Probiotics, prebiotics, and the host microbiome: the science of translation Article Published Version Creative Commons: Attribution-Noncommercial 3.0 Open Access Petschow, B., Dore, J., Hibberd, P., Dinan, T., Reid, G., Blaser, M., Cani, P. D., Degnan, F. H., Foster, J., Gibson, G., Hutton, J., Klaenhammer, T. R., Ley, R., Nieuwdorp, M., Pot, B., Relman, D., Serazin, A. and Sanders, M. E. (2013) Probiotics, prebiotics, and the host microbiome: the science of translation. Annals of the New York Academy of Sciences, 1306. pp. 1-17. ISSN 1749-6632 doi: https://doi.org/10.1111/nyas.12303 Available at http://centaur.reading.ac.uk/40449/ It is advisable to refer to the publisher’s version if you intend to cite from the work. See Guidance on citing . To link to this article DOI: http://dx.doi.org/10.1111/nyas.12303 Publisher: New York Academy of Sciences All outputs in CentAUR are protected by Intellectual Property Rights law, including copyright law. Copyright and IPR is retained by the creators or other copyright holders. Terms and conditions for use of this material are defined in the End User Agreement .
Transcript
Probiotics, prebiotics, and the host microbiome: the science of translation Article
Published Version
Creative Commons: AttributionNoncommercial 3.0
Open Access
Petschow, B., Dore, J., Hibberd, P., Dinan, T., Reid, G., Blaser, M., Cani, P. D., Degnan, F. H., Foster, J., Gibson, G., Hutton, J., Klaenhammer, T. R., Ley, R., Nieuwdorp, M., Pot, B., Relman, D., Serazin, A. and Sanders, M. E. (2013) Probiotics, prebiotics, and the host microbiome: the science of translation. Annals of the New York Academy of Sciences, 1306. pp. 117. ISSN 17496632 doi: https://doi.org/10.1111/nyas.12303 Available at http://centaur.reading.ac.uk/40449/
It is advisable to refer to the publisher’s version if you intend to cite from the work. See Guidance on citing .
To link to this article DOI: http://dx.doi.org/10.1111/nyas.12303
Publisher: New York Academy of Sciences
All outputs in CentAUR are protected by Intellectual Property Rights law, including copyright law. Copyright and IPR is retained by the creators or other copyright holders. Terms and conditions for use of this material are defined in the End User Agreement .
ANNALS OF THE NEW YORK ACADEMY OF SCIENCESIssue: Annals Reports
Probiotics, prebiotics, and the host microbiome:the science of translation
Bryon Petschow,1 Joel Dore,2 Patricia Hibberd,3 Timothy Dinan,4 Gregor Reid,5 Martin Blaser,6
Patrice D. Cani,7 Fred H. Degnan,8 Jane Foster,9 Glenn Gibson,10 John Hutton,11
Todd R. Klaenhammer,12 Ruth Ley,13 Max Nieuwdorp,14 Bruno Pot,15 David Relman,16
Andrew Serazin,17 and Mary Ellen Sanders18
1Transcend Biomedical Communications, LLC, Youngsville, North Carolina. 2Institut National de la Recherche Agronomique(INRA), Paris, France. 3Harvard Medical School and Massachusetts General Hospital, Boston, Massachusetts. 4UniversityCollege Cork, Cork, Ireland. 5Western University and Lawson Health Research Institute, London, Ontario, Canada. 6New YorkUniversity School of Medicine, New York, New York. 7Universite catholique de Louvain, Louvain Drug Research Institute,WELBIO (Walloon Excellence in Life sciences and BIOtechnology), Brussels,∗ Belgium. 8King & Spalding LLP, Washington,D.C. 9McMaster University, Hamilton, Ontario, Canada. 10University of Reading, Reading, United Kingdom. 11The Universityof York, York, United Kingdom. 12North Carolina State University, Raleigh, North Carolina. 13Cornell University, Ithaca, NewYork. 14University of Amsterdam, Amsterdam, the Netherlands. 15Institut Pasteur de Lille, Lille, France. 16Stanford UniversitySchool of Medicine, Stanford, California. 17Matatu LLC, Washington, D.C. 18Dairy and Food Culture Technologies, Centennial,Colorado
Address for correspondence: Mary Ellen Sanders, Ph.D., Dairy and Food Culture Technologies, 7119 S. Glencoe Ct.,Centennial, CO 80122. [email protected]
Recent advances in our understanding of the community structure and function of the human microbiome haveimplications for the potential role of probiotics and prebiotics in promoting human health. A group of experts recentlymet to review the latest advances in microbiota/microbiome research and discuss the implications for developmentof probiotics and prebiotics, primarily as they relate to effects mediated via the intestine. The goals of the meetingwere to share recent advances in research on the microbiota, microbiome, probiotics, and prebiotics, and to discussthese findings in the contexts of regulatory barriers, evolving healthcare environments, and potential effects on avariety of health topics, including the development of obesity and diabetes; the long-term consequences of exposureto antibiotics early in life to the gastrointestinal (GI) microbiota; lactose intolerance; and the relationship betweenthe GI microbiota and the central nervous system, with implications for depression, cognition, satiety, and mentalhealth for people living in developed and developing countries. This report provides an overview of these discussions.
Keywords: microbiome; probiotics; prebiotics; intestinal microbiota; health disorders
Introduction
Major advances have occurred in our understandingof the composition and metabolic capabilities of mi-crobial communities in the human body, gains madefrom revolutionary advances in DNA sequencing,metagenomic analytical techniques, and computa-tional biology. These strides have greatly increasedour understanding of the bacterial genomes presentin these microbial communities, the boundaries ofnormal variation, and how variations in micro-
[∗Correction added after publication 26 November 2013:an error in this affiliation was amended.]
bial composition are associated with pathology anddisease. Indeed, the number of published studieson microbiome-related research has increased four-fold between 2005 and 2012. Much remains to belearned, however, about how to translate this infor-mation to probiotic or prebiotic interventions thatmay modify the microbiome and promote humanhealth.
A one-day conference, “Probiotics, Prebiotics,and the Host Microbiome: the Science ofTranslation,”1 hosted by the New York Academy ofSciences, the Sackler Institute for Nutrition Science,and the International Scientific Association for Pro-biotics and Prebiotics (ISAPP), including experts in
Probiotics, prebiotics, and the host microbiome Petschow et al.
the fields of microbiome research, probiotics, andprebiotics, met on June 12, 2013 in New York Cityto review the latest research on these topics, dis-cuss the implications for public health, and increasecommunication and collaboration. The conferencewas divided into five sessions and included oral andvisual presentations, as well as a panel discussion.
Putting probiotics, prebiotics, and themicrobiome into translational context
The conference opened with a presentation by JohnHutton (University of York, United Kingdom) onthe economic challenges associated with probiotic-and prebiotic-based interventions. Hutton said thatduring the past 20 years economic evaluation hasbecome a widespread practice in the pharmaceu-tical sector, including cost versus benefit analysesto aid decision making about utilization and reim-bursement of disease treatments.2 In fact, healthtechnology assessment (HTA), including thedemonstration of the cost-effectiveness of newproducts, has become an essential tool for gaugingthe value of drug-based approaches worldwide. Theeconomic evaluation methods used in comprehen-sive HTA consider a number of elements, includingclinical (e.g., efficacy and safety), economic, social(equity of access), and political (incentive for inno-vation) aspects.
The value of nutrition for achieving improvedhealth outcomes has become recognized. Conse-quently, nutritional products are now consideredto be a health technology. However, the economicevaluation methods used in HTA are more diffi-cult to apply in the context of nutrition, wherereduction of disease risk is the most frequent aim.It appears unlikely that HTA methods can be usedto bridge the boundary between drug and food(including probiotics and prebiotics) in the imme-diate future because of the difference in endpointsneeded for supporting data in each category. Addi-tional clarity is needed on the business model foreconomic evaluation of general public and diseaseprevention benefits, compared to disease treatmentclaims. For disease-specific interventions, higher ev-idential standards will be expected in jurisdictionswhere healthcare system funding is sought. For moregeneral interventions to change dietary behavior, aswith many public health policies, the benefits interms of disease-risk reduction may not be realizeduntil well into the future. HTA in this context relies
on modeling and projections of costs and benefits,and is subject to many uncertainties that can only bereduced by the collection of long-term epidemiolog-ical data linking interventions with lifetime healthoutcomes. However, some HTA agencies, such asthe National Institute for Health and Clinical Excel-lence (NICE) in England and Wales, are developingmethods to address the analytical and data issues.
Assessment of the economic feasibility of pro-biotic or prebiotic interventions is further compli-cated by the variability in products, interventionprotocols, local study procedures, populations tar-geted and trial outcomes. These differences mustbe addressed across centers to allow advancementof tools in the domain of probiotics and prebiotics.Going forward, the approach described by White-head et al.3 for assessing the economic viability ofa nutritional intervention for irritable bowel syn-drome (IBS) might serve as a good model for eval-uating probiotic- or prebiotic-based interventions.a
In order for economic evaluation of probiotics andprebiotics as nutritional interventions to succeed,quality data (especially involving disease interven-tions), behavioral changes by individuals, and eco-nomic drivers must become part of the overall healthoutcome process.
Programming the microbiome
Metagenomic studies of the human intestinal mi-crobiome reveal that the human gut carries, onaverage, about 540,000 microbial genes, represent-ing the dominant microbes in this ecosystem.4 Ap-proximately 55% of these genes constitute the coremetagenome (i.e., are genes shared among at least50% of individuals), while many other genes appearto be unique and/or present in less than 20% ofindividuals.
The second conference session, moderated byDavid Mills (University of California, Davis), hadpresentations on the early development of the in-testinal microbiota and how fluctuations in thehuman microbiome can correlate with changes inhuman health. The aim of the session was to pro-vide an overview of the initial programming of the
aThese economic evaluations, however, are useful only forhealth effects that are sufficiently substantiated; for IBS,for example, see Hungin et al., 2013. Alimentary Pharma-cology & Therapeutics. doi:10.1111/apt.12460.
Petschow et al. Probiotics, prebiotics, and the host microbiome
intestinal microbiota and how environmental fac-tors or therapeutics can alter the composition andcontribute to various metabolic disease states.
David Relman (Stanford University School ofMedicine) opened the session with a perspec-tive as an infectious disease specialist and clinicalresearcher interested in variation in microbial di-versity patterns as a function of time (microbialsuccession) and in response to perturbation. Rel-man described the initial acquisition and develop-ment of the gut microbiota during the early weeksof life, and emphasized its significant role in humanhealth and disease. In addition to contributing tofood digestion and nutrition and the regulation ofmetabolism, the gut microbiota is involved in devel-opment and terminal maturation of host mucosa,regulation of immune system target recognition andresponses, and resistance to colonization and in-vasion by pathogens. Humans are born essentiallydevoid of an intestinal microbiota, but this highlyimportant ecological system is soon acquired afterbirth and eventually comprises over 90% of the cellsof the human adult. Despite the advances in toolsand techniques for studying the human gut micro-biome, a number of questions remain about its earlyacquisition and succession.
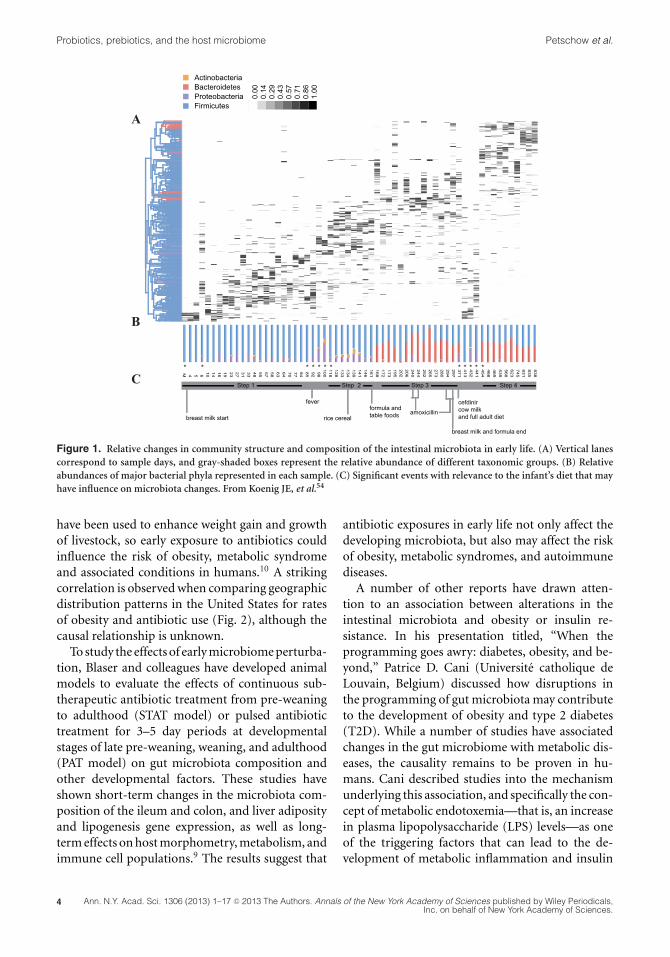
Relman and colleagues study the assembly of thegut, oral, and skin microbiota of infants duringthe first several years of life to better understandthe development of habitat specificity and the fac-tors underpinning compositional variation duringthis critical timespan. Multiple factors appear to in-fluence the early succession of the gut microbiotain individual infants, including mode and timingof birth, host genetics, diet, environmental expo-sures (other humans, animals, medications), andhealth status. Evidence from a number of studiesindicates that the fecal microbiota of the vaginallydelivered newborn resembles that of the maternalgastrointestinal tract or vagina, while that ofcesarean-delivered newborns resembles that of thematernal skin.5,6 Various taxonomic groups are ac-quired early during a window of permissivity andpersist over time, possibly undergoing shifts in pop-ulation from dominant to subdominant, while othertaxonomic groups may be acquired later or dis-appear altogether (Fig. 1). Recent studies usingcultivation-independent evaluation of the fecal mi-crobiota of premature infants have revealed a lowlevel of diversity, high inter-individual variability,
and a capacity for abrupt temporal shifts in species-and strain-level composition. The long-term effectsof these differences are not fully understood. A keyemerging question in the field is how these earlylife differences in microbiota development influencehealth through childhood and later in life.
Evaluation of the compositional structure of theinfant gut microbiota over time indicates that eco-logical states persist for days or weeks, followed byabrupt transitions to alternative states. These dis-tinctive, successive equilibrium states are observedin both preterm and term babies, although the com-position of their communities differ significantlyat the earliest days of life. Periods of stable phy-logenetic composition at the taxonomic level ofgenus and species may belie underlying shifts inthe abundances of strains with different functionalpotential.7 Bifidobacteria eventually appear in mostbabies, typically detected between 1 and 2 monthsof life. In general, members of enterobacteria andbifidobacteria appear early on, Bacteroides spp. atabout the middle of the first year, and butyrate-producing Faecalibacterium and Roseburia towardthe end of the first year of life. Considerable intra-and interpersonal variation in fecal bacterial com-munity structures is observed during the first yearof life, and intrapersonal variation decreases as afunction of age.5 The emergence of an adult-likemicrobiota pattern occurs by the third year.8 Whilechanges in diet appear to be associated with majorshifts in the structure of the infant microbial com-munities, the relative role of other variables (e.g.,medications, nutrition, phages, parasitic burden) isnot well understood.
Martin Blaser (New York University School ofMedicine) is leading a research effort on the de-velopmental pathway for the microbiome duringearly life, exploring how perturbations may in-fluence later risk of various disorders.9 Duringhis presentation, “Impact of antibiotic exposureson the developing microbiota,” Blaser remindedthe audience that medical interventions can affectthe composition of the microbiota, with poten-tial effects on metabolic and immunologic devel-opment. Examples include antibiotics administeredto mothers or infants, cesarean delivery, and use ofbasic infant formula. The collective effects of theseexposures, over time and following transmissionfrom exposed mothers to infants, may reduce thediversity of the gut microbiota. Low-dose antibiotics
Probiotics, prebiotics, and the host microbiome Petschow et al.
A
B
C
Figure 1. Relative changes in community structure and composition of the intestinal microbiota in early life. (A) Vertical lanescorrespond to sample days, and gray-shaded boxes represent the relative abundance of different taxonomic groups. (B) Relativeabundances of major bacterial phyla represented in each sample. (C) Significant events with relevance to the infant’s diet that mayhave influence on microbiota changes. From Koenig JE, et al.54
have been used to enhance weight gain and growthof livestock, so early exposure to antibiotics couldinfluence the risk of obesity, metabolic syndromeand associated conditions in humans.10 A strikingcorrelation is observed when comparing geographicdistribution patterns in the United States for ratesof obesity and antibiotic use (Fig. 2), although thecausal relationship is unknown.
To study the effects of early microbiome perturba-tion, Blaser and colleagues have developed animalmodels to evaluate the effects of continuous sub-therapeutic antibiotic treatment from pre-weaningto adulthood (STAT model) or pulsed antibiotictreatment for 3–5 day periods at developmentalstages of late pre-weaning, weaning, and adulthood(PAT model) on gut microbiota composition andother developmental factors. These studies haveshown short-term changes in the microbiota com-position of the ileum and colon, and liver adiposityand lipogenesis gene expression, as well as long-term effects on host morphometry, metabolism, andimmune cell populations.9 The results suggest that
antibiotic exposures in early life not only affect thedeveloping microbiota, but also may affect the riskof obesity, metabolic syndromes, and autoimmunediseases.
A number of other reports have drawn atten-tion to an association between alterations in theintestinal microbiota and obesity or insulin re-sistance. In his presentation titled, “When theprogramming goes awry: diabetes, obesity, and be-yond,” Patrice D. Cani (Universite catholique deLouvain, Belgium) discussed how disruptions inthe programming of gut microbiota may contributeto the development of obesity and type 2 diabetes(T2D). While a number of studies have associatedchanges in the gut microbiome with metabolic dis-eases, the causality remains to be proven in hu-mans. Cani described studies into the mechanismunderlying this association, and specifically the con-cept of metabolic endotoxemia—that is, an increasein plasma lipopolysaccharide (LPS) levels—as oneof the triggering factors that can lead to the de-velopment of metabolic inflammation and insulin
Petschow et al. Probiotics, prebiotics, and the host microbiome
Figure 2. Comparisons between the geographical distribution of obesity (left) and antibiotic use in the United States, 2010(right). Obesity trend data were from the CDC Behavioral Risk Factor Surveillance System (http://www.cdc.gov/brfss/). Antibioticprescriptions were from a national survey of out-patient pharmacy records, as described by Hicks, et al.55
resistance associated with obesity.11 High-fat di-ets have been shown to alter the gut microbiotacomposition. Cani and colleagues have previouslydemonstrated that mice treated with antibiotics areresistant to diet-induced metabolic endotoxemia,fat mass development, metabolic inflammation andinsulin resistance.12 Similarly, several studies havedemonstrated that germ-free (GF) mice are pro-tected against glucose intolerance, metabolic in-flammation, and insulin resistance, which suggeststhat the microbiota may be involved.13,14 The mi-crobiota of high-fat–fed mice or obese (ob/ob) mu-tant mice can also transfer the obesity/T2D pheno-type to ex-GF mice.13,15 Other studies have shownthat antibiotic treatment protects mice from thisdiet-induced obesity,12,16 while prebiotics lead toreduced body-weight gain and fat deposition andprotect against hepatic steatohepatitis in obese andT2D rats.17 Cani and colleagues have shown thatnutritional or genetic-induced obesity and type 2diabetic rodents display gut barrier dysfunctionsleading to the leakage of LPS and possibly othermicrobiota-derived factors.12 They found that gutmicrobiota metabolites can interact with the en-docannabinoid system,18 as well as with enteroen-docrine L cells to alter gut permeability (Fig. 3),possibly through production of GLP-1, PYY, andGLP-2.19
Additional studies have used prebiotics and pro-biotics to identify novel mechanisms of bacterial
interaction with the host that control gut perme-ability and metabolism during obesity and T2D.They showed that levels of Akkermansia muciniphilaare decreased in the gut microbiota of mice fedhigh-fat diets and in ob/ob mice, while feeding themice prebiotics (inulin-type fructans) restores thesepopulations.20 Transfer of A. muciniphila to high-fat diet–induced obese mice led to decreased fatmass gain, increased fat oxidation, and restoredgut barrier function in the colon. Viable, but notheat-killed A. muciniphila, increased mucus layerthickness, decreased plasma LPS levels, and coun-teracted the diet-induced metabolic disturbances.20
These studies demonstrate that the gut microbiotainfluences energy homeostasis; that bacterial com-pounds contribute to low-grade inflammation andgut permeability in obesity and T2D; and that cer-tain types of bacteria within the gut microbiota, suchas A. muciniphila, may play a role in preventing theseobesity-related conditions.
Another perspective on the role of the gut mi-crobiome in the development of metabolic disor-ders was presented by Max Nieuwdorp (Univer-sity of Amsterdam, the Netherlands). Nieuwdorpand collaborators have been investigating the roleof the gut microbiota in health and disease us-ing fecal transplantation. While fecal transplanta-tion has been practiced for centuries, since 1958only 500 case reports exist on treatment of patientswith recurrent Clostridium difficile infection, IBS,
Probiotics, prebiotics, and the host microbiome Petschow et al.
Figure 3. Interactions between gut microbiota and the endocannabinoid system: impact on gut barrier function and metabolicinflammation. Obesity (nutritional or genetic) is associated with changes in the gut microbiota composition and pathophysiologicalchanges, whereby the endocannabinoid system tone is altered. This phenomenon is associated with the development of gut per-meability, metabolic endotoxemia, metabolic inflammation, and altered adipose tissue metabolism (adipogenesis). From DelzenneNM, et al.56
inflammatory bowel disease (IBD), or multiplesclerosis.22 Modern procedures involve bowel lavagefor 4–6 hours followed by either colonoscopy orduodenal infusion of fecal homogenates preparedfrom healthy screened donors (following a question-naire on bowel habits, travel history, medications,and screened for an extensive list of fecal and blood-borne pathogens). A recent study by Van Noodet al.21 showed a 92% cure rate, with an increase inmicrobiota diversity for over 6 months, in patientswith recurrent C. difficile infection.
Nieuwdorp and collaborators then conducted arandomized controlled trial in obese subjects to in-vestigate the effects of fecal transplantation on in-sulin resistance using hyperinsulinemic euglycemicclamp techniques and evaluation of gut microbiotacomposition.22 Subjects were randomized to receivefecal microbiota (FM) from homogenates of theirown feces (autologic) or from healthy, lean donors(allogenic). Results showed a significant improve-ment in insulin sensitivity in subjects receiving al-logenic FM lasting for up to 6 weeks, but no changein those receiving autologic FM. Infusion of allo-genic FM also led to changes in the compositionof the fecal and small intestinal microbiota (mostlybutyrate producers) and a reduction in fecal short
chain fatty acids (SCFA) butyrate and propionate.No change was observed in food intake or weightamong groups; this is interesting, as obesity can re-sult from increased food intake as well as alterednutrient content. A more striking improvementin insulin sensitivity was observed in one subjectfollowing FM transplant from a specific allogenicdonor. This result correlated with higher levels ofEubacterium hallii (anaerobic Gram-positive Firmi-cute) in the small intestine. Recognizing that otherstudies have shown that certain groups within thegut microbiota may have diagnostic and clinicalvalue in predicting T2D in obese patients (e.g.,Roseburia species, Faecalibacterium prausnitzii,Lactobacillus gasseri),23 Nieuwdorp et al. have sinceinvestigated E. hallii and generated data suggestingthat four weeks of daily gavage with culturedE. hallii in male db/db mice is safe and has beneficialeffects on glucose metabolism, most likely throughaltered fecal SCFA production.24 Additional studiesare planned to evaluate the safety and potential ben-efits of E. hallii for improving insulin sensitivity inhumans.
Collectively, these studies suggest that the gut mi-crobiota per se and certain bacterial products inparticular, play a role in the development of obesity
Petschow et al. Probiotics, prebiotics, and the host microbiome
and changes in insulin sensitivity. Future researchneeds to confirm causality in humans and clinicalrelevance with respect to gut microbiota, as well asexplore mechanisms of action and use of probioticsor prebiotics.
Translating research to transform healthcare
Public health is confronted with complex systemsthat have outcomes with multiple determinants thatinteract in obscure ways. As a result, changes to pub-lic health systems often lead to unintended conse-quences. The third session, moderated by GregorReid (Western University and Lawson Health Re-search Institute, Canada), consisted of a keynotepresentation by Harry Burns (Chief Medical Officerfor Health, Scotland) and a panel discussion thatconsidered potential pathways and issues involvedwith translating research advances to influence pub-lic health policy. The goals of the session were tounderstand the factors involved with influencing ahealthcare system—at the country, state/province,or local community level—and discuss how best toapply this knowledge in transforming research find-ings on probiotics and (to a lesser extent) prebioticsto change public health policy.
In his keynote lecture, Burns provided examplesof how a quality improvement model under his lead-ership led to significant reductions in hospital in-fection rates (e.g., ∼90% reduction in C. difficile),hospital mortality rates, infant mortality rates, andimproved clinical record keeping. A key reasonstated for the success of this model was the involve-ment and complete buy-in from hospital personneland other key constituents. One example of this col-laboration is the Early Years Collaborative (EYC),25
whose objective is to support and drive practical ac-tion under a broader partnership program aimed atdelivering a shared commitment to give children thebest start in life and to improve the life chances ofchildren, young people, and families at risk. The EYCeffort aims to (1) deliver measurable improvementin outcomes and reduce inequalities for Scotland’svulnerable children; (2) put Scotland on course toshift the balance of public services toward early in-tervention and prevention by 2016; and (3) sustainthis change to 2018 and beyond. The strategy usedfor implementing such changes involves the follow-ing actions: understand the problem, build the will
for change, execute the change, and drive the changewith data.
Their main tactical approach is to design an inter-vention based upon sound rationale (whether med-ical, scientific, or social), test it in a real situation,measure and modify it, then continue until the im-plementation is optimized. By building the will ofall the change makers, objectives can be met andtangible improvements achieved.
Panel discussion
In translating these learnings to probiotics, Burnsand an expert panel, including Rowena Pullan(Pfizer Consumer Health Care), Bruno Pot (In-stitut Pasteur de Lille), and David Mills, restatedthe importance of public–private partnerships todrive public policy change. And while the strengthof evidence for probiotics is clear in a number ofapplications26 (such as treating necrotizing entero-colitis in premature infants,27 preventing antibiotic-associated diarrhea,28 improving urogenital healthin women,29 and countering infection and allergyrelated to respiratory health30,31), these data need tobe collated, presented to changemakers, and used asa means to shift medical practice. This is currentlya major challenge in the United States, where anyproduct (including yogurt) being tested to treat,prevent, or cure disease must be investigated as adrug, while fewer barriers exist in Canada and othercountries.
General concern was expressed by the audienceabout the regulatory view of probiotic food researchand how it can be used to substantiate market ap-proval and claims. The regulatory approval pathfor probiotic foods is complicated, particularly go-ing through the FDA. Pot responded to questionsabout the situation with probiotics at the EuropeanFood Safety Authority (EFSA). He explained thatthe EFSA position on probiotics is inconsistent withprecedent established by other dietary substances.For example, the data on the health benefits of pro-biotics are more extensive and of higher quality (i.e.,evaluating nonessential endpoints) than data formost vitamins and minerals as health supplementsthat have been accepted by the EFSA. Yet, no probi-otic claims have been approved by this governmentagency. Regulatory barriers now exist in the Euro-pean Union to use of the word probiotic on foods. Itis seen as an unapproved health claim, and as such is
Probiotics, prebiotics, and the host microbiome Petschow et al.
not allowed despite mounting scientific evidence ofbeneficial effects. There are clear challenges with theregulatory framework in the United States and theEuropean Union, but those involved with probioticsand prebiotics must continue to do quality researchand communicate the outcomes of that science inmore efficient and effective ways.
A large gap currently exists in frontline healthcare providers’ understanding of probiotics, pre-biotics and the human microbiome, and certainlyon how to interpret the vast data sets from omicsstudies. The benefits of advances in microbiome re-search and probiotics will only be realized whennew technologies from omics are integrated intomedical diagnostics, and when medical students,physicians, and administrators are educated on thehealth benefits and cost-savings potential from pro-biotics. Clear messages need to be crafted and di-rected at key target audiences, such as consumers,politicians, doctors, pharmacists, and others. Onenew initiative, Gut Microbiota for Health,32 aims tobridge this information gap with clinicians throughannual meetings and an interactive website with ba-sic information on probiotics/prebiotics and the gutmicrobiome. Lessons might be learned from similarinitiatives in the clinical field, such as the EuropeanPathoNGenTrace project, which aims to extract use-ful functional (clinically relevant) information fromthe whole genome sequence.b
David Relman inquired about the cost to collectthe data needed for regulatory approval. Cost sav-ings in health care are difficult to evaluate. Burnspointed out that although governments value cost-saving interventions, changes are often adopted sim-ply because they improve well-being and they are theright thing to do. Burns felt that the public is notimpressed by cost-effectiveness.
Impact of gut bacteria on braindevelopment, circuitry, and behavior
While many general press articles on gut microbesdiscuss the apparent link to obesity, there has alsobeen a marked increase in discussions of gut–brain interactions. Jane Foster (McMaster Univer-sity, Canada) has been using conventional and GFmice to study the gut–brain axis, which involves thecomplex interplay between the autonomic and en-
bSee http://www.patho-ngen-trace.eu/
teric nervous systems, pituitary and gut hormones,and the gut inflammatory systems (Fig. 4). Herresearch uses a well-established behavioral test—the elevated plus maze (EPM)—to examine ex-ploratory behavior and is used to assess anxiety lev-els. Typically, conventional mice spend most timein closed-arm areas (harm avoidance) of the EPMcompared to open-arm areas. Extending from ear-lier studies observing that GF mice show enhancedstress-reactivity,33 Foster et al. found that GF micedemonstrated reduced anxiety-like behavior in theEPM (more time in open arms, both in durationand numbers of visits), compared with specificpathogen–free (SPF) mice.34,35 The reduced anxietybehavior persisted when GF mice were colonizedwith SPF microflora during adulthood (ex-GF), butnot when colonized at an earlier age, demonstratingthat gut–brain interactions influence central ner-vous system wiring early in life. The low anxiety–likephenotype was accompanied by long-term changesin plasticity-related genes in the hippocampus andamygdala.
The researchers have also been examining theinterplay of leptin and central circuits for stress-reactivity and feeding in the presence or absenceof the gut microbiota. They found that leptin-insufficiency in GF mice leads to long-term changesin the expression levels of the brain’s leptin recep-tors, as well as certain peptides such as neuropep-tide Y (NPY), which influence corticosterone levelsand thus plays pivotal roles in the stress response.The response of different strains of mice and therole of serotonin or other feedback mechanisms areunclear. While many questions remain, continuedwork in this exciting area may shed light on howdevelopmental factors and the intestinal microbiotainfluence the interface between metabolic diseasesand mood disorders.
Gary Frost (Imperial College London, UnitedKingdom) reported on another area of study regard-ing the effects of gut microbiota on brain activity,namely, the effect of dietary prebiotics and asso-ciated production of SCFAs such as acetate, pro-pionate, and butyrate on hypothalamic neuronalactivity and obesity. It is widely believed that the in-creasing incidence of obesity is associated with lowconsumption of fermentable fibers and high intakeof dietary carbohydrates. Frost and colleagues are in-vestigating the mechanisms involved in the decreasein body weight and improved insulin sensitivity that
Petschow et al. Probiotics, prebiotics, and the host microbiome
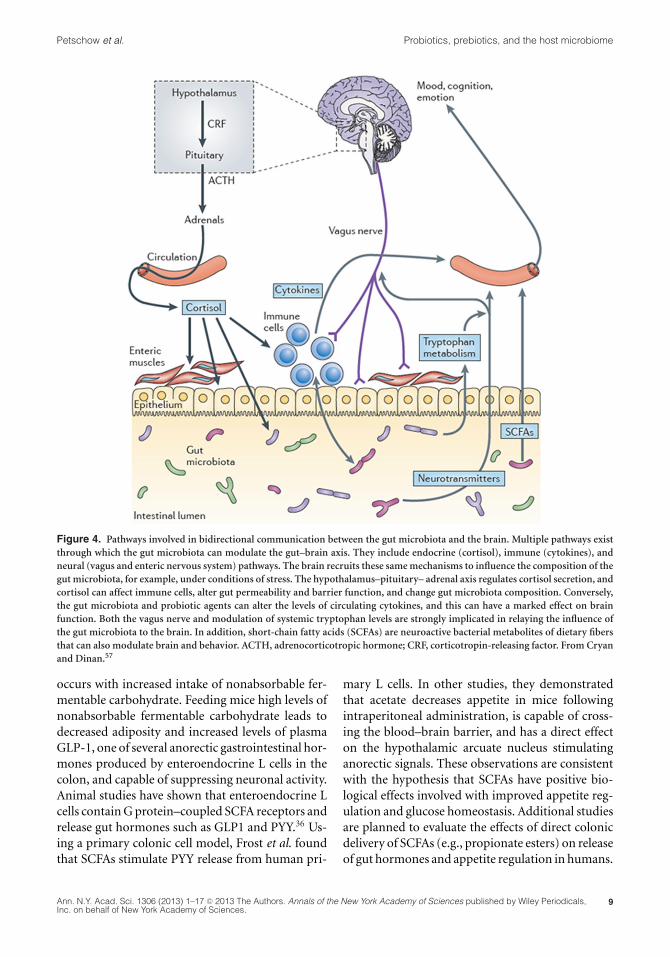
Figure 4. Pathways involved in bidirectional communication between the gut microbiota and the brain. Multiple pathways existthrough which the gut microbiota can modulate the gut–brain axis. They include endocrine (cortisol), immune (cytokines), andneural (vagus and enteric nervous system) pathways. The brain recruits these same mechanisms to influence the composition of thegut microbiota, for example, under conditions of stress. The hypothalamus–pituitary– adrenal axis regulates cortisol secretion, andcortisol can affect immune cells, alter gut permeability and barrier function, and change gut microbiota composition. Conversely,the gut microbiota and probiotic agents can alter the levels of circulating cytokines, and this can have a marked effect on brainfunction. Both the vagus nerve and modulation of systemic tryptophan levels are strongly implicated in relaying the influence ofthe gut microbiota to the brain. In addition, short-chain fatty acids (SCFAs) are neuroactive bacterial metabolites of dietary fibersthat can also modulate brain and behavior. ACTH, adrenocorticotropic hormone; CRF, corticotropin-releasing factor. From Cryanand Dinan.57
occurs with increased intake of nonabsorbable fer-mentable carbohydrate. Feeding mice high levels ofnonabsorbable fermentable carbohydrate leads todecreased adiposity and increased levels of plasmaGLP-1, one of several anorectic gastrointestinal hor-mones produced by enteroendocrine L cells in thecolon, and capable of suppressing neuronal activity.Animal studies have shown that enteroendocrine Lcells contain G protein–coupled SCFA receptors andrelease gut hormones such as GLP1 and PYY.36 Us-ing a primary colonic cell model, Frost et al. foundthat SCFAs stimulate PYY release from human pri-
mary L cells. In other studies, they demonstratedthat acetate decreases appetite in mice followingintraperitoneal administration, is capable of cross-ing the blood–brain barrier, and has a direct effecton the hypothalamic arcuate nucleus stimulatinganorectic signals. These observations are consistentwith the hypothesis that SCFAs have positive bio-logical effects involved with improved appetite reg-ulation and glucose homeostasis. Additional studiesare planned to evaluate the effects of direct colonicdelivery of SCFAs (e.g., propionate esters) on releaseof gut hormones and appetite regulation in humans.
Probiotics, prebiotics, and the host microbiome Petschow et al.
Helen E. Raybould (University of California,Davis) described her group’s work on the gut–brainaxis as it relates to regulation of GI function and foodintake. Luminal chemosensors found on enteroen-docrine cells that line the intestine transmit signalsto the central nervous system (CNS) via the vagal af-ferent neurons, which in turn influence GI functionthrough parasympathetic pathways and eating be-havior through effects on higher brain centers. Stud-ies were designed to evaluate changes in microbiotaand gut epithelial function that may be connectedwith hormonal pathways. These pathways, which in-fluence food intake and obesity, were assessed usingSprague-Dawley rats fed a high-fat diet to induce ei-ther an obesity-prone (DIO-P) or obesity-resistant(DIO-R) phenotype.37 Using bacterial 16S rRNAmeasurements, they showed a decrease in total bac-terial density and an increase in the relative propor-tion of Clostridiales in high-fat–fed rats regardlessof phenotype, while an increase in Enterobacterialeswas only seen in the microbiota of DIO-P rats. TheDIO-P group also exhibited increases in intestinalpermeability, which resulted in elevated plasma LPSlevels, and Toll-like receptor 4 activation. The datasuggest that DIO-P rats are unable to transmit sig-nals to the brain that involve cholecystokinin (CCK)regulation of vagal afferent neuron transmissionsto communicate satiety in response to food intake.Further studies show that, compared with a normaldiet, paracellular permeability, transcellular perme-ability, and mucosal inflammation are increased byfeeding a Western diet and these changes can beprevented by feeding Bifidobacterium infantis andprebiotic bovine milk oligosaccharides (MOs). Col-lectively, these results suggest that consumption of ahigh-fat diet may induce changes in the gut micro-biota and increase low-grade inflammation that ul-timately contributes to the development of diabetesand obesity. Additional investigations are expectedto clarify how specific prebiotic/probiotic combina-tions may modulate gut function and inflammatoryresponses to delay the onset of, or even prevent,diabetes and obesity.
Reaching people in need with probioticsand prebiotics
The final session, moderated by Ruth Ley (CornellUniversity), consisted of a series of short presenta-tions with a focus on translating scientific innova-tions in pro- and prebiotics to reach the people with
the greatest needs. Andrew Serazin (Matatu, LLC)set the stage by presenting the challenges involvedin these activities. Studies into the microbiome areone of the most active areas of the life sciences today,according to Serrazin, since the isolation of restric-tion enzymes nearly forty years ago. Shifting con-sumer preferences and dietary patterns, at least inthe United States, underlie drastic changes in con-sumption of major nutrient classes with concurrentrises in chronic disease. Escalating healthcare costsand demands of an aging population have led toa growing preference for self-treatment or preven-tion of disease including wellness approaches. To besuccessful in developing widely distributed probi-otic and prebiotic products, the surge of scientificinquiry into the structure and function of the micro-biome must be matched by a focused and transpar-ent effort to engage industry, health policy makers,and the general public.
Extending such products to people in need inemerging economies will present a whole new set ofchallenges. Most of the world’s 7 billion people arecurrently experiencing significant alterations in dis-ease burdens, dietary patterns, and lifestyle. This hasbeen dramatically documented in countries such asIndia and South Africa, where it is common to findhigh levels of both malnutrition-induced stuntingand obesity in the same population, which is indica-tive of nutritional deficits that manifest in oppositeforms.
Products derived from advances in our under-standing of the microbiome are part of an entirelynew field at the union of nutrition and medicine,and their applications are likely to be profound inmeeting future challenges in food and nutrition.Successful translation of advances made in micro-biome research to probiotic and prebiotic productswill require the following: (1) an open, engaged,and realistic research community with clear goals,including sharing of potential benefits throughcommitment to global access; (2) recognition that abroad number of foods and ingredients shape mi-crobiome structure and function and, in turn, canaffect the health of consumers; (3) regulation basedon meaningful biomarkers and defined outcomes;and (4) trusted products with a clear health benefitto consumers.
Gregor Reid partnered with Patricia Hibberd(Harvard Medical School and Massachusetts Gen-eral Hospital) in a presentation titled, “From yogurt
Petschow et al. Probiotics, prebiotics, and the host microbiome
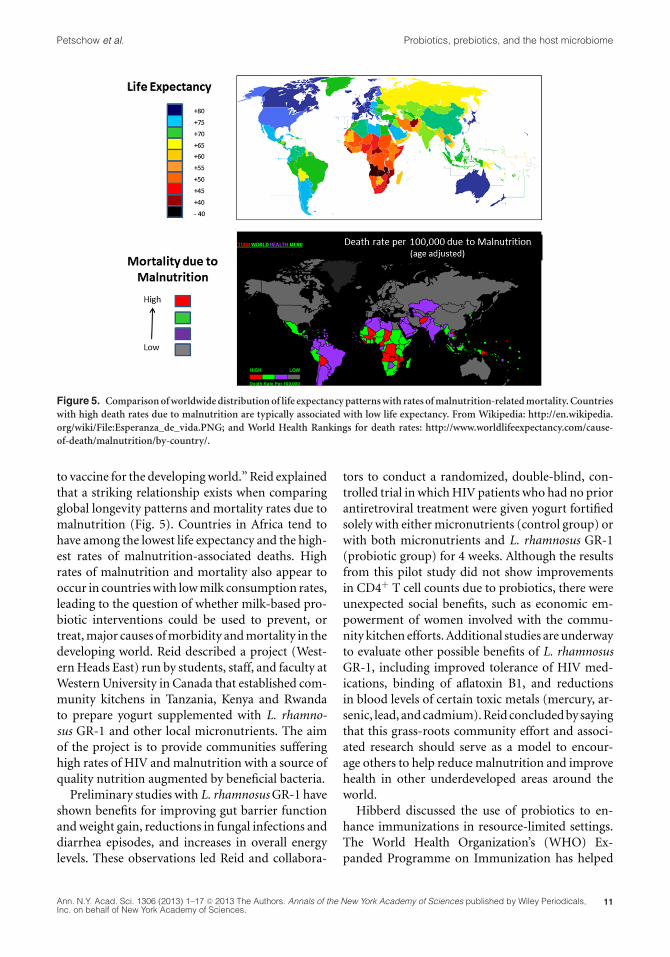
Figure 5. Comparison of worldwide distribution of life expectancy patterns with rates of malnutrition-related mortality. Countrieswith high death rates due to malnutrition are typically associated with low life expectancy. From Wikipedia: http://en.wikipedia.org/wiki/File:Esperanza_de_vida.PNG; and World Health Rankings for death rates: http://www.worldlifeexpectancy.com/cause-of-death/malnutrition/by-country/.
to vaccine for the developing world.” Reid explainedthat a striking relationship exists when comparingglobal longevity patterns and mortality rates due tomalnutrition (Fig. 5). Countries in Africa tend tohave among the lowest life expectancy and the high-est rates of malnutrition-associated deaths. Highrates of malnutrition and mortality also appear tooccur in countries with low milk consumption rates,leading to the question of whether milk-based pro-biotic interventions could be used to prevent, ortreat, major causes of morbidity and mortality in thedeveloping world. Reid described a project (West-ern Heads East) run by students, staff, and faculty atWestern University in Canada that established com-munity kitchens in Tanzania, Kenya and Rwandato prepare yogurt supplemented with L. rhamno-sus GR-1 and other local micronutrients. The aimof the project is to provide communities sufferinghigh rates of HIV and malnutrition with a source ofquality nutrition augmented by beneficial bacteria.
Preliminary studies with L. rhamnosus GR-1 haveshown benefits for improving gut barrier functionand weight gain, reductions in fungal infections anddiarrhea episodes, and increases in overall energylevels. These observations led Reid and collabora-
tors to conduct a randomized, double-blind, con-trolled trial in which HIV patients who had no priorantiretroviral treatment were given yogurt fortifiedsolely with either micronutrients (control group) orwith both micronutrients and L. rhamnosus GR-1(probiotic group) for 4 weeks. Although the resultsfrom this pilot study did not show improvementsin CD4+ T cell counts due to probiotics, there wereunexpected social benefits, such as economic em-powerment of women involved with the commu-nity kitchen efforts. Additional studies are underwayto evaluate other possible benefits of L. rhamnosusGR-1, including improved tolerance of HIV med-ications, binding of aflatoxin B1, and reductionsin blood levels of certain toxic metals (mercury, ar-senic, lead, and cadmium). Reid concluded by sayingthat this grass-roots community effort and associ-ated research should serve as a model to encour-age others to help reduce malnutrition and improvehealth in other underdeveloped areas around theworld.
Hibberd discussed the use of probiotics to en-hance immunizations in resource-limited settings.The World Health Organization’s (WHO) Ex-panded Programme on Immunization has helped
Probiotics, prebiotics, and the host microbiome Petschow et al.
increase childhood immunizations against diseasessuch as polio, diphtheria, pertussis, measles, andtetanus. While such efforts have saved an estimated20 million children’s lives over the last 20 years, in2011 over 23 million children received no immu-nizations at all. A key reason for this is that gapsexist in the cold-chain distribution systems neededto preserve the potency of the vaccines. Hibberddescribed her and collaborators’ efforts to createvaccine-delivery vehicles by genetically engineeringthe probiotic bacterium Bacillus subtilis to displayvaccine antigens; B. subtilis was chosen because itcan withstand extreme environmental conditions.Hibberd and colleagues have created non-injectable,thermo-stable vaccines for tetanus and rotavirus,with plans to extend to pertussis, diphtheria, andother major causes of childhood diseases. The engi-neered vaccines are stable at 45 C without refrig-eration for more than 1 year, and have been shownin animal safety and immunogenicity studies to besafe and capable of producing protective levels ofantibodies when administered intranasally, sublin-gually, or transdermally.38,39 Plans are underway toconduct testing of the B. subtilis vaccine platformin humans under an IND, which could eventuallylead to a viable approach for providing childhoodvaccines that do not require the cold chain, needles,or administration by skilled personnel, in resource-limited settings.
Lactose intolerance continues to be a problemfor over 40 million people in the United States.Research by Dennis Savaiano (Purdue University),Andrea Azcaarate-Peril (UNC Microbiome Center),and Todd Klaenhammer (North Carolina State Uni-versity) is evaluating the clinical effects of feed-ing a highly purified, short-chain galactooligosac-charide (GOS/RP-G28) on lactose intolerance andchanges in the composition of the colonic micro-biota (using terminal restriction fragment lengthpolymorphisms (TRFLP) and 16S rRNA pyrose-quencing). In a randomized, double-blinded study,they fed GOS/RP-G28 to lactose-intolerant adultsfor 36 days and collected stool samples at the startof the study (day 0), after GOS treatment (day 36),and 30 days after GOS feeding was stopped (day 66);consumption of dairy products was encouraged inboth placebo and intervention groups after day 36.Lactose digestion and overall symptoms of lactoseintolerance improved in subjects fed GOS/RP-G28compared to a placebo group; subjects on GOS were
six times more likely to claim they were lactose toler-ant post-treatment. When compared to the placebogroup, subjects fed GOS/RP-G28 showed only mi-nor changes in microbiota composition on day 36,but statistically significant major shifts in the micro-biota occurred at day 66. Changes in the microbiotaby day 66 included increased abundance of Fae-calibacterium prausnitzii, Lactobacillales, and Rose-buria spp. within the Firmicutes phyla; increases in(Lac+) Oscillibacter and Dorea spp.; and a reductionin some Clostridia class members. Expanded stud-ies are planned to confirm these changes in the fe-cal microbiota of lactose-intolerant individuals thatwere clinically responsive to dietary adaptation toGOS/RP-G28.
Joel Dore (Institut National de la RechercheAgronomique, France) provided an excellentoverview of research and clinical studies on theanti-inflammatory properties of F. prausnitzii, anddescribed studies conducted by his group aimedat evaluating the mechanisms of crosstalk betweenF. prausnitzii and host cells that may underlie itsrole as a mutualistic commensal. He set the stage bypointing out that research studies conducted duringthe past decade have demonstrated an associationbetween certain chronic immune diseases and dys-biosis of the intestinal microbiota40–42 A recurrenttheme in many of these studies is the observationthat such chronic immune disorders are associatedwith the presence of low-grade inflammation on thehost side and a reduction of some important antimi-crobial commensal species on the microbiome side.Most likely, other factors such as diet, genetic predis-position, environment, and lifestyle also contributeto the low-grade inflammatory state and changes inmicrobiota composition (Fig. ??). F. prausnitziii isone example of a potentially beneficial intestinalcommensal, based on anti-inflammatory proper-ties demonstrated in preclinical studies.41 Levels ofF. prausnitzii are found in low abundance in patientswith Crohn’s disease,40 colorectal cancer, obesity,43
or IBS.44 In one study, administration of F. praus-nitzii provided protection from endoscopic inflam-mation relapse 6 months after surgery in 20 patientswith active CD requiring ileocecal resection.41
Dore and colleagues are also investigatingbacteria–cell crosstalk using a functional metage-nomics approach to better understand how al-tered intestinal ecology may contribute to a chronicimmune condition. Human cell lines have been
Petschow et al. Probiotics, prebiotics, and the host microbiome
Figure 6. Alterations of the gut microbiota and low-grade inflammation may contribute to a cycle of events that induces a chronicstate in immune-mediated diseases. Interventions that target the combined modulation of gut microbiota and inflammation maybe the most effective way to manage such conditions.
engineered with stably transfected reporter genes,allowing them to assess modulation of transcrip-tion regulators such as NF-B, AP-1, or PPAR- , orproduction of proteins such as TSLP, TGF-, or Fiaf.High-throughput screening of interactions betweenover 20,000 metagenomic clones bearing large ge-nomic inserts of culturable and noncultured bacte-ria and human cells have allowed the identificationof several bioactive clones that modulate cellular ac-tivities with relevance to immune response, prolifer-ation, or metabolism. The genes involved are iden-tified and relevant bioactive signal molecules areidentified using biochemical or genetic approaches.Results from such studies may help unravel mecha-nisms by which commensal bacteria modulate cellu-lar functions, which may lead to exploration of waysto favorably modulate probiotic–host interactions.
In the closing presentation, Fred Degnan (King &Spalding, LLP) discussed the perspectives of the U.S.FDA on clinical study requirements as it relates toregulatory classifications for probiotic-containingproducts. A probiotic product can be classified indifferent regulatory categories depending on theproduct’s intended use, including drug, food, medi-cal food, food additive, or dietary supplement. Thereare implications for each classification in relation tothe nature and degree of regulatory requirementsand, ultimately, for claim substantiation and marketaccess.
As a general rule, the FDA determines the degreeof regulation for clinical trials and assigns productclassifications on the basis of the intended use for a
given product. The intended use of a product canbe determined by a number of factors, includingclaims, labeling, promotions, and by endpoints ofclinical investigation. Based primarily on these typesof communications, the product will be deemed adrug, food, dietary supplement, or medical food.The various claim structure for these different prod-ucts can be summarized as follows:
Drug/Biological product: Focus is on the cure,treatment, mitigation, or prevention of dis-ease, although these products can also affectthe structure or function of the human body(biological products are drugs derived fromlive microorganisms).
Foods or dietary supplements: Health or “qualified” health claim: Char-
acterizes the relationship between a nutri-ent, dietary supplement, or food, and thereduction in risk of a disease or health-related condition. An approved health claimmust be supported by “significant scientificagreement,” while “qualified” health claimsare based on “emerging” scientific evidence.Both must be pre-reviewed by the FDA viaa petition process or on the basis of state-ments by an authoritative body.
Structure/function claim: Describes the ef-fect of a food, food component, nutrient ordietary ingredient on the “structure or anyfunction of the body.” May not imply or
Probiotics, prebiotics, and the host microbiome Petschow et al.
express usefulness in the cure, mitigation,treatment or prevention of disease.
Medical food: For a patient under medical su-pervision for the “dietary management of adisease or condition for which distinctive nu-tritional requirements have been establishedby medical evaluation” and which cannot beaddressed by the diet alone.
A clinical study involving an FDA-regulated arti-cle can evoke certain requirements. If a company orindividual wishes to conduct a human study on aprobiotic intended for prevention or treatment of adisease, they will be required to follow a more rigor-ous regulatory pathway. Typically, this will requirefiling an IND application (21 CFR Part 312), whichmust be submitted before initiating studies in hu-mans. The IND will be expected to contain extensiveinformation for the review process, including, butnot limited to, a description of Institutional ReviewBoard review (21 CFR Part 56) and informed con-sent (21 CFR Part 50); how the product is made, toensure that safe, high-quality manufacturing pro-cesses are used; and, often, data from preclinicalanimal toxicology studies to demonstrate that it issafe to proceed with human clinical studies. As inother countries, the regulatory system has been setup so that only a drug, not a food, can treat, pre-vent, or cure disease. Questions were raised duringthe meeting as to why this bureaucratic distinctionstill remains in place today.
Guidance issued by the FDA in October, 2010on “Determining whether human research stud-ies can be conducted without an IND”45 had par-ticular relevance for studies involving probiotics.This document included language suggesting thatan IND would be required for studies in which alive organism (e.g., virus, bacterium, or fungus) isadministered to subjects to study “the pathogene-sis of disease” or “the host response to the organ-ism.” The strictest interpretation of this statementcould be that any probiotic investigation would re-quire an IND. Other guidance by the FDA issuedin February 2012 focuses on clinical trials with livebio-therapeutic agents,46 with specific reference tolanguage being “applicable” to the prevention, treat-ment, or cure of disease. However, this document ac-knowledges that a basis exists for conducting humanstudies on food (including dietary supplements)that do not require the same IND application pro-
cess as drug studies; such studies will need to avoiddrug-type endpoints. The intended use will dictateregulation, but examples of appropriate food targetsinclude human studies intended to establish healthclaims, structure/function claims, or medical foodclaims. In conclusion, to avoid FDA imposition ofa requirement for an IND, Degnan recommended(1) conceiving and designing studies that considerthe intended use and a clear understanding of reg-ulatory categories; (2) using caution in document-ing/substantiating non-biological product use; and(3) consulting with FDA’s Center for Food Safetyand Applied Nutrition.
Conclusions
The conference “Probiotics, Prebiotics, and theHost Microbiome: The Science of Translation” pro-vided a forum in which recent developments andthe potential benefits of translating research ad-vances in the human microbiome, probiotics, andprebiotics into robust nutritional and therapeu-tic applications to promote health were examined.The number of genes in the microbiome is es-timated to be more than 300-fold higher thanthe total number of human genes, which high-lights the existence of a highly complex micro-biota ecosystem with the potential for profoundeffects on metabolism and immune function. Signif-icant advances have been achieved and further stud-ies will greatly enhance our understanding of thehuman microbiota and its role in health and diseasedevelopment.
It is well established that the intestinal micro-biota plays a critical role in gastrointestinal devel-opment and function while regulating host inflam-matory responses and immune homeostasis.47,48 Arapidly growing body of evidence now also indi-cates that the microbiota acts as a metabolically ac-tive organ, capable of interacting with several hostsystems beyond the gastrointestinal tract, includingthe brain, urogenital tract, and respiratory tract.49–51
Recent research suggests the gut microbiota is capa-ble of influencing fat storage and metabolism,52,53
which may position it as a key target in the fightagainst obesity in conjunction with dietary, exer-cise, and other interventions. Disruptions in theearly programming of the gut microbiota or al-terations of adult-like microbiota may contributeto the development of obesity and T2D. Proposedmechanisms include effects on hormone-based
Petschow et al. Probiotics, prebiotics, and the host microbiome
satiety, energy salvage, appetite regulation, and LPS-induced metabolic endotoxemia. The use of probi-otics and prebiotics that target specific changes inthe microbiota and its crosstalk with the host to im-prove lipid metabolism and insulin resistance areexciting potential options for improving the man-agement of what is arguably the 21st century’s majorpublic health issue.
The gut–brain axis is a highly complex sys-tem comprising interactions that involve the au-tonomic and enteric nervous systems, pituitaryand gut hormones, and gut inflammatory systemsthat are capable of influencing nerve function andpathways, and ultimately behavior. Central ner-vous system wiring may be influenced by gut–braininteractions early in life, and alterations in the mi-crobiome may influence behaviors related to specificdisease conditions. Examples include the possibili-ties of either influencing the microbiota with pro-biotics to modulate cholecystokinin (CCK) output,which in turn regulates vagal afferent neuron trans-missions involved in communicating satiety andavoiding obesity, or providing prebiotics to encour-age fermentative production of SCFAs that stimulaterelease of gut hormones, which in turn influence hy-pothalamic neuronal activity involved with appetiteregulation.
Many questions remain regarding the develop-ment of the microbiota in the young infant, andwhether probiotic/prebiotic interventions at thistime would be effective in supporting the develop-ment of a lifelong microbiota for health. Much needsto be learned about how the microbiota is assem-bled, what influences community structure succes-sion, and which factors contribute to its long-termstability in both health and illness. It seems likelythat the future roles of probiotics and prebiotics willgo beyond traditional gastrointestinal illnesses, par-ticularly as the role of the microbiota and the CNSand other organs is better understood. Novel appli-cations in the future may include chronic immunedisorders, and anxiety-like behaviors or psychiatricillness. The development of such products is certainto face increased scrutiny over costs and benefits tosupport decisions about utilization and reimburse-ment for disease management. Influencing publichealth policy to more effectively adopt the use ofsuch products will require clear understanding andcommunication of the health benefits, building thewill for change with providers and policy makers,
executing the changes in policy, and driving thesechanges with strong, reproducible data.
As we get closer to understanding the potentialmechanisms by which particular probiotic organ-isms interact with the microbiota, it will be a missedopportunity if the quality of probiotic research stud-ies does not improve to meet the needs for eitherevidence-based medicine or nutrition. Standard-ization of probiotic/prebiotic study methods andprotocols, clear understanding of the characteris-tics, purity, and stability of test agents, and accurateand balanced reporting of study results are urgentlyneeded. In parallel, it will be important to educatehealthcare professionals, regulatory authorities, andthe public to understand the appropriate use anddocumented safety and benefits of probiotic or pre-biotic products. Presentations and discussions dur-ing the conference reiterated that there is no optingout of this paradigm shift, but rather a matterof when and how the innovations from micro-biota/microbiome and probiotic/prebiotic researchwill become part of everyday life. For those in thefield, the regulatory antiquity and recalcitrance ofmany physicians to move from a pharmaceutical-based patient-management perspective to a moreholistic one that includes recommendations of foodand supplement-based products to both generaland patient populations remain among the impedi-ments to progress. Meanwhile, regulatory issues onpro- and prebiotics remain a point of concern. High-quality human research conducted on the generalpopulation is required to convince regulators of thelegitimacy of health benefits of foods. Many con-vincing probiotic studies have been done on popu-lations that are outside the scope of foods or supple-ments. However, the safety and lack of side effectsfor these products is a strong plus. Thus, it is in-cumbent upon those in the field to help strengthenthe body of evidence and merge the knowledge in amanner that allows consumers and patients to reapthe benefits sooner rather than later.
Acknowledgments
The conference “Probiotics, Prebiotics, and the HostMicrobiome: The Science of Translation” was jointlypresented by The Sackler Institute for NutritionScience at the New York Academy of Sciences andThe International Scientific Association for Pro-biotics and Prebiotics in New York, New York,12 June 2013. The conference and open access of
Probiotics, prebiotics, and the host microbiome Petschow et al.
this publication were also generously supported (inpart) by Award Number R13AT007899 from theNational Center for Complementary and Alterna-tive Medicine (NCCAM) of the National Institutesof Health (NIH). Co-funding in support of TravelFellowships to foster the participation of early ca-reer and underrepresented minority investigatorsin this conference was provided by the following:NIH Division of Nutrition Research Coordination(DNRC), the Trans-NIH Probiotics/Prebiotics andMicrobiome Working Group (trans-NIH PPWG/T-MWG), the National Center for Complemen-tary and Alternative Medicine (NCCAM), theOffice of Dietary Supplements (ODS), and the Eu-nice Kennedy Shriver National Institute of ChildHealth and Human Development (NICHD). Theviews expressed in written conference materials orpublications and by speakers and moderators donot necessarily reflect the official policies of the De-partment of Health and Human Services; nor doesmention by trade names, commercial practices, ororganizations imply endorsement by the US Gov-ernment. The authors also want to thank JulietAnsell, Harry Burns, Megan Enos, Gary Frost, DavidMills, Rowena Pullan, Helen Reybould, and VictoriaRuiz for their participation during the conference.
Conflicts of interest
T.K. is a consultant for Ritter Pharmaceuticals andhelped plan the microbiome aspects of the projectreported on page 17. M.E.S. consults with numerousfood and dietary supplement companies conductingbusiness in the probiotic industry. She does not haveany ownership role or serve on governing boards forany company. The other authors declare no conflictsof interest.
References
1. Probiotics, Prebiotics, and the Host Microbiome: TheScience of Translation. http://www.nyas.org/events/Detail.aspx?cid=c60ea8d5-44f0-4aaa-a8ff-3e5f008186f6, 2013.
2. Trueman, P., M. Hurry, M. Bending, et al. 2009. The feasi-bility of harmonizing health technology assessments acrossjurisdictions: a case study of drug eluting stents. Int. J. Tech-nol. Assess Health Care 25: 455–462.
3. Whitehead, S.J., L. Lee, Z. Fang, et al. 2010. Can fae-cal calprotectin reduce the demand for colonoscopy inpatients with irritable bowel syndrome? Gut 59: A36;doi:10.1136/gut.2009.208991i.
4. Qin, J., R. Li, J. Raes, et al. 2010. A human gut microbial genecatalogue established by metagenomic sequencing. Nature464: 59–65.
5. Palmer, C., E.M. Bik, D.B. DiGiulio, et al. 2007. Developmentof the human infant intestinal microbiota. PLoS Biol. 5: e177.
6. Dominguez-Bello, M.G., E.K. Costello, M. Contreras, et al.2010. Delivery mode shapes the acquisition and structureof the initial microbiota across multiple body habitats innewborns. Proc. Natl. Acad. Sci. U.S.A. 107: 11971–11975.
7. Morowitz, M.J., V.J. Denef, E.K. Costello, et al. 2011. Strain-resolved community genomic analysis of gut microbialcolonization in a premature infant. Proc. Natl. Acad. Sci.U.S.A. 108: 1128–1133.
8. Yatsunenko, T., F.E. Rey, M.J. Manary, et al. 2012. Humangut microbiome viewed across age and geography. Nature486: 222–227.
9. Cho, I., S. Yamanishi, L. Cox, et al. 2012. Antibiotics inearly life alter the murine colonic microbiome and adiposity.Nature 488: 621–626.
10. Trasande, L., J. Blustein, M. Liu, et al. 2013. Infant antibioticexposures and early-life body mass. Int. J. Obes. (Lond.) 37:16–23.
11. Cani, P.D., J. Amar, M.A. Iglesias, et al. 2007. Metabolic en-dotoxemia initiates obesity and insulin resistance. Diabetes56: 1761–1772.
12. Cani, P.D., R. Bibiloni, C. Knauf, et al. 2008. Changes ingut microbiota control metabolic endotoxemia-induced in-flammation in high-fat diet-induced obesity and diabetes inmice. Diabetes 57: 1470–1481.
13. Rabot, S., M. Membrez, A. Bruneau, et al. 2010. Germ-freeC57BL/6J mice are resistant to high-fat-diet-induced insulinresistance and have altered cholesterol metabolism. FASEBJ. 24: 4948–4959.
14. Backhed, F., J.K. Manchester, C.F. Semenkovich, et al. 2007.Mechanisms underlying the resistance to diet-induced obe-sity in germ-free mice. Proc. Natl. Acad. Sci. U.S.A. 104:979–984.
15. Turnbaugh, P.J., F. Backhed, L. Fulton, et al. 2008. Diet-induced obesity is linked to marked but reversible alterationsin the mouse distal gut microbiome. Cell Host Microbe. 3:213–223.
16. Carvalho, B.M., D. Guadagnini, D.M. Tsukumo, et al. 2012.Modulation of gut microbiota by antibiotics improves in-sulin signalling in high-fat fed mice. Diabetologia 55: 2823–2834.
17. Daubioul, C., N. Rousseau, R. Demeure, et al. 2002. Dietaryfructans, but not cellulose, decrease triglyceride accumu-lation in the liver of obese Zucker fa/fa rats. J. Nutr. 132:967–973.
18. Muccioli, G.G., D. Naslain, F. Backhed, et al. 2010. The en-docannabinoid system links gut microbiota to adipogenesis.Mol. Syst. Biol. 6: 392.
19. Cani, P.D., S. Possemiers, T. Van de Wiele, et al. 2009.Changes in gut microbiota control inflammation in obesemice through a mechanism involving GLP-2-driven im-provement of gut permeability. Gut. 58: 1091–1103.
20. Everard, A., C. Belzer, L. Geurts, et al. 2013. Cross-talk be-tween Akkermansia muciniphila and intestinal epitheliumcontrols diet-induced obesity. Proc. Natl. Acad. Sci. U.S.A.110: 9066–9071.
21. van Nood, E., A. Vrieze, M. Nieuwdorp, et al. 2013. Duodenalinfusion of donor feces for recurrent Clostridium difficile.N. Engl. J. Med. 368: 407–415.
Petschow et al. Probiotics, prebiotics, and the host microbiome
22. Vrieze, A., E. Van Nood, F. Holleman, et al. 2012. Transferof intestinal microbiota from lean donors increases insulinsensitivity in individuals with metabolic syndrome. Gas-troenterology 143: 913–916, e7.
23. Karlsson, F.H., V. Tremaroli, I. Nookaew, et al. 2013. Gutmetagenome in European women with normal, impairedand diabetic glucose control. Nature 498: 99–103.
24. de Vos, W.M. & M. Nieuwdorp. 2013. Genomics: A gutprediction. Nature 498: 48–49.
25. Early Years Collaborative, The Scottish Government. http://www.scotland.gov.uk/Topics/People/Young-People/Early-Years-and-Family/early-years-collaborative.
26. Deshpande, G., S. Rao & S. Patole. 2011. Progress in thefield of probiotics: year 2011. Curr. Opin. Gastroenterol. 27:13–18.
27. Bernardo, W.M., F.T. Aires, R.M. Carneiro, et al. 2013. Ef-fectiveness of probiotics in the prophylaxis of necrotizingenterocolitis in preterm neonates: a systematic review andmeta-analysis. J. Pediatr. (Rio. J.) 89: 18–24.
28. Goldenberg, J.Z., S.S. Ma, J.D. Saxton, et al. 2013. Probioticsfor the prevention of Clostridium difficile-associated diar-rhea in adults and children. Cochrane Database Syst. Rev. 5:CD006095.
29. Macklaim, J.M., C.R. Cohen, G. Donders, et al. 2012. Explor-ing a road map to counter misconceptions about the cervico-vaginal microbiome and disease. Reprod. Sci. 19: 1154–1162.
30. Hao, Q., Z. Lu, B.R. Dong, et al. 2011. Probiotics for pre-venting acute upper respiratory tract infections. CochraneDatabase Syst. Rev. CD006895.
31. Isolauri, E., S. Rautava & S. Salminen. 2012. Probiotics in thedevelopment and treatment of allergic disease. Gastroenterol.Clin. North Am. 41: 747–762.
32. Gut Microbiota for Health. http://www.gutmicrobiotaforhealth.com/.
33. Sudo, N., Y. Chida, Y. Aiba, et al. 2004. Postnatal microbialcolonization programs the hypothalamic-pituitary-adrenalsystem for stress response in mice. J. Physiol. 558: 263–275.
34. Neufeld, K.M., N. Kang, J. Bienenstock, et al. 2011. Reducedanxiety-like behavior and central neurochemical changein germ-free mice. Neurogastroenterol. Motil. 23: 255–264,e119.
35. Neufeld, K.A., N. Kang, J. Bienenstock, et al. 2011. Effectsof intestinal microbiota on anxiety-like behavior. Commun.Integr. Biol. 4: 492–494.
36. Tazoe, H., Y. Otomo, I. Kaji, et al. 2008. Roles of short-chain fatty acids receptors, GPR41 and GPR43 on colonicfunctions. J. Physiol. Pharmacol. 59(Suppl 2): 251–262.
37. de La Serre, C.B., C.L. Ellis, J. Lee, et al. 2010. Propensityto high-fat diet-induced obesity in rats is associated withchanges in the gut microbiota and gut inflammation. Am. J.Physiol. Gastrointest Liver Physiol. 299: G440–G448.
38. Lee, S., B.R. Belitsky, J.P. Brinker, et al. 2010. Developmentof a Bacillus subtilis-based rotavirus vaccine. Clin. VaccineImmunol. 17: 1647–1655.
39. Lee, S., B.R. Belitsky, D.W. Brown, et al. 2010. Efficacy, heatstability and safety of intranasally administered Bacillus sub-tilis spore or vegetative cell vaccines expressing tetanus toxinfragment C. Vaccine 28: 6658–6665.
40. Sokol, H., P. Seksik, J.P. Furet, et al. 2009. Low counts of
Faecalibacterium prausnitzii in colitis microbiota. Inflamm.Bowel Dis. 15: 1183–1189.
41. Sokol, H., B. Pigneur, L. Watterlot, et al. 2008. Faecalibac-terium prausnitzii is an anti-inflammatory commensal bac-terium identified by gut microbiota analysis of Crohn diseasepatients. Proc. Natl. Acad. Sci. U.S.A. 105: 16731–16736.
42. Kalliomaki, M., M.C. Collado, S. Salminen, et al. 2008. Earlydifferences in fecal microbiota composition in children maypredict overweight. Am. J. Clin. Nutr. 87: 534–538.
43. Furet, J.P., L.C. Kong, J. Tap, et al. 2010. Differential adapta-tion of human gut microbiota to bariatric surgery-inducedweight loss: links with metabolic and low-grade inflamma-tion markers. Diabetes 59: 3049–3057.
44. Rajilic-Stojanovic, M., E. Biagi, H.G. Heilig, et al. 2011.Global and deep molecular analysis of microbiota signa-tures in fecal samples from patients with irritable bowelsyndrome. Gastroenterology 141: 1792–1801.
45. Guidance for Industry: Investigational New Drug Ap-plications (INDs) http://www.fda.gov/downloads/Drugs/Guidances/UCM229175.pdf.
46. Guidance for Industry: Early Clinical Trials with LiveBiotherapeutic Products http://www.fda.gov/downloads/BiologicsBloodVaccines/GuidanceComplianceRegulatory-Information/Guidances/General/UCM292704.pdf.
47. Backhed, F., R.E. Ley, J.L. Sonnenburg, et al. 2005. Host-bacterial mutualism in the human intestine. Science 307:1915–1920.
48. Cebra, J.J. 1999. Influences of microbiota on intestinal im-mune system development. Am. J. Clin. Nutr. 69: 1046S–1051S.
49. Schippa, S., V. Iebba, F. Santangelo, et al. 2013. Cystic fibrosistransmembrane conductance regulator (CFTR) allelic vari-ants relate to shifts in faecal microbiota of cystic fibrosispatients. PLoS One 8: e61176.
50. Reid, G., J.A. Younes, H.C. Van der Mei, et al. 2011. Mi-crobiota restoration: natural and supplemented recoveryof human microbial communities. Nat. Rev. Microbiol. 9:27–38.
51. Bravo, J.A., M. Julio-Pieper, P. Forsythe, et al. 2012. Commu-nication between gastrointestinal bacteria and the nervoussystem. Curr. Opin. Pharmacol. 12: 667–672.
52. Hooper, L.V., T. Midtvedt & J.I. Gordon. 2002. How host-microbial interactions shape the nutrient environment ofthe mammalian intestine. Annu. Rev. Nutr. 22: 283–307.
53. Velagapudi, V.R., R. Hezaveh, C.S. Reigstad, et al. 2010. Thegut microbiota modulates host energy and lipid metabolismin mice. J. Lipid Res. 51: 1101–1112.
54. Koenig, J.E., A. Spor, N. Scalfone, et al. 2011. Successionof microbial consortia in the developing infant gut micro-biome. Proc. Natl. Acad. Sci. U.S.A. 108(Suppl 1): 4578–4585.
55. Hicks, L.A., T.H. Taylor Jr. & R.J. Hunkler. 2013. U.S. out-patient antibiotic prescribing, 2010. N. Engl. J. Med. 368:1461–1462.
56. Delzenne, N.M., A.M. Neyrinck, F. Backhed, et al. 2011.Targeting gut microbiota in obesity: effects of prebiotics andprobiotics. Nat. Rev. Endocrinol. 7: 639–646.
57. Cryan, J.F. & T.G. Dinan. 2012. Mind-altering microor-ganisms: the impact of the gut microbiota on brain andbehaviour. Nat. Rev. Neurosci. 13: 701–712.