Potential impacts of climate change on interactions among saprotrophic cord-forming fungal mycelia and grazing soil invertebrates A. Donald A’BEAR*, T. Hefin JONES, Lynne BODDY Cardiff School of Biosciences, Cardiff University, Sir Martin Evans Building, Museum Avenue, Cardiff CF10 3AX, UK article info Article history: Received 10 September 2012 Revision received 31 January 2013 Accepted 31 January 2013 Available online - Corresponding editor: Erik Hobbie Keywords: Basidiomycete fungi CO 2 feedback Decomposition Global warming Meta-analysis Nutrient cycling Soil fauna abstract Climate change has the potential to alter the activity of, and interactions among, sapro- trophic fungi and soil invertebrate grazers, with implications for decomposer community composition, ecosystem regulation and carbon feedback. We reviewed the impacts of experimentally manipulated temperature, CO 2 concentration and soil moisture content on saprotrophic cord-forming basidiomycete growth and function, and on the abundance of soil micro-invertebrates (nematodes) and meso-invertebrates (collembola, mites and enchytraeids). In warmer and wetter conditions, mycelial growth and mycophagous invertebrate abundance are likely to increase. Grazers may either consume the extra mycelial biomass or amplify the temperature effect by stimulating fungal growth. Grazing can stimulate or inhibit decomposition of colonised woody resources and extracellular enzyme production. Future empirical study should partition saprotrophic fungi from the general microbial biomass, with particular attention focussed on enzyme activity and decomposition. Understanding how biotic and abiotic factors interact to regulate sapro- trophic fungal activity is crucial to strengthen our predictive capacity regarding decom- position and carbon feedback under climate change. ª 2013 Elsevier Ltd and The British Mycological Society. All rights reserved. Introduction Organic carbon decomposition by soil biota generates an annual global release of 60 Pg (10 15 g) C to the atmosphere (almost 10 times that of fossil fuel emissions); this is balanced by the approximately equal quantity absorbed through pri- mary production (Lal 2008). Shifts in this balance, mainly due to changes in ambient climate, have potentially far-reaching implications for CO 2 feedback and atmospheric gaseous composition. By 2100, atmospheric CO 2 concentration is pre- dicted to reach 540e970 ppm, accompanied by globally dif- ferential temperature increases in the range 1.1e6.4 C(IPCC 2007). Precipitation and the frequency of extreme events are also expected to increase, with less certainty regarding magnitude. Although key drivers of ecosystem processes (Wardle et al. 1998; Bradford et al. 2002), soil microbes and invertebrates are not explicitly considered in models predict- ing impacts of climate change on CO 2 feedbacks via, for example, decomposition and soil respiration (Cao & Woodward 1998; Cox et al. 2000; Wall et al. 2008). This is pri- marily a consequence of belowground food web complexity, the general neglect of the significance of soil interactions in climate feedback predictions, and the dearth of understanding of the direct and indirect effects of climate change in soil (Bardgett et al. 2008). Impacts of projected climatic scenarios on aboveground communities and trophic interactions (e.g. plants and their insect herbivores) have been well-studied and thoroughly * Corresponding author: Tel.: þ44 029 2087 5729. E-mail address: [email protected](A.D. A’Bear). available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/funeco 1754-5048/$ e see front matter ª 2013 Elsevier Ltd and The British Mycological Society. All rights reserved. http://dx.doi.org/10.1016/j.funeco.2013.01.009 fungal ecology xxx (2013) 1 e10 Please cite this article in press as: A’Bear AD, et al., Potential impacts of climate change on interactions among saprotrophic cord-forming fungal mycelia and grazing soil invertebrates, Fungal Ecology (2013), http://dx.doi.org/10.1016/ j.funeco.2013.01.009
increases in mycelial growth could, to some extent, be
counteracted by grazing, there is potential for increased
production and release of extracellular enzymes into the soil
environment, and accelerated primary decomposition of
colonised resources. To improve our mechanistic under-
standing of climate change impacts on decomposition, sap-
rotrophic fungal activity needs to be partitioned from that of
the general microbial biomass in empirical investigations.
The majority of studies published to date have not done this,
making functional implications of the overall trends difficult
to identify. Future research should consider interactive
effects of climate change factors on soil biota, particularly
given the influence of elevated temperature and CO2 on soil
moisture properties. Understanding how biotic and abiotic
factors interact tomediate saprotrophic fungal functioning is
crucial in enhancing our predictive capabilities regarding
primary decomposition and carbon feedback in a changing
climate.
pacts of climate change on interactions among saprotrophicrates, Fungal Ecology (2013), http://dx.doi.org/10.1016/
8 A.D. A’Bear et al.
Acknowledgements
Two anonymous reviewers made thoughtful suggestions to
improve the manuscript. Natural Environment Research
Council for provision of a studentship to ADA (NERC/I527861).
Supplementary data
Supplementary data related to this article can be found at
http://dx.doi.org/10.1016/j.funeco.2013.01.009.
r e f e r e n c e s
A’Bear AD, Boddy L, Raspotnig G, Jones TH, 2010. Non-trophic effects of oribatid mites on cord-formingbasidiomycetes in soil microcosms. Ecological Entomology 35:477e484.
A’Bear AD, Boddy L, Jones TH, 2012. Impacts of elevatedtemperature on the growth and functioning of decomposerfungi are influenced by grazing collembola. Global ChangeBiology 18: 1823e1832.
A’Bear AD, Crowther TW, Ashfield R, Chadwick DDA, Dempsey J,Meletiou L, Rees CL, Jones TH, Boddy L. Localised invertebrategrazing moderates the effect of warming on competitivefungal interactions. Fungal Ecology, in press.
Bale JS, Masters GJ, Hodkinson ID, Awmack C, Bezemer TM,Brown VK, Butterfield J, Buse A, Coulson JC, Farrar J, Good JEG,Harrington R, Hartley SE, Jones TH, Lindroth RL, Press MC,Symrnioudis I, Watt AD,Whittaker JB, 2002. Herbivory in globalclimate change research: direct effects of rising temperatureon insect herbivores. Global Change Biology 8: 1e16.
Bardgett RD, Kandeler E, Tscherko D, Hobbs PJ, Bezemer TM,Jones TH, Thompson LJ, 1999. Below-ground microbialcommunity development in a high temperature world. Oikos85: 193e203.
Bardgett RD, Freeman C, Ostle NJ, 2008. Microbial contributions toclimate change through carbon cycle feedbacks. The ISMEJournal 2: 805e814.
Bezemer TM, Jones TH, 1998. Plant-insect herbivore interactionsin elevated atmospheric CO2: quantitative analyses and guildeffects. Oikos 82: 212e222.
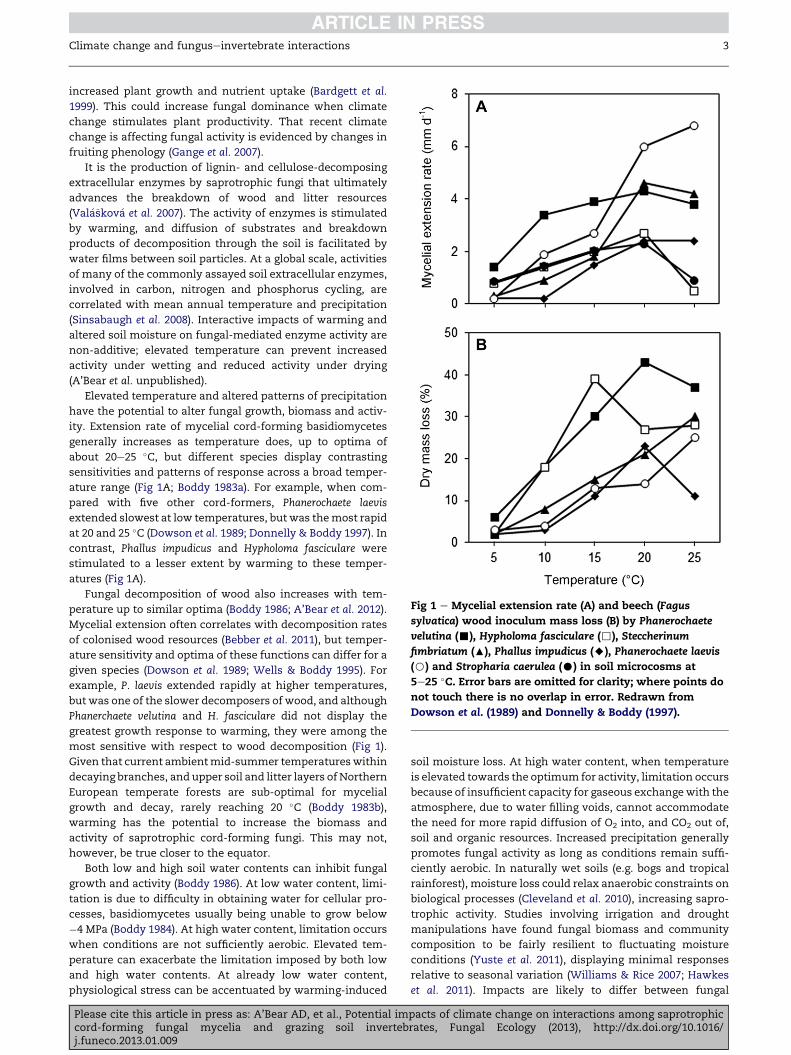
Boddy L, 1983a. The effect of temperature and water potential onthe growth rate of wood-rotting basidiomycetes. Transactionsof the British Mycological Society 80: 141e149.
Boddy L, 1983b. Microclimate and moisture dynamics of wooddecomposing in terrestrial ecosystems. Soil Biology &Biochemistry 15: 149e157.
Boddy L, 1984. The micro-environment of basidiomycete myceliain temperate deciduous woodlands. In: Jennings DH,Rayner ADM (eds), Ecology and Physiology of the Fungal Mycelium.Cambridge University Press, Cambridge, pp. 261e289.
Boddy L, 1986. Water and decomposition processes in terrestrialecosystems. In: Ayres PG, Boddy L (eds), Water, Plants andFungi. Cambridge University Press, Cambridge, pp. 375e398.
Boddy L, 1993. Saprotrophic cord-forming fungi: welfare strategiesand other ecological aspects. Mycological Research 97: 641e655.
Boddy L, Jones TH, 2008. Interactions between basidiomycota andinvertebrates. In: Boddy L, Frankland JC, van West P (eds),Ecology of Saprotrophic Basidiomycetes. Elsevier Ltd., London, pp.155e179.
Boddy L, Gibbon OM, Grungy MA, 1985. Ecology of Daldiniaconcentrica: effect of abiotic variables on mycelial extensionand mycelial interactions. Transactions of the British MycologicalSociety 85: 201e211.
Bradford MA, Jones TH, Bardgett RD, Black HIJ, Boag B,Bonkowski M, Cook R, Eggers T, Gange AC, Grayston SJ,Kandeler E, McCaig AE, Newington JE, Prosser JI, Setala H,Staddon PL, Tordoff GM, Tscherko D, Lawton JH, 2002. Impactsof soil faunal community composition on model grasslandecosystems. Science 298: 615e618.
Bretherton S, Tordoff GM, Jones TH, Boddy L, 2006. Compensatorygrowth of Phanerochaete velutina mycelial systems grazed byFolsomia candida (Collembola). FEMS Microbiology Ecology 58:33e40.
Briones MJI, Ineson P, Piearce TG, 1997. Effects of climate changeon soil fauna; responses of enchytraeids, Diptera larvae andtardigrades in a transplant experiment. Applied Soil Ecology 6:117e134.
Briones MJI, Poskitt J, Ostle N, 2004. Influence of warming andenchytraeid activities on soil CO2 and CH4 fluxes. Soil Biology &Biochemistry 36: 1851e1859.
Cairney JWG, 1992. Translocation of solutes in ectomycorrhizaland saprotrophic rhizomorphs. Mycological Research 96:135e141.
Cao MK, Woodward FI, 1998. Dynamic responses of terrestrialecosystem carbon cycling to global climate change. Nature393: 249e252.
Carrera N, Barreal ME, Gallego PP, Briones MJI, 2009. Soilinvertebrates control peatland C fluxes in response towarming. Functional Ecology 23: 637e648.
Chapela IH, Boddy L, Rayner ADM, 1988. Structure anddevelopment of fungal communities in beech logs four and ahalf years after felling. FEMS Microbiology Letters 53: 59e70.
Cleveland CC, Wieder WR, Reed SC, Townsend AR, 2010.Experimental drought in a tropical rainforest increases soilcarbon dioxide losses to the atmosphere. Ecology 91:2313e2323.
Convey P, Pugh PJA, Jackson C, Murray AW, Ruhland CT, Xiong FS,Day TA, 2002. Response of Antarctic terrestrialmicroarthropods to long-term climate manipulations. Ecology83: 3130e3140.
Cotrufo MF, Ineson P, Rowland AP, 1994. Decomposition of treeleaf litters grown under elevated CO2: effect of litter quality.Plant and Soil 163: 121e130.
Cotrufo MF, Briones MJI, Ineson P, 1998. Elevated CO2 reduces thenitrogen content of plant tissues. Soil Biology & Biochemistry 30:1565e1571.
Couteaux M, Kurz C, Bottner P, Raschi A, 1999. Influence ofincreased atmospheric CO2 concentration on quality of plantmaterial and litter decomposition. Tree Physiology 19: 301e311.
Cox PM, Betts RA, Jones CD, Spall SA, Totterdell IJ, 2000.Acceleration of global warming due to carbon-cycle feedbacksin a coupled climate model. Nature 408: 184e187.
Crowther TW, A’Bear AD, 2012. Species-specific impacts of soilfauna on decomposer fungi are not masked by density-dependence. Fungal Ecology 5: 277e281.
Crowther TW, Boddy L, Jones TH, 2011a. Species-specific effectsof soil fauna on fungal foraging and decomposition. Oecologia167: 535e545.
Crowther TW, Boddy L, Jones TH, 2011b. Outcomes of fungalinteractions are determined by soil invertebrate grazers.Ecology Letters 14: 1134e1142.
pacts of climate change on interactions among saprotrophicrates, Fungal Ecology (2013), http://dx.doi.org/10.1016/
Climate change and funguseinvertebrate interactions 9
Crowther TW, Jones TH, Boddy L, 2011c. Species-specific effects ofgrazing invertebrates on mycelial emergence and growth fromwoody resources into soil. Fungal Ecology 4: 333e341.
Crowther TW, Jones TH, Boddy L, Baldrian P, 2011d. Invertebrategrazing determines enzyme production by basidiomycetefungi. Soil Biology & Biochemistry 43: 2060e2068.
Crowther TW, Boddy L, Jones TH, 2012a. Functional and ecologicalconsequences of saprotrophic fungusegrazer interactions. TheISME Journal 6: 1992e2001.
Crowther TW, Littleboy A, Jones TH, Boddy L, 2012b. Interactiveeffects of warming and invertebrate grazing determine theoutcomes of competitive fungal interactions. FEMSMicrobiology Ecology 81: 419e426.
David JF, Gillon D, 2009. Combined effects of elevatedtemperatures and reduced leaf litter quality on the life-historyparameters of a saprophagous macroarthropod. Global ChangeBiology 15: 156e165.
David JF, Handa IT, 2010. The ecology of saprophagousmacroarthropods (millipedes, woodlice) in the context ofglobal change. Biological Reviews 85: 881e895.
Day TA, Ruhland CT, Strauss SL, Park JH, Krieg ML, Krna MA,Bryant DM, 2009. Response of plants and the dominantmicroarthropod, Cryptopygus antarcticus, to warming andcontrasting precipitation regimes in Antarctic tundra. GlobalChange Biology 15: 1640e1651.
De Deyn GB, Raaijmakers CE, Zoomer HR, Berg MP, de Ruiter PC,Verhoef HA, Bezemer TM, van der Putten WH, 2003. Soilinvertebrate fauna enhances grassland succession anddiversity. Nature 422: 711e713.
Dollery R, Hodkinson ID, Jonsdottir IS, 2006. Impact of warmingand timing of snow melt on soil microarthropod assemblagesassociated with Dryas-dominated plant communities onSvalbard. Ecography 29: 111e119.
Donnelly DP, Boddy L, 1997. Development of mycelial systems ofStropharia caerulea and Phanerochaete velutina on soil: effect oftemperature and water potential. Mycological Research 101:705e713.
Dowson CG, Rayner ADM, Boddy L, 1988a. Inoculation of mycelialcord-forming basidiomycetes into woodland soil and litter. II.Resource capture and persistence.New Phytologist 109: 343e349.
Dowson CG, Rayner ADM, Boddy L, 1988b. The form and outcomeof mycelial interactions involving cord-forming decomposerbasidiomycetes in homogeneous and heterogeneousenvironments. New Phytologist 109: 423e432.
Dowson CG, Boddy L, Rayner ADM, 1989. Development andextension of mycelial cords in soil at different temperaturesand moisture contents. Mycological Research 92: 383e391.
Dyer HC, Boddy L, Preston-Meek CM, 1992. Effect of the nematodePanagrellus redivivus on growth and enzyme production byPhanerochaete velutina and Stereum hirsutum. MycologicalResearch 96: 1019e1028.
Field CB, Jackson RB, Mooney HA, 1995. Stomatal responses toincreased CO2 e implications from the plant to the global-scale. Plant, Cell and Environment 18: 1214e1225.
Gange AC, Gange EG, Sparks TH, Boddy L, 2007. Rapid and recentchanges in fungal fruiting patterns. Science 316: 71.
Griffith GS, Boddy L, 1991. Fungal decomposition of attachedangiosperm twigs. IV. Effect of water potential on interactionsbetween fungi onagarand inwood.NewPhytologist117: 633e641.
Harsch MA, Hulme PE, McGlone MS, Duncan RP, 2009. Aretreelines advancing? A global meta-analysis of treelineresponse to climate warming. Ecology Letters 12: 1040e1049.
Please cite this article in press as: A’Bear AD, et al., Potential imcord-forming fungal mycelia and grazing soil invertebj.funeco.2013.01.009
Hawkes CV, Kivlin SE, Rocca JD, Huguet V, Thomsen MA,Suttle KB, 2011. Fungal community responses to precipitation.Global Change Biology 17: 1637e1645.
Hedges LV, Gurevitch J, Curtis PS, 1999. The meta-analysis ofresponse ratios in experimental ecology. Ecology 80:1150e1156.
Hedlund K, Augustsson A, 1995. Effects of enchytraeid grazing onfungal growth and respiration. Soil Biology & Biochemistry 27:905e909.
Hedlund K, Boddy L, Preston-Meek CM, 1991. Mycelial responsesof the soil fungus, Mortierella isabellina, to grazing byOnychiurus armatus (Collembola). Soil Biology & Biochemistry 23:361e366.
Heimann M, Reichstein M, 2008. Terrestrial ecosystem carbondynamics and climate feedbacks. Nature 451: 289e292.
Hooper DU, Adair EC, Cardinale BJ, Byrnes JEK, Hungate BA,Matulich KL, Gonzalez A, Duffy JE, Gamfeldt L, O’Connor MI,2012. A global synthesis reveals biodiversity loss as a majordriver of ecosystem change. Nature 486: 105e108.
IPCC, 2007. Climate change 2007: synthesis report. In: CoreWriting Team, Pachauri RK, Reisinger A (eds), Contribution ofWorking Groups I, II and III to the Fourth Assessment Report of theIntergovernmental Panel on Climate Change. IPCC, Geneva,Switzerland, p. 104.
Jones TH, Thompson LJ, Lawton JH, Bezemer TM, Bardgett RD,Blackburn TM, Bruce KD, Cannon PF, Hall GS, Hartley SE,Howson G, Jones CG, Kampichler C, Kandeler E, Ritchie DA,1998. Impacts of rising atmospheric carbon dioxide on modelterrestrial ecosystems. Science 280: 441e443.
Kampichler C, Rolschewski J, Donnelly DP, Boddy L, 2004.Collembolan grazing affects the growth strategy of the cord-forming fungus Hypholoma fasciculare. Soil Biology &Biochemistry 36: 591e599.
Kandeler E, Mosier AR, Morgan JA, Milchunas DG, King JY,Rudolph S, Tscherko D, 2008. Transient elevation of carbondioxide modifies the microbial community composition in asemi-arid grassland. Soil Biology & Biochemistry 40: 162e171.
King JS, Pregitzer KS, Zak DR, Kubiske ME, Ashby JA, Holmes WE,2001. Chemistry and decomposition of litter from Populustremuloides Michaux grown at elevated atmospheric CO2 andvarying N availability. Global Change Biology 7: 65e74.
Klironomos JN, Widden P, Deslandes I, 1992. Feeding preferencesof the collembolan Folsomia candida in relation to microfaunalsuccessions on decaying litter. Soil Biology & Biochemistry 24:685e692.
Klironomos JN, Rillig MC, Allen MF, Zak DR, Kubiske M,Pregitzer KS, 1997. Soil fungal-arthropod responses to Populustremuloides grown under enriched atmospheric CO2 under fieldconditions. Global Change Biology 3: 473e478.
Lal R, 2008. Carbon sequestration. Philosophical Transactions of theRoyal Society B 363: 815e830.
Lamborg MR, Hardy RWF, Paul EA, 1983. Microbial effects. In:Lemon ER (ed), CO2 and Plants: The Response of Plants to RisingLevels of Atmospheric Carbon Dioxide. Westview Press, Boulder,pp. 131e176.
Lindberg N, Bengtsson J, Persson T, 2002. Effects of experimentalirrigation and drought on the composition and diversity of soilfauna ina coniferous stand. Journal ofApplied Ecology39: 924e936.
Lindroth RL, Arteel GE, Kinney KK, 1995. Responses of threesaturniid species to paper birch grown under enriched CO2
atmospheres. Functional Ecology 9: 306e311.Loranger GI, Pregitzer KS, King JS, 2004. Elevated CO2 and O3t
concentrations differentially affect selected groups of the
pacts of climate change on interactions among saprotrophicrates, Fungal Ecology (2013), http://dx.doi.org/10.1016/
10 A.D. A’Bear et al.
fauna in temperate forest soils. Soil Biology & Biochemistry 36:1521e1524.
Maraldo K, Schmidt IK, Beier C, Holmstrup M, 2008. Can fieldpopulations of the enchytraeid, Cognettia sphagnetorum, adaptto increased drought stress? Soil Biology & Biochemistry 40:1765e1771.
Meehan TD, Crossley MS, Lindroth RL, 2010. Impacts of elevatedCO2 and O3 on aspen leaf litter chemistry and earthworm andspringtail productivity. Soil Biology & Biochemistry 42:1132e1137.
Newell K, 1984a. Interaction between two decomposerbasidiomycetes and a collembolan under Sitka spruce:distribution, abundance and selective grazing. Soil Biology &Biochemistry 16: 227e233.
Newell K, 1984b. Interaction between two decomposerbasidiomycetes and a collembolan under Sitka spruce: grazingand its potential effects on fungal distribution and litterdecomposition. Soil Biology & Biochemistry 16: 235e239.
Olsrud M, Carlsson BA, Svensson BM, Michelsen A, Melillo JM,2010. Responses of fungal root colonization, plant cover andleaf nutrients to long-term exposure to elevated atmosphericCO2 and warming in a subarctic birch forest understory. GlobalChange Biology 16: 1820e1829.
Rosenberg MS, Adams DC, Gurevitch J, 2000. MetaWin Version 2.1:Statistical Software for Meta-analysis. Sinauer, Boston.
Sadowsky MJ, Schortemeyer M, 1997. Soil microbial responses toincreased concentrations of atmospheric CO2. Global ChangeBiology 3: 217e224.
Schoeman MW, Webber JF, Dickinson DJ, 1996. The effect ofdiffusible metabolites of Trichiderma harzianum on in vitrointeractions between basidiomycete isolates at two differenttemperature regimes. Mycological Research 100: 1454e1458.
Sinsabaugh RL, Lauber CL, Weintraub MN, Ahmed B, Allison SD,Crenshaw C, Contosta AR, Cusack D, Frey S, Gallo ME,Gartner TB, Hobbie SE, Holland K, Keeler BL, Powers JS,Stursova M, Takacs-Vesbach C, Waldrop MP, Wallenstein MD,Zak DR, Zeglin LH, 2008. Stoichiometry of soil enzyme activityat global scale. Ecology Letters 11: 1252e1264.
Tordoff GM, Boddy L, Jones TH, 2008. Species-specific impacts ofcollembola grazing on fungal foraging ecology. Soil Biology &Biochemistry 40: 434e442.
Val�a�skov�a V, �Snajdr J, Bittner B, Cajthaml T, Merhautov�a V,Hofrichter M, Baldrian P, 2007. Production of lignocelluloses-degrading enzymes and degradation of leaf litter bysaprotrophic basidiomycetes isolated from a Quercus petraeaforest. Soil Biology & Biochemistry 39: 2651e2660.
van der Heijden MGA, Bardgett RD, van Straalen NM, 2008. Theunseen majority: soil microbes as drivers of plant diversity
Please cite this article in press as: A’Bear AD, et al., Potential imcord-forming fungal mycelia and grazing soil invertebj.funeco.2013.01.009
and productivity in terrestrial ecosystems. Ecology Letters 11:296e310.
Wall DH, Bradford MA, St John MG, Trofymow JA, Behan-Pelletier V, Bignell DDE, Dangerfield JM, Parton WJ, Rusek J,Voigt W, Wolters V, Gardel HZ, Ayuke FO, Bashford R,Beljakova OI, Bohlen PJ, Brauman A, Flemming S, Henschel JR,Johnson DL, Jones TH, Kovarova M, Kranabetter JM, Kutny L,Lin KC, Maryati M, Masse D, Pokarzhevskii A, Rahman H,Sabara MG, Salamon JA, Swift MJ, Varela A, Vasconcelos HL,White D, Zou XM, 2008. Global decomposition experimentshows soil animal impacts on decomposition are climate-dependent. Global Change Biology 14: 2661e2677.
Wardle DA, Verhoef HA, Clarholm M, 1998. Trophic relationshipsin the soil microfood-web: predicting the responses to achanging global environment. Global Change Biology 4: 713e727.
Wardle DA, Bardgett RD, Klironomos JN, Setala H, van derPutten WH, Wall DH, 2004. Ecological linkages betweenaboveground and belowground biota. Science 304: 1629e1633.
Wells JM, Boddy L, 1995. Effect of temperature on wood decay andtranslocation of soil-derived phosphorus in mycelial cordsystems. New Phytologist 129: 289e297.
Wells JM, Boddy L, 2002. Interspecific carbon exchange and cost ofinteractions between basidiomycete mycelia in soil and wood.Functional Ecology 16: 153e161.
Wells JM, Thomas J, Boddy L, 2001. Soil water potential shifts:developmental responses and dependence on phosphorustranslocation by the saprotrophic cord-forming basidiomycetePhanerochaete velutina. Mycological Research 105: 859e867.
Williams MA, Rice CW, 2007. Seven years of enhanced wateravailability influences the physiological, structural andfunctional attributes of a soil microbial community. AppliedSoil Ecology 35: 535e545.
Wolters V, Silver WL, Bignell DE, Coleman DC, Lavelle P, van derPutten WH, De Ruiter P, Rusek J, Wall DH, Wardle DA,Brussaard L, Dangerfield JM, Brown VK, Giller KE, Hooper DU,Sala O, Tiedje J, van Veen JA, 2000. Effects of global changes onabove- and belowground biodiversity in terrestrialecosystems: implications for ecosystem functioning. Bioscience50: 1089e1098.
Yeates GW, Bongers T, de Goede RGM, Freckman DW,Georgieva SS, 1993. Feeding habits in soil nematode familiesand genera e an outline for soil ecologists. Journal ofNematology 25: 315e331.
Yuste JC, Penuelas J, Estiarte M, Garcia-Mas J, Mattana S, Ogaya R,Pujol M, Sardans J, 2011. Drought-resistant fungi control soilorganic matter decomposition and its response totemperature. Global Change Biology 17: 1475e1486.
Zak DR, Pregitzer KS, Curtis PS, Teeri JA, Fogel R, Randlett DL,1993. Elevated atmospheric CO2 and feedback between carbonand nitrogen cycles. Plant and Soil 151: 105e117.
Zhang W, Parker KM, Luo Y, Wan S, Wallace LL, Hu S, 2005. Soilmicrobial responses to experimental warming and clipping ina tallgrass prairie. Global Change Biology 11: 266e277.
Zimmer M, 2004. Effects of temperature and precipitation on afloodplain isopod community: a field study. European Journal ofSoil Biology 40: 139e146.
pacts of climate change on interactions among saprotrophicrates, Fungal Ecology (2013), http://dx.doi.org/10.1016/