THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 257, No. 20, Issue of October 25, pp. 12121-12128, 1982 Prrnled in 1J.S.A. Products of DNA Cleavage by the 1,lO-Phenanthroline-Copper Complex INHIBITORS OF ESCHERICHIA COLI DNA POLYMERASE I* (Received for publication,March 12, 1982) Laura Marshall Pope$, Karl A. Reich, Daniel R. Graham$, and David S. Sigmang From the Department of Biological Chemistry, School of Medicine, and the Molecular Biology Institute, University of California, Los Angeles, Los Angeles, California 90024 The 2:l 1,lO-phenanthroline-cuprous complex ((OP)&u+)with hydrogen peroxide possesses an arti- ficial DNase activity that rapidly cleaves double- stranded DNA at pH 7.0 and 37 “C. The chemical ter- mini formed in the scission of the phosphodiester back- bone have been investigated in order to understand the mechanism of this unusual reaction and identify the products responsible for the inhibition of Escherichia coli DNA polymerase I. With ~o~~([~H,~’P]~A-[~H]T) as substrate, 20% of the 32P label can be isolated as inor- ganic phosphate following incubation of a digest with alkaline phosphatase. The phosphomonoesters hydro- lyzed by aIkaline phosphatase are present at both the 5’ and 3’ termini. 5’-phosphorylated ends have been demonstrated by the alkaline phosphatase-dependent labeling of the digest by polynucleotide kinase with [y-32P]ATP. The inhibition of E. coli DNA polymerase I associated with the cleavage of poly(dA-T) by (OP)zCu+ (Sigman, D. S., Graham, D. R., D’Aurora, V. D., and Stern, A. M. (1979) J. Biol. Chem 254,12269-12272) has been used to infer the formation of 3’-phosphorylated ends since alkaline phosphatase and exonuclease I11 relieve the inhibition and restore the primer-template competence of polynucleotides that have been depo- lymerized by the coordination complex. The free bases adenine and thymine are also formed in the cleavage of poly(dA-T) by (OP)zCu+. A recovery of 1 free base/every 2 phosphates liberated by alkaline phosphatase suggests that base release may be obliga- tory in one of the 2 possible pathways leading to strand scission. All of the deoxyribose carbons remain at- tached to the remnants of the phosphodiester back- bone. No derivatives of the deoxyribose reactive to NaB-, dimedone, thiobarbituric acid, hydroxylamine, or carbodiimide are formed. Based on the product anal- ysis and other properties of the reaction, the cleavage is proposed to proceed via the oxidation of the deoxy- ribose to a resonance-stabilized furan derivative by hydroxyl radicals produced from the hydrogen perox- ide oxidation of (OP)&u+ intercalated in the double- stranded DNA. The lJ0-phenanthroline-cuprous complex andhydrogen peroxide are essential co-reactants in an artificial DNase * This work was supported by United States Public Health Service Grant 21199. A preliminaryreport of these studies has been presented at the 1981 Meeting of the American Society of Biological Chemists, St. Louis, MO (1981) Fed. Proc. 40, 1730. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. $ Recipient of a grant from the United States Public Health Service Cell and Molecular Biology Program (GM 07185-05). !j To whom correspondence should be addressed. activity that makes single-stranded nicks in double-stranded DNA (1). No primary sequence specificity is apparent in the scission reaction which proceeds readily under a variety of 1‘10-phenanthroline experimental conditions at pH 7.0 and 37 “C (2). These include incubation of the DNA, OP,’ and cupric ion with: (a) thiol and molecular oxygen (1); (b) the superoxide generator, xan- thine and xanthine oxidase (3); (c) NADH and peroxide (4); and (d) thiol and hydrogen peroxide (2). The common feature of all thesereaction conditions is that they can generate kineticallysignificant concentrations of hydrogenperoxide and the 1,lO-phenanthroline-cuprous complex, the co-reac- tank directly responsible for the DNA scission reaction. The reaction is always inhibited by intercalating agents and any reagent which reduces either the concentration of the 2:11,10- phenanthroline-cuprous complex (e.g. a cuprous ion-specific chelating agent) or hydrogen peroxide (e.g. catalase) (1-3). The sensitivity of the reaction to other inhibitors depends on the pathway for the generation of (0P)&ui and H202. For example, superoxide dismutase inhibits the reaction poten- tiated by NADH andHz02 but has no effect on that activated by thiol and hydrogen peroxide (4). In this paper, we report the chemical nature of the termini formed when the phosphodiester backbone is cleaved by (OP)2Cu+ andhydrogen peroxide. A detailed understanding of this artificial DNase activity, which must include a struc- tural analysis of the ends, is of interest from several perspec- tives. The fist derives from the widespread use of 1,lO-phe- nanthroline to infer a functional role of zinc ion in DNA and RNA polymerases based on the sensitivity of these enzymes to this ligand. We had initially shown the OP inhibition of Escherichia coli DNA polymerase I and other polymerases was mediated through the cuprous complex (5, 6) which in turn was due to the cleavage reaction whose products were potent inhibitors of E. coli DNA polymerase I (1). Identifi- cation of these inhibitory structures will assist in the interpre- tation of the extensive literature on the inhibition of polym- erases by OP (1, 7-9). Secondly, hydroxyl radicals and other ’ The abbreviations and trivial names used are: OP, 1,lO-phenan- throline; (OP)L!u+, 2:l 1,lO-phenanthroline-cuprous ion complex; (OP)ZCU*+, 2:l 1,lO-phenanthroline-cupric ion complex; polymerase I, E. coli polymerase DNA I; MPA, 3-mercaptopropionic acid; 5-NacOP, 5-acetamido-1,lO-phenanthroline. 12121
Transcript
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 257, No. 20, Issue of October 25, pp. 12121-12128, 1982 Prrnled in 1J.S.A.
Products of DNA Cleavage by the 1,lO-Phenanthroline-Copper Complex INHIBITORS OF ESCHERICHIA COLI DNA POLYMERASE I*
(Received for publication, March 12, 1982)
Laura Marshall Pope$, Karl A. Reich, Daniel R. Graham$, and David S. Sigmang From the Department of Biological Chemistry, School of Medicine, and the Molecular Biology Institute, University of California, Los Angeles, Los Angeles, California 90024
The 2:l 1,lO-phenanthroline-cuprous complex ((OP)&u+) with hydrogen peroxide possesses an arti- ficial DNase activity that rapidly cleaves double- stranded DNA at pH 7.0 and 37 “C. The chemical ter- mini formed in the scission of the phosphodiester back- bone have been investigated in order to understand the mechanism of this unusual reaction and identify the products responsible for the inhibition of Escherichia coli DNA polymerase I. With ~o~~([~H,~’P]~A-[~H]T) as substrate, 20% of the 32P label can be isolated as inor- ganic phosphate following incubation of a digest with alkaline phosphatase. The phosphomonoesters hydro- lyzed by aIkaline phosphatase are present at both the 5’ and 3’ termini. 5’-phosphorylated ends have been demonstrated by the alkaline phosphatase-dependent labeling of the digest by polynucleotide kinase with [y-32P]ATP. The inhibition of E. coli DNA polymerase I associated with the cleavage of poly(dA-T) by (OP)zCu+ (Sigman, D. S., Graham, D. R., D’Aurora, V. D., and Stern, A. M. (1979) J. Biol. Chem 254,12269-12272) has been used to infer the formation of 3’-phosphorylated ends since alkaline phosphatase and exonuclease I11 relieve the inhibition and restore the primer-template competence of polynucleotides that have been depo- lymerized by the coordination complex.
The free bases adenine and thymine are also formed in the cleavage of poly(dA-T) by (OP)zCu+. A recovery of 1 free base/every 2 phosphates liberated by alkaline phosphatase suggests that base release may be obliga- tory in one of the 2 possible pathways leading to strand scission. All of the deoxyribose carbons remain at- tached to the remnants of the phosphodiester back- bone. No derivatives of the deoxyribose reactive to NaB-, dimedone, thiobarbituric acid, hydroxylamine, or carbodiimide are formed. Based on the product anal- ysis and other properties of the reaction, the cleavage is proposed to proceed via the oxidation of the deoxy- ribose to a resonance-stabilized furan derivative by hydroxyl radicals produced from the hydrogen perox- ide oxidation of (OP)&u+ intercalated in the double- stranded DNA.
The lJ0-phenanthroline-cuprous complex and hydrogen peroxide are essential co-reactants in an artificial DNase
* This work was supported by United States Public Health Service Grant 21199. A preliminary report of these studies has been presented at the 1981 Meeting of the American Society of Biological Chemists, St. Louis, MO (1981) Fed. Proc. 40, 1730. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Recipient of a grant from the United States Public Health Service Cell and Molecular Biology Program (GM 07185-05).
!j To whom correspondence should be addressed.
activity that makes single-stranded nicks in double-stranded DNA (1). No primary sequence specificity is apparent in the scission reaction which proceeds readily under a variety of
1‘10-phenanthroline
experimental conditions at pH 7.0 and 37 “C (2). These include incubation of the DNA, OP,’ and cupric ion with: ( a ) thiol and molecular oxygen (1); ( b ) the superoxide generator, xan- thine and xanthine oxidase (3); ( c ) NADH and peroxide (4); and ( d ) thiol and hydrogen peroxide (2). The common feature of all these reaction conditions is that they can generate kinetically significant concentrations of hydrogen peroxide and the 1,lO-phenanthroline-cuprous complex, the co-reac- t ank directly responsible for the DNA scission reaction. The reaction is always inhibited by intercalating agents and any reagent which reduces either the concentration of the 2:11,10- phenanthroline-cuprous complex (e.g. a cuprous ion-specific chelating agent) or hydrogen peroxide (e.g. catalase) (1-3). The sensitivity of the reaction to other inhibitors depends on the pathway for the generation of (0P)&ui and H202. For example, superoxide dismutase inhibits the reaction poten- tiated by NADH and Hz02 but has no effect on that activated by thiol and hydrogen peroxide (4).
In this paper, we report the chemical nature of the termini formed when the phosphodiester backbone is cleaved by (OP)2Cu+ and hydrogen peroxide. A detailed understanding of this artificial DNase activity, which must include a struc- tural analysis of the ends, is of interest from several perspec- tives. The fist derives from the widespread use of 1,lO-phe- nanthroline to infer a functional role of zinc ion in DNA and RNA polymerases based on the sensitivity of these enzymes to this ligand. We had initially shown the OP inhibition of Escherichia coli DNA polymerase I and other polymerases was mediated through the cuprous complex (5, 6) which in turn was due to the cleavage reaction whose products were potent inhibitors of E. coli DNA polymerase I (1). Identifi- cation of these inhibitory structures will assist in the interpre- tation of the extensive literature on the inhibition of polym- erases by OP (1, 7-9). Secondly, hydroxyl radicals and other
’ The abbreviations and trivial names used are: OP, 1,lO-phenan- throline; (OP)L!u+, 2:l 1,lO-phenanthroline-cuprous ion complex; (OP)ZCU*+, 2:l 1,lO-phenanthroline-cupric ion complex; polymerase I, E . coli polymerase DNA I; MPA, 3-mercaptopropionic acid; 5-NacOP, 5-acetamido-1,lO-phenanthroline.
12121
12122 Products of DNA Cleavage
oxygen species appear responsible for many DNA modifica- tion reactions. However, the detailed chemistry, particularly of strand scission reactions, is unclear (10). The (OP)&u+- dependent reaction is probably reflective of this chemistry and is a convenient example to study in detail. Finally, the DNase-like activity of (OP)&u+ is unique. No known biolog- ical DNase is specific for double-stranded B-form helix DNA without being highly specific for primary sequence. This prop- erty makes this reaction uniquely suited for “footprinting” analyses of protein-DNA and intercalating agent-DNA bind- ing (11) and studying conformational transitions of DNA. This chemical cleavage has recently been used in analyzing the specific distribution of nucleosomes on a given DNA sequence (12).
EXPERIMENTAL PROCEDURES’ M a t e r i a l s
Fraction- fran E. c o l i B cel ls purchased f rom Grain Processing Corp.. E. c o l i DNA Polymerase I (Pol I ) was prepared to the s tage des ignated
Muscatin , lowa (13).” 5-[~H]Acetamido-l.lO-phenanthroline was prepared by ace ty la t i on o f
5-amino-1.10-phenanthroline with C3H]acet ic anhydr ide (10 nCi /m) cata lyzed by s u l f u r i c a c i d . It was p u r i f i e d on LH-20 (Pharmacia) e lut ing wi th methanol . The product was c h r a a t o g r a p h i c a l l y i n d i s t i n g u i s h a b l e f r a n an authent ic sample (14). 5-Amino-1,lO-phenanthroline was prepared by d i th ion i te reduc t ion o f 5-nitro-1.10-phenanthroline (14).
Radiolabeled polynucleot ides were synthesized as p rev ious l y desc r ibed
on a 6-75 column e l u t i n g w i t h 0.1 M T r i s Ac. 5 nll MgAc, pH = 7.0. The f i r s t (2 ) . Poly (dA-T), approximately 400 bp i n length, was s ized by g e l f i l t r a t i o n
peak t o m e r g e was used as u b s t r a t e w en phosphorylated ends and free base re lease were quant i ta ted. [&INaBH4. [ I Hlacet ic anhydr ide and labeled nucleo- t i d e s were purchased f r a n ICN. The fol lowing chemicals were obtained i n t h e h i g h e s t g r a d e c m e r c i a l l y a v a i l a b l e and used wi thout fur ther pur i f icat ion: poly (dA-T), (Miles); 1.10-phenanthroline and 5-nitro-1.10-phenanthroline
G.F. Smith); cal f thymus DNA, t r i s . NADH and taur ine (S igna) ; e thy lenediamine
metho-p- to luene su l fonate, g lyc ine methy l ester , th iobarb i tur ic ac id , 3-mer- Eastman); 1.6-diamino-hexane. cyclohexyl-3-(2-norpholinoethyl)-carbodiinide
cap top rop ion i c ac id (A ld r i ch ) ; cup r i c su l fa te and dimedone (Mal l inckrodt ) ; po lynucleot ide k inase (P-L) , and exonuclease 111 (Bethesda Research Labs). Bleomycin was a g i f t fran Dr. U i I l i aa B radner o f B r i s to l Labora to r ies , Syracuse, NY.
Methods
Polymerization and Cleavage Assays
The assays of t h e p o l y m e r i z a t i o n a c t i v i t y o f r. c o l i P o l 1 and t h e DNA c leavage reac t ion by (DP)2Cu+ us ing rad io labeled poly--1) have been p r e v i - ously descr ibed (2).
g u a n t i t a t i o n of Phosphate Termini i Using Alkal ine Phosphatase
In a typical experiment, 5-10 ugh1 poly ([3Hl[32PldA-[3H3r) was incu- ba ted a t 37% w i t h and wi thout 20 uM W and 2 uM CuSOq in t he p resence of 7 rill MPA and 7 nll H202. A f te r 45 minu tes the reac t ion was stopped by adding Cata- l a s e ( f i n a l conc.: 0.133 mg/ml). P a r t o f t h e s o l u t i o n (0.750 m l ) was then incubated a t 65OC f o r 30 minutes wi th 0.2 u n i t s o f a l k a l i n e phosphatase added a t 15 minute in terva ls . Another 0.750 m l o f each so lut ion was incubated for 30 minutes a t 65W w i th no addi t ions.
Ice-co ld 102 t r i c h l o r o a c e t i c a c i d (2.0 m l ) was then added t o 0.5 m l of each sample. The samples were kept on i ce fo r 5 m inu tes and then cent r i fuged 11 minutes at maximum speed i n a c l i n i ca l cen t r i f uge . The supernatant (2.0 m l ) was then ex t rac ted w i th 2.5 ml buty l acetate and 0.5 m l amnonium molybdate (3.752) i n 3 N H2SO4. The butyl acetate-molybdate layer was removed and 2.0 m l was counted i n 12 m l s c i n t i l l a t i o n c o c k t a i l (RPI 3a70b). The r a t i o o f t h e t o t a l phosphate i n t h e i n i t i a l sample t o t h a t e x t r a c t e d i n t o t h e b u t y l acetate-molybdate layer was then ca lcu lated.
Polynucleot ide K inase Label ing o f Termin i i (15)
WA and 7 nll Hz0 w i t h and without 10 UW OP and 1 ut4 CuSOq. The r e a c t i o n was monitored by fo l?o* ing the loss i n a c i d p r e c i p i t a b l e c o u n t s and was stopped a f t e r 40 minutes by mak ing t he so lu t i on 0.375 nll i n 2.9-dinethyl OP. To pro- v i e a PO i t i v e c o n t r o l f o r t h e a n a l y s i s w i t h p o l y n u c l e o t i d e k i n a s e ( 1 1 ) p o l y - ([QH]dA-[lH]T) was a l so d iges ted w i th DNAase 1 (0.4 ug/ml).
described above. P a r t o f each sample was a lso incubated a t 65oC without addi- A f t e r cleavage, the samples were t r e a t e d w i t h a l k a l i n e phosphatase as
t i o n o f phosphatase. The reac t i on m ix tu re f o r po l ynuc leo t i de k inase was pre- pared by adding 0.1 ml o f a s o l u t i o n c o n t a i n i n g 3 nll sodium phosphate pH =7.0
4 uM @-92Py\TP. 7.74 x 1012 cpm/mole t o 0.1 m1 o f t h e samples p r e v i o u s l y ( t o i n h i it the alkal ine phosphatase), 10 nll d i t h i o t h r e i t o l , 20 nll MgC12, and
heated a t 65%. The samples were e q u i l i b r a t e d t o 37°C. Polynucleot ide k inase (2 u n i t s ) was added and t h samples were al lowed to incubate for 30 minutes.
each sample t o 5 ml o f i c e - c o l d 5 1 t r i c h l o r o a c e t i c a c i d . 1.52 sodium pyrophos- The incorporat ion of 12P by the k inase was assayed by adding 0.1 m l o f
phate. The samples were t h e n f i l t e r e d s l o w l y t h r o u g h n i t r o c e l l u l o s e f i l t e r s
Poly (C3H]dA-T) (6.1 ug/ml) was incubated a t 37% i n t h e presence of 7 nll
‘The “Experimental Procedures” are presented in miniprint as prepared by the authors. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are available from the Journal of Biological Chemistry, 9650 Rockville Pike, Bethesda, MD 20814. Request Document No. 82M-627, cite the authors, and include a check or money order for $2.00 per set of photocopies. Full size photocopies are also included in the microfilm edition of the Journal that is available from Waverly Press.
were washed s low ly w i th about 10 m l of t h e i c e c o l d t r i c h l o r o a c e t i c acid-pyro- ( M i l l i p o r e HAW 025) i n a 12-sample M i l l i p o r e f i l t e r apparatus. The samples
phosphate solut ion and then rap id l y w i th app rox ima te l y 100 nl o f t h e same. The f i l t e r s were then washed w i t h 20 m l o f e thano l , d r i ed and counted i n s c i n - t i l l a t i o n c o c k t a i l (RPl 3a20). All experiments were done i n d u p l i c a t e . The
a f te r sub t rac t i on o f t he cpn re ta ined i n a con t ro l l ack ing po l ynuc leo t i de r a t i o of m o l e s n u c l e o t i d e s t o m o l e s P i i n c o r p o r a t e d was then calculated.
kinase.
Free Base Release
Two so lvent systems were used i n a n a l y z i n g f o r f r e e bases with descending
(1) Solvent A - MeOH:EtOH:12 N HCl:H20-50:25:6:19; ( 2 ) Solvent 8 - 952 Et0H:l N sodium acetate-7:). The products formed
paper chromatography.
i n t h e cleavage Of Poly ([3H]dA-C3H]T) (8 ug lm l ) by OP (40 uM) and Cut+ ( 4 uM) p o t e n t i a t e d by 7 nll R A and 10 nll H 02 and by 0.1 nll NADH and 0.5 nll Hz02 were analyzed i n a single experiment. A&er 60 minutes. the react ion mix ture was quenched w i t h 2.9-dimethyl-1.10-phenanthroline and 20 u l o f edch reac t ion mix - t u r e was spot ted on Yhatman Y# paper. Following development (Solvent A), t h e paper was c u t i n t o 1 cm2 s t r i ps wh ich were soaked i n 0.01 N H C 1 f o r 2 hours and counted i n R P I 3a70b s c i n t i l l a t i o n c o c k t a i l .
(PolY([3HldA-l)) or T (poly(dA-C3HlT)). These polynucleot ides (1.2 ug/ml) were separate ly incubated a t 37% w i t h 0.5 nll Hz02 and 0.1 nll NADH w i t h and without 10 uM OP and 1 uH CuSO4. A f t e r one hour the react ions were stopped by making the so lu t ions 0.71 nll i n 2.9-dimethyl OP. A l iquots o f each r e a c t i o n mix tu re (50 u l ) were spot ted on Yhatman 3nH paper and chromatographed i n a descending chamber e q u i l i b r a t e d w i t h ( S o l v e n t 6) . Adenine and thymine Stan- dards were a l s o chromatographed and l o c a l i z e d by UV absorption. Each lane was c u t i n t o 1 cm s t r i ps wh ich were counted i n 3.0 m1 s c i n t i l l a t i o n c o c k t a i l (RPI 3a20). To quant i ta te the re lease of free adenine and thymine, the total sum of counts on the paper a f ter subt ract ing for background was determined. The percentage of these counts migrat ing equivalent ly to the adenine or thymine
re1 ase i n t h e s o l u t i o n s l a c k i n g P and copper ion. Y i t h p o l y s tandards a re re fe r red to as %A o r 2T released. No (02) A o r T was
the base only, no products were observed which contained 14C only. ([lEC][gH]dA-T) prepared f ran [U-18c] l a b e l e d dATP and dATP t r i t i u m l a b e l e d i n
Base re lease was q u a n t i t a t e d u s i n g p o l y ( a - T ) s i n g l y l a b e l e d e i t h e r i n A
F rac t i ona t ion o f O l i gonuc leo t i des on DEAE C e l l u l o s e
Digests of poly (dA-T) prepared wi th (OP)zCu+ and DNAase I were f r a c t i o n - a ted on a 10 cm x 1 an DE 52 column e q u i l i b r a t e d w i t h 20 nll T r i s HC1. pH 7.6 and 7 M ured (16.17). The column was washed w i t h 30 m l of 20 nll T r i s HC1, pH 7.6, 7 M urea and e l u t e d w i t h a 0-0.3U NaCl g r a d i e n t i n t h e same buffer.
A l i q u o t s (2 nl) o f t h e f r a c t i o n s w e r e c o u n t e d i n a l i q u i d s c i n t i l l a n t (RPI E i g h t m i l l i l i t e r f r a c t i o n s were c o l l e c t e d a t a f l o w r a t e of 38.4 ml/hour.
t i on o f t he g rad ien t , add i t i ona l nuc leo t i des cou ld be removed f r a n t h e column 3a70b). The 5 ‘ dAnP was l o c a l i z e d by i t s absorbance a t 260 MI. A f t e r CanPle-
w i t h 2.0 M NaCl i n 20 nll T r i s HCI. pH 7.6. 7 H urea.
was prepared by incubat ing 11.86 ug/ml o f p o l y (3H-dA-T) w i t h 25 u n i t s h l Of The column was s tandard ized wi th a DNAase I d i g e s t of po ly (dA-1) which
enzyme a t 200 f o r 30 m inu tes . A f te r hea t i nac t i va t i on of the enzyme a t 75O for 15 minutes, the d igest was a p p l i e d o t h e column i n 6 ml a long w i th 2.5 mg o f dAnP. The (OP)zCu+ digest of poly ( jH-dA-T) was prepared by incubat ing
nll H202 and 7 nll mercaptopropionic a c i d a t 370. Af ter quenching the react ion 11.86 ug/ml of po lynuc leo t ide w i th 20 uM DP. 2 uH CuSO4 in the p resence o f 7
w i t h 2.9-dirnethyl OP. t h e s o l u t i o n (6.0 m l ) was a p p l i e d t o t h e Column subse- quent t o t h e a d d i t i o n o f 1.3 mg of 5 ’ dAnP as a standard.
Analyses for Reaction Products Derived from Deoxyribose
a t t h e 5 ’ o r 3 ’ end i n t h e case o f hydro lys is o f the phosphod ies ter bond; b) S t rand sc i ss ion requ i res e i t he r t ha t a ) an in tact deoxyr ibose be formed
the deoxyribose be degraded to two carbon or three carbon un i ts if Oligo- n u c l e o t i d e s p h o s p h o r y l a t e d a t b o t h t h e i r 3 ‘ o r 5 ‘ t e r m i n i a r e formed; O r C) a modified deoxyribose moiety remains attached t o t h e o l i g o n u c l e o t i d e b e a r i n g a phosphate a t i t s o t h e r t e r m i n i i . S i m p l e h y d r o l y t i c r e a c t i o n s a r e e x c l u d e d b y t h e i s o l a t i o n o f f r e e bases ( p o s s i b i l i t y a); products wi th 5 ’ and 3 ’ @os- pho ry la ted t e rm in i a r i s ing f ran t he canp le e d e s t r u c t i o n Of the deoxyr ibose are not supported by t h e f a i l u r e t o f i n d l r C only products i n t h e paper chromatographic exper iments (possibi l i ty b) . I n view of the format ion O f malondialdehyde i n bleomycin cleavage. aldehydic and ketonic products were sought us ing the fo l lowing tests making use of bleanlycin digests as P o s i t i v e contro ls .
a ) Th ioba rb i tu r i c Ac id Assay.
s t r a t e t h i s d e r i v a t i v e i n b l e o n y c i n d i g e s t s (18). Poly ([3H]dA-[5H3r) (11.7 This reagent i s spec i f i c f o r ma lond ia ldehyde and has been us d t o demon-
ug/ml) was cleaved i n 20 nll phosphate pH = 7.0 b u f f e r w i t h OP (20 uM), c u p r l c s u l f a t e (2 uM). MPA (7 mH) and Hz02 (7 nll); and with b leomycin (0.23 nll)
was stopped by the addi t ion of 2.9-dimethyl OP t o a concentrat ion o f 0.5 nll (B r i s to l Labora to r ies ) and Fet+(0.23 d l ) (18). The (OP)zCu+ r e a c t i o n m i x t u r e
and t h e b l e m y c i n r e a c t i o n was stopped by making the solut ion 2.8 nll i n EDTA.
were heated f o r 20 minutes a t lOOOC i n capped tubes. The samples were cooled A l i q u o t s (0.5 m l ) were added t o 2.0 m l 0.62 th ioba rb i tu r i c ac id . The samples
t o r o m t e m p e r a t u r e and the absorbance at 532 nm was read against a 20 nll phosphate buffer. pH - 7.0 blank which had been t r e a t e d w i t h t h i o b a r b i t u r i c a c i d i n an i d e n t i c a l manner. A malondialdehyde standard was prepared by heat-
cooled and d i l u t e d w i t h 8 0 m l Hz0 t o g i v e a 1 nll nalondialdehyde standard. i n g 16 ul malondialdehyde bis (d imethylacetal) i n 20 m1 0.1 N Hcl. This was
The nalondialdehyde standard was not a f fected by OP. Cu++, Hz02 and WA i n concentrat ions necessary t o cause cleavage.
Products of DNA Cleavage 12123
A f t e r t h e samples were mad a l k a l i n e by t h e a d d i t i o n of 8 u l Of I N NaOH t 0 0.3 m l of t h e above samples, [fH]NaBH4 (1.6 x l o g cpn o f 323 C i / y ) was added. The r e s u l t i n g s o l u t i o n remained i n i c e w a t e r f o r 90 minutes before 10 U l O f g l a c i a l a c e t i c a c i d was added t o d e s t r o y any res idual [3HmaBH4. One hour 1 t e r . 1.0 a1 of acetone was added as a p recau t ion t o t rap any r m a i n i n g [3HINaBH4 and t h e samples were evaporated under a stream of n i t rogen.
EDTA. pH = 7.0. An a l iquot o f each sample was added t o 5.0 m l i c e - c o l d 5% The samples were resuspended i n 0.150 m l 20 dl s o d i m phosphate, 1 n)(
t r i c h l o r o a c e t i c a c i d - 1.52 sodiun pyrophosphate and f i l t e r e d s l o w l y t h r o u g h
then separate ly washed w i t h 0. 10, 50 and 100 m l a d d i t i o n a l t r i c h l o r o a c e t i c n i t r o c e l l u l o s e f i l t e r s ( n i l l i p o r e ) . F o u r samples o f each set of digest were
a c i d - pyrophosphate so lu t ion to rmove exchangeab le t r i t ium. The f i l t e r d isks were then washed wit 2 ml o f EtOH. d r i e d and counted i n s c i n t i l l a t i o n
5H/3EP r a t i o o f pol; (~HldA-[3H1[g~P?I) ock a i l (RPI 3620) T e $/s2P r i o s were c a l c u l a t e d and c m p a r e d t o t h e
w i t h [ 3 H j ? j p H Paper chranatographic analysis of [5HlNaBH4 reduced digests samples whi h had not been t r e a t e d
of poly( [ P] f ik ] -A-T) d id not reveal any major new peak o f t r i t i u m .
d ) Tes ts f o r Carboxy l i c Ac id and E s t e r
Termini produced by t h e c l e a v a g e o f c a l f thymus DNA by (OP)2Cu++ i n t h e presence of NADH and hydrogen peroxide were examined for the presence of car- boxy l i c ac ids o r l ac tones . F reese e t a l . (19, 20) have previously dmon- s t r a t e d t h a t d e o y r i b o n i c a c i d i s formed upon incubat ion of calf-thymus DNA with Fenton's reagent, ferrous ion and hydrogen peroxide. The f i r s t s t e p i n the fo rmat ion of deoxyr ibon ic ac id wou ld be the ox ida t ion o f the C-1 carbon of the deoxyr ibose moiety t o produce a lactone. Since f ree deoxyr ibonic acid should have been d e t e c t a b l e i n t h e paper chromatograms, deoxyr ibon ic ac id remaining i n phospcd ies te r l i nkage w i th t he po l ynuc leo t i de backbone was sought. Digests of 10 mg/ml c a l f thymus DNA were prepared using 30 ut4 OP, 3 un copper and 0.1 n)( NADH and 0.5 rdl hydrogen peroxide and quenched w i t h
a l i q u o t o f t h e r e a c t i o n m i x t u r e was hydrolyzed by a lka l ine phosphatase 2,9-dimethyl-l,lO-phenanthroline. To determine the extent of cleavage. an
r e v e a l i n g t h a t one phosphorylated terminus was formed per 100 bases.
adapted t o measure carboxy l ic ac id groups assoc iated wi th po lynucleot ides (22 ) . The d i g e s t was then i ncuba ted w i th cy lohexyl-3-(2-norpholinoethyl)- c a r b o d i i n i d e r t h o - p - t o l u e n e s u l f o n a t e (lO-&,) and e i t h e r 100 nll g l y c i n e m t h y l e s t e r , t a u r i n e o r e t h y l e n e diamine. Reaction mixtures were dialyzed, and then passed through a 6-10 Sephadex column. The o l igonuc leo t ide enr iched
c h r m a t o g r a p h y f o r any n inhyd r in pos i t i ve p r imary m ines t ha t m igh t have reac- f ract ions were lyophi l ized. hydro lyzed i n 6 W HC1 and analyzed by paper
t e d w i t h pendant carboxylate groups. Calf thymus DNA digested by pancreat ic DNAase was d e r i v a t i z e d and analyzed i n a p a r a l l e l manner as a cont ro l . No taur ine, g lyc ine or e thy lenediamine could be detected by the paper ch raa -
40:6:15. Therefore there i s no ev idence for the format ion o f carboxy l ic ac id graphs using Yhatman 3" paper developed with butano1:acetic acid:water-
groups dur ing the course of the DNA cleavage react ion mediated by (OP)&u+. However, n o a p p r o p r i a t e p o s i t i v e c o n t r o l s c a l i b r a t e d t h e s e n s i t i v i t y o f t h e assay.
f o r e s t e r s was employed (21). No c o l o r was de tec tab le i n t he d iges t . Yhen acety lcho l ine (10 uM) was added t o 1 ml o f the d igest . i t s c o l o r y i e l d was
t h e d i g e s t i n h i b i t e d t h e s e n s i t i v i t y o f t h e h y d r o x a a i c a c i d t e s t f o r t h i s i d e n t i c a l t o t h a t o f a 10 UM acety lchol ine cont ro l . Therefore no cmponent of
The ca rbod i im ide me thod fo r de te rm in ing ca rboxy l i c ac ids i n p ro te ins was
TO tes t f o r l ac tones i n t he d iges ts t he f e r r i c i on -hyd roxamic ac id assay
S t r u c t u r e Of t he ca rboxy l i c ac id , i t i s p o s s i b l e t h a t an ester , i f formed, s imp le es te r . S ince co lo r y ie lds i n t h e hydroxamic acid test depend on t h e
might not be detectable under the same condi t ions as acety lchol ine. On t h e other hand, no Pos i t i ve ev idence for the ex is tence o f an e s t e r e x i s t s .
RESULTS
Identification of Inhibitors of E. coli DNA Polymerase I- The cleavage of poly(dA-T) by (OP)2Cu+ and hydrogen per- oxide yields products that are effective inhibitors of polym- erase I and incompetent to serve as primer-template. The inhibition of polymerase I can serve as a sensitive assay of the chemical structure of the termini if free OP has no role in the mechanism of inhibition. Although our previous experiments showed the progressive increase of inhibitory products and the loss of primer-template competence as a function of time of incubation of poly(dA-T) with OP, cupric ion, and thiol under aerobic conditions (l), they did not exclude the partic- ipation of free OP in the inhibition nor did they indicate if OP covalently linked to oligonucleotide fragments was essential for the observed kinetic effects. The data summarized in Fig. 1 demonstrate that OP in any form is not required for the inhibition of polymerase I subsequent to the cleavage reaction. In this experiment, 5-[3H]acetamido-l,10-phenanthroline was used to cleave DNA with cupric ion and mercaptopropionic acid under aerobic conditions. At 5, 10, and 20 min, aliquots of the reaction mixture were quenched with 2,9-dimethyl-1,10- phenanthroline, precipitated with ethanol, and redissolved in buffer. The observed inhibition of polymerase I increased with time even though ["H]5-NacOP was quantitatively removed.
In separate experiments, the inhibitory potency of cleavage reaction mixtures which had been quenched with 2,9-di- methyl-OP and EDTA was undiminished by repeated extrac- tion with isoamyl alcohol. Virtual reaction mixtures, lacking poly(dA-T), do not produce inhibitors of polymerase I. All the available data suggest that the inhibitor(s) formed during the
DNA cleavage reaction are derived entirely from the polynu- cleotide. The oligonucleotide fragment and free ligand do not exhibit synergistic inhibition.
Relief of Inhibition and Restoration of Primer-Template Competence by Alkaline Phosphatase and Exonuclease III- DNA fragments bearing 3'-phosphorylated ends are effective inhibitors of polymerase I and are not competent to serve as primers for the polymerization reaction (23-25). To test for the formation of those termini by (OP)&u+, digests of poly(dA-T) were prepared and treated with alkaline phospha- tase and exonuclease 111. The ability of alkaline phosphatase to alter the inhibitory and primer-template competence of products of the cleavage was tested using a reaction mixture in which 20 pg/ml poly(dA-T) had been incubated with 7.5 p~ OP and 3.75 p~ cu2+ in the presence of 7 mM MPA for 30 min. After the cleavage reaction was quenched by the addition of 2,9-dimethyl-l,lO-phenanthroline and the resulting solution was heated to 65 "C, 1 unit of alkaline phosphatase was added followed by a second unit 10 min later. After an additional 10 min of incubation, the reaction mixture was heated in boiling water (10 min) to inactivate the alkaline phosphatase and the DNA was assayed as template for polymerase I in the presence of dATP and TTP (2 X M each) or as an inhibitor in the standard assay mixture. The data summarized in Fig. 2 dem- onstrate that alkaline phosphatase both restores primer-tem- plate competence to the DNA cleaved by OP and copper ion and sharply diminishes its inhibitory potency.
Parallel experiments were carried out with exonuclease 111. This enzyme has 3 characteristic catalytic activities: ( a ) 3'- phosphomonoesterase activity; ( b ) 3'-5' exonuclease activity; and ( c ) endonuclease activity at apurinic sites (26). An aliquot (0.100 m l ) of a digest of poly(dA-T) prepared as outlined above was incubated with exonuclease I11 (125 units) for 30 min at 37 "C followed by boiling to inactivate the enzyme. As a result, the primer-template competence of the DNA prod- ucts was restored and its inhibitory properties were abolished (Fig. 3). Based on the parallel results with alkaline phospha- tase, the 3'-phosphomonoesterase activity must be primarily responsible for the observed effects on the polymerization reaction. However, exonuclease I11 is more effective than alkaline phosphatase. A plausible explanation for this obser- vation is that the endonuclease activity may excise an addi- tional nonproductive furan derivative at the 3' termini whose formation is postulated on the basis of our other data (see Fig. 7).
Three other lines of investigation support the proposal that 3'-phosphorylated termini are the cleavage products respon- sible for the inhibition of polymerase I. First, preincubation of
l ' 1 ' 1 ' , ' , , , ' , ' , .
0 100 ; 0
a L' 40
ap 20 I
20 aQ
'0 2 6 I O 14 18 2 2 26 30 t , rnm
FIG. 1. Oligonucleotide products are solely responsible for polymerase I inhibition. Poly(dA-T) (IO pg/ml) was cleaved by r3H]5-NacOP (20 p ~ ) , cupric acetate (2 p ~ ) , and MPA (7 mM). At the indicated times, aliquots (20 pl ) of reaction mixtures were quenched with 2,g-dimethyl OP and the polynucleotide was precipitated with EtOH. After redissolving the oligonucleotides in assay buffer (75 pl), 10 pl were counted to determine the fraction of ["H]5-NacOP remain- ing and 50 p1 were added to a polymerase (Pol) I assay mixture (1). Polymerase I activity (0); ["H]5-NacOP removed (0).
12124 Products of DNA Cleavage
3 6 9
.""-. m "
Minutes Minutes
FIG. 2 (left). Relief of inhibition and restoration of primer- template competence of (OP)zCu+ digests of poly(dA-T) by alkaline phosphatase. Incorporation of r3H]TTP (2 x M) into acid-precipitable counts by polymerase 1 in an assay mixture also containing dATP (2 X M) and the following components: 75 pl of (OP)zCu+ digest (U); 75 pl of (OP)&u+ digest and poly(dA-T) (10 pg/ml , &--A); 75 p1 of (OP)2Cu' digest treated with alkaline phosphatase (A-A); 75 pl of (OP),Cu' digest reacted with alkaline phosphatase and poly(dA-T) (10 pg/ml, 0- - -0); control, poly(dA-T) (10 pg/ml, W). Total volume of assay is 0.25 ml, T = 37 "C, pH 7.0,0.05 M Tris-acetate buffer; specific activity of r3H]TTP is 0.2 pCi/ nmol. See text for additional details.
FIG. 3 (right). Relief of inhibition and restoration of primer- template competence of (OP)zCu+ digests of poly(dA-T) by exonuclease 111. Incorporation of ['HITTP (2 X M) into acid- precipitable counts by polymerase I in assay mixture also containing dATP (2 X M) and the following components; 75 pl of (OP)&u' digest (C - 4); 75 p1 of (OP)2Cu+ digest and poly(dA-T) (10 pg/ml, A-A); 75 pl of (0P)zCu' digest reacted with exonuclease I11 (10 pg/ml, A-A); 75 p1 of ( 0 P ) ~ c u ' digest reacted with exonuclease 111 and poly(dA-T) (10 pg/ml, M); control, poly(dA-T) (10 pg/ ml, 0- - -0).
the cleaved DNA with polymerase I fails to decrease the inhibition of the polymerization activity. Since the 5'-3' and 3-5' exonucleases are unable to excise 3'-phosphorylated ter- mini, this result is anticipated if inhibition is due to 3"phos- phorylated ends. Secondly, the products of the (OP)zCu+ cleavage of DNA are also effective inhibitors of the pyrophos- phate exchange reaction as well as net polymerization (27). This partial activity of the polymerase activity requires a competent 3"hydroxyl but is not processive and therefore would not be expected to be blocked by unusual 5' termini (28). Its inhibition by the products of the cleavage reaction is consistent with the formation of the unproductive 3'-phospho- rylated ends. Finally, the failure of a variety of physical and chemical procedures to relieve inhibition can be understood on the basis of polymerase I inhibition being attributable to 3'-phosphorylated termini. For example, the inhibition is un- changed if digests are heated at 100 "C and either quick cooled or slowly renatured. Exposure to extremes of pH (0.1 M HC1 for 6 min, 0.01 N NaOH for 5 min) similarly fails to relieve inhibition. Likewise ineffective was reaction of the digests with dimedone and sodium borohydride.
Net Phosphodiester Bond Cleavage-Our finding that 3'- phosphorylated termini are primarily responsible for the in- hibition of polymerase I prompted us to quantitate the yield of phosphorylated termini formed in the cleavage reaction at pH 7.0 and 37 "C. This was achieved by measuring the release of inorganic phosphate by alkaline phosphatase from ~ O ~ ~ ( [ ~ H , ~ ~ P ] ~ A - [ ~ H ] T ) which had been degraded by OP and Cu2+ at pH 7.0 (Tris-acetate) in the presence of 7 mM MPA, 7 mM H202. Prior to the addition of alkaline phosphatase, the reaction mixture was quenched by catalase. The "P-labeled phosphate released was extracted into butyl acetate-molyb-
date and counted. One phosphate was released for every 4-5 nucleotides in the DNA initially used as substrate (Table I). This result was the same if [CY-~~PITTP or [a-"PIdATP was the source of the label or if the cleavage reaction was poten- tiated by NADH and hydrogen peroxide. Although these results demonstrate that the phosphodiester bond of poly(dA- T) is extensively cleaved to form phosphorylated termini under physiological conditions, minor pathways generating alkaline- or acid-labile sites cannot be excluded.
5'-phosphorylated Termini-The high yield of phosphate termini immediately raises the question of whether 5"phos- phorylated termini are also produced. The restoration of the competence of cleaved poly(dA-T) as a primer-template by alkaline phosphatase and exonuclease I11 demonstrates that 3'-phosphorylated ends are formed but does not allow any estimations of their yield or exclude the possibility that 5'- phosphorylated ends are formed as well. Polynucleotide ki- nase, in conjunction with alkaline phosphatase, was used to demonstrate that 5'-phosphorylated termini are also formed in the (OP)&u+ cleavage of DNA. Although polynucleotide kinase-catalyzed phosphorylation is measurable and repro- ducible prior to alkaline phosphatase hydrolysis, the titer of ends phosphorylated by [Y-~'P]ATP is %fold greater following alkaline phosphatase treatment. However, the yield of ends that are substrates for polynucleotide kinase is low when compared to the yield of alkaline phosphatase-labile phos- phate. Only 1 phosphate is added by polynucleotide kinase per 43 nucleotides (Table 11) whereas 1 phosphate is liberated per 5 nucleotides by alkaline phosphatase from cleaved p~ly([~H,~*PldA-T) (Table I). Two experimental problems may contribute to the observed 12.5% yield of 5'-phosphoryl- ated ends. First, oligonucleotides with a higher molecular weight than average may be bound to the nitrocellulose filters used to separate the polynucleotide from excess ATP. Sec- ondly, the efficiency of the phosphorylation of the products
TABLE I Inorganic phosphate released by alkaline phosphate from cleaved
DNA Conditions were Tris-acetate buffer, 0.05 M, pH 7.0; 37 "C. Com-
parable results were obtained when DNA was cleaved in the presence of NADH and Hz02.
DNA
mm P (in- itial
DNA)/ mm P (ex- Cleavage conditions
TABLE I1 Polynucleotide kinase catalyzedphosphorylation by [y3'PJATP of
DNA cleaved by (0P)zCu' See "ExDerimental Procedures" for details. Comparable results
were obta&ed with cleavage caused by OP, Cu", NADH, and HzOZ. Retention of ni-
ters Cleavage conditions phosphatase trocellulose fil- ''W''P"
by polynucleotide kinase is uncertain. For example, unusual structures formed by the cleavage reaction (see Fig. 7 ) may be effective inhibitors of the kinase. On the basis of the available data, it is possible to conclude that both 5'- and 3'-phospho- rylated ends are formed, although the relative yield cannot be assessed.
Physical Size of Fragments-Among the possibilities which could account for the ratio of 1 phosphorylated tenninus/5 phosphates initially present in the DNA would be a penta- nucleotide with a single phosphorylated terminus, a decanu- cleotide with 2 phosphorylated termini, or equimolar mixtures of 2 pentanucleotides, one lacking terminal phosphates and one phosphorylated at both termini. To determine the phys- ical size of all the products, digests prepared from uniformly labeled ~oly([~H]dA-T) were fractionated on a DEAE-cellu- lose column developed with a linear NaCl gradient in a buffer containing 20 m~ Tris-HC1, 7 M urea. The column was cali- brated with a DNase I digest of the same DNA that was spiked with 5'-dAMP (Fig. 4). The peak which emerges just prior to the application of the salt gradient, which is minor in the DNase I digest but significant in the (OP)2Cu+ digest, is the free base adenine (see Fig. 5 ) . Approximately 15% of the total radioactivity in the (OP)2Cu+ digest was present in this peak (see Table 111). In view of the excellent resolution observed with the DNase I digest, the complexity of the chromatogram of the (OP)2Cu+ digest must reflect a hetero- geneous set of products with an average of 6 bases/fragment assuming that the 5'-phosphorylated oligonucleotides made by DNase I are appropriate standards for the unusual nucleo- tides formed in the (OP)zCu+ cleavage. Since the formation of both 5'- and 3'-phosphorylated termini has been demon- strated, the heterogeneous array of products formed from uniformly labeled DNA is anticipated. Neglecting products that might be derived from the original termini of the poly(&- T), 4 oligonucleotide products/given chain length will be
IO
4
2 ; 4.0
3.0
2 .o
I .o
0 v 0 20 40 60 80 100 I20
E 8 F m . , 8
; : 94.0
v 0 20 40 60 80 100 I20
E 8 F m . , 8
; : 94.0
Fraction Number
FIG. 4. Fractionation of oligonucleotide products on DEAE- cellulose. Top, products of DNase I digestion of ~oly([~H]dA-T). Bottom, products of (0P)~cu ' cleavage. The counts for the oligonu- cleotides have been corrected by dividing the counts/min that eluted from the column by the chain length of the oligonucleotide. It is assumed that the oligonucleotides produced in the (OP)&u+ cleavage have the same chain length as those produced in the DNase I digest even though they have different termini. The solid arrow on the bottom indicates the point a t which 2 M NaCl was applied to the column. A DE52 column (10 X 1 cm) in 20 mM Tris-HC1, pH 7.6, 7 M urea was eluted with a 0-0.3 M NaCl gradient. See "Experimental Procedures" for additional details.
A
c m from origin
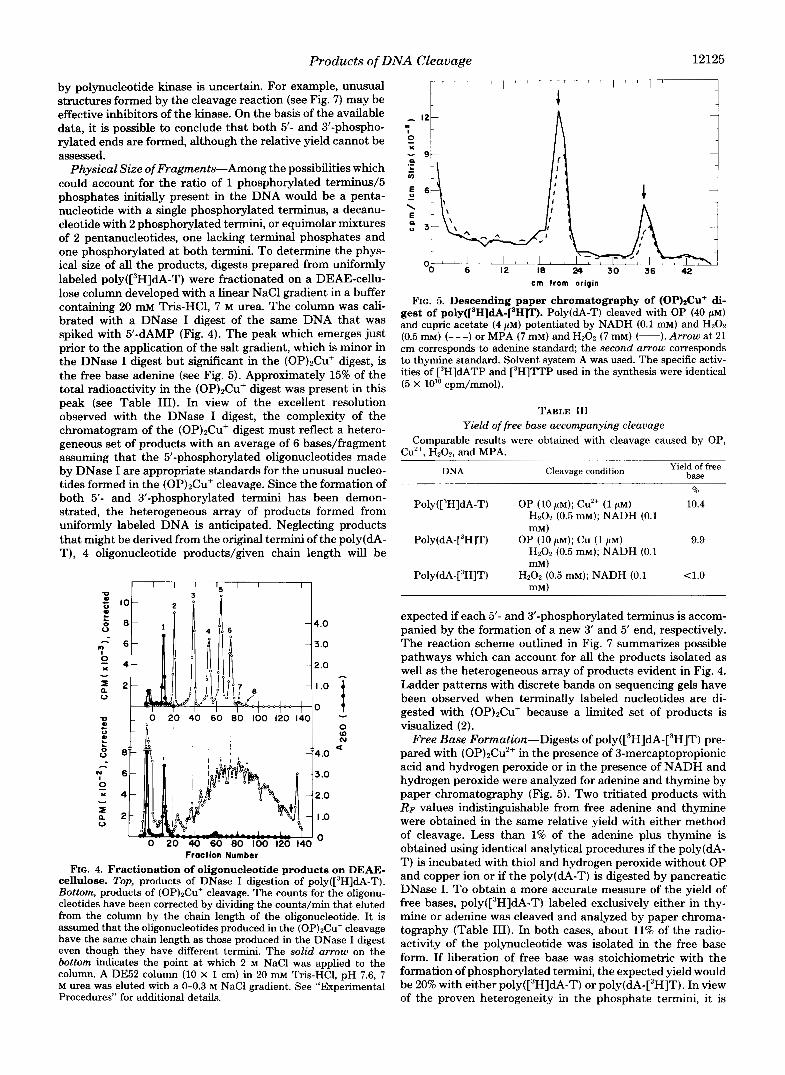
FIG. 5. Descending paper chromatography of (OP)aCu+ di- gest of p0ly(['H]dA-[~H3T). Poly(dA-T) cleaved with OP (40 p ~ ) and cupric acetate (4 PM) potentiated by NADH (0.1 m ~ ) and Hz02 (0.5 m ~ ) (- - -) or MPA (7 mM) and Hz02 (7 mM) (-).Arrow at 21 cm corresponds to adenine standard; the second arrow corresponds to thymine standard. Solvent system A was used. The specific activ- ities of r3H]dATP and r3H]TTP used in the synthesis were identical (5 X 10" cpm/mmol).
TABLE I11 Yield of free base accompanying cleavage
Comparable results were obtained with cleavage caused by OP, Cu". H202. and MPA.
DNA Cleavage condition Yield of free base
Pol~(r~H1dA-T) OP (10 p ~ ) ; Cu" (1 p ~ ) 10.4 Hz02 (0.5 mM); NADH (0.1 mM)
HzOz (0.5 mM); NADH (0.1 mM)
mM)
Poly(&-["HIT) OP (10 p ~ ) ; Cu (1 p ~ ) 9.9
Po~Y(~A-[~H]T) Hz02 (0.5 mM); NADH (0.1 4 . 0
expected if each 5'- and 3'-phosphorylated terminus is accom- panied by the formation of a new 3' and 5' end, respectively. The reaction scheme outlined in Fig. 7 summarizes possible pathways which can account for all the products isolated as well as the heterogeneous array of products evident in Fig. 4. Ladder patterns with discrete bands on sequencing gels have been observed when terminally labeled nucleotides are di- gested with (OP)2Cu+ because a limited set of products is visualized (2).
Free Base Formation-Digests of ~ o ~ ~ ( [ ~ H ] ~ A - [ ~ H ] T ) pre- pared with (0P)2Cu2+ in the presence of 3-mercaptopropionic acid and hydrogen peroxide or in the presence of NADH and hydrogen peroxide were analyzed for adenine and thymine by paper chromatography (Fig. 5). Two tritiated products with RF values indistinguishable from free adenine and thymine were obtained in the same relative yield with either method of cleavage. Less than 1% of the adenine plus thymine is obtained using identical analytical procedures if the poly(dA- T) is incubated with thiol and hydrogen peroxide without OP and copper ion or if the poly(dA-T) is digested by pancreatic DNase I. To obtain a more accurate measure of the yield of free bases, p~ly([~H]dA-T) labeled exclusively either in thy- mine or adenine was cleaved and analyzed by paper chroma- tography (Table 111). In both cases, about 11% of the radio- activity of the polynucleotide was isolated in the free base form. If liberation of free base was stoichiometric with the formation of phosphorylated termini, the expected yield would be 20% with either p~ly([~H]dA-T) or ~oly(dA-[~H]T). In view of the proven heterogeneity in the phosphate termini, it is
12126 Products of DNA Cleavage
possible that free base release accompanies one of the possible cleavage reactions (e.g. Pathway A, Fig. 7 ) .
Attempts to Identifi Fragments of Deoxyribose Moiety Kinetics and Products of Cleavage of POZY(['~C,~H]~A-T)- Poly(dA-T) was prepared using uniformly labeled [ 14C]dATP and r3H]dATP labeled in adenine only. The goal was to identify any products in the chromatography which might be derived exclusively from the deoxyribose moiety. Digests were prepared using 7 mM hydrogen peroxide, 7 mM mercaptopro- pionic acid or 0.1 mM NADH, 0.5 m~ hydrogen peroxide. Under each reaction condition, the rates of formation of the 14C and 3H acid-soluble counts were the same (Fig. 6). No material could be detected by descending paper chromatog- raphy using propanol-l/NHa/HzO that contained only 14C-
labeled material. A spot with a migration indistinguishable from that of adenine and a 3H/'4C ratio consistent with the formation of this product were present in both chromato- grams.
Lack ofAldehyde and Ketone Products-Free base release implies that the oxidation of the deoxyribose is responsible for cleavage of the phosphodiester backbone. The possibility of hydrogen peroxide, an a-nucleophile, degrading the DNA by nucleophilic attack in a reaction catalyzed by the coordi- nation complex is not consistent with the absolute specificity of the reaction for redox-active copper ion or with the lack of reactivity of the redox-stable 2:l 2,9-dimethyl OP-cuprous complex.
Since bleomycin degrades DNA by an oxidative pathway generating malondialdehyde as one of its products (29-31), digests of poly(dA-T) produced by (OP)ZCu+ have been ana- lyzed for this dialdehyde using thiobarbituric acid and for aldehydes and ketones with dimedone (32) and NaB3&. The products of the bleomycin cleavage reaction provide a positive control for the analytical techniques employed. The data summarized in Table IV indicate that no barbituric acid-
Mlnuter Minuter
FIG. 6. Cleavage of poly(dA-T) prepared from ['HJdATP la- beled in adenine only and p-'4C]dATP by OP and c ~ * + . A, formation of acid-soluble counts from poly(dA-T) (4-5 pg/ml) by OP (20 p ~ ) and Cu2+ (2 p ~ ) in the presence of MPA (7 m ~ ) and Hz02 (7 mM). B, formation of acid-soluble counts from poly(dA-T) (4-5 p g / mi) by OP (20 p ~ ) and Cu2' (2 p ~ ) in the presence of NADH (0.1 mM) and Hz02 (0.5 mM).
TABLE IV Thiobarbituric acid-reactive products in bleomycin and (OP)zCu+-
cleaved DNA See "Experimental Procedures" for details. 28.3 p g / d p01y([~H]
dA-r3H?*P1T) was the substrate for the cleavage reaction. ~
532 nm &Wac. No' Of Absorbance at
Cleavage condition teristic of mdon- tidedteminal dialdehvde phosphate
positive material is produced by (OP)&u+. Similarly, although dimedone-reactive materials were identified in bleomycin di- gests upon paper chromatography, none was observed in (OP)ZCU+ digests. Finally, no product of (OP)&u+ cleavage reducible by NaB3& could be detected on nitrocellulose fiiters or by descending chromatography using propanol-l/ammo- nia/water as the solvent developer.
Analysis for Esters and Lactones-Calf thymus DNA (10 mg/ml) was cleaved with (OP)zCu+ and H202 and oligonucle- otide products were tested for terminal carboxylic acid groups and esters as described under "Methods" (see Miniprint). None was found.
DISCUSSION
The chemical cleavage of DNA by (OP)zCu+ and H202 was initially discovered because the products of this reaction are effective inhibitors of polymerase I. The demonstration that 3'-phosphorylated termini inhibit the enzyme without the participation of OF subsequent to the DNA cleavage reaction conf i i s our previous suggestion that the zinc ion of polym- erase I is not accessible to coordination by the ligand on the time scale expected for catalysis. One mechanism of OP inhi- bition of polymerase I reflective of an essential role for zinc ion that is not directly affected by the present data is the removal of the metal ion upon prolonged incubation (33, 34). However, 3'-phosphorylated ends are potent inhibitors of polymerase I and preparations of polymerase I invariably contain traces of DNA. Therefore, any demonstration of po- lymerase I inhibition by OP claiming to reflect coordination of the zinc must be accompanied by an appropriate control excluding the formation of 3'-phosphorylated termini either during the course of incubation with the enzyme or during the subsequent assay.
Although identification of 3'-phosphorylated termini as in- hibitors of polymerase I has not provided novel insights into the active site chemistry of polymerase I, important additional properties of the reaction relevant to its potential use as a probe of DNA secondary structure have been revealed as a result of the product analysis. The most unique property of the reaction is its cleavage of double-stranded DNA without regard to primary sequence (2). In the present report, we have demonstrated that the yield of 32P-phosphorylated termini hydrolyzable by alkaline phosphate is the same whether [a-
T). This result is expected for random cleavage. In contrast, pancreatic DNase I cleaves poly(dA-T) preferentially at the Ap-T bond (35). The lack of specificity of (OP)&u+ in the cleavage of the poly(dA-T) co-polymer whose secondary struc- ture is a variant B-form helix (35-37, is reflective of the more random cleavage pattern exhibited by the coordination com- plex relative to DNase and may constitute one of its advan- tages relative to the enzyme in footprinting applications ( 11). The importance of double helical DNA for the cleavage re- action is emphasized by the observation that the average size of the oligonucleotide products after exhaustive digestion is 5 & 1 bases. These oligonucleotides are too small to form stable helical structures and as a consequence would not be sub- strates for the reaction. Other evidence of the specificity of the cleavage for double-stranded DNA includes the lack of reactivity of single-stranded poly(dA) and poly(dT) and the inhibition of scission of poly(dA-T) by intercalating agents (2).
The formation of a reactive intermediate between double- stranded DNA and (OP)ZCU+ during the cleavage provides the best explanation for the secondary structure specificity of the reaction. We have previously proposed 3 possible path- ways for strand scission after the binding of the coordination
32 PIdATP or [a-32P]TTP is used to synthesize the poly(dA-
Products of DNA Cleavage 12127
complex to the DNA. They include: ( a ) hydrogen peroxide acting as a nucleophile; ( b ) formation of a peroxy-phenan- throline intermediate; and (c) Fenton-type chemistry in which hydrogen peroxide oxidizes (OP)2Cu+ intercalated into the DNA, yielding the equivalent of a metal ion-bound hydroxyl radical (I) capable of oxidatively degrading the deoxyribose.
L
1
The fist alternative is unlikely because of the isolation of free bases during the reaction. They would not be formed if phosphodiester bond cleavage results from the attack of hy- drogen peroxide on the phosphodiester bond. The second alternative, although feasible, is not supported by our unsuc- cessful attempt to find a phenanthroline derivative produced only in the DNA cleavage reaction. The third possibility (Equation 1) is supported by the (OP)2Cu2+-catalyzed oxida- tion of both 2-electron and 1-electron donors by hydrogen peroxide (3,4) and the importance of ferry1 species (e.g. Fe(1V)
e
i' ( A - 4 Pathway E
.' * I 0.P.O 4 + :Q" 0
0 new 3'--.na 0-t.0
0 I;* F;:
*OC*,
3" 0-?a
i' ".. 5"m
0
Pathway A
from the DNA-(OP)&u+ complex. FIG. 7. Proposed scheme for the cleavage reaction starting
= 0) in the mechanism of action of cytochrome P-450s (38). Moreover, DNA is cleaved by hydroxyl radicals generated from (a) O2 and H202 by the Haber-Weiss reaction; ( b ) pulse radiolysis; and (c) UV irradiation (39). Recently, an ethidium bromide derivative with a pendant iminodiacetic acid has been demonstrated to cleave DNA rapidly in the presence of ferrous ion and molecular oxygen (40).
The scheme summarized in Fig. 7 outlines the reaction pathway which accounts for the observed products and as- sumes the Fenton-type chemistry outlined in Equation 1. The initial chemical event is the 1-electron oxidation of the deox- yribose oxygen. The subsequent removal of either the C-4' hydrogen atom ( A - I ) or C-1' hydrogen atom ( B - I ) leads to interconvertible oxenium ions. Pathway B can generate the 5'-phosphorylated ends observed in about a 15-20% yield. Pathway A which is probably predominant would proceed by liberating free base (A-3) with subsequent formation of 3'- phosphorylated ends (A-4) . In addition to accounting for all the isolated products, this scheme can also explain our failure to identify ketones, aldehydes, acids, and esters in digests. The '2 types of resonance-stabilized furans would be inert to the derivatizations attempted.
In summary, the co-reactants (OP)2Cu+ and H 2 0 n make single-stranded nicks in double-stranded DNA a t physiologi- cal temperature and pH values with no apparent primary sequence specificity. Used in conjunction with the nuclease S1, which is specific for single-stranded DNA, and DNase I, which clips both single-stranded and double-stranded DNA, this reaction should provide a useful additional tool to probe DNA-ligand interactions and the secondary structure of DNA. Although this artificial DNase-like activity does not produce single products characteristic of enzymatic reactions, it does form a noncovalent intermediate analogous to an enzyme- substrate complex. This induced approximation (41) com- bined with a productive orientation of the coordination com- plex on the surface of the DNA must contribute to the rate of this remarkable reaction.
Acknowledgments-We have profited from the insightful com- ments of Professor 0. Chapman and his co-workers from the Depart- ment of Chemistry.
REFERENCES
1. Sigman, D. S., Graham, D. R., D'Aurora, V., and Stern, A. M.
2. Marshall, L. E., Graham, D. R., Reich, K. A,, and Sigrnan, D. S.
3. Graham, D. R., Marshall, L. E., Reich, K. A., and Sigman, D. S.
4. Reich, K. A., Marshall, L. E., Graham, D. R., and Sigman, D. S.
5. D'Aurora, V., Stern, A. M., and Sigman, D. S. (1977) Biochem.
6. D'Aurora, V., Stern, A. M., and Sigman, D. S. (1978) Biochem.
7. Que, B. G., Downey, V. M., and So, A. G. (1979) Biochemistry 18,
8. Downey, V. M., Que, B. G., and So, A. G. (1980) Biochem.
9. Que, B. G., Downey, V. M., and So, A. G. (1980) Biochemistry 19,
(1979) J. Biol. Chem. 254, 12269-12272
(1981) Biochemistry 20,244-250
(1980) J. Am. Chem. SOC. 102, 5419-5421
(1981) J. Am. Chem. SOC. 103, 3582-3584
Biophys. Res. Commun. 78, 170-176
Biophys. Res. Commun. 80, 1025-1032
2064-2068
Biophys. Res. Commun. 93,264-270
5987-5991 10. Fridovich, I. (1978) Science 201, 875-880 11. Schmitz, A., and Gales, D. J . (1979) Nucleic Acids Res. 6, 111-
12. Cartwright, I. L., and Elgin, S. C. R. (1981) J. Cell Biol. 91, 64
13. Jovin, T. M., Englund, P. T., and Bertsch, L. L. (1969) J. Biol.
14. Stern, A. (1977) Ph.D. thesis, University of California, Los Angeles 15. Weiss, B., Live, T. R., and Richardson, C. C. (1968) J. Biol. Chem.
16. Tomlinson, R. V., and Tener, G. M. (1963) Biochemistry 2, 697-
137
(Abstr. 3050)
Chem. 244,2996-3008
243,4530-4542
12128 Products of DNA Cleavage
702 17. Friedman, E. A., and Smith, H. 0. (1972) J. Bwl. Chem. 247,
18. Sausville, E., Stein, R. W., Peisach, J., and Horowitz, S. B. (1978) Biochemistry 17, 2746-2754 and references cited therein
19. Rhaese, H.-J., and Freese, E. (1968) Biochim. Biophys. Acta 155,
20. Rhaese, H.-J., Freese, E., and Melzer, M. S. (1968) Biochim.
21. Hestrin, S. (1949) J. Biol. Chem. 180,249-261 22. Glazer, A. N., DeLange, R., and Sigman, D. S. (1975) Chemical
Modification of Proteins. Selected Methods and Analytical Procedures, North-Holland and American Elsevier Publishing Go., New York
23. Kornberg, A. (1980) DNA Replication, p. 107, W. H. Freeman and Co., San Francisco
24. Englund, P. T., Kelly, R. B., and Kornberg, A. (1969) J. Biol. Chem. 244, 3045-3052
25. Kappen, L. S., and Goldberg, I. H. (1978) Biochemistry 17, 729- 734
26. Weiss, B. (1976) J. Biol. Chem. 251, 1896-1901 27. D’Aurora, V. (1978) Ph.D. thesis, University of California, Los
28. Deutscher, M. P., and Kornberg, A. (1969) J. Biol. Chem. 244,
29. Burger, R. M., Berkowitz, A. R., Peisach, J., and Honvitz, S. B.
2846-2853
476-490
Biophys. Acta 155, 491-504
Angeles School of Medicine
3019-3028
(1980) J. Biol. Chem. 255, 11832-11838 30. Grollman, A. P., and Taklshlta, M. (1979) Adu. Enzyme Regul.
31. Sausville, E. A., and Honvitz, S. B. (1979) in Effects of Drugs on the Cell Nucleus (Busch, H., Brooke, S. T., and Daskul, Y., eds) pp. 181-205, Academic Press, New York
32. Glitz, D. G., and Sigman, D. S. (1972) Biochemistry 9,3433-3439 33. Poiesz, B. J., Seal, G., and Loeb, L. A. (1974) Proc. Natl. Acad.
Sci. U. S. A. 71,4892-4896 34. Springgate, C . F., Mildvan, A. S., Abramson, R., Engle, J. L., and
Loeb, L. A. (1973) J. Biol. Chem. 248,5987-5993 35. Scheffler, I. E., Elson, E. L., and Baldwin, R. L. (1968) J. Mol.
Biol. 36, 291-304 36. Lomonossoff, G. P., Butler, P. J. G., and Klug, A. (1981) J. Mol.
Biol. 149, 745-760 37. Klug, A,, Jack, A., Viswamitra, M. A., Kennard, O., Shakked, Z.,
and Steitz, T . A. (1979) J. Mol. Biol. 131,669-680 38. Walsh, C. (1979) Enzymatic Reaction Mechanisms, pp. 468-473,
W. H. Freeman and Go., San Francisco 39. Repine, J. E., Pfenninger, V. W., Talmage, D. W., Berger, E. M.,
and Pettijohn, D. E. (1981) Proc. Nutl. Acad. Sci. U. S. A . 78,
40. Hertzberg, R. P., and Dervan, P. B. (1982) J. Am. Chem. SOC.
41. Jencks, W. (1969) Catalysis in Chemistry and Enzymology,








![Supplementary InformationSynthesis and Characterization of Compounds. 1-methyl-1H-pyrazolo[3',4':5,6]pyrazino[2,3-f][1,10]phenanthroline (L1) 1,10-phenanthroline-5,6-dione (200 mg,](https://static.documents.pub/doc/80x56/5f71d2c7b455b50ab327003e/supplementary-synthesis-and-characterization-of-compounds-1-methyl-1h-pyrazolo3456pyrazino23-f110phenanthroline.jpg)

![Bis[chloridobis(1,10-phenanthroline)copper(II ... · + cations (phen is 1,10-phenanthroline), [Fe(CN) 5 NO] 2 anions and one di-methylformamide (DMF) solvent molecule of crystallization](https://static.documents.pub/doc/80x56/603d32ed89872c77881dd664/bischloridobis110-phenanthrolinecopperii-cations-phen-is-110-phenanthroline.jpg)




![Synthesis of 5,6-dihydrobenzo[1,7]phenanthroline …shodhganga.inflibnet.ac.in/bitstream/10603/37715/6/015...102 Chapter 5 Synthesis of 5,6-dihydrobenzo[1,7]phenanthroline and Quinazolinone](https://static.documents.pub/doc/80x56/5f0ccf247e708231d4373cce/synthesis-of-56-dihydrobenzo17phenanthroline-102-chapter-5-synthesis-of.jpg)








