities without any gene flow between them.The maintenance protocol and ancestry of these popula-tions are described in detail in Sheeba et al. (1998). Briefly,they were maintained as large outbred populationsunder alternating 12:12 h LD cycles (light intensity1575 mW/cm2/s) with banana–jaggery food and wateravailable ad libitum. Temperature (2571 1C) and humidity(7575%) were maintained constant throughout the study.A total of 1200 breeding adults per population, withroughly equal number of males and females were main-tained in Plexiglass cages (25 cm� 20 cm� 15 cm) on a 21-day discrete generation cycle. Eggs were collected byplacing petri dishes with food into these cages during thelight phase of LD cycle and dispensed into vials (18 cmheight� 2.4 cm diameter) with 10ml of food, at a densityof about 300 eggs per vial. Such high density of eggsresulted in staggered adult emergence over several cycles.Flies emerging between the 9th and 13th days werecollected into Plexiglass cages containing a petri dish offood. On the 18th day, a generous smear of yeast–aceticacid paste was applied on the food plates and kept in thecages. Three days later, eggs were collected to initiatethe next generation. These four populations (referred to asthe baseline populations) served as the founder populationsfor the initiation of selection lines. These four baselinepopulations were kept under 12:12 h LD cycles where lightscame on at 08:00 h and went off at 20:00 h. Four early(early1..4) and four late (late1..4) populations were initiatedfrom the four baseline populations by imposing selectionfor adult emergence during the morning (05:00–09:00 h)and evening (17:00–21:00 h) hours (henceforth will bereferred to as the morning and evening windows of
selection). Four control populations (control1..4) were alsoinitiated along with the selected populations, where noconscious selection pressure was applied on the timing ofadult emergence. Each early, control and late populationwas derived from one baseline population, thus forming amatched selected and control triplet (earlyi, controli andlatei are more closely related than earlyj, controlj and latej,i,j ¼ 1–4). For example, the early1, control1 and late1populations were initiated from baseline population 1. Thefour replicate populations with identical subscripts weretreated as random factor in the analysis of variance(ANOVA). The selected and control populations weremaintained under similar conditions, except that in eachgeneration the adult flies for the early populations werecollected between 05:00 and 09:00 h (M: morning window),for the late populations between 170:0 and 21:00 h(E: evening window), and for the control populationsthroughout the day, for 4–5 successive days. Care wastaken to maintain a large outbred structure (n�1200 withroughly equal numbers of males and females) of thepopulations.Imposition of different maintenance regimes may induce
nongenetic parental effects. Therefore all selected andcontrol populations were subjected to one generation ofcommon rearing conditions prior to the assays, duringwhich no conscious selection pressure was imposed. Suchtreatment for one generation is known to eliminatenongenetic parental effects (Prasad et al., 2001). Eggs werecollected from the running cultures and dispensed into vialswith about 10ml of food, at a density of about 300 eggs pervial. After the 12th day of egg collection, adult flies werecollected into Plexiglass cages with abundant food. Theprogeny of these flies will be hereafter referred to asstandardized flies.
2.2. Adult emergence rhythm
For the adult emergence rhythm assay, eggs of approxi-mately the same age were collected from the standardizedpopulations and dispensed into vials with 10ml of food atan egg density of �300 per vial. These vials were introducedinto short (LD 8:16 h), normal (LD 12:12 h) and long (LD16:8 h) photoperiods. Ten such vials were set up for eachreplicate population under each light regime. These vialswere monitored for the first emergence and subsequentlywere checked regularly at every 2 h interval and the numberof emerging adults was recorded. This continued for 10days. From the primary data, we estimated the mean phaserelationship (c) of the primary eclosion peak, width ofeclosion gate (g), eclosion anticipation index (AI) withrespect to lights-on and average waveform of eclosionrhythm under three different LD cycles. The c for theprimary eclosion peak under the three LD cycles wasestimated as the time interval between eclosion peak andlights-on, averaged over 10 consecutive LD cycles. The cvalues were considered to be negative if the eclosion peaksfollowed lights-on and were taken to be positive if they
ARTICLE IN PRESSS. Kumar et al. / Journal of Insect Physiology 53 (2007) 332–342 335
preceded lights-on. The gate of adult emergence (g) wasestimated as the time interval between the beginning andend of adult emergence during one complete cycle, where5% of the total flies emerging in one cycle was taken as thethreshold for defining the beginning and end of emergence.From the raw data, we obtained the AI of eclosion forindividual vials using the formula AI ¼ (b�1�b�2)� (b+1/b�1), where b�1 ¼ bin before lights-on and b+1 ¼ bin afterlights-on, (b�1�b�2) ¼ number of flies emerging in a 2 h binbefore lights-on and (b+1/b�1) denotes the ratio of numberof flies emerging in a 2 h bin immediately after and beforelights-on. This method of AI estimation of adult emergenceis a minor modification of the one proposed by Stoleru et al.(2004), because the bin size for adult emergence rhythmassay in our study was 2 h as opposed to 1 h in the proposedformula. Under DD, the free-running period (t) of eclosionrhythm was estimated (dim red light with l4640nm)by monitoring adult emergence for a minimum of 10consecutive days.
2.3. Activity/rest rhythm
The AI relative to lights-on (AIon) and lights-off (AIoff),duration of activity (a) and waveform of activity/restrhythm were estimated under the three LD cycles. Freshlyemerged adult flies were introduced individually intoactivity monitors within 24 h of their emergence, and theiractivity was monitored for a minimum of 10 consecutivedays (Sharma, 2003). Approximately 60 flies (only males)were used from each population under three different lightregimes. From the raw data we obtained AI ¼ [b�1(b�1�b�2)(b�2�b�3)/b+1], where bi ¼ activity counts inbin i, and i ¼ number of bin before (�i) and after (+i)lights-on or -off, and the anticipation of light transition isdirectly proportional to the gradual increase in activitybefore the transition [(b�1�b�2)(b�2�b�3)] and inverselyproportional to the startle effect (b+1/b�1). The a wascalculated as the average time interval between the onsetand offset of activity in one cycle. The t of the activity/restrhythm was also estimated under DD. The activity/restpattern of an individual fly was plotted and analysed usingCLOCKLAB software (Actimetrics, Evanston, IL).
2.4. Statistical analyses
The t of eclosion rhythms under DD was estimated bysubjecting the time series data collected over 10 consecutivecycles to Fourier spectral analysis using StatisticaTM
(rel.5.0B, Statistica, 1995). Statistical significance ofrhythmic contributions from different frequencies in theperiodogram was tested using Siegel’s modification ofthe Fischer test (Siegel, 1980). This method delineated thefrequency components present in the time series by defininga threshold value (Rao and Sharma, 2002). The t ofactivity/rest rhythms under DD was estimated by subject-ing the time series data collected over 10 consecutive cycles
to Lomb–Scargle periodogram analysis using CLOCK-LAB (Actimetrics, Evanston, IL).The circadian parameters of eclosion and activity/rest
rhythms were subjected to separate mixed model ANOVAtreating replicates as random factor whereas populationand light regime as fixed factors crossed with replicates.The t, AI and g values of emergence rhythm and t, AI anda values of activity/rest rhythm were treated as fixed factorscrossed with population, light regime and replicate. In allstatistical analyses, population means were used as units ofanalysis. Multiple comparisons were done using 95%confidence intervals (95%CI) around the mean.
3. Results
3.1. Adult emergence rhythm
The eclosion rhythm of the early, control and late
populations entrained to short (8:16 h), normal (12:12 h)and long (16:8 h) photoperiods maintained a stable cwithin each light regime (Fig. 1a–c). ANOVA revealed asignificant main effect of population and light regime, butpopulation� light regime interaction did not have asignificant effect on c (Table 1a). Multiple comparisonsusing 95%CI around the mean revealed that across allthree light regimes the primary peak of emergence occurredsignificantly earlier in the early populations followed by thecontrol and late populations, in that order (Fig. 1d).With an increase in the length of the photoperiod, the
width of the emergence gate (g) increased in the selectedand control populations (Fig. 1a–c and e). ANOVAshowed a significant main effect of light regime andpopulation on g, while population� light regime interac-tion did not have a significant effect (Fig. 1e; Table 1b).Multiple comparisons using 95%CI revealed that across allthree light regimes, the mean g was the shortest in the early
populations, followed by the control, and then the late
populations (Fig. 1e).Across all three light regimes, the adult emergence
rhythm of the early, control and late populations clearlyanticipated lights-on (Fig. 1a–c and f), except under thenormal LD cycle where late populations showed weakanticipation (Fig. 1a–c). ANOVA revealed a significantmain effect of population, and population� light regimeinteraction, while the light regime did not have astatistically significant effect (Table 1c). Multiple compar-isons using 95%CI revealed that under normal and longphotoperiods, the AI of the early populations wassignificantly greater than the controls, while that of thecontrols was significantly greater than the late populations(Fig. 1f). Multiple comparisons revealed that across allthree light regimes, the relative difference in the c, g and AIof the early and late populations was greater under longphotoperiod compared to short photoperiod. All statisti-cally significant comparisons are shown in the Fig. 1d–fwith asterisks (�).
ARTICLE IN PRESS
**
**
**
**
*
*
-4
-2
**
**
**
50
00
25
per
centa
ge
of
flie
s
18 00 08 16 18 00 08 16 18 00 08 16
Photo periods
**
**
**
gat
e o
f ec
losi
on
(h
r)
18
12
06
****
****
****
10.0 **
*
* early
control
late
early
control
late
0.0
-5.0
5.0
AI
****
**
**
0
-6
**
**
**
**
**
**
ph
ase-
rela
tio
nsh
ip (
hr)
**
**
**
****
****
****
shortnormal
long
shortnormal
long
shortnormal
long
Zeitgeber Time (hr)
Fig. 1. Average eclosion waveform of the selected and the control populations under short, normal and long photoperiods. Eclosion waveform under (a)
short (LD 8:16 h), (b) normal (LD 12:12 h) and (c) long (LD 16:8 h) photoperiods. Percentage of flies emerging every 2 h, averaged across 10 consecutive
cycles, is plotted along the ordinate, and time of the day in Zeitgeber Time (hours) is plotted along the abscissa. ZT0 is taken as lights-on. Filled and empty
bars represent the duration of dark and light in the LD cycles. (d) Mean phase relationship (c) of the selected and control populations. (e) Mean width of
the eclosion gate (g) (in hours) of the selected and control populations. (f) Mean eclosion anticipation index (AI) of the selected and control populations. A
total of 40 vials were used under each photoperiodic regime, of which 10 were used for each of the four replicate populations for estimation of the c, g and
AI values. Asterisks (*) in panels (d), (e) and (f) denote the significant differences. The error bars represent standard error of the means (SEM).
S. Kumar et al. / Journal of Insect Physiology 53 (2007) 332–342336
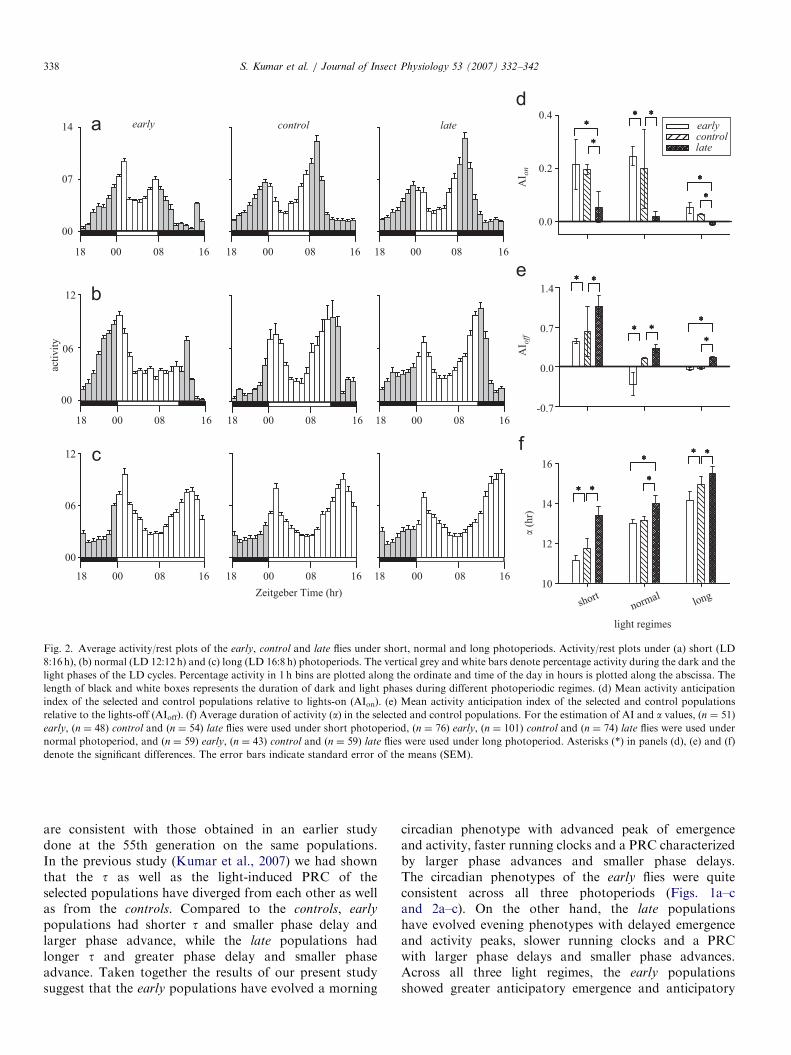
3.2. Activity/rest rhythm
The time course and waveform of the activity/rest rhythmof the selected and control populations closely mimickedthose of the adult emergence rhythm (Fig. 2a–c). ANOVA onthe lights-on (AIon) and lights-off anticipation (AIoff) indicesrevealed a significant main effect of light regime andpopulation, while the interactions were not statisticallysignificant (Table 2a and b). Multiple comparisons using95%CI revealed that across all three light regimes, the AIon of
the early and controls was significantly greater than the late
populations, while those of the early and controls differedsignificantly only under normal photoperiods (Fig. 2d). Onthe other hand, across all three light regimes, the AIoff of theearly and control populations was significantly smaller thanthat of the late populations, while the AIoff of the early
populations was significantly smaller than that of the controls
under short and normal photoperiods (Fig. 2e).Similarly, with increase in the length of photoperiod,
the a increased in the selected and control populations
ARTICLE IN PRESS
Table 1
Results of mixed model analysis of variance (ANOVA) on the (a) phase relationship (c) (b) gate (g) and (c) anticipation index (AI) of adult emergence of
the selected and control populations under three different light regimes
Effect df effect MS effect df error MS error F p-level
Populations (early, late and control) and light regime (short; LD 8:16 h, normal; LD 12:12 h, long; LD 16:8 h) were used as fixed factors, whereas replicates
(1–4) were used as random factor. In all cases, the replicate means were used as the unit of analysis and hence only the fixed factor could be tested for
statistical significance. Statistically significant values (po0.05) are shown in bold.
S. Kumar et al. / Journal of Insect Physiology 53 (2007) 332–342 337
(Fig. 2a–c and f). ANOVA revealed a significant maineffect of light regime and population on the a, whilelight regime� population interaction did not have astatistically significant effect (Table 2c). Multiple compar-isons using 95%CI revealed that across all threephotoperiods, a of the early populations were significantlylower than the controls under short and long photoperiods,while that of the late populations was significantlygreater than the early and controls (Fig. 2f, Table 2c).Multiple comparisons revealed that across all three lightregimes the relative differences in a, AIon and AIoff amongthe early and late populations were greater under longphotoperiod than under short photoperiod. All statisticallysignificant comparisons are shown in Fig. 2d–f withasterisks (�).
3.3. Periodicity of eclosion and activity/rest rhythm under
DD
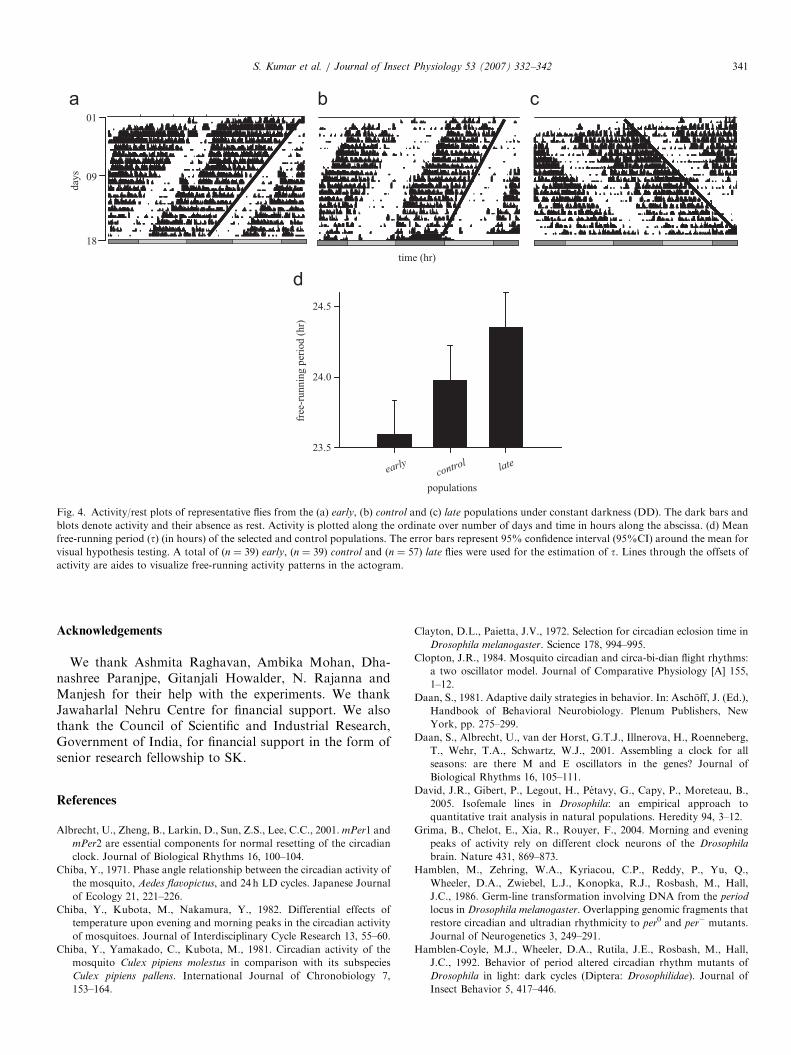
The t of the eclosion rhythm under DD differedsignificantly among the selected and control populations.ANOVA revealed a significant main effect of populationon t (F2,6 ¼ 9.90; po0.013). Multiple comparisonsusing 95%CI showed that t of the early populationswas significantly shorter than the controls, while that of
the late populations was significantly longer than thecontrols (Fig. 3a–d). ANOVA on the t of the activity/rest rhythm revealed a significant main effect of popula-tion (F2,6 ¼ 11.12; po0.009). Multiple comparisonsrevealed that the t of the early populations was signifi-cantly shorter than the controls, while that of thelate populations was significantly longer than the controls
(Fig. 4a–d).
4. Discussion
Selection on the timing of adult emergence resultedin an altered time course and waveform of eclosionand activity/rest rhythms in D. melanogaster populations.After 70 generations of selection, the primary peaks ofemergence in the early populations occurred earlierand were higher compared to the controls, whereas thoseof the late populations occurred later and were smallerthan the controls (Fig. 1a–c). In addition, the early
populations showed greater anticipation to lights-onthan the late and controls, while the late populationsdisplayed greater anticipation to lights-off compared to theearly and controls. The trend in emergence as well asthe activity/rest patterns was more or less similar underall the three light regimes. The results of our present study
ARTICLE IN PRESS
*
*
*
*
* *
**
***
*
**
**
*
*
*
*
*
*
* *
AI on
AI off
0.4
0.0
**
**
**
**
** **
0.2
1.4
0.0
-0.7
0.7
**
***
*
****
******
**
**
**
*
*
� (h
r)
shortnormal
long
10
14
16
latecontrolearly
12
****
****
**
**
Zeitgeber Time (hr)
early14
07
00
00 0818 16 00 0818 16 00 0818 16
00 0818 16 00 0818 16 00 0818 16
00 0818 16 00 0818 16 00 0818 16
control late
acti
vit
y
12
06
00
12
06
00
light regimes
Fig. 2. Average activity/rest plots of the early, control and late flies under short, normal and long photoperiods. Activity/rest plots under (a) short (LD
8:16 h), (b) normal (LD 12:12 h) and (c) long (LD 16:8 h) photoperiods. The vertical grey and white bars denote percentage activity during the dark and the
light phases of the LD cycles. Percentage activity in 1 h bins are plotted along the ordinate and time of the day in hours is plotted along the abscissa. The
length of black and white boxes represents the duration of dark and light phases during different photoperiodic regimes. (d) Mean activity anticipation
index of the selected and control populations relative to lights-on (AIon). (e) Mean activity anticipation index of the selected and control populations
relative to the lights-off (AIoff). (f) Average duration of activity (a) in the selected and control populations. For the estimation of AI and a values, (n ¼ 51)
early, (n ¼ 48) control and (n ¼ 54) late flies were used under short photoperiod, (n ¼ 76) early, (n ¼ 101) control and (n ¼ 74) late flies were used under
normal photoperiod, and (n ¼ 59) early, (n ¼ 43) control and (n ¼ 59) late flies were used under long photoperiod. Asterisks (*) in panels (d), (e) and (f)
denote the significant differences. The error bars indicate standard error of the means (SEM).
S. Kumar et al. / Journal of Insect Physiology 53 (2007) 332–342338
are consistent with those obtained in an earlier studydone at the 55th generation on the same populations.In the previous study (Kumar et al., 2007) we had shownthat the t as well as the light-induced PRC of theselected populations have diverged from each other as wellas from the controls. Compared to the controls, early
populations had shorter t and smaller phase delay andlarger phase advance, while the late populations hadlonger t and greater phase delay and smaller phaseadvance. Taken together the results of our present studysuggest that the early populations have evolved a morning
circadian phenotype with advanced peak of emergenceand activity, faster running clocks and a PRC characterizedby larger phase advances and smaller phase delays.The circadian phenotypes of the early flies were quiteconsistent across all three photoperiods (Figs. 1a–cand 2a–c). On the other hand, the late populationshave evolved evening phenotypes with delayed emergenceand activity peaks, slower running clocks and a PRCwith larger phase delays and smaller phase advances.Across all three light regimes, the early populationsshowed greater anticipatory emergence and anticipatory
ARTICLE IN PRESS
Table 2
Results of mixed model analysis of variance (ANOVA) on the (a) lights-on anticipation (AIon) (b) lights-off anticipation (AIoff) and (c) activity duration
(a) of activity/rest pattern of the selected and control populations under three different light regimes
Effect df effect MS effect df error MS error F p-level
Populations (early, late and control) and light regime (short; LD 8:16 h, normal; LD 12:12 h, long; LD 16:8 h) were used as fixed factors, whereas replicates
(1–4) were used as random factor. In all cases, the replicate means were used as the unit of analysis and hence only the fixed factor could be tested for
statistical significance. Statistically significant values (po0.05) are shown in bold.
S. Kumar et al. / Journal of Insect Physiology 53 (2007) 332–342 339
activity to lights-on and weak or no anticipation to lights-off, while the late populations show greater anticipationto lights-off and weak or no anticipation to lights-on(Figs. 1a–c and 2a–c). These results thus suggest thatthe early and late populations have evolved dominant Mand E oscillators with morning and evening peak ofemergence and activity, faster and slower runningclocks, and differential light responses. In addition, aconsistent smaller width of eclosion (g) and a in theearly and longer g and a in the late populations, observedacross all three photoperiodic regimes, suggests thatthe coupling strengths between the M and E oscillatorsin these populations are asymmetric. Such asymmetryin coupling may be critical in maintaining stable androbust morning and evening circadian phenotypes. Thefact that the relative differences among the early and late
populations were augmented under long photoperiodsuggests that long photoperiods draw the M and Eoscillators apart, whereas shorter photoperiod brings themcloser. Such effects of photoperiods on the couplingstrength between the M and E oscillators were predictedin a recent extension of the dual oscillator model (Daanet al., 2001).
In the extended M and E oscillator model (Daan et al.,2001) it was suggested that the mammalian circadian
system comprises of M and E oscillators in the form ofperiod/cryptochrome (per/cry) gene pairs; the per1/cry1 pairis assumed to regulate the morning activity whereas theper2/cry2 pair is considered to regulate the eveningactivity. The waveform of the adult emergence andactivity/rest patterns in the selected and control popula-tions in our study was left skewed in the early populationsand marginally right skewed in the late populations(Figs. 1a–c and 2a–c). This is similar to the findings of astudy on the Dmper1 (with putative E oscillator) andDmper2 (with putative M oscillator) mice (knockoutsfor M and E oscillator respectively), where mper2 andmper1 gene expressions were reported to be right andleft skewed under long and short photoperiods (Steinlech-ner et al., 2002). Furthermore, consistent with thelight responses in the early and late flies, animals runningsolely on the E oscillator (Dmper1) show only phasedelays, while those running solely on the M oscillator(Dmper2) display only phase advances (Albrecht et al.,2001).Based on previous studies, it was believed that the PDF-
positive small LNv cells regulate the morning activity andthe PDF-negative LNds regulate the evening activity(Grima et al., 2004; Stoleru et al., 2004). SubsequentlyRieger et al. (2006) demonstrated that the PDF-positive
ARTICLE IN PRESS
00
10
20
num
ber
of f
iles
time (hr)
early
control lat
e
populations
free
-run
ning
per
iod
(hr)
23.0
24.0
25.0
Fig. 3. Adult emergence patterns of representative vials from the (a) early, (b) control and (c) late populations under constant darkness (DD). The dark
and light grey bars denote subjective night and day, respectively. The percentage of flies emerging every 2 h is plotted along the ordinate and time in hours
along the abscissa. (d) Mean free-running period (t) (in hours) of eclosion rhythm of the selected and control populations. A total of 40 vials were used, of
which 10 were used for each replicate population. The error bars indicate 95% confidence interval (95%CI) around the mean for visual hypothesis
testing.
S. Kumar et al. / Journal of Insect Physiology 53 (2007) 332–342340
s-LNv cells control both the morning and eveningactivity bouts. In Drosophila, often the evening activitypeak is most prominent and it persists under DD, whilethe morning peak is reduced (Wheeler et al., 1993;Helfrich-Forster, 2000), which further suggests that mainlythe evening component constitutes the free-runningrhythm, and therefore it is not a big surprise that theneurons responsible for the free-running rhythm have someimpact on the evening component. Indeed, Veleri et al.(2003) have also demonstrated that the s-LNv show robustcycling in extended DD. This is consistent with the findingsof Stoleru et al. (2005), which suggests that the s-LNv cellsare necessary for the timing of activity peaks in DD(Stoleru et al., 2005). Therefore, in addition to theirdominance over other neuronal subgroups, the PDF-positive s-LNv cells seem to be crucial for the regulationof the normal evening activity under LD cycles (Grima etal., 2004; Stoleru et al., 2004).
Taken together our study supports the view thatcircadian clocks of Drosophila are composed of at least
two sets of oscillators. As proposed by Pittendrigh andDaan (1976), the two oscillators show a morning andevening circadian phenotype, shorter and longer circadianperiods and differential sensitivity to dawn and dusk. Suchmorning and evening circadian phenotypes could stemfrom (i) differential control of clock neurons (as suggestedby Grima et al., 2004; Stoleru et al., 2004; Rieger et al.,2006), (ii) differences in the genetic architecture ofcircadian clocks, (iii) altered temporal expression of clockand clock controlled genes, or (iv) a combination of (i) and(ii). Although our study provides compelling empiricalevidence for the M and E oscillator model of circadianclocks in D. melanogaster, further experiments should becarried out to test (a) if the M and E oscillators employdifferent pacemaking mechanisms, involving different setof core clock genes, as suggested in the case of mammals(Daan et al., 2001) or (b) whether the M and E oscillatorsare different sets of clock neurons as suggested in a fewrecent studies in Drosophila (Grima et al., 2004; Stoleruet al., 2004).
ARTICLE IN PRESS
time (hr)
early
control late
populations
free
-runnin
g p
erio
d (
hr)
23.5
24.0
24.5
day
s
01
09
18
Fig. 4. Activity/rest plots of representative flies from the (a) early, (b) control and (c) late populations under constant darkness (DD). The dark bars and
blots denote activity and their absence as rest. Activity is plotted along the ordinate over number of days and time in hours along the abscissa. (d) Mean
free-running period (t) (in hours) of the selected and control populations. The error bars represent 95% confidence interval (95%CI) around the mean for
visual hypothesis testing. A total of (n ¼ 39) early, (n ¼ 39) control and (n ¼ 57) late flies were used for the estimation of t. Lines through the offsets of
activity are aides to visualize free-running activity patterns in the actogram.
S. Kumar et al. / Journal of Insect Physiology 53 (2007) 332–342 341
Acknowledgements
We thank Ashmita Raghavan, Ambika Mohan, Dha-nashree Paranjpe, Gitanjali Howalder, N. Rajanna andManjesh for their help with the experiments. We thankJawaharlal Nehru Centre for financial support. We alsothank the Council of Scientific and Industrial Research,Government of India, for financial support in the form ofsenior research fellowship to SK.
References
Albrecht, U., Zheng, B., Larkin, D., Sun, Z.S., Lee, C.C., 2001. mPer1 and
mPer2 are essential components for normal resetting of the circadian
clock. Journal of Biological Rhythms 16, 100–104.
Chiba, Y., 1971. Phase angle relationship between the circadian activity of
the mosquito, Aedes flavopictus, and 24 h LD cycles. Japanese Journal
of Ecology 21, 221–226.
Chiba, Y., Kubota, M., Nakamura, Y., 1982. Differential effects of
temperature upon evening and morning peaks in the circadian activity
of mosquitoes. Journal of Interdisciplinary Cycle Research 13, 55–60.
Chiba, Y., Yamakado, C., Kubota, M., 1981. Circadian activity of the
mosquito Culex pipiens molestus in comparison with its subspecies
Culex pipiens pallens. International Journal of Chronobiology 7,
153–164.
Clayton, D.L., Paietta, J.V., 1972. Selection for circadian eclosion time in
Drosophila melanogaster. Science 178, 994–995.
Clopton, J.R., 1984. Mosquito circadian and circa-bi-dian flight rhythms:
a two oscillator model. Journal of Comparative Physiology [A] 155,
1–12.
Daan, S., 1981. Adaptive daily strategies in behavior. In: Aschoff, J. (Ed.),
Handbook of Behavioral Neurobiology. Plenum Publishers, New
York, pp. 275–299.
Daan, S., Albrecht, U., van der Horst, G.T.J., Illnerova, H., Roenneberg,
T., Wehr, T.A., Schwartz, W.J., 2001. Assembling a clock for all
seasons: are there M and E oscillators in the genes? Journal of