Poly rI * Poly rCERIK DE CLERCQ,1 MARC R. NUWER, AND THOMAS C. MERIGAN
Division ofInfectious Diseases, Department of Medicine, Stanford University,Stanford, California 94305
Received for publication 26 March 1970
Intraperitoneal injection of mice with mineral oil, incomplete (IFA) or completeFreund's adjuvant (CFA) increased the interferon response to endotoxin or (polyrI) -(poly rC) administered intravenously 2 days later. After endotoxin adminis-tration, circulating interferon titers were increased at several different times ofsampling and with a variety of endotoxin dosages. When injection of endotoxinwas delayed until 6 to 8 days after the administration of IFA or CFA, interferonproduction was markedly decreased. Mice treated with CFA and injected withendotoxin 2 days later became more resistant to intranasal vesicular stomatitis viruschallenge than mice injected with endotoxin alone. Hyporeactivity to the inter-feron-inducing capacity of a second injection of endotoxin 2 days after the firstinjection could not be overcome by administering CFA simultaneously with the firstdose. CFA treatment not only raised the serum interferon titers produced by endo-toxin, but also increased the number of interferon-forming cells in the spleen afteradministration of endotoxin in vivo. In addition, CFA enhanced the intravascularclearance of (poly rI) * (poly rC). The possibility that Freund's adjuvant increasedthe interferon response to endotoxin and (poly rI) * (poly rC) by stimulating theuptake and processing of the interferon inducer by lymphoreticular cells is dis-cussed.
The interferon response to endotoxin can beincreased by several factors such as preinfectionwith attenuated tubercle bacilli (BCG; references41 and 42), X-ray irradiation (11), adrenalectomy(11, 28), lead acetate (W. R. Stinebring and M.Absher, Proc. Int. Symp. Interferon, Lyons, inpress), cycloheximide (40, 43, 45), and zymosan[a cell wall extract from Saccharomyces cerevisiae(Y. Nagano and N. Maehara, Japan-U.S.A.Interferon Seminar, Tokyo, in press)]. Some ofthese factors (BCG, zymosan) have also been re-ported to stimulate the phagocytic activity of thereticuloendothelial system (5, 30) and to have anadjuvant effect on antibody formation (6, 14).Other factors such as kinetin riboside, thorotrast,corticoids, cyclophosphamide, and cytotoxicdrugs have suppressing effects on both antibodyand interferon production (as reviewed by E. DeClercq and T. C. Merigan, Annu. Rev. Med., inpress).
'Fellow of the Damon Runyon Memorial Fund for CancerResearch, "Aangesteld Navorser" of the Belgian N.F.W.O.(Nationaal Fonds voor Wetenschappelijk Onderzoek).
69
Interferon appears to be related to the immunemechanism. It is produced in vitro in humanlymphocytes from immune donors upon exposureto the specific antigen (tuberculin, tetanus ordiphtheria toxoids; reference 21) and in vivo inBCG-infected mice upon challenge with tubercu-lin (W. R. Stinebring and M. Absher, Ann. N.Y.Acad. Sci., in press). Peritoneal leukocytes ob-tained from mice immunized with virus or endo-toxin show higher interferon production thancells of control mice when exposed to the specificinducer in vitro (18; M. Yamada and M. Azuma,Japan-U.S.A. Interferon Seminar, Tokyo, inpress).As Freund's adjuvant enhances antibody for-
mation (16), partly through stimulation of thephagocytic activity of the reticuloendothelial sys-tem (26), we were interested in whether it mightalso increase the interferon response. Therefore,Freund's adjuvant [both complete (CFA) and in-complete (IFA) Freund's adjuvant] was injectedintraperitoneally into mice at several times priorto the intravenous injection of either Salmonella
enteritidis endotoxin (ESE) or polyriboinosinicacid/polyribocytidylic acid homopolymer pair[(poly rI) * (poly rC)]. Interferon titers weremeasured at various times after injection of ESEand with various doses of the inducer. The rate ofintravascular clearance of (poly rI) * (poly rC)was studied in CFA-treated and untreated mice.The influence of CFA on the interferon responseto ESE was studied in spleen cells, and the en-hanced interferon-inducing capacity of ESE inCFA-treated animals was correlated with an en-hanced effect against intranasal challenge withvesicular stomatitis virus.
MATERUILS AND METHODSAnimals. Swiss-Webster albino mice were used in
this study. They were housed in an air-conditionedroom at 20 C and given food and water ad libitum.Interferon production experiments were performedwith mice weighing 20 to 25 g and antiviral protectionstudies with mice weighing 10 to 12 g.
Materials used. ESE, CFA, and IFA were pur-chased from Difco Laboratories, Detroit, Mich. ESEwas prepared in stock solution of 1 mg/ml of normalsaline and stored at 4 C. CFA is a suspension of killedand dried Mycobacteriwn butyricum in a mixture of8.5 ml of paraffin oil (Bayol F) and 1.5 ml of mannidemonoleate (Arlacel). IFA is similar to CFA but with-out acid-fast bacilli. The paraffin oil itself is a mixtureof 42.5% paraffins, 31.4% monocyclic naphthalenes,and 26.1% polycyclic naphthalenes (39). Polyriboino-sinic and polyribocytidylic acids were purchased fromP-L Biochemicals, Inc., Milwaukee, Wis., and the(poly rI) * (poly rC) homopolymer pair was preparedand characterized as described previously (7). Theradioactive (poly rI) * (poly rC-3H) was prepared byannealing equal volumes of one time concentratedpoly rC-3H (Schwarz BioResearch, Orangeburg, N.J.)with two times concentrated poly rI (Miles Labora-tories, Elkhart, Ind.); the resultant specific radioac-tivity was 0.006 ,uc/,ug (3).
Interferon production. Interferon titers in the mouseserum were measured with a plaque reduction assayon mouse L-cell monolayers employing bovine vesicu-lar stomatitis virus (VSV), Indiana strain, as a chal-lenge virus (24). The interferon titer corresponded tothe reciprocal of the highest dilution of the samplewhich reduced virus plaque formation by 50%. Inter-feron was characterized by its sensitivity to trypsin(1.0 mg/ml, 37 C, 1 hr), thermolability (56 C, 1 hr),nonsedimentability (100,000 X g, 2 hr), and speciesspecificity (lack of activity in human skin fibroblasts).
Antiviral protection. The protective activity of ESEwas studied in mice against intranasal inoculation withVSV, an experimental virus infection, used previouslyfor measuring the antiviral efficacy of interferon in-ducers and interferon (8; E. De Clercq et al., sub-mitted for publication). Unanesthetized 20-day-oldmice (weighing 10 to 20 g) were inoculated with 5.106plaque-forming units (measured in primary chick em-bryo fibroblasts) of VSV by applying 0.02 ml of thevirus suspension in four to six drops to the externalnares from a needle (no. 27 X 0.5 inch) mounted on a
0.25-ml syringe. Mortality was recorded daily for 15days. Most animals died between the 5th and 9th days.Statistical significance was assessed by the x2 test,computed for the final numbers of mice which died orsurvived in treated and control groups.
Interferon-producing cells in the spleen. Interferonresponse of the spleen was measured by the interferon-forming cell assay technique used by Osborn andWalker (27) for the detection of interferon productionby individual spleen cells of mice after intravenous in-jection of Newcastle disease virus and Sindbis virus.The mice were anesthetized and exsanguinated; theirspleens were removed and passed through a no. 4gauge stainless steel mesh into 1 ml ofMEM (minimalEagle's medium). One to three serial dilutions of thespleen cell suspension were made in MEM, and 0.2 mlof each dilution was mixed vigorously with 2.0 ml ofan agar-MEM mixture containing three parts ofMEMfor one part of agar (1.9%) and one part of MEM(two times concentrated). The cell-agar-MEM mix-tures were poured onto confluent L-cell monolayers in60-mm petri dishes and allowed to gel at room tem-perature. After 6 hr of incubation at 37 C, cell-agar-MEM mixtures were poured off, and the monolayerswere washed with MEM to remove all traces of agar.The cells were challenged with VSV (0.1 plaque-form-ing units per cell); unadsorbed virus was removedafter 45 min of incubation at 37 C and replaced by a0.9% agar overlay. The monolayers were incubatedat 37 C for 3 days, and foci of protected cells werestained with neutral red. Foci could be counted with-out magnification, but their somewhat diffuse outlineonly permitted a semiquantitative estimation.
Clearance of (poly rI) * (poly rC) from the circulation.Two techniques were used to measure the fate of(poly rI) - (poly rC) from the blood stream: serumsamples were tested for their capacity to producecellular resistance to VSV in human skin fibroblastsor for residual (acid-insoluble) radioactivity [follow-ing injection of (poly rl) * (poly rC-'H)]. In both cases,a dose of 20 ,ug of (poly rI) - (poly rC) in 0.5 ml ofsaline or MEM was injected into a tail vein, and bloodwas obtained from the retroorbital plexus at severaltimes after the injection. The blood samples were al-lowed to clot at room temperature for 20 min and werekept at 4 C for 1 to 2 hr. Serum was collected and heldat 4 C during all manipulations preceding incubationon human skin fibroblasts (for determination of cellu-lar resistance to VSV) or precipitation with trichloro-acetic acid (for determination of residual radioac-tivity). Cellular resistance to VSV was measured byexposing human skin fibroblasts to a series of serumdilutions for 20 hr at 37 C; after removal of theseserum dilutions, the monolayers were challenged withVSV, and virus plaques were counted 2 days later.End point of the titration corresponded to the serumdilution which reduced virus plaque formation by50%. The virus plaque reduction obtained in humancells with sera of mice injected with (poly rI) * (polyrC) was attributable to residual circulating (polyrI) - (poly rC), as the antiviral activity was completelydestroyed by prior exposure of the serum sample (di-luted 1:30 in MEM) to 40 ,g/ml of pancreatic ribo-nuclease for 1 hr at 25 C. (Bovine pancreatic ribo-
nuclease-A, five times crystallized, was purchasedfrom Sigma Chemical Co., St. Louis, Mo.). For radio-activity measurements, 0.1 ml of each serum samplewas diluted to 1.0 ml with normal saline, and the ribo-nucleic acid was precipitated by adding 1.0 ml of 5%trichloroacetic acid (at 4 C). The precipitate was fil-tered through Whatman GF/A discs and washed with5% trichloroacetic acid (4 C). The filters were driedand counted in 5 ml of toluene-l 4-bis-2-(5-phenyl-oxazolyl) scintillation fluid in a liquid scintillationspectrometer.
RESULTS
Comparative effect of mineral oil, IFA, and CFAon interferon production. Intraperitoneal injectionof 0.5 ml of mineral oil, IFA, or CFA, 2 daysprior to the intravenous injection of either 2,4gof ESE or 0.2 ,ug of (poly rI) (poly rC), increasedthe interferon response to both ESE and (polyrI) * (poly rC) (Fig. 1). Mineral oil and IFA in-creased the interferon titers by 2 to 3 and CFA by6 to 8 times. Neither mineral oil, IFA, nor CFAalone was able to stimulate interferon productionunder the given conditions.
Dose-response relationship. An enhanced inter-feron response to ESE after administration ofCFA was observed with different doses of ESEtested (Fig. 2). In both CFA-treated and un-treated mice, interferon production increasedwith increasing amounts of endotoxin up to adose of 10 ,g of endotoxin, but interferon produc-tion decreased when the ESE dose was increasedto 50,ug per mouse.
Time-response relationship with interferon pro-duction measured at different times after injectionof ESE; ESE administered 2 days after injection ofCFA. Serum samples taken at different times
L 2 560
- 1280
d 640
C: 3204-
160._
f- a804
L 4,042c
0.4 2 10 50E5E doso5pg/mouso)
FIG. 2. Effect of CFA on interferon production byseveral doses of ESE. CFA (0.5 ml) injected intraperi-toneally 2 days before intravenous injection of ESE.Circulating interferon measured 2 hr after injection ofESE. Mean values (i standard deviation) for three to
five observations.
FIG. 3. Effect of CFA on) interferon production atdifferent times after the injection of ESE. CFA (0.5ml) injected intraperitoneally 2 days before intravenousinjection of 10 ,ug of ESE. Mean values (i standarddeviation) for three observations.
i
0
'4
'4t
ESE kpoly rl)- poly rC,Z,19/'mouSO 0.2 ,ug/mousc
FIG. 1. Comparative effect of mineral oil, IFA, andCFA on interferon production by ESE and (poly rI)-(poly rC); 0.5 ml of mineral oil, IFA, or CFA injectedintraperitoneally 2 days before intravenous injection ofeither ESE or (poly rI) - (poly rC). Circulating inter-feron measured 2 hr after injection of ESE and (polyrI) - (poly rC). Mean values for two observations.
after the injection of ESE showed consistentlyhigher interferon titers in CFA-tested mice whencompared to untreated mice (Fig. 3). For bothtreated and untreated animals, interferon reachedits peak value 2 hr after injection of ESE and de-clined gradually thereafter, so that 8 hr after in-jection nearly no interferon could be detected inthe blood stream. Pretreatment with CFA height-ened but did not change the shape of the time-re-sponse curve of the endotoxin stimulated inter-feron.
Time-response relationship with ESE adminis-tered at different times after injection of CFA or
]UFA; interferon production measured 2 hr after in-jection of ESE. Both CFA and IFA had a bi-phasic effect on the interferon response to ESE(Fig. 4). Interferon titers were increased when
0 2 O 0 D)OW9oftrcrifection°litervolIdoys) btweteenjection of odjuvont oand
injection ofendotoxin FIG. 5. Effect of CFA and ESE on mortality pro-
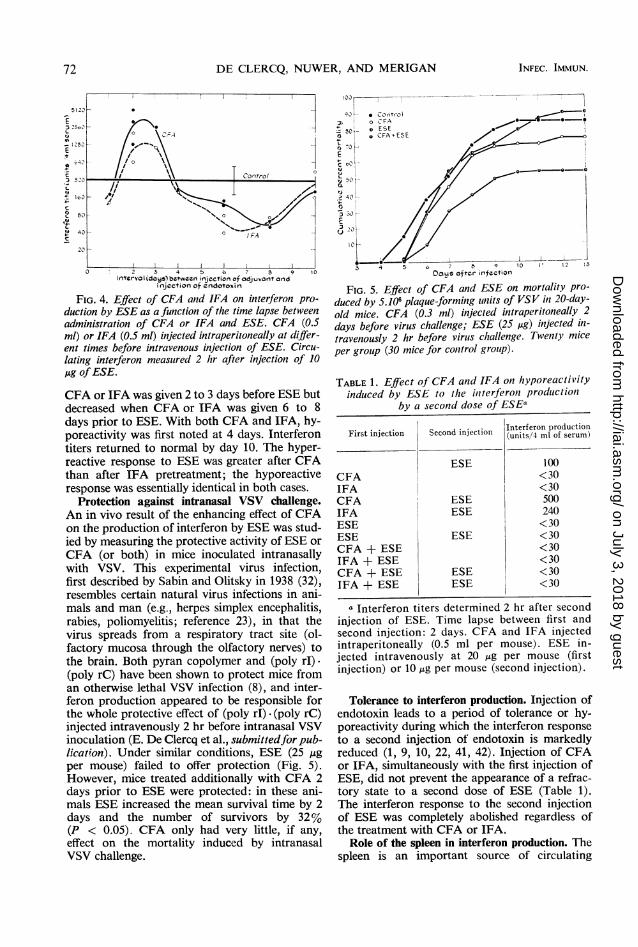
FIG. 4. Effect of CFA and IFA on interferon pro- duced by 5.106 plaque-forming units of VSV in 20-day-duction by ESE as a function of the time lapse between old mice. CFA (0.3 ml) injected intraperitoneally 2administration of CFA or IFA and ESE. CFA (0.5 days before virus challenge; ESE (25 Mg) injected in-ml) or IFA (0.5 ml) injected intraperitoneally at differ- travenously 2 hr before viruts challenge. Twenty miceent times before intravenous injection of ESE. Circu- per grouip (30 mice for conitrol grouip).lating interferon measured 2 hr after injection of 10Mug ofESE.
CFA or IFA was given 2 to 3 days before ESE butdecreased when CFA or IFA was given 6 to 8days prior to ESE. With both CFA and IFA, hy-poreactivity was first noted at 4 days. Interferontiters returned to normal by day 10. The hyper-reactive response to ESE was greater after CFAthan after IFA pretreatment; the hyporeactiveresponse was essentially identical in both cases.
Protection against intranasal VSV challenge.An in vivo result of the enhancing effect of CFAon the production of interferon by ESE was stud-ied by measuring the protective activity of ESE orCFA (or both) in mice inoculated intranasallywith VSV. This experimental virus infection,first described by Sabin and Olitsky in 1938 (32),resembles certain natural virus infections in ani-mals and man (e.g., herpes simplex encephalitis,rabies, poliomyelitis; reference 23), in that thevirus spreads from a respiratory tract site (ol-factory mucosa through the olfactory nerves) tothe brain. Both pyran copolymer and (poly rI)-(poly rC) have been shown to protect mice froman otherwise lethal VSV infection (8), and inter-feron production appeared to be responsible forthe whole protective effect of (poly rI) - (poly rC)
injected intravenously 2 hr before intranasal VSVinoculation (E. De Clercq et al., submittedfor pub-lication). Under similar conditions, ESE (25 jigper mouse) failed to offer protection (Fig. 5).However, mice treated additionally with CFA 2days prior to ESE were protected: in these ani-mals ESE increased the mean survival time by 2days and the number of survivors by 32%(P < 0.05). CFA only had very little, if any,
effect on the mortality induced by intranasalVSV challenge.
TABLE 1. Effect oJ CPA and iLA on nyporeacfU vLLyinduced by ESE to the ilnterferoni production
by a second dose of ESEa
First injection Second injection Interferon production(units/4 ml of serum)
ESE 100CFA <30IFA <30CFA ESE 500IFA ESE 240ESE <30ESE ESE <30CFA + ESE, <30IFA + ESE <30CFA + ESE ESE <30IFA + ESE ESE <30
a Interferon titers determined 2 hr after secondinjection of ESE. Time lapse between first andsecond injection: 2 days. CFA and IFA injectedintraperitoneally (0.5 ml per mouse). ESE in-jected intravenously at 20 ,ug per mouse (firstinjection) or 10lg per mouse (second injection).
Tolerance to interferon production. Injection ofendotoxin leads to a period of tolerance or hy-poreactivity during which the interferon responseto a second injection of endotoxin is markedlyreduced (1, 9, 10, 22, 41, 42). Injection of CFAor IFA, simultaneously with the first injection ofESE, did not prevent the appearance of a refrac-tory state to a second dose of ESE (Table 1).The interferon response to the second injectionof ESE was completely abolished regardless ofthe treatment with CFA or IFA.
Role of the spleen in interferon production. Thespleen is an important source of circulating
interferon in mice and rats injected with eithervirus or endotoxin (10, 17, 27, 36, 38). To eval-uate its role in the increased interferon responseto ESE in CFA-treated mice, spleens obtainedfrom mice injected with CFA or ESE (or both)were studied for their capacity to protect L cellsfrom a 100% lethal VSV challenge (27). Intra-venous injection ofESE resulted in the appearanceof about 4,000 interferon-forming (L-cell-pro-tecting) cells or cell colonies per spleen; thisnumber was increased to 16,000 if the animalswere pretreated with CFA (Table 2). Comparisonof Table 2 with Fig. 1 to 4 shows that this increasein spleen reactivity occurred in parallel to anincreased interferon production in the serum.Control mice and mice treated with CFA onlyhad much lower numbers of L-cell-protectingcells (or cell colonies) in their spleens (Table 2).Intraperitoneal injection of CFA did not cause asignificant change in the number of nucleatedspleen cells 2 days later. This suggests that theincreased number of interferon-forming cells inspleens obtained from mice injected with CFAand ESE, as compared to mice injected withESE alone, was not due to a nonspecific increaseof the total number of cells per spleen but to aspecific increase of the number or reactivity ofsplenocytes participating in the interferon re-sponse to endotoxin.
Clearance of interferon inducer from circulation.The phagocytic activity of the reticuloendothelialsystem (measured by following the rate of clear-ance of carbon from the circulation) is signifi-cantly stimulated on the first and second dayafter injection of CFA (26). As enhancement ofthe reticuloendothelial phagocytic capacity mightbe a reasonable explanation for the increasedinterferon response to ESE and (poly rI) *(poly
TABLE 2. Effect of CFA on the inzduction of spleeninterferon-forming cells by ESE-
Interferon-forming Interferon-formingFirst Second cells (or cell cells (or cell
injection injection colonies) per 10' colonies) perspleen cells spleen
CFA ESE 69.6 16,000ESE 19.7 4,100
CFA 0.4 1200.2 40
a CFA injected intraperitoneally (0.5 ml permouse), 2 days before intravenous injection of 25,ug of ESE. Spleen cell suspensions were prepared1.5 hr later; cell-agar-MEM mixtures were pre-pared and incubated over L cells for 6 hr, afterwhich the monolayers were drained, washed,and challenged with VSV. Residual L-cell fociwere stained and counted at 72 hr.
0
CcOC 100
..,0
E 30
b
Corn rol +ta a 4~~- 0 CIont,-ol*0
I I~~ '. -~204 a0 0 2 40 O 5
Tieinmnue
FIG. 6. Effect of CFA on clearance of (poly ri)-(poly rC) from the circulation. CFA (0.5 ml) injectedintraperitoneally 2 days before intravenous injection of(poly rI)- (poly rC). Circulating (poly rI)- (poly rC)measured at different times after injection of 20 MAgof (poly ri) * (poly rC), either by production of cellularresistance to VSV infection in human skin fibroblasts(A) or by trichloroacetic acid-precipitable radioactivity[after injection of (poly rI) -(poly rC-3H)] (B).
rC) in CFA-treated mice, studies were designedto compare the intravascular clearance rate ofthe interferon inducer in CFA-treated and un-treated animals. (Poly rI) - (poly rC) was chosenfor study in preference to ESE because its rate ofclearance could be measured by relatively simpleprocedures such as its ability to produce cellularresistance to VSV in human skin cells and residual(acid-insoluble) radioactivity [following injectionof tritium-labeled (poly rI) - (poly rC)]. Theclearance rate of (poly rI) -(poly rC) was sig-nificantly enhanced 2 days after injection of CFA(Fig. 6A, measured by antiviral activity; Fig. 6B,measured by radioactivity). With both techniques,the amounts of circulating (poly rI) . (poly rC)could be calculated, based on the VSV plaque-reducing capacity of (poly rI) * (poly rC) in humanskin fibroblasts or its specific radioactivity. Since(poly rI) * (poly rC) reduced VSV plaque forma-tion in human skin fibroblasts by 50% at 0.004yg/ml, whereas serum samples obtained fromCFA-treated and untreated mice 20 min afterinjection of 20 ,ug of (poly rI) * (poly rC) did soat a dilution of 1:200 and 1:800, respectively(Fig. 6A), the respective concentrations of circu-lating (poly rI) . (poly rC) were estimated at 0.8and 3.2 ,ug/ml. Based on the specific radioactivityof the input material (5,000 counts per min per,g), the residual radioactive counts found in theserum of CFA-treated and untreated mice 20 mnmafter injection of 20 ,g of (poly rI) * (poly rC-3H)(see Fig. 6B) corresponded to 2.4 ,ug/ml and 4.4,ug/ml, respectively.
antibody formation (16), stimulates phagocytosis(26), causes extensive proliferation of the lympho-reticular system (33), and produces or increasesdelayed hypersensitivity and autoallergic disease[as reviewed by White (39) ]. Because of itsstimulating effect on both cellular and humoralantibody production and on the phagocyticactivity of the reticuloendothelial system, CFAmight be expected to counteract experimentalvirus infections and tumor formations. It wasnot surprising that Berman et al. (4) foundadenovirus 12 oncogenesis reduced in CFA-treated hamsters and Gorhe (20) found foot-and-mouth-disease virus multiplication inhibited inCFA-treated mice. In other studies, however,Freund's adjuvant increased tumor development[Rous sarcoma in chickens (29), SV40 in hamsters(19), polyoma in mice and Rous sarcoma in rats(37), and adeno 12 in hamsters (2)]. In one study,a single dose of Freund's adjuvant mixed withbovine serum albumin stimulated leukemia inmice infected with Rauscher virus, whereasnumerous injections of the same preparationinhibited the development of leukemia (35). Thedivergent effects of Freund's adjuvant probablydepend on the timing and the dosage administeredand on the particular system under study.The effect of Freund's adjuvant on interferon
production by ESE was also dependent on thetiming of the interferon inducer in relationship tothe time of CFA administration. When injected2 days before ESE, CFA caused a significantincrease of interferon production both at varioustimes after injection of ESE and for a variety ofESE dosages (Fig. 2 and 3). When injection ofESE was delayed until 6 to 8 days after injectionof CFA or IFA, its interferon-inducing capacitydecreased (Fig. 4). The nature of this biphasiceffect of CFA and IFA on interferon productionis not clear, but might be related to an activationfollowed by a suppression of the endotoxin-responsive cells. A similar biphasic effect of asingle intraperitoneal injection of CFA has beenobserved when mouse spleen cells were studiedfor their (in vitro) response to phytohemag-glutinin (31).The hyperreactivity to interferon production
by ESE in Freund's adjuvant-treated mice isremarkably similar to the effects of sublethalX-irradiation (11) and BCG infection (41, 42).X-irradiation, applied from 4 hr to 4 days beforeendotoxin injection, potentiated the interferonproduction in rats (11), and mice infected withBCG 14 days prior to testing showed an increasedsensitivity to the lethal effects of endotoxin andto its interferon-producing capacity as well (41,42). The time-response curve of endotoxin-produced interferon in irradiated rats (11) and
the dose-response curve of endotoxin-producedinterferon in BCG-infected mice (42) corre-sponded closely to the time- and dose-responsecurves obtained in CFA-treated animals (Fig. 2and 3). Treatment with cycloheximide also re-sulted in an increased interferon response to ESEand (poly rI) - (poly rC) (40, 43-45), but, in con-trast to CFA, cycloheximide shifted the interferonpeak titers to later times. These findings suggestthat the mechanisms of action of BCG, CFA,and X-irradiation on the interferon response toendotoxin are fundamentally identical, thoughdifferent from the action of cycloheximide.
Mineral oil and both incomplete and completeFreund's adjuvant stimulated the interferon pro-duction by (poly rI) -(poly rC) and ESE, butCFA was a more powerful stimulant than IFAand mineral oil (Fig. 1). Similarly, CFA was moreactive than IFA in increasing the immune re-sponse [e.g., to Venezuelan equine encephalo-myelitis virus in rabbits (34)] and in stimulatingthe phagocytic activity of the reticuloendothelialsystem in mice (26). Thus, interferon production,immune response, and phagocytosis appeared tobe closely related phenomena, as they were allinfluenced by IFA and CFA in a parallel manner.The greater effect of CFA as compared to mineraloil and IFA is most likely due to the M. butyricumin CFA.The precise mechanism of action of Freund's
adjuvant is not yet settled. It has been postulatedthat mineral oil, CFA, and IFA enhance antibodyproduction by coating the antigen with a thinfilm of oil that protects it from rapid inactivation(15, 34). This hypothesis does not explain whyCFA is more active than IFA and is obviouslylimited to simultaneous injection of adjuvantand antigen by the same route. However, Freund'sadjuvant has been found capable of enhancingthe antibody response to bovine gamma globulinwhen injected by a different route than the antigen(12, 13) or when injected at several days beforethe antigen (25). In our experiments, maximalincrease of interferon production with endotoxinoccurred when Freund's adjuvant was givenintraperitoneally 2 days before the intravenousinjection of the interferon inducer. These resultssuggest that Freund's adjuvant exerts its effecton the cells involved in the processing of theinterferon inducer (or antigen) rather than on theinterferon inducer (or antigen) itself. In this re-spect, addition of acid-fast bacilli to the water-in-oil emulsion might further increase the cellreactivity or involve a greater number of cells,which could account for the greater activity ofCFA over IFA. The particular cell types, whichFreund's adjuvant acts on, are not defined yet,but it is likely that they belong to the reticulo-
endothelial system. The hypothesis that Freund'sadjuvant might enhance antibody and interferonproduction by stimulating the activity of thelymphoreticular system is suggested by theaccelerated intravascular clearance of (poly rI).(poly rC) (Fig. 6) after treatment with CFA.This explanation is directly supported by thefinding that cells of reticuloendothelial organs(such as the spleen) showed an increased inter-feron response to endotoxin in CFA-treatedanimals. However, more work is required todetermine whether the mechanism underlyingpotentiating of interferon production withendotoxin and (poly rI) - (poly rC) is identical.
ACKNOWLEDGMENT
This study was supported by Public Health grant AI-05629
fromii the National Institute of Allergy and Infectious Diseases.
LITERATURE CITED
1. Absher, M., and W. R. Stinebring. 1969. Endotoxin-like
properties of poly I. poly C, an interferon stimulator.Nature (London) 223:715-717.
2. Alford, T. C., A. Hollinshead, and R. J. Huebner. 1969.
Paradoxical effect of Freund's complete adjuvant upontransplantation efficiency of adenovirus-induced tumourcells. J. Gen. Virol. 5:541-543.
3. Bausek, G. H., and T. C. Merigan. 1969. Cell interaction
with a synthetic polynucleotide and interferon productionin vitro. Virology 39:491-498.
4. Berman, L. P., A. C. Allison, and H. G. Pereira. 1967. Effectsof Freund's adjuvant on adenovirus oncogenesis and anti-
body production in hamsters. Int. J. Cancer 2:539-543.
5. Biozzi, G., B. Benacerraf, F. Grumback, B. N. Halpern, J.
Levaditi, and N. Rist. 1954. Etude de l'activite granulo-pexique du systeme reticulo-endothelial au cours de l'infec-tion tuberculeuse experimentale de la souris. Ann. Inst.Pasteur 87:291-300.
6. Cutler, J. L. 1970. The enhancement of hemolysin productionin the rat by zymosan. J. Immunol. 84:416-419.
7. De Clercq, E., and T. C. Merigan. 1969. Requirement of a
stable secondary structure for the antiviral activity of poly-nucleotides. Nature (London) 222:1148-1152.
8. De Clercq, E., and T. C. Merigan. 1969. Local and systemicprotection by synthetic polyanionic interferon inducers in
mice against intranasal vesicular stomatitis virus. J. Gen.
Virol. 5:359-368.9. De Clercq, E., and T. C. Merigan. 1969. An active interferon
inducer obtained from Hemophilus inifluentzae type B. J.
Immunol. 103:899-906.10. De Somer, P., and A. Billiau. 1966. Interferon production by
the spleen of rats after intravenous injection of Sindbis
virus or heat-killed Escherichia coli Arch. Gesamte Vi-
rusforsch. 19:143-154.11. De Somer, P., E. De Clercq, and A. Billiau. 1968. Influence
of whole-body irradiation, cortisol treatment and adrenalec-
tomy on interferon induction in vivo in rats, p. 230-243.
Iz M. Sanders and E. H. Lennette (ed.), Medical and
applied virology. W. H. Green, Inc., St. Louis.
12. Dresser, D. W. 1960. Elimination of 13'I-labelled proteinantigens from the circulation of the mouse. Immunology3:289-295.
13. Dresser, D. W. 1961. Effectiveness of lipid and lipidophilicsubstances as adjuvants. Nature (London) 191:1169-1171.
14. Finger, H. 1966. Die Adjuvanswirkung von Mycobakterien.Klin. Wochenschr. 44:1105-111 1.
15. Freund, J. 1951. The effect of paraffin oil and mycobacteria
on antibody formation and sensitization. Amer. J. Clin.Pathol. 21:645-656.
16. Freund, J., K. J. Thomson, H. B. Hough, H. E. Sommer, and
T. M. Pisani. 1948. Antibody formation and sensitizationwith the aid of adjuvants. J. Immunol. 60:383-398.
17. Fruitstone, M. J., B. S. Michaels, D. A. C. Rudloff, andM. M. Sigel. 1966. Role of the spleen in interferon pro-
duction in mice. Proc. Soc. Exp. Biol. Med. 122:1008-1011.18. Glasgow, L. A. 1966. Leukocytes and interferon in the host
response to viral infections. II. Enhanced interferon re-
sponse of leukocytes from immune animals. J. Bacteriol.91:2185-2191.
19. Goldner, H., A. J. Girardi, and M. R. Hilleman. 1965. En-hancement in hamsters of virus oncogenesis attendingvaccination procedures. Virology 27:225-227.
20. Gorhe, D. S. 1967. Inhibition of multiplication of foot andmouth disease virus in adult mice pretreated with Freund'scomplete adjuvant. Nature (London) 216:1242-1244.
21. Green, J. A., S. R. Cooperband, and S. Kibrick. 1969. Im-mune specific induction of interferon production in cul-tures of human blood lymphocytes. Science 164:1415-1417.
22. Ho, M., Y. Kono, and M. K. Breinig. 1965. Tolerance to theinduction of interferons by endotoxins and virus: role of a
humoral factor. Proc. Soc. Exp. Biol. Med. 119:1227-1232.23. Johnson, R. T., and C. A. Mims. 1968. Pathogenesis of viral
infections of the nervous system. N. Engl. J. Med. 278:23-30.
24. Merigan, T. C., and M. S. Finkelstein. 1968. Interferon-stimulating and in vivo antiviral effects of various syn-
thetic anionic polymers. Virology 35:363-374.25. Muramatsu, S. 1964. Shortening of the period of primary
immune response by the prior injection of Freund's adju-vant. Nature (London) 201:1141-1142.
26. Nicol, T., D. C. Quantock, and B. Vernon-Roberts. 1966.Stimulation of phagocytosis in relation to the mechanismof action of adjuvants. Nature (London) 209:1142-1143.
27. Osborn, J. E., and D. L. Walker. 1969. The role of individualspleen cells in the interferon response of the intact mouse.
Proc. Nat. Acad. Sci. U.S.A. 62:1038-1045.28. Postic, B., C. De Angelis, M. K. Breinig, and M. Ho. 1967.
Effect of cortisol and adrenalectomy on induction of inter-feron by endotoxin. Proc. Soc. Exp. Biol. Med. 125:89-92.
29. Rauscher, F. J., M. A. Fink, and J. P. Kvedar. 1963. En-hancement of host response to subliminal doses of Roussarcoma virus by Freund's adjuvant. J. Nat. Cancer Inst.30:645-659.
30. Riggi, S. J., and N. R. Di Luzio. 1961. Identification of a
reticuloendothelial stimulating agent in zymosan. Amer. J.Physiol. 200:297-300.
31. Rodey, G. E., and R. A. Good. 1969. Modification of the invitro response to phytohemagglutinin of mouse spleencells by amyloidogenic agents. Proc. Soc. Exp. Biol. Med.131:457-461.
32. Sabin, A. B., and P. K. Olitsky. 1938. Influence of host fac-tors on neuro-invasiveness of vesicular stomatitis virus. III.Effect of age and pathway of infection on the character andlocalization of lesions in the central nervous system. J. Exp.Med. 67:201-228.
33. Schoenberg, M. D., P. A. Gilman, V. Mumaw, and R. D.
Moore. 1963. Proliferation of the reticuloendothelial sys-tem and phagocytosis. Exp. Molec. Pathol. 2:126-143.
34. Shepel, M., and M. R. Klugerman. 1963. Effect of adjuvantson antibody response of rabbits inoculated with Venezuelanequine encepbalomyelitis virus. J. Bacteriol. 85:1150-1155.
35. Siegel, B. V., and J. I. Morton. 1967. Influence of immuno-logic hyperstimulation on murine viral leukemogenesis.Blood 29:585-593.
36. Subrahmanyan, T. P., and C. A. Mims. 1966. Fate of intra-
venously administered interferon and the distribution ofinterferon during virus infections in mice. Brit. J. Exp.Pathol. 47:168-176.
37. Ter-Grigorov, V. S., and I. S. Irlin. 1968. The stimulating
effect of complete Freund's adjuvant on tumour inductionby polyoma virus in mice and by Rous sarcoma virus inrats. Int. J. Cancer 3:760-764.
38. Van Rossum, W., and P. De Somer. 1966. Some aspects ofthe interferon production in vivo. Life Sci. 5:105-113.
39. White, R. G. 1967. Role of adjuvants in the production ofdelayed hypersensitivity. Brit. Med. Bull. 23:39-44.
40. Youngner, J. S. 1970. Influence of inhibitors of protein syn-
thesis on interferon formation in mice. II. Comparison ofeffects of glutarimide antibiotics and tenuazonic acid.Virology 40:335-343.
41. Youngner, J. S., and W. R. Stinebring. 1965. Interferonappearance stimulated by endotoxin, bacteria, or viruses inmice pre-treated with Escherichia coli endotoxin or infected
42. Youngner, J. S., and W. R. Stinebring. 1966. Comparison ofinterferon production in mice by bacterial endotoxin andstatolon. Virology 29:310-316.
43. Youngner, J. S., and D. S. Feingold. 1967. Interferon pro-
duction in mice by cell wall mutants of Salmonella typhi-murium. J. Virol. 1:1164-1167.
44. Youngner, J. S., and J. V. Hallum. 1968. Interferon produc-tion in mice by double-stranded synthetic polynucleotides:induction or release? Virology 35:177-179.
45. Youngner, J. S., W. R. Stinebring, and S. E. Taube. 1965.Influence of inhibitors of protein synthesis on interferonformation in mice. Virology 27:541-550.