ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Aug. 1975, p. 111-116 Copyright 0 1975 American Society for Microbiology Vol. 8, No. 2 Printed in U.S.A. Potential for In Vivo Acquisition of R Plasmids by One Strain of Vibrio cholerae Biotype El Tor R. BRENT DAVEY* AND JAMES PITTARD Department of Microbiology, University of Melbourne, Parkville, Victoria, 3052, Australia Received for publication 18 March 1975 The feces of five patients admitted to a hospital during an outbreak of cholera in Melboumne, Australia, in November 1972, were examined for the presence of tetracycline-resistant coliforms and tetracycline-resistant strains of Vibrio cholerae. Despite the abundance of tetracycline-resistant coliforms able to transfer this resistance to other strains of Escherichia coli, no tetracycline-resist- ant strains of V. cholerae were detected. In vitro transfer experiments using the V. cholerae strain responsible for the outbreak as recipient revealed that it was a particularly poor host for most R plasmids. Apparently normal and healthy individuals may harbor among their bowel flora coliform organisms resistant to one or more antibiotics (8, 18). In the majority of cases such resistance has been found to be mediated by conjugative R plasmids. Oral administration of antibiotics, particularly tetracycline, may be followed by a marked increase in the numbers of organisms in the bowel flora which are resistant to the administered antibiotic (8, 15). In studies by Datta (8), most of these organisms were also found to be resistant to other antibiotics, and again in most cases the resistance was mediated by R plasmids. The serious threat presented by such R plas- mids lies in their potential ability to be trans- ferred from resident nonpathogenic organisms to an invading pathogen. At the very least, the acquistion of an R plasmid by an initially susceptible pathogen may complicate the man- agement and chemotherapy of infections caused by such organisms (1). More serious conse- quences may follow should the acquired R plasmid also mediate increased virulence, as appears to have been the case with the out- breaks of chloramphenicol-resistant Salmonella typhi and Shigella dysenteriae type 1 in Mexico and Central America in recent years (13). In November 1972, there was a limited out- break of cholera in Melbourne which occurred among passengers arriving on an international jet flight from Europe via Bahrain (6, 16, 23). Passengers found to be infected with Vibrio cholerae biotype El Tor were admitted to Fair- field Infectious Diseases Hospital, Melbourne, where they received appropriate treatment, which included a course of oral tetracycline to hasten the elimination of the pathogen from the alimentary tract (14). Naturally occurring strains of V. cholerae carrying R plasmids have been reported in Calcutta (22), and R plasmids have been trans- ferred to strains of V. cholerae in vitro (4, 17). This cholera outbreak provided an opportunity for investigating the possibility that V. cholerae might acquire R plasmids in vivo, by transfer from resistant coliforms resident in the bowel flora. MATERIALS AND METHODS Specimens. Fecal specimens were available from five patients, as paired samples taken before and after the course of tetracycline therapy (500 mg orally four times a day for 6 days; see Table 1). The pretreatment specimens were crude feces provided for primary screening when the outbreak was first reported, whereas post-treatment specimens were provided as feces immersed in buffered salts solution. All speci- mens were stored at 4 C until plate counts of V. cholerae, total coliforms, and tetracycline-resistant coliforms were performed. Patients B to E (Table 1) were not known to have received any antibiotic before the pretreatment speci- mens were collected, but patient A had been undergo- ing treatment with ampicillin for urinary tract infec- tion when the outbreak occurred. Bacteria. (i) Fecal coliform strains. For the purposes of this work, these were defined as lactose- fermenting, gram-negative rods isolated from Mac- Conkey agar after incubation for 24 h at 37 C. All strains examined gave biochemical reactions typical of either Escherichia or Klebsiella-Aerobacter species (12). Serological identification of the strains was not attempted. (ii) Laboratory Escherichia coli K-12 strains. The nalidixic acid-resistant strains JP990 (a multiple auxotroph) and JP996 (wild type, prototroph) were used as recipients in matings with R plasmid-bearing (i.e., R+) coliform and V. cholerae strains and also were used as indicators for detecting colicin produc- tion. The streptomycin-resistant multiple auxotrophs 111 on August 20, 2018 by guest http://aac.asm.org/ Downloaded from
Transcript
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Aug. 1975, p. 111-116Copyright 0 1975 American Society for Microbiology
Vol. 8, No. 2Printed in U.S.A.
Potential for In Vivo Acquisition of R Plasmids by OneStrain of Vibrio cholerae Biotype El Tor
R. BRENT DAVEY* AND JAMES PITTARD
Department of Microbiology, University of Melbourne, Parkville, Victoria, 3052, Australia
Received for publication 18 March 1975
The feces of five patients admitted to a hospital during an outbreak of cholerain Melboumne, Australia, in November 1972, were examined for the presence oftetracycline-resistant coliforms and tetracycline-resistant strains of Vibriocholerae. Despite the abundance of tetracycline-resistant coliforms able totransfer this resistance to other strains of Escherichia coli, no tetracycline-resist-ant strains of V. cholerae were detected. In vitro transfer experiments using theV. cholerae strain responsible for the outbreak as recipient revealed that it wasa particularly poor host for most R plasmids.
Apparently normal and healthy individualsmay harbor among their bowel flora coliformorganisms resistant to one or more antibiotics(8, 18). In the majority of cases such resistancehas been found to be mediated by conjugative Rplasmids. Oral administration of antibiotics,particularly tetracycline, may be followed by amarked increase in the numbers of organisms inthe bowel flora which are resistant to theadministered antibiotic (8, 15). In studies byDatta (8), most of these organisms were alsofound to be resistant to other antibiotics, andagain in most cases the resistance was mediatedby R plasmids.The serious threat presented by such R plas-
mids lies in their potential ability to be trans-ferred from resident nonpathogenic organismsto an invading pathogen. At the very least, theacquistion of an R plasmid by an initiallysusceptible pathogen may complicate the man-agement and chemotherapy of infections causedby such organisms (1). More serious conse-quences may follow should the acquired Rplasmid also mediate increased virulence, asappears to have been the case with the out-breaks of chloramphenicol-resistant Salmonellatyphi and Shigella dysenteriae type 1 in Mexicoand Central America in recent years (13).
In November 1972, there was a limited out-break of cholera in Melbourne which occurredamong passengers arriving on an internationaljet flight from Europe via Bahrain (6, 16, 23).Passengers found to be infected with Vibriocholerae biotype El Tor were admitted to Fair-field Infectious Diseases Hospital, Melbourne,where they received appropriate treatment,which included a course of oral tetracycline tohasten the elimination of the pathogen from thealimentary tract (14).
Naturally occurring strains of V. choleraecarrying R plasmids have been reported inCalcutta (22), and R plasmids have been trans-ferred to strains of V. cholerae in vitro (4, 17).This cholera outbreak provided an opportunityfor investigating the possibility that V. choleraemight acquire R plasmids in vivo, by transferfrom resistant coliforms resident in the bowelflora.
MATERIALS AND METHODSSpecimens. Fecal specimens were available from
five patients, as paired samples taken before and afterthe course of tetracycline therapy (500 mg orally fourtimes a day for 6 days; see Table 1). The pretreatmentspecimens were crude feces provided for primaryscreening when the outbreak was first reported,whereas post-treatment specimens were provided asfeces immersed in buffered salts solution. All speci-mens were stored at 4 C until plate counts of V.cholerae, total coliforms, and tetracycline-resistantcoliforms were performed.
Patients B to E (Table 1) were not known to havereceived any antibiotic before the pretreatment speci-mens were collected, but patient A had been undergo-ing treatment with ampicillin for urinary tract infec-tion when the outbreak occurred.
Bacteria. (i) Fecal coliform strains. For thepurposes of this work, these were defined as lactose-fermenting, gram-negative rods isolated from Mac-Conkey agar after incubation for 24 h at 37 C. Allstrains examined gave biochemical reactions typicalof either Escherichia or Klebsiella-Aerobacter species(12). Serological identification of the strains was notattempted.
(ii) Laboratory Escherichia coli K-12 strains.The nalidixic acid-resistant strains JP990 (a multipleauxotroph) and JP996 (wild type, prototroph) wereused as recipients in matings with R plasmid-bearing(i.e., R+) coliform and V. cholerae strains and alsowere used as indicators for detecting colicin produc-tion. The streptomycin-resistant multiple auxotrophs
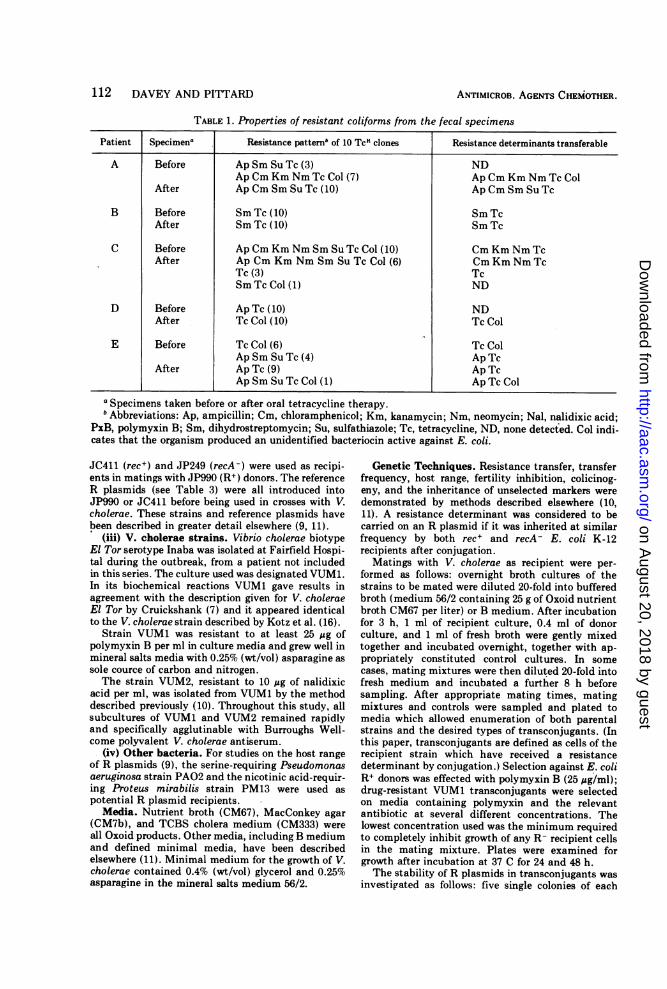
A Before Ap Sm Su Tc (3) NDApCm Km Nm Tc Col (7) Ap Cm Km Nm Tc Col
After Ap Cm Sm Su Tc (10) Ap Cm Sm Su Tc
B Before Sm Tc (10) Sm TcAfter Sm Tc (10) Sm Tc
C Before Ap Cm Km Nm Sm Su Tc Col (10) Cm Km Nm TcAfter Ap Cm Km Nm Sm Su Tc Col (6) Cm Km Nm Tc
Tc (3) TcSm Tc Col (1) ND
D Before Ap Tc (10) NDAfter Tc Col (10) Tc Col
E Before Tc Col (6) Tc ColAp Sm Su Tc (4) Ap Tc
After Ap Tc (9) Ap TcAp Sm Su Tc Col (1) Ap Tc Col
a Specimens taken before or after oral tetracycline therapy.h Abbreviations: Ap, ampicillin; Cm, chloramphenicol; Km, kanamycin; Nm, neomycin; Nal, nalidixic acid;
PxB, polymyxin B; Sm, dihydrostreptomycin; Su, sulfathiazole; Tc, tetracycline, ND, none detected. Col indi-cates that the organism produced an unidentified bacteriocin active against E. coli.
JC411 (rec+) and JP249 (recA-) were used as recipi-ents in matings with JP990 (R+) donors. The referenceR plasmids (see Table 3) were all introduced intoJP990 or JC411 before being used in crosses with V.cholerae. These strains and reference plasmids havebeen described in greater detail elsewhere (9, 11).
(iii) V. cholerae strains. Vibrio cholerae biotypeEl Tor serotype Inaba was isolated at Fairfield Hospi-tal during the outbreak, from a patient not includedin this series. The culture used was designated VUML.In its biochemical reactions VUM1 gave results inagreement with the description given for V. choleraeEl Tor by Cruickshank (7) and it appeared identicalto the V. cholerae strain described by Kotz et al. (16).
Strain VUM1 was resistant to at least 25 ug ofpolymyxin B per ml in culture media and grew well inmineral salts media with 0.25% (wt/vol) asparagine assole cource of carbon and nitrogen.The strain VUM2, resistant to 10 lsg of nalidixic
acid per ml, was isolated from VUM1 by the methoddescribed previously (10). Throughout this study, allsubcultures of VUM1 and VUM2 remained rapidlyand specifically agglutinable with Burroughs Well-come polyvalent V. cholerae antiserum.
(iv) Other bacteria. For studies on the host rangeof R plasmids (9), the serine-requiring Pseudomonasaeruginosa strain PAO2 and the nicotinic acid-requir-ing Proteus mirabilis strain PM13 were used aspotential R plasmid recipients.
Media. Nutrient broth (CM67), MacConkey agar(CM7b), and TCBS cholera medium (CM333) wereall Oxoid products. Other media, including B mediumand defined minimal media, have been describedelsewhere (11). Minimal medium for the growth of V.cholerae contained 0.4% (wt/vol) glycerol and 0.25%asparagine in the mineral salts medium 56/2.
Genetic Techniques. Resistance transfer, transferfrequency, host range, fertility inhibition, colicinog-eny, and the inheritance of unselected markers weredemonstrated by methods described elsewhere (10,11). A resistance determinant was considered to becarried on an R plasmid if it was inherited at similarfrequency by both rec+ and recA- E. coli K-12recipients after conjugation.
Matings with V. cholerae as recipient were per-formed as follows: overnight broth cultures of thestrains to be mated were diluted 20-fold into bufferedbroth (medium 56/2 containing 25 g of Oxoid nutrientbroth CM67 per liter) or B medium. After incubationfor 3 h, 1 ml of recipient culture, 0.4 ml of donorculture, and 1 ml of fresh broth were gently mnixedtogether and incubated overnight, together with ap-propriately constituted control cultures. In somecases, mating mixtures were then diluted 20-fold intofresh medium and incubated a further 8 h beforesampling. After appropriate mating times, matingmixtures and controls were sampled and plated tomedia which allowed enumeration of both parentalstrains and the desired types of transconjugants. (Inthis paper, transconjugants are defined as cells of therecipient strain which have received a resistancedeterminant by conjugation.) Selection against E. coliR+ donors was effected with polymyxin B (25 Ag/ml);drug-resistant VUM1 transconjugants were selectedon media containing polymyxin and the relevantantibiotic at several different concentrations. Thelowest concentration used was the minimum requiredto completely inhibit growth of any R- recipient cellsin the mating mixture. Plates were examined forgrowth after incubation at 37 C for 24 and 48 h.The stability of R plasmids in transconjugants was
investigated as follows: five single colonies of each
purified transconjugant to be tested were inoculatedinto separate drug-free broths. Colony counts on
media with and without appropriate antibiotics were
performed immediately and again after incubatingthe broth cultures ovemight. The ratio of the countson media with and without antibiotics provided a
measure of the stability with which a resistancedeterminant or plasmid was inherited. The resultsobtained were checked by examining colonies fromdrug-free media for the presence of resistance deter-minants, as unselected markers. Inheritance was
regarded as stable if no less than 99.5% of an initiallyresistant population retained the determinant inquestion after overnight incubation in drug-freebroth.
Antibiotic resistance testing. Minimal inhibitoryconcentrations (MIC) were established, and suscepti-ble and resistant organisms were distinguished, as
described previously (10), except that only nutrientagar, B medium, or defined minimal media were used.The following antibiotics were used: ampicillin, chlor-amphenicol, kanamycin, neomycin, nalidixic acid,polymyxin B, dihydrostreptomycin, sulphathiazole,and tetracycline.
RESULTS
Analysis of fecal flora. Examination of fecalsamples collected from the five patients at thetime of admission to hospital showed that thenumber of V. cholerae present ranged from < 10to 3 x 101 organisms per g of feces. The numberof coliforms per gram of feces was, on the otherhand, fairly constant at about 10, and a varia-ble percentage of these (ranging from 0.1 to20%) were resistant to tetracycline. No tetracy-cline-resistant strains of V. cholerae were de-tected. After treatment with tetracycline (500mg orally four times a day for 6 days), althoughthere was no significant change in the totalnumbers of coliforms present more than 50% ofthese were in each case resistant to tetracycline.This finding agrees with previous similar stud-ies (8, 15, 18). No V. cholerae were isolated fromthe post-treatment specimens.From each specimen 10 tetracycline-resistant
coliform colonies were picked at random and
examined both for their resistance to otherantibiotics and their ability to transfer resist-ance determinants to E. coli; The results aresummarized in Table 1. Most of the resistancedeterminants were transferable, the tetracy-cline (Tc) determinant being transferred atfrequencies ranging from 10-2 to 10-" per donorcell in 2-h crosses. In every case the F-fertilityinhibition (fi+) character was also inherited byTc + transconjugants.Transfer of resistance determinants from
the fecal coliforms to V. cholerae strainVUMI. The R+ coliform strains were tested fortheir ability to transfer resistance determinantsto the strain of V. cholerae responsible for theoutbreak, strain VUML. A total of 14 strains,comprising seven from specimens collected be-fore tetracycline treatment and seven fromspecimens taken after treatment, were exam-ined. Despite repeated attempts, transfer ofresistance determinants to VUM1 was detectedfrom only two strains, namely those which hadbeen isolated in the post-treatment specimensfrom patients D (CF902) and E (CF1002). Theresults of these crosses are summarized in Table2.The R+ VUM1 transconjugants formed in
these matings were able to form single colonieson media containing no more than 7.5 jig(CF902) or 5 ,ug (CF1002) of tetracycline per
ml, and in the absence of antibiotic selection theresistance markers were not stably inherited:after overnight incubation in drug-free brothonly. 0.5% (CF902) or 0.05% (CF1002) of initiallyresistant transconjugant populations still re-
tained their resistance determinants. Furthertransfer of resistance determinants from thesetransconjugants to either E. coli or the V.cholerae strain VUM2 was not detected. Theseproperties were in marked contrast to those ofthe corresponding E. coli transconjugants fromthese two donors: such transconjugants were
resistant to more than 30 ,g of tetracycline per
ml, were fertile in crosses with E. coli, and
TABLE 2. Flora coliform strains transferring resistance determinants to V. cholerae VUMI
Transfer from coliform to V. cholerae VUMlRetransfer to
Strain no. R plasmid Transfer frequency Mating Transconjugants Determinantsocharacters' to E. coli' time recovered per expressed in V. choleraerequired donor VUMi VUM2
(h)
CF902 fi+Tc Col 1.1 x 10-2 30 5 x 10- Tcc NDCF1002 fi+ApTc 6.3 x 10-4 30 2 x 10-8 ApTc ND
a Tc, Tetracycline; Ap, ampicillin, ND, not detected.Transconjugants recovered per donor after mating for 2 h.
c None of 40 Tc+ VUMl transconjugants appeared to be Col+.
maintained their resistance determinants sta-bly in the absence of antibiotic selection.
V. cholerae strain VUMI as a host for Rplasmids. Further investigation into the behav-ior of strain VUM1 as a host for R plasmids was
prompted by the difficulty experienced in dem-onstrating resistance transfer to that strain,and by the infertility and unexpectedly lowdrug resistance of the transconjugants obtained.Kuwabara et al. (17) reported that certain of
their cultures of V. cholerae were apparentlyunable to act as recipients of R plasmids.Further, Datta and Hedges (9) have shown that,for at least some of the different incompatibilitygroups of R plasmids, there is an apparentcorrelation between the incompatibility group ofa plasmid and its host range. For example,plasmids of the F group (to which the fi+plasmids from the coliforms possibly belonged)were able to form detectable transconjugants inProteus species, but not in species of Pseudomo-nas or Agrobacterium. Accordingly, the abilityof the V. cholerae strain VUM1 to act as a
recipient for reference plasmids of known in-compatibility group and host range was investi-gated. The stability of resistance determinantsin the VUM1 transconjugants recovered, andthe concentrations of antibiotics to which suchtransconjugants were resistant were also exam-ined. The results are summarized in Tables 3and 4.VUM1 transconjugants harboring R plasmids
from several of the major groups were recovered.Of the reference plasmids tested, only those ofgroup Ia (namely R64-11 and R144-3) were
apparently unable to form transconjugants inVUMi.
By comparison with their transfer frequencyto E. coli (see Table 3), the apparent frequencywith which the F group plasmids formed trans-conjugants in VUM1 was reduced by factors of101 and 10. in 2-h matings. With the plasmids
of groups P, N, and W, the reduction in ap-parent transfer frequency was only about 10fold.The low proportion of R+ transconjugants in
the population of recipient cells, in matingscontinued for 30 h (see above), suggested thatVUM1 cells newly infected with R plasmidswere inefficient donors of their R plasmids toother cells. Attempts to establish HFT cultures(see reference 24) from VUM1 transconjugantscarrying plasmids which were not already fertil-ity derepressed were unsuccessful. The onlyestablished VUM1 transconjugants capable oftransferring their plasmid to either of the E. colistrains JP990 or JP996, or to the V. choleraestrain VUM2, were those harboring the P groupplasmid RP4 or the derepressed F,, group plas-mid R1-16. VUM1 (RP4)+ transferred RP4 at afrequency of 5 x 10- 6per donor to E. coli, and of2 x 10-5 per donor to V. cholerae VUM2. Thecorresponding frequencies for VUM1 (R1-16)+were 10-6 and 10-5. In both cases mating for 30h was required before transconjugants could bedetected, and in both cases the E. coli (R+)transconjugants appeared identical to the origi-nal E. coli (R+) reference plasmid donors.When tested as described, each of the refer-
ence plasmids studied was inherited stably in E.coli transconjugants. In contrast, each plasmidwas inherited less stably in V. cholerae strainVUMl: only 60 to 80% (R1-16, 222/R4, RP4),30% (S-a), or 0.003% (R46) of initially resistanttransconjugant populations retained the resist-ance determinant tested after overnight incuba-tion in drug-free broth. In each case, coloniesfrom drug-free media were shown to carry allthe resistance determinants originally presentor none at all, suggesting that the entire plas-mid, rather than an individual determinant,was lost in the absence of antibiotic selection.Table 4 shows the MIC for each of the
antibiotics used against strain VUM1 and its R+
TABLE 3. Transfer of reference plasmids to E. coli and V. cholerae VUMI
Transconjugants recovered per donor in crosses'R plasmid Determinants Incompatibilityin E. coli carrieda group Mating To To V. cholerae
time (h) E. coli VUM1
R1-16 Km Nm (fertility F,, 2 ca. 2 x 10° 1 x 10-6derepressed)
222/R4 Cm Sm Su Tc F,l 30 >10-1 1 x 10-6RP4 ApKmNmTc P 2 3 x 10- 2 x 10-4R46 Ap Sm Su Tc N 2 4 x 10-' 1 x 10-4
30 9 x 10-2 3 x 10-7S-a Cm Km Sm Su W 2 4 x 10-4 5 x 10-5
30 ca. 10-' 2 x 10-5
aKm, Kanamycin; Nm, neomycin; Cm, chloramphenicol; Sm, dihydrostreptomycin; Su, sulfathiazole; Tc,tetracycline; Ap, ampicillin.
b Ratios of donor cells to recipient cells at the end of the mating period were between 1:6 and 1:10.
a Figures shown are MIC values (gg/ml) in nutrientagar. MIC values for sulphathiazole were determinedin defined minimal media.
The MIC for nalidixic acid against V. choleraeVUM2 was 20 Ag/ml.transconjugants. VUM1 transconjugants har-boring plasmids which carried the same resist-ance determinant showed similar MIC for therelevant drug.
found in the post-treatment specimens from twodifferent patients, were found to be able totransfer a determinant for tetracycline resist-ance to the V. cholerae strain VUMl responsiblefor the outbreak. One of these strains alsotransferred a determinant for ampicillin resist-ance to VUM1. The apparent frequency withwhich these two strains transferred the tetracy-cline-resistance determinant to VUM1 was verylow. Such transfer was achieved in antibiotic-free broth after prolonged mating, and theresulting V. cholerae (Tc+) transconjugantswere resistant to only 5 to 7 ,ug of tetracyclineper ml.
Tetracycline concentrations in excess of 100gg/ml are achieved in the contents of the smallintestine within 3 h of a single oral 500-mg dose(25) and fecal concentrations in excess of 500lsg/g would be expected from a regimen of 500mg orally four times a day (3). Any tetracycline-resistant VUM1 (R+) cells, bearing the coliformR plasmids studied here, would have been quiteunable to survive these levels of tetracycline.
Inheritance of the two fi+ plasmids from thecoliform strains, and of the reference plasmids,was notably unstable in strain VUM1. Similarunstable inheritance of R plasmids by V.cholerae has also been reported elsewhere (17;K. Rahal, G. R. Gerbaud, and D. H. Bouan-chaud, Abstr. Annu. Meet. Am. Soc. Micro-biol., 1974, G149, p 44). It is possible that suchinstability precluded the recovery of R+ VUM1transconjugants from crosses with the other 12coliform strains examined. The observed insta-
bility of the reference plasmids R46 and S-a inVUM1 probably accounts for the lower numbersof R+ transconjugants recovered when matingsinvolving these plasmids were prolonged from 2to 30 h in drug-free media (see Table 3).
It was assumed that the ability of the coliformR plasmids to be transferred to V. choleraestrain VUM1 in vitro would reflect the likeli-hood that they might be transferred to that hostin the alimentary tract. It has been shown thattransfer of R plasmids between organisms in thealimentary tract occurs at very low frequency,even where, in vitro, the R plasmid involved canbe transferred to the same recipient strain athigh frequency (2). Thus given the low fre-quency with which R plasmid transfer to VUM1from the fecal coliforms occurred in vitro, thepossibility that VUM1 might have acquired anR plasmid by conjugal transfer in the alimen-tary tract was remote. Had such an eventoccurred, the observed instability of R plasmidsin VUM1 would mean that any VUM1 (R+)cells formed could be lost in the absence ofantibiotic selective pressure.
In these five patients from this particularoutbreak at least, it must therefore be con-cluded that R plasmid transfer to the infectingV. cholerae strain was an extremely remotepossibility, and not likely to create significantproblems in the successful management andchemotherapy of these patients.Only one V. cholerae strain, the VUMl strain
responsible for this particular outbreak, wasexamined in these studies. From the results ofexperiments with the reference plasmids severalof the variables affecting the formation andproperties of VUM1 (R+) transconjugants be-come apparent. Datta and Hedges (9) haveestablished the principle that an R plasmid mayhave a limited host range, and certainly plas-mids of group Ia appeared unable to formtransconjugants in V. cholerae. F group fi+plasmids formed transconjugants in VUM1at very low frequency, which might also explainwhy so few of the fi+ coliform plasmids wereable to form detectable transconjugants in thishost.
Yet the main effect on the frequency offormation of VUM1 (R+) strains appeared to liein strain VUMl itself. This strain may not havebeen typical of V. cholerae biotype El Torstrains found in India or Southeast Asia withrespect to its ability to act as a host for Rplasmids. For example, Prescott et al. (22)readily demonstrated transfer of the R plasmid222 both into and out of V. cholerae strains.(This 222 plasmid and the 222/R4 plasmid [19]used here were probably identical, since bothwere originally provided as R222 by T. Wata-
nabe.) Their V. cholerae (222) + transconju-gants showed the same high levels of drugresistance as their E. coli (222)+ donor strain.Kuwabara et al. (17) also appeared to have nodifficulty in demonstrating R plasmid transferto and from 14 different V. cholerae biotype ElTor strains, and the resistance of the transcon-jugants, as indicated by their MIC values, wasconsiderably higher (25 to 50 ug of tetracyclineper ml than that observed for VUM1 (R+)transconjugants (5 to 7 jig of tetracycline perml).
It is not known wny VUM1 was such apoor host for R plasmids generally. (Theonly plasmid for which VUM1 appeared wellsuited as a host was the P group plasmid RP4,which has a remarkable host range [see refer-ence 21].) Okada et al. (20) observed that thetransfer frequency of an F-like R plasmid to aSalmonella host which restricted and modifiedforeign deoxyribonucleic acid was 10,000 timeslower than its transfer frequency to a nonre-stricting host strain. Although a similar reduc-tion in transfer frequency was observed herewith F-like plasmids in matings with VUM1,the results with the other reference plasmidssuggest that restriction did not have an impor-tant effect on the frequency with which R+transconjugants of VUM1 were formed. Cer-tainly restriction would not account for theinstability of R plasmids in VUM1, nor for theinefficiency with which their determinants wereapparently expressed. The possibility thatVUM1 harbors a prophage or a sex factor (seereference 5) which might affect the entry, inher-itance, and expression of R plasmids in thisstrain was not investigated.
ACKNOWLEDGMENTSThis investigation was supported by a grant from the
National Health and Medical Research Council. R.B.D. wassupported by a Commonwealth Postgraduate Researchaward.We thank J. A. Forbes, Fairfield Hospital, for providing
fecal specimens, and N. Datta and R. C. Clowes forproviding reference plasmids and bacterial strains. Ampicil-lin and nalidixic acid were generous gifts from BeechamResearch Laboratories and Winthrop Laboratories, respec-tively.
LITERATURE CITED
1. Anderson F. M., N. Datta, and E. J. Shaw. 1972. Rfactors in hospital infection Br. Med. J. 3:82-85.
2. Anderson, J. D., W. A. Gillespie, and M. H. Richmond.1973. Chemotherapy and antibiotic-resistance transferbetween enterobacteria in the human gastrointestinaltract. J. Med. Microbiol. 6:461-473.
3. Baker, H. J., and E. J. Pulaski. 1950. Effects of ter-ramycin on faecal flora. Ann. N.Y. Acad. Sci. 53:324-331.
4. Baron, L. S., and S. Falkow. 1961. Genetic transfer ofepisomes from Salmonella typhosa to Vibrio cholerae.
Genetics 46:849.5. Bhaskaran, K., and V. B. Sinha. 1971. Transmissible
plasmid factors and fertility inhibition in Vibriocholerae. J. Gen. Microbiol. 69:89-97.
6. Comment. 1972. New rules for cholera. Med. J. Austral.2:1332.
7. Cruickshank, R. (ed.). 1965. Medical microbiology. Aguide to the laboratory diagnosis and control of infec-tion, 11th ed. E. and S. Livingstone Ltd., London.
8. Datta, N. 1971. R. factors in Escherichia coli. Ann. N. Y.Acad. Sci. 182:59-64.
9. Datta, N., and R. W. Hedges. 1972. Host ranges of Rfactors. J. Gen. Microbiol. 70:453-460.
10. Davey, R B., and A. J. Pittard. 1971. Transferablemultiple antibiotic resistance amongst Shigella strainsisolated in Melbourne between 1952 and 1968. Med. J.Austral. 1:1367-1370.
11. Davey, R. B., and J. Pittard. 1974. Genetic and biophysi-cal study of R plasmids conferring sulphonamideresistance in Shigella strains isolated in 1952 and 1956.J. Bacteriol. 120:1186-1195.
12. Edwards, P. R., and W. H. Ewing. 1964. Identification ofenterobacteriaceae. Burgess, Minneapolis.
13. Gangarosa, E. J., J. V. Bennett, C. Wyatt, P. E. Pierce, J.Olarte, P. M. Hernandes, V. Vasquez, and D. M.Bessudo. 1972. An epidemic-associated episome? J.Infect. Dis. 126:215-218.
14. Greenough, W. B., III, R. S. Gordon, Jr., I. S. Rosenberg,B. I. Davies, and A. S. Benenson. 1964. Tetracycline inthe treatment of cholera. Lancet 1:355-357.
15. Hirsh, D. C., G. C. Burton, and D. C. Blenden. 1973.Effect of oral tetracycline on the occurrence of tetracy-cline-resistant strains of Escherichia coli in the intesti-nal tract of humans. Antimicrob. Agents Chemother.4:69-71.
16. Kotz., A., L. Arunachalam, N. Raush, G. Toouli, and R.Dight. 1972. A case of "gastro" which turned out to becholera. Med. J. Austral. 2:1365-1366.
17. Kuwabara, S., T. Akiba, K. Koyama, and T. Arai. 1963.Transmission of multiple drug resistance from Shigellaflexneri to Vibrio comma through conjugation. Jpr. J.Microbiol. 7:61-67.
18. Linton, K. B., P. A. Lee., M. H. Richmond, W. A.Gillespie, A. J. Rowland, and V. N. Baker. 1972.Antibiotic resistance and transferable R factors in theintestinal coliform flora of healthy adults and childrenin an urban and a rural community. J. Hyg. 70:99-104.
19. Nisioka, T., M. Mitani, and R. C. Clowes. 1970. Molecu-lar recombination between R factor deoxyribonucleicacid molecules in Escherichia coli host cells. J. Bacte-riol. 103:166-177.
20. Okada, M., T. Watanabe, and T. Miyake. 1968. On thenature of the recipient ability of Salmonellatyphimurium for foreign deoxyribonucleic acids. J.Gen. Microbiol. 50:241-252.
21. Olsen, R. H., and P. Shipley. 1973. Host range andproperties of the Pseudomonas aeruginosa R factorR1822. J. Bacteriol. 113:772-780.
22. Prescott, L. M., A. Datta, and G. C. Datta. 1968. Rfactors in Calcutta strains of Vibrio cholerae andmembers of the enterobacteriaceae. Bull. W.H.O.39:971-973.
23. Sutton, R. G. A. 1974. An outbreak of cholera in Australiadue to food served in flight on an international aircraft.J. Hyg. 72:441-451.
24. Watanabe, T. 1964. Selected methods of genetic study ofepisome mediated drug resistance in bacteria, p.202-220. In H. N. Eisen (ed.), Methods in medicalresearch, vol. 10. Year Book Medical Publishers, Inc.,Chicago.
25. Wright, L. T., and A. Prigot. 1951. Aureomycin as an aidin abdominal surgery. Ann. Surg. 133:364-373.